Submitted:
08 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
Three new species of genus Caridina are described from the northernmost part of Australia. Caridina darwin n. sp. resembles Caridina temasek Choy & Ng, 1991 from Singapore but differs in the armature of the rostrum, development of epipods on pereiopods and in lacking an appendix interna on male´s first pleopods. Caridina magnovis n. sp. resembles Caridina serratirostris de Man, 1892 but differs from this species in the armature of the ventral margin of the rostrum, a shorter stylocerite, a stouter carpus of first pereiopod, number and size of spiniform setae on third and fifth pereiopods, the shape of the preanal carina and in egg size. Caridina wilsoni n. sp. resembles Caridina gracilirostris de Man, 1892 but differs in egg size and some length to wide ratio of segments of the pereiopods. Detailed morphological descriptions of all three new species are given. A molecular phylogeny (mt DNA 16S) supports the morphospecies hypotheses and illustrates the phylogenetic relationship with morphologically similar species from outside Australia.
Keywords:
Crustacea
; Decapoda
; Atyidae
; Australia
1. Introduction
In 2007, Page et al. investigated the phylogenetic and biogeographic relationships of Australian atyid freshwater shrimps. They found several genetically distinct and putatively new species of the genus Caridina H. Milne Edwards, 1837 [1]. Subsequently, this material was also included in a paper on the molecular and conservation biogeography of Caridina shrimps from northwestern Australia [2].
This study investigates the morphological examination of some of their material. It confirms that the genetically distinct clades are distinct species. Three species are described in detail below.
2. Materials and Methods
All samples were obtained using dip and hand nets and preserved using 96% ethanol. Specimens were subsequently transferred into 70–80% ethanol for long-term preservation. A part of the preserved specimens was dissected and morphometric data were taken using a BMS 143 Trino Zoom dissecting microscope with an ocular grid. Details on setae and mouthparts were observed using a Reichert Biovar compound microscope. Rostral characters were taken from all examined specimens. Drawings were made from microphotographs using Adobe Illustrator following Coleman [3,4]. The setae terminology used mostly follows Short [5].
All material examined is deposited in museum wet collections: Museum and Art Gallery of the Northern Territory, Darwin (NTM), Western Australian Museum, Perth (WAM), Queensland Museum, Brisbane (QM) and the Museum für Naturkunde (Museum of Natural History), Berlin, Germany (ZMB). The following abbreviations are used: cl = carapace length (measured from the postorbital margin to the posterior margin of the carapace); ov. = ovigerous; juv. = juveniles; e.g. = for example.
DNA was extracted from abdominal tissue using a Qiagen Blood and Tissue Kit according to the manufacturer's instructions. A fragment of the mitochondrial 16S rRNA (16S, ~ 590 bp) gene was amplified by polymerase chain reaction (PCR) and sequenced using primers 16S-F-Car and 16S-R-Car1 [6]. Amplifications were conducted in 25 µL volumes containing 50–100 ng DNA, 1x PCR buffer, 200 mM of each dNTP, 0.5 mM of each primer, 2 mM MgCl2 and 1 U of Taq polymerase. After an initial denaturation step of 3 min at 94 °C, 35 cycles of 30 sec at 94 °C, 60 sec at 50°C and 60 sec at 72 °C were performed, followed by a final extension step of 5 min at 72 °C. PCR products were sent to Macrogen Europe for purification and sequencing of both strands of the amplified gene fragments using the primers as given above. Contigs of forward and reverse strands were assembled using Geneious Prime (v. 2019.2.1) and corrected by eye. Sequences were aligned with MAFFT [7]. To determine the best substitution model for Bayesian inference analyses (see below), hierarchical likelihood ratio tests were carried out with jModelTest (Posada 2008). Based on the Akaike Inference Criterion (AIC), the GTR + I + G model was chosen. The new sequences (n=22) have been deposited in GenBank (for accession numbers and museum voucher numbers see Table 1). Additionally, published sequences, notably twelve sequences of the three new species published by von Page et al. [1], Cook et al. [2], Short et al. [9] and de Mazancourt et al. [10], have been included in the analysis (Table1). Phylogenetic trees were reconstructed by Bayesian inference (BI) [11] using MrBayes 3.2.6 [12]. The MCMCMC-algorithm was run with four independent chains for 10,000,000 generations, samplefreq = 500, and burnin = 25%. Maximum likelihood analyses were run with IQ-TREE [13] and branch support was obtained through the implemented ultrafast bootstrap (1,000 replicates) [14]. BI and ML analyses were run with the model specified above (for IQ-TREE, see [15]). In addition, Maximum Parsimony (MP) analyses were performed using the heuristic search algorithm as implemented in PAUP* [16], with gaps treated as fifth base. Support for nodes was estimated by bootstrap analysis (1,000 bootstrap replicates with 10 random addition replicates each).
3. Results
Family ATYIDAE De Haan, 1849
Genus CaridinaH. Milne Edwards, 1837
Caridina darwinn.sp.
Caridina sp. ´NT2` Wilson, 2008
Caridina ‘sp. NT2’ Short, Page & Humphrey, 2019
Material Examined
Holotype. ♂ cl 2.1 mm, Australia, New Territory, Finnis River catchment, Howard Springs, leg. S. Choy, 29.06.1992, NTM Cr019569.
Paratypes. 1 ♂ cl 2.4 mm, same data as holotype, NTM Cr019570; 2 ♀♀ cl 3.2 and 3.7 mm, 1 specimens lacking the abdomen cl 1.8 mm, Australia, New Territory, Darwin area, Blackmore River, cultured specimens provided by D. Wilson NTM Cr019571; 2 ov. ♀♀ cl 2.1 and 2.3 mm, Australia, Northern Territory, Howard Springs, S 12° 27.345' E 131° 3.146', leg. M. Glaubrecht, N. Brinkmann, T. von Rintelen 27.06.2004, ZMB 29211.
Comparative Material Examined
Paratypes. Caridina temasek Choy & Ng, 1991. 5 specimens, Singapore, freshwater stream near Sime road, MacRitchie catchment area, ca. N 1° 20.233' E 103°48.783' leg. P.K.L. Ng and K.K.P. Lim Aug. 1990.
Other material. Caridina temasek Choy & Ng, 1991. 2 ♂♂ cl 2.1 and 2.3 mm, 2 ♀♀ cl 2.8 and 3.7 mm Singapore, Central catchment, N 1° 21.32' E 103° 48.279', leg. W. Klotz 08.12.2011, ZMB 29681.
Caridina excavatoides Johnson, 1961. 3 ♀♀ cl 2.3–3.2 mm, Indonesia, Sumatra, Jambi, Tributary of Batang Hari River, Danau Lamo Village, S 1° 28.003' E 103° 38.262', leg. M. de Bruyn 01.01.2009 ZMB 29583.
Description
Cephalothorax and cephalic appendages. Small shrimp with carapace length 1.8–3.7 mm (median 2.3 mm, n = 7) in apparently adult specimens. Carapace (Figure 1A,B) smooth, antennal spine well separated from orbital margin, pterygostomial angle broadly rounded. Rostrum (Figure 1A,B) pointed, stright or slightly sigmoidal, dorsal margin armed almost to tip of rostum with 15–21 teeth, teeth above eye larger than distal and proximal, ventral margin with 2–4 large teeth, rostrum reaching to midlenght of second segment of antennular peduncle or to distal end of antennular peduncle, 0.70–0.85 (median 0.71, n = 5) times as long as carapace. Rostrum formula based on 6 specimens 4–5(4)+11–17/2–4. Eyes (Figure 1A,B) well developed with pigmented globular cornea. Antennula (Figure 1G) and antenna (Figure 1H) well developed. Antennular peduncle (Figure 1A,B,G), 0.86–0.93 (median 0.92, n = 4) times as long as carapace in females, 1.0 times as long as carapace in male, first segment 1.69–1.92 (median 1.81, n = 5) times as long as second segment, second segment 1.86–2.67 (median 1.88, n = 5) times longer than third segment; anterior-lateral tooth on first segment well developed, acute, 0.15–0.38 (median 0.20, n = 5) times as longs as second segment. Stylocerite pointed, reaching to 0.72–0.93 (median 0.80, n = 5) times length of basal segment of antennular peduncle. Scaphocerite (Figure 1H) with well-developed distolateral tooth, 3.86–4.0 (median 3.80, n = 3) times as long as wide.
Abdominal somites, telson and uropods. Sixth abdominal somite 0.60–0.67 (median 0.63, n = 4) times length of carapace, 1.63–1.83 (median 1.72, n = 4) times as long as fifth somite, 0.87–1.0 (median 0.92, n = 4) times as long as telson. Telson (Figure 1C,D) slightly tapering distally, distal margin strongly convex to triangular, lacking a median point, lateral angles not overreaching distal margin, with 3–4 (n = 3) pairs of dorsal short spiniform setae and one pair of short spiniform setae dorsolateral; distal margin with 6–8 (n = 5) strong spiniform setae, lateral pair longest. Preanal carina (Figure 1E) rounded, low. Uropodal diaeresis (Figure 1F) with 10–13 movable spiniform setae, outermost one shorter than lateral angle.
