Submitted:
08 January 2024
Posted:
09 January 2024
You are already at the latest version
Abstract
This review explores the transformative role of induced pluripotent stem cells (iPSCs) and organoids in advancing neuropathology research. Focused on Alzheimer's disease, epilepsy, Parkinson's disease, and spinal cord injury, iPSCs offer patient-specific disease modeling with regenerative therapy potential. The versatility of iPSCs, their ability to differentiate into the main neuronal cell types, and their integration into three-dimensional organoid models enable the recreation of complex tissues in vitro. Improvement of organoid and iPSCs generation protocols and the selection of appropriate donor cell types are highlighted as crucial steps toward the application of these new technologies to overcome tumorigenic potential and other challenges. iPSCs demonstrate promise in regenerative therapies, as evidenced by successful applications in animal models.
Keywords:
iPSCs
; Organoids
; Alzheimer's disease
; epilepsy
; Parkinson's disease
; spinal cord injury.
1. Introduction
The study of neuropathologies often presents difficulties in determining proper study models. Neurodegenerative diseases such as Alzheimer's disease (AD) and Parkinson's disease (PD) present large extracellular and intracellular factors difficulting precise study models [1]. In neurodevelopmental diseases, it becomes especially complex to investigate the initial stages of the disease while replicating it in cellular and animal models, as is the case for temporal lobe epilepsies [2,3]. For tissue injuries, such as spinal cord injury (SCI) and strokes, animal models are most study models, and treatment becomes the main challenge [4,5]. For all these cases there is a common interest in the potential development of disease-specific models or therapeutic alternatives.
Induced pluripotent stem cells (iPSCs) were first generated from mouse fibroblasts [6]. Subsequent studies confirmed the ability to differentiate these cells into the main neuronal cell types, including neurons, astrocytes, and oligodendrocytes [7]. The interest in the use of iPSCs in the field of neuropathology is due to the modeling and treatment of rare and genetically altered diseases. So iPSCs have presented the possibility for a cell patient-derived disease modeling, therefore presenting somatic mutations of interest and mimeting disease phenotype, such as for AD [8]. In addition, iPSCs also present therapeutic potential in the field of regenerative therapy [9]. In this brief review, it is focused on the experimental models for neuropathologies using iPSCs, focusing on organoid models and the potential use of these cells as a therapy and in drug screening pipelines.
2. Generation and Characterization of iPSCs
Self-renewal and pluripotency stand as key characteristics of embryonic stem cells (ESCs). Self-renewal denotes the remarkable capability to continuously multiply without specifying a particular cell fate when cultured in vitro. Pluripotency, on the other hand, signifies the remarkable potential to transform into various cell types originating from the three fundamental embryonic germ layers [10,11]. The generation of iPSCs represents a remarkable feat in cellular reprogramming, offering a distinctive pathway to harness the regenerative potential of pluripotent cells.
iPSC formation was first demonstrated in the pioneering work of Shinya Yamanaka and his colleagues, who identified a set of transcription factors capable of reprogramming adult somatic cells into a pluripotent state. These factors, including Oct4, Sox2, Klf4, and c-Myc (abbreviated as OSKM), have become the hallmark of iPSC reprogramming [6,12]. In recent years, iPSCs have been used in a range of applications, including autologous cell therapy [13], experimental modeling of multiple diseases [14,15], and as platforms for drug discovery and therapeutic screening [16]. In recent years, a wide array of protocols for iPSC generation has emerged to attend to these diverse purposes. In the sections below we summarize the key differences in reprogramming techniques used to generate iPSCs.
2.1. Who Can Be Reprogrammed?
Somatic cells, which make up the vast majority of cells in the body, are specialized cells with specific functions, such as skin cells, blood cells, or nerve cells. In theory, the concept of reprogramming suggests that nearly every somatic cell in the human body has the potential to be transformed into an iPSC (Figure 1). Despite that, the efficiency and kinetics of reprogramming can vary based on the donor cell type. Fibroblasts from both mice and humans remain the most common cell types used for experimental reprogramming [17]. However, in the last years, there has been an increased interest in other cell types regarding their availability, therapeutic relevance, and ease of reprogramming. One example is CD133+ cord blood cells, which require only OCT4 and SOX2 to generate iPSCs [18]. Human primary keratinocytes can be reprogrammed two times faster and 100 times more efficiently than fibroblasts [19]. These cells also have the advantage of being obtained by culturing plucked hair from patients [20].
Urine samples can also be an easily available resource of cells that can be reprogrammed into iPSCs. Renal tubular cells and exfoliated renal epithelial cells found in urine have been successfully reprogrammed into human iPSCs (hiPSCs) [21,22]. Functional cardiomyocytes derived from urine cells in the cardiovascular system exhibited the capability to generate action potentials. This phenomenon was observed both in vitro and in vivo after reprogrammed induced pluripotent stem cells (iPSCs) underwent differentiation through lentiviral-vector gene transduction [23,24].
Another relevant source of cells for reprogramming is the blood. Peripheral Blood Mononuclear Cells (PBMCs) can be isolated from blood samples with ease and serve as one of the most popular somatic cell sources for iPSC generation [25]. Both terminally differentiated mature B and T lymphocytes gave rise to iPSCs [26,27,28], although reprogrammed T cells have been shown to induce spontaneous T cell lymphomas in mice, limiting the therapeutic applications of these cells [29]. Thus, depending on the goal it is recommended to use protocols that eliminate lymphocytes from the PBMCs [30]. One advantage of choosing PBMCs as the donor cell for reprogramming is that these cells can be cryopreserved and reprogrammed in the future without any compromise in the reprogramming kinetics and efficiency [31], allowing the use of frozen samples stored in blood banks worldwide [32].
Despite cell type, the level of differentiation and maturation also influences iPSCs generation efficiency. In mice, it has been reported that immature cells are more readily reprogrammed than terminally differentiated cells [33]. Another study demonstrated that hematopoietic stem and progenitor cells can generate up to 300 times more iPSCs colonies compared to terminally differentiated lymphocytes [34]. Therefore, selecting the appropriate cell type is a crucial factor to contemplate prior to initiating any experiment. The decision typically relies on cell accessibility and influences the necessity for external factors as well as the efficiency and kinetics of reprogramming.
2.2. The Reprogramming Recipe
The reprogramming factors OSKM are still the most used method for generating iPSCs. Still, over the recent years, multiple alternatives have been described to fine-tune the reprogramming protocols, selecting other transcription factor combinations and introducing new molecules to increase the efficiency of iPSCs generation.
Since the molecules Klf4 and c-Myc are considered proto-oncogenes, researchers searched for substitute candidates to decrease potential tumorigenic risks associated with these molecules. One of the first alternative reprogramming methods was described by Yu and colleagues, where Oct4 and Sox2 were used in combination with Nanog and Lin28 leading to a reprogramming efficiency similar to that obtained by Yamanaka’s OSKM combination [35]. An approach to differentiate mouse embryonic fibroblasts (MEFs) into iPSCs using three factors was also developed [36]. This technique included Oct4, Sox2, and the orphan nuclear receptor Esrrb, and obtained similar efficiency compared to the OSKM protocol. Subsequent studies further developed strategies to reprogram cells using only two transcription factors, including multiple combinations of Oct3/4, Sox2, Klf4, and c-Myc [37,38,39,40,41]. (Figure 1).
Other molecules known to modulate cellular processes that are relevant for iPSCs generation and maintenance have also been used to increase reprogramming efficiency. Proteins that induce proliferation, such as telomerase reverse transcriptase (TERT) and the SV40 large T antigen (SV40LT) increase the appearance of colonies when combined with OSKM [42]. Chemical compounds that also positively regulate cell cycle progression, such as mitogen-activated protein kinase kinase (MAPKK) inhibitors, increased the number of iPSCs colonies obtained from reprogrammed neural precursor cells [43]. MicroRNAs (miRNAs) are also known to influence pluripotency and reprogramming [44], and several miRNAs have been tested for their capacity to increase iPSCs generation. Among these, a number of miRNAs from the miR-290 cluster were able to increase the number of colonies following reprogramming compared to cells using the OSKM factors alone [45]. These miRNAs are believed to be downstream effectors of c-Myc signaling but induce a population of iPSCs more homogeneous compared to c-Myc [46]. Numerous cell signaling pathways are regulated by miRNAs and their potential effects on iPSCs production have been extensively reviewed elsewhere [47,48].
Epigenetic modifications, which include DNA methylation and histone modifications, regulate gene expression without changing the underlying DNA sequence. During iPSC reprogramming, these epigenetic marks are erased to resemble those of embryonic stem cells (ESCs) [49,50]. This resetting of the epigenetic landscape is essential for the successful conversion of differentiated cells into pluripotent stem cells [51]. Therefore, applying chemical molecules that regulate DNA methylation or chromatin modifications can improve reprogramming in many cell types [51,52,53]. Treatment with histone deacetylase (HDACs) inhibitors, including hydroxamic acid (SAHA), trichostatin A (TSA), valproic acid (VPA), and butyrate improves reprogramming in MEFs [51,54,55]. VPA also induced pluripotency in dermal fibroblasts and neonatal human foreskin fibroblasts (HFFs) in combination with Oct4 and Sox2 [56].
In summary, identifying molecules that enhance pluripotency and maintain stem cell states is crucial due to the low success rates in current iPSC generation protocols. Considering the cell's transcriptome and epigenetic profile is essential for selecting appropriate molecules, ensuring the reprogramming process yields an adequate number of pluripotent cell colonies.
2.3. Reprogramming Factors Delivery Systems
Originally, the OSKM transcription factors have been delivered into a mouse and human fibroblasts using Moloney murine leukemia virus (MMLV)-derived retroviruses [57,58]. Subsequently, reprogramming was also reported using Lentivirus-based vectors [58]. They are generally derived from HIV, present a higher cloning capacity, and allow for the infection of both dividing and non-dividing cells, usually having higher infection efficiency rates compared to MMLV-based models [59]. Furthermore, Tet-inducible lentiviruses for reprogramming enable the controlled expression of reprogramming factors [60]. Despite achieving an accepted efficiency, their integration into the host genome raised safety concerns.
Since then, an array of new delivery systems has emerged, using viral vectors that are non-integrative, including Sendai virus and adenovirus, as well as non-viral methods, including liposomes and vectors based on piggyBac transposon [61]. (Figure 1).
General delivery systems employed in iPSCs reprogramming have been extensively reviewed elsewhere [59,60,61]. Each delivery method presents both advantages and limitations, making the selection of an appropriate delivery system an important issue to resolve before proceeding to reprogram somatic cells into iPSCs.
3. Organoid Models:
3.1. What They Are, How They Work, and Applications
Organoid models are three-dimensional (3D) in vitro structures of cells under specific conditions. Such structures shape and behave in such a way that they end up expressing characteristics similar to tissues and organs such as kidneys, lungs, intestines, brain and retina, as already obtained in different studies [62,63]. (Figure 2). 3D organoids are formed from human pluripotent stem cells (hPSCs), such as iPSCs, but can be formed from ESCs [64]. In the case of organoids made from iPSCs, there is a possibility of combining the potential for self-organization and the capacity for differentiation through genetic tools, to direct these cells and structures to any specific organ, which could be fundamental in the treatment of diseases [65]. More practically, organoids can be applied in advanced therapies such as organ repair, through transplantable structures, and drug studies, as well as being used to understand pathological mechanisms of certain diseases. [66].
Regarding the use of iPSC organoids for transplantation therapies, different protocols have been developed, working with neural differentiation to treat neurological disorders such as Alzheimer's disease, Parkinson's disease, and epilepsies (Figure 2). In this case, the addition of genome editing techniques through the CRISPR/Cas9 system can help modulate gene expression, making it a promising therapeutic approach for these diseases [67].
In the field of neurosciences, neural organoid models have become important for studying aspects of the brain such as neurodegenerative diseases. This importance is because they present characteristics of human brain development that cannot be well analyzed in animal models [64,65]. The application of organoids enabled the formation of “mini-brains” with very specialized zones and structures such as radial glial cells and cerebral cortex to model human microcephaly [68,70].
This methodology has advanced in such a way that it is possible to generate highly specialized cells such as oligodendrocytes and astrocytes and that it is subsequently possible to develop even more specialized structures in the advanced stages of neural development [71,72]. In addition to 3D organoid models, 2D models are used to study brain structures, since these models form neural networks [69]. However, 3D models represent better candidates for studying and treating neurological diseases [Table 1] [67].
3.2. Applications of Organoid Models in Neurological Diseases
3.2.1. Alzheimer's Disease
Alzheimer's disease (AD) is a neurodegenerative disease characterized by progressive memory loss, problems in cognitive functions, and loss of independence in carrying out daily activities, being the main cause of dementia [73]. AD arises due to the accumulation of Tau protein and the formation of amyloid beta plaques, which are responsible for changes in the brain that result in the destruction of neurons and impairment of synapses. Because of this, the diagnosis of AD is defined by the presence of amyloid β and phosphorylated tau protein.
Another aspect of AD is its multifactorial nature, with a substantial genetic component, where the apolipoprotein E (APOE) gene and its alleles (APOE2, APOE3, and APOE4) are risk factors for Alzheimer's [74]. The APOE4 allele was particularly related to AD presence and its heritability and progression [75]. Different experimental approaches, including in vivo, in silico, and in vitro have been used to understand and possibly treat AD. However, none have managed to fully mimic the pathological features observed in the human AD brain, although much progress and positive results have been achieved [76]. Currently, there are three commonly used methods to model AD in cerebral organoids:
Aftin-5 (Aβ42 agonist): In this model, there is an induction of APP amyloid precursor protein (Aβ) using Aftin-5 (an Aβ42 inducer that increases the production and secretion of soluble extracellular amyloid peptides). Aftin-5 treatment leads to a reproducible disruption of the physiological balance between Aβ42 and Aβ40, generating an AD-like condition in human cerebral organoids [77].
