
Submitted:
08 January 2024
Posted:
10 January 2024
You are already at the latest version
Abstract
Hypervirulent Klebsiella pneumoniae (hvKp) is a variant that has been increasingly linked to severe, life threatening infections including pyogenic liver abscess and bloodstream infections. HvKp belonging to the capsular serotypes K1 and K2 have been reported worldwide, however very scarce studies are available on the their genomics and virulence. In the current study we report four hypermucoviscous hvKp ESBL clinical strains of capsular serotype K1 and K2 isolated from pus and urine of critically ill patients in tertiary care hospitals in Oman. These strains belong to diverse sequence types (STs) namely ST-23(K1), ST-231(K2), ST-881(K2), and ST-14(K2). To study their virulence, Galleria Mellonella model and resistance to human serum killing were used. The G. mellonella model revealed that the K1/ST-23 isolate is the most virulent as 50% of the larvae died in the first day, followed by isolate K2/ST-231 for which 75% of the larvae died in the second day and isolate K2/ST-14 for which 50% of the larvae died in the second day. Resistance to human serum killing showed there was complete inhibition of bacterial growth of all four isolates by the end of the first hour and up to the third hour. For ST-23(K1), ST-231(K2), ST-881(K2), and ST-14(K2), there was a gradual decrease in the number of colonies up to the second to third hour, which indicates neutralization of bacterial cells by the serum components. However, this was followed by sudden increase of bacterial growth, indicating possible resistance of bacteria against human serum bactericidal activity. Whole genome sequencing (WGS) revealed that hvKp strains display unique genetic arrangement of k-loci. Whole genome SNP-based phylogenetic analysis revealed that these hvKp isolates are phylogenetically distinct, belonging to diverse clades and belong to different STs in comparison to global isolates. This is the first report from Oman detailing the WGS of hvKp clinical isolates and assessing their resistance and virulence genomics, which reinforce our understanding of their epidemiology and dissemination in clinical settings.
Keywords:
Hypermucoviscous1
; Klebsiella pneumoniae2
; Hypervirulent3
; Mobile genetic elements4
; capsule serotypes5
; Virulence6
; Whole genome sequencing7
; sequence types8
1. Introduction
Klebsiella pneumoniae is considered to be the most common cause of hospital-acquired infections (HAIs) accounting for 10% of all nosocomial infections worldwide [1]. Immunocompromised patients are at higher risk of HAIs comprising 8% to 12% of hospitalized patients particularly ventilator-associated pneumonia [2]. This can result in life-threatening illnesses with mortality rates ranging from 50% to 100% 3,4. K. pneumoniae have evolved over the years from the classical type into a hypervirulent K. pneumoniae (hvKp), which has been observed in many countries around the globe [4]. However, hypervirulent K. pneumoniae strains (hvKp) can also be acquired from the community and have been shown to infect otherwise healthy and immunocompetent individuals [5]. There has been an increase trend in emerging of new clonal lineages of hypervirulent strains with co-occurrence of multidrug resistance genotype, known as MDR- hvKp. The most common circulating hvKp serotypes include K1, K2, K20, K54 and K57, with K1 and K2 being the most virulent and accounting for 70% of hvKp isolates [7]. HvKp strains producing hyper-mucoviscous exopolysaccharide coating capsule which contributes significantly to the pathogenicity and virulence of the bacteria and have been associated with dissemination of severe infections such as pyogenic liver abscess mainly in K1 and ST-23 serotypes 8,9. Moreover, hvKp can cause other invasive infections particularly serotypes K2 and ST-65 [4]. However, it is well established that the hypermucoviscosity and hypervirulence are two distinct phenotypes and do not necessarily co-exist in virulent strains [10].
In addition to the capsule, other virulence factors in K. pneumoniae can increase the virulence including the mucoid phenotype regulators (rmpA and rmpA2), siderophores including: yersiniabactin irp2, ybt and Aerobactin iucA, outer membrane porin (KpnO), and phospholipase D family protein (PLD1) [11]. Other virulence factors which have not been thoroughly characterized including efflux pumps, iron transport systems and genes involved in allantoin metabolism [12,13,14]. Recent studies have shown possible contribution of allantoin related genes to progression of colorectal cancer [15]. In addition, some studies have demonstrated that the virulome of the same species could change depending on the host factors [16]. The host factors that expose hospitalized individuals to colonization and infection including the admission into the intensive care unit (ICU), poor adherence to infection control strategies, prolonged use of invasive devices and broad-spectrum antibiotics as well as immunocompromised state especially diabetics and alcoholics [17]. However, as discussed earlier, K. pneumoniae infection can also be acquired from the community especially hypervirulent strains that proved to infect people who are healthy and immunocompetent [14].
Apart from the intrinsic resistance to ampicillin in hvKp, antibiotic resistance phenotype in hvKp is still not very prevalent and the reason is still unclear [4]. However, there has been an increasing trend in resistance in hvKp isolates due to dissemination via mobile genetic elements such as types of NDM, OXA-48 and K. pneumoniae carbapenamase (KPC). Recently, the hvKp serotypes K1 and K2 have been detected from clinical isolates in Oman. Therefore, this study aims to characterize their virulence and antibiotic resistance genomically. We aimed to study host responses against infection with K. pneumoniae K1 and K2 serotypes circulating in Oman.
2. Results
2.1. ST-231 and ST-395 Are the Two Prominent Sequence Types
Initial PCR screening of MDR K. pneumoniae was performed for the purpose of assessing the prevalence of hvKp capsular serotypes [Supplementary figure 1]. This screening revealed that only four out of 129 K. pneumoniae isolates were hypervirulent capsular strains: Kp124 (K1/ST-23), Kp 125 (K2/ST-231), Kp 126 (K2/ST-881) and Kp 83 (K2/ST-14) [Figure 1].
Three of these isolates were invasive K. pneumoniae strains with hypermucoviscous phenotype (HMV K. pneumoniae) and one of these (K2/ST-881) was carbapenemase-producing (CRE) isolate. WGS analysis was performed for these four isolates in addition to the other isolates for detrmining the ST and k-loci composition. MLST was performed on the isolates from WGS data to determine their sequence type (ST) using the CGE website [Table 1] [18]. One isolate from a pyogenic infection, belonged to the ST-23, similar to global trends associating these features [Lee et al., 2017]. On the other hand, the three other isolates belonged to three different STs including ST-14, ST-231 and ST-881 [Table 1]. One third (32%, n=10) of the isolates belonged to ST-231 and 19% of the isolates (n=6) belonged to ST-395 [Table 1]. Moreover, 6% of the isolates (n=2) had ST-23 and another 6% of the isolates (n=2) had ST-405.
2.2. K-loci Analysis Shows Unique Genetic Arrangement for STs with hvKp
Initial screening of MDR K. pneumoniae revealed that four isolates displayed hvKp phenotype. These four isolates were isolated mainly from urine and one from wound swab. We conducted a comparison of the k-loci in these isolates to known K. pneumoniae references in the database using the Kaptive online tool to analyze similarities and variabilities [19]. The genetic arrangement of different k-loci in each isolate as well as percentage of similarity between the isolate to known references showed that most of the screened K. pneumoniae clinical isolates have similar genetic arrangement [Supplementary figure 3-30]. On the other hand, it was shown that the isolates with unique k-loci genetic arrangements belong to the hvKp isolates with serotypes K1/ST-23, K2/ST-231, K2/ST-881 and K2/ST-14 [Table 2].
The phenotypic antimicrobial susceptibility profiles of the hvKp isolates were performed using disk diffusion [Table 3 and 4]. All hvKp isolates in this study (Kp 83, Kp 125 and Kp 126) that are K2 positive and Kp 124 that is K1 positive are ESBL producers. Kp 83 (K2/ST-14) showed resistance to AMP, TZP, FEP, CTX, IMP, CAZ and CIP. Kp 126 (K2/ST-881) was resistant to AMP, FEP, CTX, IMP, CAZ, CIP. Whereas Kp124 (K1/ST-23) was resistant to AMP, TZP, FEP, CTX, FOX, CAZ. On the other hand, Kp 125 (K2/ST-231), is KPC producer with resistance to AMP, TZP, FEP, CTX, CAZ.
2.3. Plasmid compositions of hvKp K1/ST-23 (Kp124), K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83)
The four isolates belonging to K1/ST-23, K2/ST-231, K2/ST-881and K2/ST-14 were collected during possible cross-transmission episodes in SQUH, Muscat, Oman between 2019 to 2020. Whole genome sequences of these isolates comprised 5569468 (Kp125 K2/ST-231) bases, 5572070 (K1/ST-23) bases, 5638661 (Kp126 K2/ST-881) bases, and 5447756 (Kp83 K2/ST-14) bases with 57% GC content in all strains [Supplementary data 3]. There has been an increase in reporting MDR in hvKp clinical strains over the last few years, therefore designated as MDR- hvKp. This was attributed to the horizontal gene transfer mediated by plasmids (type I and type II) [5]. For example, type I plasmids were reported to carry blaNDM-1 and blaKPC-3 20–23. In Silico detection of plasmids using plasmidFinder demonstrated that the plasmids in these strains carry mobilizable pathogenicity and resistance-conferring genes [24,25] [Table 3]. The isolate Kp 125 belonging to K1/ST-231 contain seven plasmids; pColKP3 which carries one copy of OXA-181 and KPC-3 conferring carbapenem and cephalosporin resistance by mechanism of antibiotic inactivation. The other plasmids are pIncFII(pAMA11 67-NDM- 5), carrying New Delhi metallo-β-lactamase (NDM)-producing enzymes, and pIncFIB(pQil). Isolate belonging to K2/ST-23 carries three plasmids, including pIncFIB(pQil), containing KPC-3, which has been attributed to the global spread of bla-KPC.
In K1/ST-23 Col440II shared 100% with the reference plasmid (CP023921). On the other hand, pIncFIB(K) shares 98% identity with the reference plasmids. Isolate belonging to K2/ST-231 has three plasmids which are: IncFIB(pQil), IncFII(pAMA1167-NDM-5) and RepB. pRepB is a megaplasmid (244,152 bp) associated with the IncFIB family and has been known to integrate massive accessory modules. In ST-231 pRepB carries a very large composite transposon (15,832 bp) and insertion sequences (ISEc27) and IS102-like among other mobile elements like integrons, and MDR conferring genes [26].
Isolate Kp 83 (K2/ST-14) has four plasmids including, pIncFIA (HI1) that carries resistance genes for several antibiotic categories such as tetracyclines, phenicols, folate pathway antagonists and aminoglycosides. pIncFIB(K) harbors resistance genes against sulfonamides, macrolides and beta lactams. pIncFIB(pNDM-Mar) carries antimicrobial genes against phenicols, beta lactams, quinolones and aminoglycosides. Finally, pIncR carries antibiotic genes conferring resistance to beta lactams, quinolones and aminoglycosides [Table 3]. While most of these plasmids share common sequences, there are wide areas of mosaic genetic structures which might indicate multiple recombination events [Supplementary figures 31 and 32].
2.4. The Pathogenicity Genes in hvKp Strains
WGS of the K124 (K1/ST-23), Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14), were compared to the reference genomes of K. pneumoniae strain RJF293 (accession number CP014008.1 ) which belongs to serotype K2 assigned to ST-374. WGS analysis revealed that genes encoding for pathogenicity are conserved in the analyzed strains at the same relative chromosomal position. There are four main virulence classes in K. pneumoniae including the capsule, lipopolysaccharide (LPS), siderophores and fimbriae [supplementary file 3]. Aerobactin, but not Yersiniabactin, Salmochelin, or Enterobactin, enables the growth and survival of hvKp Ex-vivo and In-vivo [4]. Enterobactin (entB) is detected in all the four hvKp isolates, whereas Aerobactin (iucA) is found in Kp 124 (K1/ST-23), Kp 125 (K2/ST-231), Kp 126 (K2/ST-881), but not Kp83 (K2/ST-14). yersiniabactin (irp2 and ybt) and salmochelin (iroB) were missing in all isolates. Capsule biosynthesis gene rcsB was detected in all isolates but not the regulators rmpA and rmpA2. LPS, adhesins and type 3 fimbriae were detected in all isolates. These findings indicate that our hvKp are well equipped with the virulence machinery which could under the right conditions enable hypervirulent phenotype.
2.5. Clinical hvKp Were Susceptible to Human Serum and Associated with Increased Virulence in G. Mellonella Model
The isolates K124 (K1/ST-23), Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14), were tested for survival in normal human serum in comparison to the ATCC 1705 K. pneumoniae control, [27]. In isolate K124 (K1/ST-23) [Figure 2], there is a slight decrease in the growth indicating the death of bacterial cells followed by a slight increase in the bacterial growth in at T=3. In isolate Kp125 (K2/ST-231), in the first hour there is a decrease in the number of colonies, however, in the second and third hours there is a significant increase in bacterial growth. In isolate Kp126 (K2/ST-881), there is a decrease in growth of bacteria in the first hour but in the second and third hour there is a noticable increase in the number of colonies, which is higher compared to the previous two isolates. In Kp83 (K2/ST-14), is a slight decrease in the number of colonies in the first and second hours. However, after the second hour and by the third hour, there is a dramatic increase in the bacterial growth. In all replicates of the four tested isolates, p value showed statistical significance (p-value of 0.0007).
