Submitted:
24 January 2024
Posted:
25 January 2024
You are already at the latest version
Abstract
In order to explore the effect of fertilization patterns on the growth of rapeseed seedlings under waterlogging stress, three fertilization patterns (conventional fertilization, supplemental organic fertilization, and supplemental microbial fertilization) were set up using the variety Xiangyou 708 as the material, and waterlogging treatment was carried out during the seedling stage of rapeseed. The effects of fertilization patterns on the growth of rapeseed seedlings and rhizosphere microorganisms under waterlogging stress were investigated. The results showed that all three fertilization patterns exhibited that waterlogging stress inhibited the growth of rapeseed seedlings, inhibited root activity, and changed the structure of rhizosphere bacterial colonies. However, supplemental organic fertilization and microbial fertilization could better promote the growth of rapeseed seedlings, reduce the impact of waterlogging stress on the growth of rapeseed seedlings, and accelerate the recovery of rapeseed seedlings after waterlogging stress. In addition, supplemental microbial fertilization could also change the structure of rhizosphere bacterial colonies. Under normal water supply, supplemental organic fertilization enriched P_Bacteroidota, P_Actinobacteriota, P_Chloroflexi, and G_Flavisolibacter in the rhizosphere soil of rapeseed, while supplemental microbial fertilization enriched P_Bacteroidota and G_Flavisolibacter in the rhizosphere soil of rapeseed. After 7 days of waterlogging treatment, supplemental organic fertilization enriched P_Verrucomicrobiota in the rhizosphere soil of rapeseed, while supplemental microbial fertilization enriched P_Actinobacteriota, G_SC-I-84, and G_Ellin6067 in the rhizosphere soil of rapeseed. The enrichment of these bacteria may be related to the growth promotion and waterlogging tolerance of rapeseed. This study provides evidence that microbial fertilizer and organic fertilizer can promote the growth of rapeseed and enhance its waterlogging tolerance, as well as evidence that some rhizosphere microorganisms have a potential role in promoting the growth and waterlogging tolerance of rapeseed.
Keywords:
fertilization model
; rapeseed
; waterlogging tolerance
; rhizosphere microorganisms
; seedling growth
1. Introduction
Rapeseed is one of the most important oil crops in China, mainly using the planting mode of rapeseed and rice rotation. This planting mode accounts for more than 75% of China's total rapeseed production. The rapeseed production areas in the Yangtze River Basin have more rainfall in spring and autumn, which often leads to higher soil viscosity and poor ventilation for rapeseed growth due to the previous crop of rice. Coupled with the high groundwater level caused by previous crops, waterlogging is extremely prone to occur. [1,2,3,4].
When waterlogging occurs, due to the high water content in the soil, it has a significant impact on crop root respiration and normal metabolism, coupled with the accumulation of toxic substances in the roots, which can lead to slow growth or even death of crops. In addition, due to the impact of waterlogging on soil, the physicochemical properties of soil change, which also has an impact on plant rhizosphere soil microorganisms. Studies have shown that when crops are under stress, beneficial microorganisms can enhance the resistance of crops [5,6]. When adversity strikes, plant roots release signals to recruit microorganisms that have a positive effect on resisting adversity. This mechanism plays an important role in plants suffering from waterlogging stress [7,8,9]. After the end of waterlogging, plant roots are often damaged, and due to the slow recovery of roots, this leads to a significant lag effect of waterlogging[10].
In recent years, research on the tolerance of rapeseed to waterlogging has mainly focused on the genetic level, while there have been few reports on studying the tolerance of rapeseed to waterlogging by adjusting the fertilization mode. Microbial fertilizer is a kind of fertilizer made by mixing a variety of beneficial microorganisms, substrates, and some carriers that maintain microbial survival. The application of microbial fertilizer can improve the microbial environment in soil, increase the number of beneficial strains in soil, stimulate crop growth, and enhance stress resistance [11]. Organic fertilizer is made from animal waste and plant residues after fermentation and decomposition [12]. The most significant effect of applying organic fertilizer is to increase the content of organic matter in soil and regulate the balance of soil nutrients [13]. In addition, organic fertilizer can also improve soil texture, which indirectly promotes plant root absorption of nutrients in soil, promotes soil microbial activity, and increases biomass in soil [14]. These effects directly or indirectly promote crop growth and stress resistance. In recent years, with the use of microbial fertilizers and organic fertilizers in agricultural production, the effects of these new fertilizers on crop yield increase and stress resistance enhancement have been increasingly recognized [13,15]. Therefore, it is very meaningful to explore the application of organic fertilizers and microbial fertilizers to study the tolerance of rapeseed to waterlogging. In this experiment, different fertilization modes and whether to carry out flooding treatment were used as variables to study the physiological indicators and rhizosphere soil bacterial species and community structure of rapeseed. The aim is to explore the effects of fertilization modes on the tolerance of rapeseed to waterlogging and rhizosphere soil microorganisms, providing scientific evidence for enhancing the tolerance of rapeseed to waterlogging by changing the fertilization mode.
2. Materials and Methods
2.1. Overview of the experiment site
The experiment was conducted in 2022-2023 at the Hunan Agricultural University Internship Base in Liuyang City, Hunan Province (113°83′46′′E, 28°30′93′′N). The soil was red soil with a pH value of 6.2, organic matter of 23.37 g⋅kg-1, total nitrogen (N) of 1.41 g⋅kg-1, total phosphorus (P) of 0.52 g⋅kg-1, alkali-hydrolyzed nitrogen of 153.2 mg⋅kg-1, available phosphorus of 36.98 mg⋅kg-1, available potassium of 109.69 mg⋅kg-1.
2.2. Experimental materials
The test variety is Xiangyou 708, provided by the Hunan branch of the National Oil Crops Improvement Center. The test fertilizer is: 45% potassium sulfate compound fertilizer (N: P2O2 : K2O=15:15:15); organic fertilizer (organic matter content ≥30%) ;microbial bacterial fertilizer (multi-resistant functional bacterial fertilizer, effective viable bacteria count ≥200 million/g)
2.3. experiment design
Three fertilization modes: 1. 40kg/mu compound fertilizer (Conventional fertilization); 2. 40kg/mu compound fertilizer + 40kg/mu organic fertilizer (Supplementary organic fertilizer); 3. 40kg/mu compound fertilizer + 40kg/mu microbial bacterial fertilizer (Supplementary microbial fertilizer); all three fertilization modes are applied as base fertilizer before sowing.
Two water treatment methods: 1. Natural precipitation, no irrigation; 2. After two months of normal growth of rapeseed from sowing, start to flood the soil with irrigation water that is greater than the maximum field capacity. The irrigation water should flood the soil surface by 1-2 cm for seven days, and the growth condition should be observed and recorded daily.
The experiment began sowing on September 30, 2022, using a split-plot design with irrigation as the main area and fertilization as the secondary area. The split area was 8m2 (two meters wide and four meters long), with three replicates. The planting density was 450,000 plants per hectare. Field sampling was conducted on November 30, 2022. After the first sampling, the flooding treatment was stopped and the natural precipitation state was restored. The second field sampling was conducted on December 14, 2022.
2.4. Sample collection and determination
2.4.1. Harvesting of rapeseed seedlings:
After the treatment, the plants were taken out of the field, with the root system as intact as possible. The plants were cleaned and the surface moisture was wiped off with paper towels. The plants were divided into above-ground and underground parts. They were then placed in sealed bags and frozen for subsequent testing.
2.4.2. Collection of rapeseed rhizosphere soil
Samples were taken on November 30, 2022 and December 14, 2022, respectively. The roots were dug out and gently shaken to disperse the loose soil attached to the plant roots. Sterile hairbrushes were used to collect soil that was tightly bound to the roots, and the soil was mixed evenly through a 1 mm sieve and then stored in sterile bags for refrigeration. It was sent to Allwegene, a company in Beijing, China, for soil microbial extraction and sequencing.
2.4.3. Determination of plant growth indicators
Use a ruler to measure the stem diameter and plant height of rapeseed seedlings, and use an electronic balance to determine the fresh weight of the aboveground and underground parts of the plant. After measuring, blanch the plant at 105°C for 30 minutes, then dry it at 80°C until constant weight, and weigh its dry weight; use TTC colorimetric method to determine the root vigor of the plant.
2.4.4. Rhizosphere microorganism determination
Use the E.Z.N.A. Soil DNA Kit (Omega Bio-tek, Inc., USA) kit to extract genomic DNA from soil samples. Use Nanodrop 2000 (ThermoFisher Scientific, Inc., USA) to detect DNA quality and concentration. The V3-V4 region of the 16S rRNA gene of bacteria was amplified using universal primers 338F (5’-ACTCCTACGGGAGGCAGCAG-3’) and 806R (5’-GGACTACHVGGGT-WTCTAAT-3’).Add 8bp barcode sequences at the 5' ends of the primers upstream and downstream to distinguish different samples. Finally, universal primers with barcode sequences were synthesized and amplified on an ABI 9700 PCR instrument (Applied Biosystems, Inc., USA).Use 1% agarose gel electrophoresis to detect the size of the amplified target band, 170V for 30 min. The PCR products were automatically purified using the Agency AMPure XP (Beckman Coulter, Inc., USA) nucleic acid purification kit. The concentration of the library was roughly determined using a Nanodrop 2000 (ThermoFisher Scientific, Inc., USA), and the size of the library fragments was detected using an Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., USA).The concentration of the library was accurately quantified using the ABI StepOnePlus Real-Time PCR System (Applied Biosystems, Inc., USA).Finally, the library was sequenced on the Illumina Miseq/ Nextseq 2000/Novaseq 6000 (Illumina, Inc., USA) platform using the PE250/PE300 sequencing strategy.
2.5. Statistical analysis
The physiological experimental data were entered and collated using Microsoft Excel 2013, and the statistical analysis software SPSS (Statistical Product and Service Solutions) was used to perform a significant difference test on the data. The Origin 2021 software was used for plotting.
