
Submitted:
05 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
CRISPR/Cas9-based gene therapy has gotten a lot of interest during last decade. This treatment protocol tries to fix disease-causing traits by changing the DNA code in the exact spot on the chromosome. Therefore, we have tried to elaborate the most current information that shows how well CRISPR/Cas9-based gene therapy works. In vivo methods, like the ones described in the next two lines, are used to study cells in live creatures, like babies that are growing or animals that are already grown up. It has also been suggested to use ex-vivo methods to change somatic stem cells or progenitor cells in a culture outside of the body before putting them back into the patient. CRISPR/Cas is a good way to change specific genes in the DNA. Clustered Regularly Interspaced Short Palindromic Repeats is the full name for CRISPR. Biotechnologists can use them to fix DNA instead of the body's natural ways of doing so. This helps physicians in treatment diseases that run in families more successfully. Cas9 is an enzyme that, along with a guide RNA, is part of the CRISPR/Cas system. As this new technology grows quickly, it is replacing normal medical processes with treatments that are more cutting-edge and can change people's lives. CRISPR-Cas technology has changed biology by making it possible to change genes quickly. CRISPR molecular tools (Cas9 or Cas12a) have a lot of potential, but they are not very useful right now because they depend on the target cell's own DNA repair systems. With or without a template, the body's natural DNA repair processes can fix DNA breaks caused by Cas9 and Cas12a-based technologies. People use these methods a lot, but their effectiveness ranges from cell type to cell type. HDR-mediated DNA repair is a part of cell division, so tools that target it don't work on cells that don't divide, like neurons. CRISPR-associated transposase (CAST) has recently been studied, which suggests that it may offer new ways to change genes with CRISPR. The CRISPR activator of type V-K that is part of CAST has the same structure as the transposase. This is because CRISPR systems can put DNA in the right place without the help of the cell's own DNA repair systems. But a lot of work is being done right now to improve Cas9 and Cas12a so that DNA can be put into a target gene more precisely. Experts are still working hard to come up with better and more reliable ways to change genes, so even though there may be a problem, both methods have useful uses.
Keywords:
CRISPR-Cas
; Endonucleases
; Biogenesis
; DNAse
Introduction
As a result of these studies, CRISPR/Cas9-based gene therapy has gotten a lot of interest. This treatment tries to fix disease-causing traits by changing the DNA code in the exact spot on the chromosome. In this study, we look at the most current data that shows how well CRISPR/Cas9-based gene therapy works. In vivo methods, like the ones described in the next two lines, are used to study cells in live creatures, like babies that are growing or animals that are already grown up. It has also been suggested to use methods called "ex-vivo" to change somatic stem cells or progenitor cells in a culture outside of the body before putting them back into the patient. CRISPR/Cas is a good way to change specific genes in the DNA. Clustered Regularly Interspaced Short Palindromic Repeats is the full name for CRISPR. Biotechnologists can use them to fix DNA instead of the body's natural ways of doing so. This helps doctors treat diseases that run in families more successfully. Cas9 is an enzyme that, along with a guide RNA, is part of the CRISPR/Cas system. As this new technology grows quickly, it is replacing normal medical processes with treatments that are more cutting-edge and can change people's lives.
History, Development and Scientific Advancement
The genome editing was a dream of health care researcher to cure the challenging disease. This is another potential method for genetic engineering because it makes it possible to add, change, or take out specific pieces of DNA from the genome of an organism. Because the testing is different, the traits of the genes change. In the end, this causes the change in how the tissues and organs work. Because of this, the healing uses of this technology have grown to include areas as different as biology and synthetic nanotechnology. The history of development and advancements of CRISPR Cas is also interesting. That highlights the chronological review and scientific achievements in this field as mentioned in Table 1.