To correlate between resistance to serum killing and virulence, the same strains were injected into G. mellonella larvae. The survival curve of the three replicates separately, are shown for comparison [Figure 3]. Isolates K124 (K1/ST-23) and Kp83 (K2/ST-14) seem to be the most virulent isolates as most larvae died in the first two days. However, isolates K125 (K2/ST-231) and Kp126 (K2/ST-881) seem to be less virulent as the number of larvae gradually decreased throughout the experiment. In the three replicates [Figure 3 (A)], the Control 1 larvae, consisted of the untouched larvae. In one replicate of the control group, there was a gradual decline in the survival of the larvae from day 2 to 5 as the first larva died in the second day.
Isolate Kp 124 is represented in green, isolate Kp 125 in purple straight line in (C) and dotted in panel (D) for clarity, isolate Kp 126 in cyan, and isolate Kp 83 in yellow as indicated in the colour legend in the right side of each image.
However, in the other two replicates, there were no deaths throughout the 5 observation days but by the fifth day the larvae slow down as cocoon formation commenced. In Control 2 larvae, consisted of PBS only infected larvae, there is also a gradual decline in the survival of the larvae throughout the 5 days in replicates one, however no death was noticed in the second replicate until the larvae slowed down in motility as cocoon formation commenced. However, in the third replicate, there was 100% death, which could suggest the poor quality of that batch of control larvae. In Control 3 larvae, consisting of larvae injected with isolate Kp 99 that is negative for all hypervirulent capsular serotypes, in the first day about 20- 50% of the larvae died with no death in the remaining 50-60% of larvae until the end of the experiment as observed in the three replicates. In strain K124 (K1/ST-23) test group, more than 50% in two replicates and 100% in the other replicate of the larvae died in the first day of the experiment. The remainder of larvae were dead by the fourth day. For strain K125 (K2/ST-231) test group, about 50-75% of the larvae died in the second day and by the 4th day all larvae were dead, in almost all three replicates. In isolate Kp 126 (K2/ST-881) test group, the death ranged between 25% to 60% of the larvae in the first day but overall, the survival rate declines gradually throughout the 5 observation days, in the three replicates. In isolate Kp 83 (K2/ST-14), more than 50%-100% of larvae died by the second day of the experiment in the three replicates.
2.6. Phylogenetic Analysis Shows Diverse Sequence Types
A whole-genome single nucleotide polymorphisms (SNPs) calling phylogenetic tree of 131 isolates including 30 strains in this study (from CPHL and Kp 83, represented in blue) as well as 101 strains from Genbank global strains (represented in black) [27] were plotted to compare genetic relatedness (Figure 4). The hypervirulent isolates are marked with differently colored arrows to highlight their position in the tree amongst other isolates.
A separate phylogenetic tree of 31 isolates for a close-up inspection, including the hypervirulent isolates of this study Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14), as well as 27 other non-hypervirulent K. pneumoniae local clinical isolates [Figure 5]. The unique STs and are highlighted in different colors. Group A in blue is representing isolates with ST-395, group B in yellow belong to ST-405, group C in green belong to ST-23, group D in red has ST-231 and finally the unhighlighted isolates belong to miscellaneous STs. The hypervirulent isolates Kp124 (K1/ST-23), Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14) are indicated with magenta arrows. Hypervirulent Kp83 (K2/ST-14) and Kp124 (K1/ST-23), are branching from the same clade but belong to two different clusters, C and D, respectively. Following constructing the k loci in Kaptive, it was evident from the phylogenetic tree that the clinical strains with similar STs have high similarity in the genetic arrangement of k-loci [Figure 1] and [supplementary figures 3-30]. The hypervirulent serotypes positive for K2, namely K2/ST-231 (Kp125), K2/ST-881 (Kp126), and K2/ST-14 (Kp83), exhibit diversity in their STs, resulting in distinct k-loci gene compositions.
The K. pneumoniae capsular synthesis loci, k-loci, in all our isolates generally consists of the following conserved genes including galF, ORF2 (cpsACP), wzi, wza, wzb, wzc, and gnd which are chromosomally encoded core and similar to what has been reported in the literature [28,29] [Figure 1] as these are necessary for capsular synthesis along with other genes. Moreover, galF, ORF2 and gnd are involved in carbohydrates metabolism, wzi (orfX), wza, wzb, and wzc are responsible for the capsule translocation and surface assembly [30,31]. In our isolates ICEfinder showed that Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14), have 7, 3, 2, and 1 putative Type 4 secretion system with putative ICEs in, respectively [Supplementary figure 33].
The hypervirulent strains carry a wide range of acquired antimicrobial genes [Figure 5]. Kp83 (K2/ST-14) carries a variety of resistance-conferring genes as compared to the other isolates except for qnrS1, blaOXA-2, blaAP and tet(A) genes. Kp 124 (K1/ST-23) carries the least number of resistance-conferring genes and harbors no resistance genes for some antibiotic classes including folate pathway antagonist, quaternary ammonium compound, streptogramin b, rifamycin, aminocyclitol, macrolide, lincosamide and tetracycline. Moreover, isolate Kp125 (K2/ST-231), carries resistance genes for all antibiotic classes in [Figure 4] except glycosamide and tetracycline. In addition, Kp 126 (K2/ST-881), seems to lack resistance-conferring genes to aminoglycosides, amphenicol, streptogramin b, rifamycin, aminocyclitol, macrolide and lincosamide.
3. Discussion
This study aims to investigate the capsular serotypes in hvKp K1 and K2 high risk clones of ST-23, ST-231, ST-881 and ST14 clinical isolates from Oman from 2016-2021. These clones were compared to the non-outbreak clones and less common STs. This comparison facilitated further exploration of genomic landscape of resistance and virulence co-existence in a wide K. pnuemoniae population. Virulence markers in addition to their spread and resistome was also investigated. The hvKp are known to be associated with increased virulence as well as their association with severe human infections [32]. Only 3% of the clinical isolates tested positive for K1 and K2 during the same period in 2019 at SQUH, from urine and pus samples, and one K2 case was reported in 2017 from a urine specimen. This finding is consistent with several studies which showed that the most commonly circulating hypervirulent capsular serotypes are K1 and K2 and has been attributed to disruption of epithelial and mucosal barriers such as endotracheal tubes, catheters, and surgical wounds in hospital settings [21,27,33,34,35]. Phenotypically, all the hypervirulent isolates in this study are ESBLs, which are known to be associated with increased length of stay and mortality rates. In contrast to previous studies, the prevalence rate of ESBLs were higher in non-hypermucoviscous K. pneumoniae isolates 37. The possibility of an opportunistic infection in these patients cannot be excluded as the majority are immunocompromised [37,38].
The k-loci of the capsular operon share similar genetic organization, in which major operons are conserved in most isolates with few exceptions. MLST analysis showed that most of the isolates with similar STs have similar gene arrangements with a similar reference genome in Kaptive. Moreover, these strains were in the same cluster of in the Whole Genome SNP calling phylogenetic tree. The remaining strains belonged to miscellaneous STs and therefore, variable k-loci genetic arrangement figures were observed. To the best of our knowledge, this observation has not been previously reported, making this study the first to indicate a correlation between genetic k-loci and their STs.
In addition to the conserved genes in the k-loci, there are other genes that are commonly present in the including manC and manB that are involved in the synthesis of GDP-D-mannose as well as RmlA, RmlB, RmlC and RmlD proteins involved in the conversion of glucose 1-phosphate to dTDP-L-rhamnose [40]. Previous studies proved that hypervirulent K. pneumoniae strains are associated with increased antibiotic resistance, therefore, recognizing their global dissemination is an urgent priority 5. According to ResFinder and CARD online tools findings, the hypervirulent strains in this study including Kp124 (K1/ST-23), Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14) are in fact highly resistant as they carry a wide variety of antibiotic resistance genes for multiple antimicrobial classes. All these strains carry incompatibility group (Inc) FIB plasmids that are known to encode both virulence and antimicrobial resistance genes in a wide variety of intestinal bacteria including E. coli, Salmonella and Klebsiella spp. including IncFIA and IncFIB that represent one of the most common plasmid types that play a major role in the dissemination of AMR in Enterobacteriaceae [39,40,41]. Furthermore, IncFIB (K) is carried by both Kp 83 (K2/ST-14) and Kp 126 (K2/ST-881) which suggests likelihood of HGT of these plasmids between different bacteria [42]. Moreover, Kp 125 (K2/ST-231) carries ColKP3 plasmid that is known to harbor OXA-232 carbapenemase responsible for the rapidly increasing carbapenem resistance in K. pneumoniae strains worldwide [43]. Moreover, Kp 125 (K2/ST-231) and Kp 124 (K1/ST-23) both carry IncFIB(pQil) that is known to be harbored by K. pneumoniae and other pathogens responsible for nosocomial infections [44]. This finding suggests the likelihood of HGT of this plasmid amongst K. pneumoniae bacterial isolates since both Kp 125 (K2/ST-231) and Kp 124 (K1/ST-23) were isolated in the same period in 2019 in SQUH.
SNP-calling phylogenetic trees performed on WGS data shows that K. pneumoniae strains isolated from different geographical regions (n=131) are arranged with distinct clustering [27]. The strains in our study were found to be phylogenetically diverse belonging to various STs. The hvKp Kp124 (K1/ST-23), Kp125 (K2/ST-231) are present in a clade A, whereas Kp126 (K2/ST-881) is in clade B. The close-up tree showed that our strains are grouped in four main clusters highlighted with different colors indicating the diverse group of strains circulating in our local hospitals. However, there are clustering in these four main groups indicating a possibility of dissemination at possibly larger scale [45,46]. Isolates belonging to ST-231 have been associated with MDR spread in Europe with the potential to cause a future epidemic possessing a public threat. These strains produce OXA-232 as well as RmtF which are associated with high-level carbapenem resistance [47,48]. Furthermore, K. pneumoniae belonging to ST-395 are associated with the production of OXA-48 which is the most common enzyme causing healthcare associated infections globally. OXA-48 enzyme hydrolyses carbapenems and shows weak activity against extended spectrum cephalosporins such as cefepime and ceftazidime, making it difficult to treat these infections [49,50]. This clone of K. pneumoniae has been detected in many countries of the Middle east including Oman, Yemen and Saudi Arabia [51,52,53]. Furthermore, Kp 117 and Kp 105 belong to ST 405 (blue cluster) that have also been associated with the production of OXA-48 just like clones belonging to ST 395 and thus has been responsible for many hospital- associated outbreaks and war-related outbreaks (in Germany) and some of which are resistant to colistin [54,55,56].
Several studies have shown that infection of G. mellonella with K. pneumoniae has resulted in host responses similar to the innate immune responses in murine models including cell death, inhibition of phagocytosis and antimicrobial peptide production. Furthermore, G. mellonella can differentiate between pathogenic and non-pathogenic Klebsiella strains and showed that the more virulent strains were associated with increased survival and host cellular damage as compared to avirulent strains 59
In the present study, it was shown that Kp 124 (K1/ST-23) is the most virulent as 50% of the larvae died in the first day followed by K2/ST-231where 75% of the larvae died in the second day and Kp 83 (K2/ST-14) where 50% of the larvae died in the second day. Similarly in Europe, K1 positive K. pneumoniae belonging to ST-23 has been associated with severe, and fatal infections associated with liver abscesses60–62
However, in Kp 126 (K2/ST-881) there was a gradual decline in the number of larvae over the course of 5 days perhaps making it the least virulent as the death was slower and steadier. It was expected for Kp 126 (K2/ST-881), although it was a hypervirulent strain, it belongs to ST-881 that according to the literature has not been associated with severe human infections as compared to the other STs of Kp 83 (K2/ST-14), Kp 124 (K1/ST-23) and Kp 125 (K2/ST-231). The G. mellonella experiment was repeated three times to allow for comparison and to check how the quality of larvae might affect the results. The results of the experiments though all indicate the higher virulence of hypervirulent K1 and K2 capsular serotype isolates as compared to non-hypervirulent strains [61,62]. However, the main limitation in this assay was due to shipping and delivery restrictions caused by the COVID-19 pandemic as the larvae used in this study were not ideal.