Use the Pear (v0.9.6) software to filter and splice the sequencing data, remove sequences containing ambiguous base N, remove sequences with quality values below 20, set the minimum overlap to 10bp during splicing, and set the p-value to 0.0001.After splicing, the sequences shorter than 230 bp were removed using Vsearch (v2.7.1) software, and the chimeric sequences were removed using theuchime method based on the Gold Database.Use Vsearch (v2.7.1) software to perform OTU clustering (Operational Taxonomic Units) on high-quality sequences with a sequence similarity threshold of 97%. Use the BLAST algorithm to align the representative sequences of OTU with Silva138 database, with an e-value threshold set to 1e-5, to obtain the corresponding species classification information for each OTU. Based on the OTU and its abundance results.Use QIIME (v1.8.0) software to calculate α diversity index, and use R (v3.6.0) software to draw the plot.Based on the species annotation and relative abundance results, R (v3.6.0) software was used for species composition histogram analysis.Based on the OTU and abundance results, QIIME (v1.8.0) software was used to calculate the beta diversity distance matrix, and R (v3.6.0) software was used for PCoA analysis and plotting based on the distance matrix. Phython (v2.7) software was used for LEfSe analysis.
3. Results
3.1. Effects of different fertilization patterns and water treatments on the growth of rapeseed during the seedling stage
As shown in Table 2, compared with the treatment without flooding for 7 days, the plant height, stem diameter, dry weight, and fresh weight of rapeseed seedlings subjected to flooding treatment were affected to varying degrees. After 7 days of flooding treatment, the stem diameter, aboveground fresh weight, aboveground dry weight, root fresh weight, total fresh weight, and total dry weight of rapeseed seedlings supplemented with organic fertilizer and microbial fertilizer were significantly higher than those of the conventional fertilization group (P<0.05). This indicates that the supplementary application of microbial fertilizer and organic fertilizer can increase the tolerance to waterlogging stress. After 14 days of recovery from flooding stress, the rapeseed seedlings supplemented with organic fertilizer and microbial fertilizer had significantly higher fresh and dry weight of the shoots, fresh weight of roots, total fresh weight, and total dry weight than those of the rapeseed seedlings with conventional fertilization (P<0.05). Under conventional fertilization conditions, compared with the 14-day recovery from flooding treatment, the rapeseed seedlings that were not subjected to flooding treatment and were further cultivated for 14 days showed a decrease in stem diameter, dry weight of shoots, fresh weight of roots, dry weight of roots, and total dry weight of 31.6%, 40.9%, 28.9%, 34.5%, and 40.0%, respectively. This is the largest decrease compared to the other two fertilization modes. This indicates that supplementing microbial fertilizer and organic fertilizer can promote the recovery of rapeseed after suffering from flooding stress.

Table 1.
Experimental design.
| Treatment cord | Fertilization mode | Water treatment | Period |
|---|---|---|---|
| CZ-7 | Conventional fertilization | waterlogging stress | Flooded for 7 days |
| YZ-7 | Supplement organic fertilizer | waterlogging stress | Flooded for 7 days |
| WZ-7 | Supplement microbial fertilizer | waterlogging stress | Flooded for 7 days |
| CW-7 | Conventional fertilization | Normal water supply | Control |
| YW-7 | Supplement organic fertilizer | Normal water supply | Control |
| WW-7 | Supplement microbial fertilizer | Normal water supply | Control |
| CZ-14 | Conventional fertilization | waterlogging stress | End flooding and restore 14 days |
| YZ-14 | Supplement organic fertilizer | waterlogging stress | End flooding and restore 14 days |
| WZ-14 | Supplement microbial fertilizer | waterlogging stress | End flooding and restore 14 days |
| CW-14 | Conventional fertilization | Normal water supply | Control |
| YW-14 | Supplement organic fertilizer | Normal water supply | Control |
| WW-14 | Supplement microbial fertilizer | Normal water supply | Control |
Table 2.
The effect of different fertilization patterns and water treatments on physical indicators of rapeseed seedlings at different stages. Treatments the same as Table 1, Different lowercase letters indicate significant differences at the 0.05 level between different treatments for the same indicator.
Table 2.
The effect of different fertilization patterns and water treatments on physical indicators of rapeseed seedlings at different stages. Treatments the same as Table 1, Different lowercase letters indicate significant differences at the 0.05 level between different treatments for the same indicator.
| Treatment | Plant height | Stem diameter | Aboveground part/g | Root/g | Total weight /g | ||||
|---|---|---|---|---|---|---|---|---|---|
| (cm) | (cm) | Fresh weight | Dry weight | Fresh weight | Dry weight | Total fresh weight | Total dry weight | ||
| YZ-7 | 39.03 g | 0.94 f | 77.89 h | 7.72 h | 7.55 g | 1.61 f | 85.44 h | 9.33 h | |
| CZ-7 | 37.90 g | 0.80 e | 67.08 i | 6.34 i | 5.99 h | 1.22 f | 73.07 i | 7.57 i | |
| WZ-7 | 40.43 fg | 1.09 d | 75.98 h | 7.76 h | 7.59 g | 1.49 f | 83.57 h | 9.25 h | |
| YW-7 | 45.00 de | 1.24 c | 94.80 f | 11.55 g | 10.99 e | 2.30 de | 105.79 f | 13.85 g | |
| CW-7 | 43.33 ef | 1.09 d | 86.37 g | 11.06 g | 9.51 f | 2.09 e | 95.88 g | 13.15 g | |
| WW-7 | 46.50 cde | 1.22 cd | 89.65 fg | 11.12 g | 11.42 e | 2.35 de | 101.07 fg | 13.46 g | |
| YZ-14 | 47.67cd | 1.47 b | 152.40 d | 19.08 d | 14.97 c | 3.33 c | 167.37 d | 22.41 d | |
| CZ-14 | 45.03de | 1.26 c | 137.47 e | 15.13 f | 12.85 d | 2.71 d | 150.32 e | 17.84 f | |
| WZ-14 | 45.77de | 1.40 b | 147.10 d | 17.02 e | 14.98 c | 3.26 c | 162.08 d | 20.27 e | |
| YW-14 | 56.13a | 1.92 a | 196.66 a | 28.37 a | 20.27 a | 4.67 a | 216.93 a | 33.04 a | |
| CW-14 | 50.20bc | 1.85 a | 175.22 c | 25.59 c | 18.08 b | 4.13 b | 193.30 c | 29.72 c | |
| WW-14 | 51.90b | 1.92 a | 188.89 b | 27.09 b | 20.54 a | 4.68 a | 209.43 b | 31.77 b | |
3.2. Effect of different fertilization patterns and water treatments on root vigor of rapeseed during seedling stage
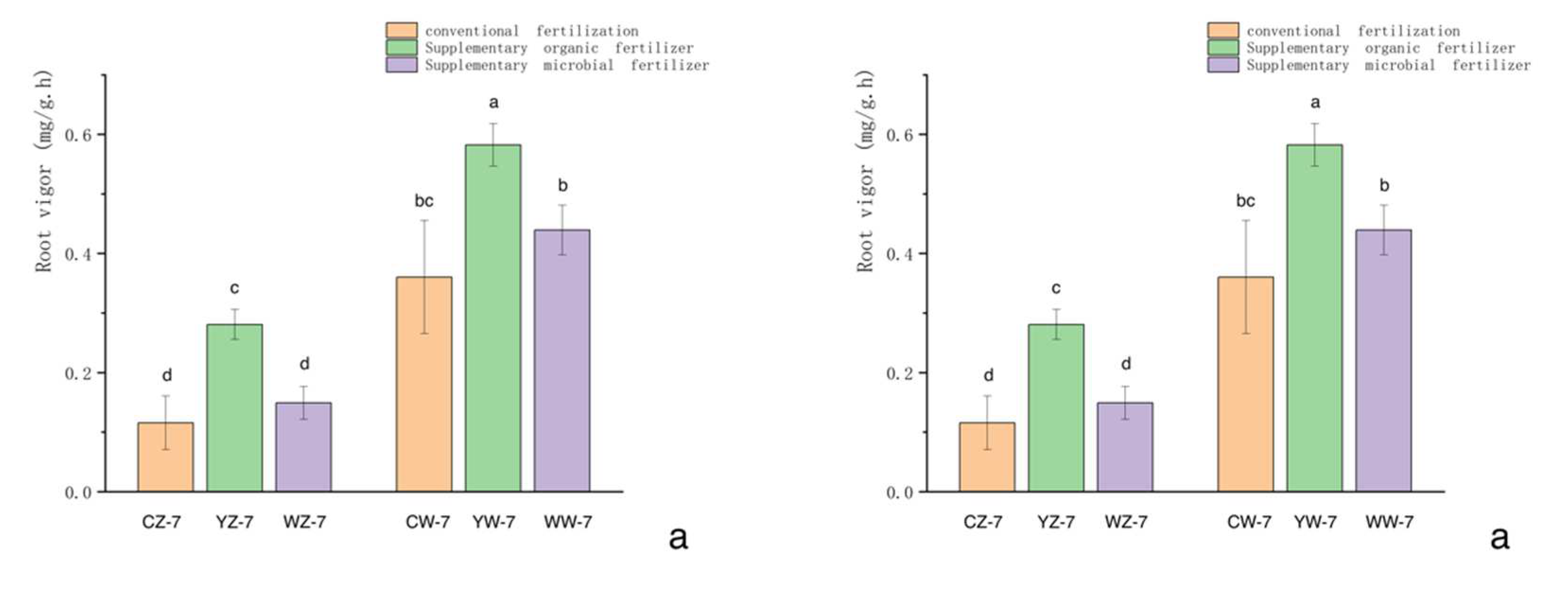
When plants are subjected to waterlogging stress, their root respiration is blocked, and the accumulation of toxic metabolites leads to a decrease in root vigor. As shown in Figure 1-a, compared with no flooding treatment, the root vigor of rapeseed under different fertilization treatments significantly decreased after 7 days of flooding (P<0.05). After 14 days of recovery from flooding, there were significant differences in root vigor between the conventional fertilization and the supplemented microbial fertilizer treatments for rapeseed seedlings (P<0.05), while there were no significant differences in the other three treatments (P>0.05). However, the application of organic fertilizer significantly increased root vigor under all water treatment conditions compared with the other two fertilization modes (P<0.05).
As shown in Figure 1-b, At the end of the flooding treatment, After 14 days of recovery, The rapeseed seedlings that had been subjected to flooding treatment began to resume normal growth, The root vitality also increased. After 14 days of recovery from flooding treatment, The root activity of rapeseed supplemented with microbial fertilizer, conventional fertilizer, and organic fertilizer increased by 83.7%, 49.3%, and 81.2%, respectively, compared to the previous conditions. This suggests that supplemental organic fertilizer and microbial fertilizer are more helpful in restoring the root vigor of rapeseed after being subjected to flooding stress.