Organoids derived from Familial AD (FAD): This model is achieved through the formation of iPSC-derived brain organoids from patients with FAD who carry APP duplications or have mutations in the presenilin1 (PSEN1) gene. This model can demonstrate key aspects of the pathology such as the presence of beta-amyloid plaques and tau protein, in addition to showing a timeline relating to the increase in P-tau levels [78,79]. Another variation of this same model is the use of stem cells from patients carrying a missense mutation in the PSEN1 gene linked to early-onset AD. In this case, the cerebral organoids exhibit the same Aβ and P-tau protein aggregates as the previous model [80,81].
Model with APOE3 allele: Due to the strong relationship between the apoE gene and its effects on AD, organoid models with induced mutations in this gene have been developed. This model consists of using gene editing (CRISPR/Cas9) to convert APOE3 to APOE4 in iPSCs derived from patients with sporadic (or idiopathic) Alzheimer's disease since the APOE4 variant has a greater genetic influence on AD compared to other variants. In this study, it was observed that APOE4 neurons had a greater number of synapses and Aβ42 secretion compared to APOE3 cells [82].
3.2.2. Epilepsy
Epilepsy is a diverse set of central nervous system disorders that have in common an increased tendency for seizures [83]. Seizures occur when neural networks are formed irregularly or are disturbed by abnormal structural, infectious, or metabolic problems, which end up generating abnormal firing patterns in one point of the brain (focal epilepsy), or throughout the brain (generalized epilepsy) [84]. Around 50 million people of all ages worldwide have their lives negatively affected by epilepsy [85].
The use of iPSCs-derived neurons from epileptic individuals can help better understand the molecular and pathological mechanisms in some epilepsy phenotypes [70]. Hence, it is possible to study neuronal behavior without the need for resected brain tissue. The use of editing techniques can further intensify findings in studies of iPSCs for epilepsy models [67]
For studies of diseases related to known mutations, cellular models of iPSCs become especially attractive. While for epileptogenic cortical malformations or developmental epileptic encephalopathies experimental models using organoids become attractive to generate a natural environment-like culture.
3.2.3. Huntington’s Disease
Huntington's Disease (HD) is a neurodegenerative disease caused by a CAG trinucleotide repeat increase in the huntingtin gene (HTT) [86] The HTT mutation can lead to protein aggregation, disrupting cellular processes particularly the basal ganglia and cortical regions of the brain [87,88]. The neuronal dysfunction ultimately leads to choreiform movements, psychiatric symptoms, dystonia, bradykinesia, and dementia [88,89].
Since HD is caused by a genetic mutation, current treatments primarily focus on managing symptoms through pharmacological interventions targeting mainly dopamine modulation [90,91]. Still, a better understanding of the pathophysiology and progression of the disease is needed to develop better treatment options. Multiple in vitro and in vivo models have been used to study the mechanisms of disease, including introducing the HTT mutation or inserting CAG repeats in the cells of both invertebrate and vertebrate animal models [92,93]. These models have been important in understanding the basic mechanisms of neuronal dysfunction and dysregulation of neurotransmitters but fail to recapitulate more complex and clinical manifestations of the disease.
One promising alternative has been studying the disease using cells from patients that carry the mutated gene. The lack of biological material can be overcome by introducing iPSC technology into these models. In 2008, the Daley laboratory pioneered the creation of human iPSC-based models for Huntington's disease (HD). Notably, they successfully developed the initial iPSC line from an HD patient with 72 CAG repeats. The cells were then transformed into GABAergic, DARPP32-positive neurons, highlighting the potential of iPSCs to be directed into striatal neurons, a crucial cell type susceptible to degeneration in HD [94,95,96]. Subsequently, other groups developed additional iPSCs cell lines from different patients [97,98,99].
Using iPSCs from patients to model HD has revealed many cellular alterations caused by the mutation. Genes related to DNA damage control pathways were downregulated in neurons derived from iPSCs (iNeurons) from patients with high CAG repeat mutations [100]. The malfunction of DNA damage repair systems can be connected to the somatic instability and mosaicism observed in HD [101]. HD-derived iNeurons showed multiple abnormalities in neuronal patterning [102,103] and an observed persistent mitotic population [104]. These changes in neuronal differentiation patterns can be linked to alterations in neurodevelopmental gene expression profiles linked to HD.
Changes in gene expression are believed to be one of the mechanisms that cause neurodegeneration in HD. Comparisons in gene expression from iNeurons derived from HD iPSCs and gene-corrected control lines demonstrated that transforming growth factor beta (TGF-b) pathway were upregulated in HD [105]. Studies in iNeurons form other HD patients further support this finding [106,107,108]. Recently, the National Institute of Health (NIH) formed a consortium called “HD iPSC Consortium”, to investigate gene expression and functional changes associated with HD. RNA sequencing (RNA-seq) analysis from this group reported transcriptomic alterations in numerous pathways involving development and master regulators of neurogenesis [108].
Overall, the use of iPSCs to understand the pathophysiology of HD unfolded many pathways that can be therapeutically targeted to try to control the disease. Since iPSCs can be divided into other cell subtypes from brain tissue, including microglia and astrocytes, it will be interesting to examine if the alterations observed in iNeurons are preserved in cell-cell interaction models.
3.2.4. Parkinson's Disease
Parkinson's disease (PD) is a devastating neurodegenerative disease, preferentially involving progressive loss of dopaminergic neurons in the substantia nigra, resulting in loss of dopamine and dysregulation of fine motor coordination. Ultimately, the death of dopaminergic neurons manifests clinically in parkinsonian symptoms, including bradykinesia, muscle rigidity, and resting tremors, and pathologically involves the presence of Lewy body aggregates composed of α-synuclein. Furthermore, PD is influenced by genetic aspects, where several genes have been linked to dominant or recessive familial forms of PD including SCNA, LRRK2, PINK1, PARK2 (parkin), and GBA1, as well as additional genes such as DJ-1, PARK9 (ATP13A2 ), SJ-1 and VPS35. Taking these points into account, there has been much debate about possible in vitro techniques to seek therapeutic measures and/or manipulate and understand the different aspects of PD [109].
Although there are still no therapeutic measures to modify the course of the disease, recent work using iPSC and 3D brain organoid models has provided more information about pathogenesis and potential therapeutic targets for several diseases and may be the case for PD. Among the uses, neurons from patients with PD can be used to form organoids to analyze pathogenic mechanisms in detail and test drugs against PD. Thus, several iPSC-derived neuron organoid models from PD patients carrying mutations in the genes mentioned above have been proposed to seek to understand the pathology of PD and possible therapeutic interventions [110]. Some organoid models will be presented below:
α-synuclein (SNCA) model: α-synuclein is a protein that assumes different conformations dictated by cellular stress and is involved in neurodegenerative diseases such as PD [111]. Mutations of A53T mutant α-synuclein or α-synuclein in neurons lead to increased nitrosative stress, mitochondrial dysfunction, disrupted synaptic connectivity, transcriptional changes in synaptic signaling genes, and reduced ratio of α-synuclein tetramer to monomer], important factors in the pathogenesis of PD [112]. The iPSC-derived neuron model has triplicated levels of α-synuclein and could be a good model to understand the morphophysiological divergences between healthy neurons and mutant neurons from PD patients.
LRRK2 model: Leucine-rich repeat kinase 2 (LRRK2) is a multikinase involved in roles in neurite outgrowth, phosphorylation of multiple proteins, and endocytic sorting via interactions with Rab-GTPases. Mutations in the gene encoding LRRK2 imply a significant risk for PD as well as other factors [113]. LRRK2 organoid models showed increased levels of oxidized dopamine and lysosomal receptor for chaperone-mediated autophagy. Also, neurons in this model have greater apoptotic activity, reducing neurite growth. Interestingly, LRRK2 also demonstrated irregularities in synaptic vesicle recycling, leading to disrupted synaptic vesicle endocytosis and decreased vesicle density in neurons.
PINK1 model: PINK1 (PTEN-induced kinase 1) is a phosphatase and tensin (PTEN) homologous protein/kinase. PINK1 localizes to the mitochondrial membrane after its depolarization, where it phosphorylates Parkin. Together, PINK1 and Parkin regulate mitochondrial health, and mutations in either related gene are associated with autosomal recessive diseases and early-onset forms of PD. A model with iPSC-derived neurons from patients expressing nonsense (Q456X) or missense (V170G) PINK1 exhibit mitochondrial defects, including impaired recruitment of Parkin to mitochondria [114].
GBA model: The GBA1 gene is responsible for encoding glucocerebrosidase (GCase or β-glucosidase, a lysosomal enzyme that catalyzes the hydrolysis of glucosylceramide (GlcCer) into glucose and ceramide, but also the hydrolysis of D-glucosyl-N-acylsphingosine into D-glucose and N-acylsphingosine. Studies have confirmed that there is a relationship between PD and GBA mutations including insertion, deletion, frameshift, and point mutations in GBA. Approximately 5–10% of PD patients carry GBA1 mutations. The result of the GBA mutation is the accumulation of lipids in neurons. In this PD model, alterations of GBA and GCase substrates, glycolipids glucosylceramide (GlcCer), and glucosylsphingosine (GlcSph) are found at increased levels, resulting in defective action of cellular organelles of neurons, making neurons more vulnerable to apoptosis [110].
Idiopathic Parkinson's: The neuron-based organoid model of idiopathic Parkinson's seeks to understand aspects of the disease that do not necessarily involve genetic aspects while also seeking to understand PD. Interestingly, it has been shown that neurons derived from patients with idiopathic PD have decreased mitochondrial respiration, increased levels of oxidized dopamine and oxidized DJ-1, and decreased GCase enzyme activity [115].
3.3. Limitations in the Use of Organoid Models
Even though they have numerous important uses, it is important to highlight that the use of organoid models presents certain limitations in treatment approaches for diseases. An important point to take into account is their effectiveness since they are not found in typical environments that are generally found in tissues and organs and this can be a limiting factor in treatments [66].
In addition, iPSC-derived organoids still exhibit other general deficiencies. These include lack of reproducibility, lack of specificity regarding cell type composition of a certain tissue/organ; uncontrolled size, and shape heterogeneity, absence of vascular, immunological, and innervation components and specific morphological characteristics, and lack of functionality. Based on these points, it has become a current objective to improve organoid protocols to both reduce these in vitro and organ effects and to promote better adaptation and expression of organ characteristics [65].
4. iPSCs-Based Therapies for Neurological Diseases
It is likely possible to model iPSCs into all somatic cell types [139]. Studies based on this possibility turned to modeling specific diseases at the cellular level, particularly those dependent on mutations. Patient-derived iPSCs have become of interest for high-throughput developmental-level screening of several pathologies [140]. Therefore, iPSCs have been widely used in research strategies for pharmacological screening and regenerative therapy [141].
The use of stem cell treatment strategies for regenerative therapy is objective remission of the changes present in pathogenic tissue, in addition to the possibility of recovering the function lost due to local damage [142]. The proliferation and differentiation capacity of this cell type provides tissue regeneration for diseases treated by surgical resection or loss of tissue, such as spinal cord injury (SCI) [143]. iPSCs in the context of regenerative therapy become valuable due to the patient-derived cells, which prevent tissue rejection, in addition to promoting recovery of function [144].
There are currently several studies investigating the regenerative strategies of iPSCs for neuropathologies. In animal models, studies for SCI present high success rates with recovery of motor function [145,146,147]. There have been groups that have attempted to prevent loss of function in animal models of stroke and ischemia [148,149]. In temporal lobe epilepsy, it was carried out to graft modified iPSCs to non-epileptogenic GABAergic neurons [150,151,152]. Even the use of iPSCs has become the target of study as a potential treatment of diseases by replacing tissue in which surgical intervention would not occur, such as the case of PD, metachromatic leukodystrophy (MLD), and HD [153,154,155]. Within all these pathologies, iPSCs have great regenerative and replacement potential for damaged tissue due to the possibility of editing cells to overcome the alterations resulting from the pathology [142].
Pharmacological tests carried out in in vitro assays based on different human cells present difficulties in mimicking the pathology. iPSCs provide the opportunity to generate patient-specific cellular models to investigate the cellular mechanisms of disease [156]. Despite being less used in terms of drug screening, iPSCs have properties of great value in research due to the patient-derived cells being extremely specific and the less invasive acquisition of in vitro study models [157].
The modeling of specific diseases by iPSCs has been increasing intensely given the interest in more precise investigations of pathologies. For better-understood pathologies such as AD, tests of well-established drugs have already been applied to investigate the efficiency of these models [158]. Currently, the attempt to use new drugs directly in this model is unusual, but the strategies of using iPSCs as proof of drug efficiency and the same for cellular models are growing in use [Table 2] [159].
iPSCs are still being studied to overcome their biggest difficulties. Their pluripotent capacity promotes the ability to form teratomas, which is even used to prove their nature in early stages of studies. However, its tumorigenic capacity is extremely undesirable for cell therapy. In graft therapy studies, the formation of gliomas in the brain has already been reported [159]. Since the beginning of research with iPSCs, large groups have elucidated points of great danger for these cells related to the tumorigenic capacity of transgenic C-MYC and viral integration. Therefore, current studies in the interest of applying iPSCs for grafting tend to deeply investigate changes in the generated cellular model to avoid any unwanted modification [160,161]. Despite problems identified in the application of the technique, well-established groups indicate interest in the application of hPSCs for treatment in humans [162].