Human serum resistance experiments were performed for the hypervirulent strains Kp124 (K1/ST-23), Kp125 (K2/ST-231), Kp126 (K2/ST-881) and Kp83 (K2/ST-14). For each isolate, 3 replicates were done for accurate comparison and reproducibility. There was complete inhibition of bacterial growth in all four isolates by the end of the first hour and up to the third hour. This is due to the presence of complement proteins and lysozyme which have a bactericidal effect [63]. Furthermore, unexpected pattern of bacterial growth was observed in all replicas for all four strains. For Kp124 (K1/ST-23), there was a gradual decrease in the number of colonies up to the second hour which indicates neutralization of bacterial cells by the serum components, however, in the third hour there was a sudden splurge of bacterial growth. For Kp125 (K2/ST-231), the same decrease in bacterial growth was noticed by the end of the first hour but the dramatic increase in the bacterial growth starting from the second hour unlike in Kp124 (K1/ST-23). Furthermore, for Kp126 (K2/ST-881) and Kp83 (K2/ST-14) similar patterns of sudden growth of bacterial cells commenced in the third hour. Therefore, the sudden increase in the bacterial growth despite the unfavorable serum environment for growth indicates possible resistance of bacteria against human serum bactericidal activity [64]. Human serum, unlike blood, contains antibodies and complement proteins that contribute to the neutralization of bacteria but does not contain immune cells such as phagocytes. Previous studies discuss the bactericidal activity that human serum possesses against Gram-negative bacteria including K. pneumoniae. Moreover, they highlight that the resistance against human serum may be a significant virulence factor. Therefore, the strains used in this assay might have acquired the resistance against human serum as one of their virulence factors [65]. Furthermore, earlier studies have shown that K. pneumoniae LPS O antigen and the capsule polysaccharide contributes strongly to the resistance against human serum antibodies and complement proteins. The isolates tested in this experiment are hypervirulent and possess a thick capsule therefore, their resistance to human serum could be explained by this theory [66,67]. In future studies it would be worthy to test the hypervirulent strains of this study in human blood that contains immune cells such as phagocytes to investigates how it compares to human serum. In previous studies K. pneumoniae has shown its ability to resist killing by phagocytosis in human blood killing assays [68].
4. Materials and Methods
4.1. Bacterial Isolates
The study was commenced after obtaining approval by the Medical Ethics Research committee (MREC#1896), College of Medicine and Health Sciences, Sultan Qaboos University, Muscat, Oman. Initially, we screened 129 K. pneumoniae clinical isolates for K1, K2, K20, K54 and K57 from various samples (urine, wound, tracheal aspirate, blood, sputum), from patients from Central Public Health Laboratories (CPHL) representing various areas from Oman collected between 2015 to 2020. Another 30 isolates were screened from Sultan Qaboos University Hospital (SQUH), Muscat Oman collected between 2019 to 2021 (Supplementary Table 1). PCR preliminary data is presented in supplementary table 5 and 6 including PCR cycling conditions and primers. Colonies were collected from purity plates of CLED agar (Oxoid, Basingstoke Hampshire, UK) were used to make frozen stock of our samples in cryotubes containing beads according to the manufacturer’s instructions (Mast Diagnostics, Derby, UK). The samples were frozen at −80 ◦C for future use.
4.2. String Test for Hypermucoviscosity
An inoculation loop was used to stretch a colony on a Cystine Lactose Electrolyte Deficient (Oxoid, Basingstoke Hampshire, UK) agar plate and the immediate formation of a viscous string of > 5mm in length was indicative of a hypermucoviscous strain of K. pneumoniae [69].
4.3. Antibiotic Susceptibility Testing
Antimicrobial susceptibility profiles of the isolates were carried out using disk diffusion method [72]. Firstly, from an overnight pure culture, three to five colonies were suspended in 5 mL of normal saline (Fisher Scientific, United Kingdom). The suspension was adjusted to 0.5 McFarland (approximately 1–2 × 108 CFU/mL) using CrystalSpec nephlometer (BD Diagnostics, USA) following the manufacturer’s protocol. The antibiotic disks include ampicillin (AMP 10 mg), piperacillin/tazobactam (TZP 110 mg), cefepime (FEP 30 mg), cefotaxime (CTX 30 mg), cefoxitin (FOX 30 mg), ceftazidime (CAZ 30 mg), imipenem (IPM 10 mg), meropenem (MEM 10mg), amikacin (AK 30 mg), gentamicin (CN 10 mg), and ciprofloxacin (CIP 5 mg), (Liofilchem, Italy and BioMérieux, France. The susceptibility assay was carried out according to the CLSI standards [70] were placed on the nutrient agar plates (Oxoid, United Kingdom) and inoculated using sterile forceps. The plates were then incubated for 18-24 hours at 37°C. E. coli (ATCC 25922) and P. aeruginosa (ATCC 27853) were used as susceptible control strains. The readings were taken after 18-24 hours and the interpretive categories and zone diameter breakpoints are listed in Table 4.
4.4. DNA Extraction and WGS
DNA was extracted from an overnight culture using QIAamp DNA Mini Kit (Qiagen, Germany) for whole genome sequencing. The manufacturer’s protocol was followed with slight modifications. For highly mucoid K. pneumoniae isolates, a pre-lysis step was performed which includes the preparation of a pre-lysis buffer constituting of 100 μl of Tris//EDTA (TE) buffer (ThermoFisher Scientific, USA), 1.00 μl lysozyme 10mg/mL (final concentration 0.1mg/mL) (Sigma-Aldrich, USA), 0.20 μl lysostaphin 10mg/mL (final concentration 0.02mg/mL) (ThermoFisher Scientific, USA), 0.10 μl RNAse A 100mg/mL (final concentration 0.1mg/mL) (Qiagen, Germany). The bacterial suspension from an overnight culture was centrifuged at 8000 x g for 2 minutes and the pelleted bacterial cells were resuspended in 100 μl of the pre-lysis buffer by pipetting up and down several times with a P200 pipette. The suspension was then incubated for 60 minutes. After incubation, 1.00 μl of Proteinase K (Sigma-Aldrich, USA) was added as a final step in the pre-lysis stage. The sample was then ready for the first step of the Qiagen DNA extraction protocol as discussed in the previous section.
After DNA quantification by NanoDrop (ThermoFisher Scientific 1000 NanoDrop Spectrophotometer), the samples were sent to microbesNG in the United Kingdom for WGS by Illumina next-generation sequencing (https://microbesng.co.uk, Birmingham, United Kingdom) [71]. The extracted DNA was prepared following the manufacturer’s protocol as described in the company’s website as follow: ILLUMINA SEQUENCING (SGS and EGS) Genomic DNA libraries are prepared using the Nextera XT Library Prep Kit (Illumina, San Diego, USA) following the manufacturer’s protocol with the following modifications: input DNA is increased 2-fold, and PCR elongation time is increased to 45 s. DNA quantification and library preparation are carried out on a Hamilton Microlab STAR automated liquid handling system (Hamilton Bonaduz AG, Switzerland). Pooled libraries are quantified using the Kapa Biosystems Library Quantification Kit for Illumina. Libraries are sequenced using Illumina sequencers (HiSeq/NovaSeq) using a 250bp paired end protocol. Reads are adapter trimmed using Trimmomatic 0.30 with a sliding window quality cutoff of Q15 [72].De novo assembly is performed on samples using SPAdes version 3.7 76, and contigs are annotated using Prokka 1.11[77].
4.5. Bioinformatics Analysis
For visualization of the K. pneumoniae contigs as well as the investigation of main virulence genes, Artemis [75] and SnapGene Viewer (https://snapgene.com) software were used. Moreover, Artemis was used to identify Klebsiella capsule synthesis loci in the different isolates. Basic Local Alignment Search Tool (BLAST) was used to identify the similarities between the loci against a reference K. pneumoniae RJF293 (GenBank accession number CP014008) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) [76]. In addition, Genbank was used to search for similar DNA sequences or genes from the DNA database to the sequenced genomes of this study [77]. Kaptive was used for locus typing and variant analysis on the pre-assembled sequences against a K. pneumoniae database (https://kaptive-web.erc.monash.edu/) [78]. The Klebsiella capsule synthesis loci (k-loci) of each of the sequenced genomes were generated and were used for comparison between the different isolates (Supplementary figures 3-30). Kleborate v2.1.0 was used to investigate other virulence factors including yersiniabactin, colibactin, aerobactin and salmochelin [79]. ICEfinder online tool was used to detect integrative and conjugative elements (https://bioinfo-mml.sjtu.edu.cn/ICEfinder/ICEfinder.html) [80]. The multilocus sequence typing (MLST) was determined using the MLST tool in the Center for genomic epidemiology (CGE) (http://cge.cbs.dtu.dk/services) [81]. PlasmidFinder and ResFinder in the CGE website were used to identify plasmids and acquired antimicrobial resistance genes, respectively [82]. Additionally, Comprehensive Antibiotic Resistance Database (CARD) (https://card.mcmaster.ca/) was used to detect putative antimicrobial resistance genes using Resistance Gene Identifier (RGI) tool. This tool identifies the antibiotic resistome(s) as well as point mutations within the resistance-conferring genes [83]. Furthermore, the phylogeny tool in the CGE server was used to investigate the genetic relatedness between the different sequences by identifying Single Nucleotide Polymorphisms (SNPs) [84]. The tree files were visualized using Mega7 that performs automatic sequence alignment and infers phylogenetic trees [88] .In addition, a combined phylogenetic tree of 131 isolates that includes the isolates in this study as well as published global isolates was created using R, a statistical programming software for comparison and visualization of genetic similarities or differences between the project’s isolates and the published ones [86]. iTOL; an online tool, was used to display, annotate, and manage the phylogenetic tree 90. A whole genome SNP alignment was generated using Snippy v4.4.5 [Seemann 2015] and the AE015929 genome as a reference. Then, iqtree v1.6.12, using model finder and ultrafast bootstrap 92, was used to produce a maximum likelihood phylogenetic tree. A phylogenetic tree of the whole genome SNP was constructed and linked to the gene analysis heat map using the R platform [93,94]. List of Clinical isolate serial numbers of phylogenetic trees in Figure 4 and Figure 5 are presented in supplementary table 4. For statistical analysis, two-way ANOVA was used to analyze variations in bactericidal activity of human serum against K. pneumoniae clinical isolates using GraphPap Prism software (10.0.3). P-value of < 0.05 is considered as statistically significant. Observation tables are presented in Supplementary table 3.
4.6. G. mellonella Virulence Assays
Virulence assay was performed using the larvae of G. mellonella as a model of infection. The protocol was modified from [92]. To ensure quality, the larvae were manually picked through local beekeepers in Oman and were kept at room temperature (25 °C) in the dark and wood shavings were provided as a food source. Stage 6 larvae, approximately 2-3 cm in size were selected for the purpose of the experiment. For each isolate and the controls, a replicate of 10 and 5 larvae were used, respectively. The larvae were distributed in Petri dishes lined with wood shavings before injecting them with bacteria the next day. In day 1, the selected isolates for the experiment were sub-cultured into CLED agar plates (Oxoid, Basingstoke Hampshire, UK) from cryo-beads. In day 2, three to five colonies were picked up and resuspended in 10 mL of liquid broth then incubated overnight at 37 °C. In day 3, a 1:100 dilution (1 mL bacterial suspension + 9 mL liquid broth) was grown for 3-4 hours. The bacterial suspension was then centrifuged, the supernatant discarded, and the pelleted bacteria was washed three times with sterile 1x Phosphate buffered saline (PBS) to remove excess salts (Invitrogen, USA). The bacterial pellet was then resuspended in PBS to an absorbance of 0.2 (OD600) which was repeated for each isolate. The absorbance was measured using a spectrophotometer (Eppendorf BioPhotometer Spectrophotometer UV/VIS, USA) and the colony-forming unit (CFU) counts were determined for each sample. For the test larvae, 10 μL of each sample (undiluted 0.2 OD600 suspension) was injected into the larvae. For Control 1 larvae, PBS only was injected and for Control 2 larvae, they were kept untouched, for Control 3 larvae, Kp 99 non-hypermucovirulent (negative for any capsular serotype) bacterial suspension was injected. All larvae were injected through the haemocoel via the rear left pro-leg using an insulin syringe. Post injection, the larvae were incubated at 37 °C and their mortality was assessed every 24 hours for 5 days. Over the course of 5 days, the larvae were examined with forceps by flipping them into their backs and checking if there are signs of motility. Live larvae will quickly flip back whereas sick larvae will be slow, and the exterior will be hardened and darkened, and they will usually die within the coming 1-2 days. Dead larvae will have a hard, dry, and very dark exterior and will shrink in size. The results were observed and noted, and survival graphs were generated using GraphPad prism 10.0.3 software (La Jolla California USA, www.graphpad.com). Observation data is presented in Supplementary table 3.