Figure 1.
Effect of different fertilization patterns and different water treatments on the root vigor of rapeseed seedlings. Treatments the same as Table 1, Different lowercase letters indicate significant differences at the 0.05 level between different treatments.
Figure 1.
Effect of different fertilization patterns and different water treatments on the root vigor of rapeseed seedlings. Treatments the same as Table 1, Different lowercase letters indicate significant differences at the 0.05 level between different treatments.
3.3. Analysis of high-throughput sequencing results and relative abundance and diversity of rhizosphere bacteria
To investigate the differences in rhizosphere bacteria in rape at different stages under different fertilization and water treatment modes, The V3-V4 region of bacterial 16S rRNA was sequenced from rhizosphere soil samples of 12 treatment groups in Table 1,A total of 15,240 OTUs were generated through clustering, and 13,975 were retained after normalization (OTU clustering similarity level ≥97%).A total of 7287 OTUs were found in each treatment group supplemented with organic fertilizer, of which 2216 OTUs were common among the four groups (Figure 2-a);A total of 8017 OTUs were found in the various treatment groups of conventional fertilization, of which 4076 OTUs were common to all four groups (Figure 2-b);A total of 6,488 OTUs were found in the various treatment groups supplemented with microbial fertilizer, of which 2,347 OTUs were common to all four groups (Figure 2-c);The chao1 and shannon indices of rhizosphere bacteria in different treatment groups (Figure 3-a, b) showed that there were no significant differences in the richness of rhizosphere microorganisms in rapeseed at different stages between flooding treatment, fertilization mode, and control treatment (P>0.05).
3.4. Analysis of rhizosphere bacteria Bate in rapeseed seedling stage under different fertilization modes and water treatments
To verify the differences and similarities in fungal communities between different samples, the ANOSIM showed that there were significant differences (r=0.40, P<0.05) in the rhizosphere bacterial community structure among the 12 treatment groups. Beta analysis was conducted on 36 groups of samples using PLS-DA, The results showed that under normal water supply, rapeseed seedlings supplemented with microbial fertilizer. After 14 days of continuous cultivation, the bacterial community in the rhizosphere soil did not change significantly. On the second principal component PC2 (with a differential explanatory rate of 10.73%), compared with no flooding treatment, flooding for 7 days caused the greatest difference in rhizosphere bacterial communities in rapeseed supplemented with microbial fertilizer, which was similar to the results of the other two fertilization methods (Figure 4-a, b, c).Under the conventional fertilization model, the first principal component PC2 (with a difference interpretation rate of 10.95%)After 14 days of continuous cultivation without flooding treatment, the rhizosphere soil community of rapeseed showed the greatest difference, which was similar to the difference shown by the addition of organic fertilizer (Figure 4-b, c).
3.5. Analysis of rhizosphere soil community composition of rapeseed seedlings under different fertilization patterns and water treatments
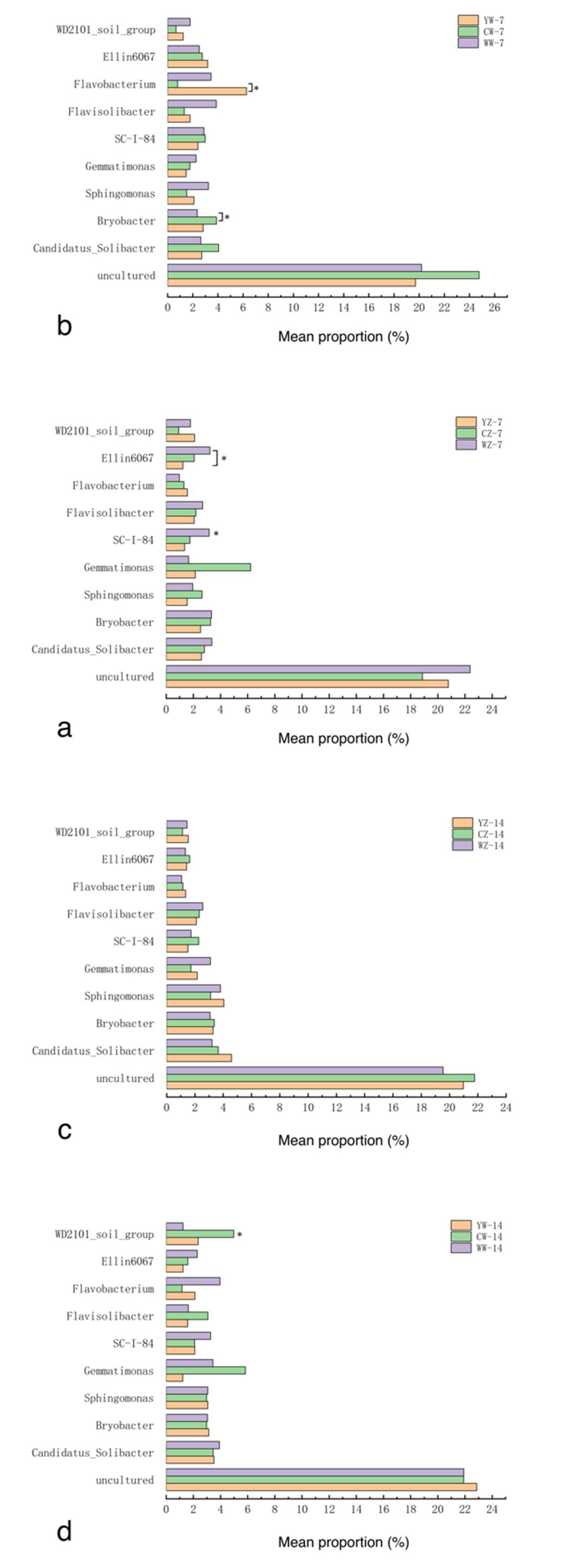
Samples were taken from the rhizosphere soil of 12 groups of rapeseed treatments (Table 1). Three replicates were set up for each treatment group, resulting in a total of 36 soil samples.After sequencing these samples, a total of 53 phyla were detected in the 12 treatment groups, of which Proteobacteria, Acidobacteriota, Bacteroidota, Gemmatim-onadota, and Chloroflexi were the dominant phyla. Their relative abundances are 30.6% ~ 38.8%, 14.0% ~ 20.4%, 10.4% ~ 20.7%, 2.8% ~ 10.8%, and 2.4% ~ 8.6% (Figure 5). The results of the differential analysis are shown in Figure 4-a-d. After 7 days of flooding treatment, compared with conventional fertilization, the relative abundance of Verrucomicrobiota in the rapeseed rhizosphere soil supplemented with organic fertilizer was higher (P<0.05). Under normal water supply, compared with conventional fertilization, the relative abundance of Bacteroidota in the rapeseed rhizosphere soil supplemented with organic fertilizer and microbial fertilizer was higher (P<0.05). After 14 days of recovery from flooding treatment, compared with conventional fertilization, the relative abundance of Actinobacteriota in the rapeseed rhizosphere soil supplemented with microbial fertilizer was higher (P<0.05). After 14 days of continuous cultivation without flooding treatment, compared with conventional fertilization, the relative abundance of Chloroflexi and Actinobacteriota in the rapeseed rhizosphere soil supplemented with organic fertilizer was higher (P<0.05).
At the genus level, the top 10 dominant bacterial genera with relative abundance were: Flavobacterium, Gemmatimonas, uncultured, Bryobacter, Sphingomonas, Flavisolibacter, SC-I-84, WD2101_soil_group, Ellin6067, and Candidatus_Solibacter. After 7 days of flooding treatment, Compared with the supplementary application of organic fertilizer, The relative abundance of Ellin6067 was higher in the rapeseed rhizosphere soil supplemented with microbial fertilizer (P<0.05), compared with conventional fertilization. The relative abundance of SC-I-84 was higher in the rapeseed rhizosphere soil supplemented with microbial fertilizer (P<0.05).Under normal water supply, Compared with conventional fertilization, The relative abundance of Flavobacterium was higher in the rapeseed rhizosphere soil supplemented with organic fertilizer and microbial bacterial fertilizer (P<0.05);After 14 days of continuous cultivation without flooding, Compared with the supplementary application of organic fertilizer and microbial fertilizer, The relative abundance of WD2101_soil_group was higher in the conventionally fertilized rapeseed rhizosphere soil (P<0.05) (Figure 5, a-d).
Figure 7.
Changes in relative abundance of bacterial genera in the rhizosphere of rapeseed in different treatment groups, with treatments as in Table 1; *: P<0.05.
Figure 7.
Changes in relative abundance of bacterial genera in the rhizosphere of rapeseed in different treatment groups, with treatments as in Table 1; *: P<0.05.
Based on the abundance distribution of taxa or the similarity between samples, clustering analysis was conducted on the top 20 genera in different samples. Compared with normal water supply, the relative abundance of Anaeromyxobacter, Citrifermentans, and Flavisolibacter was higher in flooded rapeseed rhizosphere soil, while the relative abundance of Edaphobaculum, Subgroup_7, SC−I−84, and Flavobacterium was lower (Figure 8).