5. Conclusion and Future Perspectives
The use of iPSCs in the study of neuropathologies presents a favourable approach for understanding complex diseases and exploring potential therapeutic interventions. The introduction of iPSCs marked a significant breakthrough in cellular reprogramming, allowing for the generation of patient-specific models that recapitulate the characteristics of various neurological disorders [189].
The versatility of iPSCs in differentiating into key neuronal cell types, neurons, astrocytes, and oligodendrocytes, has facilitated the development of disease models patient-derived for the highlighted conditions (AD, PD, epilepsy, and SCI). The ability to manipulate iPSCs through gene editing techniques has further expanded their utility, enabling researchers to study specific disease-related mutations [109,190].
It emphasizes the importance of selecting appropriate donor cell types and optimizing reprogramming protocols. It is fundamental to maintain continuous efforts to enhance efficiency and safety in iPSC generation. There are several concerns about its applicability. The use of iPSCs as therapy is controversial mainly due to their tumorigenesis ability. However, several studies using animal models report positive results, without tumor formation [Table 2].
The integration of iPSCs into organoid models represents a significant advancement, allowing for a 3D tissue recreation in vitro. Organoids derived from iPSCs offer a unique platform for studying complex structures, such as the brain, and have provided insights into the pathogenesis of neurodegenerative diseases like AD and PD. The detailed exploration of organoid models for each neurological condition, including specific genetic mutations and treatment approaches, demonstrates the potential of this technology in disease modeling and drug screening.
While iPSCs hold great promise for regenerative therapies, there are challenges and limitations to be addressed. Improvements in organoid protocols to enhance reproducibility, specificity, and functional characteristics. Additionally, considerations about the tumorigenic potential of iPSCs emphasize the importance of thorough investigations and safety measures in their application for cell therapy [191,192].
In summary, the comprehensive exploration of iPSCs and their applications in neurological disease modeling, organoid development, and potential therapeutic interventions provides a valuable overview of the current state of research in the field. The ongoing efforts to refine techniques and address limitations underscore the commitment to advancing iPSC-based approaches for a deeper understanding of neuropathologies and the development of innovative treatments.
Author Contributions
Conceptualization, J. G.; investigation, D. P., T. P., J. G., and F. X.; resources, J. da C.; writing—original draft preparation, D. P., T. P., J. G., and F. X.; writing—review and editing, D. P., T. P., and J. G.; supervision, D. M.; funding acquisition, J. da C. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) — Brazil, Finance Code 001.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trejo-Lopez, J.A.; Yachnis, A.T.; Prokop, S. Neuropathology of Alzheimer’s Disease. Neurotherapeutics 2022, 19, 173–185. [CrossRef]
- Curatolo, P.; Specchio, N.; Aronica, E. Advances in the Genetics and Neuropathology of Tuberous Sclerosis Complex: Edging Closer to Targeted Therapy. Lancet Neurol. 2022, 21, 843–856. [CrossRef]
- Chang, B.-L.; Chang, K.-H. Stem Cell Therapy in Treating Epilepsy. Front. Neurosci. 2022, 16, 934507. [CrossRef]
- Toman, N.G.; Grande, A.W.; Low, W.C. Neural Repair in Stroke. Cell Transplant. 2019, 28, 1123–1126. [CrossRef]
- Ribeiro, B.F.; da Cruz, B.C.; de Sousa, B.M.; Correia, P.D.; David, N.; Rocha, C.; Almeida, R.D.; Ribeiro da Cunha, M.; Marques Baptista, A.A.; Vieira, S.I. Cell Therapies for Spinal Cord Injury: A Review of the Clinical Trials and Cell-Type Therapeutic Potential. Brain J. Neurol. 2023, 146, 2672–2693. [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [CrossRef]
- Huang, Y.; Tan, S. Direct Lineage Conversion of Astrocytes to Induced Neural Stem Cells or Neurons. Neurosci. Bull. 2015, 31, 357–367. [CrossRef]
- McKinney, C.E. Using Induced Pluripotent Stem Cells Derived Neurons to Model Brain Diseases. Neural Regen. Res. 2017, 12, 1062–1067. [CrossRef]
- Paredes-Espinosa, M.B.; Paluh, J.L. Human Stem Cell-Derived Neurons and Neural Circuitry Therapeutics: Next Frontier in Spinal Cord Injury Repair. Exp. Biol. Med. Maywood NJ 2022, 247, 2142–2151. [CrossRef]
- Evans, M.J.; Kaufman, M.H. Establishment in Culture of Pluripotential Cells from Mouse Embryos. Nature 1981, 292, 154–156. [CrossRef]
- Martello, G.; Smith, A. The Nature of Embryonic Stem Cells. Annu. Rev. Cell Dev. Biol. 2014, 30, 647–675. [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [CrossRef]
- Madrid, M.; Sumen, C.; Aivio, S.; Saklayen, N. Autologous Induced Pluripotent Stem Cell-Based Cell Therapies: Promise, Progress, and Challenges. Curr. Protoc. 2021, 1, e88. [CrossRef]
- Ho, B.X.; Pek, N.M.Q.; Soh, B.-S. Disease Modeling Using 3D Organoids Derived from Human Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2018, 19, 936. [CrossRef]
- Hong, L.; Zhang, M.; Ly, O.T.; Chen, H.; Sridhar, A.; Lambers, E.; Chalazan, B.; Youn, S.-W.; Maienschein-Cline, M.; Feferman, L.; et al. Human Induced Pluripotent Stem Cell-Derived Atrial Cardiomyocytes Carrying an SCN5A Mutation Identify Nitric Oxide Signaling as a Mediator of Atrial Fibrillation. Stem Cell Rep. 2021, 16, 1542–1554. [CrossRef]
- Pasteuning-Vuhman, S.; de Jongh, R.; Timmers, A.; Pasterkamp, R.J. Towards Advanced iPSC-Based Drug Development for Neurodegenerative Disease. Trends Mol. Med. 2021, 27, 263–279. [CrossRef]
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem Cell Rev. Rep. 2020, 16, 3–32. [CrossRef]
- Giorgetti, A.; Montserrat, N.; Aasen, T.; Gonzalez, F.; Rodríguez-Pizà, I.; Vassena, R.; Raya, A.; Boué, S.; Barrero, M.J.; Corbella, B.A.; et al. Generation of Induced Pluripotent Stem Cells from Human Cord Blood Using OCT4 and SOX2. Cell Stem Cell 2009, 5, 353–357. [CrossRef]
- Aasen, T.; Raya, A.; Barrero, M.J.; Garreta, E.; Consiglio, A.; Gonzalez, F.; Vassena, R.; Bilić, J.; Pekarik, V.; Tiscornia, G.; et al. Efficient and Rapid Generation of Induced Pluripotent Stem Cells from Human Keratinocytes. Nat. Biotechnol. 2008, 26, 1276–1284. [CrossRef]
- Aasen, T.; Belmonte, J.C.I. Isolation and Cultivation of Human Keratinocytes from Skin or Plucked Hair for the Generation of Induced Pluripotent Stem Cells. Nat. Protoc. 2010, 5, 371–382. [CrossRef]
- Zhou, T.; Benda, C.; Dunzinger, S.; Huang, Y.; Ho, J.C.; Yang, J.; Wang, Y.; Zhang, Y.; Zhuang, Q.; Li, Y.; et al. Generation of Human Induced Pluripotent Stem Cells from Urine Samples. Nat. Protoc. 2012, 7, 2080–2089. [CrossRef]
- Xue, Y.; Cai, X.; Wang, L.; Liao, B.; Zhang, H.; Shan, Y.; Chen, Q.; Zhou, T.; Li, X.; Hou, J.; et al. Generating a Non-Integrating Human Induced Pluripotent Stem Cell Bank from Urine-Derived Cells. PLOS ONE 2013, 8, e70573. [CrossRef]
- Jiang, Y.-F.; Chen, M.; Zhang, N.-N.; Yang, H.-J.; Rui, Q.; Zhou, Y.-F. In Vitro and in Vivo Differentiation of Induced Pluripotent Stem Cells Generated from Urine-Derived Cells into Cardiomyocytes. Biol. Open 2018, 7, bio029157. [CrossRef]
- Scalise, M.; Marino, F.; Salerno, L.; Cianflone, E.; Molinaro, C.; Salerno, N.; De Angelis, A.; Viglietto, G.; Urbanek, K.; Torella, D. From Spheroids to Organoids: The Next Generation of Model Systems of Human Cardiac Regeneration in a Dish. Int. J. Mol. Sci. 2021, 22, 13180. [CrossRef]
- Su, R.-J.; Baylink, D.J.; Neises, A.; Kiroyan, J.B.; Meng, X.; Payne, K.J.; Tschudy-Seney, B.; Duan, Y.; Appleby, N.; Kearns-Jonker, M.; et al. Efficient Generation of Integration-Free Ips Cells from Human Adult Peripheral Blood Using BCL-XL Together with Yamanaka Factors. PloS One 2013, 8, e64496. [CrossRef]
- Hanna, J.; Markoulaki, S.; Schorderet, P.; Carey, B.W.; Beard, C.; Wernig, M.; Creyghton, M.P.; Steine, E.J.; Cassady, J.P.; Foreman, R.; et al. Direct Reprogramming of Terminally Differentiated Mature B Lymphocytes to Pluripotency. Cell 2008, 133, 250–264. [CrossRef]
- Nagano, S.; Maeda, T.; Ichise, H.; Kashima, S.; Ohtaka, M.; Nakanishi, M.; Kitawaki, T.; Kadowaki, N.; Takaori-Kondo, A.; Masuda, K.; et al. High Frequency Production of T Cell-Derived iPSC Clones Capable of Generating Potent Cytotoxic T Cells. Mol. Ther. Methods Clin. Dev. 2020, 16, 126–135. [CrossRef]
- Suwanpitak, S.; Promnakhon, N.; Netsrithong, R.; Wattanapanitch, M. Efficient Generation of iPSC-Derived Hematoendothelial Progenitors and Specification Toward T Cell Lineage. Methods Mol. Biol. Clifton NJ 2022, 2454, 423–442. [CrossRef]
- Serwold, T.; Hochedlinger, K.; Swindle, J.; Hedgpeth, J.; Jaenisch, R.; Weissman, I.L. T-Cell Receptor-Driven Lymphomagenesis in Mice Derived from a Reprogrammed T Cell. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 18939–18943. [CrossRef]
- Dowey, S.N.; Huang, X.; Chou, B.-K.; Ye, Z.; Cheng, L. Generation of Integration-Free Human Induced Pluripotent Stem Cells from Postnatal Blood Mononuclear Cells by Plasmid Vector Expression. Nat. Protoc. 2012, 7, 2013–2021. [CrossRef]
- Zhou, H.; Martinez, H.; Sun, B.; Li, A.; Zimmer, M.; Katsanis, N.; Davis, E.E.; Kurtzberg, J.; Lipnick, S.; Noggle, S.; et al. Rapid and Efficient Generation of Transgene-Free iPSC from a Small Volume of Cryopreserved Blood. Stem Cell Rev. Rep. 2015, 11, 652–665. [CrossRef]
- Staerk, J.; Dawlaty, M.M.; Gao, Q.; Maetzel, D.; Hanna, J.; Sommer, C.A.; Mostoslavsky, G.; Jaenisch, R. Reprogramming of Human Peripheral Blood Cells to Induced Pluripotent Stem Cells. Cell Stem Cell 2010, 7, 20–24. [CrossRef]
- González, F.; Boué, S.; Izpisúa Belmonte, J.C. Methods for Making Induced Pluripotent Stem Cells: Reprogramming à La Carte. Nat. Rev. Genet. 2011, 12, 231–242. [CrossRef]
- Eminli, S.; Foudi, A.; Stadtfeld, M.; Maherali, N.; Ahfeldt, T.; Mostoslavsky, G.; Hock, H.; Hochedlinger, K. Differentiation Stage Determines Potential of Hematopoietic Cells for Reprogramming into Induced Pluripotent Stem Cells. Nat. Genet. 2009, 41, 968–976. [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [CrossRef]