4.7. Serum Resistance Assay
The susceptibility of bacteria to human serum was investigated using the method of [93] with some modifications. In day 1, the selected isolates for the experiment were sub-cultured into agar plates from cryo-beads as well as K. pneumoniae ATCC 1705 that was used as a positive control in this experiment. In day 2, 3-5 colonies were picked up and resuspended in 10 mL of liquid broth then incubated overnight at 37 °C. In day 3, the bacterial suspension was diluted to 2 x 106 CFU/mL. The Then 25 µL of bacterial suspension and 75 µL of human serum (Sigma- Aldrich, USA) were dispensed in microtitration trays and mixed then incubated at 37 °C. To assess viability, the samples were streaked onto agar plates immediately at time zero, after 1 hour, after 2 hours and finally after 3 hours. Each isolate was tested 3 times. The streaked agar plates were incubated overnight. In day 4, CFU counts of each sample and the control at 4 different time points were recorded and graphs were made using Microsoft Excel, 2010. (Supplementary tables 6-10)
5. Conclusions
In summary, to our knowledge this is the first study in Oman to depict the molecular and genomic characteristics of hypervirulent K. pneumoniae isolates collected from all around the country, where analysis of WGS of the isolates showed distinct pattern of k-loci depending on the particular STs. The hvKp strains in this study are diverse in the phylogenetic origin but the likelihood of transmission and future outbreaks is key to detect possible spread of hvKp isolates in health-care settings via active surveillance.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
“Conceptualization, Z.J. and R.F; methodology, B.B.; software, B.B. and Z.S.; validation, M.R. and Z.J.; formal analysis, B.B. and Z.S.; investigation, B.B.; resources, A.R., M.M. and A.J.; data curation, Z.J and Z.S.; writing—original draft preparation, B.B.; writing—review and editing, Z.J.; visualization, M.R.; supervision, Z.J.; project administration, Z.J.; funding acquisition, Z.J. All authors have read and agreed to the published version of the manuscript.”.
Funding
“This research was funded by Funding was provided by Sultan Qaboos University internal grant (Project code: IG/MED/MICR/21/01).
Institutional Review Board Statement
The study was commenced after obtaining approval by the Medical Ethics Research committee (MREC#1896), College of Medicine and Health Sciences, Sultan Qaboos University, Muscat, Oman.
Informed Consent Statement
“Not applicable.”.
Data Availability Statement
All supporting data can be found in the supplementary files. All whole genome sequencing data are deposited in Genbank at accession number PRJNA999478. The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Acknowledgments
We would like to express our sincere thanks to the sequencing company pro- vided by MicrobesNG (Birmingham, UK) (https://microbesng.com (accessed on 6th June 2023) for performing WGS and bioinformatics analysis. We would like to thank supervising staff, College of Agriculture, Department of Food Microbiology for providing instrumental support to conduct some experiments for cultivating the bacterial isolates.
Conflicts of Interest
“The authors declare no conflict of interest.”.
Appendix A
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
References
- Choby, J. E.; Howard-Anderson, J.; Weiss, D. S. Hypervirulent Klebsiella Pneumoniae – Clinical and Molecular Perspectives. Journal of Internal Medicine. Blackwell Publishing Ltd March 1, 2020, pp 283–300. [CrossRef]
- Prokesch, B. C.; TeKippe, M.; Kim, J.; Raj, P.; TeKippe, E. M. E.; Greenberg, D. E. Primary Osteomyelitis Caused by Hypervirulent Klebsiella Pneumoniae. The Lancet Infectious Diseases. Lancet Publishing Group 2016, pp e190–e195. [CrossRef]
- Esposito, E. P.; Cervoni, M.; Bernardo, M.; Crivaro, V.; Cuccurullo, S.; Imperi, F.; Zarrilli, R. Molecular Epidemiology and Virulence Profiles of Colistin-Resistant Klebsiella Pneumoniae Blood Isolates from the Hospital Agency “Ospedale Dei Colli,” Naples, Italy. Front Microbiol 2018, 9 (JUL). [CrossRef]
- Li, D.; Huang, X.; Rao, H.; Yu, H.; Long, S.; Li, Y.; Zhang, J. Klebsiella Pneumoniae Bacteremia Mortality: A Systematic Review and Meta-Analysis. Frontiers in Cellular and Infection Microbiology. Frontiers Media S.A. 2023. [CrossRef]
- Lee, C. R.; Lee, J. H.; Park, K. S.; Jeon, J. H.; Kim, Y. B.; Cha, C. J.; Jeong, B. C.; Lee, S. H. Antimicrobial Resistance of Hypervirulent Klebsiella Pneumoniae: Epidemiology, Hypervirulence-Associated Determinants, and Resistance Mechanisms. Frontiers in Cellular and Infection Microbiology. Frontiers Media S.A. November 21, 2017. [CrossRef]
- Zhu, Z.; Zhang, B.; Wang, Y.; Jing, S.; Ning, W.; Liu, C.; Chen, C. A Wide Clinical Spectrum of Pulmonary Affection in Subjects with Community-Acquired Klebsiella Pneumoniae Liver Abscess (CA-KPLA). Journal of Infection and Chemotherapy 2023, 29 (1), 48–54. [CrossRef]
- Yang, Z.; Zhou, R.; Chen, Y.; Zhang, X.; Liu, L.; Luo, M.; Chen, J.; Chen, K.; Zeng, T.; Liu, B.; Wu, Y.; Huang, J.; Liu, Z.; Ouyang, J. Clinical and Molecular Characteristics and Antibacterial Strategies of Klebsiella Pneumoniae in Pyogenic Infection. Microbiol Spectr 2023. [CrossRef]
- Russo, T. A.; Olson, R.; MacDonald, U.; Beanan, J.; Davidsona, B. A. Aerobactin, but Not Yersiniabactin, Salmochelin, or Enterobactin, Enables the Growth/Survival of Hypervirulent (Hypermucoviscous) Klebsiella Pneumoniae Ex Vivo and in Vivo. Infect Immun 2015, 83 (8), 3325–3333. [CrossRef]
- Cerdeira, L.; Nakamura-Silva, R.; Oliveira-Silva, M.; Sano, E.; Esposito, F.; Fuga, B.; Moura, Q.; Miranda, C. E. S.; Wyres, K.; Lincopan, N.; Pitondo-Silva, A. A Novel Hypermucoviscous Klebsiella Pneumoniae ST3994-K2 Clone Belonging to Clonal Group 86. Pathog Dis 2021, 79 (8). [CrossRef]
- Catalán-Nájera, J. C.; Garza-Ramos, U.; Barrios-Camacho, H. Hypervirulence and Hypermucoviscosity: Two Different but Complementary Klebsiella Spp. Phenotypes? Virulence. Taylor and Francis Inc. October 3, 2017, pp 1111–1123. [CrossRef]
- Catalán-Nájera, J. C.; Humberto, B. C.; Josefina, D. B.; Alan, S. P.; Rigoberto, H. C.; García-Méndez, J.; Rayo, M. O.; Velázquez-Larios, M. del R.; Vianney, O. N.; Lourdes, G. X.; Celia, A. A.; Jesús, S. S.; Ulises, G. R. Molecular Characterization and Pathogenicity Determination of Hypervirulent Klebsiella Pneumoniae Clinical Isolates Serotype K2 in Mexico. Diagnostic Microbiology and Infectious Disease. Elsevier Inc. July 1, 2019, pp 316–319. [CrossRef]
- Bagley, S. T. Habitat Association of Klebsiella Species. Infection Control. 1985. [CrossRef]
- Rock, C.; Thom, K. A.; Masnick, M.; Johnson, J. K.; Harris, A. D.; Morgan, D. J. Frequency of Klebsiella Pneumoniae Carbapenemase (KPC)–Producing and Non-KPC-Producing Klebsiella Species Contamination of Healthcare Workers and the Environment . Infect Control Hosp Epidemiol 2014, 35 (4). [CrossRef]
- Paczosa, M. K.; Mecsas, J. Klebsiella Pneumoniae: Going on the Offense with a Strong Defense. Microbiology and Molecular Biology Reviews 2016, 80 (3). [CrossRef]
- Kaur, C. P.; Iyadorai, T.; Sears, C.; Roslani, A. C.; Vadivelu, J.; Samudi, C. Presence of Polyketide Synthase (PKS) Gene and Counterpart Virulence Determinants in Klebsiella Pneumoniae Strains Enhances Colorectal Cancer Progression In-Vitro. Microorganisms 2023, 11 (2). [CrossRef]
- Rodríguez-Pallares, S.; Mateo-Vargas, M. A.; Rodríguez-Iglesias, M. A.; Galán-Sánchez, F. Molecular Characterization of Consecutive Isolates of OXA-48-Producing Klebsiella Pneumoniae: Changes in the Virulome Using next-Generation Sequencing (NGS). Microbes Infect 2023, 105217. [CrossRef]
- Ashurst, J. v.; Dawson, A. Pneumonia, Klebsiella; 2018.
- Larsen, J.; Enright, M. C.; Godoy, D.; Spratt, B. G.; Larsen, A. R.; Skov, R. L. Multilocus Sequence Typing Scheme for Staphylococcus Aureus: Revision of the Gmk Locus. Journal of Clinical Microbiology. July 2012, pp 2538–2539. [CrossRef]
- Wick, R. R.; Heinz, E.; Holt, K. E.; Wyres, K. L. Kaptive Web: User-Friendly Capsule and Lipopolysaccharide Serotype Prediction for Klebsiella Genomes. J Clin Microbiol 2018, 56 (6). [CrossRef]
- Han, M.; Liu, C.; Xie, H.; Zheng, J.; Zhang, Y.; Li, C.; Shen, H.; Cao, X. Genomic and Clinical Characteristics of Carbapenem-Resistant Enterobacter Cloacae Complex Isolates Collected in a Chinese Tertiary Hospital during 2013–2021. Front Microbiol 2023, 14. [CrossRef]
- Guo, X.; Chen, R.; Wang, Q.; Li, C.; Ge, H.; Qiao, J.; Li, Y. Global Prevalence, Characteristics, and Future Prospects of IncX3 Plasmids: A Review. Frontiers in Microbiology. 2022. [CrossRef]
- Bilal, H.; Zhang, G.; Rehman, T.; Han, J.; Khan, S.; Shafiq, M.; Yang, X.; Yan, Z.; Yang, X. First Report of Blandm-1 Bearing Incx3 Plasmid in Clinically Isolated St11 Klebsiella Pneumoniae from Pakistan. Microorganisms 2021, 9 (5). [CrossRef]
- Hameed, M. F.; Chen, Y.; Wang, Y.; Shafiq, M.; Bilal, H.; Liu, L.; Ma, J.; Gu, P.; Ge, H. Epidemiological Characterization of Colistin and Carbapenem Resistant Enterobacteriaceae in a Tertiary: A Hospital from Anhui Province. Infect Drug Resist 2021, 14. [CrossRef]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M. V.; Lund, O.; Villa, L.; Aarestrup, F. M.; Hasman, H. PlasmidFinder and PMLST: In Silico Detection and Typing of Plasmid. Antimicrob Agents Chemother 2014, 58.