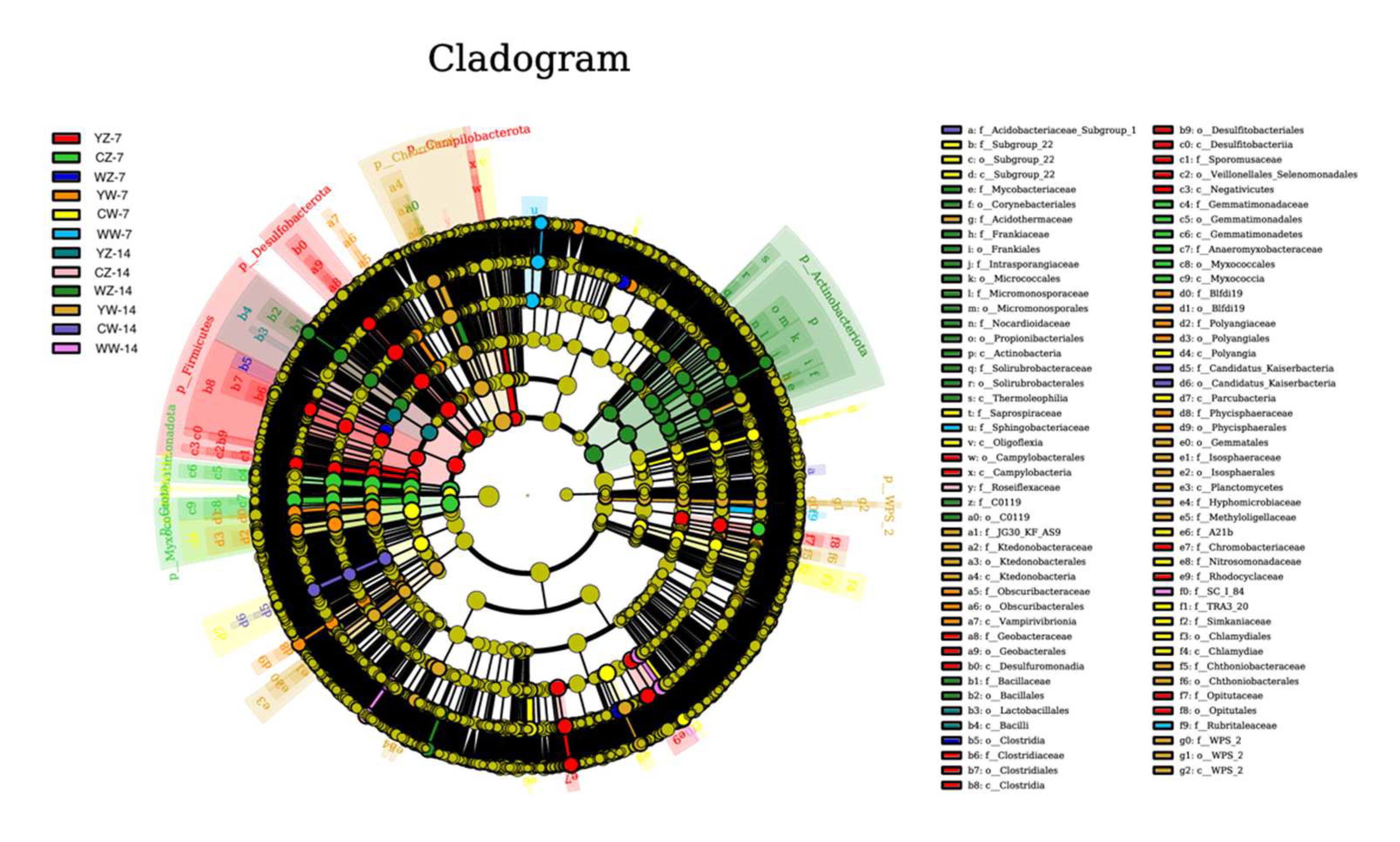
Besides, The threshold value of the screening criteria was set to 3 using LEfSe, To find biomarkers with statistical differences between different samples, Examine the contribution relationship between different treatments. As shown in Figure 9, in the case of, A total of 89 differentially expressed biomarker groups were found in 12 treatment groups. Among them, 44 differential groups were found in the rhizosphere soil supplemented with organic fertilizer, However, the number of differentially expressed taxa found in the supplementary application of microbial fertilizer and conventional fertilization was relatively small. The LEfSe analysis results showed that Fertilization patterns can change the bacterial community structure of soil, The different groups found in the supplementary application of organic fertilizer and microbial fertilizer may have a positive effect on improving the waterlogging tolerance of rapeseed and promoting its growth during the seedling stage. However, further research is still needed to understand how these dominant bacteria respond to waterlogging in rapeseed.
4. Discussion
Winter rapeseed grown in southern China often suffers from waterlogging damage. When rapeseed is affected by waterlogging, the roots are the first to be affected. The aerobic respiration of the rapeseed roots under waterlogging stress is weakened, nutrient transport is blocked, and toxic substances accumulate, which affects the normal growth of rapeseed during the seedling stage. This experiment obtained similar results, that is, waterlogging has seriously affected the growth of rapeseed, and plant height, stem diameter, plant dry weight, and plant fresh weight decreased (Table 2). The plant root activity decreased and it was difficult to recover to normal level in a short period of time (Figure 1). These similar phenomena have also appeared in previous reports. [16,17,18].
An important role of organic fertilizers and microbial fertilizers in the application process is to improve soil structure and increase nutrients in the soil [12,15,19].The experimental results showed that under normal water conditions, there was no significant difference in dry matter accumulation between rapeseed seedlings supplemented with organic fertilizer and microbial fertilizer (Table 2);When rapeseed seedlings were subjected to waterlogging stress, Compared with the rapeseed seedlings treated with conventional fertilization, the seedlings treated with additional microbial fertilizer and organic fertilizer showed significant differences in growth, more dry matter accumulation. Although there are few reports on the ability of microbial fertilizers and organic fertilizers to promote rape resistance to waterlogging, Zhang and Zhao believe that microbial fertilizers and organic fertilizers do enhance the ability of crops to resist stress and disease [17,20]. This may be related to the increase of nutrients in the soil, the change of soil structure, and the impact on the community relationship of soil microorganisms. Another experimental result showed that, After 7 days of flooding stress, Compared with conventional fertilization, The root system is more vigorous, And during the later recovery process, Supplementing organic fertilizer and microbial fertilizer can restore rapeseed root vigor faster. This may promote the metabolism of the root of rapeseed after suffering from waterlogging, thereby better allowing the rapeseed seedlings to accumulate dry matter. It is worth noting that, The effect of supplementing organic fertilizer on improving the root activity of rapeseed under waterlogging stress is better than that of supplementing microbial fertilizer.
As the most dominant microbial group in soil, bacteria account for about 80% of the total microbial biomass in soil. They can effectively promote the decomposition of organic matter and the release of nutrients in the soil, Among them, bacteria in the rhizosphere soil are of great significance for plant growth [21,22]. Changes in soil moisture conditions can affect the changes in rhizosphere microbial communities. According to existing reports, microbial fertilizers and organic fertilizers can affect the community changes of rhizosphere microorganisms [13,15].This study aims to reveal the effects of waterlogging stress and fertilization patterns on the rhizosphere bacteria of rapeseed.
Based on OUT clustering analysis and Alpha analysis, Among the 12 groups of treated rhizosphere soils, There were no significant differences in bacterial relative abundance and diversity. Prior to this, the results of Azarbad et al. [23] showed that waterlogging reduced the abundance of bacteria in the rhizosphere of wheat. However, other studies have also found that waterlogging increases the number of bacteria in the rhizosphere of maize [24]. Obviously, the results of the two are contradictory, which may be related to the type of crops.
ANOSIM and PLS-DA showed that, The flooding treatment and fertilization mode significantly changed the bacterial community structure in the rhizosphere of rapeseed. According to previous studies, When waterlogging occurs, due to differences in the intensity of root respiration, environmentally sensitive microorganisms undergo changes [23,25,26,27].In addition, After 7 days of flooding treatment, The differences in the bacterial community structure of the root system between the supplementary application of microbial fertilizer and organic fertilizer were relatively small. The rhizosphere bacterial community structure of the two treatments and the conventional fertilization treatment showed significant differences, This is similar to the differences in the growth of rapeseed seedlings under different fertilization patterns.
Based on the gate level analysis, The dominant bacterial phyla in all soil samples were Proteobacteria, Acidobacteriota, Bacteroidota, Gemmatimonadota, and Chloroflexi. According to previous studies, Proteobacteria, Acidobacteriota, Gemmatimonadota, and Bacteroidota are the dominant bacterial phyla in the rhizosphere of crops.This study is consistent with previous research [28,29,30,31].The Proteobacteria phylum is the most abundant bacterial group in rhizospheric soil, mainly due to its ability to utilize a variety of plant root exudates and its rapid growth rate [32].A large number of studies have shown that Acidobacteriota mainly degrades plant residue polymers [33] and participates in iron cycling [34,35].In addition, Acidobacteriota is a stable component of the bacterial ecosystem and has good adaptability to the environment [36,37]. The Bacteroidota is now believed to be significantly associated with the oxidation and reduction of nitrates [38,39].Under normal water supply,Compared with conventional fertilization, the relative abundance of Bacteroidetes in the rhizosphere soil was higher under the application of organic fertilizer and microbial fertilizer.This may be related to the promotion of seedling growth by applying organic fertilizer and topdressing with microbial fertilizer. Gemmatimonadota can generally convert various sugar molecules into vitamins [40].Relevant studies have shown that appropriately increasing the vitamin content in the roots can promote the growth of wheat [41].After 7 days of flooding treatment, compared to the supplement of microbial fertilizer, the conventional fertilization method resulted in higher Gemmatimonadota levels in the rhizosphere soil.After 14 days without flooding treatment, the conventional fertilization method had higher Gemmatimonadota levels in the rhizosphere soil compared to the supplementary organic fertilization method.This may be related to the secretion of more sugar substances by the root system under conventional fertilization.Zhao et al. found that Chloroflexi prefer to live in nutrient-rich environments, and adequate nutrients are more conducive to the growth and reproduction of Chloroflexi [42].After 14 days without flooding treatment,Compared with conventional fertilization and supplementary microbial fertilizer,The Chloroflexi were higher in the rhizosphere soil,This indicates that the application of organic fertilizer increases the nutrients in the soil.
In addition, the experimental results also showed that during the recovery period after the end of flooding, the supplementary application of microbial fertilizer and organic fertilizer could better enrich Actinobacteriota.According to previous reports,Actino-bacteriota can decompose cellulose and lignin, and abundant actinomycetes are beneficial to the decomposition of plant residues in soil [43].The supplementary application of microbial fertilizer and organic fertilizer during the experiment can help rapeseed recover faster from flooding stress, which may be related to the enrichment of Actinobacteriota in the rhizosphere soil of rapeseed by the supplementary application of microbial fertilizer and organic fertilizer.Orellana [44] believed that the Verrucomicrobiota was related to the degradation of polysaccharides.Through the degradation of polysaccharides, the polysaccharides in the soil are converted into small molecule sugars that can be absorbed by plant roots. Our results also showed that,after 7 days of flooding treatment, the supplementary application of organic fertilizer enriched the Verrucomicrobiota, which may be related to promoting the growth of rapeseed under flooding stress.