CRISPR-Cas System’s Groups and Classes
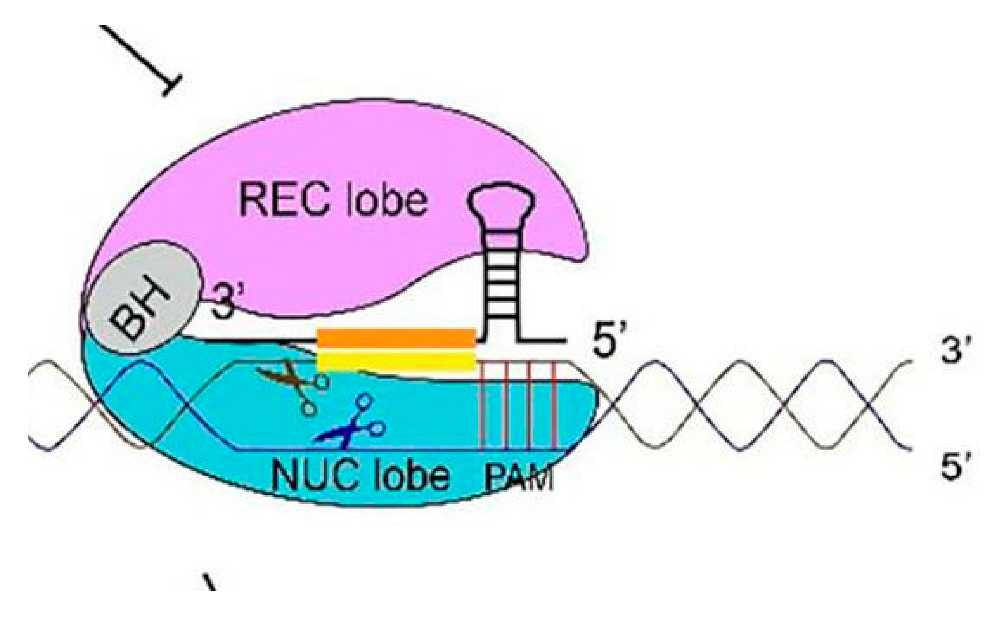
There are two major and different kinds of CRISPR-Cas systems that fits into six different groups. In class 1 (I, III, and IV) CRISPR ribonucleoprotein work as effector nucleases and there is more than one Cas protein. In the class 2 (II, V, and VI) CRISPR effector nucleases with one Cas protein. About 90% of all CRISPR-Cas genes are part of Class 1 CRISPR-Cas systems. Both bacteria and archaea have these kinds of systems. The only things that have class 2 CRISPR-Cas systems are bacteria. These make up about 10% of all CRISPR-Cas systems. They do their jobs by putting together combinations of crRNA and Cas protein, which are called ribonucleoproteins. The crRNA tells the cell where on the genome to go. When the crRNA and the target DNA have the same PAM (Protospacer Adjacent Motif) sequence, these multidomain effector proteins are turned on. These ribonucleoprotein complexes improve the guide sequence used to change DNA by using crRNA that has been changed. This makes the editing of genomes more accurate. The CRISPR-Cas system of Streptococcus pyogenes, especially the Type II subtype II-A system, has been studied a lot. In class 2, there are four different