
Submitted:
24 January 2024
Posted:
24 January 2024
You are already at the latest version
Abstract
The DNA damage response (DDR) system is a complicated network of signaling pathways that detects and repairs DNA damage or induces apoptosis. Critical regulators of the DDR network include the DNA damage kinases ataxia telangiectasia mutated Rad3-related kinase (ATR) and ataxia-telangiectasia mutated (ATM). The ATR pathway coordinates processes such as replication stress response, stabilization of replication forks, cell cycle arrest, and DNA repair. ATR inhibition disrupts these functions, causing reduction of DNA repair, accumulation of DNA damage, replication fork collapse, inappropriate mitotic entry and mitotic catastrophe. Recent data have shown that the inhibition of ATR can lead to synthetic lethality in ATM-deficient malignancies. In addition, ATR inhibition plays a significant role in the activation of the immune system by increasing the tumor mutational burden and neoantigen load as well as by triggering the accumulation of cytosolic DNA and subsequently inducing the cGAS-STING pathway and the type I IFN response. Taken together, we review stimulating data showing that ATR kinase inhibition can alter the DDR network, the immune system and their interplay and therefore potentially provide a novel strategy to improve the efficacy of antitumor therapy, using ATR inhibitors as monotherapy or in combination with genotoxic drugs and/or immunomodulators.
Keywords:
ATR
; ATR inhibitor
; ceralasertib (AZD6738)
; DNA damage response
; immune system
; synthetic lethality
; ATR-ATM interplay
1. Introduction
Cells face constant exposure to multiple DNA damage sources, both endogenous (e.g., oxidation, alkylation, hydrolysis, mismatch of DNA bases) and exogenous (genotoxic chemicals, UV light, ionizing radiation, etc.) [1,2,3,4,5]. To neutralize these threats and ensure genomic stability, cells have developed several mechanisms, collectively called the DNA damage response (DDR) network [6]. The DDR system includes damage sensors, transducer kinases, and effectors to maintain genomic integrity. Interestingly, recent data have shown that the deregulated DDR network is capable of activating the host immune system [7]. These results potentially provide a novel strategy for enhancing the efficacy of immunotherapy.
On the other hand, deregulated DDR pathways trigger mutagenesis and genomic instability, thus getting implicated in the onset and progression of cancer. Cancer cells divide rapidly and continuously due to a breakdown of the mechanisms regulating the cell cycle. The increased proliferation rate and the DNA repair defects in cancer cells make these cells more vulnerable to specific DDR inhibition [8]. Hence, DDR inhibitors, a class of drugs that can modify the DDR network, have recently gained great attention in the research of cancer treatment. The known DDR inhibitors include drugs that inhibit different DNA repair pathways or factors, such as the polyADP-ribose polymerase (PARP), the ataxia telangiectasia mutated kinase (ATM), the ataxia telangiectasia and Rad3 related kinase (ATR), the Checkpoint kinases 1 and 2 (CHK1/2), the Cyclin-dependent kinases 4 and 6, (DK4/6), the cell-cycle checkpoint kinase WEE1, and the DNA-dependent protein kinase (DNA-PK)[8].
Particularly, ATM and ATR kinases have a critical role in the activation of the DDR network. As for ATR, following the formation of the stable replication protein A (RPA)-single-stranded DNA (ssDNA) complex at sites of DNA damage, the ATR-interacting protein (ATRIP) will bind directly to RPA, resulting in the localization of the ATR kinase to these sites [9]. Next, to give more time for the DNA repair mechanism to proceed, the ATR-CHK1 signaling pathway causes cell-cycle arrest at G2-M phase. As for ATM, this kinase is activated via the MRN (meiotic recombination protein 11 - MRE11, Nijmegen breakage syndrome protein 1 - NBS1) complex, a DNA double-strand breaks (DSBs) sensor [10]. Then, ATM phosphorylates the H2A histone family member X (H2AX) at S139 (γH2AX), and induces the CHK2 kinase, resulting in the activation of the G1-S and intra-S-phase. Based on the above, ATR and ATM kinases may be promising molecular targets in the treatment of cancer. Currently, numerous small molecule ATM/ATR inhibitors have been discovered and are undergoing preclinical and clinical evaluation.
Herein, we present a review of the current literature summarizing the role of ATR inhibition in the modification of the DDR network, the immune system and their interplay. The latest advances of ATR inhibitors in preclinical and clinical states are also elucidated.
2. The ATR Pathway in the DNA Damage Response Network
The DNA damage response network is activated following the detection of DNA damage by specific sensors [6]. The next step is the activation of a signal transduction cascade which leads to the induction of genome protection mechanisms, such as DNA repair pathways, cell cycle checkpoints, or the initiation of apoptosis. Deregulated DDR may also result in mutagenesis and genomic instability. Since DDR is an important cellular network of molecular pathways that regulates the cell's decision to repair the DNA damage or to undergo apoptosis, it is implicated in both the onset and progression of a disease, as well as in the outcome of therapeutic treatment.
There are several DNA repair mechanisms active throughout the cell cycle, including the Fanconi Anemia (FA) pathway, which is implicated in the repair of interstrand crosslinks (ICLs), the Nucleotide Excision Repair (NER), which removes adducts that disrupt the DNA double-helix, the Base Excision Repair (BER), coping with alkylated, oxidized and deaminated bases, the Mismatch Repair (MMR) pathway that resolves mismatched bases that may occur during DNA replication, the homologous recombination (HR) repair and the non-homologous DNA end joining (NHEJ), two major subpathways for the repair of DSB, the most lethal type of DNA lesion [11].
It is generally accepted that in cells with dysfunctional DDR, such as cancer cells, DNA integrity is often compromised. During the S phase, the replication fork is usually stalled by DNA lesions and if these remain unresolved, the replication machinery eventually collapses [12]. This condition is referred to as “replication stress” and is a common characteristic of tumor cells due to chronic proliferation, being also the main cause of genomic instability in cancer. Nonetheless, it may also be noticed in normal cells, on account of oxidative stress or other endogenous damage [12]. DDR also needs to modulate cell cycle progression, as cell cycle arrest is required for the resolution of DNA lesions. Two major kinases appear to be the key players organizing the response right after DNA damage recognition, ATM and ATR (Figure 1)[13].
ATM is mostly activated in response to DSBs during all phases of cell cycle [6,14], while ATR is involved in the recognition of single-strand breaks (SSBs), occurring as a response to numerous mechanisms (e.g., during replication fork stalling or as NER and DSBs repair intermediates) [13,15,16,17]. The broad involvement of ATR in various processes (replication stress response, SSBs and DSBs repair, interstrand crosslink repair, meiosis) is highlighted by that ATR, and not ATM, is indispensable for cell survival [18,19]. Particularly, ATR is an essential protein with scarce loss-of-function mutations in cancer [20], while it has been observed that impaired ATR function in mouse models leads to tumorigenesis resistance [21].
Of note, previous studies have shown crosstalk between the ATR and the ATM pathways [22,23]. As far as DSBs repair is concerned, ATM-dependent activation of ATR has been shown to occur [24,25]. DSBs are primarily detected by the MRN complex that is vital for the activation of ATM [26,27,28,29,30]. During the DSBs repair that is mediated by ATM signaling, ssDNA fragments are often accumulated as a result of the resection of DSBs by exo- and endo-nucleases [24]. These ssDNA fragments stimulate the ATR pathway, forming the ATR-ATM interplay during DSBs repair [23].
ATR pathway initiates with the RPA protein recognizing and coating ssDNA, followed by the binding of ATR-interacting protein (ATRIP) and the assembly of ATR-ATRIP complex at the DNA damage sites. However, this is not sufficient for the activation of ATR [31]. Several additional regulatory proteins, including the Rad17 complex, the Rad9–Rad1–Hus1 (9-1-1) complex, and the 9-1-1 interacting nuclear orphan (RHINO), need to be implicated [23,32,33] in order to recruit the DNA topoisomerase 2-binding protein 1 (TOPBP1) that finally stimulates the kinase activity of ATR [19,23,33]. Consequently, with the aid of mediators such as claspin [34], ATR phosphorylates the downstream Checkpoint Kinase 1 (CHK1). This pathway can result in cell cycle arrest either in the intra-S-phase or in the G2/M phase [12,13], as CHK1 is responsible for the phosphorylation of multiple substrates including phosphatases CDC25A, CDC25B and CDC25C [19]. This results in their inhibition, preventing them from keeping active the kinases CDK2 and CDK1, thus blocking cell cycle progression [35,36].
3. The ATR Pathway and the Interplay between the DDR Network and the Immune System
The Immune System and the DDR network are important mechanisms that are implicated in the survival of the living organisms. Interestingly, a growing number of data have shown that these two systems play a crucial role in the onset and progression of cancer, as well as in the outcome of anticancer therapy [37]. Traditionally, conventional chemotherapy has been considered immunosuppressive and several chemotherapeutics are used to treat autoimmune conditions. On the other hand, accumulating data suggest that DNA damaging agents can promote immunogenicity in a variety of ways, some of which have the potential to be exploited in relation to immunotherapy. Several mechanisms are implicated in the DDR-mediated activation of the immune system, including the following:
- The induction of immunogenic cell death, i.e., cell death which elicits an immune response [38]. Not all modes of cell death induce such a response which requires, in addition to neoantigen exposure, the presence of additional danger signals [39]. Such signals are provided by damage-associated molecular patterns (DAMPs), i.e., molecules released from dying tumor cells that stimulate the recruitment of antigen-presenting cells to the site, where they process and present tumor neoantigens, thereby priming an adaptive immune response. DAMPs released during chemotherapy-induced immunogenic cell death include, among others, DNA release in the cytoplasm where it leads to activation of stimulator-of-interferon genes (STING) and induction of type I interferon (IFN) and pro-inflammatory cytokines [40].
- The increase in antigen presentation through the upregulation of MHC-1 (major histocompatibility complex type 1) expression on tumor cells and promotion of dendritic cell maturation, priming them for an adaptive immune response [41].
- Increase of the tumor neoantigen burden. There are indications that genotoxic drugs may enhance tumor immunogenicity by causing, thanks to their mutagenicity, an increase of tumor neoantigens, which appear to play a critical role in the effectiveness of immune checkpoint blockade immunotherapy [48,49,50].
