Submitted:
05 January 2024
Posted:
08 January 2024
You are already at the latest version
Abstract
: Protease-producing yeasts were isolated from potato wastes and screened for protease production on skim milk agar plates The best producer of protease isolate was identified as Clavispora lusitaniae. The strain showed better enzyme production using tomato pomace and bread waste mix as a solid fermentation substrate. The optimized conditions improved enzyme activity and showed a maximal production of 33450±503 IU/g compared with the initial activity of 11205,78 ±360 without medium optimization. A threefold increase in protease activity after medium optimization proved the reliability of using the PBD and CCD design. A 19.76-fold purified enzyme and a yield of 32.94% were obtained after purification. The protease showed maximum activity at pH 4 and 60 °C and was resistant to Tween 20, Tween 40, SDS, and β-mercaptoethanol, Ca2+, and Mg2+ stimulated it. The protease activity was strongly inhibited in the presence of urea, and EDTA. The results revealed Clavispora lusitaniae protease's ability to degrade wheat seeds and flour gluten by 98.7% and 97% respectively under pH 4 for 24 hours at 40 °C. According to this study, this enzyme could be a potential candidate for the food industry, particularly for treating wheat seed and flour to reduce the immunogenicity of gluten.
Keywords:
Clavispora lusitaniae PC3
; protease
; solid- state fermentation
; enzyme purification
; gluten degradation
1. Introduction
Proteases (E.C. 3.4.21-24.x) are a significant group of industrial enzymes and represent 60% of the enzyme market, estimated to be $7 billion in 2023 [1,2,3]. They hydrolyze proteins into small polypeptides or amino acids [4]. Protease enzymes degrade proteins at different sites [5]. All proteases in clans and families can be classified into asparagine proteases, aspartic proteases, cysteine proteases, glutamic proteases, mixed proteases, metalloproteases, threonine proteases, serine proteases, and unknown proteases based on these phylogenetic relationships and mechanisms of action [6,7].
A variety of enzymes are used in place of chemicals, with microbial enzymes—particularly proteases—being used most frequently in the industrial sectors for the production of food products (particularly in cheese making). They are also used in detergents, paper and waste management strategies [7,8,9], pharmaceutical, textile, feather bioconversion, leather industries [10,11], as well as in silver recovery silver extraction from cast-off X-ray films [12,13] and medical sectors [1,14].
Microbial proteases are the leading catalysts with a sharp rise in global demand in recent decades [15] because of their low cost of production, their extracellular nature, availability, and stability. Furthermore, given that the growing medium's estimated cost accounts for 30–40% of the industrial enzyme production cost [16], protease production necessitates lower-cost media, which can be obtained by utilizing easily accessible and reasonably priced agro-industrial wastes. However, the composition of tomato pomace represents a very significant source of lycopene, lipids, ascorbic acid, fiber, and protein [17]. In addition, bread waste contains a significant amount of starch, which is easily hydrolyzed into monomer sugars with the help of enzymes. This allows the exploitation of this waste in several areas, in particular, animal feed and the production of enzymes and vitamins.
The optimization of the nutritional and physicochemical conditions of the culture medium has been realized by the application of statistical designs. The Plackett-Burman designs (1946) [18] are used for the selection of the factors with significant effect on proteolytic production, and the Box-Wilson designs (1951) [19] are applied to determine the optima of the factors selected during the first design.
Molds and bacteria have historically been used in the production of industrial enzymes. In recent years, the use of the production of proteases from yeast has increased in demand [20]. As there aren't many highly proteolytic yeast species [21], it would be interesting to isolate new protease-producing yeast strains. However, only a few studies have been performed on the production of enzymes by the yeast Clavispora lusitaniae. Thid yeast has shown its ability to produce phytase [22], amylopullulanase [23] and alpha amylase [24].
Because of its viscoelasticity, gluten is essential to the baking and food processing industries. Conversely, wheat proteins—particularly gluten—have a negative impact on those who are genetically susceptible to conditions like celiac disease (CD), wheat allergy, gluten intolerance, asthma in bakers. Patients with CD currently follow a stringent lifelong gluten-free diet. An alternate study aimed at lowering the immunostimulatory celiac hazardous peptide sequence in wheat prolamins is now underway. through the application of contemporary technologies like proteolytic enzymes that specifically cleave proline [25].
Currently, CD patients strictly stick on to life-long gluten-free diet. An alternative investigation is in progress to reduce the immunostimulatory celiac toxic peptide sequence in wheat prolamins. By using modern technologies such as proline cleaving specific proteolytic enzymes, sourdough technology, chemical deamination, and enzymatic modification gluten [25]. It would be interesting if the acid protease produced by Clavispora lusitaniae had the capacity to hydrolyze.
The aim of this research is to study the production, purification, characterization, and application of yeast protease isolated from potato waste. The strain Clavispora lusitaniae isolated from potato peels was used for protease production on a low-cost medium. Production optimization was realized by the statistical design of experiments via Plackett-Burman design and RSM followed by the protease's purification and characterization. Furthermore, the gluten-digesting ability of the enzyme is performed.
2. Materials and Methods
2.1. Isolation and Identification of Protease-Producing Strain
2.1.1. Sampling and Yeast Strains Isolation
Potato wastes (PW) were collected from waste dumping sites in restaurants. Ten grams of PW was put into a sterile 250 ml flask Erlenmeyer containing 90 ml of sterile Yeast Malt (YM) liquid medium (glucose 2%, malt extract 1% and yeast extract 1%). After 48h of incubation in a shaker at 28°C and 150 rpm, the presence of yeast cells was examined using light microscopy. After several dilutions with sterile distilled water, 0.1 ml of the culture was spread out on the surface of the YMA medium supplemented with 0.01% chlortetracycline (Sigma), and 0.01% chloramphenicol (Sigma) to prevent bacterial growth.
After incubation at 28 ◦C for 5-7 days, strain colonies were periodically checked; the representative colonies of each morphological type were purified, and maintained on YPGA: yeast extract 1%, peptone 1%, glucose 2%, and agar 2% stored at 4 ◦C. The ability of isolated strains to produce protease is studied on Skim Milk Agar (SMA) medium:1% skim milk, 2% agar. The inoculated plates were incubated at 30°C for 4 to 5 days. Yeasts showing a clear zone around the colonies were selected and, considered protease-positive [7,26].
The yeast colonies were purified and conserved on YMA, and YPGA media at 4°C. They were also, stored in cryo-beads at -80°C, for long storage.
2.1.2. Characterization and Identification of Strain PC3
Morphological studies of yeast isolates were performed using YM agar and YM broth incubated at 28°C for 72 h. To induce the sporulation of yeasts, RAT medium is used to study the presence of mycelium.
Physiological and biochemical characteristics of the yeast isolates were determined according to the API test (API 32C AIX; BioMerieux S.A., Marcyl’Etoile, France).
The selected yeast strain was submitted for identification via molecular approach (sequencing of the D1/D2 domain of 26S rRNA gene). DNA extraction was carried out according to Bennamoun et al., (2016) [17]. DNA was first amplified as a template by the PCR method using the primers V9G (50-TGCGTTGATTACGTCCCTGC-30) and RLR3R (50-GGTCCGTGTTTCAAGAC-30; Sigma-Aldrich Co). A 600-650 bp region was sequenced by the forward primer (50-GCATATCAATAAGCGGAGGAAAAG-30) and the reverse primer NL4 (50-GGTCCGTGTTTCAAGACGG-30; Sigma-Aldrich Co). The PCR products were sequenced using a commercial sequencing facility (Macrogen, Amsterdam, Netherlands). The sequences obtained were compared with those included in the GenBank database (Blast freeware from http://www.ncbi.nlm.nih.gov/BLAST) [17].
2.2. Protease Production
2.2.1. Substrates Preparation
Tomato pomace (TP) was obtained from the tomato-processing plant Ben Azzouz (Skikda, Algeria). It was dried in the open air (25–30 ◦C, 4 days). The bread waste (BW) was collected from restaurants, in Constantine, Algeria, and was cut into pieces, and dried at room temperature for 3 days [27]. The dried bread waste and tomato pomace were milled with the Pulverisette 14-rotor mill Fritsch (France). The milled TP and BW were sieved (2mm), and preserved in hermetically sealed boxes.
The composition of tomato pomace used in this study (in dry weight basis) was as follows: 22.4% total sugars, 21.2% protein, and 6.1% minerals and that of bread is 48.46% carbohydrate, 46.3 % starch, 8.4% proteins (gluten), 1.3% minerals.
2.2.2. Fermentation
The production of Clavispora lusitaniae protease is carried out by solid fermentation and liquid fermentation in the presence of TW or BW or a mixture of both.
For solid-state fermentation (SSF), 05 Five grams of tomato waste, leftover bread, and a mixture of both (2.5g tomato waste and 2.5g waste bread) are introduced, separately, into 250 ml Erlenmeyer flasks (3 tests are carried out for each waste) then, a quantity of distilled water is added to obtain moisture level of 60% [20]. After sterilization, the microbial inoculum of 105 cells/ml was used, and contents must be mixed well using a sterile rod (the operation was carried out aseptically) and then incubated for 48 h at 30°C.
After incubation, fermented substrate is mixed with 50 ml of Tween 80 solution (0.1%) for 10 min and centrifuged at 10000 g at 4°C for 20 min. The supernatant was filtered using Whatman paper n°1, and the filtrate was used as a crude enzyme.
For submerged fermentation (SmF), the flour from tomato waste and bread waste and a mixture of both was diluted in distilled water at a concentration of 5%. Media were divided at a rate of 40 ml per 250 ml Erlenmeyer flasks and were autoclaved at 121°C for 20 min. The inoculated sterile media (105 cells/g) were incubated in a shaker incubator at 30°C for 48h at 150 rpm. After incubation, the culture media are centrifuged at 10000 rpm at 4°C for 20 min, and the supernatant represents the enzymatic extract.
2.3. Production Optimization
Protease production is influenced by various production parameters, including nutritional and environmental parameters. For this, the composition of the culture medium was optimized according to statistical methods using two experimental designs.
Screening of significant factors: Plakett and Burman's design (PBD) (1946) [18] was applied to determine factors with a significant positive or negative effect on protease production, using a minimum number of experiments [28]. For this, a matrix of 12 experiments was used to evaluate the effect of the three dummy variables (D, H, and K) and eight reel variables (factors) studied. Each factor is studied at two levels high (+1) and low (- 1) [29,30] namely, A: Temperature, B: Moisture, C: Inoculum, E: Glucose, F: «Corn steep liquor»: G: (NH₄) SO₄, I: NaCl, and J: Time fermentation (Table 1).
Optimum determination of selected factors
The Central composite design (CCD) of Box-Wilson (1951) [19] under RSM is an effective experimental tool allowing the study of the interactions of the effective factors and the selection of the optimal conditions of the enzyme production [31].
The three most significant variables (factors): Humidity (B), inoculum (C) and fermentation time (J) selected by Plackett and Burman’s design were optimized by the CCD under RSM. Each factor in the design was studied at five levels (Table 2). Statistical analysis is performed using Minitab 19 software.
2.4. Analytical Methods
2.4.1. Protease Activity Assay
The proteolytic activity in enzymatic extract of Clavispora lusitaniae PC3 was measured according to method described by Benkahoul et al., (2020) [28] with minor modifications using casein as substrate. The mixture containing 0.625 ml of 2.5% casein as the substrate and 0.25 ml of enzymatic extract and 0.375 ml of buffer was prepared. It was incubated at 40°C for 30 min, and then 2.5 ml of trichloroacetic acid (TCA) 4% were added to the reaction mixture to terminate the enzyme reaction. After 10 min, the reaction mixture was filtrate and to 0.5 ml of filtrate, 2.5 ml of Na2CO3 2% and 0.25 ml of 25% Folin Ciocaltreu reagent (before use, 25 ml of reagent diluted with 100 ml of distilled water) were added and the mixture was incubated at room temperature. After 30 min, the protease activity was read at 750 nm in a spectrophotometer (UV/VIS) using a tyrosine standard curve as a reference. Simultaneous controls containing enzyme, heat-denatured enzyme and substrate were maintained. One unit of protease activity was defined as the amount of enzyme required to liberate 1 µmole tyrosine/min.
