Submitted:
03 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Leukemia encompasses diverse blood cancers marked by the unchecked growth and developmental blockage of myeloid or lymphoid precursor cells within the bone marrow and the bloodstream. Treatment traditionally involves intensive chemotherapy or hematopoietic stem cell transplantation, albeit these approaches often yield severe side effects and encounter resistance from leukemia cells. Hence, the quest for novel therapies targeting leukemic cells selectively is paramount to enhancing leukemia outcomes. Exploring natural products is a promising avenue for pursuing effective chemotherapy and preventive measures against leukemia. These compounds serve as a crucial wellspring for drug development and offer a source of biologically active elements for potential therapeutic interventions in leukemia. The enzyme telomerase plays a vital role in stabilizing chromosomes by adding DNA sections to the end of chromosomes during the mitotic cycle. Notably, hTERT mRNA expression strongly correlates with telomerase activity, making it a potential target for therapeutic intervention without adverse systemic effects. Telomerase inhibition is a promising target in cancer therapy, especially in combination with natural ingredients, demonstrating less cytotoxicity than chemotherapies.
Keywords:
leukemia
; natural substances
; telomerase
; antioxidant
; hematological malignancy
; genomic instability
; DNA damage
; polyphenols
; vitamins
; in vitro
; in vivo and clinical trials
1. Introduction
Telomerase is a chromosome stabilizing enzyme, active at the end of a mitotic cycle – its main purpose is to add the missing section of the DNA on the delayed strand. It works on telomeres located at the 3′-ends of eukaryotic chromosomes, whose role is to protect the DNA against possible damage during copying, as shown in Figure 1. In somatic cells it is impossible to completely replicate them, resulting in their shortening during every cell division [1]. Telomerase is a large complex composed of RNA and proteins. It creates multiple copies of a repetitive sequence, called the telomeric repeat, which is rich in guanine, using telomerase reverse transcriptase (in humans called hTERT) and its telomerase RNA (TER), which includes the template that coordinates repeat synthesis (hTERC – human telomerase RNA component) [2]. These repeats are combined by the shelterin complex, composed of six proteins: protection of telomeres 1 (POT1), telomere repeat factor 1 and 2 (TRF1, TRF2), tripeptidyl peptidase I (TPP1), TRF1-interacting nuclear protein 2 (TIN2), and repressor/activator protein 1 (RAP1) [3].
In most somatic cells, telomerase activity is not found, which makes their prolific ability limited. Enzyme expression was found in germ cells and self-renewing tissues, such as the intestinal epithelium, the ovary, and hematopoietic stem cells [5]. A significant activity of telomerase is expressed in approximately 90% of cancer cells and tumor tissues, which leads to suspect the correlation between the addition of telomeric DNA and the infinite proliferation of cancer cells [6]. Further research into the mechanism underlying its regulation can be beneficial and used for better cancer diagnosis and treatment.
The extent of hTERT mRNA expression strongly corresponds to cellular telomerase activity. This indicates, that this specific subunit is a key determinant of telomerase engagement in replication and may be the best option to choose while attacking this enzyme [5]. Understanding this protein allows the creation of more precise therapies that do not affect negatively the whole body. TERT function varies and may influence the process of elongation due to the creation of structural variants and genetic or epigenetic alterations. Moreover, it also affects the nucleus, cytoplasm, and mitochondrion, causing DNA-damage response, stress protection, gene expression regulation, and a decrease in apoptosis [4]. TERT and TERC are accompanied by auxiliary proteins, that are necessary for telomere homeostasis, which is essential in the regulation of aging and cancer formation such as structure stabilizing protein named dyskerin (DKC1) and non-histone chromosome protein 2 (NHP2). Other proteins also found are telomerase Cajal body protein 1 (TCAB1), heat shock protein 90 (HSP90), serine, and arginine-rich splicing factor 11 (SRSF11) [7].