Mouthparts and branchiae. Mandibles (Figure 1I) without palp, incisor process ending in irregular teeth, molar process truncated. Lower lacinia of maxillula (Figure 1J) broadly rounded with long pappose setae and few rows of serrate setae near margin and scattered cuspidate setae proximal, upper lacinia elongate, with numerous distinct cuspidate setae flanked by a row of serrate setae on inner margin and few pappose setae subterminal, palp slender with few simple setae and one conical spiniform seta near tip. Upper endites of maxilla (Figure 1K) subdivided, furnished with numerous pappose, simple and papposerrate setae arranged in rows, palp slender, scaphognathite tapering posteriorly, fringed with long, curved serrulate setae at truncated posterior margin and plumose setae inferior. Palp of first maxilliped (Figure 1L,M) ending in a finger-like extension, caridean lobe and flagellum with plumose setae, basis with numerous pappose, papposerrate and serrate setae arranged in rows and tufts. Podobranch on second maxilliped (Figure 1N) well developed; exopod slender, overreaching propodal segment, with long plumose setae distally; basis and merus with rows of pappose setae, distolateral margin of penultimate segment with several pappose and serrate setae; dactylus with numerous serrate setae flanked by a row of pappose setae. Third maxilliped (Figure 2A) slender, with two arthrobranches; exopod slender, overreaching the distal margin of antepenultimate segment, with few long plumose setae distally; antepenultimate segment of endopod slender, with few simple setae basally; penultimate segment slender, with straight lateral margins, with few simple setae distally, inner margin with rows of short serrate setae; ultimate segment slightly shorter than penultimate segment, tapering distally, ending in a claw-like curved spiniform setae (nail), proximal to the nail about few cuspidate setae; inner margin dense beset with rows of serrated setae; epipod well developed, with distal hook. Branchial apparatus with two arthrobranches on third maxilliped and one arthrobranch on first pereiopod, podobranch on second maxilliped well developed. Pleurobranches present on all pereiopods. Well-developed (with distal hooks) epipods present on third maxilliped and first pereiopod. Vestigial epipods (only visible after clearing and dying appendages) on second and third pereiopod. One setobranch on first pereiopod.
Pereiopods. First and second pereiopods chelate. Chela of first pereiopod (Figure 2B) caridinoid in shape, slender, with well-developed palm, 2.44–2.90 (median 2.67, n = 2) times as long as wide, 1.30–1.32 (median 1.31, n = 2) times as long as carpus; tips of fingers rounded, without hooks; dactylus 3.67–4.00 (median 3.83, n = 2) times as long as wide, 1.22–1.44 (median 1.33, n = 2) times as long as palm, with tufts of setae distally; carpus slender, hardly excavated distally, 2.73–2.75 (median 2.74, n = 2) times as long as wide, 1.29–1.36 (median 1.33, n = 2) times as long as merus. Merus not inflated, 2.44–2.62 (median 2.53, n = 2) times as long as wide, shorter than ischium. Chela of second pereiopod (Figure 2C) very slender, caridinoid in shape, with well-developed palm, 3.65–3.75 (median 3.70, n = 2) times as long as wide, 0.73–0.84 (median 0.78, n = 2) times as long as carpus; tips of fingers rounded, without hooks, dactylus 6.17–6.67 (median 6.42, n = 2) times as long as wide, 1.42–1.67 (median 1.54, n = 2) times as long as palm, with tufts of setae distally; carpus slender, 6.17–6.63 (median 6.50, n = 2) times as long as wide, 1.64–1.67 (median 1.70, n = 2) times as long as merus. Merus not inflated, 3.82–4.55 (median 4.18, n = 2) times as long as wide, shorter than ischium. Third pereiopod (Figure 2D,E) with dactylus (Figure 2E) 5.00–5.33 (median 5.17, n = 2) times as long as wide (terminal claw and spiniform setae on flexor margin included), terminating in one claw, with 5 or 6 accessory spiniform setae on flexor margin; propodus not dilated or curved distally, 12.0 (n = 2) times as long as wide, 3.0 (n = 2) times as long as dactylus; carpus 4.33–4.71 (median 4.52, n = 2) times as long as wide, 0.54–0.55 (median 0.55, n = 2) times as long as propodus, 0.45–0.49 (median 0.47, n = 2) times as long as merus, with one large spiniform setae near distal margin and 3 small spiniform setae proximal; merus 7.07–9.73 (median 8.40, n = 2) times as long as wide, 2.04–2.21 (median 2.13, n = 2) times as long as carpus, bearing 3 or 4 strong movable spiniform setae on externo-inferior margin; ischium with one spiniform setae. Fourth pereiopod similar to third but slightly smaller. Fifth pereiopod (Figure 2F,G) slender; dactylus (Figure 2G) 4.0–5.50 (median 4.75, n = 2) times as long as wide (terminal claw and spiniform setae on flexor margin included), terminating in one large claw, with 21–49 spiniform setae on flexor margin; propodus with small spiniform setae on ventral margin, distal pair of spiniform setae not enlarged, propodus 12.50–16.40 (median 14.45, n = 2) times as long as wide, 2.27–4.10 (median 3.19, n = 2) times as long as dactylus; carpus 5.0–7.83 (median 6.42, n = 2) times as long as wide, 0.50–0.57 (median 0.54, n = 2) times as long as propodus, 0.61–0.67 (median 0.64, n = 2) times as long as merus, with 1 strong spiniform setae near distal margin and 2–5 small spiniform setae proximal; merus 9.11–9.33 (median 5.22, n = 2) times as long as wide, 1.49–1.64 (median 1.56, n = 2) times as long as carpus, bearing 3 strong appressed movable spiniform setae on externo-inferior margin. Ischium without or with one spiniform seta.
Pleopods. Endopod of male´s first pleopod (Figure 2H) leaf-like, lacking an appendix interna, 3.2 times as long as proximal width, 0.38 times as long as exopod, with long plumose setae on outer and distal margin, with few pappose setae on inner margin. Appendix masculina on male second pleopod (Figure 2I) stick-like, 8.3 (n = 1) times as long as wide, 0.63 (n = 1) times as long as endopod, armed with strong spiniform setae on inner margin and distal margin; appendix interna arising from about 0.4 times length of appendix masculina, reaching to about 0.75 times lenght of appendix masculina.
Sexual dimorphism. Males similar to females except size, relative length of antennular peduncle and sexual appendages. The main morphological differences from females lead in the structure of the endopod of first pleopod as well as the presence of an appendix masculina on second pleopod.
Reproductive biology. Ovigerous females with medium-sized eggs; size of undeveloped eggs (without eyes) 0.59–0.64 x 0.37–0.39 mm.
Size. Postorbital carapace length of apparently adult specimens 1.8–3.7 mm.
Colouration. Transparent to brownish with cream transversal stripes on abdomen (Wilson 2008; Short, Page and Humphrey 2019).
Distribution. The species occurs in rivers around Darwin, Northern Territories, Australia.
Etymology. The species name darwin was derived from the Darwin river catchment, the natural habitat of the proposed new species and is used as a noun in apposition.
Common name. Blackmore River Caridina.
Remarks. Based on morphology, Caridina darwin n. sp. is related to a group of species including Caridina propinqua de Man, 1908 and Caridina laevis Heller, 1862 as most well-known members. Within this group two species with large eggs, Caridina temasek Choy & Ng, 1991 and Caridina excavatoides Johnson, 1961, both inhabiting freshwaters of the Malayan peninsula, are most closely related to the proposed new species. Caridina darwin n. sp. differs from C. temasek in a higher number of teeth on dorsal margin of the rostrum (15–21 vs. 14–16 in C. temasek), the development of epipods (well developed (with distal hooks) on third maxilliped and first pereiopod, vestigial arthropod (only visible after clearing and dying appendages) on second and third pereiopod vs. well developed (with distal hooks) epipods on third maxilliped and first and second pereiopod, reduced epipod on third and vestigial epipod on fourth pereiopod in C. temasek). Further the proposed new species is different from C. temasek by lacking an appendix interna on first pleopod of males (vs. with a well developed epipod in C. temasek and smaller egg size (0.59–0.64 x 0.37–0.39 mm vs. 0.70–0.80 x 0.44–0.54 mm in C. temasek according to the original description). There is still some confusion about the shape of the preanal carina in C. temasek. Choi & Ng [17] did not give a drawing of this part but in the verbal description thy wrote “Pre-anal carina with about 10 setae” suggesting that they did not observe a distinct hook-like spine. Cai, Ng and Choy in their paper on Atyidae from peninsular Malaysia and Singapore [18] wrote that the preanal carina in C. temasek is without or with a spine. 4 of 5 paratypes of C. temasek observed for the present study are showing a prominent hook-like spine on the preanal carina like all 4 topotypical specimens from Singapore listed above. The holotype of C. temasek was not available for the present study. It seems likely that specimens assigned to C. temasek with and without a hook-like spine on the preanal carina might belong to different species. All specimens of Caridina darwin n. sp. are lacking a spine on the preanal carina.
Caridina darwin n. sp. is also similar to Caridina excavatoides Johnson, 1961. Both species do not bear an appendix interna on endopod of first pleopod in males. The proposed new species is different from C. excavatoides by a longer sixth abdominal segment (0.60–0.67 times as long as carapace vs. less than 0.6 times as long as carapace in C. excavatoides). Further, Caridina darwin n. sp. Is different from this species by the shape of the palp of first maxilliped (with a distinct finger-like extension vs. ending in a triangular extension in C. excavatoides). In development of epipods on third maxilliped and pereiopods Caridina darwin n. sp. differs from C. excavatoides in the same way as from C. temasek. The dactylus of fifth pereiopod bears a smaller number of spiniform setae on flexor margin (21–49 vs. 50–60 in C. excavatoides).
Caridinamagnovis n.sp.
Caridina sp. WA 4 Page, von Rintelen & Hughes 2007
Caridina sp. ´WA 4` Wilson, 2008
Caridina sp. WA 4 Cook, Page & Hughes 2011
Caridina ‘sp. WA4’ Short, Page & Humphrey, 2019
Material Examined
Holotype. ♂ cl 2.9 mm, Australia, Western Australia, Fitzroy River catchment, Calder Yard, FIT12-6, leg. M. Scanlon 29.04.1998, WAM C81392.