2.4.2. Protein Assay
Total protein content was determined by Lowry et al. (1951) [32] using bovine serum albumin as reference.
2.5. Purification of Protease
Clavispora lusitaniae PC3 protease was partially purified by ammonium sulfate precipitation and gel filtration chromatography. Crude enzyme was precipitated by ammonium sulfate (60% saturation) and kept overnight at 4°C. The precipitate was collected by centrifugation at 10,000 rpm for 30 min, dissolved in 0.1 M phosphate buffer (pH 5), and then dialyzed overnight at 4°C against the same buffer to eliminate residual ammonium sulfate. The concentrated dialysate was loaded in the Sephadex G-100 column equilibrated with the same phosphate buffer (pH5). Fractions (1.5ml) were collected and examined, for protein content and protease activity.
2.6. Biochemical Characterization of Clavispora lusitaniae PC3 Protease
2.6.1. Effect of Temperature on Protease Activity
The optimal temperature for the enzyme activity was determined at temperatures of 30 to 80°C.
2.6.2. Effects of pH on Protease Activity
The effect of pH on the enzyme activity was determined by incubating the purified enzyme between pH 3 and 12 using the standard assay condition. The buffers used were 0.2M citrate-sodium buffer (pH3 to pH7), 0.1M glycine-NaOH buffer (pH 8 –12).
2.6.3. Effects of Metal Ions and Chemicals on Enzyme Activity
The effect of ions on protease activity was studied in the presence of 5mM salts such as CuSO4, FeSO4, KCl, MnSO4, ZnSO4, MgSO4, NaCl, and CaCl2. EDTA, urea, and 5mM SDS, β-marcaptoethanol, tween 80 and tween 20 were also tested for enzymatic activity. After incubating the enzyme with each metal ion or reagent at 60°C for 10 min, the activities are measured and compared with that of the control incubated, under the same conditions without effectors [23]. All assays were repeated three times, and the values presented are averages of the three experiments.
2.7. Gluten Digestion
2.7.1. Proteolytic Degradation of Gluten
Wheat flour was weighed into conical flasks (25 g/flask) containing 12 to 15 ml of Clavispora lusitaniae PC3 protease (pH 4.0), for hydrolysis. The procedure used by Luoto et al. (2012) [33] was followed with a few minor modifications. The flasks were incubated in a water bath that were shaken at 40 °C for 8, 16 and 24 h incubation. For the control, the enzymatic extract was replaced by distilled water without incubation.
25 g of wheat seeds were mixed in 50 ml of enzymatic extract and then incubated at 40°C for 8, 16, and 24 hours. After drying out, the mixture is crushed to make flour, which is then combined with 12 to 13 ml of distilled water. For the control, the wheat seeds are crushed without enzymatic treatment, and 25 g of flour is mixed with 12 to 15 ml of distilled water.
2.7.2. Gluten Determination
The dosage of gluten is based on its solubility in salt water and on its agglomeration during mixing under a stream of water [34].
After the addition of the enzymatic extract and distilled water to 25g of wheat flour. Form a homogeneous dough that, after two to three minutes of kneading, should not adhere to the walls of the container. After resting for 15 minutes, this dough is kneaded by hand to transform it into a homogeneous piece of dough that stretches perfectly. This dough is kneaded under a stream of water by compressing it slightly, or by placing it above a sieve intended to retain the gluten fragments that are entrained, to recover them (the starch is therefore eliminated while the gluten gradually welds itself to itself). When the gluten forms a homogeneous mass, the water flow is increased to carry out the washing until transparent washing water is obtained. The gluten is then squeezed out by compressing it strongly several times between the palms of the hands, which have been carefully wiped until it adheres to the hand. Finally, the wet gluten is quickly weighed.
Dry gluten is obtained by drying wet gluten at a temperature of 102°C, until a constant weight is obtained, after approximately 18 hours.
Results expression
Wet gluten weight (WG) is expressed in grams per 100g of flour by:
WG=100(Mw /25)
Dry gluten (DG) is expressed in grams per 100g of flour by:
DG= 100(Md /25)
Mw: mass in g of wet gluten; Md: mass in g of dry gluten.
3. Results and Discussion
3.1. Isolation and Identification of Yeast Isolate
Proteases are a group of enzymes that find wide applications in industries. Due to the enhancement of continuous demand for protease utilization for specific properties, researchers are interested in novel protease sources.
Among 70 isolated strains from potato wastes (PW), 10 showed their ability to hydrolyze proteins and were considered. One strain was selected for further studies based on the maximum zone of clearance.
The isolated strain seems to be yeast according to their cultural, and biochemical characteristics. By analysis of the 26S rRNA gene D1/D2 region sequencing revealed that the isolate was Clavispora lusitaniae. The identified strain was submitted to GenBank with the accession number PP057739 (Table 3).
Different yeast species were reported to produce protease such as Rhodotorula mucilaginose [35], Pichia anomala CO-1 [36], Candida humicola [37], Yarrowia lipolytica [20,38], Candida albicans [39], Wickerhamomyces anomalus and Metschnikovia pulcherrima [40], Pichia membranifaciens [41], Candida tropicalis [21].

Table 3.
Identification from biochemical (ID 32C), microscopic and molecular biology characters.
| Strain Code | Biochemical Characters (ID 32C) | Morphological Characteristics of the Selected Isolate | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Colony Characteristics | Cell Shape | Vegetative Reproduction | Mycelium (RAT) |
Sporulation | ||||||
| PC3 | 5157 3701 17 lac- et ESC+ |
Smooth, glistening, butyrous, White to cream colored, entire margin. | Sub-globose, ovoid to elongate | Budding (unipolar and bipolar) | Pseudo mycelium |
+ | ||||
| Protease production | Strain on YPGA | Microscopic Characteristics | ||||||||
 |
 |
 |
||||||||
| Blast results | ||||||||||
| Id sequences | Query (bp) | Species | Accession number | % max ident. | ||||||
| ITS_2156ZAB059 | ITS 692 | Clavispora lusitaniae strain CBS 6936 T | PP057739 | 676/676(100%) | ||||||
3.2. Fermentation and Substrate Selection
Clavispora lusitaniae PC3 protease production was studied in SmF and SSF on three media: tomato pomace (TP), bread waste (BW), and the TP and BW mix. The protease activity was assayed at different pH.
From Figure 1 and Figure 2, it appears that Clavispora lusitaniae isolated from potato peels shows protease activities when it is grown in solid fermentation and liquid fermentation of different substrates studied.
In SMF, in the presence of tomato pomace, bread waste, and a mixture of the two substrates, the synthesis of acid protease is the most dominant followed by Less important neutral protease activity and a minor quantity of alkaline protease (Figure 1). Nevertheless, the best production of acid protease (456.2 IU) is obtained by the mixture of TP and BW.
In SSF, we notice that Clavispora lusitaniae PC3 cultivated on the different substrates (tomato pomace, bread waste, and a mixture of both substrates) mainly produces acidic protease followed by a low rate of neutral protease and no alkaline protease synthesis. However, acidic protease production is the most important (11205.78 ±88.5 IU/g) for TP and BW mix (Figure 2). It appears that the TP and BW mix used constitute a good substrate for protease production by Clavispora lusitaniae PC3 in SmF because an interesting result may be obtained without any nutriment supplementation, owing to its enough biochemical composition, particularly in proteins.
This observation is in agreement with the experimental results obtained by Hisham et al. (2017) [16] who found that among 23 strains isolated from different natural sources collected from Abha region, Kingdom of Saudi Arabia, five successful yeasts for protease production were selected. At pH6 H. uvarum KKU-M19c produces neutral protease, whereas the other four yeast isolates, R. mucilaginosa KKU-M12c, C. albidus KKU-M13c, P. membranifaciens KKU-M18c, and C. californica KKU-M20c, produced acidic proteases at pH 4 and neutral proteases at pH 6. However, none of the five yeasts produces alkaline proteases.
According to the results (Figure 2), the non-existence of alkaline protease production can be explained by the fact that the pH of the medium is not favorable. The pH of TP, BW, and TP-BW mix media is acid (pH 4.2, pH 5.4, and pH 4.7 respectively). The synthesis of protease depends on the pH of the medium as it plays a role in regulating its yield and affects the type of protease produced [42]. Similar results were also obtained by Rucha et al., (1982) [43], after their study applied for C. albicans protease production and found the strain produced acid protease activity, but not neutral or alkaline protease activity when it was grown at acidic pH. During his study, Kim (2019) [36] showed that When the yeast Pichia anomala CO-1 was grown in an acidic medium (pH 4) at 30°C, secreted an extracellular neutral protease with optimum pH 7.
By the results obtained with Y. lipolytica and C. olea, it was suggested that pH seems to be a major factor in regulation and neutral or alkaline proteases may not be produced at acidic pH [21]. the presence of a low level of neutral protease from Clavispora lusitaniae is probably due to the change in pH of the medium during fermentation. Aspartic peptidases are predominantly active at acidic pH, however some also demonstrate activity at neutral pH, according to James (2004) [44], whereas carboxypeptidases catalyze reactions in the pH range of 6 to 9 [20,45,46].
The diversity of protease production in Clavispora lusitaniea PC3 grown in different media can be explained by the adaptation of the strain to the culture medium, particle size, composition, and the pattern of degradation of the substrate [21].
It also appears that the combination of tomato pomace and bread waste is the best for both types of fermentation, allowing a more important enzyme yield than each of the substrates used separately. This agrees with the results of Sharma et al., (2017) [14] who found that a mixture of two or three different substrates gives greater enzyme yields than use of the substrates individually [14]. It has previously been investigated that reducing the cost of the culture medium can be realized by using wastes of the food industry had previously been investigated. Various agricultural or industrial waste such as sugar cane bagasse, grape waste, wheat or barley straw, pineapple waste, orange peel waste, sugar cane bagasse, wheat bran, and rice bran can be used as a substrate and carrier for the production of microbial protease by SSF [9,35,47].
Clavispora lusitaniae PC3 showed good production of protease acid. This type of enzyme is an active proteolytic enzyme present everywhere in acidic environments. It was also, revealed that yeast proteases are extracellular and biotechnology applicable. Currently, interest in using their acid aspartic proteases in different industrial applications is increasing [40]. Several research showed that many non-saccharomyces yeasts produce extracellular acid proteases and the rate of protease produced could vary significantly from strain to strain [21,40].
Yarrowia lipolytica was studied for extracellular protease production. The yeast secreted the alkaline and acid proteases and less neutral protease production. It was demonstrated that although most Y. lipoIytica strains produce an alkaline protease and no neutral protease, Y. lipoIytica strain 37-1 produces a neutral protease and no alkaline protease. Another study reported that Candida oleu (Y. lipolytica) grown in a glucose BSA-based medium produced a single acid protease, a single alkaline protease, and no neutral protease [48].
Murao et al. (1972) [49] reported that Rhodotorula glutinis is a producer of extracellular acid proteases and found the enzyme to be stable between pH 2.4 and 6.5 after 20 h incubation at 37°C. The yeasts Wickerhamomyces anomalus and Metschnikovia pulcherrima also produce acid proteases [40].
The strain is very efficient in the production of acid protease (Aspartic protease) in the presence of the three media for both types of fermentation. Compared to SmF, SSF has several advantages such as simplicity, low cost, higher yield of enzymes, the concentrated nature of the solid substrate, the reduction of contamination due to the low moisture content, increased volumetric productivity [3], as well as the possibility of using several agro-industrial wastes as a substrate relatively cheap [2]. Moreover, SSF is especially appropriate for fungal enzyme production [3].
For this, we will proceed with the production of acid protease from Clavispora lusitaniae in solid fermentation.
3.3. Optimization of Protease Production from Clavispora lusitaniae PC3
3.3.1. Screening of Significant Factors
The Plackett-Burman Design (PBD) [18] was used for screening factors that affect protease synthesis. Protease activities of the PB experimental design for 12 assays are shown in Table 4.