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M. V.; Lund, O.; Villa, L.; Aarestrup, F. M.; Hasman, H. In Silico Detection and Typing of Plasmids Using Plasmidfinder and Plasmid Multilocus Sequence Typing. Antimicrob Agents Chemother 2014, 58 (7). [CrossRef]
- Nazir, A.; Zhao, Y.; Li, M.; Manzoor, R.; Tahir, R. A.; Zhang, X.; Qing, H.; Tong, Y. Structural Genomics of Repa, Repb1-Carrying Incfib Family Pa1705-Qnrs, P911021-Teta, and P1642-Teta, Multidrug-Resistant Plasmids from Klebsiella Pneumoniae. Infect Drug Resist 2020, 13. [CrossRef]
- Wyres, K. L.; Lam, M. M. C.; Holt, K. E. Population Genomics of Klebsiella Pneumoniae. Nature Reviews Microbiology. 2020. [CrossRef]
- Wang, X.; Xie, Y.; Li, G.; Liu, J.; Li, X.; Tian, L.; Sun, J.; Ou, H. Y.; Qu, H. Whole-Genome-Sequencing Characterization of Bloodstream Infection-Causing Hypervirulent Klebsiella Pneumoniae of Capsular Serotype K2 and ST374. Virulence 2018, 9 (1), 510–521. [CrossRef]
- Pan, Y. J.; Lin, T. L.; Chen, Y. H.; Hsu, C. R.; Hsieh, P. F.; Wu, M. C.; Wang, J. T. Capsular Types of Klebsiella Pneumoniae Revisited by Wzc Sequencing. PLoS One 2013, 8 (12). [CrossRef]
- Pan, Y. J.; Fang, H. C.; Yang, H. C.; Lin, T. L.; Hsieh, P. F.; Tsai, F. C.; Keynan, Y.; Wang, J. T. Capsular Polysaccharide Synthesis Regions in Klebsiella Pneumoniae Serotype K57 and a New Capsular Serotype. J Clin Microbiol 2008, 46 (7). [CrossRef]
- Whitfield, C.; Roberts, I. S. Structure, Assembly and Regulation of Expression of Capsules in Escherichia Coli. Molecular Microbiology. 1999. [CrossRef]
- Rahn, A.; Drummelsmith, J.; Whitfield, C. Conserved Organization in the Cps Gene Clusters for Expression of Escherichia Coli Group 1 K Antigens: Relationship to the Colanic Acid Biosynthesis Locus and the Cps Genes from Klebsiella Pneumoniae. J Bacteriol 1999, 181 (7). [CrossRef]
- Qian, C.; Zhang, S.; Xu, M.; Zeng, W.; Chen, L.; Zhao, Y.; Zhou, C.; Zhang, Y.; Cao, J.; Zhou, T. Genetic and Phenotypic Characterization of Multidrug-Resistant Klebsiella Pneumoniae from Liver Abscess. Microbiol Spectr 2023, 11 (1). [CrossRef]
- Struve, C.; Roe, C. C.; Stegger, M.; Stahlhut, S. G.; Hansen, D. S.; Engelthaler, D. M.; Andersen, P. S.; Driebe, E. M.; Keim, P.; Krogfelt, K. A. Mapping the Evolution of Hypervirulent Klebsiella Pneumoniae. mBio 2015, 6 (4). [CrossRef]
- Cubero, M.; Grau, I.; Tubau, F.; Pallarés, R.; Dominguez, M. A.; Liñares, J.; Ardanuy, C. Hypervirulent Klebsiella Pneumoniae Clones Causing Bacteraemia in Adults in a Teaching Hospital in Barcelona, Spain (2007-2013). Clinical Microbiology and Infection 2016, 22 (2). [CrossRef]
- Zheng, R.; Zhang, Q.; Guo, Y.; Feng, Y.; Liu, L.; Zhang, A.; Zhao, Y.; Yang, X.; Xia, X. Outbreak of Plasmid-Mediated NDM-1-Producing Klebsiella Pneumoniae ST105 among Neonatal Patients in Yunnan, China. Ann Clin Microbiol Antimicrob 2016, 15 (1). [CrossRef]
- Lee, C. H.; Liu, J. W.; Su, L. H.; Chien, C. C.; Li, C. C.; Yang, K. D. Hypermucoviscosity Associated with Klebsiella Pneumoniae-Mediated Invasive Syndrome: A Prospective Cross-Sectional Study in Taiwan. International Journal of Infectious Diseases 2010, 14 (8). [CrossRef]
- Chung, D. R.; Lee, H.; Park, M. H.; Jung, S. I.; Chang, H. H.; Kim, Y. S.; Son, J. S.; Moon, C.; Kwon, K. T.; Ryu, S. Y.; Shin, S. Y.; Ko, K. S.; Kang, C. I.; Peck, K. R.; Song, J. H. Fecal Carriage of Serotype K1 Klebsiella Pneumoniae ST23 Strains Closely Related to Liver Abscess Isolates in Koreans Living in Korea. European Journal of Clinical Microbiology and Infectious Diseases 2012, 31 (4), 481–486. [CrossRef]
- Gorrie, C. L.; Mirc Eta, M.; Wick, R. R.; Edwards, D. J.; Thomson, N. R.; Strugnell, R. A.; Pratt, N. F.; Garlick, J. S.; Watson, K. M.; Pilcher, D. V.; McGloughlin, S. A.; Spelman, D. W.; Jenney, A. W. J.; Holt, K. E. Gastrointestinal Carriage Is a Major Reservoir of Klebsiella Pneumoniae Infection in Intensive Care Patients. Clinical Infectious Diseases 2017, 65 (2), 208–215. [CrossRef]
- Shu, H. Y.; Fung, C. P.; Liu, Y. M.; Wu, K. M.; Chen, Y. T.; Li, L. H.; Liu, T. T.; Kirby, R.; Tsai, S. F. Genetic Diversity of Capsular Polysaccharide Biosynthesis in Klebsiella Pneumoniae Clinical Isolates. Microbiology (N Y) 2009, 155 (12). [CrossRef]
- Johnson, T. J.; Siek, K. E.; Johnson, S. J.; Nolan, L. K. DNA Sequence of a ColV Plasmid and Prevalence of Selected Plasmid-Encoded Virulence Genes among Avian Escherichia Coli Strains. J Bacteriol 2006, 188 (2). [CrossRef]
- Dolejska, M.; Vill, L.; Dobiasova, H.; Fortini, D.; Feudi, C.; Carattoli, A. Plasmid Content of a Clinically Relevant Klebsiella Pneumoniae Clone from the Czech Republic Producing CTX-M-15 and QnrB1. Antimicrob Agents Chemother 2013, 57 (2). [CrossRef]
- Khajanchi, B. K.; Hasan, N. A.; Choi, S. Y.; Han, J.; Zhao, S.; Colwell, R. R.; Cerniglia, C. E.; Foley, S. L. Comparative Genomic Analysis and Characterization of Incompatibility Group FIB Plasmid Encoded Virulence Factors of Salmonella Enterica Isolated from Food Sources. BMC Genomics 2017, 18 (1). [CrossRef]
- Pedersen, T.; Tellevik, M. G.; Kommedal, Ø.; Lindemann, P. C.; Moyo, S. J.; Janice, J.; Blomberg, B.; Samuelsen, Ø.; Langeland, N. Horizontal Plasmid Transfer among Klebsiella Pneumoniae Isolates Is the Key Factor for Dissemination of Extended-Spectrum β-Lactamases among Children in Tanzania. mSphere 2020, 5 (4). [CrossRef]
- Lutgring, J. D.; Zhu, W.; De Man, T. J. B.; Avillan, J. J.; Anderson, K. F.; Lonsway, D. R.; Rowe, L. A.; Batra, D.; Rasheed, J. K.; Limbago, B. M. Phenotypic and Genotypic Characterization of Enterobacteriaceae Producing Oxacillinase-48-like Carbapenemases, United States. Emerg Infect Dis 2018, 24 (4). [CrossRef]
- Ragupathi, N. K. D.; Bakthavatchalam, Y. D.; Mathur, P.; Pragasam, A. K.; Walia, K.; Ohri, V. C.; Veeraraghavan, B. Plasmid Profiles among Some ESKAPE Pathogens in a Tertiary Care Centre in South India. Indian Journal of Medical Research 2019, 149 (2). [CrossRef]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P. A. D.; Brisse, S. Multilocus Sequence Typing of Klebsiella Pneumoniae Nosocomial Isolates. J Clin Microbiol 2005, 43 (8). [CrossRef]
- Choi, M.; Hegerle, N.; Nkeze, J.; Sen, S.; Jamindar, S.; Nasrin, S.; Sen, S.; Permala-Booth, J.; Sinclair, J.; Tapia, M. D.; Johnson, J. K.; Mamadou, S.; Thaden, J. T.; Fowler, V. G.; Aguilar, A.; Terán, E.; Decre, D.; Morel, F.; Krogfelt, K. A.; Brauner, A.; Protonotariou, E.; Christaki, E.; Shindo, Y.; Lin, Y. T.; Kwa, A. L.; Shakoor, S.; Singh-Moodley, A.; Perovic, O.; Jacobs, J.; Lunguya, O.; Simon, R.; Cross, A. S.; Tennant, S. M. The Diversity of Lipopolysaccharide (O) and Capsular Polysaccharide (K) Antigens of Invasive Klebsiella Pneumoniae in a Multi-Country Collection. Front Microbiol 2020, 11. [CrossRef]
- Shen, S.; Han, R.; Yin, D.; Jiang, B.; Ding, L.; Guo, Y.; Wu, S.; Wang, C.; Zhang, H.; Hu, F. A Nationwide Genomic Study of Clinical Klebsiella Pneumoniae Carrying Bla OXA-232 and RmtF in China . Microbiol Spectr 2023, 11 (3). [CrossRef]
- Mancini, S.; Poirel, L.; Tritten, M. L.; Lienhard, R.; Bassi, C.; Nordmann, P. Emergence of an MDR Klebsiella Pneumoniae ST231 Producing OXA-232 and RmtF in Switzerland. Journal of Antimicrobial Chemotherapy. 2018. [CrossRef]
- Potron, A.; Kalpoe, J.; Poirel, L.; Nordmann, P. European Dissemination of a Single OXA-48-Producing Klebsiella Pneumoniae Clone. Clinical Microbiology and Infection 2011, 17 (12). [CrossRef]
- Gijón, D.; Tedim, A. P.; Valverde, A.; Rodríguez, I.; Morosini, M.-I.; Coque, T. M.; Manrique, M.; Pareja, E.; Tobes, R.; Ruiz-Garbajosa, P.; Cantón, R. Early OXA-48-Producing Enterobacterales Isolates Recovered in a Spanish Hospital Reveal a Complex Introduction Dominated by Sequence Type 11 (ST11) and ST405 Klebsiella Pneumoniae Clones . mSphere 2020, 5 (2). [CrossRef]
- Al-Agamy, M. H.; Aljallal, A.; Radwan, H. H.; Shibl, A. M. Characterization of Carbapenemases, ESBLs, and Plasmid-Mediated Quinolone Determinants in Carbapenem-Insensitive Escherichia Coli and Klebsiella Pneumoniae in Riyadh Hospitals. J Infect Public Health 2018, 11 (1). [CrossRef]
- Alsharapy, S. A.; Gharout-Sait, A.; Muggeo, A.; Guillard, T.; Cholley, P.; Brasme, L.; Bertrand, X.; Moghram, G. S.; Touati, A.; De Champs, C. Characterization of Carbapenem-Resistant Enterobacteriaceae Clinical Isolates in Al Thawra University Hospital, Sana’a, Yemen. Microbial Drug Resistance 2020, 26 (3). [CrossRef]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like Carbapenemases: The Phantom Menace. Journal of Antimicrobial Chemotherapy 2012, 67 (7). [CrossRef]
- López-Camacho, E.; Paño-Pardo, J. R.; Ruiz-Carrascoso, G.; Wesselink, J. J.; Lusa-Bernal, S.; Ramos-Ruiz, R.; Ovalle, S.; Gómez-Gil, R.; Pérez-Blanco, V.; Pérez-Vázquez, M.; Gómez-Puertas, P.; Mingorance, J. Population Structure of OXA-48-Producing Klebsiella Pneumoniae ST405 Isolates during a Hospital Outbreak Characterised by Genomic Typing. J Glob Antimicrob Resist 2018, 15. [CrossRef]
- Kutlu, H. H.; Dolapçı, İ.; Avcı, M.; Tekeli, A. The Emergence of Klebsiella Pneumoniae Sequence Type 395 Non-Susceptible to Carbapenems and Colistin from Turkey. Indian J Med Microbiol 2023, 46. [CrossRef]
- Sandfort, M.; Hans, J. B.; Fischer, M. A.; Reichert, F.; Cremanns, M.; Eisfeld, J.; Pfeifer, Y.; Heck, A.; Eckmanns, T.; Werner, G.; Gatermann, S.; Haller, S.; Pfennigwerth, N. Increase in NDM-1 and NDM-1/OXA-48-Producing Klebsiella Pneumoniae in Germany Associated with the War in Ukraine, 2022. Eurosurveillance 2022, 27 (50). [CrossRef]
- Ménard, G.; Rouillon, A.; Cattoir, V.; Donnio, P. Y. Galleria Mellonella as a Suitable Model of Bacterial Infection: Past, Present and Future. Frontiers in Cellular and Infection Microbiology. Frontiers Media S.A. December 22, 2021. [CrossRef]
- Decré, D.; Verdet, C.; Emirian, A.; Le Gourrierec, T.; Petit, J. C.; Offenstadt, G.; Maury, E.; Brisse, S.; Arlet, G. Emerging Severe and Fatal Infections Due to Klebsiella Pneumoniae in Two University Hospitals in France. J Clin Microbiol 2011, 49 (8). [CrossRef]
- Konagaya, K.; Yamamoto, H.; Suda, T.; Tsuda, Y.; Isogai, J.; Murayama, H.; Arakawa, Y.; Ogino, H. Ruptured Emphysematous Prostatic Abscess Caused by K1-ST23 Hypervirulent Klebsiella Pneumoniae Presenting as Brain Abscesses: A Case Report and Literature Review. Front Med (Lausanne) 2022, 8. [CrossRef]
- Nakamura, K.; Nomoto, H.; Harada, S.; Suzuki, M.; Yomono, K.; Yokochi, R.; Hagino, N.; Nakamoto, T.; Moriyama, Y.; Yamamoto, K.; Kutsuna, S.; Ohmagari, N. Infection with Capsular Genotype K1-ST23 Hypervirulent Klebsiella Pneumoniae Isolates in Japan after a Stay in East Asia: Two Cases and a Literature Review. Journal of Infection and Chemotherapy 2021, 27 (10). [CrossRef]
- Shon, A. S.; Bajwa, R. P. S.; Russo, T. A. Hypervirulent (Hypermucoviscous) Klebsiella Pneumoniae: A New and Dangerous Breed. Virulence 2013, 4 (2). [CrossRef]
- Zhao, Y.; Zhang, X.; Torres, V. V. L.; Liu, H.; Rocker, A.; Zhang, Y.; Wang, J.; Chen, L.; Bi, W.; Lin, J.; Strugnell, R. A.; Zhang, S.; Lithgow, T.; Zhou, T.; Cao, J. An Outbreak of Carbapenem-Resistant and Hypervirulent Klebsiella Pneumoniae in an Intensive Care Unit of a Major Teaching Hospital in Wenzhou, China. Front Public Health 2019, 7. [CrossRef]
- Bugla-Płoskońska, G.; Kiersnowski, A.; Futoma-Kołoch, B.; Doroszkiewicz, W. Killing of Gram-Negative Bacteria with Normal Human Serum and Normal Bovine Serum: Use Of Lysozyme and Complement Proteins in the Death Of Salmonella Strains O48. Microb Ecol 2009, 58 (2). [CrossRef]
- Tichaczek-Goska, D.; Witkowska, D.; Cisowska, A.; Jankowski, S.; Hendrich, A. B. The Bactericidal Activity of Normal Human Serum against Enterobacteriaceae Rods with Lipopolysaccharides Possessing O-Antigens Composed of Mannan. Advances in Clinical and Experimental Medicine 2012, 21 (3).