Paratypes. 1 ♂ cl 2.6 mm, 1 ♀ cl 3.7 mm, same data as holotype WAM C81393; 1 ♂ cl 2.7 mm, Australia, Western Australia, Ord River catchment, Mantinea Flats, leg. M. Scanlon 18.09.1998, WAM C81394; 2 ov. ♀♀ cl 4.0 and 4.3 mm, 1 ♀ cl 3.7 mm, Darwin Area, Howard River, Gunn Pt. Rd Xing, cultured specimens provided by D. Wilson NTM Cr019573; 3 ♀♀ cl 1.8–2.2 mm, 2 ♂♂ cl 2.3 and 2.4 mm, Fitzroy River catchment, Dimond Gorge, leg. M. Scanlon 14.04.1998 ZMB 33101; 1 ♂ cl 3.0 mm, Darwin Area, Blackmore River, cultured specimens provided by D. Wilson ZMB 33112; 1♀ cl 2.1 mm, 1 ov. ♀ cl. 3.1 mm, 4 juv. cl 1.6–1.8 mm, Australia, Northern Territory, Top End, Coomalie Creek, S 13°0.884´, E 131°7.371´, 85-04, leg. N. Brinkmann und T. von Rintelen 28.06.2004, ZMB 29152; 2 ♀♀ cl 2.5 und 2.7 mm, 2 ov. ♀♀ cl 2.8 und 2.9 mm, Australia, Northern Territory, Howard Springs, S 12° 27.345' E 131° 3.146', 79-04, leg. M. Glaubrecht, N. Brinkmann, T. von Rintelen 27.06.2004, ZMB 32120.
Comparative Material Examined
Lectotype. Caridina serratirostris de Man, 1892. Ov. ♀ cl 5.0 mm, Bangkalanvir River, Saleyer, RMNH.CRUS.D.57023. Syntypes. Caridina serratirostris de Man, 1892. 2 ov. ♀♀ cl 4.4 and 4.6 mm, 2 ♂♂ cl 3.1 and 3.3 mm, Bangkalanvir River, Saleyer, ZMA.CRUS.D.102625. 4 ov. ♀♀ cl 3.3–4.4 mm, 2 ♀♀ cl 2.4 and 3.5 mm, 2 ♂♂ cl 2.4 and 2.6 mm, Indonesia, Rivier bij Reo, Flores eil. coll. 1888.
Description
Cephalothorax and cephalic appendages. Small shrimp with carapace length 2.1–4.3 mm (median 2.9 mm, n = 16) in apparently adult specimens. Carapace (Figure 3A,B) smooth, antennal spine almost fused with the inferior orbital angle, pterygostomial angle rounded. Rostrum (Figure 3A,B) pointed, stright or slightly convex, dorsal margin armed to tip of rostum with 17–29 teeth, teeth above eye larger than distal and ventral, ventral margin with 3–6 teeth placed in the distal half of the rostrum, reaching to end of second segment of antennular peduncle or to distal end of antennular peduncle, 0.53–0.76 (median 0.64, n = 8) times as long as carapace. Rostrum formula based on 12 specimens 5–8+12–24/3–6. Eyes (Figure 3A,B) well developed with pigmented globular cornea. Antennula (Figure 3G) and antenna (Figure 2H) well developed. Antennular peduncle (Figure 3A,B,G), 0.60–0.71 (median 0.67, n = 9) times as long as carapace in females, 0.91 times as long as carapace in male, first segment 1.69–2.10 (median 2.0, n = 9) times as long as second segment, second segment 2.20–2.75 (median 2.31, n = 8) times longer than third segment; antero-lateral tooth on first segment short (0.14–0.23 times as long as second segment), well developed. Stylocerite pointed, reaching to 0.89–1.05 (median 1.00, n = 9) times length of basal segment of antennular peduncle. Scaphocerite (Figure 3H) with well-developed distolateral tooth, 2.92–3.7 (median 3.14, n = 5) times as long as wide.
Abdominal somites, telson and uropods. Sixth abdominal somite 0.44–0.54 (median 0.46, n = 7) times length of carapace, 1.23–1.69 (median 1.55, n = 6) times as long as fifth somite, 0.80–0.89 (median 0.87, n = 5) times as long as telson. Telson (Figure 3C,D) slightly tapering distally, distal margin convex, with a prominent median point, lateral angles not overreaching distal margin, with 4–5 (n = 7) pairs of dorsal short spiniform setae and one pair of short spiniform setae dorsolateral; distal margin with one pair of strong strong spiniform setae and 7–9 plumose setae clearly overreaching lateral pair of spiniform setae. Preanal carina (Figure 3E) rounded, high, with a small tooth in 2 of 7 specimens. Uropodal diaeresis (Figure 3F) with 12–15 movable spiniform setae, outermost one shorter than lateral angle.
Mouthparts and branchiae. Mandible (Figure 3I,J) without palp, incisor process ending in comparably well developed and numerous irregular teeth, molar process truncated. Lower lacinia of maxillula (Figure 3K) broadly rounded with long pappose setae and few rows of serrate setae near margin and scattered cuspidate setae proximal, upper lacinia elongate, with numerous distinct cuspidate setae flanked by a row of serrate setae on inner margin and few papose setae subterminal, palp slender with few simple setae and one conical spiniform seta near tip. Upper endites of maxilla (Figure 3L) subdivided, furnished with numerous pappose, simple and papposerrate setae arranged in rows, palp slender, scaphognathite tapering posteriorly, fringed with long, curved serrulate setae at truncated posterior margin and plumose setae inferior. Palp of first maxilliped (Figure 3M,N) ending in a triangular extension, caridean lobe narrow, this part and flagellum with plumose setae, basis with numerous pappose, papposerrate and serrate setae arranged in rows and tufts. Podobranch on second maxilliped (Figure 3O) well developed; exopod slender, overreaching propodal segment, with long plumose setae distally; basis and merus with rows of papose setae, distolateral margin of penultimate segment with several pappose and serrate setae; dactylus with numerous serrate setae flanked by a row of pappose setae. Third maxilliped (Figure 4A) slender, with one well developed arthrobranch, second one reduced to a small stick-like structure; exopod slender, overreaching the distal margin of antepenultimate segment, with few long plumose setae distally; antepenultimate segment of endopod slender, with few simple setae basally; penultimate segment slender, with straight lateral margins, with few simple setae distally, inner margin with rows of short serrate setae; ultimate segment slightly shorter than penultimate segment, tapering distally, ending in a claw-like curved spiniform setae (nail), proximal to the nail about few cuspidate setae; inner margin dense beset with rows of serrated setae; epipod well developed, with distal hook. Branchial apparatus with one well developed and one reduced arthropods on third maxilliped and one arthropod on first periopod, podobranch on second maxilliped well developed. Pleurobranchs present on all pereiopods. Well-developed (with distal hooks) epipods present on third maxilliped and first to fourth pereiopod. Two setobranch on first pereiopod, one setobranch on each posterior pereiopods.
Pereiopods. First and second pereiopods chelate. Chela of first pereiopod (Figure 4B) caridinoid in shape, slender, with well-developed palm, 1.75–2.84 (median 2.21, n = 5) times as long as wide, 0.93–1.17 (median 1.12, n = 5) times as long as carpus; tips of fingers rounded, without hooks; dactylus 2.73–4.25 (median 3.57, n = 5) times as long as wide, 1.00–1.64 (median 1.25, n = 5) times as long as palm, with tufts of setae distally; carpus slender, hardly excavated distally, 2.12–3.22 (median 2.78, n = 5) times as long as wide, 1.09–1.25 (median 1.17, n = 5) times as long as merus. Merus not inflated, 2.75–4.33 (median 3.00, n = 0) times as long as wide, 0.95–1.18 (median 1.0, n = 5) times as long as ischium. Chela of second pereiopod (Figure 4C) very slender, caridinoid in shape, with well-developed palm, 2.75–4.40 (median 3.30, n = 5) times as long as wide, 0.66–0.71 (median 0.69, n = 5) times as long as carpus; tips of fingers rounded, without hooks, dactylus 4.44–5.45 (median 5.00, n = 5) times as long as wide, 1.25–1.36 (median 1.33, n = 5) times as long as palm, with tufts of setae distally; carpus slender, 6.36–8.17 (median 7.14, n = 5) times as long as wide, 1.04–1.40 (median 1.35, n = 5) times as long as merus. Merus not inflated, 5.83–7.83 (median 6.24, n = 5) times as long as wide, 1.06–1.17 (median1.13, n = 4) times as long as ischium. Third pereiopod (Figure 4D–G) with dactylus (Figure 4E–G) slightly sexually dimorph, spiniform setae on flexor margin more curved, 3.78 times as long as wide (terminal claw and spiniform setae on flexor margin included) in male (n = 1), 4.00–5.00 (median 4.56, n = 4) times as long as wide in females, terminating in one claw, with 6–8 accessory spiniform setae on flexor margin, first spiniform setae on flexor margin strong and curved in some specimens; propodus not dilated or curved distally, distal spiniform setae not enlarged, 10.6–14.29 (median 12.50, n = 5) times as long as wide, 3.24–3.73 (median 3.33, n = 5) times as long as dactylus; carpus 5.67–6.80 (median 5.86, n = 4) times as long as wide, 0.63–0.70 (median 0.66, n = 4) times as long as propodus, 0.50–0.59 (median 0.57, n = 4) times as long as merus, with one large spiniform setae near distal margin and 3–6 small spiniform setae proximal; merus 6.84–9.08 (median 8.47, n = 4) times as long as wide, 1.69–2.00 (median 1.74, n =4) times as long as carpus, bearing 3–5 strong movable spiniform setae on externo-inferior margin; ischium without or with one spiniform setae. Fourth pereiopod similar to third but slightly smaller. Fifth pereiopod (Figure 4H,I) slender; dactylus (Figure 4I) 3.6–5.00 (median 4.30, n = 2) times as long as wide (terminal claw and spiniform setae on flexor margin included), terminating in one large claw, with 16–18 spiniform setae on flexor margin; propodus with small spiniform setae on ventral margin, distal pair of spiniform setae not enlarged, propodus 12.40–13.80 (median 12.50, n = 3) times as long as wide, 3.44–3.75 (median 3.60, n = 2) times as long as dactylus; carpus 5.83–7.09 (median 6.62, n = 3) times as long as wide, 0.56–0.57 (median 0.57, n = 3) times as long as propodus, 0.66–0.68 (median 0.67, n = 3) times as long as merus, with 1 strong spiniform setae near distal margin and 5 small spiniform setae proximal; merus 7.43–8.43 (median 8.40, n = 3) times as long as wide, 1.47–1.51 (median 1.49, n = 3) times as long as carpus, bearing 2–4 strong appressed movable spiniform setae on externo-inferior margin. Ischium without or with one spiniform seta.