kinds of V. These are written from A to U in the order of the alphabet. Neither the V-C nor the V-U shapes, nor how they are made, are talked about in detail. The V-A gene sets the amount of Cas12a that is made. Recent studies of the structure of Cas12a have given us important information about how the protein works.
Characteristics of CRISPR-Cas Technology
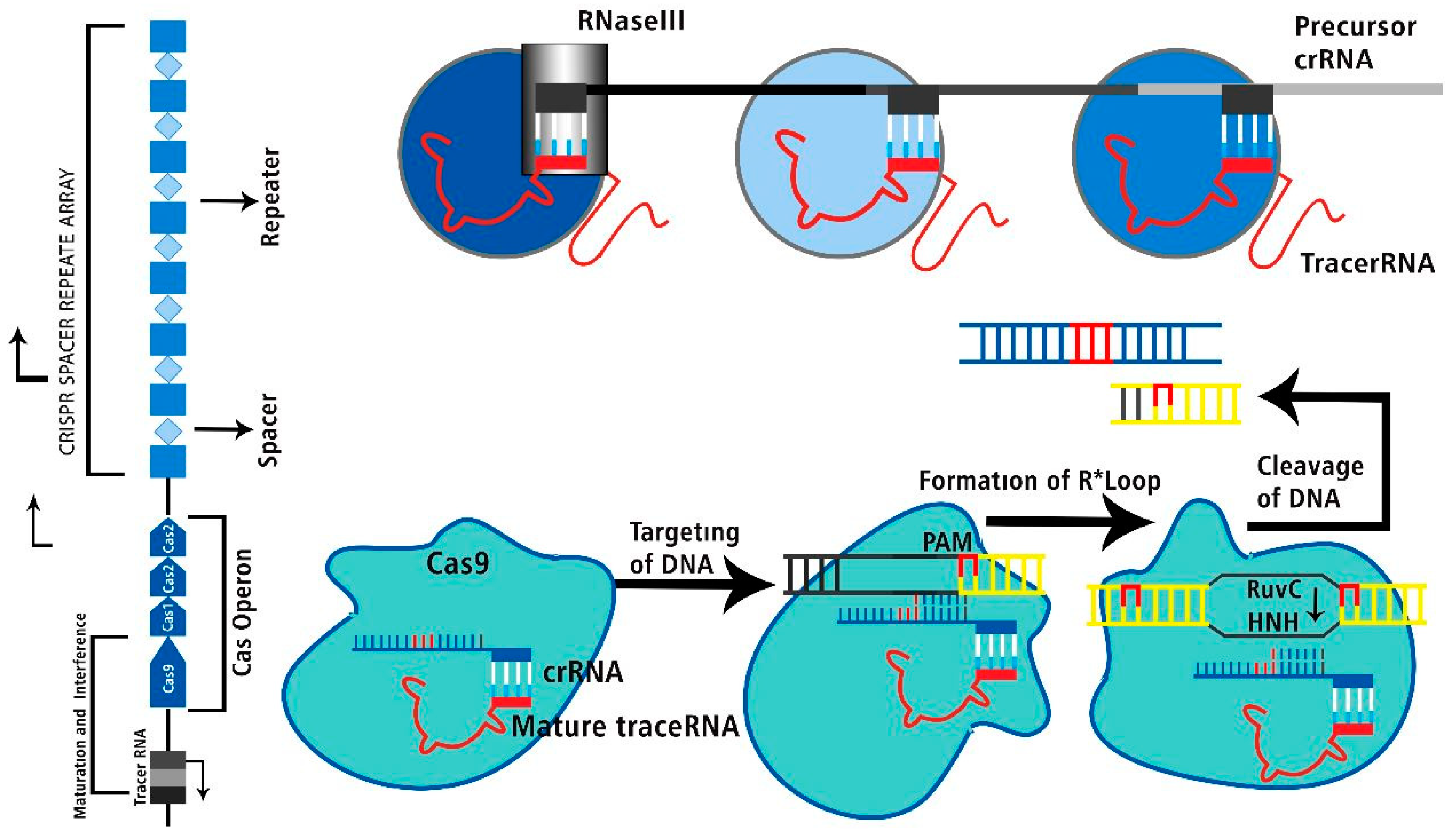
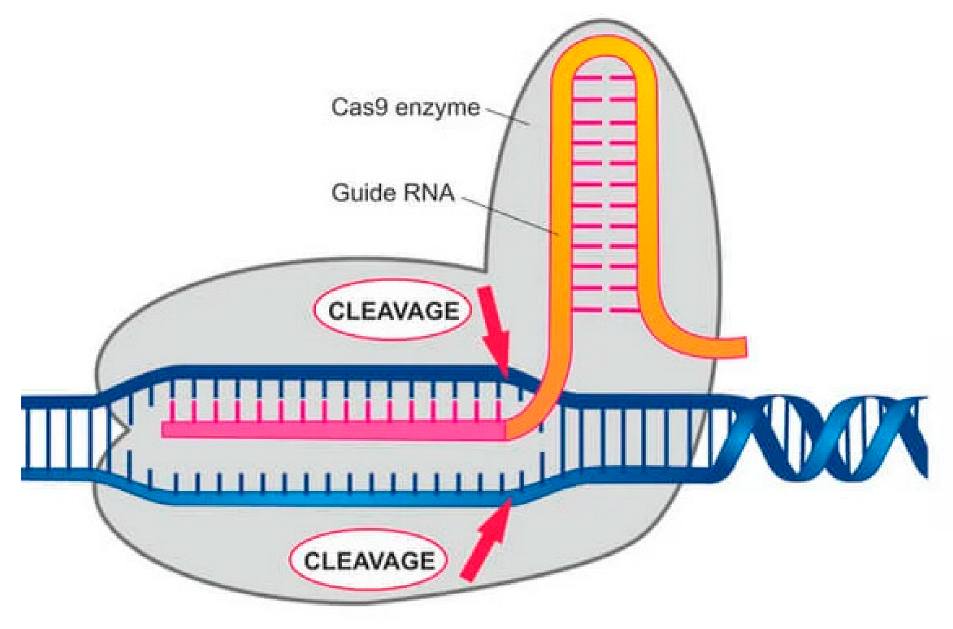
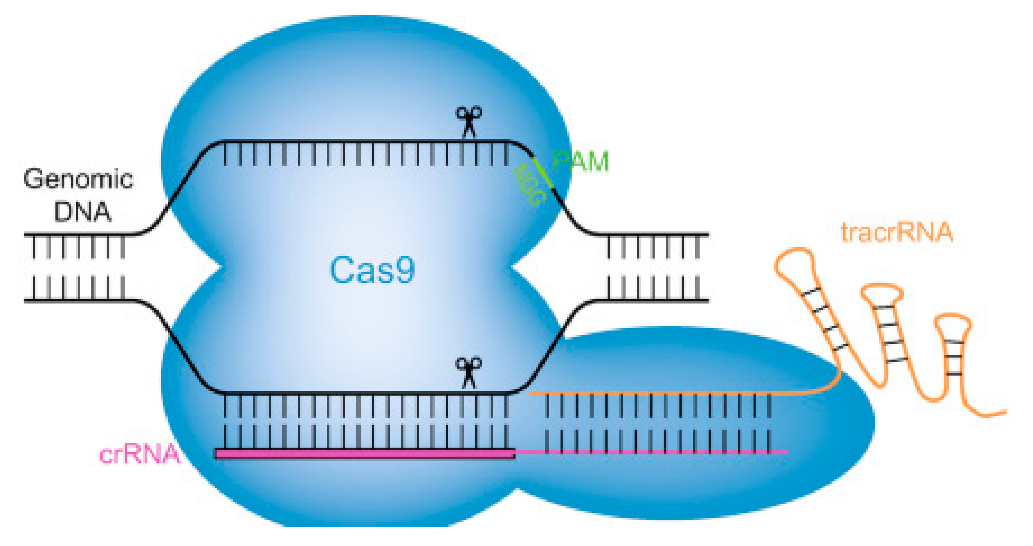
Clustered regularly interspaced short palindromic repeats, or CRISPRs, are a type of repeating DNA structure that helps bacteria and archaea fight off phages and plasmids when they work with the Cas genes. Most study has been done on Type II CRISPR/Cas systems, but there are two other types. The CRISPR gene is changed by adding pieces of alien DNA to it. This is the first step of the defense system being used. The locus DNA is turned into a single copy of noncoding precursor CRISPR RNA (Pre-crRNA), which is then processed into the final crRNA. When the crRNA joins with another non-coding RNA called trans-activating CRISPR RNA (tracrRNA), an endonuclease called Cas9 is made. This is a ribonucleoprotein complex. After Cas9 finds the alien DNA, it starts to cut it.