- Feng, B.; Ng, J.-H.; Heng, J.-C.D.; Ng, H.-H. Molecules That Promote or Enhance Reprogramming of Somatic Cells to Induced Pluripotent Stem Cells. Cell Stem Cell 2009, 4, 301–312. [CrossRef]
- Kim, J.B.; Zaehres, H.; Wu, G.; Gentile, L.; Ko, K.; Sebastiano, V.; Araúzo-Bravo, M.J.; Ruau, D.; Han, D.W.; Zenke, M.; et al. Pluripotent Stem Cells Induced from Adult Neural Stem Cells by Reprogramming with Two Factors. Nature 2008, 454, 646–650. [CrossRef]
- Huangfu, D.; Osafune, K.; Maehr, R.; Guo, W.; Eijkelenboom, A.; Chen, S.; Muhlestein, W.; Melton, D.A. Induction of Pluripotent Stem Cells from Primary Human Fibroblasts with Only Oct4 and Sox2. Nat. Biotechnol. 2008, 26, 1269–1275. [CrossRef]
- Kim, J.B.; Greber, B.; Araúzo-Bravo, M.J.; Meyer, J.; Park, K.I.; Zaehres, H.; Schöler, H.R. Direct Reprogramming of Human Neural Stem Cells by OCT4. Nature 2009, 461, 649–653. [CrossRef]
- Kim, J.B.; Sebastiano, V.; Wu, G.; Araúzo-Bravo, M.J.; Sasse, P.; Gentile, L.; Ko, K.; Ruau, D.; Ehrich, M.; van den Boom, D.; et al. Oct4-Induced Pluripotency in Adult Neural Stem Cells. Cell 2009, 136, 411–419. [CrossRef]
- Tsai, S.Y.; Bouwman, B.A.; Ang, Y.S.; Kim, S.J.; Lee, D.F.; Lemischka, I.R.; Rendl, M. Single Transcription Factor Reprogramming of Hair Follicle Dermal Papilla Cells to Induced Pluripotent Stem Cells. Stem Cells 2011, 29, 964–971. [CrossRef]
- Park, I.-H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of Human Somatic Cells to Pluripotency with Defined Factors. Nature 2008, 451, 141–146. [CrossRef]
- Lee, J.; Park, Y.-J.; Jung, H. Protein Kinases and Their Inhibitors in Pluripotent Stem Cell Fate Regulation. Stem Cells Int. 2019, 2019, e1569740. [CrossRef]
- Li, N.; Long, B.; Han, W.; Yuan, S.; Wang, K. microRNAs: Important Regulators of Stem Cells. Stem Cell Res. Ther. 2017, 8, 110. [CrossRef]
- Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple Targets of miR-302 and miR-372 Promote Reprogramming of Human Fibroblasts to Induced Pluripotent Stem Cells. Nat. Biotechnol. 2011, 29, 443–448. [CrossRef]
- Çağlayan, E.S.; Güran, Ş. Importance of Myc-Related microRNAs in Induced Pluripotency. Cell Biol. Int. 2015, 39, 987–994. [CrossRef]
- Lakshmipathy, U.; Davila, J.; Hart, R.P. miRNA in Pluripotent Stem Cells. Regen. Med. 2010, 5, 545–555. [CrossRef]
- Gomes, K.M.S.; Costa, I.C.; Santos, J.F.D.; Dourado, P.M.M.; Forni, M.F.; Ferreira, J.C.B. Induced Pluripotent Stem Cells Reprogramming: Epigenetics and Applications in the Regenerative Medicine. Rev. Assoc. Medica Bras. 1992 2017, 63, 180–189. [CrossRef]
- Brix, J.; Zhou, Y.; Luo, Y. The Epigenetic Reprogramming Roadmap in Generation of iPSCs from Somatic Cells. J. Genet. Genomics Yi Chuan Xue Bao 2015, 42, 661–670. [CrossRef]
- van den Hurk, M.; Kenis, G.; Bardy, C.; van den Hove, D.L.; Gage, F.H.; Steinbusch, H.W.; Rutten, B.P. Transcriptional and Epigenetic Mechanisms of Cellular Reprogramming to Induced Pluripotency. Epigenomics 2016, 8, 1131–1149. [CrossRef]
- Papp, B.; Plath, K. Epigenetics of Reprogramming to Induced Pluripotency. Cell 2013, 152, 1324–1343. [CrossRef]
- Huangfu, D.; Maehr, R.; Guo, W.; Eijkelenboom, A.; Snitow, M.; Chen, A.E.; Melton, D.A. Induction of Pluripotent Stem Cells by Defined Factors Is Greatly Improved by Small-Molecule Compounds. Nat. Biotechnol. 2008, 26, 795–797. [CrossRef]
- Mali, P.; Chou, B.-K.; Yen, J.; Ye, Z.; Zou, J.; Dowey, S.; Brodsky, R.A.; Ohm, J.E.; Yu, W.; Baylin, S.B.; et al. Butyrate Greatly Enhances Derivation of Human Induced Pluripotent Stem Cells by Promoting Epigenetic Remodeling and the Expression of Pluripotency-Associated Genes. STEM CELLS 2010, 28, 713–720. [CrossRef]
- Kang, S.-J.; Park, Y.-I.; So, B.; Kang, H.-G. Sodium Butyrate Efficiently Converts Fully Reprogrammed Induced Pluripotent Stem Cells from Mouse Partially Reprogrammed Cells. Cell. Reprogramming 2014, 16, 345–354. [CrossRef]
- Shi, Y.; Desponts, C.; Do, J.T.; Hahm, H.S.; Schöler, H.R.; Ding, S. Induction of Pluripotent Stem Cells from Mouse Embryonic Fibroblasts by Oct4 and Klf4 with Small-Molecule Compounds. Cell Stem Cell 2008, 3, 568–574. [CrossRef]
- Kitamura, T.; Koshino, Y.; Shibata, F.; Oki, T.; Nakajima, H.; Nosaka, T.; Kumagai, H. Retrovirus-Mediated Gene Transfer and Expression Cloning: Powerful Tools in Functional Genomics. Exp. Hematol. 2003, 31, 1007–1014. [CrossRef]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of Pluripotent Stem Cells from Fibroblast Cultures. Nat. Protoc. 2007, 2, 3081–3089. [CrossRef]
- Varas, F.; Stadtfeld, M.; de Andres-Aguayo, L.; Maherali, N.; di Tullio, A.; Pantano, L.; Notredame, C.; Hochedlinger, K.; Graf, T. Fibroblast-Derived Induced Pluripotent Stem Cells Show No Common Retroviral Vector Insertions. Stem Cells Dayt. Ohio 2009, 27, 300–306. [CrossRef]
- Yao, S.; Sukonnik, T.; Kean, T.; Bharadwaj, R.R.; Pasceri, P.; Ellis, J. Retrovirus Silencing, Variegation, Extinction, and Memory Are Controlled by a Dynamic Interplay of Multiple Epigenetic Modifications. Mol. Ther. J. Am. Soc. Gene Ther. 2004, 10, 27–36. [CrossRef]
- Sommer, C.A.; Stadtfeld, M.; Murphy, G.J.; Hochedlinger, K.; Kotton, D.N.; Mostoslavsky, G. Induced Pluripotent Stem Cell Generation Using a Single Lentiviral Stem Cell Cassette. Stem Cells Dayt. Ohio 2009, 27, 543–549. [CrossRef]
- Woltjen, K.; Michael, I.P.; Mohseni, P.; Desai, R.; Mileikovsky, M.; Hämäläinen, R.; Cowling, R.; Wang, W.; Liu, P.; Gertsenstein, M.; et al. piggyBac Transposition Reprograms Fibroblasts to Induced Pluripotent Stem Cells. Nature 2009, 458, 766–770. [CrossRef]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [CrossRef]
- Lee, C.-T.; Bendriem, R.M.; Wu, W.W.; Shen, R.-F. 3D Brain Organoids Derived from Pluripotent Stem Cells: Promising Experimental Models for Brain Development and Neurodegenerative Disorders. J. Biomed. Sci. 2017, 24, 59. [CrossRef]
- Andrews, M.G.; Kriegstein, A.R. Challenges of Organoid Research. Annu. Rev. Neurosci. 2022, 45, 23–39. [CrossRef]
- Garreta, E.; Kamm, R.D.; Chuva De Sousa Lopes, S.M.; Lancaster, M.A.; Weiss, R.; Trepat, X.; Hyun, I.; Montserrat, N. Rethinking Organoid Technology through Bioengineering. Nat. Mater. 2021, 20, 145–155. [CrossRef]
- Turhan, A.G.; Hwang, J.W.; Chaker, D.; Tasteyre, A.; Latsis, T.; Griscelli, F.; Desterke, C.; Bennaceur-Griscelli, A. iPSC-Derived Organoids as Therapeutic Models in Regenerative Medicine and Oncology. Front. Med. 2021, 8.
- Niu, W.; Parent, J.M. Modeling Genetic Epilepsies in a Dish. Dev. Dyn. 2020, 249, 56–75. [CrossRef]
- Camp, J.G.; Badsha, F.; Florio, M.; Kanton, S.; Gerber, T.; Wilsch-Bräuninger, M.; Lewitus, E.; Sykes, A.; Hevers, W.; Lancaster, M.; et al. Human Cerebral Organoids Recapitulate Gene Expression Programs of Fetal Neocortex Development. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 15672–15677. [CrossRef]
- Wray, S. Modelling Neurodegenerative Disease Using Brain Organoids. Semin. Cell Dev. Biol. 2021, 111, 60–66. [CrossRef]
- Hirose, S.; Tanaka, Y.; Shibata, M.; Kimura, Y.; Ishikawa, M.; Higurashi, N.; Yamamoto, T.; Ichise, E.; Chiyonobu, T.; Ishii, A. Application of Induced Pluripotent Stem Cells in Epilepsy. Mol. Cell. Neurosci. 2020, 108, 103535. [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [CrossRef]
- Renner, M.; Lancaster, M.A.; Bian, S.; Choi, H.; Ku, T.; Peer, A.; Chung, K.; Knoblich, J.A. Self-organized Developmental Patterning and Differentiation in Cerebral Organoids. EMBO J. 2017, 36, 1316–1329. [CrossRef]
- 2022 Alzheimer’s Disease Facts and Figures. Alzheimers Dement. 2022, 18, 700–789. [CrossRef]
- Gatz, M.; Reynolds, C.A.; Fratiglioni, L.; Johansson, B.; Mortimer, J.A.; Berg, S.; Fiske, A.; Pedersen, N.L. Role of Genes and Environments for Explaining Alzheimer Disease. Arch. Gen. Psychiatry 2006, 63, 168. [CrossRef]
- Lee, S.J. van der; Wolters, F.J.; Ikram, M.K.; Hofman, A.; Ikram, M.A.; Amin, N.; Duijn, C.M. van The Effect of APOE and Other Common Genetic Variants on the Onset of Alzheimer’s Disease and Dementia: A Community-Based Cohort Study. Lancet Neurol. 2018, 17, 434–444. [CrossRef]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; Van Der Flier, W.M. Alzheimer’s Disease. The Lancet 2021, 397, 1577–1590. [CrossRef]
- Ranjan, V.D.; Qiu, L.; Tan, E.K.; Zeng, L.; Zhang, Y. Modelling Alzheimer’s Disease: Insights from in Vivo to in Vitro Three-Dimensional Culture Platforms. J. Tissue Eng. Regen. Med. 2018, 12, 1944–1958. [CrossRef]
- Bi, F.-C.; Yang, X.-H.; Cheng, X.-Y.; Deng, W.-B.; Guo, X.-L.; Yang, H.; Wang, Y.; Li, J.; Yao, Y. Optimization of Cerebral Organoids: A More Qualified Model for Alzheimer’s Disease Research. Transl. Neurodegener. 2021, 10, 27. [CrossRef]
- Pavoni, S.; Jarray, R.; Nassor, F.; Guyot, A.-C.; Cottin, S.; Rontard, J.; Mikol, J.; Mabondzo, A.; Deslys, J.-P.; Yates, F. Small-Molecule Induction of Aβ-42 Peptide Production in Human Cerebral Organoids to Model Alzheimer’s Disease Associated Phenotypes. PLoS ONE 2018, 13, e0209150. [CrossRef]
- Raja, W.K.; Mungenast, A.E.; Lin, Y.-T.; Ko, T.; Abdurrob, F.; Seo, J.; Tsai, L.-H. Self-Organizing 3D Human Neural Tissue Derived from Induced Pluripotent Stem Cells Recapitulate Alzheimer’s Disease Phenotypes. PLoS ONE 2016, 11, e0161969. [CrossRef]
- Yan, Y.; Song, L.; Bejoy, J.; Zhao, J.; Kanekiyo, T.; Bu, G.; Zhou, Y.; Li, Y. Modeling Neurodegenerative Microenvironment Using Cortical Organoids Derived from Human Stem Cells. Tissue Eng. Part A 2018, 24, 1125–1137. [CrossRef]
- Lin, Y.-T.; Seo, J.; Gao, F.; Feldman, H.M.; Wen, H.-L.; Penney, J.; Cam, H.P.; Gjoneska, E.; Raja, W.K.; Cheng, J.; et al. APOE4 Causes Widespread Molecular and Cellular Alterations Associated with Alzheimer’s Disease Phenotypes in Human iPSC-Derived Brain Cell Types. Neuron 2018, 98, 1141-1154.e7. [CrossRef]
- Fisher, R.S.; Boas, W. van E.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel Jr., J. Epileptic Seizures and Epilepsy: Definitions Proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 2005, 46, 470–472. [CrossRef]
- Falco-Walter, J. Epilepsy—Definition, Classification, Pathophysiology, and Epidemiology. Semin. Neurol. 2020, 40, 617–623. [CrossRef]
- Organization, W.H. Epilepsy: a public health imperative: summary. 2019.
- MacDonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; Groot, N.; et al. A Novel Gene Containing a Trinucleotide Repeat That Is Expanded and Unstable on Huntington’s Disease Chromosomes. Cell 1993, 72, 971–983. [CrossRef]
- Arrasate, M.; Finkbeiner, S. Protein Aggregates in Huntington’s Disease. Exp. Neurol. 2012, 238, 1–11. [CrossRef]
- Andhale, R.; Shrivastava, D. Huntington’s Disease: A Clinical Review. Cureus 2022, 14, e28484. [CrossRef]
- Tabrizi, S.J.; Flower, M.D.; Ross, C.A.; Wild, E.J. Huntington Disease: New Insights into Molecular Pathogenesis and Therapeutic Opportunities. Nat. Rev. Neurol. 2020, 16, 529–546. [CrossRef]
- Bates, G.P.; Dorsey, R.; Gusella, J.F.; Hayden, M.R.; Kay, C.; Leavitt, B.R.; Nance, M.; Ross, C.A.; Scahill, R.I.; Wetzel, R.; et al. Huntington Disease. Nat. Rev. Dis. Primer 2015, 1, 15005. [CrossRef]
- Ross, C.A.; Aylward, E.H.; Wild, E.J.; Langbehn, D.R.; Long, J.D.; Warner, J.H.; Scahill, R.I.; Leavitt, B.R.; Stout, J.C.; Paulsen, J.S.; et al. Huntington Disease: Natural History, Biomarkers and Prospects for Therapeutics. Nat. Rev. Neurol. 2014, 10, 204–216. [CrossRef]
- Pouladi, M.A.; Morton, A.J.; Hayden, M.R. Choosing an Animal Model for the Study of Huntington’s Disease. Nat. Rev. Neurosci. 2013, 14, 708–721. [CrossRef]
- Ramaswamy, S.; McBride, J.L.; Kordower, J.H. Animal Models of Huntington’s Disease. ILAR J. 2007, 48, 356–373. [CrossRef]
- Park, I.-H.; Lerou, P.H.; Zhao, R.; Huo, H.; Daley, G.Q. Generation of Human-Induced Pluripotent Stem Cells. Nat. Protoc. 2008, 3, 1180–1186. [CrossRef]
- Park, I.-H.; Arora, N.; Huo, H.; Maherali, N.; Ahfeldt, T.; Shimamura, A.; Lensch, M.W.; Cowan, C.; Hochedlinger, K.; Daley, G.Q. Disease-Specific Induced Pluripotent Stem Cells. Cell 2008, 134, 877–886. [CrossRef]
- Zhang, N.; An, M.C.; Montoro, D.; Ellerby, L.M. Characterization of Human Huntington’s Disease Cell Model from Induced Pluripotent Stem Cells. PLoS Curr. 2010, 2, RRN1193. [CrossRef]
- Chae, J.-I.; Kim, D.-W.; Lee, N.; Jeon, Y.-J.; Jeon, I.; Kwon, J.; Kim, J.; Soh, Y.; Lee, D.-S.; Seo, K.S.; et al. Quantitative Proteomic Analysis of Induced Pluripotent Stem Cells Derived from a Human Huntington’s Disease Patient. Biochem. J. 2012, 446, 359–371. [CrossRef]
- HD iPSC Consortium Induced Pluripotent Stem Cells from Patients with Huntington’s Disease Show CAG-Repeat-Expansion-Associated Phenotypes. Cell Stem Cell 2012, 11, 264–278. [CrossRef]
- Mehta, S.R.; Tom, C.M.; Wang, Y.; Bresee, C.; Rushton, D.; Mathkar, P.P.; Tang, J.; Mattis, V.B. Human Huntington’s Disease iPSC-Derived Cortical Neurons Display Altered Transcriptomics, Morphology, and Maturation. Cell Rep. 2018, 25, 1081-1096.e6. [CrossRef]
- Moss, D.J.H.; Pardiñas, A.F.; Langbehn, D.; Lo, K.; Leavitt, B.R.; Roos, R.; Durr, A.; Mead, S.; TRACK-HD investigators; REGISTRY investigators; et al. Identification of Genetic Variants Associated with Huntington’s Disease Progression: A Genome-Wide Association Study. Lancet Neurol. 2017, 16, 701–711. [CrossRef]
- Telenius, H.; Kremer, B.; Goldberg, Y.P.; Theilmann, J.; Andrew, S.E.; Zeisler, J.; Adam, S.; Greenberg, C.; Ives, E.J.; Clarke, L.A. Somatic and Gonadal Mosaicism of the Huntington Disease Gene CAG Repeat in Brain and Sperm. Nat. Genet. 1994, 6, 409–414. [CrossRef]
- Świtońska, K.; Szlachcic, W.J.; Handschuh, L.; Wojciechowski, P.; Marczak, Ł.; Stelmaszczuk, M.; Figlerowicz, M.; Figiel, M. Identification of Altered Developmental Pathways in Human Juvenile HD iPSC With 71Q and 109Q Using Transcriptome Profiling. Front. Cell. Neurosci. 2018, 12, 528. [CrossRef]
- Mattis, V.B.; Tom, C.; Akimov, S.; Saeedian, J.; Østergaard, M.E.; Southwell, A.L.; Doty, C.N.; Ornelas, L.; Sahabian, A.; Lenaeus, L.; et al. HD iPSC-Derived Neural Progenitors Accumulate in Culture and Are Susceptible to BDNF Withdrawal Due to Glutamate Toxicity. Hum. Mol. Genet. 2015, 24, 3257–3271. [CrossRef]
- Smith-Geater, C.; Hernandez, S.J.; Lim, R.G.; Adam, M.; Wu, J.; Stocksdale, J.T.; Wassie, B.T.; Gold, M.P.; Wang, K.Q.; Miramontes, R.; et al. Aberrant Development Corrected in Adult-Onset Huntington’s Disease iPSC-Derived Neuronal Cultures via WNT Signaling Modulation. Stem Cell Rep. 2020, 14, 406–419. [CrossRef]
- An, M.C.; Zhang, N.; Scott, G.; Montoro, D.; Wittkop, T.; Mooney, S.; Melov, S.; Ellerby, L.M. Genetic Correction of Huntington’s Disease Phenotypes in Induced Pluripotent Stem Cells. Cell Stem Cell 2012, 11, 253–263. [CrossRef]
- Ring, K.L.; An, M.C.; Zhang, N.; O’Brien, R.N.; Ramos, E.M.; Gao, F.; Atwood, R.; Bailus, B.J.; Melov, S.; Mooney, S.D.; et al. Genomic Analysis Reveals Disruption of Striatal Neuronal Development and Therapeutic Targets in Human Huntington’s Disease Neural Stem Cells. Stem Cell Rep. 2015, 5, 1023–1038. [CrossRef]
- Xu, X.; Tay, Y.; Sim, B.; Yoon, S.-I.; Huang, Y.; Ooi, J.; Utami, K.H.; Ziaei, A.; Ng, B.; Radulescu, C.; et al. Reversal of Phenotypic Abnormalities by CRISPR/Cas9-Mediated Gene Correction in Huntington Disease Patient-Derived Induced Pluripotent Stem Cells. Stem Cell Rep. 2017, 8, 619–633. [CrossRef]
- HD iPSC Consortium Developmental Alterations in Huntington’s Disease Neural Cells and Pharmacological Rescue in Cells and Mice. Nat. Neurosci. 2017, 20, 648–660. [CrossRef]
- Bose, A.; Petsko, G.A.; Studer, L. Induced Pluripotent Stem Cells: A Tool for Modeling Parkinson’s Disease. Trends Neurosci. 2022, 45, 608–620. [CrossRef]
- Marotta, N.; Kim, S.; Krainc, D. Organoid and Pluripotent Stem Cells in Parkinson’s Disease Modeling: An Expert View on Their Value to Drug Discovery. Expert Opin. Drug Discov. 2020, 15, 427–441. [CrossRef]
- Burré, J.; Sharma, M.; Südhof, T.C. Cell Biology and Pathophysiology of α-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8, a024091. [CrossRef]
- Dettmer, U.; Newman, A.J.; Soldner, F.; Luth, E.S.; Kim, N.C.; Von Saucken, V.E.; Sanderson, J.B.; Jaenisch, R.; Bartels, T.; Selkoe, D. Parkinson-Causing α-Synuclein Missense Mutations Shift Native Tetramers to Monomers as a Mechanism for Disease Initiation. Nat. Commun. 2015, 6, 7314. [CrossRef]
- Jeong, G.R.; Lee, B.D. Pathological Functions of LRRK2 in Parkinson’s Disease. Cells 2020, 9, 2565. [CrossRef]
- Seibler, P.; Graziotto, J.; Jeong, H.; Simunovic, F.; Klein, C.; Krainc, D. Mitochondrial Parkin Recruitment Is Impaired in Neurons Derived from Mutant PINK1 Induced Pluripotent Stem Cells. J. Neurosci. 2011, 31, 5970–5976. [CrossRef]
- Burbulla, L.F.; Song, P.; Mazzulli, J.R.; Zampese, E.; Wong, Y.C.; Jeon, S.; Santos, D.P.; Blanz, J.; Obermaier, C.D.; Strojny, C.; et al. Dopamine Oxidation Mediates Mitochondrial and Lysosomal Dysfunction in Parkinson’s Disease. Science 2017, 357, 1255–1261. [CrossRef]
- Chen, X.; Sun, G.; Tian, E.; Zhang, M.; Davtyan, H.; Beach, T.G.; Reiman, E.M.; Blurton-Jones, M.; Holtzman, D.M.; Shi, Y. Modeling Sporadic Alzheimer’s Disease in Human Brain Organoids under Serum Exposure. Adv. Sci. Weinh. Baden-Wurtt. Ger. 2021, 8, e2101462. [CrossRef]
- Ghatak, S.; Dolatabadi, N.; Trudler, D.; Zhang, X.; Wu, Y.; Mohata, M.; Ambasudhan, R.; Talantova, M.; Lipton, S.A. Mechanisms of Hyperexcitability in Alzheimer’s Disease hiPSC-Derived Neurons and Cerebral Organoids vs Isogenic Controls. eLife 2019, 8, e50333. [CrossRef]
- Gonzalez, C.; Armijo, E.; Bravo-Alegria, J.; Becerra-Calixto, A.; Mays, C.E.; Soto, C. Modeling Amyloid Beta and Tau Pathology in Human Cerebral Organoids. Mol. Psychiatry 2018, 23, 2363–2374. [CrossRef]
- Arber, C.; Lovejoy, C.; Harris, L.; Willumsen, N.; Alatza, A.; Casey, J.M.; Lines, G.; Kerins, C.; Mueller, A.K.; Zetterberg, H.; et al. Familial Alzheimer’s Disease Mutations in PSEN1 Lead to Premature Human Stem Cell Neurogenesis. Cell Rep. 2021, 34, 108615. [CrossRef]
- Abud, E.M.; Ramirez, R.N.; Martinez, E.S.; Healy, L.M.; Nguyen, C.H.H.; Newman, S.A.; Yeromin, A.V.; Scarfone, V.M.; Marsh, S.E.; Fimbres, C.; et al. iPSC-Derived Human Microglia-like Cells to Study Neurological Diseases. Neuron 2017, 94, 278-293.e9. [CrossRef]
- Zhao, J.; Fu, Y.; Yamazaki, Y.; Ren, Y.; Davis, M.D.; Liu, C.-C.; Lu, W.; Wang, X.; Chen, K.; Cherukuri, Y.; et al. APOE4 Exacerbates Synapse Loss and Neurodegeneration in Alzheimer’s Disease Patient iPSC-Derived Cerebral Organoids. Nat. Commun. 2020, 11, 5540. [CrossRef]
- Park, J.-C.; Jang, S.-Y.; Lee, D.; Lee, J.; Kang, U.; Chang, H.; Kim, H.J.; Han, S.-H.; Seo, J.; Choi, M.; et al. A Logical Network-Based Drug-Screening Platform for Alzheimer’s Disease Representing Pathological Features of Human Brain Organoids. Nat. Commun. 2021, 12, 280. [CrossRef]
- Pérez, M.J.; Ivanyuk, D.; Panagiotakopoulou, V.; Di Napoli, G.; Kalb, S.; Brunetti, D.; Al-Shaana, R.; Kaeser, S.A.; Fraschka, S.A.-K.; Jucker, M.; et al. Loss of Function of the Mitochondrial Peptidase PITRM1 Induces Proteotoxic Stress and Alzheimer’s Disease-like Pathology in Human Cerebral Organoids. Mol. Psychiatry 2021, 26, 5733–5750. [CrossRef]
- Zhou, L.-T.; Liu, D.; Kang, H.-C.; Lu, L.; Huang, H.-Z.; Ai, W.-Q.; Zhou, Y.; Deng, M.-F.; Li, H.; Liu, Z.-Q.; et al. Tau Pathology Epigenetically Remodels the Neuron-Glial Cross-Talk in Alzheimer’s Disease. Sci. Adv. 2023, 9, eabq7105. [CrossRef]
- Lin, Y.-T.; Seo, J.; Gao, F.; Feldman, H.M.; Wen, H.-L.; Penney, J.; Cam, H.P.; Gjoneska, E.; Raja, W.K.; Cheng, J.; et al. APOE4 Causes Widespread Molecular and Cellular Alterations Associated with Alzheimer’s Disease Phenotypes in Human iPSC-Derived Brain Cell Types. Neuron 2018, 98, 1141-1154.e7. [CrossRef]