Pleopods. Endopod of male´s first pleopod (Figure 4J) leaf-like, lacking an appendix interna, 2.0 (n = 1) times as long as proximal width, 0.40 times as long as exopod, with long plumose setae on distal margin, and few shorter plumose setae on inner and outer margin. Appendix masculina on male second pleopod (Figure 4K) stick-like, 10.0 (n = 1) times as long as wide, 0.83 (n = 1) times as long as endopod, armed with strong spiniform setae on inner margin and distal margin; appendix interna arising from about 0.4 times length of appendix masculina, reaching to about 0.65 times length of appendix masculina.
Sexual dimorphism. Males similar to females except size, relative length of antennular peduncle and sexual appendages. The main morphological differences from females lead in the structure of the endopod of first pleopod as well as the presence of an appendix masculina on second pleopod. Third and fourth pereiopods slightly dimorph, dactyli slightly shorter and spiniform setae on dactyli slightly more curved in males.
Reproductive biology. Ovigerous females with medium-sized eggs; size of eggs 0.66–0.76 x 0.39–0.49 mm (n = 6). 25 and 30 eggs counted in two ovigerous specimens.
Size. Postorbital carapace length of apparently adult specimens 2.1–4.3 mm.
Colouration. Variable. Some specimens reddish with a pale median dorsal line reaching from the rostrum to 6th abdominal segment, some specimens mottled with reddish and cream chromatophores, smaller specimens mostly transparent [9,19].
Distribution. The species is known from rivers draining to the Timor Sea. Westwards from the Fitzroy River in Western Australia eastwards to the Howard River in Northern Territory.
Etymology. The species name magnovis derives from the rather large egg size of the proposed new species compared to C. serratirostris.
Common name. North Australian Chameleon Shrimp.
Remarks. The general morphology of Caridina magnovis n.sp. with a straight rostrum, armed throughout close to the tip and a high number of postorbital teeth (5–8), a long stylocerite, slender chelipeds and distal margin of the telson armed with long plumose setae distinctly overreaching a lateral pair of strong spiniform setae and the variable coloration of living specimens resembles that of Caridina serratirostris de Man, 1892. The proposed new species differs from C. serratirostris in the armature of the rostrum, i.e., ventral teeth placed in the distal half of the rostrum vs. ventral teeth placed at midlength of the rostrum in C. serratirostris. The stylocerite of Caridina magnovis n.sp. is shorter, reaching to 0.89–1.05 of first segment of antennular peduncle vs. clearly overreaching this segment (1.09–1.47 times length of first segment). Carpus of first cheliped is 2.12.–3.22 times as long as wide in Caridina magnovis n.sp. vs. 2.63–4.67 times in C. serratirostris. Dactylus of third pereiopod with 6–8 spiniform setae vs. with 4–5 in C. serratirostris. Dactylus of fifth pereiopod with 16–18 spiniform setae on flexor margin, first setae not remarkably enlarged vs. with 11–17 spiniform setae, first setae enlarged in C. serratirostris. Uropodal diaeresis with 12–15 movable spiniform setae vs. with 16–21 movable spiniform setae in C. serratirostris. Preanal carina rounded, without a distinct tooth in most specimens (vs. preanal carina with a prominent hook-like tooth in all specimens of C. serratirostris. Eggs 0.66–0.76 x 0.39–0.49 mm vs. 0.37–0.44 x 0.20–0.26 mm in C. serratirostris.
Caridinawilsoni n.sp.
Caridina sp. Gulf 1 Page, von Rintelen & Hughes, 2007
Caridina sp. ‘Gulf 1’ Wilson, 2008
Caridina sp. Gulf 1 Cook, Page & Hughes, 2011
Caridina ‘sp. Gulf1’ Short, Page & Humphrey, 2019
Material Examined
Holotype. ♂ cl 3.4 mm, Australia, Queensland, Archer River catchment, Archer River at telegraph line, FNARH5/216, leg. DERM, 10.04.2000, QM W29816.
Paratypes. 1ov. ♀ cl 4.3 mm, same data as holotype, QM W29817; ♂ cl 3.0 mm, Australia, Queensland, Wenlock River catchment, Wenlock River at Stones Xing, leg. DERM, 10.04.2000, QM W29818; 1 ♀ cl 3.4 mm, Commercial production ponds (most likely Blackmore R., could be Darwin R.), Darwin Area, cultured specimens provided by D. Wilson NTM Cr019572; 7 ov. ♀♀ cl 3.2–4.0 mm 3 ♂♂ cl 2.4–2.5 mm, Australia, Northern Territory, Top End, Darvin River crossing; S 12°49,089´, E 130°57,634´, 83-04, coll. N. Brinkmann & T. von Rintelen 27.6.2004, ZMB 29180; 2 ♂♂ cl 1.9 and 2.2 mm, 4 ♀♀ cl 2.3–2.7 mm, Australia, Northern Territory, Top End, Coomalie Creek, S 13°0,884´, E 131°7,371´; coll. N. Brinkmann & T. von Rintelen 28.06.2003, ZMB 29240.
Comparative Material Examined
Syntypes. Caridina gracilirostris De Man, 1892. 2 ov. ♀♀ cl 5.2 and 5.3 mm, Indonesia, Celebes (=Sulawesi): river near Balangnipa 1888-1889, leg. M. Weber: RMNH.CRUS.D.2557; 1 ov. ♀ cl 5.3 mm, 2 ♂♂ cl 4.0 and 4.5 mm, Indonesia, Celebes (=Sulawesi), river near Balangnipa 1888-1889, leg. M. Weber, RMNH.CRUS.D.1076.
Other material. Caridina gracillima Lanchester, 1901. 3 specimens, Malaysia, Sabah, Kinabatangan River at Sukau, N 5° 31' 37.67'' E 118° 17' 39.23'', leg. M. de Bruyn 01.01.2009; 2 specimens. Thailand, Phitsanulok, Tributary of Kaek River, c. 28km E of Phitsanulok, N 16° 50.866' E 100° 30.709', leg. F. Köhler 27.03.2006, ZMB 29579.
Description
Cephalothorax and cephalic appendages. Small shrimp with carapace length 1.9–4.3 mm (median 2.8 mm, n = 22) in apparently adult specimens. Carapace (Figure 5A,B) smooth, antennal spine well separated from the inferior orbital angle, pterygostomial angle broadly rounded. Rostrum (Figure 5A,B) long, curved upwards, dorsal margin armed with 7–9 widely spaced teeth at the proximal part of the rostrum, no teeth behind orbital margin (1 tooth in one specimen examined), one subapical tooth near tip, ventral margin with 35 teeth, rostrum well overreaching distal margin of the scaphocerite, 1.04–2.31 (median 1.97, n = 8) times as long as carapace. Rostrum formula based on 18 specimens 0–1+8–10/19–35. Eyes (Figure 5A,B) well developed with pigmented globular cornea. Antennula (Figure 5G) and antenna (Figure 5H) well developed. Antennular peduncle (Figure 5A,B,G), 0.86–1.02 (median 0.91, n = 7) times as long as carapace in females, 0.89–1.08 (median 1.00, n = 6) times as long as carapace in males, first segment 1.85–2.33 (median 2.16, n = 12) times as long as second segment, second segment 1.61–2.75 (median 2.05, n = 12) times longer than third segment; antero-lateral tooth on first segment acute, well-developed, 0.23–0.39 (median 0.33, n = 11) times as long as second segment. Stylocerite pointed, reaching to 0.70–1.04 (median 0.77, n = 11) times length of basal segment of antennular peduncle. Scaphocerite (Figure 5H) with well-developed distolateral tooth, 3.87–4.95 (median 4.66, n = 5) times as long as wide.
Abdominal somites, telson and uropods. Sixth abdominal somite 0.86–1.02 (median 0.92, n = 17) times length of carapace, 1.81–2.41 (median 2.18, n = 17) times as long as fifth somite, 1.08–1.30 (median 1.13, n = 15) times as long as telson. Telson (Figure 5C,D) slightly tapering distally, distal margin broadly convex, lacking a median point, lateral angles not overreaching distal margin, with 3 or 4 (n = 10) pairs of dorsal short spiniform setae and one pair of short spiniform setae dorsolateral; distal margin with two pairs of spiniform setae, lateral pair stronger and longer compared to inner setae. Preanal carina (Figure 5E) with a prominent hook-like tooth. Uropodal diaeresis (Figure 5F) with 5–9 movable spiniform setae, outermost one distinctly shorter than lateral angle.