The statistical analysis of the results is shown in Table 5.
The effect was calculated by changing the response as the factor changes from its lower (−1) level to its higher (+1) level using the student’s t-test. The P value of each factor was also evaluated. The factors with P values less than 0.05 (P value <0.05) were considered as significant factors for protease production [29,50]
Protease production is influenced by the factors with a significant positive or negative effect (p ≤0.05) (Table 3) namely B: humidity (p=0.049), C: inoculum (p= 0.026), and J: incubation time (p=0.004). The reduced polynomial equation of protease production (Y) is as follows:
Y = 16040-1919 Moisture-4909 inoculum+10025 incubation period
The Fisher of the model F-value = 15 with p = 0.024 (≤ 0.05) is very significant, it allows us to conclude that the model is adequate and that the production of the protease is well explained by humidity, inoculum, and incubation period. This is also supported by the value of R2 (coefficient of determination) of the model is 0.9756 (a value > 0.75 indicates aptness of the model) [51] this means that 97.56% of the production of the protease is influenced by the selected factors (Table 6).
3.3.2. Determination of the Significant Factors' Optimum
RMS is a statistical method used to determine the optimum of significant factors in order to develop the best growth medium for Clavispora lusitaniae PC3 and protease production. The results of the optimization experiments are presented in Table 7 and are analyzed using Minitab 17 software.
The P-value and F-value in the ANOVA (Table 8) were determined, and the lesser the P-value significantly the greater the importance of the corresponding coefficient [31,50].
The table indicates a highly significant Fisher value of 52.40 with p=0.000 which means that the chosen model is adequate for the production of the enzyme.
Table 8.
ANOVA for response surface quadratic model.
| Source | DF | Sum of Squares | Mean Square | F-Value | P-Value |
|---|---|---|---|---|---|
| Model | 9 | 333643381 | 37071487 | 52,40 | 0,000 |
| Linear | 3 | 154916915 | 51638972 | 72,99 | 0,000 |
| B | 1 | 114548527 | 114548527 | 161,91 | 0,000 |
| C | 1 | 8457389 | 8457389 | 11,95 | 0,006 |
| J | 1 | 57185579 | 57185579 | 80,83 | 0,000 |
| Square | 3 | 173532796 | 57844265 | 81,76 | 0,000 |
| B2 | 1 | 46462590 | 46462590 | 65,67 | 0,000 |
| C2 | 1 | 113539208 | 113539208 | 160,48 | 0,000 |
| J2 | 1 | 70951960 | 70951960 | 100,29 | 0,000 |
| 2-factor interaction | 3 | 25845123 | 8615041 | 12,18 | 0,001 |
| B*C | 1 | 5888080 | 5888080 | 8,32 | 0,016 |
| B*J | 1 | 8721639 | 8721639 | 12,33 | 0,006 |
| C*J | 1 | 4624970 | 4624970 | 6,54 | 0,029 |
| Error | 10 | 7074816 | 707482 | ||
| Lack of fit | 6 | 5055665 | 842611 | 1,67 | 0,323 |
| Pure error | 4 | 2019151 | 504788 | ||
| Total | 19 | 340718197 |
The results of the statistical analysis of the CCD showed that proteolytic activity could be presented by the following regression equation:
Proteolytic activity = 32584 + 5177 C1-1339 C2 -3557 C3 -5548 C12 -8291 C22 -6841 C32 + 2477 C1C2 + 3361 C1× C3 +2712 C2C3
The analysis of variance gave a highly significant Fisher value of 52.40 with p= 0 confirming a strong relationship between the production of the enzyme and the three selected factors and demonstrating the model is significant and adequate for the enzymatic production [16].
According to Adetunji and Olaniran (2020) [52], the lack of fit determines the model's incapacity to accurately represent data at points that are excluded from the regression. The study's lack of fit value (p =0.323) was not statistically significant which is favorable since it indicates that the model equation might accurately predict the protease production for any interaction between variable's values.
The value of the coefficient of determination (R2) is 0.9792 shows that 97.92% of the variation in acid protease production can be explained by the three factors studied. The adjusted R2 was 0.9605, which has a close relationship with R2. The predicted R2 was 0.9194 indicates a close resemblance with R2 and adjusted R2. This reveals the goodness of the model and the factors optimized [50,52].
Protease production by solid-state fermentation (SSF) was evaluated with TP and BW mix as a substrate for Clavispora lusitaniae PC3. It has been reported that enzyme production is strongly influenced by different culture conditions (physical factors) such as moisture content, inoculum size and fermentation time.
The iso-response plot of protease production is generated from equation (2) using Minitab software version 17. Figure 3. represents the regression equations in 2D and 3D.
Each contour curve represents an infinite number of combinations of two test variables with the third held at its respective zero level. Ellipses are obtained when there is a perfect interaction between independent variables [52]. Contour plots visually interpret the interaction between the two variables, and make it easier to locate optimal experimental conditions. The dark areas provide information on the conditions that optimize the production of the enzyme.
3.3.3. Moisture
For maximum enzyme productivity, the optimum moisture content is required [9]. Low humidity has been reported to reduce the solubility of substrate nutrients and the degree of swelling. However, the increase in the rate of moisture leads to a decrease in the porosity of the substrate and thus limits the transfer of oxygen. A maximum yield of protease from Cl. lusitaniae PC3 was obtained when the humidity was 76.63%. A moisture content of 50% appears to be best for the production of protease from Candida utilis [53], Penicillium sp. and Aspergillus oryzae NRRL 2217 [14]. A humidity rate of 60% was obtained for maximum protease production in Penicillium godlewskii SBSS 25 [54], and for growth and protease production in Y. lipolytica [20]. The moisture of 65% was used for Rhizopus oryzae [55], 80% for T. thalpophilus PEE 14 [14], 120% for Beauveria felina [56], 90-170% for B. circulans, and 100% for B. subtilis [14].
3.3.4. Inoculum Size
The production of enzymes depends on the size of the inoculum, which plays an important role in the rate of fermentation [57]. The value of the inoculum rate of 8.22 x 106 cells/g is within the range of values (106 to 108 cells/ml) provided by the literature [58]. This inoculum rate is also close to 2 x 107 cells/g 2.5 x 107 cells/g used for protease production in Y. lipolytica [20] and Candida utilis [53]. In other fungi, the inoculum of 105 spores /g was used for the production of the protease of Aspergillus oryzae MTCC 5341 [59] and Rhizopus oryzae [55].
The decrease in enzyme production with increasing inoculum size could be due to the rapid initial growth of the microorganism, increased competition for insufficient nutrients in the medium, and reduced dissolved oxygen. Whereas a low amount of inoculum leads to reduced production of proteases in microorganisms, this may be due to a deficit of microbial cells, insufficient to better consume the fermentation medium [60]. There is no precise rate of microbial inoculum suitable for the production of an enzyme.
Media composition and culture conditions are important for microbial growth and enzyme production. To obtain a high production of proteases, it is crucial to find the significant conditions of growth and induction. It was noted that there is no common medium suitable for all producer microorganisms. Each strain has unique specific conditions for the maximum production of an enzyme [61].
3.3.5. Fermentation Time
Enzyme production is greatly affected by the incubation period, which varies from 24 hours to 9 days depending on the type of microorganism, the size of the inoculum, the pH, and the temperature. The maximum production of the acid protease of Clavispora lusitaniae PC3 is obtained after 56.5 hours of incubation. It has been reported that a very prolonged incubation period leads to low enzyme activity, possibly due to reaching the decline phase and death of microorganisms, depletion of nutrients in the medium, or release of toxic metabolites or inhibitors [60].
In fungi, the incubation time for maximum protease production in SSF is 48 h for A. oryzae NRRL 2217, 72 h for A. niger MTCC 281, Penicillium sp., A. flavus and A. terreus [14]. Other studies found that better protease production was obtained at 96 h for P. godlewskii SBSS 25 [54], 120 h fermentation period under SSF in M. circinelloides, A. oryzae MTCC 5341, and Aspergillus spp. [62], and 168 h for Beauveria felina [56].
The predicted value of maximum protease production is 33970 IU/g (Figure 3). Under the optimum fermentation conditions of moisture (76.63%), inoculum (8.22 x 106 cell/g), and fermentation time (56.5h), maximum protease production of 33450± 503 IU/g was obtained. This result is in close agreement with the prediction of the statistical model. and this was found to be 3-fold higher than protease yield recorded in the unoptimized medium (11205.78 ±360 IU/g) (Figure 4). It is concluded that the statistical plans are effective for the optimization of the culture medium for acidic protease production from Clavispora lusitaniae PC3.
Culture medium optimization produced an increase of about three times in protease activity when compared with the activity of the initial production medium (33450±503 IU/g, 11205.75±360 IU/g respectively),
In biotechnology, there is a growing recommendation of use for statistical experimental designs and several scientists were satisfied optimization of protease production from microbial sources using the statistical designs [28,30,52].
In this study, PBD and CCD with RSM were shown to be efficient for optimizing enzyme production. Because of the commercial interest of the enzyme, media production cost was always a primary concern [51]. Protease production by Cl. lusitaniae PC3 in the initial medium was deficient compared with the optimized medium. Therefore, we conclude that Optimization of the initial culture medium showed that it is possible to increase yeast's protease production and reduce the cost of the culture medium using the food wastes.
3.4. Purification of Protease
Purification of protease from Clavispora lusitaniae PC3 led to a recovery of 32.94% protease with a purification rate of 19.76 (Table 9).
3.5. Protease Characteristics
3.5.1. Effect of pH on Protease Activity
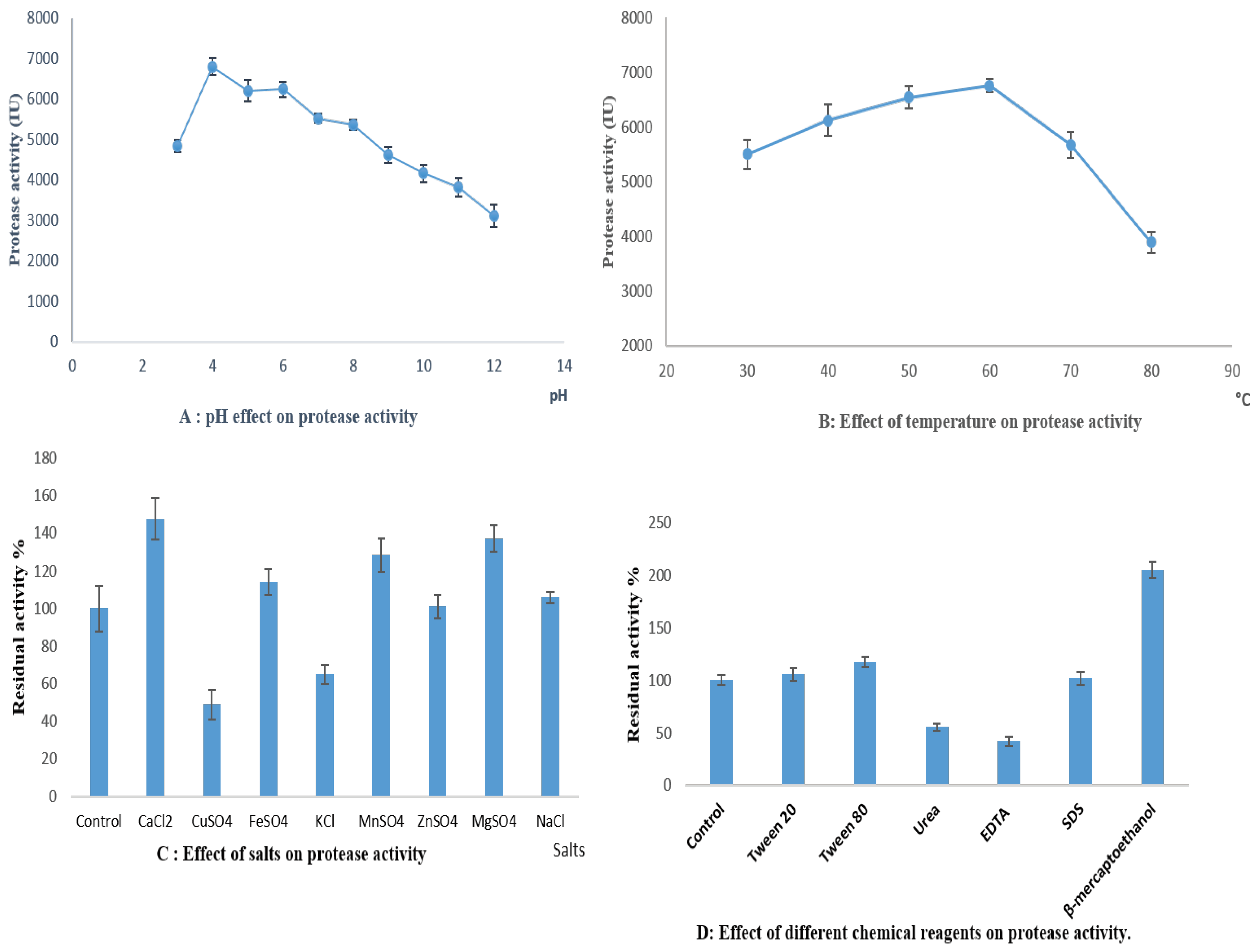
Analysis of the experimental results of the protease activity by the ANOVA method reveals that the pH affects the protease activity (F=58,13; p=0.000). The study showed a wide range of activity from pH 3 to 12, with a maximum at pH 4 (Figure 4A). These results suggested that the Cavispora lusitaniae PC3’s enzyme was an acidic protease, which is expectedfor protease s produced by fungi [63]. Considerable activity was observed at pH 6 (95, 6%), Beyond this pH value, a decline in activity is observed (Figure 5A).