- Benge, G. R. Bactericidal Activity of Human Serum against Strains of Klebsiella from Different Sources. J Med Microbiol 1988, 27 (1). [CrossRef]
- Yeh, K. M.; Chiu, S. K.; Lin, C. L.; Huang, L. Y.; Tsai, Y. K.; Chang, J. C.; Lin, J. C.; Chang, F. Y.; Siu, L. K. Surface Antigens Contribute Differently to the Pathophysiological Features in Serotype K1 and K2 Klebsiella Pneumoniae Strains Isolated from Liver Abscesses. Gut Pathog 2016, 8 (1). [CrossRef]
- Tomas, J. M.; Benedi, V. J.; Ciurana, B.; Jofre, J. Role of Capsule and O Antigen in Resistance of Klebsiella Pneumoniae to Serum Bactericidal Activity. Infect Immun 1986, 54 (1). [CrossRef]
- DeLeo, F. R.; Kobayashi, S. D.; Porter, A. R.; Freedman, B.; Dorward, D. W.; Chen, L.; Kreiswirth, B. N. Survival of Carbapenem-Resistant Klebsiella Pneumoniae Sequence Type 258 in Human Blood. Antimicrob Agents Chemother 2017, 61 (4). [CrossRef]
- Kumabe, A.; Kenzaka, T. String Test of Hypervirulent Klebsiella Pneumonia. QJM. Oxford University Press December 1, 2014, p 1053. [CrossRef]
- Ruangpan, L. Chapter 3. Minimal Inhibitory Concentration (MIC) Test and Determination of Antimicrobial Resistant Bacteria. Laboratory manual of standardized methods for antimicrobial sensitivity tests for bacteria isolated from aquatic animals and environment 2004, No. Mic.
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing A CLSI Supplement for Global Application. Clinical and Laboratory Standards Institute 2020.
- Bolger, A. M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30 (15), 2114–2120. [CrossRef]
- Bolger, A. M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30 (15). [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; Stepanauskas, R.; McLean, J.; Lasken, R.; Clingenpeel, S. R.; Woyke, T.; Tesler, G.; Alekseyev, M. A.; Pevzner, P. A. Assembling Genomes and Mini-Metagenomes from Highly Chimeric Reads. In Lecture Notes in Computer Science (including subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); 2013; Vol. 7821 LNBI. [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30 (14). [CrossRef]
- Carver, T.; Harris, S. R.; Berriman, M.; Parkhill, J.; McQuillan, J. A. Artemis: An Integrated Platform for Visualization and Analysis of High-Throughput Sequence-Based Experimental Data. Bioinformatics 2012, 28 (4). [CrossRef]
- Altschul, S. F.; Gish, W.; Miller, W.; Myers, E. W.; Lipman, D. J. Basic Local Alignment Search Tool. J Mol Biol 1990, 215 (3). [CrossRef]
- Agarwala, R.; Barrett, T.; Beck, J.; Benson, D. A.; Bollin, C.; Bolton, E.; Bourexis, D.; Brister, J. R.; Bryant, S. H.; Canese, K.; Cavanaugh, M.; Charowhas, C.; Clark, K.; Dondoshansky, I.; Feolo, M.; Fitzpatrick, L.; Funk, K.; Geer, L. Y.; Gorelenkov, V.; Graeff, A.; Hlavina, W.; Holmes, B.; Johnson, M.; Kattman, B.; Khotomlianski, V.; Kimchi, A.; Kimelman, M.; Kimura, M.; Kitts, P.; Klimke, W.; Kotliarov, A.; Krasnov, S.; Kuznetsov, A.; Landrum, M. J.; Landsman, D.; Lathrop, S.; Lee, J. M.; Leubsdorf, C.; Lu, Z.; Madden, T. L.; Marchler-Bauer, A.; Malheiro, A.; Meric, P.; Karsch-Mizrachi, I.; Mnev, A.; Murphy, T.; Orris, R.; Ostell, J.; O’Sullivan, C.; Palanigobu, V.; Panchenko, A. R.; Phan, L.; Pierov, B.; Pruitt, K. D.; Rodarmer, K.; Sayers, E. W.; Schneider, V.; Schoch, C. L.; Schuler, G. D.; Sherry, S. T.; Siyan, K.; Soboleva, A.; Soussov, V.; Starchenko, G.; Tatusova, T. A.; Thibaud-Nissen, F.; Todorov, K.; Trawick, B. W.; Vakatov, D.; Ward, M.; Yaschenko, E.; Zasypkin, A.; Zbicz, K. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res 2018, 46 (D1). [CrossRef]
- Wick, R. R.; Heinz, E.; Holt, K. E.; Wyres, K. L. Kaptive Web: User-Friendly Capsule and Lipopolysaccharide Serotype Prediction for Klebsiella Genomes. J Clin Microbiol 2018, 56 (6). [CrossRef]
- Lam, M. M. C.; Wick, R. R.; Watts, S. C.; Cerdeira, L. T.; Wyres, K. L.; Holt, K. E. A Genomic Surveillance Framework and Genotyping Tool for Klebsiella Pneumoniae and Its Related Species Complex. Nat Commun 2021, 12 (1). [CrossRef]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H. Y. ICEberg 2.0: An Updated Database of Bacterial Integrative and Conjugative Elements. Nucleic Acids Res 2019, 47 (D1), D660–D665. [CrossRef]
- Larsen, M. v.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R. L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D. W.; Aarestrup, F. M.; Lund, O. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J Clin Microbiol 2012, 50 (4). [CrossRef]
- Bortolaia, V.; Kaas, R. S.; Ruppe, E.; Roberts, M. C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R. L.; Rebelo, A. R.; Florensa, A. F.; Fagelhauer, L.; Chakraborty, T.; Neumann, B.; Werner, G.; Bender, J. K.; Stingl, K.; Nguyen, M.; Coppens, J.; Xavier, B. B.; Malhotra-Kumar, S.; Westh, H.; Pinholt, M.; Anjum, M. F.; Duggett, N. A.; Kempf, I.; Nykäsenoja, S.; Olkkola, S.; Wieczorek, K.; Amaro, A.; Clemente, L.; Mossong, J.; Losch, S.; Ragimbeau, C.; Lund, O.; Aarestrup, F. M. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. Journal of Antimicrobial Chemotherapy 2020, 75 (12). [CrossRef]
- Alcock, B. P.; Raphenya, A. R.; Lau, T. T. Y.; Tsang, K. K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A. L. v.; Cheng, A. A.; Liu, S.; Min, S. Y.; Miroshnichenko, A.; Tran, H. K.; Werfalli, R. E.; Nasir, J. A.; Oloni, M.; Speicher, D. J.; Florescu, A.; Singh, B.; Faltyn, M.; Hernandez-Koutoucheva, A.; Sharma, A. N.; Bordeleau, E.; Pawlowski, A. C.; Zubyk, H. L.; Dooley, D.; Griffiths, E.; Maguire, F.; Winsor, G. L.; Beiko, R. G.; Brinkman, F. S. L.; Hsiao, W. W. L.; Domselaar, G. v.; McArthur, A. G. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res 2020, 48 (D1). [CrossRef]
- Kaas, R. S.; Leekitcharoenphon, P.; Aarestrup, F. M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS One 2014, 9 (8). [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0. Molecular Biology and Evolution. Mol Biol Evol 2016, 33 (7). [CrossRef]
- R Core Team. R Core Team (2014). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/. 2014.
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL): An Online Tool for Phylogenetic Tree Display and Annotation. Bioinformatics 2007, 23 (1). [CrossRef]
- Jesper, L.; C, E. M.; Daniel, G.; G, S. B.; R, L. A.; L, S. R. Multilocus Sequence Typing Scheme for Staphylococcus Aureus: Revision of the Gmk Locus. J Clin Microbiol 2012, 50 (7), 2538–2539. [CrossRef]
- Nguyen, L.-T.; Schmidt, H. A.; von Haeseler, A.; Minh, B. Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol Biol Evol 2015, 32 (1), 268–274. [CrossRef]
- Yu, G.; Smith, D. K.; Zhu, H.; Guan, Y.; Lam, T. T.-Y. Ggtree: An r Package for Visualization and Annotation of Phylogenetic Trees with Their Covariates and Other Associated Data. Methods Ecol Evol 2017, 8 (1), 28–36. [CrossRef]
- Hoang, D. T.; Chernomor, O.; von Haeseler, A.; Minh, B. Q.; Vinh, L. S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol Biol Evol 2018, 35 (2), 518–522. [CrossRef]
- Harding, C. R.; Schroeder, G. N.; Collins, J. W.; Frankel, G. Use of Galleria Mellonella as a Model Organism to Study Legionella Pneumophila Infection. J Vis Exp 2013, No. 81. [CrossRef]
- Bilal, S.; Volz, M. S.; Fiedler, T.; Podschun, R.; Schneider, T. Klebsiella Pneumoniae– Induced Liver Abscesses, Germany. Emerging Infectious Diseases. 2014. [CrossRef]
Figure 1.
k-loci gene arrangement of hvKP isolates. (A) ATCC 1705 (B) Kp 125 (K2/ST-231), (C) Kp124 (K1/ST-23, (D) Kp 126 (K2/ST-881) and Kp 83 (K2/ST-14). Gene names are indicated above the colored arrows and each group of genes with shared function or operon are colored with same colors. Chromosomal genes are conserved in all isolates including capsular synthesis, capsule translocation and surface assembly genes galF, ORF2, wzi, wza, wzb, wzc, and gnd. wcoZ is uniquely found in K. pneumoniae ATCC 1705 control strain and one additional isolate from our study (Kp102) Supplementary [Figure 5]. GDP-D-mannose synthesis genes manC and manB (C and D), and glucose conversion genes rmlA, rmlB, rmlC and rmlD (B and C).
Figure 1.
k-loci gene arrangement of hvKP isolates. (A) ATCC 1705 (B) Kp 125 (K2/ST-231), (C) Kp124 (K1/ST-23, (D) Kp 126 (K2/ST-881) and Kp 83 (K2/ST-14). Gene names are indicated above the colored arrows and each group of genes with shared function or operon are colored with same colors. Chromosomal genes are conserved in all isolates including capsular synthesis, capsule translocation and surface assembly genes galF, ORF2, wzi, wza, wzb, wzc, and gnd. wcoZ is uniquely found in K. pneumoniae ATCC 1705 control strain and one additional isolate from our study (Kp102) Supplementary [Figure 5]. GDP-D-mannose synthesis genes manC and manB (C and D), and glucose conversion genes rmlA, rmlB, rmlC and rmlD (B and C).