Mouthparts and branchiae. Mandible (Figure 5I) without palp, incisor process ending in irregular teeth, molar process truncated. Lower lacinia of maxillula (Figure 5J) broadly rounded to subrectangular with long pappose setae and few rows of serrate setae near margin and scattered cuspidate setae proximal, upper lacinia elongate, with numerous distinct cuspidate setae flanked by a row of serrate setae on inner margin and few pappose setae subterminal, palp slender with few simple setae and one conical spiniform seta near tip. Upper endites of maxilla (Figure 5K) subdivided, furnished with numerous pappose, simple and papposerrate setae arranged in rows, palp slender, scaphognathite tapering posteriorly, fringed with long, curved serrulate setae at truncated posterior margin and plumose setae inferior. Palp of first maxilliped (Figure 5L,M) ending in a short blunt extension, caridean lobe narrow, this part and flagellum with plumose setae, basis with numerous pappose, papposerrate and serrate setae arranged in rows and tufts. Podobranch on second maxilliped (Figure 5N) well developed; exopod slender, overreaching propodal segment, with long plumose setae distally; basis and merus with rows of pappose setae, distolateral margin of penultimate segment with several pappose and serrate setae; dactylus with numerous serrate setae flanked by a row of pappose setae. Third maxilliped (Figure 6A) slender, with two well developed arthrobranches, exopod slender, overreaching the distal margin of antepenultimate segment, with few long plumose setae distally; antepenultimate segment of endopod slender, with few simple setae basally; penultimate segment slender, with straight lateral margins, with few simple setae distally, inner margin with rows of short serrate setae; ultimate segment shorter than penultimate segment, tapering distally, ending in a claw-like curved spiniform setae (nail), proximal to the nail about few cuspidate setae; inner margin dense beset with rows of serrated setae; epipod well developed, with distal hook. Branchial apparatus with two arthropods on third maxilliped and one arthropod on first periopod, podobranch on second maxilliped well developed. Pleurobranchs present on all pereiopods. Well-developed (with distal hooks) epipods present on third maxilliped and first to fourth pereiopod. One setobranch on all pereiopods.
Pereiopods. First and second pereiopods chelate. Chela of first pereiopod (Figure 6B) caridinoid in shape, with well-developed palm, 1.75–2.32 (median 2.10, n = 9) times as long as wide, 1.16–1.54 (median 1.41, n = 9) times as long as carpus; tips of fingers rounded, with prominent hooks hidden by tufts of setae; dactylus 3.00–3.80 (median 3.29, n = 7) times as long as wide, 0.80–1.36 (median 1.13, n = 9) times as long as palm; carpus slender, hardly excavated distally, 1.73–2.27 (median 2.00, n = 9) times as long as wide, 0.65–1.27 (median 1.14, n = 9) times as long as merus. Merus not inflated, 2.33–4.58 (median 2.48, n = 9) times as long as wide, 0.96–1.96 (median 1.06, n = 8) times as long as ischium. Chela of second pereiopod (Figure 6C) slender, caridinoid in shape, with well-developed palm, 2.21–2.75 (median 2.47, n = 9) times as long as wide, 0.80–0.94 (median 0.88, n = 9) times as long as carpus; tips of fingers rounded, with prominent hooks hidden by tufts of setae, dactylus 3.80–4.44 (median 4.17, n = 7) times as long as wide, 1.25–1.73 (median 1.33, n = 9) times as long as palm; carpus slender, 3.73–4.94 (median 4.31, n = 9) times as long as wide, 1.17–1.41 (median 1.32, n = 9) times as long as merus. Merus not inflated, 3.50–5.00 (median 4.40, n = 9) times as long as wide, 0.91–1.06 (median 0.97, n = 8) times as long as ischium. Third pereiopod (Figure 6D–E) slender, dactylus (Figure 6E) not sexually dimorph, 2.71–3.75 (median 3.75, n = 3) times as long as wide (terminal claw and spiniform setae on flexor margin included), terminating in one claw, with 6–8 accessory spiniform setae on flexor margin; propodus not dilated or curved distally, distal spiniform setae not enlarged, 7.88–10.60 (median 10.00, n = 3) times as long as wide, 3.00–3.53 (median 3.32, n = 3) times as long as dactylus; carpus 5.38–6.67 (median 5.65, n = 4) times as long as wide, 0.62–0.64 (median 0.63, n = 2) times as long as propodus, 0.50–0.56 (median 0.52, n = 4) times as long as merus, with one large spiniform setae near distal margin and one small spiniform setae proximal; merus 8.56–9.67 (median 8.85, n = 4) times as long as wide, 1.79–2.00 (median 1.93, n =4) times as long as carpus, bearing 2 or 3 strong movable spiniform setae on externo-inferior margin; ischium with one spiniform setae (lacking in one specimen examined). Fourth pereiopod similar to third but slightly smaller. Fifth pereiopod (Figure 6F,G) slender; dactylus (Figure 6G) 3.00–3.80 (median 3.63, n = 6) times as long as wide (terminal claw and spiniform setae on flexor margin included), terminating in one large claw, with 21–35 spiniform setae on flexor margin; propodus with small spiniform setae on ventral margin, distal pair of spiniform setae not remarkable enlarged, propodus 10.75–12.14 (median 11.54, n = 5) times as long as wide, 3.07–3.54 (median 3.33, n = 5) times as long as dactylus; carpus 5.17–5.75 (median 5.43, n = 3) times as long as wide, 0.51–0.55 (median 0.53, n = 4) times as long as propodus, 0.53–0.61 (median 0.56, n = 4) times as long as merus, with 1 strong spiniform setae near distal margin; merus 8.33–10.73 (median 9.57, n = 3) times as long as wide, 1.63–1.90 (median 1.79, n = 4) times as long as carpus, bearing 2 or 3 strong appressed movable spiniform setae on externo-inferior margin. Ischium without a spiniform seta.
Pleopods. Endopod of male´s first pleopod (Figure 6H) elongated triangular, lacking an appendix interna, 2.00—2.57 (medina 2.45, n = 4) times as long as proximal width, 0.23–0.26 (median 0.24, n = 3) times as long as exopod, with long pappose setae on distal, inner and outer margin. Appendix masculina on male second pleopod (Figure 6I) stick-like, 9.33–11.50 (median 10.42, n = 2) times as long as wide, 0.68–0.72 (median 0.70, n = 2) times as long as endopod, armed with strong spiniform setae on inner and distal margin; appendix interna arising from about 0.4 times length of appendix masculina, reaching to 0.74–0.81 (median 0.77, n = 2) times length of appendix masculina.
Sexual dimorphism. Males similar to females except size, relative length of antennular peduncle and sexual appendages. The main morphological differences from females lead in the structure of the endopod of first pleopod as well as the presence of an appendix masculina on second pleopod.
Reproductive biology. Ovigerous females with medium-sized eggs; size of eggs 0.59–0.66 x 0.34–0.43 mm (n = 8). 25–108 eggs counted in 4 ovigerous specimens.
Size. Postorbital carapace length of apparently adult specimens 1.9–4.3 mm.
Colouration. Body transparent. Rostrum red, red chromatophores also arranged to a narrow line running along the abdominal sternites, at the distal part of the exopod of uropod and a red transversal line at dorsal margin of third abdominal segment. Eggs greenish (Wilson 2008; Short, Page and Humphrey 2019).
Distribution. The species is known from rivers draining to the Gulf of Carpentaria (Queensland) and to the Timor Sea at the Darwin area (Northern Territory).
Etymology. The species is named after the biologist and aquarist Dave Wilson, collector of a part of the type material and an enthusiast on Australian fish and shrimp fauna.
Common name. Darwin Red Nose Shrimp.
Remarks. Caridina wilsoni n. sp. resembles Caridina gracilirostris De Man, 1892 both in coloration as well as in general morphology (body very slender; rostrum strongly curved upwards with few widely spaced teeth on proximal part of dorsal margin, no postorbital teeth; preanal carina with a hook-like tooth; distal margin of telson with a low number of spiniform setae). The proposed new species differs from this species by its larger egg size (0.59–0.66 x 0.34–0.43 mm vs. 0.34–0.39 x 0.21–0.25 mm in C. gracilirostris), a slightly stouter carpus of second cheliped (3.73–4.94 times as long as wide vs. 4.69–5.26 times as long as wide in C. gracilirostris), merus of third pereiopod 8.56–9.67 times as long as wide vs. 9.55–10.22 in C. gracilirostris, propodus of fifth pereiopod 3.07–3.54 times as long as dactylus vs. 4.0–4.25 in C. gracilirostris. The proposed new species also resembles C. gracillima Lanchester, 1901 known from the Malayan peninsula especially in egg size (0.59–0.66 x 0.34–0.43 mm vs. 0.55–0.66 x 0.35–0.40 mm in C. gracillima) [20], but differs from this species by its preanal carina bearing a hook-like spine vs. preanal carina rounded without a spine in C. gracillima.
Molecular Systematics
In the molecular phylogeny based on a fragment of the mitochondrial 16S rRNA gene, the sequences of each of the three new species form a strongly supported clade (Figure 7). Their relationship to the next respective relative is also well supported in each case. Caridina darwin n.sp. is closely related to Caridina excavatoides, which is its sister species, and the allied species Caridina malayensis and C. temasek. Caridina magnovis n.sp. is found in a sister group relationship to C. serratirostris. Caridina wilsoni n.sp. is the sister species to C. gracillima and also closely related to Caridina gracilirostris and C. neglecta. These relationships closely reflect the morphological similarities of the new Australian species as described above. Each species is also differing strongly genetically from its closest relative (minimum p-distances: 6.5% Caridina wilsoni n.sp. – C. gracillima; 9.1% C. darwin n.sp. – C. excavatoides; 12.6% C. magnovis n.sp. – C. serratirostris), which offers additional clear support for their status as distinct species.
Page et al. [1] remarked on the close evolutionary relationships of Australian species with non-Australian taxa from the surrounding regions and hinted at the endemism of Caridina wilsoni (“Gulf1”) and C. magnovis (“W WA4”) in Australia. This is hardly surprising for amphidromous species and also not in cases such as Caridina magnovis n.sp., whose closest relative, C. serratirostris, is a species that is widespread throughout the Indo-Australian Archipelago (IAA) and from which C. magnovis n.sp. seems to have evolved into an Australian endemic species. The sister species of Caridina wilsoni n.sp., C. gracillima, has a similar egg size to C. wilsoni n.sp., but both are closely related to the very widespread amphidromous species Caridina gracilirostris (the extent of the distribution of the equally closely related C. neglecta is poorly known), so it is not overly surprising that Caridina gracillima and C. wilsoni n.sp. occur on the western and eastern end of the IAA, respectively. Most interesting perhaps in this context is C. darwin n.sp., which has relatively small eggs in comparison to its closest relatives with medium to large sized eggs and a distribution confined to the Sunda shelf area. Although an Australian endemism of all three new species is likely, their medium egg size differs from other endemic Australian species with large eggs indicative of highly suppressed larval development, such as Caridina biyiga from Leichhardt Springs, Kakadu National Park, Northern Territory (egg size 0.75-1.0 mm) [9].