Interestingly, previous studies have demonstrated that a shift in the balance between DNA damage and repair causes the accumulation of cytosolic DNA that can act as potent immune-stimulator via the induction of the cGAS/STING pathway and the subsequent activation of the type-I interferon (IFN) signaling pathway [51,52,53]. Other studies have also shown that the progression of the cell cycle through mitosis in the presence of DNA DSBs results in the generation of micronuclei and the activation of the immune system [54,55]. Foreign DNA detection is a crucial step in the induction of immunity in many organisms. In mammalian cells, activation of the immune responses is contributed mainly by the cyclic GMP-AMP synthase (cGAS) –STING pathway, which plays an important role for coupling the detection of the DNA to the activation of the innate immune defence mechanisms [56]. In this pathway, the binding of cGAS to dsDNA induces its catalytic activity and results in the production of 2′,3′-cyclic GMP–AMP (cGAMP), a second messenger molecule, acting as a potent agonist of STING [57,58]. The synthesis of cGAMP is an important first step that results in the activation of the cGAS-mediated antiviral effects in several species [59]. Indeed, the cGAS molecule is activated by bacterial and viral DNA as well as by mitochondrial DNA and phagocytosed DNA that are abnormally localized in the cytosol. The induction of cGAS produces cGAMP that activates STING and leads to the induction of TANK-binding kinase 1 (TBK1), IkB kinase (IKK) and NF-kB inducing kinase (NIK) [60,61]. Together, induction of these kinases leads to the activation and nuclear translocation of IFN regulatory factor 3 (IRF3) and NF-kB, resulting in the expression of type I IFN, interferon-stimulated genes (ISGs) and inflammatory cytokines-further connecting the DDR network with the immune system [62,63]. On the other hand, extensive observations suggest that chronic activation of cGAS/STING can induce an immune suppressive tumor microenvironment (TME) that promotes the progression of the tumor [64,65,66]. In line with these data, activation of cGAS/STING pathway may have either a pro-tumor or an anti-tumor effect, depending on the stage of tumor progression and the tissue-specific context.
Since cytoplasmic dsDNA can activate STING, chemotherapies that result in the accumulation of cytoplasmic dsDNA may be an alternative strategy for STING activation. Indeed, genotoxic therapies including radiotherapy, cytotoxic chemotherapy, inhibitors of PARP and/or ATR augmented cytosolic DNA damage-induced dsDNA and activate the cGAS-STING-IFN response [67,68,69,70] with S-phase DNA damage being a particularly potent activator [71]. The activation of cGAS/STING inflammatory responses following DNA damage by PARP [72] or ATR [73] inhibition may also induce the formation of micronuclei with subsequent leaking of DNA from the micronuclei able to activate the innate immune response [52,74]. Micronuclei are small organelles that contain DNA and are produced as a result of genotoxic stress and chromosome missegregation in subsequent cell division [75]. Although these organelles are formed with a nuclear envelope (NE), after mitosis they lose compartmentalization as their NE ruptures [76]. A consequence of micronuclei rupture is that chromosomal DNA become accessible to cGAS and the subsequent induction of immune responses [52,54,55,77,78,79].
Concerning the ATR inhibition, an accumulating body of evidence suggests that the ATR pathway modulates the antitumor immunity (Figure 2). Indeed, ATR is induced in response to replication stress, single-stranded DNA and increased R-loops, and activates a kinase signaling cascade that involves CHK1 and WEE1 kinases that, in turn, leads to the activation of a checkpoint and the arrest of the cell cycle in order to give more time to the DNA repair mechanism to remove lesions [80]. In line with these data, the inhibition of ATR disrupts these functions of ATR, resulting in inappropriate mitotic entry and mitotic catastrophe. Moreover, the cytosolic DNA released may induce the cGAS-STING pathway and a type I IFN response. In addition, the inhibition of ATR plays an important role in augmenting tumor mutational burden (TMB) and neoantigen repertoire. Indeed, previous reports have studied the role of DDR inhibition as a means of increasing the TMB and the production of neoantigens[54] which may, in turn, increase the sensitivity to immune checkpoint blockade by elevated antigen presentation. Interestingly, samples harboring mutations in DNA damage signaling genes, such as ATR, showed increased neoantigen levels, thus enhancing the rationale for combination therapies using PD-1/PD-L1 blocking and ATR inhibitors. This is supported by preliminary data in a syngeneic mouse model of head and neck squamous cell carcinoma (HNSCC), where ATR inhibition by AZD6738 resulted in cGAS/STING pathway activation and induced tumor infiltration of cytotoxic T cells that eventually achieved tumor growth arrest and prolonged survival [81].
In line with these results, previous studies have shown that inhibitors of ATR potentiate immune stimulation following exposure to radiotherapy. Indeed, combined treatment with radiotherapy and ATR inhibitor induced type I/II IFN signaling and infiltration of CD8+ T-cells in a manner dependent on cGAS/STING [82,83,84]. While ATR inhibitors do not directly induce DNA damage, one may assume that the higher immunogenicity observed in irradiated tumors in the context of ATR inhibition is due to overriding of the G2/M cell cycle checkpoint. As a result, an increased proportion of cells with non-repaired DNA lesions enter mitosis, resulting in DNA fragmentation and micronuclei formation that trigger innate immune responses[54,85,86]. Furthermore, the inhibition of the ATR effector kinase CHK1 has been observed to abrogate the G2/M checkpoint after irradiation, resulting in the formation of micronuclei and type I IFN signaling in cancer cells [87]. Additionally, administering a combination of radiotherapy and the CHK1 inhibitor AZD7762 to mice led to increased CD8+ T-cell infiltration and a reduction in tumor volume compared to individual treatments with these agents [87]. Furthermore, ATR inhibition may enhance tumor immunogenicity by reducing the expression of programmed cell death 1 ligand 1 (PD-L1) in irradiated cancer cells [82,88,89]. Also, preclinical studies have indicated that cells that experience high replication stress may be selectively eliminated by ATR inhibition [90]. Indeed, the researchers observed that as the level of single-stranded DNA increased, a greater proportion of cells treated with ATR inhibitors underwent mitotic catastrophe. This finding suggests that the degree of cellular replication stress and the extent of ATR inhibitor-induced single-stranded DNA could potentially serve as predictive factors for the sensitivity to ATR inhibition.
Lately, it has been shown that except for the induction of immune response through canonical cGAS/STING signaling, the combination of irradiation therapy and ATR inhibition can also activate non-canonical STING pathway. As a result, a more robust immune activation was achieved leading to increased type I interferon-related gene expression and T cell infiltration turning the “cold” tumor microenvironment into “hot” and, thus, restoring sensitivity to PD-L1 immunotherapy [91].
Taken together, DDR-targeted therapies, including the inhibition of the ATR kinase, have the potential to enhance the antitumor immune response through various mechanisms, including the augmentation of antigenicity, the promotion of genomic instability in tumor cells, the activation of cytosolic immunity, as well as the modulation of different components that influence the interaction between tumor and immune cells [92].
4. The ATR Pathway as a Therapeutic Target
Chemotherapy resistance is a common challenge in cancer treatment, as the activation of a functional DDR can lead to cell cycle arrest and prolonged DNA repair [6]. Blocking the ATR pathway can reverse this state and enhance the cytotoxicity of genotoxic drugs by abrogating the cell cycle checkpoint [22,93].
4.1. ATM Functionality as a Predictive Biomarker of ATR-Blockade Response
A large-scale screening in in vitro and in vivo preclinical models of colorectal cancer has indicated that DDR inhibitors in general and ATR inhibitors specifically, are strong candidates for immunotherapy alternatives and has also suggested various response-predictive biomarkers for ATR inhibition, such as ATM protein loss [94]. Interestingly, recent data confirmed preclinical findings that the inhibition of ATR can lead to synthetic lethality in ATM-deficient malignancies. In fact, a study in ATM-deficient/p53-null cancer cells showed that ATR inhibition with VE-821 resulted in increased cytotoxicity after treatment with a variety of genotoxic agents, including platinum-based drugs, radiation, antimetabolites (gemcitabine), and topoisomerase inhibitors (camptothecin and etoposide). Importantly, VE-821 demonstrated a synergistic effect in tumor cells, but not in normal cells [95]. Another study has also presented synergy between cisplatin and the ATR inhibitor ceralasertib (AZD6738) in ATM-deficient NSCLC (Non-small cell lung cancer) cells [96]. Together, these data suggest that inhibition of ATR can lead to synthetic lethality in ATM-deficient/p53-null cancer cells that depends on alternative pathways to repair DSBs [97]. Strikingly, combining ceralasertib with cisplatin resulted in an enhanced cytotoxic effect even in ATM-proficient cell lines [98]. Of note, although previous studies have shown that ATM is implicated in preserving replication fork integrity and maintaining DNA replication [99], in ATM-proficient tumors, the ATR pathway also plays the most important role in replication stress management. For example, tumors expressing oncogenes (e.g., Ras, Myc) known to induce high replication stress [100], exhibited a strong response to ATR inhibition even without additional genotoxic treatment [101,102,103,104]. In fact, ATR appears to be crucial for the survival of those tumors, rendering ATR inhibition monotherapy a potential anticancer treatment [12,33].
4.2. ATR Inhibitors Synergy with Other Anti-Tumor Therapies
ATR inhibitor AZD6738 has been proved to synergize with chemotherapy agents like cisplatin in various solid tumor preclinical models, resulting in augmented antitumor activity [98,105]. Likewise, berzosertib (VE-822, VX-970, M6620) has been found to increase cell death both in cell lines and in patient-derived primary lung xenografts after cisplatin treatment, while also exhibiting a strong effect in tumor growth arrest in NSCLC models [106,107]. Other chemotherapy drugs may be also combined with ATR inhibition. Indeed, a recent study has shown synergism of the ATR inhibitor AZD6738 with the topoisomerase I inhibitor belotecan in ovarian cancer models [108], while combination with the antimetabolite gemcitabine in pancreatic models has been shown to instigate high replication stress leading to increased cell death and tumor shrinkage [109].
Recently, it has also been reported that AZD6738 can result in augmented cytotoxicity in vitro and tumor regression in vivo, when combined with Trastuzumabderuxtecan (T-DXd), an anti-HER2 antibody-topoisomerase I inhibitor hybrid [110], as well as improve the effectiveness of PI3K inhibitors, probably by DSBs-induced apoptosis as shown both in vitro and in vivo in preclinical models of breast cancer [111]
An intriguing idea has led to testing the combination of ATR inhibition with poly (ADP-ribose) polymerase (PARP) inhibitors. PARP is an essential protein for multiple DDR pathways and several inhibitors, such as olaparib, have been synthesized and are currently used in the clinical practice. Olaparib induces DNA damage and activates BRCA1/2-dependent homologous recombination. Thus, it is used to cause synthetic lethality in BRCA1/2-deficient cancers or with synchronous administration of HR-blocking agents [112]. Accumulating data show that the ATR inhibitor AZD6738 synergizes with olaparib to overcome resistance and achieve induced cytotoxicity in ATM-deficient tumors and/or tumors with impaired HR repair [113,114]. However, it has also been proved that AZD6738 combined with olaparib and radiotherapy can benefit therapeutically even HR-proficient tumors, through “PARP trapping” and the formation of PARP-DNA complexes that impede DNA replication [115].
A recent study underlined the significance of ATR inhibition scheduling during therapy [116]. The authors reported that to achieve increased cytotoxic T cells in the tumor-draining lymph node (DLN), radiation therapy or immune checkpoint inhibition must be followed by a short ATR inhibition, rather than a prolonged one.