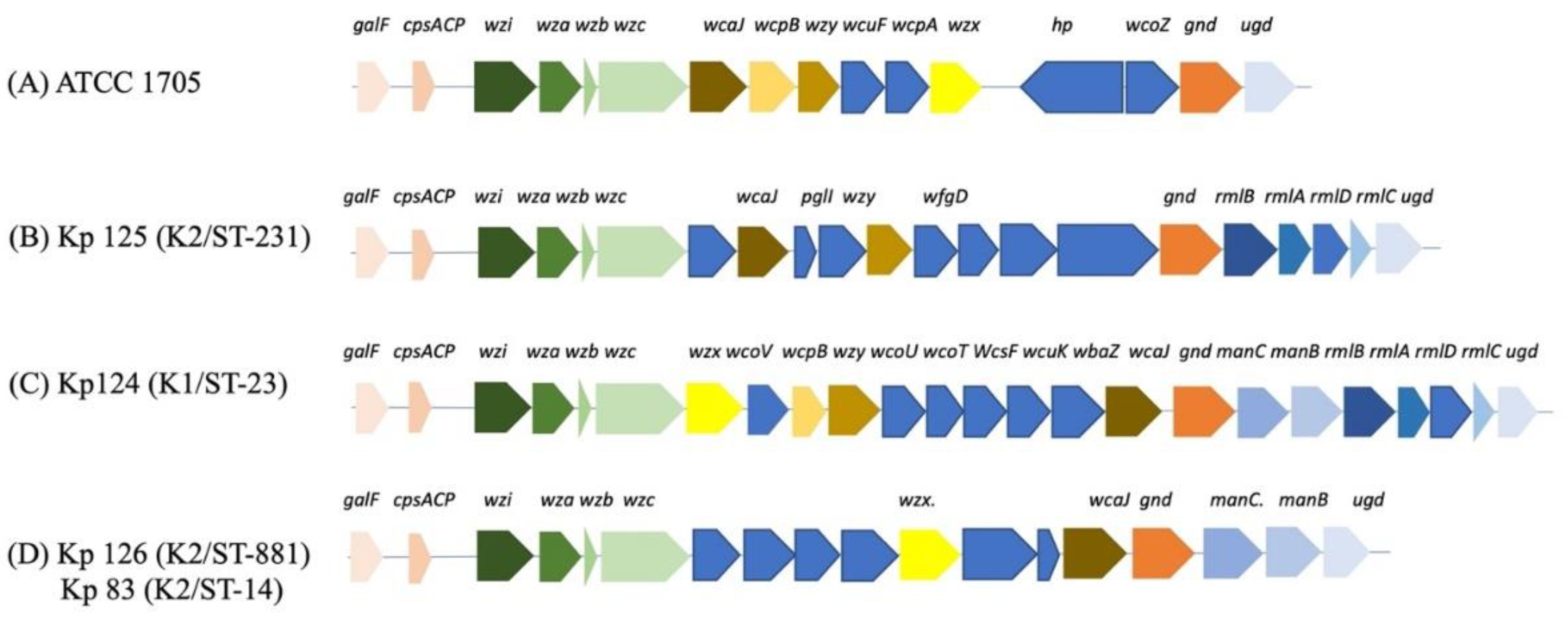
Figure 2.
Bactericidal activity of human serum samples against hvKp clinical strains. The CFU counts of all strains including the control were tested at four time points, T=0, T=1, T=2 and T=3 (time in hours). Each strain represents mean for three replicates, K. pneumoniae ATCC 1705 (control), K1/ST-23(Kp 124), K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83). P-value of 0.0007 for CFU counts which is statistically significant as shown by One-way ANOVA. Error bars represents SEM. A slight dip in the bacterial growth at T=2 is followed by a slight to significant increase in the bacterial growth in all three replicates at T=3, For Kp125, Kp 126 and Kp 38.
Figure 2.
Bactericidal activity of human serum samples against hvKp clinical strains. The CFU counts of all strains including the control were tested at four time points, T=0, T=1, T=2 and T=3 (time in hours). Each strain represents mean for three replicates, K. pneumoniae ATCC 1705 (control), K1/ST-23(Kp 124), K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83). P-value of 0.0007 for CFU counts which is statistically significant as shown by One-way ANOVA. Error bars represents SEM. A slight dip in the bacterial growth at T=2 is followed by a slight to significant increase in the bacterial growth in all three replicates at T=3, For Kp125, Kp 126 and Kp 38.
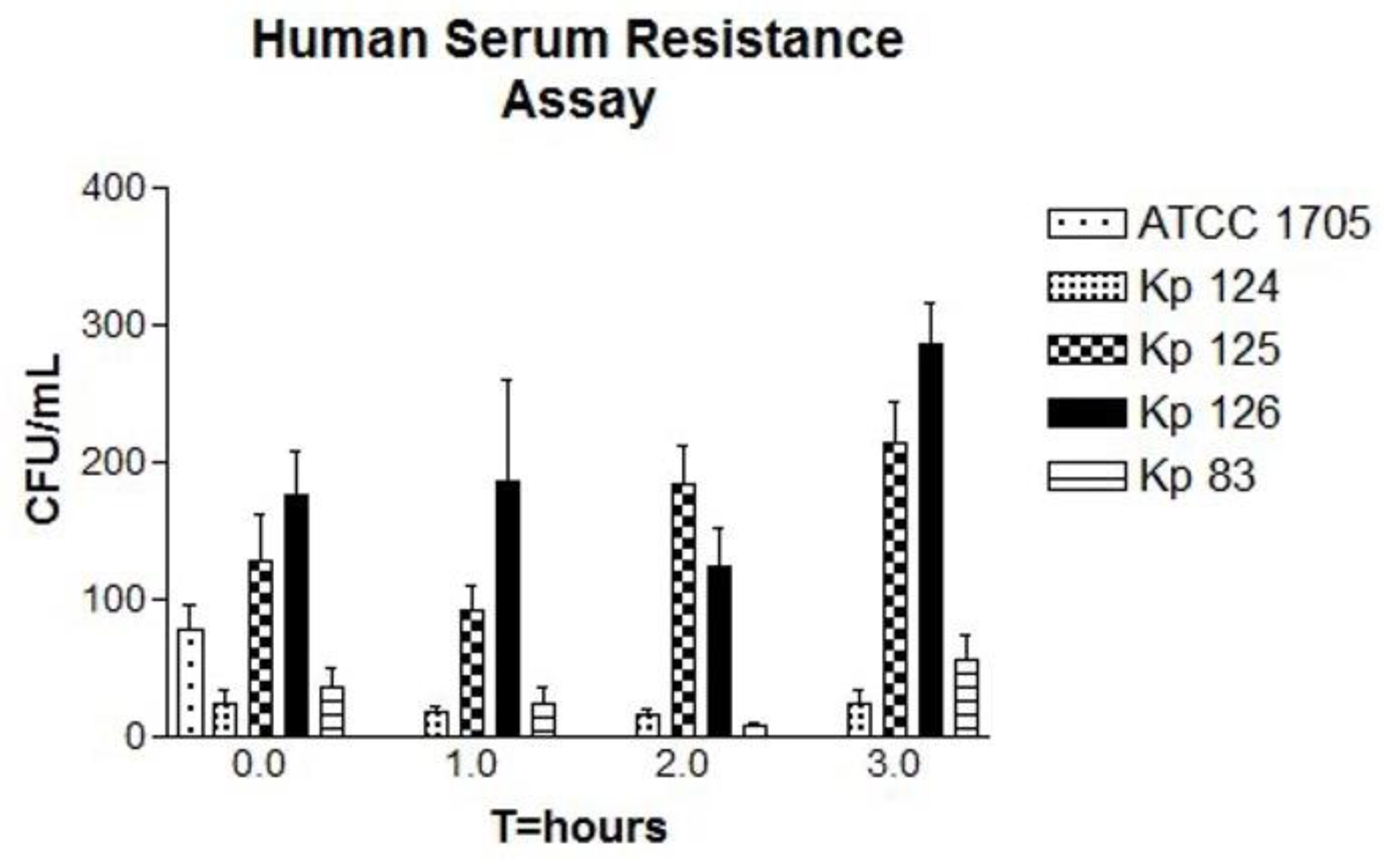
Figure 3.
Kaplan-Meier survival curve of K. pneumoniae clinical isolates. (A) Phenotypic characteristics of G. mellonella larvae pre and post injection and throughout the 5 observation days. (B, C and D) Survival of G. mellonella larvae infected with K1/ST-23(Kp 124), K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83) in comparison to controls 1 and 2 over the course of 5 days. Control 1 – untouched, uninfected larvae is represented in red. Control 2 – larvae injected with PBS is represented in blue. A third control, Kp 99 in replicate (C) and (D), was used which was a non-hypervirulent K. pneumoniae isolate just to compare the rate of survival to the hypervirulent strains. .
Figure 3.
Kaplan-Meier survival curve of K. pneumoniae clinical isolates. (A) Phenotypic characteristics of G. mellonella larvae pre and post injection and throughout the 5 observation days. (B, C and D) Survival of G. mellonella larvae infected with K1/ST-23(Kp 124), K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83) in comparison to controls 1 and 2 over the course of 5 days. Control 1 – untouched, uninfected larvae is represented in red. Control 2 – larvae injected with PBS is represented in blue. A third control, Kp 99 in replicate (C) and (D), was used which was a non-hypervirulent K. pneumoniae isolate just to compare the rate of survival to the hypervirulent strains. .
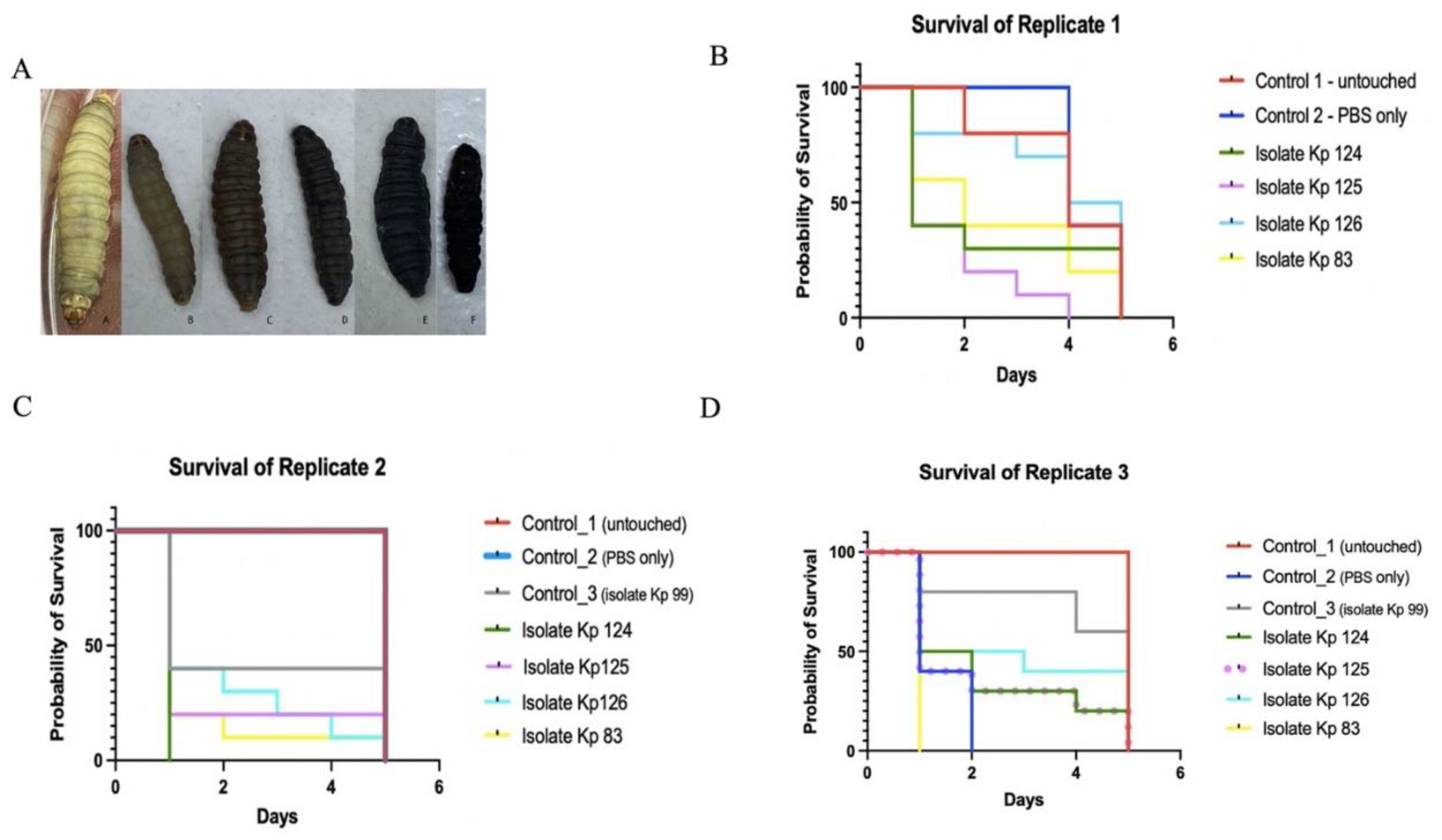
Figure 4.
A whole-genome single nucleotide polymorphisms (SNPs) calling phylogenetic tree of K. pneumoniae isolates (A) (n= 131). A maximum-likelihood tree showing genetic relatedness between K. pneumoniae strains based on WGS data. The strains are labelled with serial numbers and grouped according to their STs. K. pneumoniae RJF293 (GenBank accession number CP014008) was used as a reference. The isolates starting with NZ and CP (in black) are global published isolates retrieved from Genbank, were used for comparison. The isolates in blue are strains from this study. The hypervirulent isolates are pointed at with arrows; red for K2/ST-881 (Kp126), green for K2/ST-231 (Kp125), purple for K1/ST-23(Kp 124) and pink for K2/ST-14 (Kp83). iTOL was used to visualize and annotate the tree. Clades are indicated in A, B, C and D. This figure compares the local strains in this study (represented in blue) as well as global strains, which clearly demonstrates the diverse clonality of clinical isolates from Oman, belonging to various STs.
Figure 4.