The rather restricted molecular dataset presented here aimed at providing support for morphospecies in an integrative taxonomy approach is not suited to discuss evolutionary and biogeographic aspects in more detail, but the few observations made above underline once again the diverse biogeographic origin of the Australian atyid fauna and their potential for more extensive studies on the adaptations going along with the colonization of that continent.
Author Contributions
Conceptualization, W.K. and K.vR.; morphological examination and drawings, W.K.; phylogenetical analysis, T.vR.; writing—original draft preparation, W.K.; writing—review and editing, T.vR. and K. vR.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors thank Timothy J. Page, Australia, for providing the majority of specimens examined for this study. We further thank Suzanne Horner (MAGNT), Marissa McNamara QM and Andrew Hosie (WAM) for providing museum numbers for deposited material and Sammy de Grave for providing a free waiver for this publication.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Page, T. J., von Rintelen, K., Hughes, J. M. An island in the stream: Australia’s place in the cosmopolitan world of Indo-West Pacific freshwater shrimp (Decapoda: Atyidae: Caridina). Mol. Phylogenetics Evol. 2007, 43, 645–659. [CrossRef] [PubMed]
- Cook, B.D., Page, T.J.; Hughes, J.M. (2011). Molecular and conservation biogeography of freshwater caridean shrimps in north-western Australia. In C. Held, S. Koenemann & C. D. Schubart (Eds.), Phylogeography and Population Genetics in Crustacea (pp. 273–289). CRC Press.
- Coleman, Charles, Oliver “Digital inking”: How to make perfect line drawings on computers. Org. Divers. Evol. Electron. Suppelment 2003, 14, 1–14.
- Coleman, Charles, Oliver Substituting time-consuming pencil drawings in arthropod taxonomy using stacks of digital photographs. Zootaxa 2006, 1360, 61–68.
- Short, J.W. A revision of Australian river prawns, Macrobrachium (Crustacea: Decapoda: Palaemonidae). Hydrobiologia 2004, 525, 1–100. [Google Scholar] [CrossRef]
- von Rintelen, K., von Rintelen, T.; Glaubrecht, M. Molecular phylogeny and diversification of freshwater shrimps (Decapoda, Atyidae, Caridina) from ancient Lake Poso (Sulawesi, Indonesia) – the importance of being colourful. Mol. Phylogenetics Evol. 2007, 45, 1033–1041. [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Short, J.W., Page, T.J.; Humphrey, C.L. Caridina biyiga sp. nov., a new freshwater shrimp (Crustacea: Decapoda: Atyidae) from Leichhardt Springs, Kakadu National Park, Australia, based on morphological and molecular data, with a preliminary illustrated key to Northern Territory Caridina. Zootaxa 2019, 4695, 1–25. [CrossRef]
- de Mazancourt, V., Klotz, W., Marquet, G., Mos, B., Rogers, D.C.; Keith, P. The complex study of complexes: the first well-supported phylogeny of two species complexes within genus Caridina (Decapoda: Caridea: Atyidae) sheds light on evolution, biogeography, and habitat. Mol. Phylogenetics Evol. 2019, 131, 164–180. [CrossRef]
- Huelsenbeck, J.P., Ronquist, F., Nielsen, R.; Bollback, J. P. Bayesian inference of phylogeny and its impact on evolutionary biology. Science 2001, 294, 2310–2314. [CrossRef] [PubMed]
- Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [CrossRef] [PubMed]
- Nguyen, L.T., Schmidt, H.A., Von Haeseler, A.; Minh, B.Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [CrossRef] [PubMed]
- Hoang, D.T., Vinh, L.S., Flouri, T., Stamatakis, A., von Haeseler, A.; Minh, B.Q. MPBoot: fast phylogenetic maximum parsimony tree inference and bootstrap approximation. BMC Evol. Biol. 2018, 18, 11. [CrossRef]
- Chernomor, O., Von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [CrossRef] [PubMed]
- Swofford, D. L. (2002). PAUP* (version 4.0). Phylogenetic analysis using parsimony (*and other methods). Sinauer.
- Choy, S.C.; Ng, P.K.L. A new species of freshwater atyid shrimp, Caridina temasek (Decapoda: Caridea:Atyidae) from Singapore. Raffles Bull. Zool. 1991, 39, 265–277. [Google Scholar]
- Cai, Y., Ng, P.K.L.; Choy, S. Freshwater shrimps of the family Atyidae (Crustacea: Decapoda: Caridea) from Peninsular Malaysia and Singapore. Raffles Bull. Zool. 2007, 55, 277–309.
- Wilson, D. Freshwater Shrimps from the Top End. Australian Aquarium Magazine, 2008, 51–53.
- Cai, Y.; Ng, P.K.L. A revision of the Caridina gracilirostris De Man, 1892, species group, with descriptions of two new taxa (Decapoda; Caridea; Atyidae). J. Nat. Hist. 2007, 41, 1585–1602. [Google Scholar] [CrossRef]
Figure 1.
A, C, E–N Caridina darwin n.sp. paratype ♀ cl 3.2 mm, NTM Cr019571; B, D Caridina darwin n.sp. paratype ♂ cl 2.4 mm, NTM Cr019570. A, B carapace; C telson, D distal margin of telson; E preanal carina; F uropodal diaeresis; G antennular peduncle; H scaphocerite; I mandibles; J maxillulae; K maxillae; L first maxilliped; M palp of first maxilliped; N second maxilliped.
Figure 1.
A, C, E–N Caridina darwin n.sp. paratype ♀ cl 3.2 mm, NTM Cr019571; B, D Caridina darwin n.sp. paratype ♂ cl 2.4 mm, NTM Cr019570. A, B carapace; C telson, D distal margin of telson; E preanal carina; F uropodal diaeresis; G antennular peduncle; H scaphocerite; I mandibles; J maxillulae; K maxillae; L first maxilliped; M palp of first maxilliped; N second maxilliped.

Figure 2.
A, B, D–G Caridina darwin n.sp. paratype ♀ cl 3.2 mm, NTM Cr019571; C, H, I Caridina darwin n.sp. paratype ♂ cl 2.4 mm, NTM Cr019570. A third maxilliped, B first pereiopod, C second pereiopod, D third pereiopod, E dactylus of third pereiopod, F fifth pereiopod, G dactylus of fifth pereiopod, H first pleopod, I second pleopod.
Figure 2.
A, B, D–G Caridina darwin n.sp. paratype ♀ cl 3.2 mm, NTM Cr019571; C, H, I Caridina darwin n.sp. paratype ♂ cl 2.4 mm, NTM Cr019570. A third maxilliped, B first pereiopod, C second pereiopod, D third pereiopod, E dactylus of third pereiopod, F fifth pereiopod, G dactylus of fifth pereiopod, H first pleopod, I second pleopod.
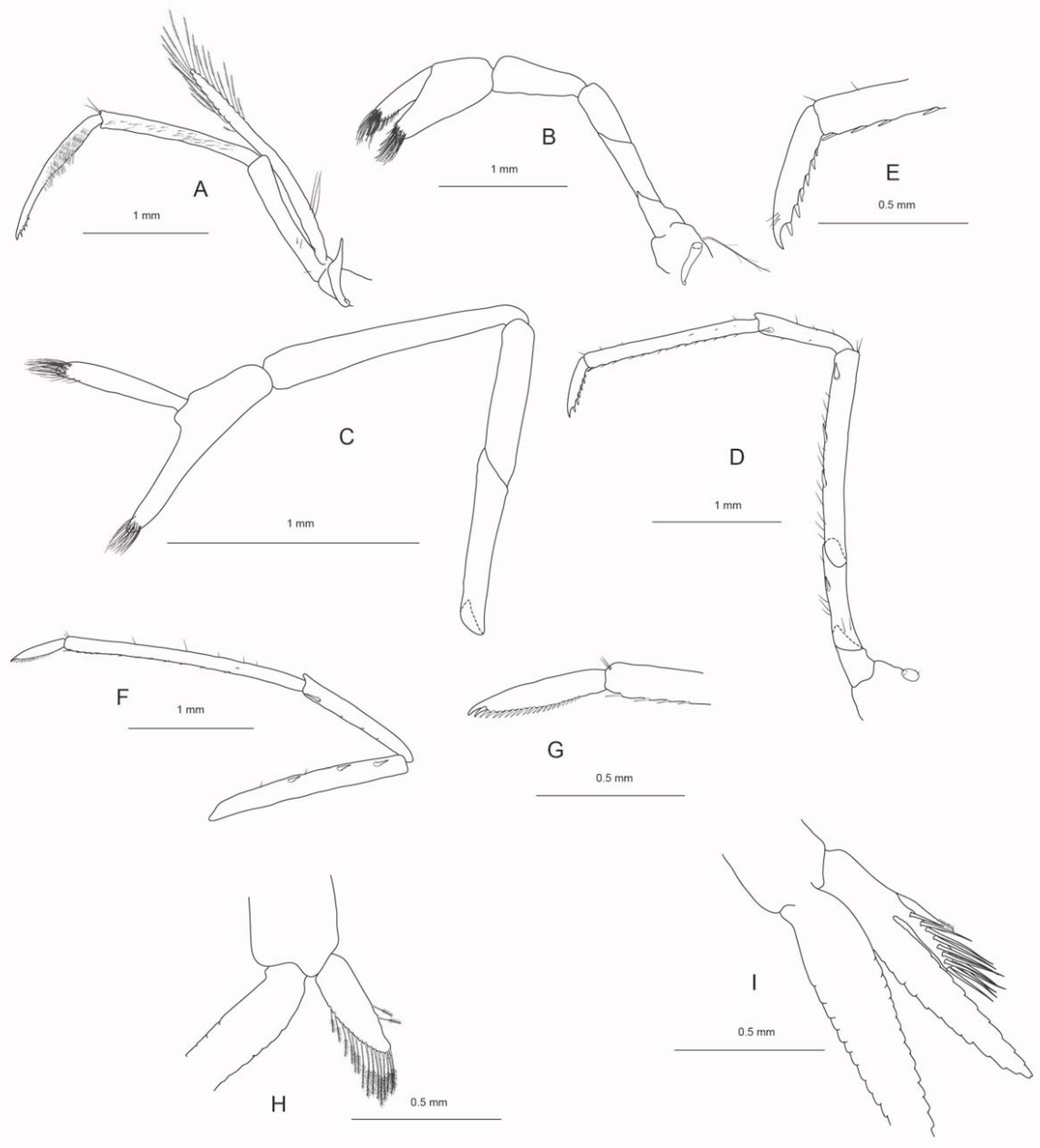
Figure 3.