4.3. ATR Inhibition in Clinical Studies
Several clinical trials of ATR inhibitors are ongoing and promise to radically alter the treatment landscape in a variety of solid tumors, either as monotherapies or in combinational treatments. As we anticipate the results of next phase trials in the upcoming years, we briefly present some of the most encouraging data from the clinic. One of the agents further along in clinical development is ceralasertib (AZD6738), with several phase 2 trials having completed recruitment and reporting clinical outcomes. Relevant clinical trials of ceralasertib in patients with solid tumors are listed in Table 1.
4.3.1. Breast Cancer
A phase I trial of ceralasertib and olaparib included patients with pretreated, HRR-wild type metastatic triple negative breast cancer (mTNBC) (n=25) or BRCA-mutated HER2-negative metastatic breast cancer (n=37). In the first cohort, no responses were observed, with a median progression-free survival (PFS) of 3.1 months (80% confidence interval [CI]: 2.0-3.9 months). In the BRCA-mutated cohort, the overall response rate (ORR) was 35%, with a median PFS of 7.7 months (80% CI: 5.8-11.4 months) [117]. The results of this study were further evaluated in VIOLETTE (NCT03330847), a randomized phase 2 trial evaluating the combination of ceralasertib and olaparib in comparison to olaparib monotherapy or the combination of olaparib and adavosertib, a WEE1 inhibitor, in patients with pretreated mTNBC. In this study (n=273 patients), the combination of ceralasertib and olaparib did not improve PFS over olaparib monotherapy (7.3 vs. 7.4 months, hazard ratio [HR]: 1.02, 90% CI: 0.63-1.66, p = 0.94). Interestingly, while response rates were similar between the combination therapy and olaparib monotherapy (50% vs. 44%), the ORR with the combination therapy was higher in patients without HRR gene mutations (15% vs. 4%, odds ratio: 4.45; 90% CI: 1.30-21.20, p = 0.04) [118]. In the plasmaMATCH trial, which included patients with pretreated mTNBC (n=70), the ORR was 17.1% (95% CI: 10.4%-25.5%), with a median PFS of 4.3 months. Responses were observed in patients without BRCA1/2 mutations that had functional HR-deficiency (HRD) by RAD51 foci [119]. This may account for the responses observed in patients without HRR gene mutations in the VIOLETTE study, and this subset of patients requires further evaluation in future clinical trials.
The combination of ceralasertib and olaparib, among other combinations, is being evaluated in mTNBC in another phase 2 trial that is actively recruiting patients (NCT03801369). Other than mTNBC, this combination is also being evaluated in HER2-negative, germline BRCA mutated advanced or metastatic breast cancer patients pre-treated with PARP inhibitors (NCT04090567). Trials evaluating other drug combinations include ATRiBRAVE (NCT05582538), an open-label phase 2 trial of mTNBC patients that have experienced disease progression after locoregional therapy that included chemotherapy and immunotherapy. In this trial, patients will receive a priming therapy by ceralasertib followed by the combination of paclitaxel and durvalumab, aiming to restore sensitivity to immunotherapy.
4.3.2. Lung Cancer
The HUDSON trial (NCT03334617) is an open-label, biomarker-directed trial for patients with metastatic NSCLC (mNSCLC) after progression on chemotherapy and immunotherapy. In this trial, the combination of durvalumab with ceralasertib demonstrated early signals of efficacy in patients with ATM alterations (n=18 patients, 6-month overall survival [OS]: 100%, 6-month PFS: 61.2%, ORR: 13.3%) and in unselected patients (n=20 patients, 6-month OS: 74.8%, 6-month PFS: 53.8%, ORR: 11.1%) [120]. As a result, a randomized phase 2 trial (NCT03833440) and a randomized phase 3 trial (NCT05450692) are ongoing, which compare the combination of durvalumab and ceralasertib in mNSCLC patients who have progressed on chemotherapy and immunotherapy with the standard of care in this indication (docetaxel) and may be practice-changing in this setting. In another open-label, biomarker-directed clinical trial, the National Lung Matrix Trial (NLMT), which included pretreated, KRAS-mutated or KRAS-wild type mNSCLC patients that had received prior immunotherapy, outcomes were numerically higher in patients with KRAS mutations (ORR: 13.8% vs. 4.8%, mPFS 5.95 vs. 3.9 months, mOS 30.9 vs 13.2 months).
In extensive-stage small cell lung cancer (ES-SCLC), a small phase 2 trial is ongoing in the first-line setting that evaluates the efficacy of maintenance therapy with ceralasertib plus durvalumab after 4 cycles of induction therapy with platinum-etoposide-based chemotherapy and durvalumab (NCT04699838). Results have been reported from an open-label phase 2 trial that included 21 patients with platinum-refractory ES-SCLC that received ceralacertib plus olaparib (NCT02937818), the ORR was 4.8%, with a 12-week disease control rate of 38.1%. Interestingly, despite the disappointing response rate, the median OS for patients receiving the combination therapy was 7.56 months, which is in line with approved therapies for this indication. Similar results were seen in SUKSES, a phase 2 umbrella trial that included patients with refractory ES-SCLC that received the combination therapy. In this study, the ORR was 3.8%, median PFS was 2.75 months (95% CI: 1.77–5.44 months), and median OS was 7.18 months (95% CI: 5.97-10.79 months) [121].
4.3.3. Gynaecological cancers
The combination of ceralasertib and olaparib has shown early clinical activity in patients with advanced, high-grade serous ovarian cancer (HGSOC) that have progressed after treatment with a PARP inhibitor. In OLAPCO, a basket trial of olaparib combinations in heavily pre-treated patients, responses or prolonged disease stabilization with ceralasertib plus olaparib were observed in patients with ATM mutations and patients who had received prior treatment with a PARP inhibitor [122]. In CAPRI, an open-label phase 2 trial of the same combination, promising clinical activity was seen in patients with HGSOC enrolled immediately after progression on a PARP inhibitor (n=13), with an ORR of 46% [123]. No responses were observed in patients with platinum-resistant disease (mPFS was 4.2 months overall (90% CI: 3.5–8.2 months) [124].
The combination of ceralasertib and olaparib has also been evaluated in rare gynaecological cancers in the open label phase 2 ATARI trial (n=78). In this study, outcomes were similar in patients with clear cell histology with or without AT-rich interactive domain-containing protein 1A (ARID1A) loss (ORR: 14% vs. 14%, median PFS: 3.6 vs. 3.5 months), while the clinical activity of the combination therapy may be higher in patients with non-clear cell histologies (ORR: 24%, median PFS: 5.6 months) [125].
4.3.4. Other Solid Tumors
In PATRIOT, the first-in-human trial of ceralasertib in patients with advanced solid tumors, a subset of patients with ARID1A-deficiency derived greater benefit from treatment [126]. Furthermore, an open-label phase 2 trial in patients with advanced solid tumors also included a cohort of ARID1A-deficient tumors. In this cohort, durable complete responses were achieved in 2/10 patients, for an ORR of 20% [127]. ARID1A-deficiency represents a promising target for ATR inhibition that warrants future evaluation.
The combination of ceralasertib plus durvalumab has been evaluated in various solid tumors. In an open label phase 2 trial of the combination therapy in patients with pretreated advanced gastric cancer (n=31), the ORR was 22.6% (95% CI: 9.6%-41.1%), the median PFS 3.0 months (95% CI: 2.1-3.9 months), and the median OS 6.7 months (95% CI: 3.8-9.6 months). The benefit was limited to patients with ATM deficiency or a mutational signature attributable to HRD (median PFS: 5.60 vs 1.65 months, HR: 0.13, 95% CI: 0.045-0.39, p < 0.001) [128]. This study also included a cohort of patients with melanoma who had progressed on treatment with a PD-1 inhibitor (n=30). In this cohort, the ORR was 31.0%, the median PFS was 7.1 months (95% CI: 3.6-10.6 months), and the median OS was 14.2 months (95% CI: 9.3-19.1 months) [129].
In addition to PARP inhibitors and immune checkpoint inhibitors, ceralasertib is also being evaluated in combination with chemotherapy. In a phase I trial of ceralasertib plus weekly paclitaxel in patients with advanced solid tumors, the combination was safe and showed preliminary signs of efficacy, with a ORR of 25.4%, including one complete response in a patient with melanoma [130]. A phase I trial of ceralasertib plus gemcitabine in patients with advanced solid tumors is ongoing (NCT03669601).
5. Conclusions
Taken together, data present in this report demonstrate that the inhibition of the ATR kinase can modify the DNA damage response network and the immune system. These results potentially offer a new approach to improving the effectiveness of anticancer therapy using combinations of an ATR inhibitor with genotoxic drugs and/or immunomodulators.
Author Contributions
Conceptualization, D.M. and V.L.S.; writing—review and editing, D.M, V.L.S., E.P. and A.G.; supervision, K.N.S. and V.L.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
The data presented in this study are openly available in the reference section.
Acknowledgments
None
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cadet, J.; Wagner, J.R. DNA Base Damage by Reactive Oxygen Species, Oxidizing Agents, and UV Radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559–a012559. [Google Scholar] [CrossRef]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef]
- Ganai, R.A.; Johansson, E. DNA Replication—A Matter of Fidelity. Mol. Cell 2016, 62, 745–755. [Google Scholar] [CrossRef]
- Tubbs, A.; Nussenzweig, A. Endogenous DNA Damage as a Source of Genomic Instability in Cancer. Cell 2017, 168, 644–656. [Google Scholar] [CrossRef]
- Van Houten, B.; Santa-Gonzalez, G.A.; Camargo, M. DNA repair after oxidative stress: Current challenges. Curr. Opin. Toxicol. 2018, 7, 9–16. [Google Scholar] [CrossRef]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef]
- Xu, Y.; Nowsheen, S.; Deng, M. DNA Repair Deficiency Regulates Immunity Response in Cancers: Molecular Mechanism and Approaches for Combining Immunotherapy. Cancers 2023, 15, 1619. [Google Scholar] [CrossRef]
- Cheng, B.; Pan, W.; Xing, Y.; Xiao, Y.; Chen, J.; Xu, Z. Recent advances in DDR (DNA damage response) inhibitors for cancer therapy. Eur. J. Med. Chem. 2022, 230, 114109. [Google Scholar] [CrossRef]
- Sundar, R.; Brown, J.; Ingles Russo, A.; Yap, T.A. Targeting ATR in cancer medicine. Curr. Probl. Cancer 2017, 41, 302–315. [Google Scholar] [CrossRef]
- Lavin, M.; Kozlov, S.; Gatei, M.; Kijas, A. ATM-Dependent Phosphorylation of All Three Members of the MRN Complex: From Sensor to Adaptor. Biomolecules 2015, 5, 2877–2902. [Google Scholar] [CrossRef]
- Carusillo, A.; Mussolino, C. DNA Damage: From Threat to Treatment. Cells 2020, 9, 1665. [Google Scholar] [CrossRef]
- Da Costa, A.A.B.A.; Chowdhury, D.; Shapiro, G.I.; D’Andrea, A.D.; Konstantinopoulos, P.A. Targeting replication stress in cancer therapy. Nat. Rev. Drug Discov. 2023, 22, 38–58. [Google Scholar] [CrossRef]
- Smith, H.L.; Southgate, H.; Tweddle, D.A.; Curtin, N.J. DNA damage checkpoint kinases in cancer. Expert Rev. Mol. Med. 2020, 22, e2. [Google Scholar] [CrossRef]
- Smith, J.; Mun Tho, L.; Xu, N.; A. Gillespie, D. The ATM–Chk2 and ATR–Chk1 Pathways in DNA Damage Signaling and Cancer. In Advances in Cancer Research; Elsevier, 2010; Vol. 108, pp. 73–112 ISBN 978-0-12-380888-2.