At the genus level,Differential analysis was conducted on the top 10 bacterial groups with relatively high richness.The four genera of Flavisolibacter, SC-I-84, Ellin6067, and WD2101_soil_group showed significant differences in different water treatments and different fertilization patterns. According to Yoon's[45] research, Flavisolibacter can increase carbon-related functional groups, further increasing carbon sequestration, which may be related to promoting plant growth. In the control group treated with flooding for 7 days,Compared to conventional fertilization,The Flavisolibacter content in the rapeseed rhizosphere soil supplemented with organic fertilizer and microbial fertilizer was higher,This may be one of the reasons why the application of organic fertilizer and microbial fertilizer can promote the growth of rapeseed seedlings.Ellin6067 is an ammonia-oxidizing bacterium that can oxidize ammonia to nitrite [46].This indicates that Ellin6067 can promote the absorption of nitrogen by plants.In the control group that was treated with flooding for 7 days,Compared to the application of organic fertilizer,The addition of microbial fertilizer to rapeseed rhizosphere soil resulted in higher Ellin6067 levels,This may be the reason why the application of microbial fertilizer can promote the growth of rapeseed when suffering from waterlogging.Existing research has shown that WD2101_soil_group can participate in the decomposition of polysaccharides [47].The experimental results showed that under conventional fertilization conditions, after the rape was subjected to flooding treatment, the relative abundance of WD2101_soil_group was low.After 14 days of recovery, the relative abundance of WD2101_soil_group increased significantly.This may be related to the normal growth of rapeseed after the flooding is lifted.SC-I-84 is known as anaerobic ammonia oxidation bacteria [48].This is also beneficial for the absorption of nitrogen by plant roots,After 7 days of flooding treatment, the microbial fertilizer enriched SC-I-84.This may also be related to the promotion of rapeseed growth.
5. Conclusions
To sum up, After being subjected to flooding stress, rapeseed will affect the seedling height, stem diameter, dry matter accumulation, root activity, and other aspects. Supplementing microbial fertilizer and organic fertilizer can make rapeseed grow better under normal water supply, reduce the impact of flooding stress on rapeseed seedlings, and allow rapeseed to recover faster after suffering from flooding stress during the seedling stage. Through the Beta analysis of bacterial communities in the rhizosphere of rapeseed, it was found that different water treatment and fertilization patterns can change the community structure of bacteria in the rhizosphere soil. Through the analysis of the differences between different treatment groups at the level of genus and family, It was found that under normal water supply, the application of organic fertilizer enriched the rhizosphere soil of rapeseed with P_Bacteroidota, P_Actinobacteriota, P_Chloroflexi, and G_Flavisolibacter, and the application of microbial fertilizer enriched the rhizosphere soil of rapeseed with P_Bacteroidota and G_Flavisolibacter. After flooding treatment, the application of organic fertilizer enriched the rhizosphere soil of rapeseed with P_Verrucomicrobiota, and the application of microbial fertilizer enriched the rhizosphere soil of rapeseed with P_Actinobacteriota, G_SC-I-84, and G_Ellin6067. The enrichment of these bacterial groups may be related to the growth promotion and waterlogging tolerance of rapeseed, but the specific functions of these bacterial groups remain to be further verified.
- Author Contributions: Conceptualization, C.G. and M.G.; methodology, X.W. and B.H.; software, J.W. and D.Z. and B.Z.; investigation, X.W.; writing—original draft preparation, X.W. ; writing—review and editing, M.G.; visualization, X.W.; supervision, C.G. All authors have read and agreed to the published version of the manuscript.
- Funding: This research was funded by the National Rapeseed Industrial Technology System (CARS- 12) and the Hunan Agriculture Research System of DARA (Xiangnongfa (2022)).
- Data Availability Statement: Not applicable.
- Conflicts of Interest: The authors declare no conflict of interest.
References
- Li, C-J. A statistical analysis of the storm flood disasters in China. J Catastrophol, (in Chinese with English abstract). 1996, 11, 59–63. [Google Scholar]
- Liu H-L. Practical Cultivation of Rapeseed. S: Shanghai, 1987.
- 3. Oil Crops Research Institute Chinese Academy of Agricultural Sciences. Rapeseed Cultivation in China. Beijing: Agriculture Press, 1990. (in Chinese)
- Hu Q, Hua W, Yin Y; et al. Rapeseed research and production in China. The Crop Journal 2017, 5, 127–135. [Google Scholar] [CrossRef]
- Barnawal D, Bharti N, Maji D; et al. 1-Aminocyclopropane-1-carboxylic acid (ACC) deaminase-containing rhizobacteria protect Ocimum sanctum plants during waterlogging stress via reduced ethylene generation. Plant Physiology and Biochemistry 2012, 58, 227–235. [Google Scholar] [CrossRef]
- Grichko V P, Glick B R. Flooding tolerance of transgenic tomato plants expressing the bacterial enzyme ACC deaminase controlledby the 35S, rolD or PRB-1b promoter[J]. Plant Physiology and Biochemistry 2001, 39, 19–25. [Google Scholar] [CrossRef]
- Canarini A, Dijkstra F A. Dry-rewetting cycles regulate wheat carbon rhizodeposition, stabilization and nitrogen cycling. Soil Biology and Biochemistry 2015, 81, 195–203. [Google Scholar] [CrossRef]
- 8. Castrillo G, Teixeira P J P L, Paredes S H, et al. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [CrossRef]
- Grayston S J, Wang S, Campbell C D; et al. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biology and Biochemistry 1998, 30, 369–378. [Google Scholar] [CrossRef]
- Tang Z C, Kozlowski T T. Water relations, ethylene production, and morphological adaptation of Fraxinus pennsylvanica seedlings to flooding. Plant and Soil 1984, 77, 183–192. [Google Scholar] [CrossRef]
- Nabti E, Jha B, Hartmann A. Impact of seaweeds on agricultural crop production as biofertilizer[J]. International Journal of Environmental Science and Technology 2017, 14, 1119–1134. [Google Scholar] [CrossRef]
- NING C, GAO P, WANG B; et al. Impacts of chemical fertilizer reduction and organic amendments supplementation on soil nutrient, enzyme activity and heavy metal content[J]. Journal of Integrative Agriculture 2017, 16, 1819–1831. [Google Scholar] [CrossRef]
- Meng X, Liu B, C; et al. Effect of pig manure on the chemical composition and microbial diversity during co-composting with spent mushroom substrate and rice husks. Bioresource technology 2018, 251, 22–30. [Google Scholar] [CrossRef]
- Kaje V V, Sharma D K, Shivay Y S; et al. Long-term impact of organic and conventional farming on soil physical properties under rice (Oryza sativa)-wheat (Triticum aestivum) crop** system in north-western Indo-Gangetic plains. Indian J. Agric. Sci. 2018, 88, 107–113. [Google Scholar] [CrossRef]
- Zheng M Y, Li H M, Zhao J S; et al. Current situation and developmental trend of microbial fertilizer researches. Acta Agriculturae Jiangxi 2018, 30, 52–56. [Google Scholar]
- Hong-peng Z, Pan-pan Z, Dong L I B L I; et al. Effects of uniconazole on alleviation of waterlogging stress in soybean. CHINESE JOURNAL OF OIL CROP SCIENCES, 2017, 39, 655. [Google Scholar]
- Zhang, Zhi-Yong, et al.; et al. Effects of irrigation at flowering stage on soil nutrient and root distribution in wheat field. The Journal of Applied Ecology 2022, 12, 3328–3336. [Google Scholar]