- Jin, M.; Xu, R.; Wang, L.; Alam, M.M.; Ma, Z.; Zhu, S.; Martini, A.C.; Jadali, A.; Bernabucci, M.; Xie, P.; et al. Type-I-Interferon Signaling Drives Microglial Dysfunction and Senescence in Human iPSC Models of Down Syndrome and Alzheimer’s Disease. Cell Stem Cell 2022, 29, 1135-1153.e8. [CrossRef]
- Choi, H.; Kim, H.J.; Yang, J.; Chae, S.; Lee, W.; Chung, S.; Kim, J.; Choi, H.; Song, H.; Lee, C.K.; et al. Acetylation Changes Tau Interactome to Degrade Tau in Alzheimer’s Disease Animal and Organoid Models. Aging Cell 2020, 19, e13081. [CrossRef]
- Kim, H.; Park, H.J.; Choi, H.; Chang, Y.; Park, H.; Shin, J.; Kim, J.; Lengner, C.J.; Lee, Y.K.; Kim, J. Modeling G2019S-LRRK2 Sporadic Parkinson’s Disease in 3D Midbrain Organoids. Stem Cell Rep. 2019, 12, 518–531. [CrossRef]
- Becerra-Calixto, A.; Mukherjee, A.; Ramirez, S.; Sepulveda, S.; Sinha, T.; Al-Lahham, R.; De Gregorio, N.; Gherardelli, C.; Soto, C. Lewy Body-like Pathology and Loss of Dopaminergic Neurons in Midbrain Organoids Derived from Familial Parkinson’s Disease Patient. Cells 2023, 12, 625. [CrossRef]
- Zheng, X.; Han, D.; Liu, W.; Wang, X.; Pan, N.; Wang, Y.; Chen, Z. Human iPSC-Derived Midbrain Organoids Functionally Integrate into Striatum Circuits and Restore Motor Function in a Mouse Model of Parkinson’s Disease. Theranostics 2023, 13, 2673–2692. [CrossRef]
- Raja, W.K.; Neves, E.; Burke, C.; Jiang, X.; Xu, P.; Rhodes, K.J.; Khurana, V.; Scannevin, R.H.; Chung, C.Y. Patient-Derived Three-Dimensional Cortical Neurospheres to Model Parkinson’s Disease. PloS One 2022, 17, e0277532. [CrossRef]
- Wulansari, N.; Darsono, W.H.W.; Woo, H.-J.; Chang, M.-Y.; Kim, J.; Bae, E.-J.; Sun, W.; Lee, J.-H.; Cho, I.-J.; Shin, H.; et al. Neurodevelopmental Defects and Neurodegenerative Phenotypes in Human Brain Organoids Carrying Parkinson’s Disease-Linked DNAJC6 Mutations. Sci. Adv. 2021, 7, eabb1540. [CrossRef]
- Sabate-Soler, S.; Nickels, S.L.; Saraiva, C.; Berger, E.; Dubonyte, U.; Barmpa, K.; Lan, Y.J.; Kouno, T.; Jarazo, J.; Robertson, G.; et al. Microglia Integration into Human Midbrain Organoids Leads to Increased Neuronal Maturation and Functionality. Glia 2022, 70, 1267–1288. [CrossRef]
- Chlebanowska, P.; Tejchman, A.; Sułkowski, M.; Skrzypek, K.; Majka, M. Use of 3D Organoids as a Model to Study Idiopathic Form of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 694. [CrossRef]
- Walter, J.; Bolognin, S.; Poovathingal, S.K.; Magni, S.; Gérard, D.; Antony, P.M.A.; Nickels, S.L.; Salamanca, L.; Berger, E.; Smits, L.M.; et al. The Parkinson’s-Disease-Associated Mutation LRRK2-G2019S Alters Dopaminergic Differentiation Dynamics via NR2F1. Cell Rep. 2021, 37, 109864. [CrossRef]
- Samarasinghe, R.A.; Miranda, O.A.; Buth, J.E.; Mitchell, S.; Ferando, I.; Watanabe, M.; Allison, T.F.; Kurdian, A.; Fotion, N.N.; Gandal, M.J.; et al. Identification of Neural Oscillations and Epileptiform Changes in Human Brain Organoids. Nat. Neurosci. 2021, 24, 1488–1500. [CrossRef]
- Steinberg, D.J.; Repudi, S.; Saleem, A.; Kustanovich, I.; Viukov, S.; Abudiab, B.; Banne, E.; Mahajnah, M.; Hanna, J.H.; Stern, S.; et al. Modeling Genetic Epileptic Encephalopathies Using Brain Organoids. EMBO Mol. Med. 2021, 13, e13610. [CrossRef]
- Eichmüller, O.L.; Corsini, N.S.; Vértesy, Á.; Morassut, I.; Scholl, T.; Gruber, V.-E.; Peer, A.M.; Chu, J.; Novatchkova, M.; Hainfellner, J.A.; et al. Amplification of Human Interneuron Progenitors Promotes Brain Tumors and Neurological Defects. Science 2022, 375, eabf5546. [CrossRef]
- Lewitzky, M.; Yamanaka, S. Reprogramming Somatic Cells towards Pluripotency by Defined Factors. Curr. Opin. Biotechnol. 2007, 18, 467–473. [CrossRef]
- Yagi, T.; Ito, D.; Okada, Y.; Akamatsu, W.; Nihei, Y.; Yoshizaki, T.; Yamanaka, S.; Okano, H.; Suzuki, N. Modeling Familial Alzheimer’s Disease with Induced Pluripotent Stem Cells. Hum. Mol. Genet. 2011, 20, 4530–4539. [CrossRef]
- Yamanaka, S. A Fresh Look at iPS Cells. Cell 2009, 137, 13–17. [CrossRef]
- Okano, H.; Yamanaka, S. iPS Cell Technologies: Significance and Applications to CNS Regeneration and Disease. Mol. Brain 2014, 7, 22. [CrossRef]
- Nori, S.; Okada, Y.; Yasuda, A.; Tsuji, O.; Takahashi, Y.; Kobayashi, Y.; Fujiyoshi, K.; Koike, M.; Uchiyama, Y.; Ikeda, E.; et al. Grafted Human-Induced Pluripotent Stem-Cell-Derived Neurospheres Promote Motor Functional Recovery after Spinal Cord Injury in Mice. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 16825–16830. [CrossRef]
- Yamanaka, S. Pluripotent Stem Cell-Based Cell Therapy-Promise and Challenges. Cell Stem Cell 2020, 27, 523–531. [CrossRef]
- Kwokdinata, C.; Ramanujam, V.; Chen, J.; de Oliveira, P.N.; Nai, M.H.; Chooi, W.H.; Lim, C.T.; Ng, S.Y.; David, L.; Chew, S.Y. Encapsulation of Human Spinal Cord Progenitor Cells in Hyaluronan-Gelatin Hydrogel for Spinal Cord Injury Treatment. ACS Appl. Mater. Interfaces 2023. [CrossRef]
- Sieber-Blum, M. Epidermal Neural Crest Stem Cells and Their Use in Mouse Models of Spinal Cord Injury. Brain Res. Bull. 2010, 83, 189–193. [CrossRef]
- Obara, K.; Shirai, K.; Hamada, Y.; Arakawa, N.; Yamane, M.; Takaoka, N.; Aki, R.; Hoffman, R.M.; Amoh, Y. Chronic Spinal Cord Injury Functionally Repaired by Direct Implantation of Encapsulated Hair-Follicle-Associated Pluripotent (HAP) Stem Cells in a Mouse Model: Potential for Clinical Regenerative Medicine. PloS One 2022, 17, e0262755. [CrossRef]
- Polentes, J.; Jendelova, P.; Cailleret, M.; Braun, H.; Romanyuk, N.; Tropel, P.; Brenot, M.; Itier, V.; Seminatore, C.; Baldauf, K.; et al. Human Induced Pluripotent Stem Cells Improve Stroke Outcome and Reduce Secondary Degeneration in the Recipient Brain. Cell Transplant. 2012, 21, 2587–2602. [CrossRef]
- Kawai, H.; Yamashita, T.; Ohta, Y.; Deguchi, K.; Nagotani, S.; Zhang, X.; Ikeda, Y.; Matsuura, T.; Abe, K. Tridermal Tumorigenesis of Induced Pluripotent Stem Cells Transplanted in Ischemic Brain. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2010, 30, 1487–1493. [CrossRef]
- Upadhya, D.; Hattiangady, B.; Castro, O.W.; Shuai, B.; Kodali, M.; Attaluri, S.; Bates, A.; Dong, Y.; Zhang, S.-C.; Prockop, D.J.; et al. Human Induced Pluripotent Stem Cell-Derived MGE Cell Grafting after Status Epilepticus Attenuates Chronic Epilepsy and Comorbidities via Synaptic Integration. Proc. Natl. Acad. Sci. 2019, 116, 287–296. [CrossRef]
- Cunningham, M.; Cho, J.-H.; Leung, A.; Savvidis, G.; Ahn, S.; Moon, M.; Lee, P.K.J.; Han, J.J.; Azimi, N.; Kim, K.-S.; et al. hPSC-Derived Maturing GABAergic Interneurons Ameliorate Seizures and Abnormal Behavior in Epileptic Mice. Cell Stem Cell 2014, 15, 559–573. [CrossRef]
- Martin, P.; Wagh, V.; Reis, S.A.; Erdin, S.; Beauchamp, R.L.; Shaikh, G.; Talkowski, M.; Thiele, E.; Sheridan, S.D.; Haggarty, S.J.; et al. TSC Patient-Derived Isogenic Neural Progenitor Cells Reveal Altered Early Neurodevelopmental Phenotypes and Rapamycin-Induced MNK-eIF4E Signaling. Mol. Autism 2020, 11, 2. [CrossRef]
- Cai, J.; Yang, M.; Poremsky, E.; Kidd, S.; Schneider, J.S.; Iacovitti, L. Dopaminergic Neurons Derived from Human Induced Pluripotent Stem Cells Survive and Integrate into 6-OHDA-Lesioned Rats. Stem Cells Dev. 2010, 19, 1017–1023. [CrossRef]
- Meneghini, V.; Frati, G.; Sala, D.; De Cicco, S.; Luciani, M.; Cavazzin, C.; Paulis, M.; Mentzen, W.; Morena, F.; Giannelli, S.; et al. Generation of Human Induced Pluripotent Stem Cell-Derived Bona Fide Neural Stem Cells for Ex Vivo Gene Therapy of Metachromatic Leukodystrophy. Stem Cells Transl. Med. 2017, 6, 352–368. [CrossRef]
- Yoon, Y.; Kim, H.S.; Hong, C.P.; Li, E.; Jeon, I.; Park, H.J.; Lee, N.; Pei, Z.; Song, J. Neural Transplants From Human Induced Pluripotent Stem Cells Rescue the Pathology and Behavioral Defects in a Rodent Model of Huntington’s Disease. Front. Neurosci. 2020, 14, 558204. [CrossRef]
- Chang, C.-Y.; Ting, H.-C.; Liu, C.-A.; Su, H.-L.; Chiou, T.-W.; Lin, S.-Z.; Harn, H.-J.; Ho, T.-J. Induced Pluripotent Stem Cell (iPSC)-Based Neurodegenerative Disease Models for Phenotype Recapitulation and Drug Screening. Mol. Basel Switz. 2020, 25, 2000. [CrossRef]
- Aboul-Soud, M.A.M.; Alzahrani, A.J.; Mahmoud, A. Induced Pluripotent Stem Cells (iPSCs)-Roles in Regenerative Therapies, Disease Modelling and Drug Screening. Cells 2021, 10, 2319. [CrossRef]
- Kondo, T.; Asai, M.; Tsukita, K.; Kutoku, Y.; Ohsawa, Y.; Sunada, Y.; Imamura, K.; Egawa, N.; Yahata, N.; Okita, K.; et al. Modeling Alzheimer’s Disease with iPSCs Reveals Stress Phenotypes Associated with Intracellular Aβ and Differential Drug Responsiveness. Cell Stem Cell 2013, 12, 487–496. [CrossRef]
- Miura, K.; Okada, Y.; Aoi, T.; Okada, A.; Takahashi, K.; Okita, K.; Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Ohnuki, M.; et al. Variation in the Safety of Induced Pluripotent Stem Cell Lines. Nat. Biotechnol. 2009, 27, 743–745. [CrossRef]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of Induced Pluripotent Stem Cells without Myc from Mouse and Human Fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [CrossRef]
- Okita, K.; Nakagawa, M.; Hyenjong, H.; Ichisaka, T.; Yamanaka, S. Generation of Mouse Induced Pluripotent Stem Cells without Viral Vectors. Science 2008, 322, 949–953. [CrossRef]