The extracellular acidic protease produced by Rhodotorula glutinis, Candida parapsilosis, Candida tropicalis and Candida humicola, Rhodotorula mucilaginosa L7 and Rhodotorula oryzicola has a pH optimum of 2.8 to 3.5, 4.3, 3.4 to 3.8, 4, 5 and 6.51 respectively [37,64,65].
Furthermore, It was revealed that in addition to producing an alkaline serine protease, C. albicans [37,66], Candida olea [67], Saccharomycopsis lipolytica [68], and Rhodotorula glutinis [35,69] secrete acidic proteases with a molecular mass ranging from 30 to 45 kDa and an optimal pH range of 2.5 to 3.9. Similarly, Schlander et al., (2016) [40] showed that the pH optimum of W. anomalus 227 and M. pulcherrima 446 was 3 and 4 respectively.
Srividya (2012) [70], reported that a pH acid of 5 was ideal for the fungal protease activity of Aspergillus sp. The pH optimum of Aspergillus niger protease was 3 - 4 [71,72,73].
As suggested by their name, acidic proteases are aspartic proteases that are mostly active in the acidic pH range, which is between pH 3.8 and 5.6. Their ideal pH is between 3 and 4 [11,74].
The principal producers of acid proteases are fungi, as opposed to bacteria, which mostly generate alkaline proteases. The most frequently documented genera of acid proteases are Aspergillus, Penicillium, Endothia, Mucor, etc. [75]. Numerous reports have indicated that certain yeasts and molds secrete new acid proteases [11,76].
Figure 5.
Characteristics of Clavispora lusitaniae PC3 protease.
3.5.2. Effect of Temperature on Protease Activity
Temperature is one of the major factors affecting the enzyme activity. The influence of temperature was examined by reaction of the enzyme at temperatures ranging from 30℃to 80℃. The profile of protease activities as a function of temperature presents a somewhat broad aspect (Figure 5. B). The enzyme activity highly depends on the temperature (highly significant difference with F=12,6 (p 0.001). Clavispora lusitaniae PC3 protease showed maximum activity at 60°C. however, considerable activity was observed at 30°C (81.5%) and more than 90% of the maximal activity between 40℃ and 50℃, while at 70℃ and 80°C, the enzyme activity was 84.2% and 58% respectively (Figure 5B).
A similar result was detected for protease produced by Rhodotorula glutinis that showed maximum activity at 60°C and the enzyme preserved 80 % of its maximal activity after incubation at 70 °C [69]. Also, Rhodotorula oryzicola protease reached its maximum at 63.04°C [65].
Proteases with optimal temperatures between 50 and 60 °C have also been reported to be produced by a variety of cold-adapted organisms [77,78,79]; however, the mechanisms underlying these enzymes' thermal stability remain undetermined [64].
This temperature is quite different from the optimal values reported for protease from Rhodotorula mucilaginosa L7 with an optimal temperature of 50°C [64]. Aguilar and Sato, (2018) [80] showed that acid proteases (Aspartic proteases) activity has an optimum pH ranging between 3 and 5, and an optimum temperature of 40 to 55°C. Clavispora lusitaniae PC3 protease is an acid protease with an optimum pH of 4 and an optimum temperature of 60°C. No study on the protease of the species Clavispora lusitaniae has been reported in the literature However, the study by Nakamura et al., 2000 [22] revealed that Clavispora lusitaniae produced a phytase having an optimal temperature of 70°C. Amylopullulanase from Clavispora lusitaniae ABS7 isolated from arid zone wheat has an optimal pH of 75-80°C [23].
3.5.3. Effect of Metal Ions and Additives on Protease Activity
According to an assessment of the effects of different metal ions on enzyme activity, the Clavispora lusitaniae PC3's protease activity was stimulated by Ca2+, Mg2+, Mn2+, and Fe2+ to 47, 67%, 37, 16%, 28,5, and 14,15%, respectively (Figure 5C), indicating that these ions had a functional role in the molecular structure of the enzyme [81]. Previous research investigations have mentioned the role that Ca2+ and Mg2+ ions play in maximizing the production of enzymes [9]. The best enzyme activity and stability were obtained with Ca2+, with an increase in activity suggesting that metal ions had compatibility with the enzyme. The same observation had been made, according to Srividya (2012) [70]. Whereas enzyme activity was significantly reduced by the addition of Cu2+, and K+, compared to the control (Figure 3C), and was stable against Zn2+ and Na+.
From the Figure 5D, it appears that mercaptoethanol significantly increased protease activity (2 fold), followed by tween 80 (17.7%). The enzyme is inhibited by urea and EDTA and remains stable in the presence of tween 20 and SDS.
EDTA inhibits protéase activity, with a 58,12% loss of its residual activity. This result indicates that the protease is a metalloenzyme and the activity is CaCl2 dependent. Although they are not necessary for their catalytic functions, Calcium ions act as inducers and stabilizers of many enzymes against thermal denaturation and self-digestion and protect them from conformational changes [63]. Different ions have shown a positive influence on the protease activity of different bacterial strains such as Ca2+, Mg2+, and Mn+2 on the enzyme of B. circulans [14] and Aspergillus braziliensis [63]. Mn2+ enhanced Pseudomonas thermaerum GW1 protease activity five-fold, while Cu2+, Mg2+, and Ca2+ activated it moderately [82]. The presence of CaCl2 enhanced the alkaline protease activity of A. niger by 105.3% [83].
3.6. Action of Clavispora lusitaniae Protease on Gluten
The food and beverage industries depend heavily on the Aspartic proteases (EC 3.4.23), also known as acidic proteases. They are a subfamily of endopeptidases that have been isolated from diverse sources such as fungi, bacteria, plants, and animals. Microbial origin proteases are being employed in place of animal origin enzymes, such as rennin, in the process of clotting milk for the making of cheese [76]. Likewise, acid proteases are employed in bakeries to enhance dough properties. The acid proteases are also used to re remove the haze from juices and beer [74].
The aspartic protease of Clavispora lusitaniae PC3 exhibits the best activity at low pH (pH 4) and temperature of 60°C, suggesting that the enzyme is active at acidic pH and suitable for the food industry and beverage industry [62].
Because gliadin, one of the toxic protein fractions of gluten, is responsible for the development of celiac disease due to the presence of celiac disease eliciting epitopes in gluten [84], gluten proteins play an active role in celiac disease and their ingestion leads to damage to the villi of the small intestine. Therefore, enzymatic treatments of wheat gliadins appear to be an alternative method for reducing celiac activity.
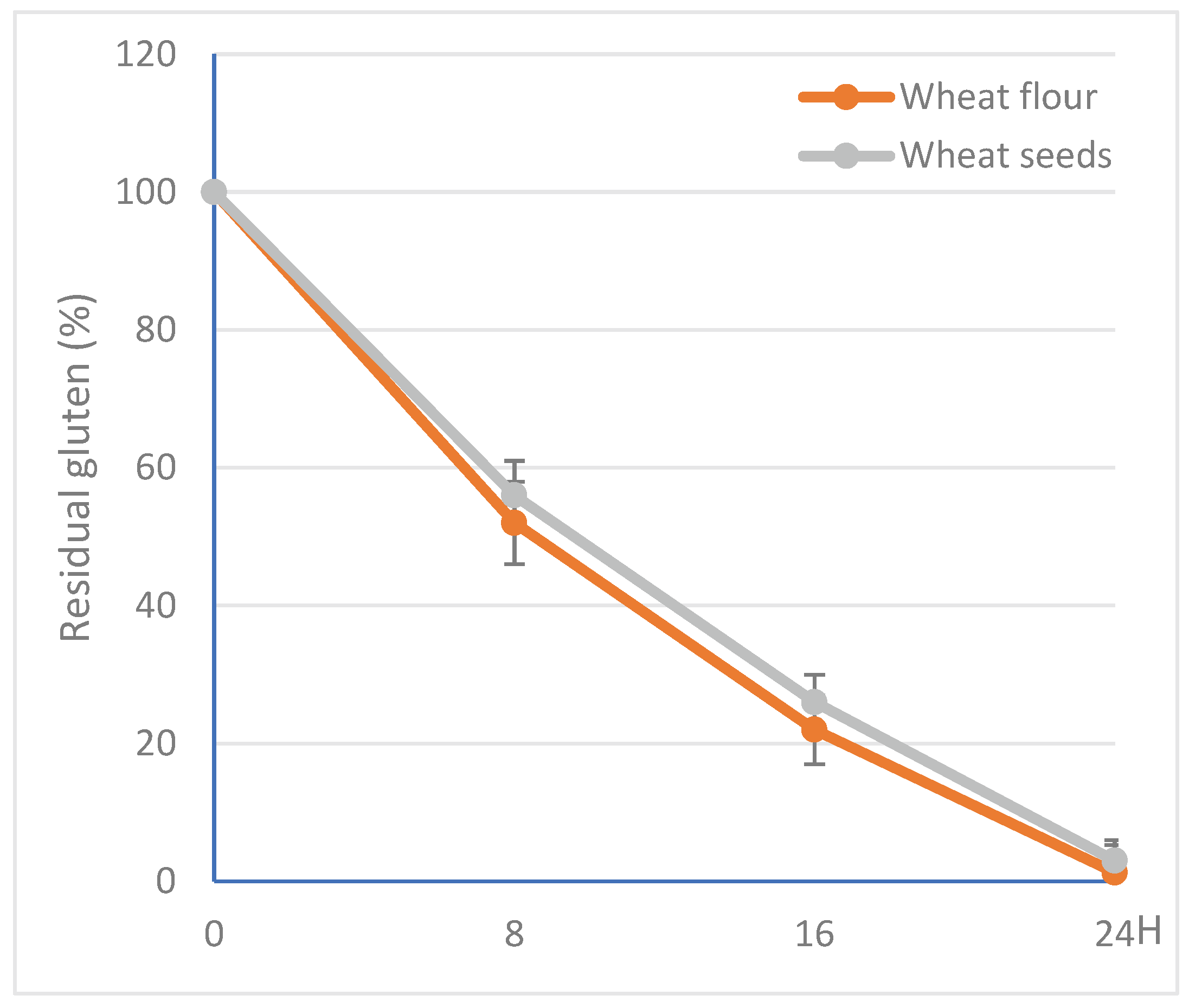
The action of Clavispora lusitaniae PC3 protease on gluten was studied on Wheat seeds and flour. The results (Figure 6) show a reduction in gluten over time and reach 97% for wheat seeds and 98.7% for wheat flour after 24 hours of incubation. Our results corroborate those of Luoto et al., (2012) [33] who revealed that the protease treatment of native wheat, barley, and rye malt allowed the reduction of prolamins by 99.95%, 99.17%, and 99.95% respectively. After 48 hours of incubation, Walter et al. (2014) [85] noted a reduction of 99% of the gluten content of wheat bran. Heredia-Sandoval et al. (2018) [86] also obtained a reduction in gluten content of 98% in wheat flour at 8 h of incubation.