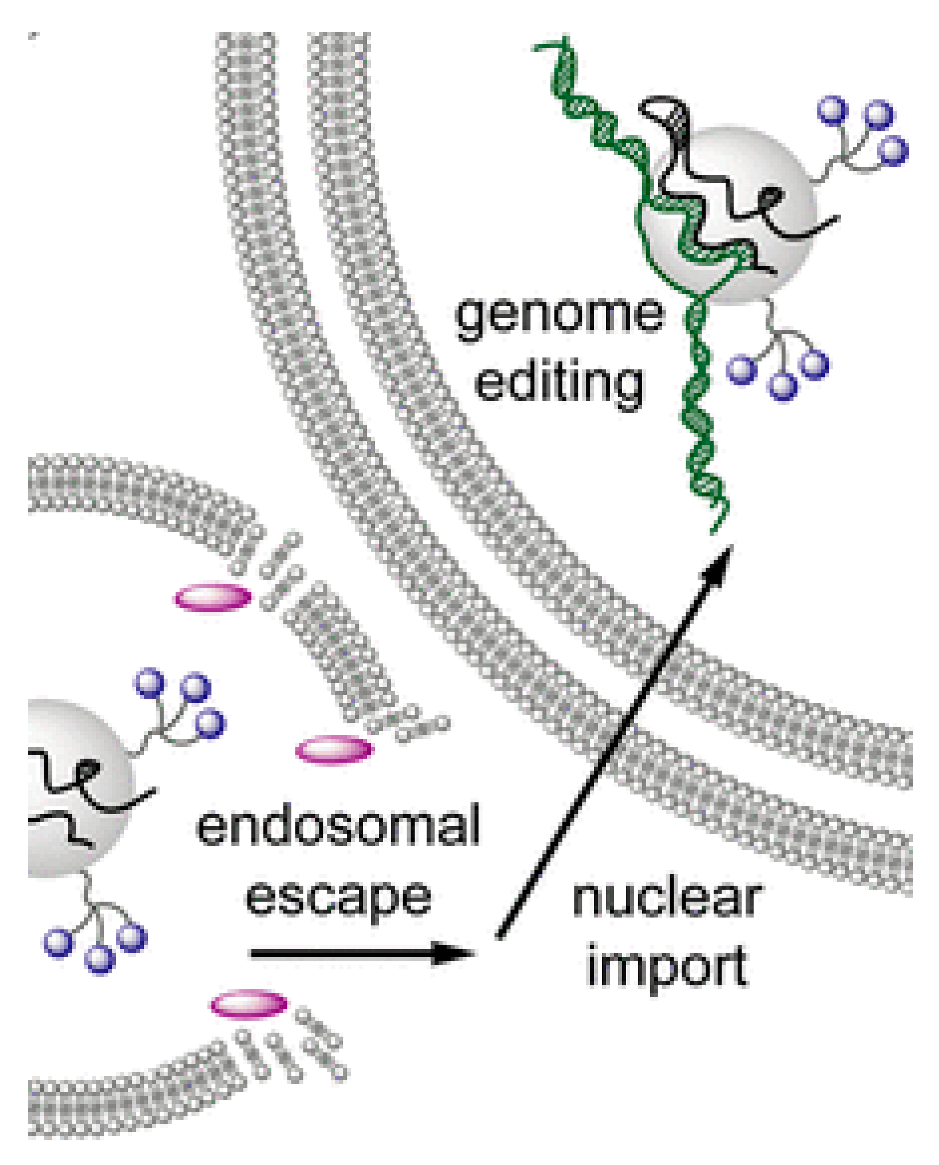
Figure 1.
The fundamental four steps of CRIPR Cas to precisely inset the desired DNA at targeted place.
Figure 1.
The fundamental four steps of CRIPR Cas to precisely inset the desired DNA at targeted place.