- Gusho, E.; Laimins, L. Human Papillomaviruses Target the DNA Damage Repair and Innate Immune Response Pathways to Allow for Persistent Infection. Viruses 2021, 13, 1390. [Google Scholar] [CrossRef]
- Hossain, Md.; Lin, Y.; Yan, S. Single-Strand Break End Resection in Genome Integrity: Mechanism and Regulation by APE2. Int. J. Mol. Sci. 2018, 19, 2389. [Google Scholar] [CrossRef]
- Zou, L.; Elledge, S.J. Sensing DNA Damage Through ATRIP Recognition of RPA-ssDNA Complexes. Science 2003, 300, 1542–1548. [Google Scholar] [CrossRef]
- Durocher, D.; Jackson, S.P. DNA-PK, ATM and ATR as sensors of DNA damage: variations on a theme? Curr. Opin. Cell Biol. 2001, 13, 225–231. [Google Scholar] [CrossRef]
- Saldivar, J.C.; Cortez, D.; Cimprich, K.A. The essential kinase ATR: ensuring faithful duplication of a challenging genome. Nat. Rev. Mol. Cell Biol. 2017, 18, 622–636. [Google Scholar] [CrossRef]
- Martin, J.C.; Hoegel, T.J.; Lynch, M.L.; Woloszynska, A.; Melendy, T.; Ohm, J.E. Exploiting Replication Stress as a Novel Therapeutic Intervention. Mol. Cancer Res. 2021, 19, 192–206. [Google Scholar] [CrossRef]
- Lecona, E.; Fernandez-Capetillo, O. Targeting ATR in cancer. Nat. Rev. Cancer 2018, 18, 586–595. [Google Scholar] [CrossRef]
- Fokas, E.; Prevo, R.; Hammond, E.M.; Brunner, T.B.; McKenna, W.G.; Muschel, R.J. Targeting ATR in DNA damage response and cancer therapeutics. Cancer Treat. Rev. 2014, 40, 109–117. [Google Scholar] [CrossRef]
- Shiotani, B.; Zou, L. ATR signaling at a glance. J. Cell Sci. 2009, 122, 301–304. [Google Scholar] [CrossRef]
- Jazayeri, A.; Falck, J.; Lukas, C.; Bartek, J.; Smith, G.C.M.; Lukas, J.; Jackson, S.P. ATM- and cell cycle-dependent regulation of ATR in response to DNA double-strand breaks. Nat. Cell Biol. 2006, 8, 37–45. [Google Scholar] [CrossRef]
- Myers, J.S.; Cortez, D. Rapid Activation of ATR by Ionizing Radiation Requires ATM and Mre11. J. Biol. Chem. 2006, 281, 9346–9350. [Google Scholar] [CrossRef]
- Berkovich, E.; Monnat, R.J.; Kastan, M.B. Roles of ATM and NBS1 in chromatin structure modulation and DNA double-strand break repair. Nat. Cell Biol. 2007, 9, 683–690. [Google Scholar] [CrossRef]
- Falck, J.; Coates, J.; Jackson, S.P. Conserved modes of recruitment of ATM, ATR and DNA-PKcs to sites of DNA damage. Nature 2005, 434, 605–611. [Google Scholar] [CrossRef]
- Kitagawa, R.; Bakkenist, C.J.; McKinnon, P.J.; Kastan, M.B. Phosphorylation of SMC1 is a critical downstream event in the ATM–NBS1–BRCA1 pathway. Genes Dev. 2004, 18, 1423–1438. [Google Scholar] [CrossRef]
- Lee, J.-H.; Paull, T.T. Activation and regulation of ATM kinase activity in response to DNA double-strand breaks. Oncogene 2007, 26, 7741–7748. [Google Scholar] [CrossRef]
- You, Z.; Bailis, J.M.; Johnson, S.A.; Dilworth, S.M.; Hunter, T. Rapid activation of ATM on DNA flanking double-strand breaks. Nat. Cell Biol. 2007, 9, 1311–1318. [Google Scholar] [CrossRef]
- MacDougall, C.A.; Byun, T.S.; Van, C.; Yee, M.; Cimprich, K.A. The structural determinants of checkpoint activation. Genes Dev. 2007, 21, 898–903. [Google Scholar] [CrossRef]
- Delacroix, S.; Wagner, J.M.; Kobayashi, M.; Yamamoto, K.; Karnitz, L.M. The Rad9–Hus1–Rad1 (9–1–1) clamp activates checkpoint signaling via TopBP1. Genes Dev. 2007, 21, 1472–1477. [Google Scholar] [CrossRef]
- Karnitz, L.M.; Zou, L. Molecular Pathways: Targeting ATR in Cancer Therapy. Clin. Cancer Res. 2015, 21, 4780–4785. [Google Scholar] [CrossRef]
- Osborn, A.J.; Elledge, S.J.; Zou, L. Checking on the fork: the DNA-replication stress-response pathway. Trends Cell Biol. 2002, 12, 509–516. [Google Scholar] [CrossRef]
- Boutros, R.; Dozier, C.; Ducommun, B. The when and wheres of CDC25 phosphatases. Curr. Opin. Cell Biol. 2006, 18, 185–191. [Google Scholar] [CrossRef]
- Karlsson-Rosenthal, C.; Millar, J.B.A. Cdc25: mechanisms of checkpoint inhibition and recovery. Trends Cell Biol. 2006, 16, 285–292. [Google Scholar] [CrossRef]
- Dunn, G.P.; Bruce, A.T.; Ikeda, H.; Old, L.J.; Schreiber, R.D. Cancer immunoediting: from immunosurveillance to tumor escape. Nat. Immunol. 2002, 3, 991–998. [Google Scholar] [CrossRef]
- Bezu, L.; Gomes-de-Silva, L.C.; Dewitte, H.; Breckpot, K.; Fucikova, J.; Spisek, R.; Galluzzi, L.; Kepp, O.; Kroemer, G. Combinatorial Strategies for the Induction of Immunogenic Cell Death. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef]
- Galluzzi, L.; Buqué, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunogenic cell death in cancer and infectious disease. Nat. Rev. Immunol. 2017, 17, 97–111. [Google Scholar] [CrossRef]
- Barber, G.N. STING: infection, inflammation and cancer. Nat. Rev. Immunol. 2015, 15, 760–770. [Google Scholar] [CrossRef]
- Liu, W.M.; Fowler, D.W.; Smith, P.; Dalgleish, A.G. Pre-treatment with chemotherapy can enhance the antigenicity and immunogenicity of tumours by promoting adaptive immune responses. Br. J. Cancer 2010, 102, 115–123. [Google Scholar] [CrossRef]
- Chatzinikolaou, G.; Karakasilioti, I.; Garinis, G.A. DNA damage and innate immunity: links and trade-offs. Trends Immunol. 2014, 35, 429–435. [Google Scholar] [CrossRef]
- Malaquin, N.; Carrier-Leclerc, A.; Dessureault, M.; Rodier, F. DDR-mediated crosstalk between DNA-damaged cells and their microenvironment. Front. Genet. 2015, 6. [Google Scholar] [CrossRef]
- Banissi, C.; Ghiringhelli, F.; Chen, L.; Carpentier, A.F. Treg depletion with a low-dose metronomic temozolomide regimen in a rat glioma model. Cancer Immunol. Immunother. 2009, 58, 1627–1634. [Google Scholar] [CrossRef]
- Vincent, J.; Mignot, G.; Chalmin, F.; Ladoire, S.; Bruchard, M.; Chevriaux, A.; Martin, F.; Apetoh, L.; Rébé, C.; Ghiringhelli, F. 5-Fluorouracil Selectively Kills Tumor-Associated Myeloid-Derived Suppressor Cells Resulting in Enhanced T Cell–Dependent Antitumor Immunity. Cancer Res. 2010, 70, 3052–3061. [Google Scholar] [CrossRef]
- Sheng, J.; Fang, W.; Yu, J.; Chen, N.; Zhan, J.; Ma, Y.; Yang, Y.; Huang, Y.; Zhao, H.; Zhang, L. Expression of programmed death ligand-1 on tumor cells varies pre and post chemotherapy in non-small cell lung cancer. Sci. Rep. 2016, 6, 20090. [Google Scholar] [CrossRef]
- Ghebeh, H.; Lehe, C.; Barhoush, E.; Al-Romaih, K.; Tulbah, A.; Al-Alwan, M.; Hendrayani, F.; Manogaran, P.; Alaiya, A.; Al-Tweigeri, T.; et al. RDesoeaxrcoh arrtuiclbe icin downregulates cell surface B7-H1 expression and upregulates its nuclear expression in breast cancer cells: role of B7-H1 as an anti-apoptotic molecule. 2010.