In the study by Luoto et al., (2012) [33], Aspergillus niger prolyl endoprotease was required to reduce germinated wheat products' gluten. The proteolytic activity of strain Clavispora lusitaniae PC3 presented an interesting capacity to degrade gluten.
Figure 6.
Action of Clavispora lusitaniae PC3 protease on gluten.
4. Conclusions
The present study selected Clavispora lusitaniae PC3 isolated from potato wastes for its ability to produce protease. It gave the best production of acidic protease in SSF on tomato pomace and bread waste mix (11205.78±360 IU/g). The optimization of the production of the protease was carried out by the Plackett-Burman plan for the selection of factors with a significant effect on protease production. Box-Wilson's plan made it possible to determine their optima, namely the humidity at 76.63%, the inoculum of 8.22 x 106, and the fermentation time of 56.5 hours, and the production increased 3 fold (33450±503 IU/g). The enzyme is 19.76-fold purified with a yield of 32.94%. Its maximum activity is obtained at pH 4 and 60°C. This acid protease is activated by CaCl2 and inhibited by EDTA. This leads to the conclusion that it is probably a Ca2+-dependent metalloenzyme.
As per our knowledge, acidic protease production by Clavispora lusitaniae species has not been reported. Thus, according to this study, the Clavispora lusitaniae PC3 is a very promising strain for biotechnological application. Its acid protease was produced in a low-cost medium, providing a novel and effective alternative for producing a higher-value product. These yeast extracellular enzymes have the potential to stabilize beer and wine. The use of acid proteases is important to prevent haze formation in wine. To minimize the loss of volume and aroma of the wine, these enzymes represent an interesting alternative to bentonite fining by their degradation of haze-forming proteins. This study showed the possibility of using this enzyme as a natural alternative to chemicals in food processing industries in particular to produce low-gluten wheat and so to reduce gluten in cereal products for celiac patients as a step forward for clinical studies to treat celiac disease.
Author Contributions
S.D.D. performed the experiments and wrote the paper; A.A-K. and E-O. helped in the statistical analysis and data interpretation, B.L., helped in data interpretation; D.A. helped in wrote the paper and creation of figures; N.T. contributed to the laboratory, reagents, and materials and L.G. supervised the study. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The authors are thankful to the Algerian Ministry of Higher Education and Scientific Research for funding this work. The author Scheherazed Dakhmouche-Djekrif is grateful to all the people for their helpful comments on the manuscript.
Declaration of Competing Interest
None.
References
- Nadeem, F.; Mehmood, T.; Naveed, M.; Shamas, S.; Saman, T.; Anwar, Z. Protease Production from Cheotomium globusum Through Central Composite Design Using Agricultural Wastes and Its Immobilization for Industrial Exploitation. Waste Biomass Valori. 2019, 12, 6529–6539. [Google Scholar] [CrossRef]
- Gimenes, N. C.; Silveira, E.; Tambourgi, E. B. An Overview of Proteases: Production, Downstream Processes and Industrial Applications. Sep. Purif. Rev. 2021, 50, 223–243. [Google Scholar] [CrossRef]
- Alahmad Aljammas, H.; Yazji, S.; Azizieh, A. Optimization of protease production from Rhizomucor miehei Rm4 isolate under solid-state fermentation. J. Genet. Eng. Biotechnol. 2022, 20, 82. [Google Scholar] [CrossRef]
- Bhavikatti, J. S.; Bodducharl, S. M.; Kamagond, R. S.; Desai, S. V.; Shet, A. R. Statistical optimisation of protease production using a freshwater bacterium Chryseobacterium cucumeris SARJS-2 for multiple industrial applications. 3 Biotech. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Marathe, S.K.; Vashistht, M.A.; Prashanth, A.; Praveen, N.; Chakraborty, S.; Nair, S.S. Isolation, partial purification, biochemical characterization and detergent compatibility of alkaline protease produced by Bacillus subtilis, Alcaligenes faecalis and Pseudomonas aeruginosa obtained from sea water samples. J. Genet. Eng. Biotechnol. 2018, 16, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N. D.; Barrett, A. J.; Thomas, P. D.; Huang, X.; Bateman, A.; Finn, R. D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the Panther database. Nucleic Acids Res. 2017, 46, D624–D632. [Google Scholar] [CrossRef] [PubMed]
- Song., P.; Zhang, X.; Wang, S.; Xu, W.; Wang, F.; Fu, R.; Wei, F. Microbial proteases and their applications. Front. Microbiol. 2023, 14, 1236368. [Google Scholar] [CrossRef] [PubMed]
- Aruna, V.; Chandrakala, V.; Angajala, G.; Nagarajan, E.R. Proteases: An overview on its recent industrial developments and current scenario in the revolution of biocatalysis. Mater. to day proc. 2023, 92, 565–573. [Google Scholar] [CrossRef]
- Balachandran, C.; Vishali, A.; Arun Nagendran, N.; Baskar, K.; Hashem, A.; Abd_Allah E., F. Optimization of protease production from Bacillus halodurans under solid state fermentation using agrowastes. Saudi J. Biol.Sci 2021, 28, 4263–4269. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, A. I.; Olaitan, M. O.; Erasmus, M.; Olaniran A., O. Microbial proteases: A next generation green catalyst for industrial, environmental and biomedical sustainability. Food Mater. Res. 2023, 3, 1–15. [Google Scholar] [CrossRef]
- Solanki, P.; Putatunda, C.; Kumar, A.; Bhatia, R.; Walia, A. Microbial proteases: ubiquitous enzymes with innumerable uses. 3 Biotech 2021, 11, 428. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, S.; Iqbal, S.; Ahmed, I.; Janjua, H. A. Production, Optimization, and Partial Purification of Alkali- Thermotolerant Proteases from Newly Isolated Bacillus subtilis S1 and Bacillus amyloliquefaciens KSM12. Processes 2022, 10, 6, 1050. [Google Scholar] [CrossRef]
- Wahab, W. A. A.; Ahmed, S. A. Response surface methodology for production, characterization and application of solvent, salt and alkali-tolerant alkaline protease from isolated fungal strain Aspergillus niger WA 2017. Int. J. Biol. Macromol 2018, 115, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. M.; Kumar, R.; Panwar, S.; Kumar, A. Microbial alkaline proteases: Optimization of production parameters and their properties. J. Genet. Eng. Biotechnol. 2017, 15, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, A. I.; Olaitan, M. O.; Erasmus, M.; Olaniran, A. O. Microbial proteases: A next generation green catalyst for industrial, environmental and biomedical sustainability. Food Mater. Res. 2023, 3, 12. [Google Scholar] [CrossRef]
- Hesham, A. E. L.; Alrumman, S. A.; Al-Dayel, M. A.; Salah, H. A. Screening and genetic identification of acidic and neutral protease-producing yeasts strains by 26S rRNA gene sequencing. Cytol. Genet. 2017, 51, 221–229. [Google Scholar] [CrossRef]
- Bennamoun, L.; Hiligsmann, S.; Dakhmouche, S.; Ait-Kaki, A.; Labbani, F.-Z. K.; Nouadri, T.; Meraihi, Z.; Turchetti, B.; Buzzini, P.; Thonart, P. Production and Properties of a Thermostable, pH—Stable Exo-Polygalacturonase Using Aureobasidium pullulans Isolated from Saharan Soil of Algeria Grown on Tomato Pomace. Foods 2016, 5, 72. [Google Scholar] [CrossRef]
- Plackett, R. L.; Burman, J. P. The design of optimum multifactorial experiments. Biometrika 1956, 33, 305–325. [Google Scholar] [CrossRef]
- Box, G. E. P.; Wilson, K. B. On the Experimental Attainment of Optimum Conditions. J. R. Stat. Soc. Ser. B (Methodological) 1951, 1, 1–38. [Google Scholar] [CrossRef]
- López-Trujillo, J.; Mellado-Bosque, M.; Ascacio-Valdés, J.A.; Prado-Barragán, L.A.; Hernández-Herrera, J.A.; Aguilera-Carbó, A.F. Temperature and pH optimization for protease production fermented by Yarrowia lipolytica from Agro-Industrial Waste. Fermentation 2023, 9, 819. [Google Scholar] [CrossRef]
- Ogrydziak, D. M. Yeast Extracellular Proteases. Crit. Rev. Biotechnol. 1993, 13, 1–55. [Google Scholar] [CrossRef]
- Nakamura, Y.; Fukuhara, H.; Sano, K. Secreted phytase activities of yeasts. Biosci. Biotechnol. Biochem. 2000, 64, 841–844. [Google Scholar] [CrossRef]
- Dakhmouche Djekrif, S.; Bennamoun, L.; Labbani, F.-Z.K.; Ait Kaki, A.; Nouadri, T.; Pauss, A.; Meraihi, Z.; Gillmann, L. An Alkalothermophilic Amylopullulanase from the Yeast Clavispora lusitaniae ABS7. Purification, Characterization and Potential Application in Laundry Detergent. Catalysts 2021, 11, 12. [Google Scholar] [CrossRef]
- Ranjan, K.; Ahmad, L.; Sahay, S.M. Detergent compatible cold-active alkaline amylases from Clavispora lusitaniae CB13. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 306–310. [Google Scholar] [CrossRef]
- Mohan Kumar, B.V.; Sarabhai, S.; Prabhasankar, P. Targeted degradation of gluten proteins in wheat flour by prolyl endoprotease and its utilization in low immunogenic pasta for gluten sensitivity population. J. Cereal Sci. 2019, 87, 59–67. [Google Scholar] [CrossRef]
- Shaikh, I. A.; Turakani, B.; Malpani, J.; Goudar, S. V.; Mahnashi, M. H.; Hamed Al-Serwi, R.; et al. Extracellular protease production, optimization, and partial purification from Bacillus nakamurai PL4 and its applications. J. King Saud. Univ. Sci. 2023, 35, 102429. [Google Scholar] [CrossRef]
- Svensson, S.E.; Bucuricova, L.; Ferreira, J.A.; Souza Filho, P.F.; Taherzadeh, M.J.; Zamani, A. Valorization of Bread Waste to a Fiber and Protein-Rich Fungal Biomass. Fermentation 2021, 7, 91. [Google Scholar] [CrossRef]
- Benkahoul, M.; Bramki, A.; Belmessikh, A.; Mechakra-Maza, A. Statistical optimization of an acid protease production by a local Aspergillus niger MH109542 using a medium based on decommissioned dates. Asia Pac. J. Mol. Biol. Biotechnol. 2020, 28, 68–76. [Google Scholar] [CrossRef]
- Djekrif-Dakhmouche, S.; Gheribi-Aoulmi, Z.; Meraihi, Z.; Bennamoun, L. Application of a statistical design to the optimization of culture medium for α-amylase production by Aspergillus niger ATCC 16404 grown on orange waste powder. J. Food Eng. 2006, 73, 190–197. [Google Scholar] [CrossRef]
- Shrestha, S.; Chio, C.; Khatiwada, J. R.; Kognou, A. L. M.; Qin, W. Optimization of multiple enzymes production by fermentation using lipid-producing Bacillus sp. Front. Microbiol. 2022, 13, 1049692. [Google Scholar] [CrossRef] [PubMed]
- Shafique, T.; Shafique, J.; Zahid, S.; Kazi, M.; Alnemer, O.; Ahmad, A. Screening, selection and development of Bacillus subtilis apr-IBL04 for hyper production of macromolecule alkaline protease. Saudi J. Biol. Sci. 2021, 28, 1949–1951. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O. H.; Rosebrough, N. J.; Farr, A.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Luoto, S.; Jiang, Z.; Brinck, O.; Sontag-Strohm, T.; Kanerva, P.; Bruins, M.; Edens, L.; Salovaara, H.; Loponen, J. Malt hydrolysates for gluten-free applications: autolytic and proline endopeptidase assisted removal of prolamins from wheat, barley and rye. J. Cereal Sci. 2012, 56, 504–509. [Google Scholar] [CrossRef]
- Sayaslan, A. Wet-milling of wheat flour: industrial processes and small scale test methods. LWT. Food Sci. Technol. 2004, 37, 499–515. [Google Scholar] [CrossRef]
- Lario, L. D.; Pillaca-Pullo, O. S.; Sette, L. D.; Converti, A.; Casati, P.; Spampinato, C.; Pessoa, A. Optimization of protease production and sequence analysis of the purifed enzyme from the cold adapted yeast Rhodotorula mucilaginosa CBMAI 1528. Biotechnol. Rep. 2020, 28, e00546. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. Y. Isolation of the Protease-producing Yeast Pichia anomala CO-1 and Characterization of Its Extracellular Neutral Protease. J. Life Sci. 2019, 29, 1126–1135. [Google Scholar] [CrossRef]