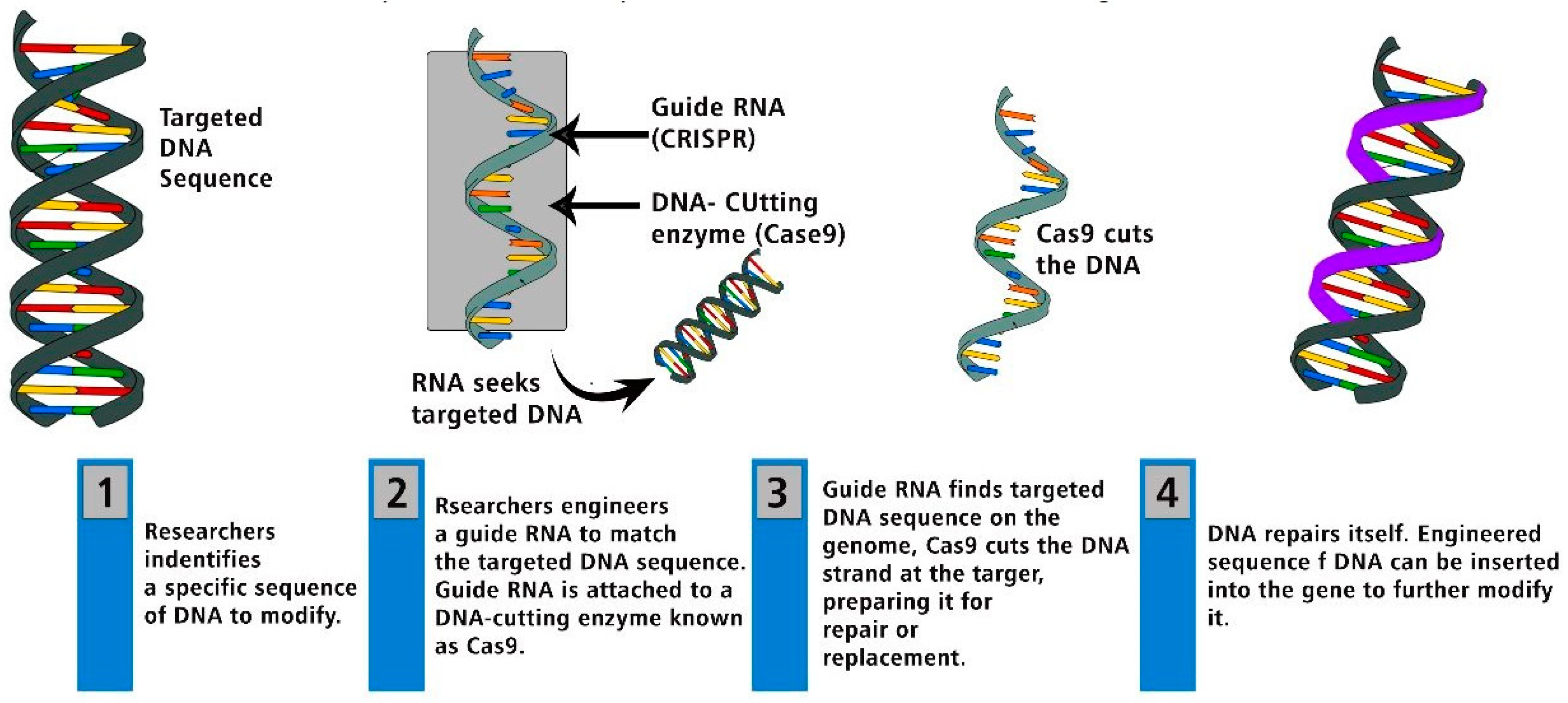
When the CRISPR/Cas9 complex makes Site-Specific Double-Strand Breaks in the DNA, there are two things that could happen. First of all, there isn't a single DNA code. A process called Nonhomologous End Joining (NHEJ) can add or remove small pieces of DNA to fix Double-Strand Breaks. Second, when a synthetic repair template is available, any changes to base pairs can be made during homology-directed repair (HDR) of double-strand breaks. Before RNA-guided designer nucleases like CRISPR/Cas9 were made, DNA was edited with programmable DNA-binding nucleases like Zinc Finger Nucleases (ZNFs) and transcription activator-like effector nucleases (TALENs). But sequence-specific DNA-binding proteins aren't used as much as they could be because it's hard and takes a long time to make them. CRISPR/Cas9 has become the go-to way to change DNA. Due to how easy it is to make CRISPR-guided nucleases, many studies have been done on different animal species, including mammals and monkeys, to change their genes.
Figure 2.
Classification of CRISPR Cas system and its application in tomato breeding.
Genome Modification, Cleavage Reliability and Specificity
Many of the nucleases used to change the genome depend on cleavage dependability very much. Because changes in other parts of the genome could have effects that were not expected, the best tool would only change the part that was wanted to be changed. So, uses that involve changing the genome put a high value on precision and the products that come from cleavage. Nuclease processing and deep sequencing was recently used to compare Cas12a and Cas9 from different species. This showed that both enzymes have similar types of specificity and can handle similar mistakes, even though in vivo studies have shown that Cas12a has less of an effect on things that aren't its target. Maybe the difference between notice and cutting is what makes this seem unreal, but these cutters might act differently in a chromatin environment. That arise query of whether in vitro or in vivo methods should be used to rethink nucleases methodology.
Additionally, the RNA-guided interference has become a powerful biology tool that can be used to change the genome. Changing the RNA guide sequence of a nuclease is all it takes to change its job. Cas9, a part of the Type II CRISPR-Cas interference system, is the most common tool used to change DNA.
Biomedical Applications
All of these tools are kept up-to-date so that scientists can continue to use them. Researchers have been looking into how CRISPR-Cas nucleases could be used to make smart materials like hydrogels. These water-filled molecules are wrapped in DNA. In a recent study, Cas12a was used to break down the DNA structure of DNA hydrogels in a specific way. This lets the smart tool be turned into a device whose contents are released at a certain time and place from DNA-encased hydrogels. Cas12a could be used because the way it breaks up makes it easier to direct the path of the bomb. It's hard to think about all of the possible uses because there are so many, but experts can now make knowledgeable guesses about how certain methods and mixtures of them might be used.
Conclusions
CRISPR-Cas technology has surprisingly changed the biology by making it possible to change the genes promptly. CRISPR molecular tools have a lot of potential, but they are not very useful right now because they depend on the particular target cells or DNA systems. The researchers have to move with or without a template; the body's natural DNA repair processes can fix DNA breaks caused by Cas9 and Cas12a-based technologies. However, scientists use these methods frequently but their effectiveness ranges from cell type to cell type. HDR-mediated DNA repair is a part of cell division, so tools that target it don't work on cells that don't divide, like neurons. CRISPR-associated transposase has recently been studied, which suggests that it may offer new ways to change genes with CRISPR. The CRISPR activator is part of CRISPR-associated transposase has the same structure as the transposase. This is because CRISPR systems can put DNA in the right place without the help of the cell's own DNA repair systems. Thus, a lot of work is being completed to improve technology. So the DNA can be added into a target place within the gene more precisely. We are still working hard to come up with better and more reliable ways to change genes.