- Sugai, K.; Sumida, M.; Shofuda, T.; Yamaguchi, R.; Tamura, T.; Kohzuki, T.; Abe, T.; Shibata, R.; Kamata, Y.; Ito, S.; et al. First-in-Human Clinical Trial of Transplantation of iPSC-Derived NS/PCs in Subacute Complete Spinal Cord Injury: Study Protocol. Regen. Ther. 2021, 18, 321–333. [CrossRef]
- Hoveizi, E.; Mohammadi, T.; Moazedi, A.A.; Zamani, N.; Eskandary, A. Transplanted Neural-like Cells Improve Memory and Alzheimer-like Pathology in a Rat Model. Cytotherapy 2018, 20, 964–973. [CrossRef]
- Armijo, E.; Edwards, G.; Flores, A.; Vera, J.; Shahnawaz, M.; Moda, F.; Gonzalez, C.; Sanhueza, M.; Soto, C. Induced Pluripotent Stem Cell-Derived Neural Precursors Improve Memory, Synaptic and Pathological Abnormalities in a Mouse Model of Alzheimer’s Disease. Cells 2021, 10, 1802. [CrossRef]
- Yahata, N.; Asai, M.; Kitaoka, S.; Takahashi, K.; Asaka, I.; Hioki, H.; Kaneko, T.; Maruyama, K.; Saido, T.C.; Nakahata, T.; et al. Anti-Aβ Drug Screening Platform Using Human iPS Cell-Derived Neurons for the Treatment of Alzheimer’s Disease. PloS One 2011, 6, e25788. [CrossRef]
- Brownjohn, P.W.; Smith, J.; Portelius, E.; Serneels, L.; Kvartsberg, H.; De Strooper, B.; Blennow, K.; Zetterberg, H.; Livesey, F.J. Phenotypic Screening Identifies Modulators of Amyloid Precursor Protein Processing in Human Stem Cell Models of Alzheimer’s Disease. Stem Cell Rep. 2017, 8, 870–882. [CrossRef]
- Al-Gharaibeh, A.; Culver, R.; Stewart, A.N.; Srinageshwar, B.; Spelde, K.; Frollo, L.; Kolli, N.; Story, D.; Paladugu, L.; Anwar, S.; et al. Induced Pluripotent Stem Cell-Derived Neural Stem Cell Transplantations Reduced Behavioral Deficits and Ameliorated Neuropathological Changes in YAC128 Mouse Model of Huntington’s Disease. Front. Neurosci. 2017, 11, 628. [CrossRef]
- Cho, I.K.; Hunter, C.E.; Ye, S.; Pongos, A.L.; Chan, A.W.S. Combination of Stem Cell and Gene Therapy Ameliorates Symptoms in Huntington’s Disease Mice. NPJ Regen. Med. 2019, 4, 7. [CrossRef]
- Park, H.J.; Han, A.; Kim, J.Y.; Choi, J.; Bae, H.S.; Cho, G.-B.; Shin, H.; Shin, E.J.; Lee, K.-I.; Kim, S.; et al. SUPT4H1-Edited Stem Cell Therapy Rescues Neuronal Dysfunction in a Mouse Model for Huntington’s Disease. NPJ Regen. Med. 2022, 7, 8. [CrossRef]
- Doerr, J.; Böckenhoff, A.; Ewald, B.; Ladewig, J.; Eckhardt, M.; Gieselmann, V.; Matzner, U.; Brüstle, O.; Koch, P. Arylsulfatase A Overexpressing Human iPSC-Derived Neural Cells Reduce CNS Sulfatide Storage in a Mouse Model of Metachromatic Leukodystrophy. Mol. Ther. J. Am. Soc. Gene Ther. 2015, 23, 1519–1531. [CrossRef]
- Alekseenko, Z.; Dias, J.M.; Adler, A.F.; Kozhevnikova, M.; van Lunteren, J.A.; Nolbrant, S.; Jeggari, A.; Vasylovska, S.; Yoshitake, T.; Kehr, J.; et al. Robust Derivation of Transplantable Dopamine Neurons from Human Pluripotent Stem Cells by Timed Retinoic Acid Delivery. Nat. Commun. 2022, 13, 3046. [CrossRef]
- Brot, S.; Thamrin, N.P.; Bonnet, M.-L.; Francheteau, M.; Patrigeon, M.; Belnoue, L.; Gaillard, A. Long-Term Evaluation of Intranigral Transplantation of Human iPSC-Derived Dopamine Neurons in a Parkinson’s Disease Mouse Model. Cells 2022, 11, 1596. [CrossRef]
- Guo, Y.; Guan, Y.; Zhu, H.; Sun, T.; Wang, Y.; Huang, Y.; Ma, C.; Emery, R.; Guan, W.; Wang, C.; et al. Therapeutic Function of iPSCs-Derived Primitive Neuroepithelial Cells in a Rat Model of Parkinson’s Disease. Neurochem. Int. 2022, 155, 105324. [CrossRef]
- Hiller, B.M.; Marmion, D.J.; Thompson, C.A.; Elliott, N.A.; Federoff, H.; Brundin, P.; Mattis, V.B.; McMahon, C.W.; Kordower, J.H. Optimizing Maturity and Dose of iPSC-Derived Dopamine Progenitor Cell Therapy for Parkinson’s Disease. NPJ Regen. Med. 2022, 7, 24. [CrossRef]
- Grinand, L.; Takahashi, J. Automated Measurement of Fluorescence Signals Reveals a Significant Increase of the Graft-Derived Neurite Extension in Neonates Compared to Aged Rats. Regen. Ther. 2022, 19, 97–106. [CrossRef]
- Nakamura, R.; Nonaka, R.; Oyama, G.; Jo, T.; Kamo, H.; Nuermaimaiti, M.; Akamatsu, W.; Ishikawa, K.-I.; Hattori, N. A Defined Method for Differentiating Human iPSCs into Midbrain Dopaminergic Progenitors That Safely Restore Motor Deficits in Parkinson’s Disease. Front. Neurosci. 2023, 17, 1202027. [CrossRef]
- Guo, Y.; Zhu, H.; Wang, Y.; Sun, T.; Xu, J.; Wang, T.; Guan, W.; Wang, C.; Liu, C.; Ma, C. Miniature-Swine iPSC-Derived GABA Progenitor Cells Function in a Rat Parkinson’s Disease Model. Cell Tissue Res. 2023, 391, 425–440. [CrossRef]
- Zheng, X.; Han, D.; Liu, W.; Wang, X.; Pan, N.; Wang, Y.; Chen, Z. Human iPSC-Derived Midbrain Organoids Functionally Integrate into Striatum Circuits and Restore Motor Function in a Mouse Model of Parkinson’s Disease. Theranostics 2023, 13, 2673–2692. [CrossRef]
- Lavoie, N.S.; Truong, V.; Malone, D.; Pengo, T.; Patil, N.; Dutton, J.R.; Parr, A.M. Human Induced Pluripotent Stem Cells Integrate, Create Synapses and Extend Long Axons after Spinal Cord Injury. J. Cell. Mol. Med. 2022, 26, 1932–1942. [CrossRef]
- Kitagawa, T.; Nagoshi, N.; Kamata, Y.; Kawai, M.; Ago, K.; Kajikawa, K.; Shibata, R.; Sato, Y.; Imaizumi, K.; Shindo, T.; et al. Modulation by DREADD Reveals the Therapeutic Effect of Human iPSC-Derived Neuronal Activity on Functional Recovery after Spinal Cord Injury. Stem Cell Rep. 2022, 17, 127–142. [CrossRef]
- Shibata, T.; Tashiro, S.; Shibata, S.; Shinozaki, M.; Shindo, T.; Hashimoto, S.; Kawai, M.; Kitagawa, T.; Ago, K.; Matsumoto, M.; et al. Rehabilitative Training Enhances Therapeutic Effect of Human iPSC-Derived Neural Stem/Progenitor Cells Transplantation in Chronic Spinal Cord Injury. Stem Cells Transl. Med. 2023, 12, 83–96. [CrossRef]
- Zheng, Y.; Gallegos, C.M.; Xue, H.; Li, S.; Kim, D.H.; Zhou, H.; Xia, X.; Liu, Y.; Cao, Q. Transplantation of Human Induced Pluripotent Stem Cell-Derived Neural Progenitor Cells Promotes Forelimb Functional Recovery after Cervical Spinal Cord Injury. Cells 2022, 11, 2765. [CrossRef]
- Deng, P.; Wang, L.; Zhang, Q.; Chen, S.; Zhang, Y.; Xu, H.; Chen, H.; Xu, Y.; He, W.; Zhang, J.; et al. Therapeutic Potential of a Combination of Electroacupuncture and Human iPSC-Derived Small Extracellular Vesicles for Ischemic Stroke. Cells 2022, 11, 820. [CrossRef]
- Kaiser, E.E.; Waters, E.S.; Yang, X.; Fagan, M.M.; Scheulin, K.M.; Sneed, S.E.; Cheek, S.R.; Jeon, J.H.; Shin, S.K.; Kinder, H.A.; et al. Tanshinone IIA-Loaded Nanoparticle and Neural Stem Cell Therapy Enhances Recovery in a Pig Ischemic Stroke Model. Stem Cells Transl. Med. 2022, 11, 1061–1071. [CrossRef]
- Chen, Y.; Song, F.; Tu, M.; Wu, S.; He, X.; Liu, H.; Xu, C.; Zhang, K.; Zhu, Y.; Zhou, R.; et al. Quantitative Proteomics Revealed Extensive Microenvironmental Changes after Stem Cell Transplantation in Ischemic Stroke. Front. Med. 2022, 16, 429–441. [CrossRef]
- Arakawa, M.; Sakamoto, Y.; Miyagawa, Y.; Nito, C.; Takahashi, S.; Nitahara-Kasahara, Y.; Suda, S.; Yamazaki, Y.; Sakai, M.; Kimura, K.; et al. iPSC-Derived Mesenchymal Stem Cells Attenuate Cerebral Ischemia-Reperfusion Injury by Inhibiting Inflammatory Signaling and Oxidative Stress. Mol. Ther. Methods Clin. Dev. 2023, 30, 333–349. [CrossRef]
- Cao, S.-Y.; Tao, M.-D.; Lou, S.-N.; Yang, D.; Lin, Y.-H.; Wu, H.-Y.; Chang, L.; Luo, C.-X.; Xu, Y.; Liu, Y.; et al. Functional Reconstruction of the Impaired Cortex and Motor Function by hMGEOs Transplantation in Stroke. Biochem. Biophys. Res. Commun. 2023, 671, 87–95. [CrossRef]
- Martinez-Curiel, R.; Jansson, L.; Tsupykov, O.; Avaliani, N.; Aretio-Medina, C.; Hidalgo, I.; Monni, E.; Bengzon, J.; Skibo, G.; Lindvall, O.; et al. Oligodendrocytes in Human Induced Pluripotent Stem Cell-Derived Cortical Grafts Remyelinate Adult Rat and Human Cortical Neurons. Stem Cell Rep. 2023, 18, 1643–1656. [CrossRef]
- Hulme, A.J.; Maksour, S.; St-Clair Glover, M.; Miellet, S.; Dottori, M. Making Neurons, Made Easy: The Use of Neurogenin-2 in Neuronal Differentiation. Stem Cell Rep. 2022, 17, 14–34. [CrossRef]
- Barak, M.; Fedorova, V.; Pospisilova, V.; Raska, J.; Vochyanova, S.; Sedmik, J.; Hribkova, H.; Klimova, H.; Vanova, T.; Bohaciakova, D. Human iPSC-Derived Neural Models for Studying Alzheimer’s Disease: From Neural Stem Cells to Cerebral Organoids. Stem Cell Rev. Rep. 2022, 18, 792–820. [CrossRef]
- Andrews, P., Cavagnaro, J., Deans, R. et al. Harmonizing standards for producing clinical-grade therapies from pluripotent stem cells. Nat Biotechnol, 2014, 32, 724–726.
- Okano H, Nakamura M, Yoshida K, Okada Y, Tsuji O, Nori S, Ikeda E, Yamanaka S, Miura K. Steps toward safe cell therapy using induced pluripotent stem cells. Circ Res. 2013, 112, 523-533.
Figure 1.
Primary cultures can be obtained from different tissues such as skin, blood, neurons and urine. Cellular reprogramming to generate iPSC can be carried out using different combinations of transcription factors, with Yamanaka factors (OSKM) still being the most used. Likewise, there are different tools to promote the transfection of cells to be reprogrammed, such as MMLV, SENDAI, Liposome and Transposons.
Figure 1.
Primary cultures can be obtained from different tissues such as skin, blood, neurons and urine. Cellular reprogramming to generate iPSC can be carried out using different combinations of transcription factors, with Yamanaka factors (OSKM) still being the most used. Likewise, there are different tools to promote the transfection of cells to be reprogrammed, such as MMLV, SENDAI, Liposome and Transposons.
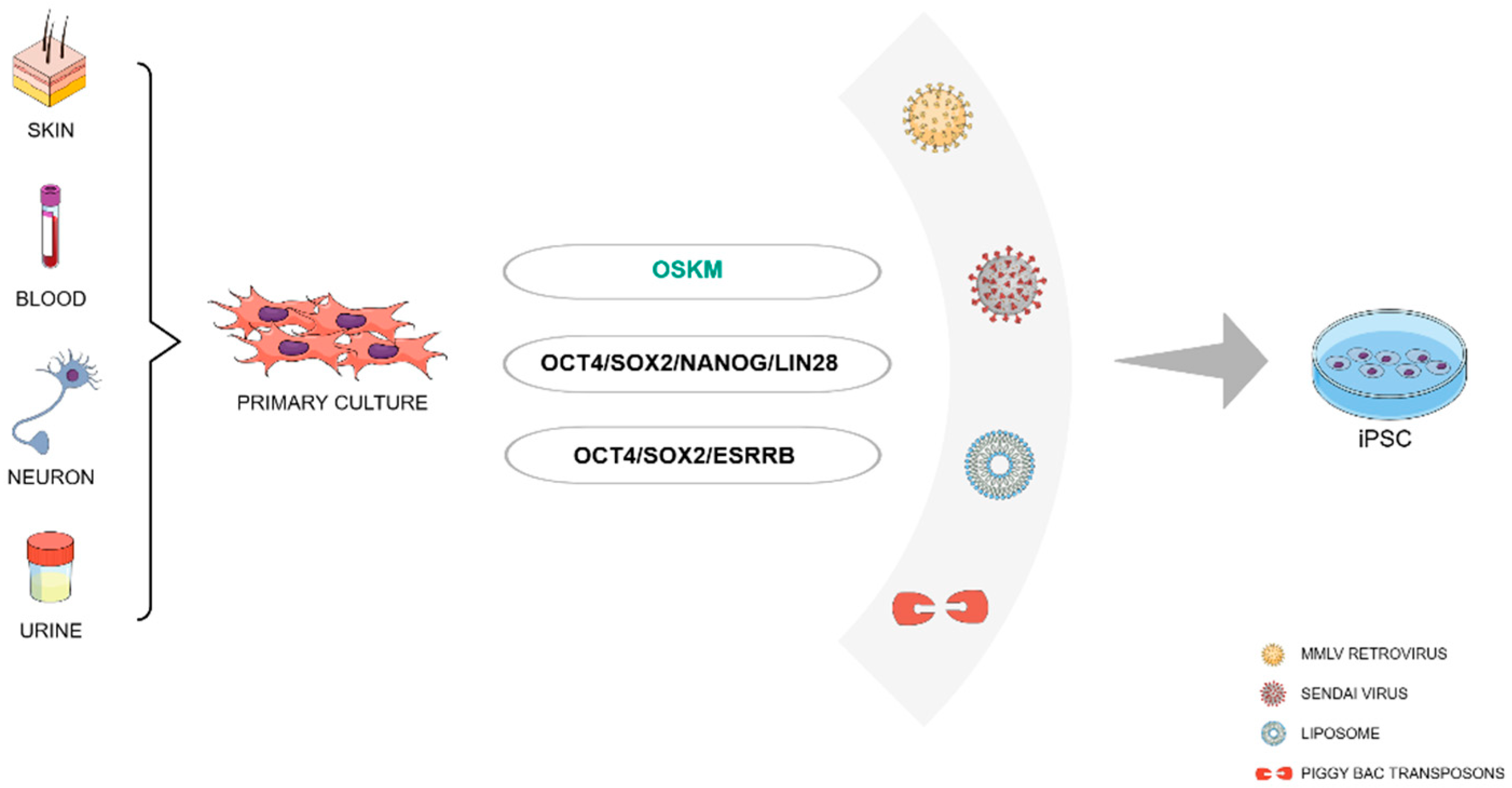
Figure 2.