- Ray, M. K.; Uma Devi, K.; Seshu Kumar, G.; Shivaji, S. Extracellular Protease from the Antarctic Yeast Candida humicola. Appl. Environ. Microbiol. 1992, 58, 1918–1923. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Lopez, C. I.; Szabo, R.; Blanchin-Roland, S.; Gaillardin, C. Genetic control of extracellular protease synthesis in the yeast Yarrowia lipolytica. Genetics 2002, 160, 417–427. [Google Scholar] [CrossRef]
- Theron, L.W.; Divol, B. Microbial aspartic proteases: Current and potential applications in industry. Appl. Microbiol. Biotechnol 2014, 98, 8853–8858. [Google Scholar] [CrossRef]
- Schlander, M.; Distler, U.; Tenzer, S.; Thines, E.; Claus, H. Purification and Properties of Yeast Proteases Secreted by Wickerhamomyces anomalus 227 and Metschnikovia pulcherrima 446 during Growth in a White Grape Juice. Fermentation 2016, 3, 2. [Google Scholar] [CrossRef]
- Fernández, M.; Úbeda, J.F.; Briones, A.I. Typing of non-Saccharomyces yeasts with enzymatic activities of interest in wine-making. Int. J. Food. Microbiol. 2000, 59, 29–36. [Google Scholar] [CrossRef]
- Abdelal, A. T. H.; Kennedy, E. H.; Ahearn, G. Purification and characterization of a neutral protease from Saccharomycopsis lipolytica. J. Bacteriol. 1977, 130, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Ruchel, R.; Tegeler, R.; Trost, M. A comparison of secretory proteinases from different strains of Candida albicans. Sabouraudia 1982, 20, 233–244. [Google Scholar] [CrossRef] [PubMed]
- James, M.N. Catalytic pathway of aspartic peptidases. In Handbook of Proteolytic Enzymes, 2nd ed.; Beynon, R., Bond, J.S., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 12–18. [Google Scholar] [CrossRef]
- Tunga, R.; Shrivastava, B.; Banerjee, R. Purification and characterization of a protease from solid state cultures of Aspergillus parasiticus. Process Biochem 2003, 38, 1553–1558. [Google Scholar] [CrossRef]
- Wang, R.; Law, R.C.; Webb, C. Protease production and conidiation by Aspergillus oryzae in flour fermentation. Process Biochem. 2005, 40, 217–227. [Google Scholar] [CrossRef]
- Chaud, L. C. S.; Lario, L. D.; Bonugli-Santos, R. C.; Sette, L. D.; Pessoa Junior, A.; Felipe, M.; das Graças, A. Improvement in extracellular protease production by the marine antarctic yeast Rhodotorula mucilaginosa L7. N. Biotechnol. 2016, 33, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Barnett, J. A.; Payne, R. W.; Yarrow, D. Yeasts Characteristics and Identification; Cambridge University Press: London, 1983. [Google Scholar]
- Murao, S.; Kamada, M.; Nakase, T.; Ogura, S.; Oda, K. Studies on the Extracellular Protease of Yeasts Part I. J. agric. chem. Soc. Jpn. 1972, 46, 167–170. [Google Scholar] [CrossRef]
- Tennalli, G. B.; Garawadmath, S.; Sequeira, L.; Murudi, S.; Patil, V.; Divate, M. N.; Hungund, B. S. Media optimization for the production of alkaline protease by Bacillus cereus PW3A using response surface methodology. J. Appl. Biol. Biotechnol. 2022, 10, 17–26. [Google Scholar] [CrossRef]
- Ferracini-Santos, L.; Sato, H. H. Production of alkaline protease from Cellulosimicrobium cellulans. Braz. J. Microbiol. 2009, 40, 54–60. [Google Scholar] [CrossRef]
- Adetunji, A. I.; Olaniran, A. O. Statistical modelling and optimization of protease production by an autochthonous Bacillus aryabhattai Ab15-ES: A response surface methodology approach. Biocatal. Agric. Biotechnol. 2020, 24, 101528. [Google Scholar] [CrossRef]
- Moftah, O. A. S.; Grbavčić, S.; Žuža, M.; Luković, N.; Bezbradica, D.; Knežević-Jugović, Z. Adding Value to the Oil Cake as a Waste from Oil Processing Industry: Production of Lipase and Protease by Candida utilis in Solid State Fermentation. Appl. Biochem. Biotechnol. 2012, 166, 348–364. [Google Scholar] [CrossRef]
- Sindhu, R.; Suprabha, G. N.; Shashidhar, S. Optimization of process parameters for the production of alkaline protease from Penicillium godlewskii SBSS 25 and its application in detergent industry. Afr. J. Microbiol. Res. 2009, 3, 515–522. [Google Scholar]
- Benabda, O.; M’hir, S.; Kasmi, M.; Mnif, W.; Hamdi, M. Optimization of Protease and Amylase Production by Rhizopus oryzae Cultivated on Bread Waste Using Solid-State Fermentation. J. Chem. 2019, 3738181. [Google Scholar] [CrossRef]
- Agrawal, D.; Patidar, P.; Banerjee, T.; Patil, S. Alkaline protease production by a soil isolate of Beauveria feline under SSF condition: parameter optimization and application to soy protein hydrolysis. Process Biochem. 2005, 40, 1131–1136. [Google Scholar] [CrossRef]
- Elmansy, E. A.; Asker, M. S.; El-Kady, E. M.; Hassanein, S. M.; El-Beih, F. M. Production and optimization of α-amylase from thermo-halophilic bacteria isolated from different local marine environments. Bull. Natl. Res. Cent. 2018, 42, 31. [Google Scholar] [CrossRef]
- Takeyama, M. M.; de Carvalho, M. C.; Carvalho, H. S; et al. Pectinases Secretion by Saccharomyces cerevisiae: Optimization in Solid-State Fermentation and Identification by a Shotgun Proteomics Approach. Molecules 2022, 27, 4981. [Google Scholar] [CrossRef] [PubMed]
- Vishwanatha, K. S.; Appu Rao, A. G.; Singh S., A. Acid protease production by solid-state fermentation using Aspergillus oryzae MTCC 5341: optimization of process parameters. J. Ind. Microbiol. Biotechnol. 2010, 37, 129–138. [Google Scholar] [CrossRef]
- Arekemase, M.O.; Alfa, O.P.; Agbabiaka, T.O.; Ajide-Bamigboye, N.T.; Aderoboye, O.Y.; Orogu, J.O.; Ahmed, T. Optimization of amylase produced from bacteria isolated from cassava peel dumpsite using submerged fermentation. Sci. World J. 2020, 15, 64–75. [Google Scholar]
- Rozanov, A. S.; Shekhovtsov, S. V.; Bogacheva, N. V.; Pershina, E. G.; Ryapolova, A.V.; Bytyak, D. S.; Peltek, S. E. Production of subtilisin proteases in bacteria and yeast. Vavilovskii Zh. Genet 2021, 25, 125–134. [Google Scholar] [CrossRef]
- Usman, A.; Mohammed, S.; Mamo, J. Production, optimization, and characterization of an acid protease from a filamentous fungus by solid-state fermentation. Int. J. Microbiol. 2021, 6685963. [Google Scholar] [CrossRef]
- Chimbekujwo, K. I.; Ja’afaru, M. I.; Adeyemo, O. M. Purification, characterization and optimization conditions of protease produced by Aspergillus brasiliensis strain BCW2. Sci. Afr. 2020, 8, e00398. [Google Scholar] [CrossRef]
- Lario, L. D.; Chaud, L.; Almeida M., G.; Converti, A.; Sette L., D.; Pessoa Jr, A. Production, purification, and characterization of an extracellular acid protease from the marine Antarctic yeast Rhodotorula mucilaginosa L7. Fungal Biol. 2015, 119, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Mota de Oliveira, J.; Fernandes, P.; Benevides, R. G.; Aparecida de Assis, S. Production, characterization, and immobilization of protease from the yeast Rhodotorula oryzicola. Biotechnol. Appl. Biochem. 2021, 68, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Remold, H.; Fasold, H.; Staib, F. Purification and characterization of a proteolytic enzyme from Candida albicans. Biochim. Biophys. Acta 1968, 167, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Young, T. W. Extracellular acid and alkaline proteases from Candida olea. J. Gen. Microbiol. 1987, 133, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Ogrydziak, O.M. Extracellular acid proteases produced by Saccharomycopsis lipolytica. J. Bacteriol. 1983, 154, 23–31. [Google Scholar] [CrossRef]
- Kamada, M.; Oda, K.; Murao, S. The purification of the extracellular acid protease of Rhodotorula glutinis K-24 and its general properties. Agric. Biol. Chem. 1972, 36, 1095e1101. [Google Scholar] [CrossRef]
- Srividya, S. Production and characterization of an acid protease from a local Aspergillus sp. By solid substrate fermentation. Arch. Appl. Sci. Res. 2012, 1, 188–199. [Google Scholar]
- Wei, M.; Chen, P.; Zheng, P.; et al. Purification and characterization of aspartic protease from Aspergillus niger and its efficient hydrolysis applications in soy protein degradation. Microb. Cell Factories 2023, 22, 42. [Google Scholar] [CrossRef]
- Purushothaman, K.; Bhat, S. K.; Annapurna Singh, S.; Kedihithlu Marathe, G.; Appu Rao, G.; Appu, R. Aspartic protease from Aspergillus niger: Molecular characterization and interaction with pepstatin A, Int. J. Biol. Macromol. 2019, 139, 199–212. [Google Scholar] [CrossRef]
- Mohan Kumar, B.V.; Sarabhai, S.; Prabhasankar, P. Targeted degradation of gluten proteins in wheat flour by prolyl endoprotease and its utilization in low immunogenic pasta for gluten sensitivity population. J. Cereal Sci. 2019, 87, 59–67. [Google Scholar] [CrossRef]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial Proteases Applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef]
- Mamo, J.; Assefa, F. The role of microbial aspartic protease enzyme in food and beverage industries. J. Food Qual. 2018, 2018, 7957269. [Google Scholar] [CrossRef]
- Mandujano-Gonzalez, V.; Villa-Tanaca, L.; Anducho-Reyes, M. A.; Mercado-Flores, Y. Secreted fungal aspartic proteases: a review. Rev. Iberoam Micol. 2016, 33, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Zhang, Y. Z.; Gao, P. J.; Luan, X.W. Two different proteases produced by a deep-sea psychrophic bacterial strain, Pseudoalteromonas sp SM9913. Mar. Biol. 2003, 143, 989–993. [Google Scholar] [CrossRef]
- Kim, J.I.; Lee, S.M.; Jung, H.J. Characterization of calcium activated bifunctional peptidase of the psychrotrophic Bacillus cereus. J. Microbiol. 2005, 43, 237–243. [Google Scholar] [PubMed]
- Vazquez, S.; Ruberto, L.; Mac Cormack, W. Properties of extracellular proteases from three psychrotolerant Stenotrophomonas maltophilia isolated from Antarctic soil. Polar Biol. 2005, 28, 319–325. [Google Scholar] [CrossRef]
- Aguilar, dos S. JG; Sato, H. H. Microbial proteases: Production and application in obtaining protein hydrolysates. Food Res. Int. 2018, 103, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, W. C. A.; Martins, M. L. L. Production and properties of an extracellular protease from thermophilic Bacillus sp. Braz. J. Microbiol. 2004, 35, 91–96. [Google Scholar] [CrossRef]
- Gaur, S.; Agrahari, S.; Wadhwa, N. Purification of Protease from Pseudomonas thermaerum GW1 Isolated from Poultry Waste Site. Open Microbiol. J. 2010, 4, 67–74. [Google Scholar] [CrossRef]
- Devi, M. K.; Banu, A. R.; Gnanaprabhal, G. R.; Pradeep, B.V.; Palaniswamy, M. Purification, characterization of alkaline protease enzyme from native isolate Aspergillus niger and its compatibility with commercial detergents. Indian J. Sci. Technol. 2008, 1, 1–6. [Google Scholar] [CrossRef]
- Sharma, N.; Bhatia, S.; Chunduri, V.; Kaur, S.; Sharma, S.; Kapoor, P.; Kumari, A.; Garg, M. Pathogenesis of celiac disease and other gluten related disorders in wheat and strategies for mitigating them. Front. Nutr. 2020, 7, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Walter, T.; Wieser, H.; Koehler, P. Degradation of gluten in wheat bran and bread drink by means of a proline-specific peptidase. J. Nutr. Food Sci. 2014, 4, 2–15. [Google Scholar] [CrossRef]
- Heredia-Sandoval, N.G.; de la Barca, A.C.; Carvajal-Millán, E.; Islas-Rubio, A.R. Amaranth addition to enzymatically modified wheat flour improves dough functionality, bread immunoreactivity and quality. Food Funct 2018, 9, 534–540. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Production of Clavispora lusitaniae protease on different solid media in SmF.