Author Contributions
This work was carried out in collaboration among all authors. Taha Nazir designed the study of proposed hypothesis and compile the scientific contents. Nida Taha elaborated study to make it more credible. Whereas, Hameed A Mirza managed the literature searches and citation part of the manuscript. Thus, all authors have read and approved the final manuscript for publication in this journal.
Funding
This project is not-funded from any local and/ or international organization.
Ethical Approval and Consent to participate
All procedures performed in studies are not involving human participants. Therefore there is no need of the ethical approval of the institutional and/or national research committee and 1964 Helsinki declaration and its later amendments or comparable ethical standards. For type of studies no formal consent is required.
Animal rights
Additionally, this research studies no animals involved. The authors indicate the procedures followed are in accordance with the standards set forth in the eighth edition of Guide for the Care and Use of Laboratory Animals; published by the National Academy of Sciences, The National Academies Press, Washington, D.C.).
Informed Consent Statement
Authors agree and grant consent to publish this article in this research journal.
Data Availability Statement
All study information and possible research data successfully incorporated for publication.
Acknowledgements
We acknowledge the technical and scientific support of A.S. Chemical Laboratories Inc., Concord, ON L4K4M4 Canada and Advanced Multiple Inc., Mississauga ON, L5T2M9 Canada
Conflicts of Interest
The authors also declare that they are no any potential and/ or completing conflict of interest.
References
- Abudayyeh, O.O.; et al. (2017) RNA targeting with CRISPR–Cas13. Nature 550, 280–28. [CrossRef]
- Ali, Z.; et al. (2016) CRISPR/Cas9-mediated immunity to geminiviruses: differential interference and evasion. Sci. Rep. 6, 26912. [CrossRef]
- Andersson, M.; et al. (2017) Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas. [CrossRef]
- Bayat, H.; et al. (2018) The conspicuity of CRISPR[1]Cpf1 system as a significant breakthrough in genome editing. Curr. Microbiol. 75, 107–115. [CrossRef]
- Braatz, J.; et al. (2017) CRISPR-Cas9 targeted mutagenesis leads to simultaneous modification of different homoeologous gene copies in polyploid oilseed rape (Brassica napus). Plant Physiol. 174, 935–942. [CrossRef]
- Char, S.N.; et al. (2017) An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol. J. 15, 257–268. [CrossRef]
- Che, P.; et al. (2018) Developing a flexible, high[1]efficiency Agrobacterium-mediated sorghum transformation system with broad application. Plant Biotechnol. J. 16, 1388–1395. [CrossRef]
- Chen, X.; et al. (2017) Targeted mutagenesis in cotton (Gossypium hirsutum L.) using the CRISPR/ Cas9 system. Sci. Rep. 7, 44304. [CrossRef]
- Cho, S.; et al. (2018) Applications of CRISPR/Cas system to bacterial metabolic engineering. Int. J. Mol. Sci. 19, 1089. [CrossRef]
- Dreissig, S.; et al. (2017) Live-cell CRISPR imaging in plants reveals dynamic telomere movements. Plant J. 91, 565–573. [CrossRef]
- Enciso-Rodriguez, F.; et al. (2019) Overcoming self[1]incompatibility in diploid potato using CRISPR[1]Cas9. Front. Plant Sci. 10, 376. [CrossRef]
- Endo, M.; et al. (2019) Genome editing in plants by engineered CRISPR–Cas9 recognizing NG PAM. Nat. Plants 5, 14–17. [CrossRef]
- Gallego-Bartolome´, J.; et al. (2018) Targeted DNA demethylation of the Arabidopsis genome using the human TET1 catalytic domain. Proc. Natl. Acad. Sci. U. S. A. 115, E2125–E2134. [CrossRef]
- Gao, L.; et al. (2017) Engineered Cpf1 variants with altered PAM specificities. Nat. Biotechnol. 35, 789–792. [CrossRef]
- Gaudelli, N.M.; et al. (2017) Programmable base editing of A, T to G, C in genomic DNA without DNA cleavage. Nature 551, 464. [CrossRef]
- Gehrke, J.M.; et al. (2018) An APOBEC3A-Cas9 base editor with minimized bystander and off-target activities. Nat. Biotechnol. 36, 977–982. [CrossRef]
- Harrington, L.B.; et al. (2018) Programmed DNA destruction by miniature CRISPR-Cas14 enzymes. Science 362, 839–842. [CrossRef]
- (2013) DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 31, 827–832. [CrossRef]
- Hu, J.H.; et al. (2018) Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 556, 57–63. [CrossRef]
- Hua, K.; et al. (2019) Genome engineering in rice using Cas9 variants that recognize NG PAM sequences. Mol. Plant 12, 1003–1014. [CrossRef]
- Jansing, J.; et al. (2018) CRISPR/Cas9-mediated knockout of six glycosyltransferase genes in Nicotiana benthamiana for the production of recombinant proteins lacking b-1, 2-xylose and core a-1, 3-fucose. Plant Biotechnol. J. 17, 350–361. [CrossRef]