- SHANGGUAN, *!!! REPLACE !!!*. SHANGGUAN. "Effect of nitrogen on root vigor and growth in different genotypes of wheat under drought stress. 37.6. Journal of Triticeae Crops 2017, 820–827. [Google Scholar]
- Lu W T, Jia Z K, Zhang P; et al. Effects of organic fertilization on winter wheat photosynthetic characteristics and water use efficiency in semi-arid areas of southern Ningxia. Plant Nutrition and Fertilizer Science 2011, 17, 1066–1074. [Google Scholar]
- Zhao J, Ni T, Li J; et al. Effects of organic–inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice—wheat crop** system. Applied soil ecology 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Shang Q, Yang G, Wang Y; et al. Illumina-based analysis of the rhizosphere microbial communities associated with healthy and wilted Lanzhou lily (Lilium davidii var. unicolor) plants grown in the field. World Journal of Microbiology and Biotechnology 2016, 32, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fan M, Li J, Yan W; et al. Shifts in the structure and function of wheat root-associated bacterial communities in response to long-term nitrogen addition in an agricultural ecosystem. Applied Soil Ecology 2021, 159, 103852. [Google Scholar] [CrossRef]
- Azarbad H, Constant P, Giard-Laliberté C; et al. Water stress history and wheat genotype modulate rhizosphere microbial response to drought. Soil Biology and Biochemistry 2018, 126, 228–236. [Google Scholar] [CrossRef]
- Guo T Z, Yuan L Z, Zhao Y Q. Liu J Y,Gu C. Effects of waterlogging on maize yield and the rhizosphere soil microorganism. Hubei Agric Sci 2014, 53, 505–507, (in Chinese with English abstract). [Google Scholar]
- Evans S E, Wallenstein M D. Soil microbial community response to drying and rewetting stress: does historical precipitation regime matter ? Biogeochemistry 2012, 109, 101–116. [Google Scholar] [CrossRef]
- Unger I M, Kennedy A C, Muzika R M. Flooding effects on soil microbial communities. Applied Soil Ecology 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Meisner A, Leizeaga A, Rousk J; et al. Partial drying accelerates bacterial growth recovery to rewetting. Soil Biology and Biochemistry 2017, 112, 269–276. [Google Scholar] [CrossRef]
- HU R, ZHENG L, LIU H. Effects of straw returning on microbial diversity in rice rhizosphere and occurrence of rice sheath blight. Acta Phytophylacica Sinica 2020, 47, 1261. [Google Scholar]
- Zhang J, Ke W J, Liu J; et al. Influence of water controlling depth on soil microflora and bacterial community diversity in paddy soil. Zhongguo Shengtai Nongye Xuebao/Chinese Journal of Eco-Agriculture 2019, 27, 277–285. [Google Scholar]
- Fierer N, Bradford M A, Jackson R B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Wang G P, Ayiguli T H T, Wang R. Study on the diversity and community structure of salt tolerant bacteria in saline alkali soil in Wuerhe. Xinjiang J Microbiol 2021, 41, 17–26, (in Chinese with English abstract). [Google Scholar]
- AI C, SUN J, WANG X; et al. Advances in the study of the relationship between plant rhizodeposition and soil microorganism. Journal of Plant Nutrition and Fertilizers 2015, 21, 1343–1351. [Google Scholar]
- Pankratov T A, Ivanova A O, Dedysh S N; et al. Bacterial populations and environmental factors controlling cellulose degradation in an acidic Sphagnum peat. Environmental Microbiology 2011, 13, 1800–1814. [Google Scholar] [CrossRef]
- Lu S, Gischkat S, Reiche M; et al. Ecophysiology of Fe-cycling bacteria in acidic sediments. Applied and environmental microbiology, 2010, 76, 8174–8183. [Google Scholar] [CrossRef] [PubMed]
- Coates J D, Ellis D J, Gaw C V; et al. Geothrix fermentans gen. nov., sp. nov., a novel Fe (III)-reducing bacterium from a hydrocarbon-contaminated aquifer. International journal of systematic and evolutionary microbiology 1999, 49, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Kielak A, Pijl A S, Van Veen J A; et al. Differences in vegetation composition and plant species identity lead to only minor changes in soil-borne microbial communities in a former arable field. FEMS microbiology ecology 2008, 63, 372–382. [Google Scholar] [CrossRef]
- Zhang Y, Cong J, Lu H; et al. Community structure and elevational diversity patterns of soil Acidobacteria. Journal of Environmental Sciences 2014, 26, 1717–1724. [Google Scholar] [CrossRef]
- García-López M, Meier-Kolthoff J P, Tindall B J; et al. Analysis of 1,000 type-strain genomes improves taxonomic classification of Bacteroidetes. Frontiers in microbiology 2019, 10, 2083. [Google Scholar]
- Feng D, Wu Z, Xu S. Nitrification of human urine for its stabilization and nutrient recycling. Bioresource technology 2008, 99, 6299–6304. [Google Scholar] [CrossRef]
- Jia-ying X U, **-rong Z, Jie W U. Medium-and long-term effects of the veterinary antibiotic sulfadiazine on soil microorganisms in a rice field. Journal of Agro-Environment Science 2020, 39, 1757–1766. [Google Scholar]
- Wang K, Yang Y A, Yuan K N. Effects of three vitamins on the growth and physiological properties in wheat. Bull Sci Technol 1995, 11, 301–305, (in Chinese with English abstract). [Google Scholar]
- Zhao L J, Liu Y G, Wang Y; et al. Bacterial community structure and diversity of sediments in a typical plateau lakeshore. Microbiol. China 2020, 47, 401–410. [Google Scholar]
- Kanokratana P, Uengwetwanit T, Rattanachomsri U; et al. Insights into the phylogeny and metabolic potential of a primary tropical peat swamp forest microbial community by metagenomic analysis. Microbial ecology 2011, 61, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Orellana L H, Francis T B, Ferraro M; et al. Verrucomicrobiota are specialist consumers of sulfated methyl pentoses during diatom blooms. The ISME Journal 2022, 16, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Yoon M H, Im W T. Flavisolibacter ginsengiterrae gen. nov., sp. nov. and Flavisolibacter ginsengisoli sp. nov., isolated from ginseng cultivating soil. International journal of systematic and evolutionary microbiology 2007, 57, 1834–1839. [Google Scholar] [CrossRef] [PubMed]
- Wei W, Guan D, Ma M; et al. Long-term fertilization coupled with rhizobium inoculation promotes soybean yield and alters soil bacterial community composition. Frontiers in Microbiology 2023, 14, 1161983. [Google Scholar] [CrossRef]
- Kovaleva O L, Merkel A Y, Novikov A A; et al. Tepidisphaera mucosa gen. nov., sp. nov., a moderately thermophilic member of the class Phycisphaerae in the phylum Planctomycetes, and proposal of a new family, Tepidisphaeraceae fam. nov., and a new order, Tepidisphaerales ord. nov. International journal of systematic and evolutionary microbiology 2015, 65(Pt. 2), 549–555. [Google Scholar] [CrossRef]
- J Jiang Z, Huang X, Wang S; et al. Divalent manganese stimulates the removal of nitrate by anaerobic sludge. RSC advances 2024, 14, 2447–2452. [Google Scholar] [CrossRef]
Figure 2.
Venn diagram of the number of OTUs in different fertilization patterns; Treatments the same as Table 1.
Figure 2.
Venn diagram of the number of OTUs in different fertilization patterns; Treatments the same as Table 1.
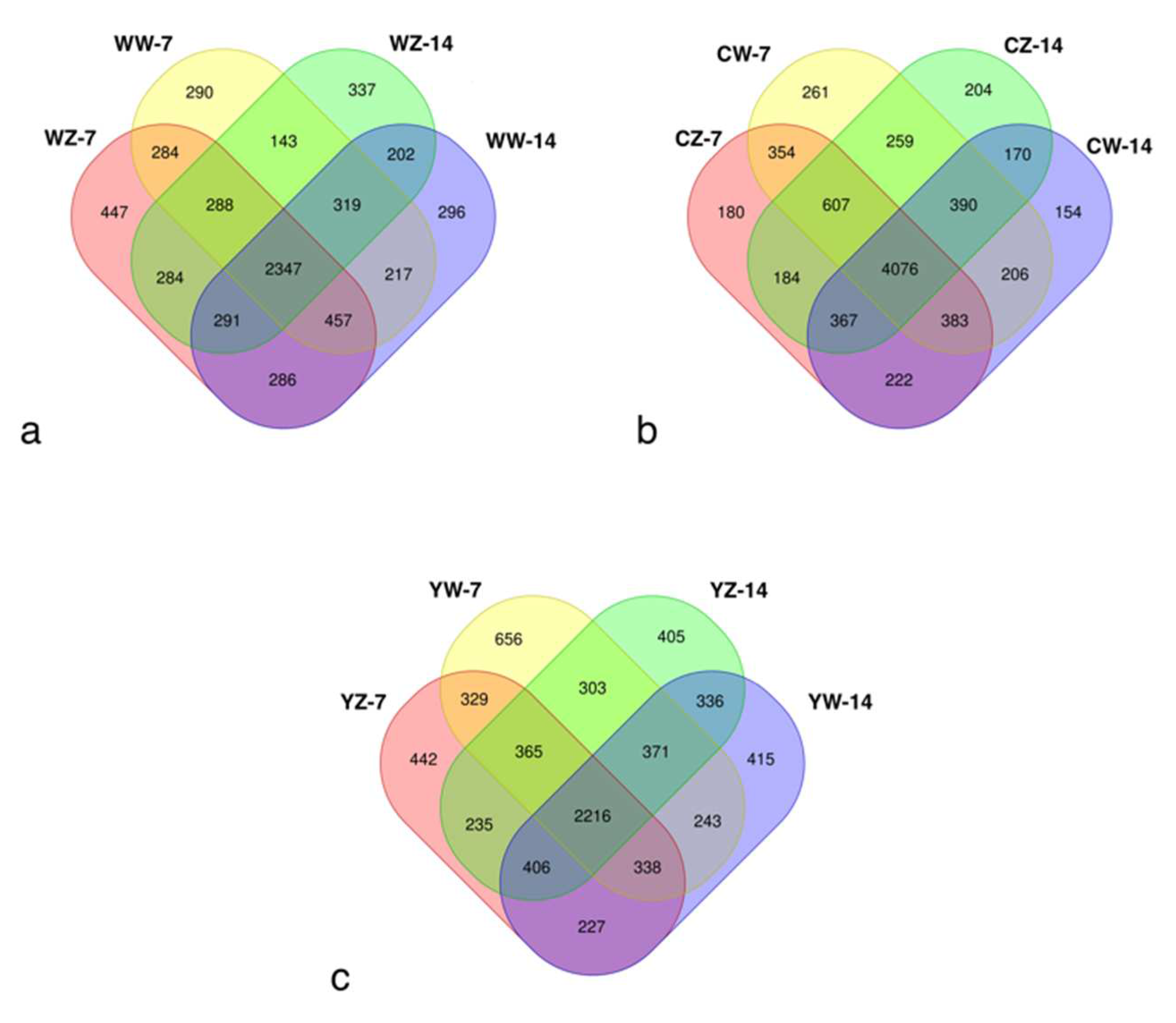
Figure 3.
Effect of fertilization pattern and water treatment on Chao1 abundance (a) and Shannon (b); Different letters indicate significant differences between the two treatments, and the treatments are the same as Table 1.
Figure 3.
Effect of fertilization pattern and water treatment on Chao1 abundance (a) and Shannon (b); Different letters indicate significant differences between the two treatments, and the treatments are the same as Table 1.
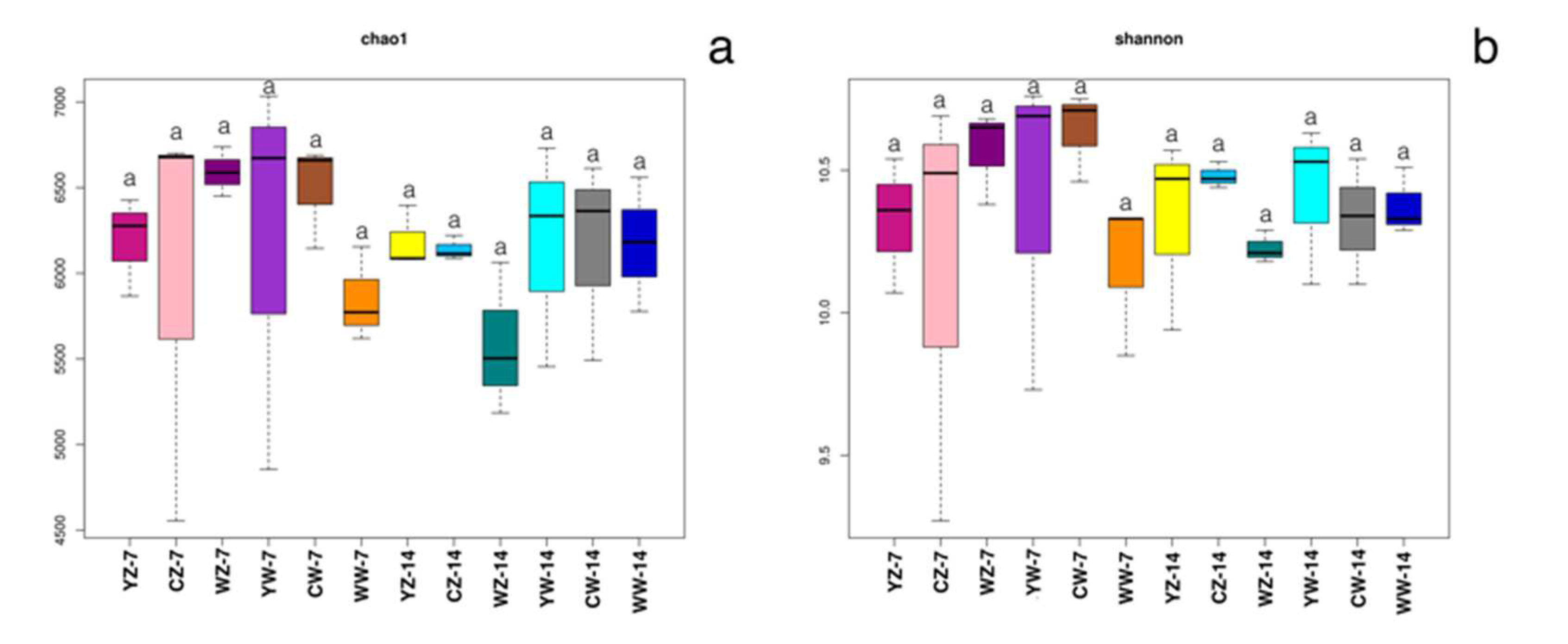
Figure 4.