More than 80% of tumors use diverse regulatory techniques, known as telomere maintenance mechanisms (TMMs) because they sustain telomere length by reactivating telomerase. TMMs are unique for tissue histotype, cancer type, and cell lines. The most important TTMs are TERT and TERC gene amplification, TERT gene rearrangements, TERTp somatic mutations, and transcription factor binding. Also exists epigenetic alterations, alternative splicing, and polymorphic variants within the TERT gene body and TERTp. 10-15% of cancer cells achieve immortality utilizing a telomerase-independent mechanism, called alternative lengthening of telomeres (ALT) [8]. The lack of telomerase activity results in the remodeling of the catalytic proteins by chromatinization or methylation [9]. The remaining percentage of tumors proliferates based on a non-defined telomere maintenance mechanism (NDTMM), in which both TERT expression and ALT are absent [4].
Telomerase inhibitors may be divided into two categories: natural or synthetic. The main substances which draw the attention of the researchers are: in the case of natural inhibitors - curcumin and indole-3-carbinol, and when it comes to synthetic ones - BIBR1532 and geldanamycin. In this study, we will discuss natural inhibitors of telomerase, which are included in Figure 2 and Table 1.
The inhibition process varies depending on the substance – it may bind directly to the enzyme and downregulate its activity just like oxoisoaporphine [10] or it activates intracellular pathways affecting telomerase, which happens in the instance of genistein [11]. We have remarked that each substance’s inhibitory ability is different.
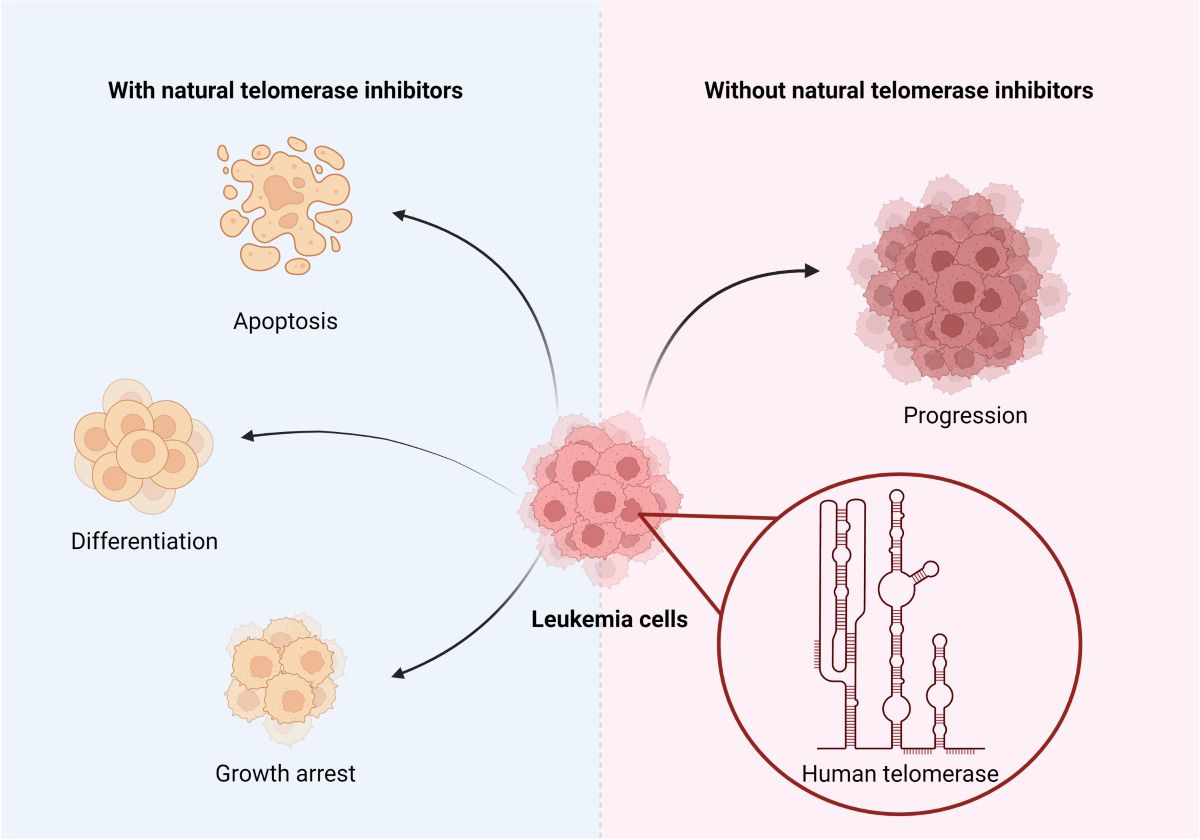
Telomerase inhibition is a promising target in cancer therapy, especially in combination with natural ingredients demonstrating less cytotoxicity than chemotherapies. That is why we would like to review the inhibitory properties of telomerase in leukemias natural substances, as it is displayed in Figure 3.