- Jeon, Y.; et al. (2018) Direct observation of DNA target searching and cleavage by CRISPR-Cas12a. Nat. Commun. 9, 2777. [CrossRef]
- Jia, H.; et al. (2016) Modification of the PthA4 effector binding elements in type I CsLOB1 promoter using Cas9/sg RNA to produce transgenic Duncan grapefruit alleviating XccDpthA4:dCsLOB1.3 infection. Plant Biotechnol. J. 14, 1291–1301. [CrossRef]
- Kanazashi, Y.; et al. (2018) Simultaneous site[1]directed mutagenesis of duplicated loci in soybean using a single guide RNA. Plant Cell Rep. 37, 553–563. [CrossRef]
- Kaur, N.; et al. (2018) CRISPR/Cas9- mediated efficient editing in phytoene desaturase (PDS) demonstrates precise manipulation in banana cv. Rasthali genome. Funct. Integr. Genom 18, 89–99. [CrossRef]
- Khan, M.Z.; et al. (2019) Targeting plant ssDNA viruses with engineered miniature CRISPR-Cas14a. Trends Biotech. 37, 800–804. [CrossRef]
- Khanday, I.; et al. (2019) A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. Nature 565, 91–95. [CrossRef]
- Kim, E.; et al. (2017) In vivo genome editing with a small Cas9 orthologue derived from. Campylobacter jejuni. Nat. Commun. 8, 14500. [CrossRef]
- Kim, Y.B.; et al. (2017) Increasing the genome[1]targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nat. Biotechnol. 35, 371–376. [CrossRef]
- Kleinstiver, B.P.; et al. (2015) Engineered CRISPR[1]Cas9 nucleases with altered PAM specificities. Nature 523, 481–485. [CrossRef]
- Kleinstiver, B.P.; et al. (2019) Engineered CRISPR–Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing. Nat. Biotechnol. 37, 276–282. [CrossRef]
- Kocak, D.D.; et al. (2019) Increasing the specificity of CRISPR systems with engineered RNA secondary structures. Nat. Biotechnol. 37, 657–666. [CrossRef]
- Komor, A.C.; et al. (2017) Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:G-to-T:A base editors with higher efficiency and product purity. Sci. Adv. 3, eaao4774. [CrossRef]
- Lee, C.M.; et al. (2016) The Neisseria meningitides CRISPR-Cas9 system enables specific genome editing in mammalian cells. Mol. Ther. 24, 645–654. [CrossRef]
- Lee, K.; et al. (2018) Activities and specificities of CRISPR/Cas9 and Cas12a nucleases for targeted mutagenesis in maize. Plant Biotechnol. J. 17, 362–372. [CrossRef]
- Li, A.; et al. (2018) Editing of an alpha-kafirin gene family increases, digestibility and protein quality in Sorghum. Plant Physiol. 177, 1425–1438. [CrossRef]
- Li, B.; et al. (2017) Targeted mutagenesis in the medicinal plant. Salvia miltiorrhiza. Sci. Rep. 7, 43320. [CrossRef]
- Pessina, S.; et al. (2016) Knockdown of MLO genes reduces susceptibility to powdery mildew in grapevine. Hort. Res. 3, 16016. [CrossRef]
- Li, C.; et al. (2017) A high-efficiency CRISPR/Cas9 system for targeted mutagenesis in cotton (Gossypium hirsutum L.). Sci. Rep. 7, 43902. [CrossRef]
- Macovei, A.; et al. (2018) Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to rice tungro spherical virus. Plant. [CrossRef]
- Makarova, K.S.; et al. (2015) An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 13, 722–736. [CrossRef]
- Malnoy, M.; et al. (2016) DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 7, 1904. [CrossRef]
- Mao, Y.; et al. (2017) Heritability of targeted gene modifications induced by plant-optimized CRISPR systems. Cell. Mol. Life Sci. 74, 1075–1093. [CrossRef]
- Mercx, S.; et al. (2017) Inactivation of the b (1, 2)- xylosyltransferase and the a (1, 3)-fucosyltransferase genes in Nicotiana tabacum BY-2 cells by a multiplex CRISPR/Cas9 strategy results in glycoproteins without plant-specific glycans. Front. Plant Sci. 8, 403. [CrossRef]
- Mir, A.; et al. (2018) Heavily and fully modified RNAs guide efficient SpyCas9-mediated genome editing. Nat. Commun. 9, 2641. [CrossRef]
- Moon, S.B.; et al. (2018) Highly efficient genome editing by CRISPR-Cpf1 using CRISPR RNA with a uridinylate-rich 3ʹ-overhang. Nat. Commun. 9, 3651. [CrossRef]
- Nishimasu, H.; et al. (2018) Engineered CRISPR-Cas9 nuclease with expanded targeting space. Science 361, 1259–1262. [CrossRef]
- Peng, C.; et al. (2018) High-throughput detection and screening of plants modified by gene editing using quantitative real-time PCR. Plant J. 95, 557–567. [CrossRef]
- Puchta, H.; et al. (2017) Applying CRISPR/Cas for genome engineering in plants: the best is yet to come. Curr. Opin. Plant Biol. 36, 1–8. [CrossRef]
- Qin, L.; et al. (2019) High efficient and precise base editing of C, G to T, A in the allotetraploid cotton (Gossypium hirsutum) genome using a modified CRISPR/Cas9 system. Plant Biotechnol. J. Published online May 22, 2019. [CrossRef]