PLS-DA analysis of bacterial communities in rhizosphere soils under different treatments; Treatments the same as Table 1.
Figure 4.
PLS-DA analysis of bacterial communities in rhizosphere soils under different treatments; Treatments the same as Table 1.
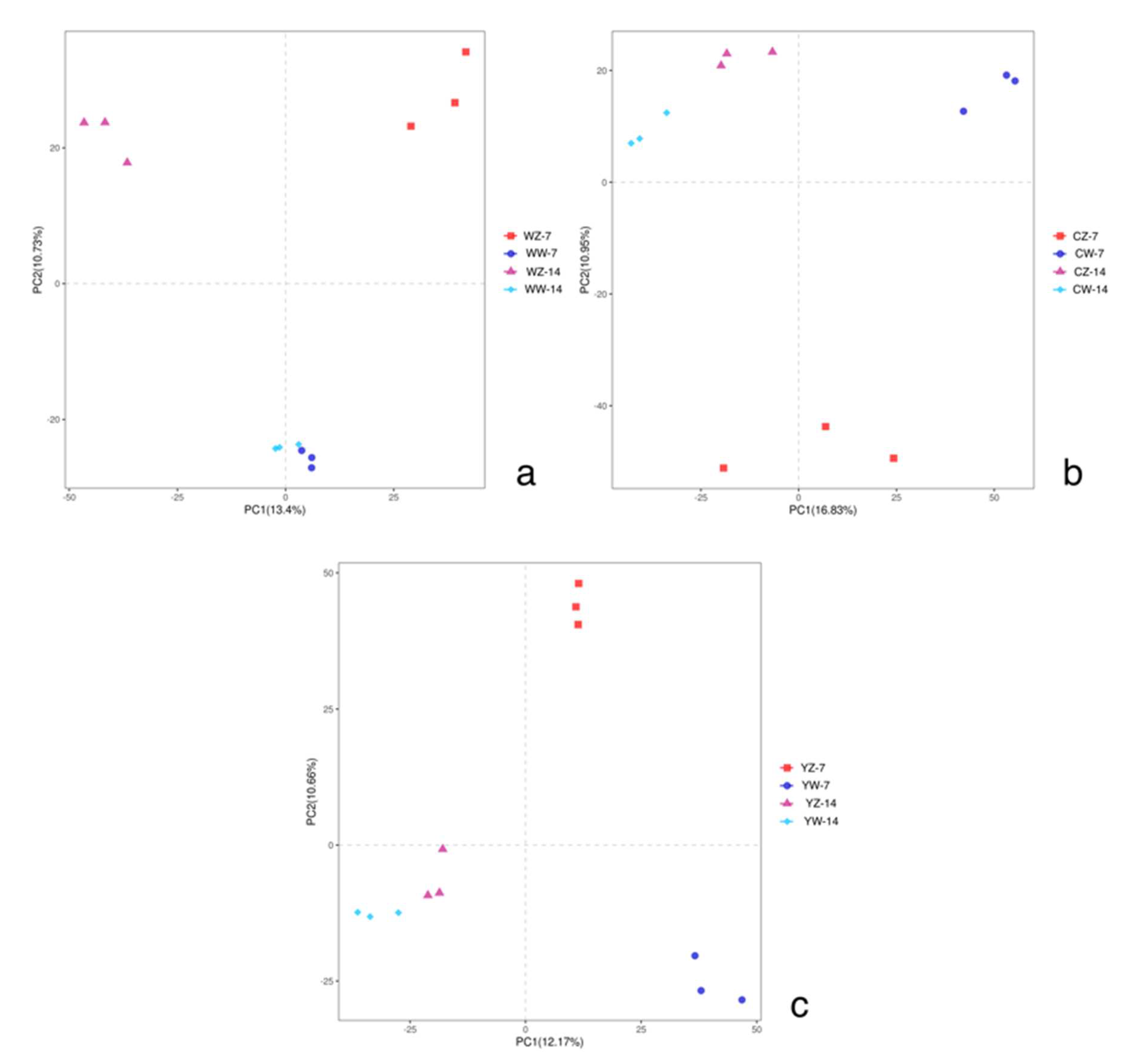
Figure 5.
Relative richness of doors under different treatments; Treatments the same as Table 1.
Figure 5.
Relative richness of doors under different treatments; Treatments the same as Table 1.
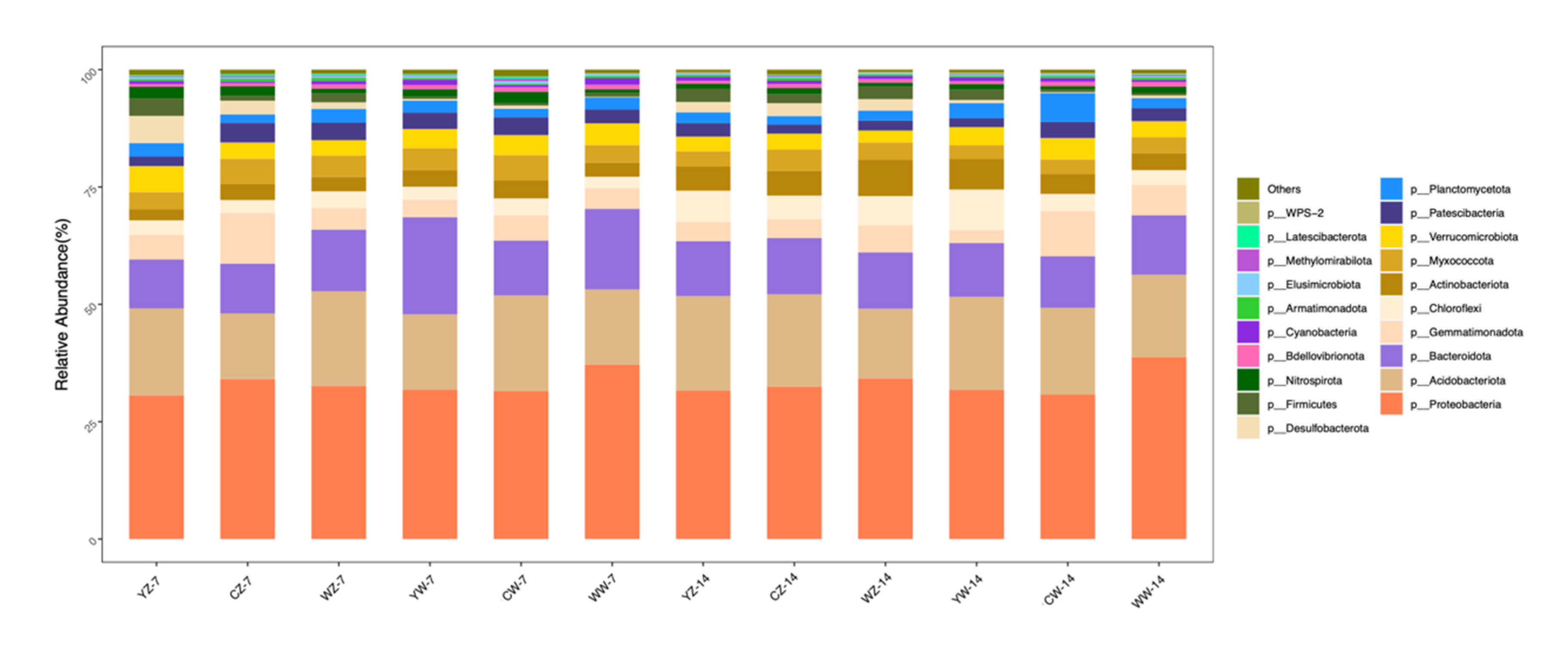
Figure 6.
Changes in relative abundance of bacterial phyla in the rhizosphere of rapeseed in different treatment groups, with treatments as in Table 1; *: P<0.05.
Figure 6.
Changes in relative abundance of bacterial phyla in the rhizosphere of rapeseed in different treatment groups, with treatments as in Table 1; *: P<0.05.