2. Materials and Methods.
Telomerase inhibition is an interesting prospect, which has been inspiring numerous scientists to the usage of various substances. Electronic searches were conducted to find them. After searching on PubMed.gov keywords ‘telomerase‘, ‘inhibition’, ‘leukemia’, and ‘natural substances’ there weren’t any articles found. The search was then changed, and the keywords were ‘leukemia’ and ‘telomerase’. This procedure allowed us to find 822 results, as shown in Figure 4. The data was narrowed down to articles focused solely on natural inhibitors. Through some articles, we found even more substances, which are mentioned in the paragraphs below.
3. Natural substances inhibiting telomerase
3.1. Polyphenols
Polyphenols are the substances found in fruits and vegetables. Over the past years, researchers around the world studied their beneficial properties, currently especially concerning cancers. Their anti-inflammatory, anti-carcinogenic and antioxidant properties have been proven in numerous studies [12]. In our review, we wanted to draw attention to their telomerase inhibition property in leukemias.
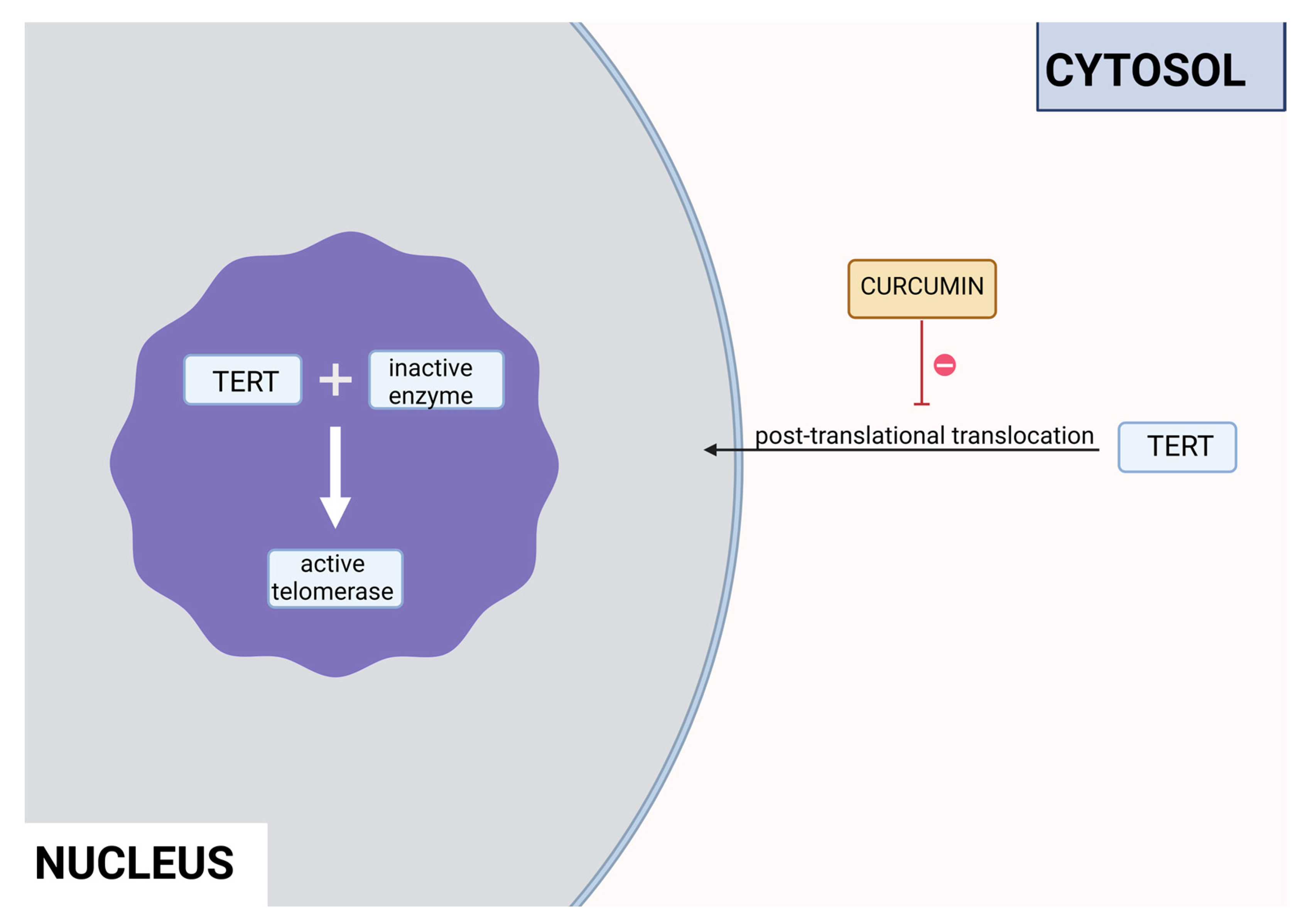
Curcumin is a polyphenol produced by plants from the Curcuma longa species and may be found in turmeric. It has many beneficial properties for the human organism such as anti-inflammatory, anti-carcinogenic, anti-mutagenic and anti-proliferative. In the study performed by S. Mukherjee et al. on acute myeloblastic leukemia HL-60 cell line inhibition of telomerase was observed. Different concentrations of curcumin were applied to the cells that were incubated for 24 hours. After this time, telomerase activity was measured using TRAP assay. It has been noticed that inhibition of telomerase activity increases with the dose of curcumin – 55% and 78% using 10μM and 55μM, respectively. The IC50 value was estimated at 9,8μM. It was concluded that there is a linear relationship between inhibition of telomerase activity and induction of apoptosis, as curcumin also induced cytochrome c release from mitochondria to