- Rizvi, N.A.; Hellmann, M.D.; Snyder, A.; Kvistborg, P.; Makarov, V.; Havel, J.J.; Lee, W.; Yuan, J.; Wong, P.; Ho, T.S.; et al. Mutational landscape determines sensitivity to PD-1 blockade in non–small cell lung cancer. Science 2015, 348, 124–128. [Google Scholar] [CrossRef]
- Chae, Y.K.; Davis, A.A.; Raparia, K.; Agte, S.; Pan, A.; Mohindra, N.; Villaflor, V.; Giles, F. Association of Tumor Mutational Burden With DNA Repair Mutations and Response to Anti–PD-1/PD-L1 Therapy in Non–Small-Cell Lung Cancer. Clin. Lung Cancer 2019, 20, 88–96. [Google Scholar] [CrossRef]
- Strickland, K.C.; Howitt, B.E.; Shukla, S.A.; Rodig, S.; Ritterhouse, L.L.; Liu, J.F.; Garber, J.E.; Chowdhury, D.; Wu, C.J.; D’Andrea, A.D.; et al. Association and prognostic significance of BRCA1/2-mutation status with neoantigen load, number of tumor-infiltrating lymphocytes and expression of PD-1/PD-L1 in high grade serous ovarian cancer. Oncotarget 2016, 7, 13587–13598. [Google Scholar] [CrossRef]
- Härtlova, A.; Erttmann, S.F.; Raffi, F.Am.; Schmalz, A.M.; Resch, U.; Anugula, S.; Lienenklaus, S.; Nilsson, L.M.; Kröger, A.; Nilsson, J.A.; et al. DNA Damage Primes the Type I Interferon System via the Cytosolic DNA Sensor STING to Promote Anti-Microbial Innate Immunity. Immunity 2015, 42, 332–343. [Google Scholar] [CrossRef]
- Mackenzie, K.J.; Carroll, P.; Martin, C.-A.; Murina, O.; Fluteau, A.; Simpson, D.J.; Olova, N.; Sutcliffe, H.; Rainger, J.K.; Leitch, A.; et al. cGAS surveillance of micronuclei links genome instability to innate immunity. Nature 2017, 548, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Erdal, E.; Haider, S.; Rehwinkel, J.; Harris, A.L.; McHugh, P.J. A prosurvival DNA damage-induced cytoplasmic interferon response is mediated by end resection factors and is limited by Trex1. Genes Dev. 2017, 31, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Harding, S.M.; Benci, J.L.; Irianto, J.; Discher, D.E.; Minn, A.J.; Greenberg, R.A. Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature 2017, 548, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, K.; Knittler, K.; Borowski, C.; Rudnik, S.; Damme, M.; Aden, K.; Spehlmann, M.E.; Frey, N.; Saftig, P.; Chalaris, A.; et al. Absence of RNase H2 triggers generation of immunogenic micronuclei removed by autophagy. Hum. Mol. Genet. 2017, 26, 3960–3972. [Google Scholar] [CrossRef]
- Ablasser, A.; Chen, Z.J. cGAS in action: Expanding roles in immunity and inflammation. Science 2019, 363, eaat8657. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ma, Z.; Barber, G.N. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009, 461, 788–792. [Google Scholar] [CrossRef]
- Diner, E.J.; Vance, R.E. Taking the STING out of cytosolic DNA sensing. Trends Immunol. 2014, 35, 1–2. [Google Scholar] [CrossRef]
- Bai, J.; Liu, F. The cGAS-cGAMP-STING Pathway: A Molecular Link Between Immunity and Metabolism. Diabetes 2019, 68, 1099–1108. [Google Scholar] [CrossRef]
- Li, X.; Shu, C.; Yi, G.; Chaton, C.T.; Shelton, C.L.; Diao, J.; Zuo, X.; Kao, C.C.; Herr, A.B.; Li, P. Cyclic GMP-AMP Synthase Is Activated by Double-Stranded DNA-Induced Oligomerization. Immunity 2013, 39, 1019–1031. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, J.; Du, F.; Xu, H.; Sun, L.; Chen, Z.; Brautigam, C.A.; Zhang, X.; Chen, Z.J. The Cytosolic DNA Sensor cGAS Forms an Oligomeric Complex with DNA and Undergoes Switch-like Conformational Changes in the Activation Loop. Cell Rep. 2014, 6, 421–430. [Google Scholar] [CrossRef]
- Pilger, D.; Seymour, L.W.; Jackson, S.P. Interfaces between cellular responses to DNA damage and cancer immunotherapy. Genes Dev. 2021, 35, 602–618. [Google Scholar] [CrossRef]
- Paludan, S.R.; Reinert, L.S.; Hornung, V. DNA-stimulated cell death: implications for host defence, inflammatory diseases and cancer. Nat. Rev. Immunol. 2019, 19, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Xia, T.; Konno, H.; Konno, K.; Ruiz, P.; Barber, G.N. Inflammation-driven carcinogenesis is mediated through STING. Nat. Commun. 2014, 5, 5166. [Google Scholar] [CrossRef]
- Kwon, J.; Bakhoum, S.F. The Cytosolic DNA-Sensing cGAS–STING Pathway in Cancer. Cancer Discov. 2020, 10, 26–39. [Google Scholar] [CrossRef]
- Bruand, M.; Barras, D.; Mina, M.; Ghisoni, E.; Morotti, M.; Lanitis, E.; Fahr, N.; Desbuisson, M.; Grimm, A.; Zhang, H.; et al. Cell-autonomous inflammation of BRCA1-deficient ovarian cancers drives both tumor-intrinsic immunoreactivity and immune resistance via STING. Cell Rep. 2021, 36, 109412. [Google Scholar] [CrossRef]
- Shevtsov, M.; Sato, H.; Multhoff, G.; Shibata, A. Novel Approaches to Improve the Efficacy of Immuno-Radiotherapy. Front. Oncol. 2019, 9, 156. [Google Scholar] [CrossRef]
- Van Limbergen, E.J.; De Ruysscher, D.K.; Olivo Pimentel, V.; Marcus, D.; Berbee, M.; Hoeben, A.; Rekers, N.; Theys, J.; Yaromina, A.; Dubois, L.J.; et al. Combining radiotherapy with immunotherapy: the past, the present and the future. Br. J. Radiol. 2017, 90, 20170157. [Google Scholar] [CrossRef]
- Lee, E.K.; Konstantinopoulos, P.A. Combined PARP and Immune Checkpoint Inhibition in Ovarian Cancer. Trends Cancer 2019, 5, 524–528. [Google Scholar] [CrossRef]
- Ngoi, N.Y.L.; Peng, G.; Yap, T.A. A Tale of Two Checkpoints: ATR Inhibition and PD-(L)1 Blockade. Annu. Rev. Med. 2022, 73, 231–250. [Google Scholar] [CrossRef]
- Parkes, E.E.; Walker, S.M.; Taggart, L.E.; McCabe, N.; Knight, L.A.; Wilkinson, R.; McCloskey, K.D.; Buckley, N.E.; Savage, K.I.; Salto-Tellez, M.; et al. Activation of STING-Dependent Innate Immune Signaling By S-Phase-Specific DNA Damage in Breast Cancer. J. Natl. Cancer Inst. 2017, 109, djw199. [Google Scholar] [CrossRef]
- Chabanon, R.M.; Muirhead, G.; Krastev, D.B.; Adam, J.; Morel, D.; Garrido, M.; Lamb, A.; Hénon, C.; Dorvault, N.; Rouanne, M.; et al. PARP inhibition enhances tumor cell–intrinsic immunity in ERCC1-deficient non–small cell lung cancer. J. Clin. Invest. 2019, 129, 1211–1228. [Google Scholar] [CrossRef]
- Schoonen, P.M.; Kok, Y.P.; Wierenga, E.; Bakker, B.; Foijer, F.; Spierings, D.C.J.; Van Vugt, M.A.T.M. Premature mitotic entry induced by ATR inhibition potentiates olaparib inhibition-mediated genomic instability, inflammatory signaling, and cytotoxicity in BRCA2-deficient cancer cells. Mol. Oncol. 2019, 13, 2422–2440. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Mann, C.C.; Kranzusch, P.J. cGAS Conducts Micronuclei DNA Surveillance. Trends Cell Biol. 2017, 27, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Kirsch-Volders, M.; Natarajan, A.T.; Surralles, J.; Crott, J.W.; Parry, J.; Norppa, H.; Eastmond, D.A.; Tucker, J.D.; Thomas, P. Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis 2011, 26, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Hatch, E.M.; Fischer, A.H.; Deerinck, T.J.; Hetzer, M.W. Catastrophic Nuclear Envelope Collapse in Cancer Cell Micronuclei. Cell 2013, 154, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Dou, Z.; Ghosh, K.; Vizioli, M.G.; Zhu, J.; Sen, P.; Wangensteen, K.J.; Simithy, J.; Lan, Y.; Lin, Y.; Zhou, Z.; et al. Cytoplasmic chromatin triggers inflammation in senescence and cancer. Nature 2017, 550, 402–406. [Google Scholar] [CrossRef]
- Glück, S.; Guey, B.; Gulen, M.F.; Wolter, K.; Kang, T.-W.; Schmacke, N.A.; Bridgeman, A.; Rehwinkel, J.; Zender, L.; Ablasser, A. Innate immune sensing of cytosolic chromatin fragments through cGAS promotes senescence. Nat. Cell Biol. 2017, 19, 1061–1070. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Ren, J.; Chen, Q.; Chen, Z.J. cGAS is essential for cellular senescence. Proc. Natl. Acad. Sci. 2017, 114. [Google Scholar] [CrossRef]
- Wayne, J.; Brooks, T.; Landras, A.; Massey, A.J. Targeting DNA damage response pathways to activate the STING innate immune signaling pathway in human cancer cells. FEBS J. 2021, 288, 4507–4540. [Google Scholar] [CrossRef]