- Rees, H.A.; Liu, D.R. (2018) Base editing: precision chemistry on the genome and transcriptome of living cells. Nat. Rev. Genet. 19, 770–788. [CrossRef]
- Shrestha, A.; et al. (2018) Cis-trans engineering advances and perspectives on customized transcriptional regulation in plants. Mol. Plant 11, 886–898. [CrossRef]
- Tang, X.; et al. (2017) A CRISPR–Cpf1 system for efficient genome editing and transcriptional repression in plants. Nat. Plants 3, 17018. [CrossRef]
- Wang, P.; et al. (2018) High efficient multisites genome editing in allotetraploid cotton (Gossypium hirsutum) using CRISPR/Cas9 system. Plant Biotechnol J. 16, 137–150. [CrossRef]
- Wang, X.; et al. (2018) CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 16, 844–855. [CrossRef]
- Wilson, L.O.; et al. (2018) The current state and future of CRISPR-Cas9 gRNA design tools. Front. Pharmacol. 9, 749. [CrossRef]
- Wolter, F.; Puchta, H. (2018) The CRISPR/Cas revolution reaches the RNA world: Cas13, a new Swiss Army knife for plant biologists. Plant J. 94, 767–775. [CrossRef]
- Yang, B.; et al. (2017) APOBEC: from mutator to editor. J. Genet. Genomics 44, 423–437 Doudna, J.A. and. [CrossRef]

Table 1.
CRISPR Cas system - Chronological review and scientific advancement.
| Advancement/ Discovery | Advancement/ Discovery | ||
| 2020 Nobel prize for CRISPR Cas9 genome editing |
 |
2019 First invivo CRISPR clinically trialed for the treatment monogenic disorders. The invention of nCATS by CRISPR/Cas9 |
 |
| 2018 1st CRISPR clinical trial for cancer immunotherapy |
 |
2016 The Base editor (BE) was made after Cas13a (C2c2) was found. |
 |
| 2015 Therapeutic proof of concept based on the discovery of Cas13a (C2c2), which is a part of the Multiplex CRISPR/Cas9 system, and its use to fix the dystrophin gene in vitro and then put it into a mouse model. The Base editor (BE) was made after Cas12a (Cpf1) was found. |
 |
2014 Cas9 has been turned into a solid, and libraries for screening the whole genome have been made. |
 |
| 2013 Using CRISPR Cas9, cells from mammals were successfully changed. When two Cas9n Nickases are used together, they work better for off-site targeting. Evidence for genome editing in eukaryotic cells using CRISPR/Cas9. Cas9-based functional screening of the whole genome. For the first time, it has been shown that eukaryotic cells can use Cas9 to change their DNA. |
 |
2014 Mammalian cells' genomes are being changed. Mammalian cells' genomes are being changed. The finds of dCas9, CRISPRi, and CRISPRa. Cas9 RNA makes site-specific editing of the genome possible in human cells and other living creatures. The finds of dCas9, CRISPRi, and CRISPRa. Cas9 RNA makes site-specific editing of the genome possible in human cells and other living creatures. |
 |
| 2012 In live species, Cas9 was shown to be an RNA-guided endonuclease, and its ability to target DNA was defined. Cas9-driven DNA targeting: a look in the lab. CRISPR Cas9 is a DNA endonuclease that is directed by RNA and has been studied in a lab dish.. |
 |
2011 There are three parts to the CRISPR system, and each one is made up of a different mix of tracrRNA, crRNA, and Cas9. The finds of tracRNA and Type II CRISPR Even though all cas genes have tracRNA, only the cas9 gene is needed for type II defence. |
 |
| 2010 Cas9 is led to the place where the DNA will be cut by protospacer sequences. Gameau (2010) found that Cas9 cuts target DNA and that Type II CRISPR Cas also cuts target DNA (67). |
 |
2009 Cmr complex cleaves ssRNA |
 |
| 2008 CRISPR system types III-A targets DNA, Discovered the function of crRNA |
 |
The first experimental proof that CRISPR gives adaptive immunity (Barrangou et al., 2007) suggests that CRISPR ioci may give their hosts adaptive immunity. |  |
| 2005 Due to the fact that the alien origin of the spaceship was found, the adaptive immune function was proposed. Cas genes found in CRISPRs include virus sequences. (2005-2006) |
 |
2002 Identification of Cas genes, Coined the CRISPR acronym, CRISPR name adopted and signature Cas genes identified. Coined CRISPR name defined signature Cas genes, Coined CRISPR name, discovery of Cas genes |
 |
| 2000 Discovered that CRISPR families are widespread in procaryotes |
 |
1993 Discovery of CRISPR clustered repeats in M. tuberculosis |
 |
| 1987 Escherichia coli was the first to find CRISPR grouped repeats. The CRISPR-associated grouped repeats were found. In a 1987 study by Ishino et al., CRISPR clustered repeats (CRISPR) were first described. This study was also the first to describe CRISPR clustered repeats. |
 |
1979 Gene Replacement in yeast. |
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