A, C–F, I Caridina magnovis n.sp. paratype ov. ♀ cl 4.0 mm, NTM Cr019573; B, G, H, J–O Caridina magnovis n.sp. paratype ♂ cl 2.7 mm, WAM C81394. A, B carapace; C telson, D distal margin of telson; E preanal carina; F uropodal diaeresis; G antennular peduncle; H scaphocerite; I, J mandibles; K maxillulae; L maxillae; M first maxilliped; N palp of first maxilliped; O second maxilliped.
Figure 3.
A, C–F, I Caridina magnovis n.sp. paratype ov. ♀ cl 4.0 mm, NTM Cr019573; B, G, H, J–O Caridina magnovis n.sp. paratype ♂ cl 2.7 mm, WAM C81394. A, B carapace; C telson, D distal margin of telson; E preanal carina; F uropodal diaeresis; G antennular peduncle; H scaphocerite; I, J mandibles; K maxillulae; L maxillae; M first maxilliped; N palp of first maxilliped; O second maxilliped.

Figure 4.
A–E, G–K Caridina magnovis n.sp. paratype ♂ cl 2.7 mm, WAM C81394. F Caridina magnovis n.sp. paratype ov. ♀ cl 4.0 mm, NTM Cr019573. A third maxilliped, B first pereiopod, C second pereiopod, D third pereiopod, E, F dactylus of third pereiopod, G dactylus of fourth pereiopod, H fifth pereiopod, I dactylus of fifth pereiopod, J first pleopod, K second pleopod.
Figure 4.
A–E, G–K Caridina magnovis n.sp. paratype ♂ cl 2.7 mm, WAM C81394. F Caridina magnovis n.sp. paratype ov. ♀ cl 4.0 mm, NTM Cr019573. A third maxilliped, B first pereiopod, C second pereiopod, D third pereiopod, E, F dactylus of third pereiopod, G dactylus of fourth pereiopod, H fifth pereiopod, I dactylus of fifth pereiopod, J first pleopod, K second pleopod.

Figure 5.
A, Caridina wilsoni n.sp. paratype ov. ♀ cl 3.5 mm, ZMB 29180; B–N Caridina wilsoni n.sp. paratype ♂ cl 3.0 mm, QM W29818. A, B carapace; C telson, D distal margin of telson; E preanal carina; F uropodal diaeresis; G antennular peduncle; H scaphocerite; I mandible; J maxillulae; K maxillae; L first maxilliped; M palp of first maxilliped; N second maxilliped.
Figure 5.
A, Caridina wilsoni n.sp. paratype ov. ♀ cl 3.5 mm, ZMB 29180; B–N Caridina wilsoni n.sp. paratype ♂ cl 3.0 mm, QM W29818. A, B carapace; C telson, D distal margin of telson; E preanal carina; F uropodal diaeresis; G antennular peduncle; H scaphocerite; I mandible; J maxillulae; K maxillae; L first maxilliped; M palp of first maxilliped; N second maxilliped.
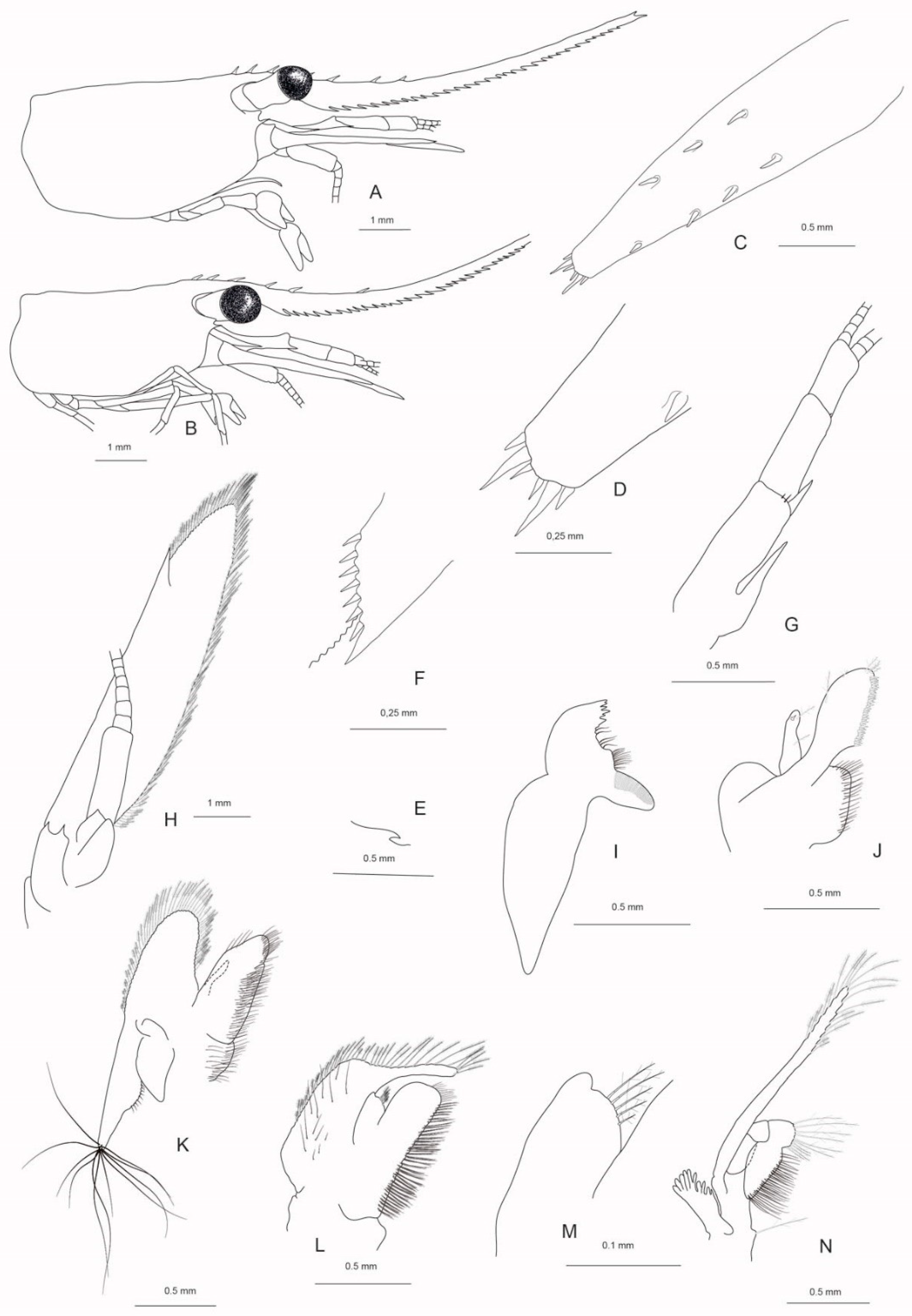
Figure 6.
A–I Caridina wilsoni n.sp. paratype ♂ cl 3.0 mm, QM W29818. A third maxilliped, B first pereiopod, C second pereiopod, D third pereiopod, E dactylus of third pereiopod, F fifth pereiopod, G dactylus of fifth pereiopod, H first pleopod, I second pleopod.
Figure 6.
A–I Caridina wilsoni n.sp. paratype ♂ cl 3.0 mm, QM W29818. A third maxilliped, B first pereiopod, C second pereiopod, D third pereiopod, E dactylus of third pereiopod, F fifth pereiopod, G dactylus of fifth pereiopod, H first pleopod, I second pleopod.