- Osman, A.A.; da Costa, F.H.B.; Adebayo, A.A.; Bartels, M.D.; Xie, T.; Amit, M.; Kawabe, M.; Rangel, R.; Frederick, M.J.; Sandulache, V.; et al. Abstract PO-085: Inhibition of ATR as a therapeutic strategy to enhance immunotherapy in head and neck cancer. Clin. Cancer Res. 2023, 29, PO–085. [Google Scholar] [CrossRef]
- Vendetti, F.P.; Karukonda, P.; Clump, D.A.; Teo, T.; Lalonde, R.; Nugent, K.; Ballew, M.; Kiesel, B.F.; Beumer, J.H.; Sarkar, S.N.; et al. ATR kinase inhibitor AZD6738 potentiates CD8+ T cell–dependent antitumor activity following radiation. J. Clin. Invest. 2018, 128, 3926–3940. [Google Scholar] [CrossRef]
- Dillon, M.T.; Bergerhoff, K.F.; Pedersen, M.; Whittock, H.; Crespo-Rodriguez, E.; Patin, E.C.; Pearson, A.; Smith, H.G.; Paget, J.T.E.; Patel, R.R.; et al. ATR Inhibition Potentiates the Radiation-induced Inflammatory Tumor Microenvironment. Clin. Cancer Res. 2019, 25, 3392–3403. [Google Scholar] [CrossRef]
- Sheng, H.; Huang, Y.; Xiao, Y.; Zhu, Z.; Shen, M.; Zhou, P.; Guo, Z.; Wang, J.; Wang, H.; Dai, W.; et al. ATR inhibitor AZD6738 enhances the antitumor activity of radiotherapy and immune checkpoint inhibitors by potentiating the tumor immune microenvironment in hepatocellular carcinoma. J. Immunother. Cancer 2020, 8, e000340. [Google Scholar] [CrossRef]
- Ruiz, S.; Mayor-Ruiz, C.; Lafarga, V.; Murga, M.; Vega-Sendino, M.; Ortega, S.; Fernandez-Capetillo, O. A Genome-wide CRISPR Screen Identifies CDC25A as a Determinant of Sensitivity to ATR Inhibitors. Mol. Cell 2016, 62, 307–313. [Google Scholar] [CrossRef]
- Dok, R.; Glorieux, M.; Bamps, M.; Nuyts, S. Effect of ATR Inhibition in RT Response of HPV-Negative and HPV-Positive Head and Neck Cancers. Int. J. Mol. Sci. 2021, 22, 1504. [Google Scholar] [CrossRef]
- Chao, H.-H.; Karagounis, I.V.; Thomas, C.; François, N.B.; Facciabene, A.; Koumenis, C.; Maity, A. Combination of CHEK1/2 inhibition and ionizing radiation results in abscopal tumor response through increased micronuclei formation. Oncogene 2020, 39, 4344–4357. [Google Scholar] [CrossRef]
- Sato, H.; Niimi, A.; Yasuhara, T.; Permata, T.B.M.; Hagiwara, Y.; Isono, M.; Nuryadi, E.; Sekine, R.; Oike, T.; Kakoti, S.; et al. DNA double-strand break repair pathway regulates PD-L1 expression in cancer cells. Nat. Commun. 2017, 8, 1751. [Google Scholar] [CrossRef]
- Sun, L.-L.; Yang, R.-Y.; Li, C.-W.; Chen, M.-K.; Shao, B.; Hsu, J.-M.; Chan, L.-C.; Yang, Y.; Hsu, J.L.; Lai, Y.-J.; et al. Inhibition of ATR downregulates PD-L1 and sensitizes tumor cells to T cell-mediated killing. Am J Cancer Res 2018, 8, 1307–1316. [Google Scholar]
- Buisson, R.; Boisvert, J.L.; Benes, C.H.; Zou, L. Distinct but Concerted Roles of ATR, DNA-PK, and Chk1 in Countering Replication Stress during S Phase. Mol. Cell 2015, 59, 1011–1024. [Google Scholar] [CrossRef]
- Liu, C.; Wang, X.; Qin, W.; Tu, J.; Li, C.; Zhao, W.; Ma, L.; Liu, B.; Qiu, H.; Yuan, X. Combining radiation and the ATR inhibitor berzosertib activates STING signaling and enhances immunotherapy via inhibiting SHP1 function in colorectal cancer. Cancer Commun. 2023, 43, 435–454. [Google Scholar] [CrossRef]
- Chabanon, R.M.; Rouanne, M.; Lord, C.J.; Soria, J.-C.; Pasero, P.; Postel-Vinay, S. Targeting the DNA damage response in immuno-oncology: developments and opportunities. Nat. Rev. Cancer 2021, 21, 701–717. [Google Scholar] [CrossRef]
- Lewis, K.A.; Lilly, K.K.; Reynolds, E.A.; Sullivan, W.P.; Kaufmann, S.H.; Cliby, W.A. Ataxia telangiectasia and rad3-related kinase contributes to cell cycle arrest and survival after cisplatin but not oxaliplatin. Mol. Cancer Ther. 2009, 8, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Durinikova, E.; Reilly, N.M.; Buzo, K.; Mariella, E.; Chilà, R.; Lorenzato, A.; Dias, J.M.L.; Grasso, G.; Pisati, F.; Lamba, S.; et al. Targeting the DNA Damage Response Pathways and Replication Stress in Colorectal Cancer. Clin. Cancer Res. 2022, 28, 3874–3889. [Google Scholar] [CrossRef] [PubMed]
- Reaper, P.M.; Griffiths, M.R.; Long, J.M.; Charrier, J.-D.; MacCormick, S.; Charlton, P.A.; Golec, J.M.C.; Pollard, J.R. Selective killing of ATM- or p53-deficient cancer cells through inhibition of ATR. Nat. Chem. Biol. 2011, 7, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Menezes, D.L.; Holt, J.; Tang, Y.; Feng, J.; Barsanti, P.; Pan, Y.; Ghoddusi, M.; Zhang, W.; Thomas, G.; Holash, J.; et al. A Synthetic Lethal Screen Reveals Enhanced Sensitivity to ATR Inhibitor Treatment in Mantle Cell Lymphoma with ATM Loss-of-Function. Mol. Cancer Res. 2015, 13, 120–129. [Google Scholar] [CrossRef]
- Curtin, N.J. Targeting the DNA damage response for cancer therapy. Biochem. Soc. Trans. 2023, 51, 207–221. [Google Scholar] [CrossRef]
- Vendetti, F.P.; Lau, A.; Schamus, S.; Conrads, T.P.; O’Connor, M.J.; Bakkenist, C.J. The orally active and bioavailable ATR kinase inhibitor AZD6738 potentiates the anti-tumor effects of cisplatin to resolve ATM-deficient non-small cell lung cancer in vivo. Oncotarget 2015, 6, 44289–44305. [Google Scholar] [CrossRef]
- Olcina, M.M.; Foskolou, I.P.; Anbalagan, S.; Senra, J.M.; Pires, I.M.; Jiang, Y.; Ryan, A.J.; Hammond, E.M. Replication Stress and Chromatin Context Link ATM Activation to a Role in DNA Replication. Mol. Cell 2013, 52, 758–766. [Google Scholar] [CrossRef]
- Halazonetis, T.D.; Gorgoulis, V.G.; Bartek, J. An Oncogene-Induced DNA Damage Model for Cancer Development. Science 2008, 319, 1352–1355. [Google Scholar] [CrossRef]
- Gilad, O.; Nabet, B.Y.; Ragland, R.L.; Schoppy, D.W.; Smith, K.D.; Durham, A.C.; Brown, E.J. Combining ATR Suppression with Oncogenic Ras Synergistically Increases Genomic Instability, Causing Synthetic Lethality or Tumorigenesis in a Dosage-Dependent Manner. Cancer Res. 2010, 70, 9693–9702. [Google Scholar] [CrossRef] [PubMed]
- Murga, M.; Campaner, S.; Lopez-Contreras, A.J.; Toledo, L.I.; Soria, R.; Montaña, M.F.; D’Artista, L.; Schleker, T.; Guerra, C.; Garcia, E.; et al. Exploiting oncogene-induced replicative stress for the selective killing of Myc-driven tumors. Nat. Struct. Mol. Biol. 2011, 18, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Schoppy, D.W.; Ragland, R.L.; Gilad, O.; Shastri, N.; Peters, A.A.; Murga, M.; Fernandez-Capetillo, O.; Diehl, J.A.; Brown, E.J. Oncogenic stress sensitizes murine cancers to hypomorphic suppression of ATR. J. Clin. Invest. 2012, 122, 241–252. [Google Scholar] [CrossRef]
- Toledo, L.I.; Murga, M.; Zur, R.; Soria, R.; Rodriguez, A.; Martinez, S.; Oyarzabal, J.; Pastor, J.; Bischoff, J.R.; Fernandez-Capetillo, O. A cell-based screen identifies ATR inhibitors with synthetic lethal properties for cancer-associated mutations. Nat. Struct. Mol. Biol. 2011, 18, 721–727. [Google Scholar] [CrossRef]
- Kim, H.; Min, A.; Im, S.; Jang, H.; Lee, K.H.; Lau, A.; Lee, M.; Kim, S.; Yang, Y.; Kim, J.; et al. Anti-tumor activity of the ATR inhibitor AZD6738 in HER2 positive breast cancer cells. Int. J. Cancer 2017, 140, 109–119. [Google Scholar] [CrossRef]
- Leibrandt, R.C.; Tu, M.-J.; Yu, A.-M.; Lara, P.N.; Parikh, M. ATR Inhibition in Advanced Urothelial Carcinoma. Clin. Genitourin. Cancer 2023, 21, 203–207. [Google Scholar] [CrossRef]
- Hall, A.B.; Newsome, D.; Wang, Y.; Boucher, D.M.; Eustace, B.; Gu, Y.; Hare, B.; Johnson, M.A.; Li, H.; Milton, S.; et al. Potentiation of tumor responses to DNA damaging therapy by the selective ATR inhibitor VX-970. Oncotarget 2014, 5, 5674–5685. [Google Scholar] [CrossRef]
- Hur, J.; Ghosh, M.; Kim, T.H.; Park, N.; Pandey, K.; Cho, Y.B.; Hong, S.D.; Katuwal, N.B.; Kang, M.; An, H.J.; et al. Synergism of AZD6738, an ATR Inhibitor, in Combination with Belotecan, a Camptothecin Analogue, in Chemotherapy-Resistant Ovarian Cancer. Int. J. Mol. Sci. 2021, 22, 1223. [Google Scholar] [CrossRef] [PubMed]