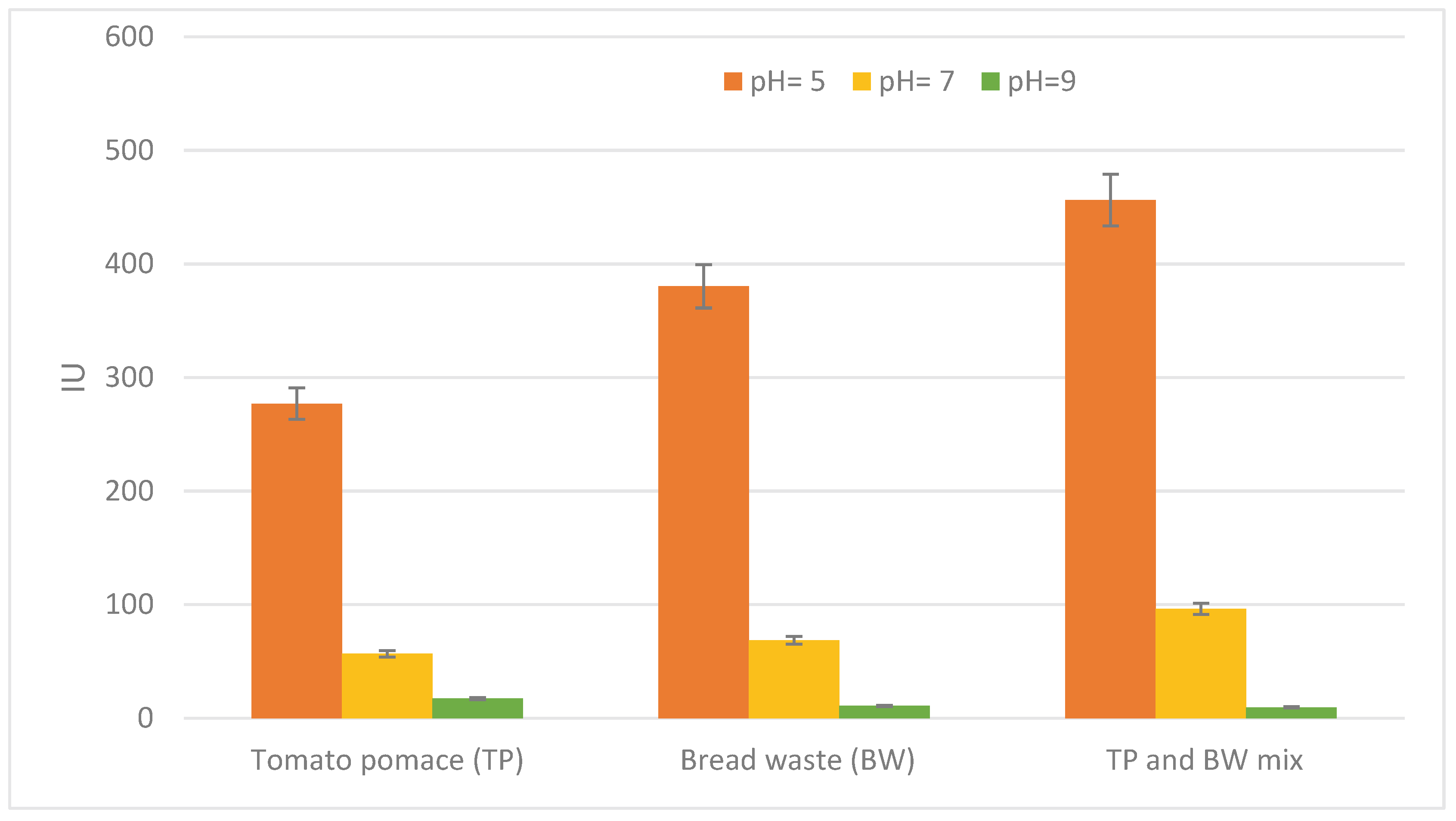
Figure 2.
Production of Clavispora lusitaniae PC 3 protease on different solid media in SSF.
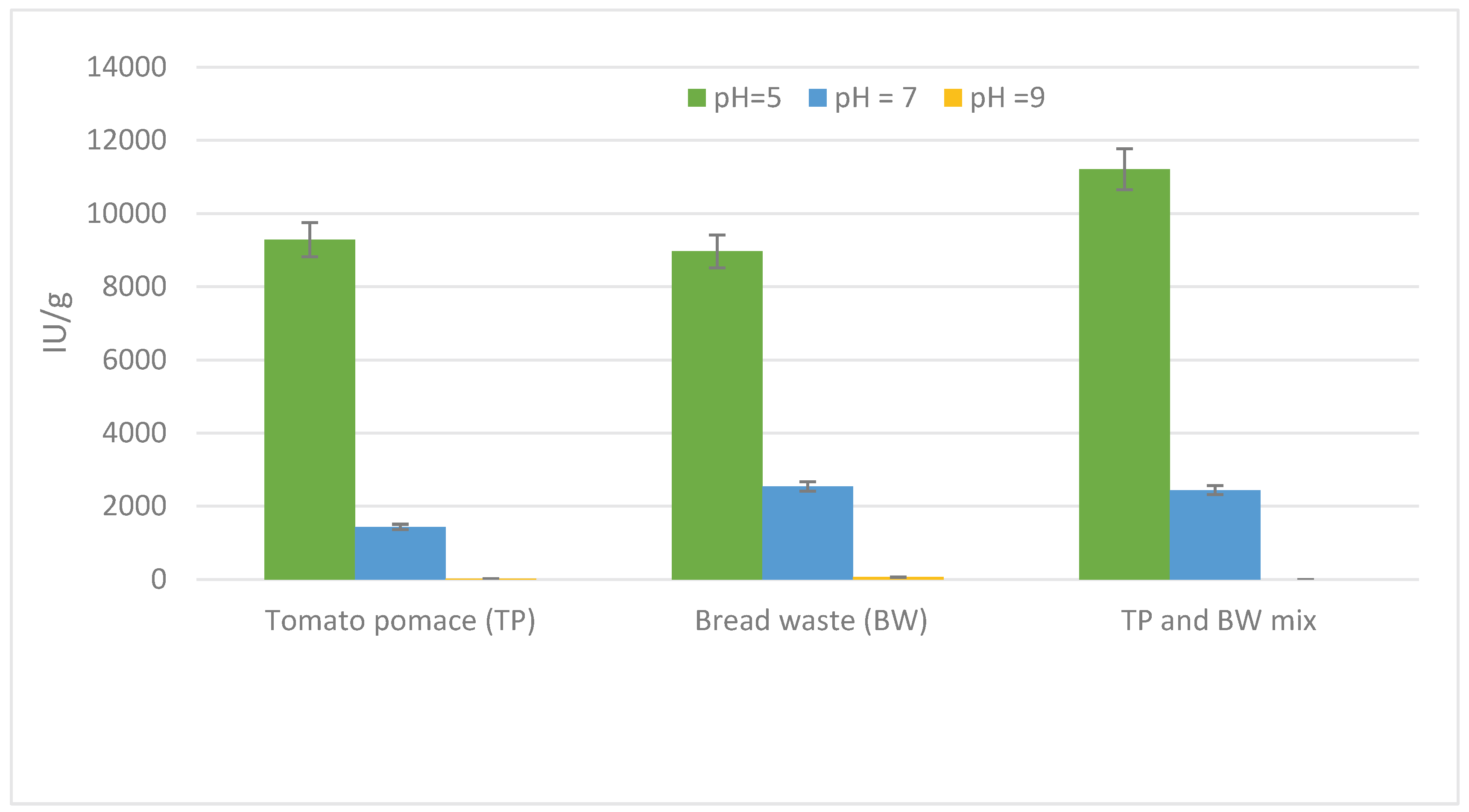
Figure 3.
Response surface graphs showing (A) Moisture vs inoculum; (B) Inoculum vs fermentation time; (C) Moisture vs Fermentation time.
Figure 3.
Response surface graphs showing (A) Moisture vs inoculum; (B) Inoculum vs fermentation time; (C) Moisture vs Fermentation time.