Organoids can be generated from iPSC cultures using three-dimensional cultivation tools. Organoids can serve as models for studies of various diseases, including neurological diseases such as Alzheimer Disease, Epilepsy, Huntington Disease and Parkinson Disease.
Figure 2.
Organoids can be generated from iPSC cultures using three-dimensional cultivation tools. Organoids can serve as models for studies of various diseases, including neurological diseases such as Alzheimer Disease, Epilepsy, Huntington Disease and Parkinson Disease.
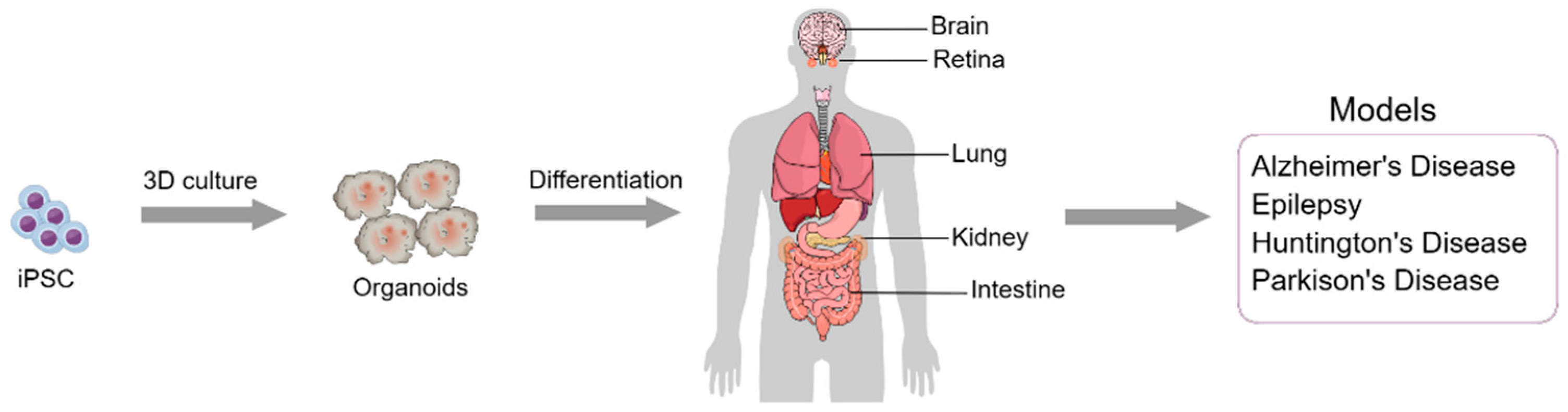

Table 1.
Organoid models generated for studying several neuropathological diseases.
| Organoid Type | Disease | Cell Type | Result | Reference |
|---|---|---|---|---|
| Cerebral Organoid | AD | iPSC | Modeling Sporadic Alzheimer’s Disease in Human Brain Organoids under Serum Exposure | [116] |
| Cerebral Organoid | AD | hiPSC | Mechanisms of hyperexcitability in Alzheimer’s disease hiPSC-derived neurons and cerebral organoids vs isogenic controls | [117] |
| Cerebral Organoid | AD | iPSC | Modeling amyloid beta and tau pathology in human cerebral organoids | [118] |
| Disease Stem Cell | AD | iPSC | Familial Alzheimer’s Disease Mutations in PSEN1 Lead to Premature Human Stem Cell Neurogenesis | [119] |
| Disease Stem Cell | AD | iPSC and hiPSC | iPSC-derived human microglia-like cells to study neurological diseases | [120] |
| Cerebral Organoid | AD | iPSC | APOE4 exacerbates synapse loss and neurodegeneration in Alzheimer’s disease patient iPSC-derived cerebral organoids | [121] |
| Cerebral Organoid | AD | iPSC | A logical network-based drug-screening platform for Alzheimer’s disease representing pathological features of human brain organoids | [122] |
| Cerebral Organoid | AD | iPSC | Loss of function of the mitochondrial peptidase PITRM1 induces proteotoxic stress and Alzheimer’s disease-like pathology in human cerebral organoids | [123] |
| Cerebral Organoid | AD | iPSC | Tau pathology epigenetically remodels the neuron-glial cross-talk in Alzheimer’s disease | [124] |
| Disease Stem Cell | AD | iPSC | APOE4 causes widespread molecular and cellular alterations associated with Alzheimer’s disease phenotypes in human iPSC-derived brain cell types | [125] |
| Disease Stem Cell | AD | iPSC | Type I Interferon Signaling Drives Microglial Dysfunction and Senescence in Human iPSC Models of Down Syndrome and Alzheimer’s Disease | [126] |
| Cerebral Organoid | AD | iPSC | Acetylation changes tau interactome to degrade tau in Alzheimer’s disease animal and organoid models | [127] |
| Cerebral Organoid | PD | hiPSC | Modeling G2019S-LRRK2 Sporadic Parkinson’s Disease in 3D Midbrain Organoids | [128] |
| Cerebral Organoid | PD | hiPSC | Lewy Body-like Pathology and Loss of Dopaminergic Neurons in Midbrain Organoids Derived from Familial Parkinson’s Disease Patient | [129] |
| Midbrain Organoid | PD | hiPSC | Human iPSC-derived midbrain organoids functionally integrate into striatum circuits and restore motor function in a mouse model of Parkinson’s disease | [130] |
| Neurospheres | PD | hiPSC and iPSC | Patient-derived three-dimensional cortical neurospheres to model Parkinson’s disease | [131] |
| Midbrain Organoid | PD | hiPSC and iPSC | Neurodevelopmental defects and neurodegenerative phenotypes in human brain organoids carrying Parkinson’s disease linked DNAJC6 mutations | [132] |
| Midbrain Organoid | PD | iPSC | Microglia integration into human midbrain organoids leads to increased neuronal maturation and functionality | [133] |
| Cerebral Organoid | PD | iPSC | Use of 3D Organoids as a Model to Study Idiopathic Form of Parkinson’s Disease | [134] |
| Cerebral Organoid | PD | iPSC | The Parkinson’s-disease-associated mutation LRRK2-G2019S alters dopaminergic differentiation dynamics via NR2F1 | [135] |
| Cerebral Organoid | Rett syndrome | hiPSC | Identification of neural oscillations and epileptiform changes in human brain organoids | [136] |
| Cerebral Organoid | TLE | iPSC | Modeling genetic epileptic encephalopathies using brain organoids | [137] |
| Cerebral Organoid | TSC | hiPSC | Amplification of human interneuron progenitors promotes brain tumors and neurological defects | [138] |
Legend: AD - Alzheimer's Disease; PD - Parkinson's Disease; TLE - Temporal Lobe Epilepsy; TSC – Tuberous Sclerosis Complex.
Table 2.
iPSCs usage in cell therapy and drug screening for several neuropathologies.
| Trial Type | Disease | Target | Result | Reference |
|---|---|---|---|---|
| Cell Therapy | AD | Rat | The transplanted rats rescued Alzheimer's cognition. | [163] |
| Cell Therapy | AD | Mouse | Grafted mice showed improved memory, synaptic plasticity, and reduced AD brain pathology, including a reduction of amyloid and tangles deposits. | [164] |
| Drug Screening | AD | hiPSC | β-secretase inhibitor IV (BSI) and γ-secretase inhibitor XXI/Compound E (GSI) showed similar effects as screening in other models. | [165] |
| Drug Screening | AD | hiPSC | Docosahexaenoic acid (DHA) treatment alleviated the stress responses in the AD neural cells. | [158] |
| Drug Screening | AD | hiPSC | The anthelminthic avermectins increases the relative production of short forms of Aβ and reduces the relative production of longer Aβ fragments in human cortical neurons. | [166] |
| Cell Therapy | HD | Mice | iPSCs survived and differentiated into region-specific neurons in both mice groups without tumor formation. | [167] |
| Cell Therapy | HD | Mice | Grafted mice showed a significant increase in lifespan. In iPSCs groups, animals showed significant improvement in motor functions and grip strength. | [168] |
| Cell Therapy | HD | Rat | Grafted rats showed significant behavioral improvements for up to 12 weeks. iPSCs enhanced endogenous neurogenesis and reconstituted the damaged neuronal connections. | [155] |
| Cell Therapy | HD | Mice | Improved neuronal dysfunction by SUPT4H1-edited iPSCs grafts. | [169] |
| Cell Therapy | MLD | Mice | Transplantation of ARSA-overexpressing precursors into ARSA-deficient mice significantly reduced sulfatide storage up to 300 µm from grafted cells. | [170] |
| Cell Therapy | MLD | Mice | Grafts of iPSCs into neonatal and adult immunodeficient MLD mice stably restored arylsulfatase A (ARSA) activity in the whole central nervous system and a significant decrease of sulfatide storage when ARSA-overexpressing cells were used. | [154] |
| Cell Therapy | PD | Rat | iPSC graft differentiated into mature mDA neurons that survive over long term and restored motor function. | [171] |
| Cell Therapy | PD | Mice | hiPSCs differentiated into mDA neurons and there was long-term motor functional recovery after transplantation. | [172] |
| Cell Therapy | PD | Rat | Grafted iPSCs could survive in Parkinsonian rat brain for at least 150 days, and many of them differentiated into tyrosine hydroxylase (TH)-positive cells. | [173] |
| Cell Therapy | PD | Rat | Intranigral engraftment to the ventral midbrain demonstrated that mDA progenitors cryopreserved on day 17, cells had a greater capacity than immature mDA neuron cells to innervate over long distances to forebrain structures. | [174] |
| Cell Therapy | PD | Rat | hiPSCs-derived dopaminergic progenitor cells integrate better into the striatum of neonates than older rats. | [175] |
| Cell Therapy | PD | Mice | More than 90% of the engrafted cells differentiated into the lineage of mDA neurons, and approximately 15% developed into mature mDA neurons without tumor formation. | [176] |
| Cell Therapy | PD | Rat | There was a neural remodel of basal ganglia circuitry and no tumorigenicity. | [177] |
| Cell Therapy | PD | Mice | iPSCs matured into mDA neurons and reverse motor function and establishment of bidirectional connections with natural brain target regions, without tumor formation. | [178] |
| Cell Therapy | SCI | Rat | Transplanted cells displayed robust integration properties including synapse formation and myelination by host. | [179] |
| Cell Therapy | SCI | Mice | Due to DREADD expression, it was shown a significant decrease in locomotor dysfunction in SCI-grafted mice, which was exclusively observed following the neurons maturation. | [180] |
| Cell Therapy | SCI | Mice | The combination of iPSCs graft and rehabilitative training therapy significantly improved motor functions. | [181] |
| Cell Therapy | SCI | Rat | Neuro pluripotent cells derived from iPSC were able to survive and differentiate into both neurons and astrocytes and, improved forelimb locomotor function. | [182] |
| Cell Therapy | Stroke | Mice | Combination of electroacupuncture and iPSC-derived extracellular vesicles treatment ameliorated neurological impairments and reduced the infarct volume and neuronal apoptosis in MCAO mice. | [183] |
| Cell Therapy | Stroke | Pig | Tanshinone IIA nanoparticles increased iPSCs engraftment, enhanced cellular and tissue recovery, and improved neurological function in a translational pig stroke model. | [184] |
| Cell Therapy | Stroke | Rat | Increased glucose metabolism and neurofunctional in iPSCs-transplanted rats. | [185] |
| Cell Therapy | Stroke | Rat | Graft of iPSCs inhibited microglial activation and expression of proinflammatory cytokines and suppressed oxidative stress and neuronal death in the cerebral cortex at the ischemic border zone. | [186] |
| Cell Therapy | Stroke | Mice | Graft survived well and primarily differentiated into GABAergic interneurons and significantly restored the sensorimotor deficits of stroke mice for a long time. | [187] |
| Cell Therapy | Stroke | Rat | Generated oligodendrocytes survived and formed myelin-ensheathing human axons in the host tissue after grafting onto adult human cortical organotypic cultures. | [188] |
| Cell Therapy | TLE | Mice | A much-reduced frequency of spontaneous recurrent seizures in grafted animals. | [150] |
Legend: AD - Alzheimer's Disease; HD - Huntington's disease; MLD - Metachromatic leukodystrophy; PD - Parkinson's Disease; SCI - Spinal Cord Injury; TLE - Temporal Lobe Epilepsy; mDA – Midbrain dopaminergic; SCI and PD papers presented are only from 2023 and 2022 due to the large number of publications.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