- Wallez, Y.; Dunlop, C.R.; Johnson, T.I.; Koh, S.-B.; Fornari, C.; Yates, J.W.T.; Bernaldo De Quirós Fernández, S.; Lau, A.; Richards, F.M.; Jodrell, D.I. The ATR Inhibitor AZD6738 Synergizes with Gemcitabine In Vitro and In Vivo to Induce Pancreatic Ductal Adenocarcinoma Regression. Mol. Cancer Ther. 2018, 17, 1670–1682. [Google Scholar] [CrossRef] [PubMed]
- Wallez, Y.; Proia, T.; Cheraghchi-Bashi-Astaneh, A.; Karmokar, A.; Wilson, Z.; Randle, S.; Anderton, M.; Durant, S.; Leo, E.; Lau, A.; et al. Abstract 5298: Activity and tolerability of combinations of trastuzumab deruxtecan (T-DXd) with inhibitors of the DNA damage response in preclinical models. Cancer Res. 2022, 82, 5298–5298. [Google Scholar] [CrossRef]
- Moon, Y.W.; Gosh, M.; Park, N.; Pandey, K.; Katwal, N.B.; Hong, S.D. Abstract P2-26-09: Synergistic activity of PI3K inhibitor in combination with AZD6738, ATR inhibitor in breast cancer preclinical model via DNA damage response pathway. Cancer Res. 2023, 83, P2–26. [Google Scholar] [CrossRef]
- Nakhjavani, M.; Hardingham, J.E.; Palethorpe, H.M.; Price, T.J.; Townsend, A.R. Druggable Molecular Targets for the Treatment of Triple Negative Breast Cancer. J. Breast Cancer 2019, 22, 341. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, R.L.; Wijnhoven, P.W.G.; Ramos-Montoya, A.; Wilson, Z.; Illuzzi, G.; Falenta, K.; Jones, G.N.; James, N.; Chabbert, C.D.; Stott, J.; et al. Combined PARP and ATR inhibition potentiates genome instability and cell death in ATM-deficient cancer cells. Oncogene 2020, 39, 4869–4883. [Google Scholar] [CrossRef] [PubMed]
- Wilson, Z.; Odedra, R.; Wallez, Y.; Wijnhoven, P.W.G.; Hughes, A.M.; Gerrard, J.; Jones, G.N.; Bargh-Dawson, H.; Brown, E.; Young, L.A.; et al. ATR Inhibitor AZD6738 (Ceralasertib) Exerts Antitumor Activity as a Monotherapy and in Combination with Chemotherapy and the PARP Inhibitor Olaparib. Cancer Res. 2022, 82, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Parsels, L.A.; Engelke, C.G.; Parsels, J.; Flanagan, S.A.; Zhang, Q.; Tanska, D.; Wahl, D.R.; Canman, C.E.; Lawrence, T.S.; Morgan, M.A. Combinatorial Efficacy of Olaparib with Radiation and ATR Inhibitor Requires PARP1 Protein in Homologous Recombination–Proficient Pancreatic Cancer. Mol. Cancer Ther. 2021, 20, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Vendetti, F.P.; Pandya, P.; Clump, D.A.; Schamus-Haynes, S.; Tavakoli, M.; diMayorca, M.; Islam, N.M.; Chang, J.; Delgoffe, G.M.; Beumer, J.H.; et al. The schedule of ATR inhibitor AZD6738 can potentiate or abolish antitumor immune responses to radiotherapy. JCI Insight 2023, 8, e165615. [Google Scholar] [CrossRef]
- Dean, E.; Krebs, M.G.; Im, S.-A.; Campone, M.; Postel-Vinay, S.; Arkenau, T.; Lopez, J.; Abida, W.; Jodrell, D.; Lee, K.-W.; et al. Abstract PS11-18: Ceralasertib (cer) in combination with olaparib (ola) in patients (pts) with advanced breast cancer (BC): Results of phase I expansion cohorts. Cancer Res. 2021, 81, PS11–18. [Google Scholar] [CrossRef]
- Tutt, A.; Nowecki, Z.; Szoszkiewicz, R.; Im, S.A.; Arkenau, H.T.; Armstrong, A.C.; Jacot, W.; Kim, J.H.; Webster, M.; Balmana, J.; et al. VIOLETTE: Randomised phase II study of olaparib (ola) plus ceralasertib (cer) or adavosertib (ada) vs ola alone in patients (pts) with metastatic triple-negative breast cancer (mTNBC). 2022. [CrossRef]
- Ring, A.; Kilburn, L.S.; Pearson, A.; Moretti, L.; Afshari-Mehr, A.; Wardley, A.M.; Gurel, B.; Macpherson, I.R.; Riisnaes, R.; Baird, R.D.; et al. Olaparib and Ceralasertib (AZD6738) in Patients with Triple-Negative Advanced Breast Cancer: Results from Cohort E of the plasmaMATCH Trial (CRUK/15/010). Clin. Cancer Res. 2023, 29, 4751–4759. [Google Scholar] [CrossRef] [PubMed]
- Besse, B.; Awad, M.; Forde, P.; Thomas, M.; Park, K.; Goss, G.; Rizvi, N.; Huemer, F.; Hochmair, M.; Bennouna, J.; et al. OA07.08 HUDSON: An Open-Label, Multi-Drug, Biomarker-Directed, Phase II Platform Study in Patients with NSCLC, who Progressed on Anti-PD(L)1 Therapy. J. Thorac. Oncol. 2021, 16, S118–S119. [Google Scholar] [CrossRef]
- Park, S.; Mortimer, P.G.; Smith, S.; Kim, H. (Rosa); Jung, H.A.; Sun, J.-M.; Ahn, J.S.; Ahn, M.-J.; Park, W.-Y.; Lee, S.-H.; et al. The clinical efficacy of olaparib monotherapy or combination with ceralasertib (AZD6738) in relapsed small cell lung cancer. J. Clin. Oncol. 2021, 39, 8562–8562. [Google Scholar] [CrossRef]
- Mahdi, H.; Hafez, N.; Doroshow, D.; Sohal, D.; Keedy, V.; Do, K.T.; LoRusso, P.; Jürgensmeier, J.; Avedissian, M.; Sklar, J.; et al. Ceralasertib-Mediated ATR Inhibition Combined With Olaparib in Advanced Cancers Harboring DNA Damage Response and Repair Alterations (Olaparib Combinations). JCO Precis. Oncol. 2021, 1432–1442. [Google Scholar] [CrossRef]
- Wethington, S.L.; Shah, P.D.; Martin, L.P.; Tanyi, J.L.; Latif, N.A.; Morgan, M.A.; Torigian, D.A.; Pagan, C.; Rodriguez, D.; Domchek, S.M.; et al. Combination of PARP and ATR inhibitors (olaparib and ceralasertib) shows clinical activity in acquired PARP inhibitor-resistant recurrent ovarian cancer. J. Clin. Oncol. 2021, 39, 5516–5516. [Google Scholar] [CrossRef]
- Shah, P.D.; Wethington, S.L.; Pagan, C.; Latif, N.; Tanyi, J.; Martin, L.P.; Morgan, M.; Burger, R.A.; Haggerty, A.; Zarrin, H.; et al. Combination ATR and PARP Inhibitor (CAPRI): A phase 2 study of ceralasertib plus olaparib in patients with recurrent, platinum-resistant epithelial ovarian cancer. Gynecol. Oncol. 2021, 163, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Leary, A.; Stewart, J.R.; Dewan, M.; Lheureux, S.; Clamp, A.R.; Ray-Coquard, I.L.; Selle, F.; Gourley, C.; Glasspool, R.M. 34O ATR inhibitor alone (ceralasertib) or in combination with olaparib in gynaecological cancers with ARID1A loss or no loss: Results from the ENGOT/GYN1/NCRI ATARI trial. ESMO Open 2023, 8. [Google Scholar] [CrossRef]
- Dillon, M.T.; Guevara, J.; Mohammed, K.; Patin, E.C.; Smith, S.A.; Dean, E.; Jones, G.N.; Willis, S.E.; Petrone, M.; Silva, C.; et al. Durable responses to ATR inhibition with ceralasertib in tumors with genomic defects and high inflammation. J. Clin. Invest. 2023. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Umetsu, S.; Dhawan, M.; Grabowsky, J.; Carnevale, J.; Howell, M.; Wilch, L.; Chapman, J.; Alvarez, E.; Calabrese, S. 512O Interim results from a phase II study of the ATR inhibitor ceralasertib in ARID1A-deficient and ARID1A-intact advanced solid tumor malignancies. Ann. Oncol. 2021, 32, S583. [Google Scholar] [CrossRef]
- Kwon, M.; Kim, G.; Kim, R.; Kim, K.-T.; Kim, S.T.; Smith, S.; Mortimer, P.G.S.; Hong, J.Y.; Loembé, A.-B.; Irurzun-Arana, I.; et al. Phase II study of ceralasertib (AZD6738) in combination with durvalumab in patients with advanced gastric cancer. J. Immunother. Cancer 2022, 10, e005041. [Google Scholar] [CrossRef]
- Kim, R.; Kwon, M.; An, M.; Kim, S.T.; Smith, S.A.; Loembé, A.B.; Mortimer, P.G.S.; Armenia, J.; Lukashchuk, N.; Shah, N.; et al. Phase II study of ceralasertib (AZD6738) in combination with durvalumab in patients with advanced/metastatic melanoma who have failed prior anti-PD-1 therapy. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2022, 33, 193–203. [Google Scholar] [CrossRef]
- Lee, J.; Tae Kim, S.; Smith, S.; Mortimer, P.G.; Loembé, B.; Hong, J.; Kozarewa, I.; Pierce, A.; Dean, E. Results from a phase I, open-label study of ceralasertib (AZD6738), a novel DNA damage repair agent, in combination with weekly paclitaxel in refractory cancer (NCT02630199). J. Clin. Oncol. 2020, 38, 3503–3503. [Google Scholar] [CrossRef]
Figure 1.
Schematic overview of the ATR/ATM pathways (Figure was created with BioRender.com).
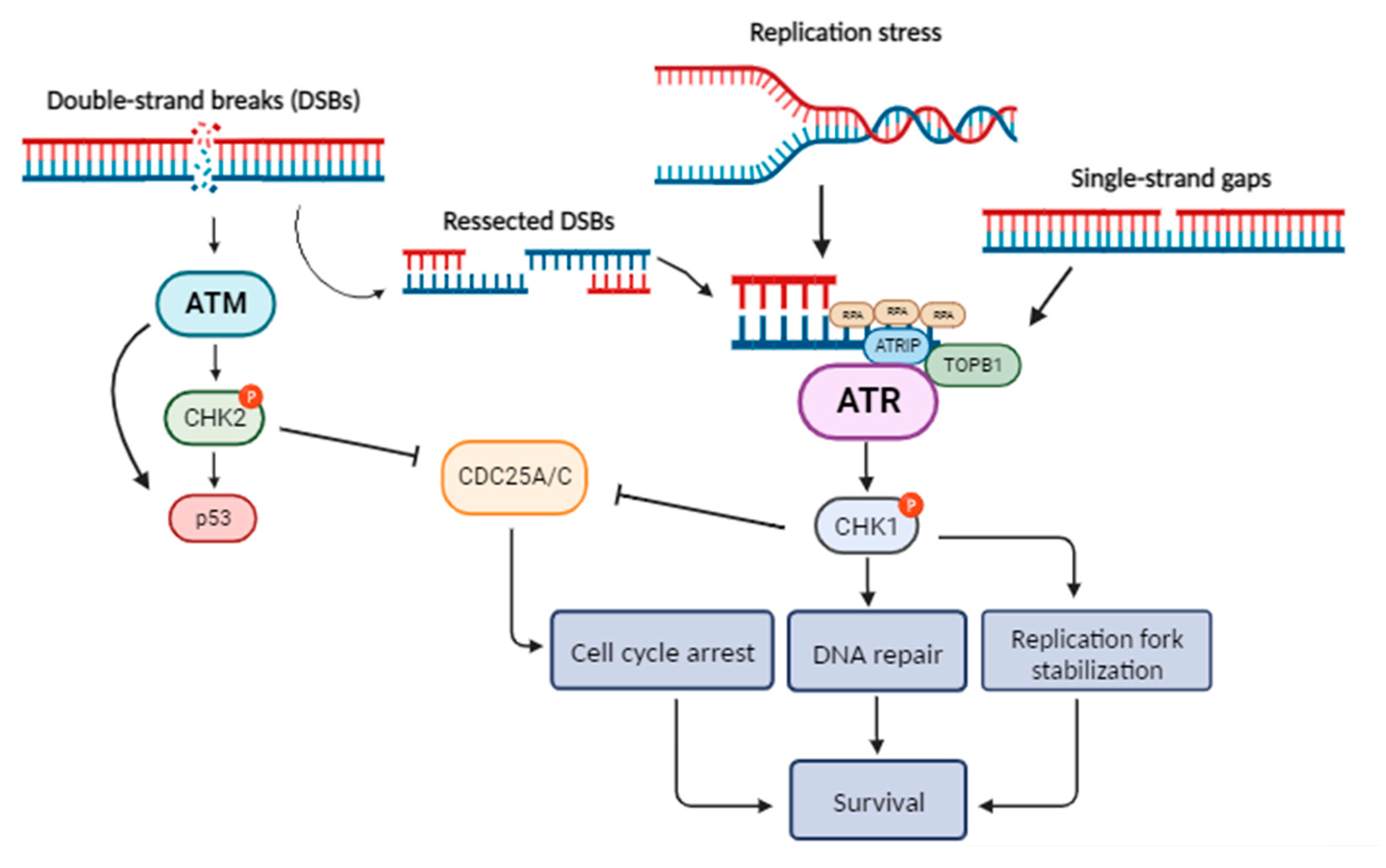
Figure 2.
ATR pathway implication in the antitumor immunity (Figure was created with BioRender.com).
Figure 2.
ATR pathway implication in the antitumor immunity (Figure was created with BioRender.com).