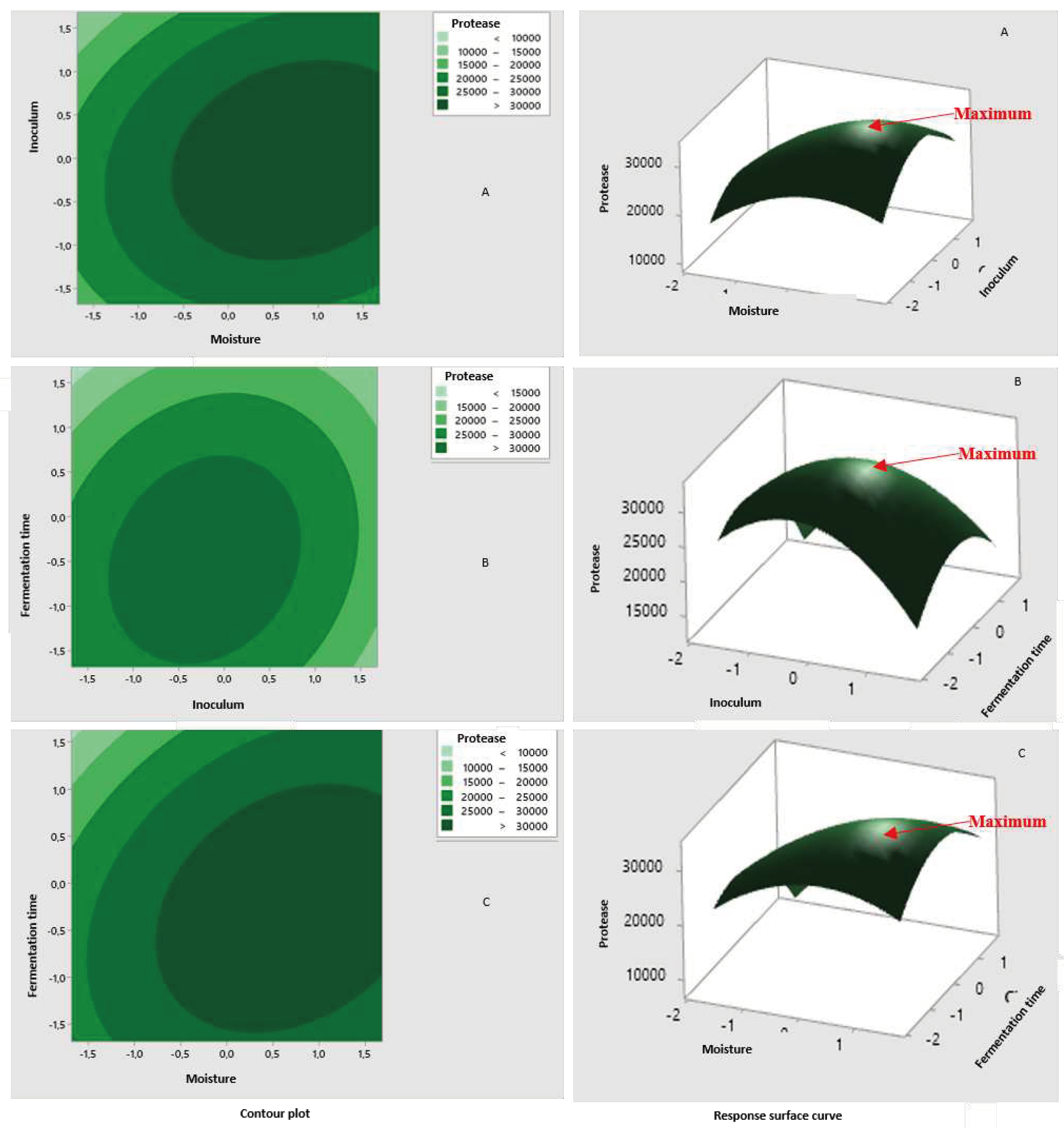
Figure 4.
Multiple response optimization of protease production in Clavispora lusitaniae PC3.
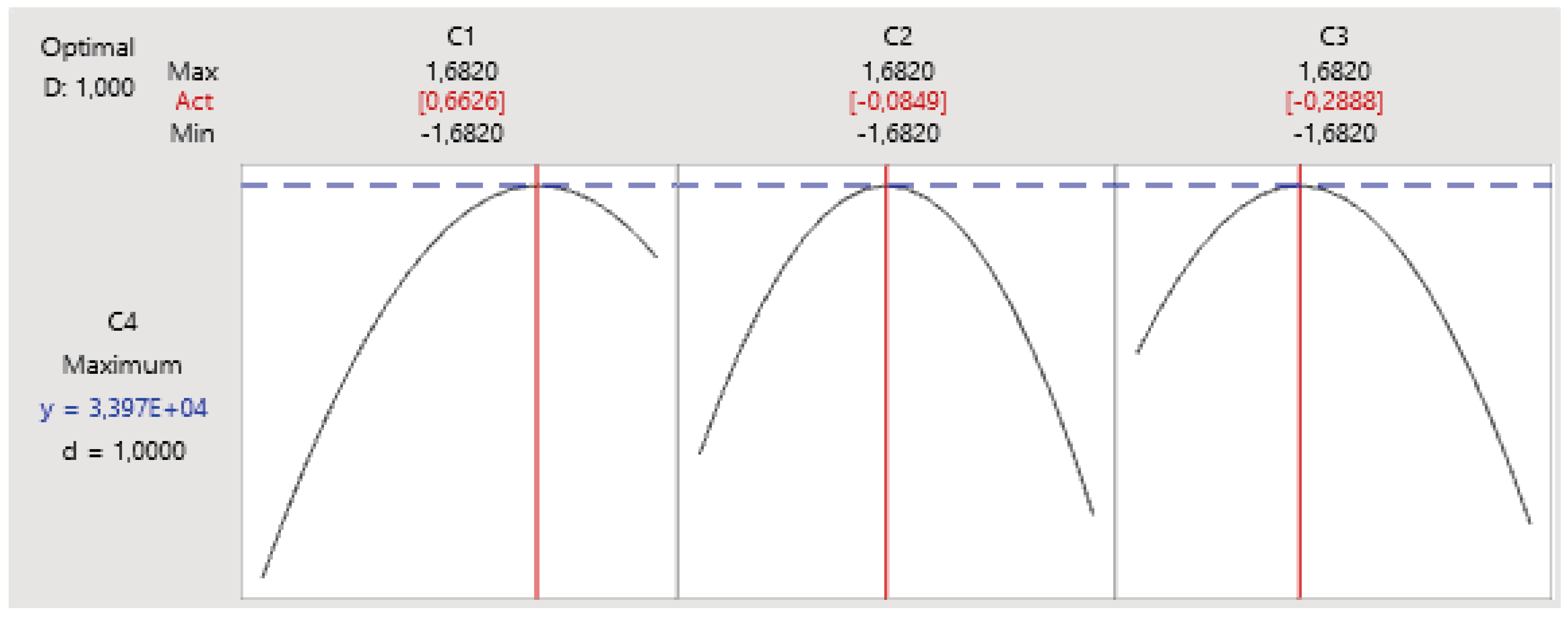
Table 1.
Relationship between the coded levels and the real levels of the factors studied in the design of Plackett and Burman.
Table 1.
Relationship between the coded levels and the real levels of the factors studied in the design of Plackett and Burman.
| Factors | Level | Unit | |
|---|---|---|---|
| -1 | +1 | ||
| A:Temperature | 30 | 50 | °C |
| B:Moisture | 50 | 80 | % |
| C:Inoculum | 104 | 108 | Cells/g |
| D:Error | - | - | - |
| E:Glucose | 0 | 0.5 | % |
| F:Corn Steep Liquor | 0 | 0.5 | % |
| G:(NH4)2SO4 | 0 | 0.5 | % |
| H:Error | - | - | - |
| I:NaCl | 0 | 0.1 | g/l |
| J:Time fermentation | 24 | 72 | H |
| K:Error | - | - | - |
Table 2.
Actual and coded values of experimental variables used CCD.
| -1.682 | -1 | 0 | +1 | +1.682 | |
|---|---|---|---|---|---|
| B | 53,18 | 60 | 70 | 80 | 86,82 |
| C | 2,08 x 105 | 106 | 107 | 108 | 0,5 x109 |
| J | 39.81 | 48 | 60 | 72 | 80.18 |
| B: Moisture (%), | C: Inoculum (Cells/ml), | J: Fermentation time (H) | |||
Table 4.
Plackett–Burman design for the study of 11 factors with 12 experiments.
| Experiments | A | B | C | (D) | E | F | G | (H) | I | J | (K) | Protease Activity (IU/g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | +1 | +1 | -1 | +1 | +1 | +1 | -1 | -1 | -1 | +1 | -1 | 18305,3 |
| 2 | -1 | +1 | +1 | -1 | +1 | +1 | +1 | -1 | -1 | -1 | +1 | 6567,55 |
| 3 | +1 | -1 | +1 | +1 | -1 | +1 | +1 | +1 | -1 | -1 | -1 | 13621,2 |
| 4 | -1 | +1 | -1 | +1 | +1 | -1 | +1 | +1 | +1 | -1 | -1 | 9917,55 |
| 5 | -1 | -1 | +1 | -1 | +1 | +1 | -1 | +1 | +1 | +1 | -1 | 15332,15 |
| 6 | -1 | -1 | -1 | +1 | -1 | +1 | +1 | -1 | +1 | +1 | +1 | 25011,8 |
| 7 | +1 | -1 | -1 | -1 | +1 | -1 | +1 | +1 | -1 | +1 | +1 | 30662,8 |
| 8 | +1 | +1 | -1 | -1 | -1 | -1 | +1 | -1 | +1 | -1 | +1 | 12932,05 |
| 9 | +1 | +1 | +1 | -1 | -1 | -1 | +1 | -1 | +1 | +1 | -1 | 21830,3 |
| 10 | -1 | +1 | +1 | +1 | -1 | -1 | -1 | +1 | -1 | +1 | +1 | 15173,05 |
| 11 | +1 | -1 | +1 | +1 | +1 | -1 | -1 | -1 | +1 | -1 | +1 | 8991,2 |
| 12 | -1 | -1 | -1 | -1 | -1 | -1 | -1 | -1 | -1 | -1 | -1 | 14138,3 |
| A, B, C, E, F, G, I and J are real variables | ||||||||||||
| D, H and K are dummy variables | ||||||||||||
Table 5.
Statistical analysis of the Plackett-Burman design (PBD).
| Variables | Effect | Coefficient | t-Value | P-Value | Confidence Level % |
|---|---|---|---|---|---|
| Constant | - | 16040 | 26.76 | 0 | - |
| A : Temperature | 3367 | 1684 | 2.81 | 0.067 | 93.3 |
| B : Humidity | -3839 | -1919 | -3.2 | 0.049* | 95.1* |
| C : Inoculum | -4909 | -2454 | -4.09 | 0.026* | 97.4* |
| E : Glucose | -2155 | -1078 | -1.8 | 0.17 | 83 |
| F: Corn steep liquor | -1491 | -745 | -1.24 | 0.302 | 69.8 |
| G : (NH4)2SO4 | 3790 | 1895 | 3.16 | 0.051 | 94.9 |
| I : NaCl | -742 | -371 | -0.62 | 0.058 | 94.2 |
| J : Incubation period | 10025 | 5012 | 8.36 | 0.004* | 99.6* |
Coef.: coefficient; t-value: Student’s test value; p-value: probability value;
* Significant effect at p < 0.05;.
Table 6.
ANOVA of PBD for Clavispora lusitaniae protease production.
| Source | DF | Sum of Squares | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 8 | 517319398 | 64664925 | 15,00 | 0.024 |
| Error | 3 | 12935581 | 431477314 | ||
| Total | 11 | 530254980 | |||
|
R2 = 0.9756; Adjusted-R2 = 0.9106. | |||||
Table 7.
CCD for optimization of protease production by Clavispora lusitaniae PC3.
| Run | B: Moisture | C: Inoculum | J: Fermentation Time | Protease Activity (IU/g) |
|---|---|---|---|---|
| 1 | 1,682 | 0 | 0 | 31856,3 |
| 2 | 0 | 0 | 0 | 33831,6 |
| 3 | 0 | 0 | 1,682 | 21550,6 |
| 4 | 1 | 1 | -1 | 28422,3 |
| 5 | 1 | -1 | 1 | 26156,6 |
| 6 | 0 | 0 | 0 | 31879,4 |
| 7 | 1 | -1 | -1 | 30514,1 |
| 8 | 0 | 0 | 0 | 32626,5 |
| 9 | -1 | -1 | 1 | 19636,1 |
| 10 | -1 | -1 | -1 | 27337,0 |
| 11 | 0 | 0 | 1 | 29101,4 |
| 12 | -1 | 1 | 0 | 22548,0 |
| 13 | 0 | 0 | 0 | 32492,0 |
| 14 | 0 | -1,682 | 0 | 25896,2 |
| 15 | 0 | 1,682 | 0 | 23030,7 |
| 16 | -1,682 | 0 | -1 | 24251,0 |
| 17 | -1 | 1 | 0 | 22731,3 |
| 18 | 0 | 0 | -1 | 31088,8 |
| 19 | 1 | 1 | 1 | 28721,6 |
| 20 | 0 | 0 | -1,682 | 29864,7 |
Table 9.
Purification of acid protease from Clavispora lusitaniae PC3.
| Sample | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Crude extract | 31589.2 | 168 | 188.03 | 1 | 100 |
| Ammonium sulfate (60%) | 29276.1 | 53 | 552.4 | 2.938 | 92.67 |
| Dialysis | 25821.5 | 28 | 922.2 | 4.904 | 81.74 |
| Sephadex G100 | 10406 | 2.8 | 3716.4 | 19.76 | 32.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