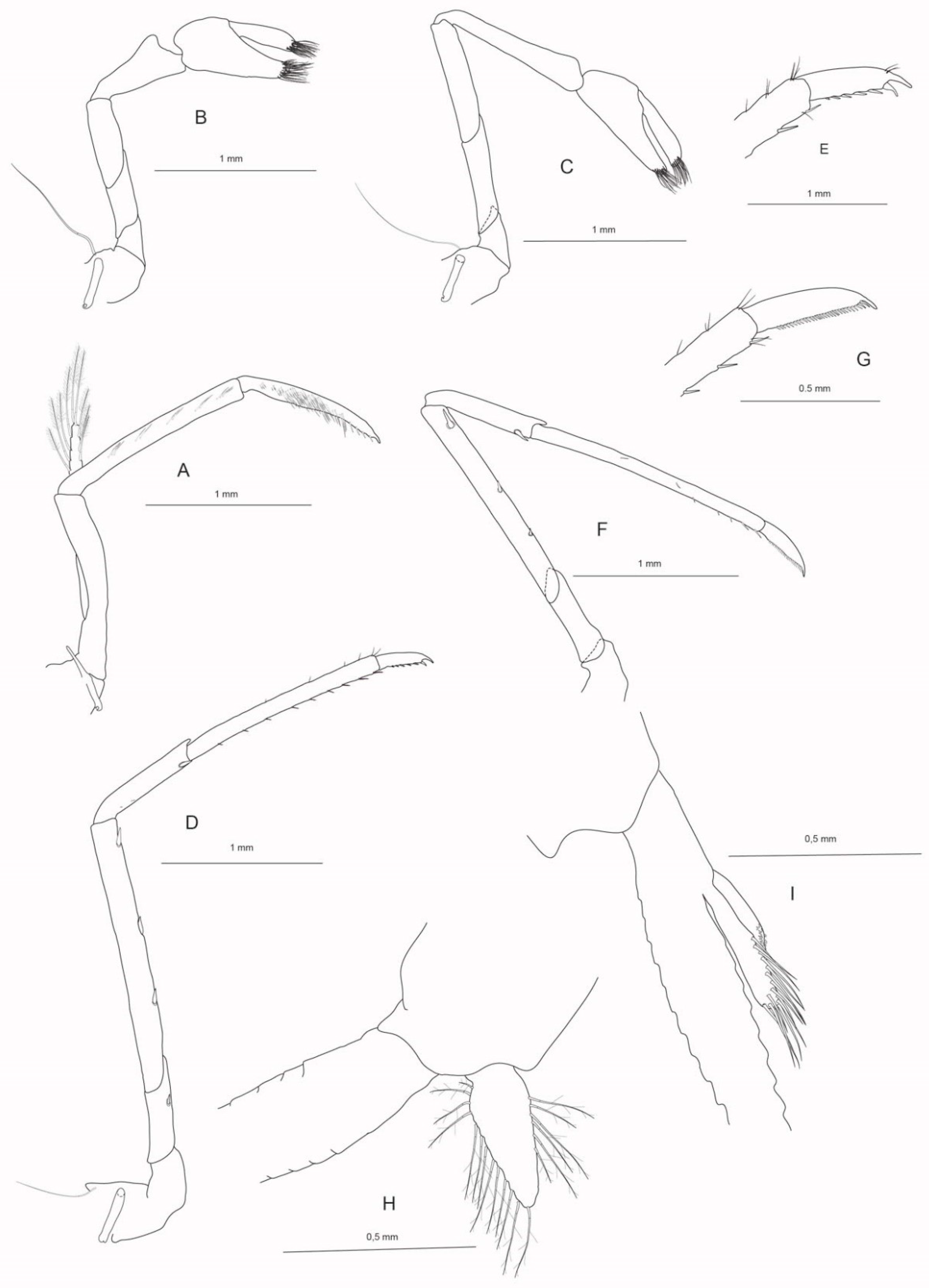
Figure 7.
Molecular phylogeny (BI, 16S) showing the relationship of the three new species to morphologically similar congeners. Numbers on branches are, from the top, Bayesian posterior probabilities and ML/MP bootstrap values. The scale bar indicates the substitution rate.
Figure 7.
Molecular phylogeny (BI, 16S) showing the relationship of the three new species to morphologically similar congeners. Numbers on branches are, from the top, Bayesian posterior probabilities and ML/MP bootstrap values. The scale bar indicates the substitution rate.
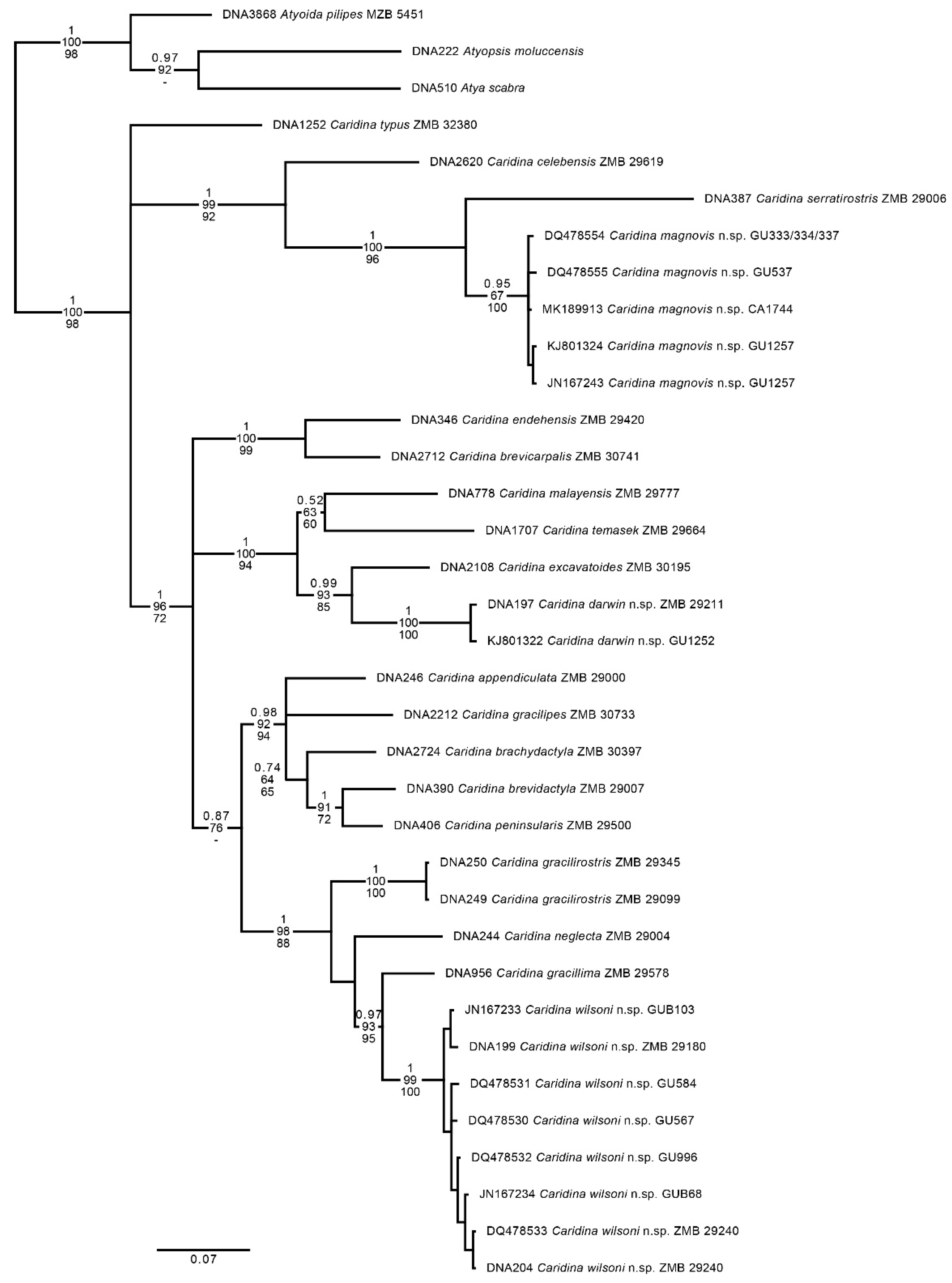

Table 1.
List of sequenced specimens with geographic origin, voucher information and data source. “ZMB DNA” vouchers are DNA samples from specimens with an origin in the pet trade without any specimen remains.
Table 1.
List of sequenced specimens with geographic origin, voucher information and data source. “ZMB DNA” vouchers are DNA samples from specimens with an origin in the pet trade without any specimen remains.
| Species | Origin | Museum Voucher | GenBank Accession | Sequence Source |
|---|---|---|---|---|
| Atya scabra | Panama | ZMB DNA 510 | # | This study |
| Atyoida pilipes | Indonesia: Sulawesi | MZB Cru 5451 | # | This study |
| Atyopsis moluccensis | Indo-Pacific | ZMB DNA 222 | # | This study |
| Caridina appendiculata | Indonesia: Sulawesi | ZMB 29000 | # | This study |
| Caridina brachydactyla | Indonesia: Sulawesi | ZMB 30397 | # | This study |
| Caridina brevicarpalis | Indonesia: Sulawesi | ZMB 30741 | # | This study |
| Caridina brevidactyla | Indonesia: Sulawesi | ZMB 29007 | # | This study |
| Caridina celebensis | Philippines: Mindoro | ZMB 29619 | # | This study |
| Caridina darwin n.sp. | Australia: NT | ZMB 29211 GU1252 |
# KJ801322 |
This study Short et al. 2019 |
| Caridina endehensis | Indonesia: Sulawesi | ZMB 29420 | # | This study |
| Caridina excavatoides | Thailand | ZMB 30195 | # | This study |
| Caridina gracilipes | Indonesia: Sulawesi | ZMB 30733 | Klotz et al. 2023 | |
| Caridina gracilirostris | Indonesia: Bali | ZMB 29345 ZMB 29099 |
# # |
This study This study |
| Caridina gracillima | Malaysia (peninsular) | ZMB 29578 | # | This study |
| Caridina magnovis n.sp. | Australia: WA Australia: NT |
GU333/334/337 GU537 GU1257 CA1744 |
DQ478554 DQ478555 KJ801324 JN167243 MK189913 |
Page et al. 2007 Short et al. 2019 De Mazancourt et al. 2019 |
| Caridina malayensis | Malaysia (peninsular) | ZMB 29777 | # | This study |
| Caridina neglecta | Indonesia: Sulawesi | ZMB 29004 | # | This study |
| Caridina peninsularis | Malaysia (peninsular) | ZMB 29500 | # | This study |
| Caridina serratirostris | Indonesia: Sulawesi | ZMB 29006 | # | This study |
| Caridina temasek | Singapore | ZMB 29664 | # | This study |
| Caridina typus | Reunion | ZMB 32380 | # | This study |
| Caridina wilsoni n.sp. | Australia: NT Australia: QLD Australia: NT/QLD |
ZMB 29180 ZMB 29240 GU567 GU584 GU996 GUB68 GUB103 |
# DQ478533 DQ478530 DQ478531 DQ478532 JN167234 JN167233 |
This study This study Page et al. 2007 Page et al. 2007 Page et al. 2007 Page et al. 2007 Cook et al. 2011 Cook et al. 2011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