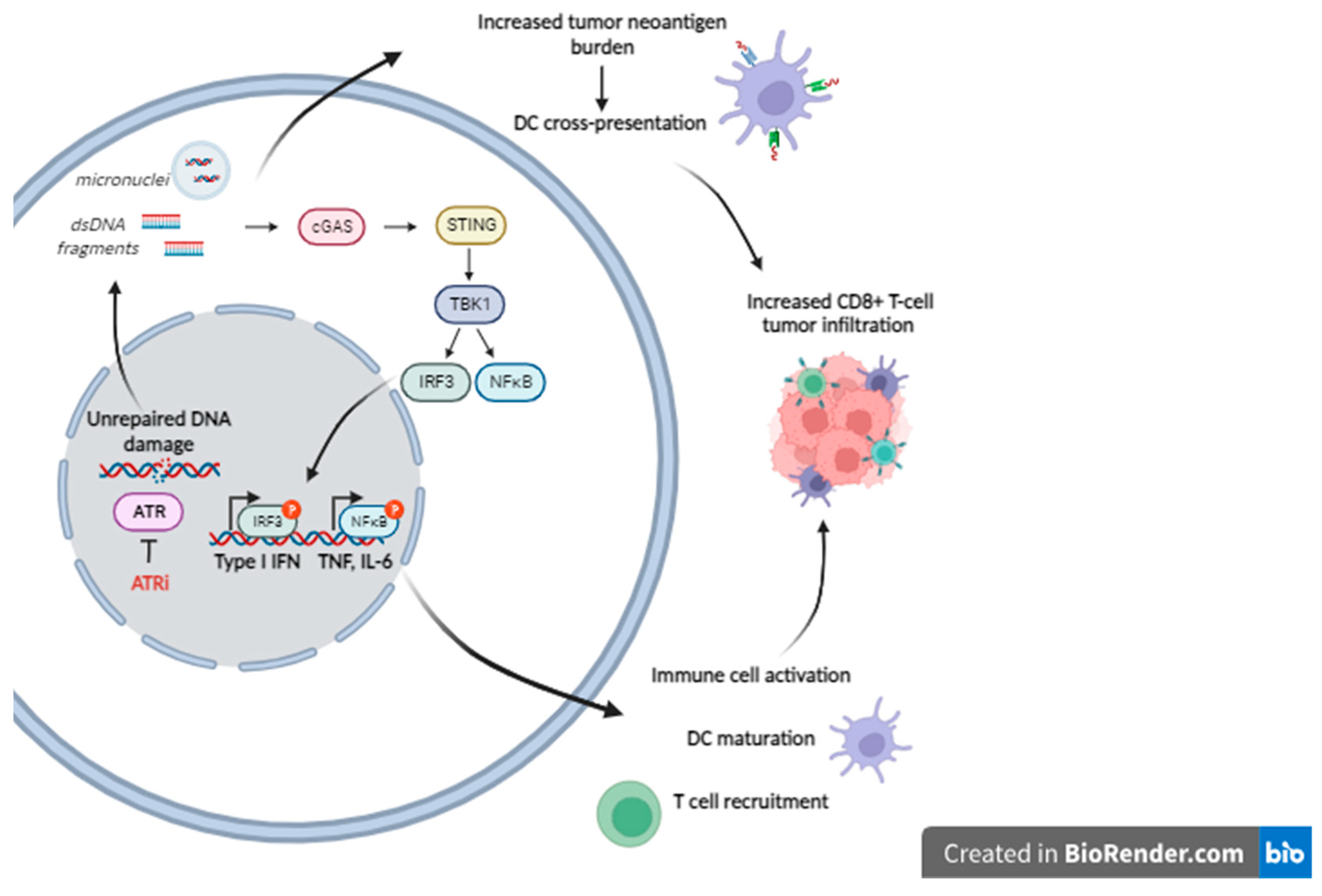

Table 1.
Clinical trials of Ceralasertib (AZD6738) in patients with solid tumors.
| NCT Number | Study Status | Conditions | Interventions | Primary Outcome | Phase | Enrollment | Completion Date |
|---|---|---|---|---|---|---|---|
| NCT03330847 | Active, not recruiting | mTNBC | Ceralasertib + Olaparib | PFS | 2 | 273 | Sep-24 |
| NCT03801369 | Recruiting | mTNBC | Ceralasertib + Olaparib | ORR | 2 | 132 | Dec-27 |
| NCT03740893 | Recruiting | Operable TNBC | Ceralasertib | Biomarker | 2 | 81 | Dec-25 |
| NCT03182634 | Completed | mBC | Ceralasertib + Olaparib | ORR | 2 | 70 | Nov-23 |
| NCT04090567 | Recruiting | HER2-, BRCA+ mBC | Ceralasertib + Olaparib | ORR | 2 | 60 | Mar-25 |
| NCT05582538 | Recruiting | mTNBC | Ceralasertib followed by Durvalumab/nab-Paclitaxel | PFS | 2 | 37 | Nov-25 |
| NCT05450692 | Recruiting | mNSCLC | Ceralasertib + Durvalumab | OS | 3 | 580 | May-25 |
| NCT03334617 | Active, not recruiting | mNSCLC | Ceralasertib, Ceralasertib + Durvalumab | 12-week ORR | 2 | 531 | Sep-24 |
| NCT02664935 | Active, not recruiting | mNSCLC | Ceralasertib + Durvalumab | ORR, PFS, 24-week DCR | 2 | 423 | Sep-23 |
| NCT03833440 | Recruiting | mNSCLC | Ceralasertib + Durvalumab | 12-week DCR | 2 | 120 | Feb-24 |
| NCT05941897 | Recruiting | mNSCLC | Ceralasertib + Durvalumab | ORR | 2 | 38 | Jun-25 |
| NCT02937818 | Active, not recruiting | ES-SCLC | Ceralasertib + Olaparib | ORR | 2 | 72 | Dec-23 |
| NCT04361825 | Active, not recruiting | ES-SCLC | Ceralasertib + Durvalumab | ORR | 2 | 45 | Dec-23 |
| NCT04699838 | Recruiting | ES-SCLC | Platinum-Etoposide-Durvalumab + maintenance Ceralasertib/Durvalumab | PFS | 2 | 30 | May-24 |
| NCT03428607 | Completed | ES-SCLC | Ceralasertib + Olaparib | ORR | 2 | 26 | Jan-21 |
| NCT03579316 | Recruiting | Ovarian Cancer | Ceralasertib + Olaparib | ORR | 2 | 104 | Dec-24 |
| NCT04239014 | Withdrawn | Ovarian Cancer | Ceralasertib + Olaparib | PFS | 2 | 0 | Jan-21 |
| NCT04065269 | Active, not recruiting | Gynaecological Cancers | Ceralasertib + Olaparib | ORR | 2 | 168 | Mar-23 |
| NCT05061134 | Active, not recruiting | Melanoma | Ceralasertib, Ceralasertib + Durvalumab | ORR | 2 | 186 | Apr-24 |
| NCT03780608 | Active, not recruiting | Melanoma, Gastric cancer | Ceralasertib + Durvalumab | ORR | 2 | 61 | Dec-23 |
| NCT04298021 | Active, not recruiting | Biliary Tract Cancer | Ceralasertib + Durvalumab, Ceralasertib + Olaparib | DCR | 2 | 74 | Dec-24 |
| NCT04298008 | Recruiting | Biliary Tract Cancer | Ceralasertib + Durvalumab | DCR | 2 | 26 | Dec-24 |
| NCT04417062 | Recruiting | Osteosarcoma | Ceralasertib + Olaparib | 4-month EFS | 2 | 63 | Jun-25 |
| NCT03787680 | Active, not recruiting | Prostate Cancer | Ceralasertib + Olaparib | ORR | 2 | 49 | Jan-27 |
| NCT03022409 | Completed | HNSCC | Ceralasertib | Biomarker | 1 | 21 | Jan-21 |
| NCT04704661 | Recruiting | HER2+ GEJ/CRC | Ceralasertib + T-DXd | Toxicity | 1 | 15 | Mar-26 |
| NCT02264678 | Recruiting | Advanced Solid Tumors | Ceralasertib + Olaparib | Toxicity | 1/2 | 466 | Jul-26 |
| NCT03682289 | Recruiting | Advanced Solid Tumors | Ceralasertib, Ceralasertib + Olaparib | ORR | 2 | 89 | Jul-25 |
| NCT02223923 | Active, not recruiting | Advanced Solid Tumors | Ceralasertib | MTD | 1 | 87 | Dec-23 |
| NCT02576444 | Terminated | Advanced Solid Tumors | Ceralasertib + Olaparib | ORR | 2 | 67 | Nov-19 |
| NCT02630199 | Completed | Advanced Solid Tumors | Ceralasertib + Paclitaxel | Toxicity, MTD | 1 | 65 | Apr-21 |
| NCT03669601 | Recruiting | Advanced Solid Tumors | Ceralasertib + Gemcitabine | DLT | 1 | 55 | Sep-24 |
| NCT04564027 | Active, not recruiting | Advanced Solid Tumors | Ceralasertib | ORR | 2 | 54 | Feb-24 |
| NCT05514132 | Active, not recruiting | Advanced Solid Tumors | Ceralasertib + Olaparib | DLT | 1 | 14 | Apr-25 |
| NCT05469919 | Active, not recruiting | Advanced Solid Tumors | Ceralasertib | DLT | 1 | 12 | Dec-24 |
| NCT03878095 | Suspended | IDH1/2mut Advanced Solid Tumors | Ceralasertib + Olaparib | ORR | 2 | 50 | Mar-24 |
| NCT03330847 | Active, not recruiting | mTNBC | Ceralasertib + Olaparib | PFS | 2 | 273 | Sep-24 |
| NCT03801369 | Recruiting | mTNBC | Ceralasertib + Olaparib | ORR | 2 | 132 | Dec-27 |
| NCT03740893 | Recruiting | Operable TNBC | Ceralasertib | Biomarker | 2 | 81 | Dec-25 |
| NCT03182634 | Completed | mBC | Ceralasertib + Olaparib | ORR | 2 | 70 | Nov-23 |
| NCT04090567 | Recruiting | HER2-, BRCA+ mBC | Ceralasertib + Olaparib | ORR | 2 | 60 | Mar-25 |
| NCT05582538 | Recruiting | mTNBC | Ceralasertib followed by Durvalumab/nab-Paclitaxel | PFS | 2 | 37 | Nov-25 |
| NCT05450692 | Recruiting | mNSCLC | Ceralasertib + Durvalumab | OS | 3 | 580 | May-25 |
| NCT03334617 | Active, not recruiting | mNSCLC | Ceralasertib, Ceralasertib + Durvalumab | 12-week ORR | 2 | 531 | Sep-24 |
* Abbreviations: CRC, colorectal carcinoma; DCR, disease control rate; DLT, dose-limiting toxicity; EFS, event-free survival; ES-SCLC, extensive-stage small cell lung cancer; GEJ, gastroesophageal junction; HNSCC, head and neck squamous cell carcinoma; IDH1/2mut, isocitrate dehydrogenase 1/2-mutated; mBC, metastatic breast cancer; mNSCLC, metastatic non-small cell lung cancer; MTD, maximum tolerated dose; mTNBC, metastatic triple negative breast cancer; ORR, overall response rate; OS, overall survival; PFS, progression-free survival; T-DXd, trastuzumabderuxtecan; TNBC, triple negative breast cancer; trAE, treatment-related adverse events.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