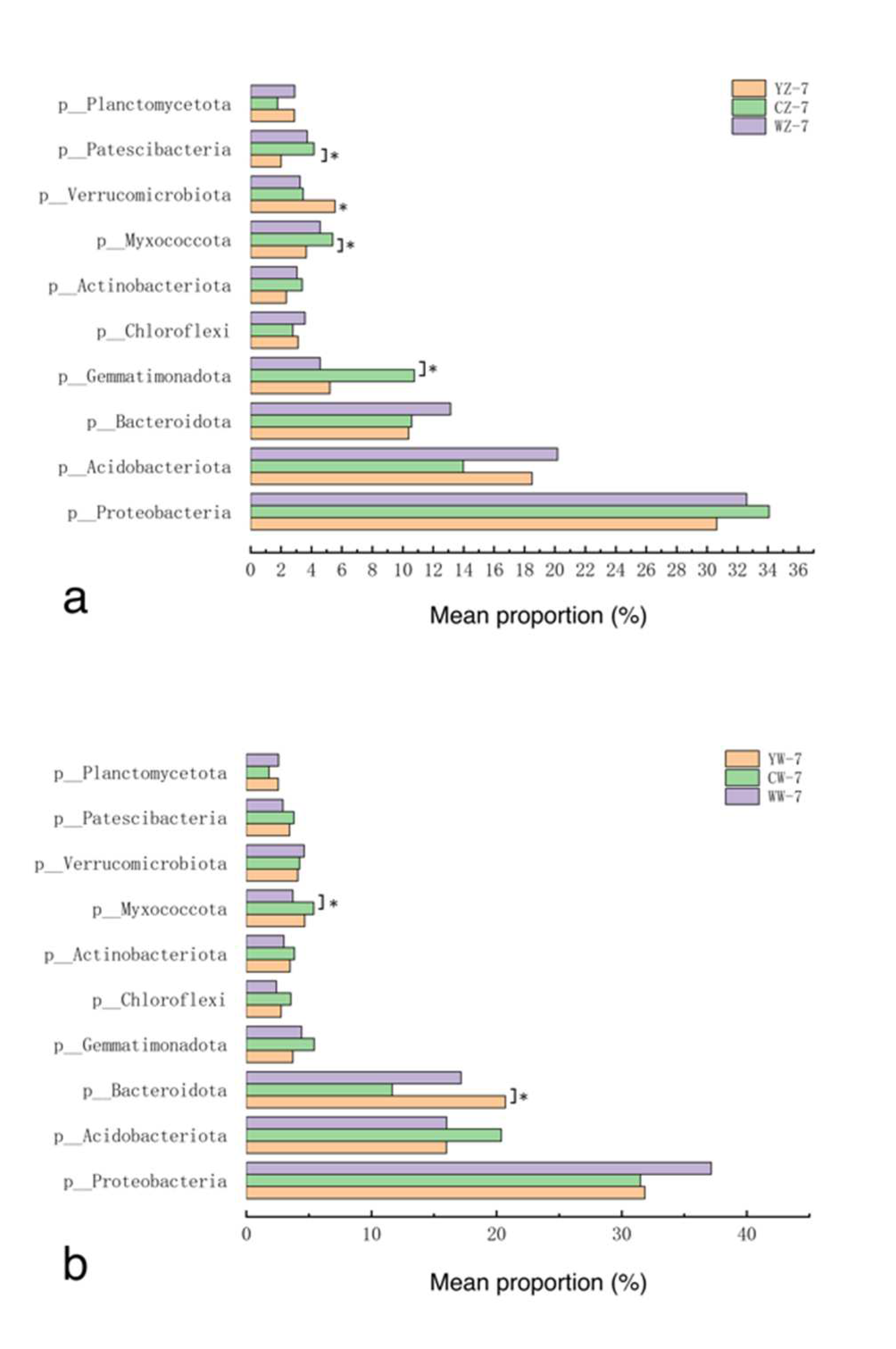
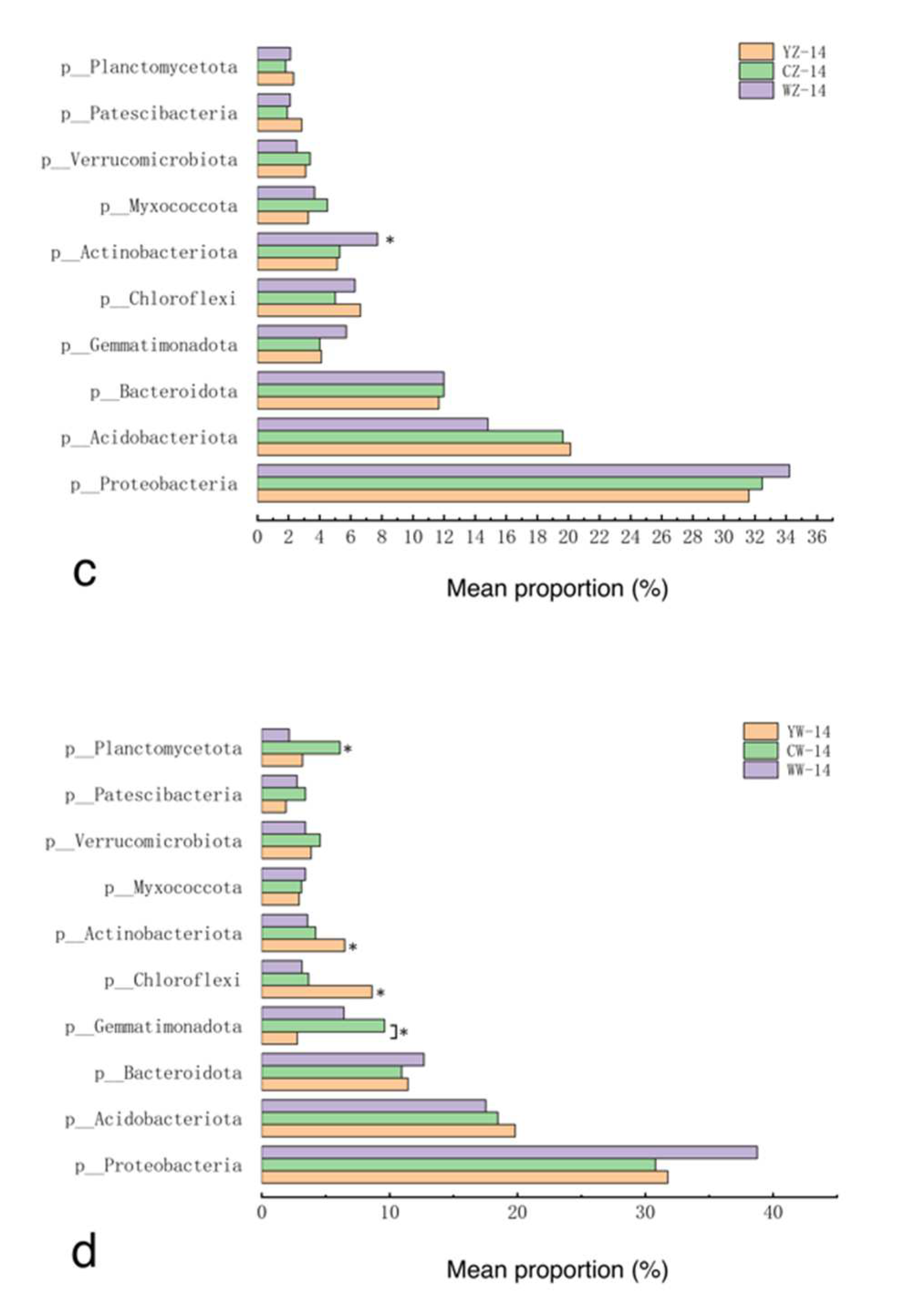
Figure 8.
heat map and cluster analysis of the composition of the integrated population, with the same table as Table 1.
Figure 8.
heat map and cluster analysis of the composition of the integrated population, with the same table as Table 1.
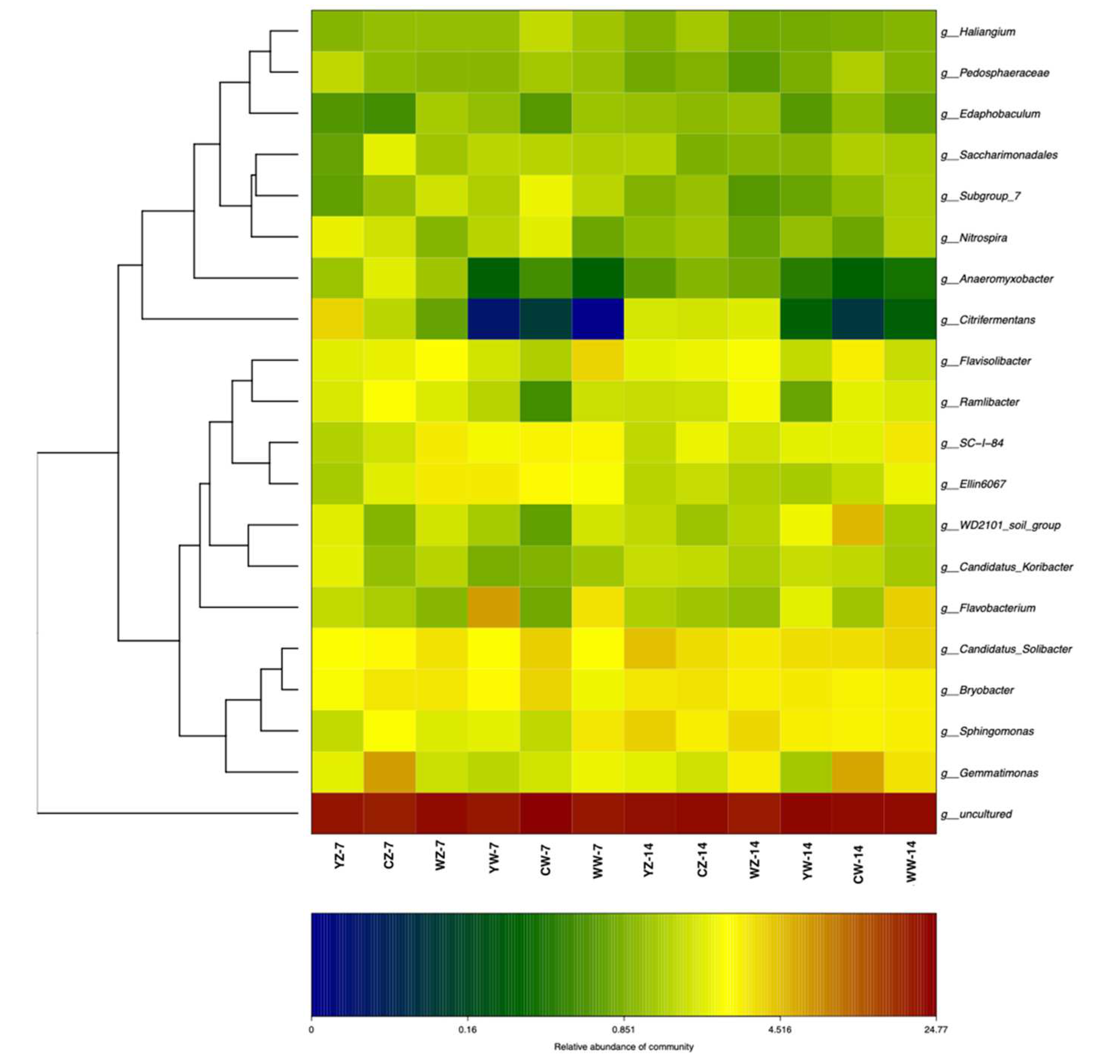
Figure 9.
Analysis of species differences between different groups. Treatments the same as Table 1.
Figure 9.
Analysis of species differences between different groups. Treatments the same as Table 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