A whole-genome single nucleotide polymorphisms (SNPs) calling phylogenetic tree of K. pneumoniae isolates (A) (n= 131). A maximum-likelihood tree showing genetic relatedness between K. pneumoniae strains based on WGS data. The strains are labelled with serial numbers and grouped according to their STs. K. pneumoniae RJF293 (GenBank accession number CP014008) was used as a reference. The isolates starting with NZ and CP (in black) are global published isolates retrieved from Genbank, were used for comparison. The isolates in blue are strains from this study. The hypervirulent isolates are pointed at with arrows; red for K2/ST-881 (Kp126), green for K2/ST-231 (Kp125), purple for K1/ST-23(Kp 124) and pink for K2/ST-14 (Kp83). iTOL was used to visualize and annotate the tree. Clades are indicated in A, B, C and D. This figure compares the local strains in this study (represented in blue) as well as global strains, which clearly demonstrates the diverse clonality of clinical isolates from Oman, belonging to various STs.

Figure 5.
A whole-genome single nucleotide polymorphisms (SNPs) calling phylogenetic tree of K. pneumoniae isolates (n= 31) and heat map of resistance-encoding genes distribution and their antimicrobial classes. A maximum-likelihood tree showing genetic relatedness between K. pneumoniae strains based on WGS results. The isolates are grouped and highlighted according to their STs (Blue = ST 395, Red= ST 231, Green= ST 405, Yellow= ST 23, Unhighlighted= miscellaneous STs). The hypervirulent isolates K1/ST-23(Kp 124, K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83) are pointed at using magenta arrows. K. pneumoniae RJF293 (GenBank accession number CP014008) reference was used. iTOL was used to visualize and annotate the tree. Hypervirulent Kp83 (K2/ST-14) and Kp124 (K1/ST-23), are branching from the same clade but belong to two different clusters, C and D, respectively. The relation between their STs and resistance phenotype indicates the diversity in the acquired antimicrobial genes in these hypervirulent strains.
Figure 5.
A whole-genome single nucleotide polymorphisms (SNPs) calling phylogenetic tree of K. pneumoniae isolates (n= 31) and heat map of resistance-encoding genes distribution and their antimicrobial classes. A maximum-likelihood tree showing genetic relatedness between K. pneumoniae strains based on WGS results. The isolates are grouped and highlighted according to their STs (Blue = ST 395, Red= ST 231, Green= ST 405, Yellow= ST 23, Unhighlighted= miscellaneous STs). The hypervirulent isolates K1/ST-23(Kp 124, K2/ST-231 (Kp125), K2/ST-881 (Kp126) and K2/ST-14 (Kp83) are pointed at using magenta arrows. K. pneumoniae RJF293 (GenBank accession number CP014008) reference was used. iTOL was used to visualize and annotate the tree. Hypervirulent Kp83 (K2/ST-14) and Kp124 (K1/ST-23), are branching from the same clade but belong to two different clusters, C and D, respectively. The relation between their STs and resistance phenotype indicates the diversity in the acquired antimicrobial genes in these hypervirulent strains.
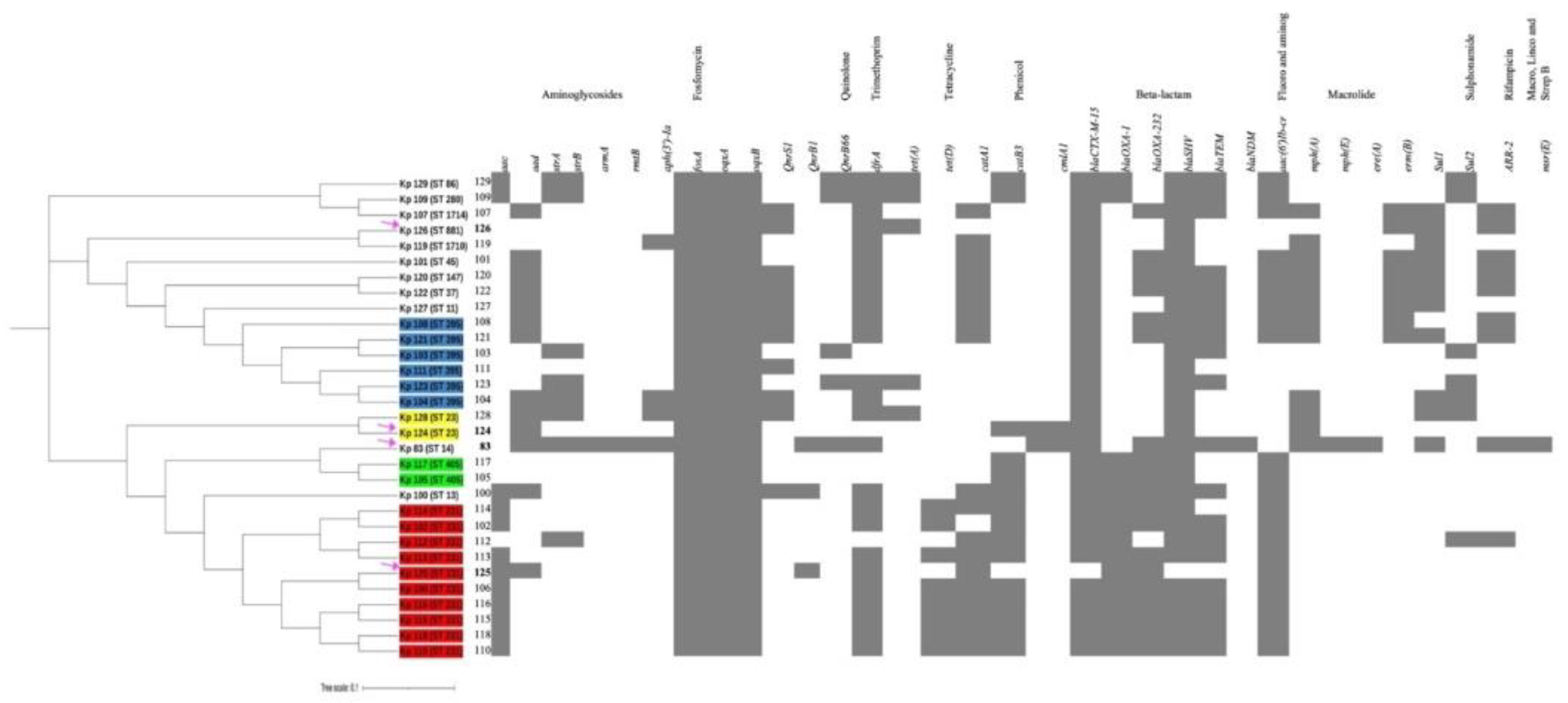

Table 1.
Sequence types of local K. pneumoniae clinical isolates (n=31).
| Isolate (Kp) | Sequence type | Percentage (%) |
|---|---|---|
| 102, 106, 110, 112, 113, 114, 115, 116, 118, 125 | 231 | 32 |
| 103, 104, 108, 111,121, 123 | 395 | 19 |
| 124 and 128 | 23 | 6 |
| 105 and 117 | 405 | 6 |
| 100 | 13 | 1 |
| 101 | 45 | 1 |
| 107 | 1714 | 1 |
| 109 | 280 | 1 |
| 119 | 1710 | 1 |
| 120 | 147 | 1 |
| 122 | 37 | 1 |
| 126 | 881 | 1 |
| 127 | 11 | 1 |
| 83 | 14 | 1 |
| 129 | 86 | 1 |
Table 2.
Source and molecular genotype of clinical hypervirulent Klebsiella pneumoniae isolates.
| Isolate | Serotype | ST | Source | Cluster |
|---|---|---|---|---|
| Kp 83 | K2 | 14 | Urine | D |
| Kp 124 | K1 | 23 | Pus | C |
| Kp 125 | K2 | 231 | Urine | B |
| Kp 126 | K2 | 881 | Urine | A |
Table 3.
Hypervirulent Klebsiella pneumoniae genotype and phenotype.
| Strain name, serotype, and ST | Plasmid | Resistance Phenotype | Resistance genotype | Resistance Mechanism | Drug Class |
|---|---|---|---|---|---|
| Kp 83 K2/ST-14 | IncFIA (HI1) 204,189 |
ESBL, | tet (B), tetR | Antibiotic efflux, target alteration | Tetracycline |
| AMP, TZP, FEP, CTX, IMP, CAZ, CIP | |||||
| IncFIB(K) 208,191 bp |
catI | Antibiotic inactivation | Phenicol | ||
| drfA12 | Antibiotic target replacement | Folate pathway antagonist | |||
| aadA2 | Antibiotic inactivation | Aminoglycoside | |||
| qacEdelta1 | Efflux | Disinfecting agents and intercalating dyes | |||
| sul1 | Target replacement | Sulfonamides | |||
| mphA | Antibiotic inactivation | Macrolides | |||
| IncFIB(pNDM-Mar) 372 826 bp |
OXA-1 | Antibiotic inactivation | Carbapenem, Cephalosporin | ||
| NDM-1 | Antibiotic inactivation | Carbapenem, Cephalosporin | |||
| QnrB1 | Target protection | Quinolones | |||
| catI | Antibiotic inactivation | Phenicol | |||
| CTX-M-15 | Antibiotic inactivation | Cephalosporin | |||
| AAC(6’)-lb-cr6 | Antibiotic inactivation | Quinolones, Aminoglycoside | |||
| IncR 68 649 bp |
tet(D) | Efflux | Tetracycline | ||
| sul2 | Target replacement | Sulfonamides | |||
| drfA14 | Target replacement | Folate pathway antagonist | |||
| SHV-2 | Inactivation | Carbapenem, Cephalosporin | |||
| QnrS1 | Target protection | Quinolones | |||
| APH(6)-Id | Inactivation | Aminoglycoside | |||
| Kp125 K2/ST-231 | ColKP3 5095 bp |
KPC | OXA-181 | Antibiotic Inactivation | Carbapenem, Cephalosporin |
| IncFIB(pQil) 115.300 bp |
AMP, TZP, FEP, CTX, CAZ | KPC-3 | Antibiotic Inactivation | Carbapenem, Cephalosporin | |
| TEM-1 | Antibiotic Inactivation | Carbapenem, Cephalosporin | |||
| APH(3’)-la | Antibiotic Inactivation | Aminoglycoside | |||
| IncFII(pAMA1167-NDM-5) 175,879 bp |
sul1 | Target replacement | Sulfonamide | ||
| qacEdelta1 | Efflux | Disinfecting agents and intercalating dyes | |||
| aadA5 | Antibiotic Inactivation | Aminoglycoside | |||
| NDM-5 | Antibiotic Inactivation | Carbapenem, Cephalosporin | |||
| mphA | Antibiotic Inactivation | Macrolides | |||
| drA17 | Target replacement | Folate pathway antagonist | |||
| tetR | Antibiotic efflux, target alteration | Tetracylines | |||
| Kp124 K1/ST-23 | IncFIB(pQil) 115.300 bp |
ESBL/KPC | KPC-3 | Antibiotic Inactivation | Carbapenem, Cephalosporin |
| AMP, TZP, FEP, CTX, FOX, CAZ | |||||
| Kp126 K2/ST-881 | IncFIB(K) 208,191 bp |
ESBL, | catI | Antibiotic inactivation | Phenicol |
| AMP, FEP, CTX, IMP, CAZ, CIP | drfA12 | Antibiotic target replacement | Folate pathway antagonist | ||
| aadA2 | Antibiotic inactivation | Aminoglycoside | |||
| qacEdelta1 | Efflux | Disinfecting agents and intercalating dyes | |||
| sul1 | Target replacement | Sulfonamides | |||
| mphA | Antibiotic inactivation | Macrolides |
Table 4.
Interpretive categories and zone diameter breakpoints according to CLSI guidelines.
| Antibiotic | Abbreviation | Disk content (μg) | Zone diameter breakpoints (mm) | ||
| Susceptible | Intermediate | Resistant | |||
| Ampicilin | AMP | 10 | ≥ 17 | 14-16 | ≤ 13 |
| Piperacillin-tazobactam | TZP | 110 | ≥ 21 | 18-20 | ≤ 17 |
| Cefepime | FEP | 30 | ≥ 25 | 19-24 | ≤ 18 |
| Cefotaxime | CTX | 30 | ≥ 26 | 23-25 | ≤ 22 |
| Cefoxitin | FOX | 30 | ≥18 | 15-17 | ≤ 14 |
| Ceftazidime | CAZ | 30 | ≥ 21 | 18-20 | ≤ 17 |
| Imipenem | IMP | 10 | ≥ 23 | 20-22 | ≤ 19 |
| Meropenem | MEM | 10 | ≥ 23 | 20-22 | ≤ 18 |
| Gentamicin | CN | 30 | ≥ 15 | 13-14 | ≤ 12 |
| Amikacin | AK | 10 | ≥ 17 | 15-16 | ≤ 14 |
| Ciprofloxacin | CIP | 5 | ≥ 31 | 21-30 | ≤ 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