cytosol, which is an indicator of apoptosis, as well as Bcl-2 expression decreased, whereas Bax level increased [13]. Another study also indicated the induction of apoptosis by curcumin in cells from the HL- 60 cell line. M. Dikmen et al. have established that apoptosis is at a significant level using 15μM of curcumin (81,1% of late apoptotic cells) and higher concentrations are more effective – at 40μM 88,6% of late apoptotic cells. Higher caspase 3 activity was also observed [14]. In the human chronic myelogenic leukemia cells from the K-562 cell line, curcumin-induced telomerase inhibition was also examined by S. Chakraborty et al. Different concentrations of curcumin were applied and telomerase activity was measured after 48 hours by TRAP assay. It was found that inhibition grows in a dose and time dependant manner. After 48 hours inhibition was at 45% and 83,5% in the cells with 10μM and 50μM of curcumin, respectively. Whereas, after 24 hours with 50μM concentration, the inhibition was at 66%. The correlation of telomerase inhibition with the proportion of apoptotic cells, DNA content in the sub-G1 peak, caspase 3 and caspase 8 activations. Furthermore, the TERT subunit was detected by Western blot. The inhibition of post-translational translocation of the subunit from the cytosol to the nucleus was observed (the process presented in Figure 4). The expression of the cytosolic fraction of TERT was more pronounced – 73,8% with the presence of curcumin, whereas without curcumin it was at 43,2% [15].
Figure 5.
Inhibition of the post-translational translocation of the TERT cytosolic fraction in the presence of curcumin.
Figure 5.
Inhibition of the post-translational translocation of the TERT cytosolic fraction in the presence of curcumin.
Epigallocatechin-3-gallate (EGCG) is the main polyphenol present in green tea. In the study on the HL-60 cell line, no hTERT gene down-regulation was observed, however, apoptosis did occur due to other pathways – perhaps due to higher Bax-Bcl2 ratio or ROS production [16]. In another study, on ALL Jurkat E6.1 cells, apoptosis was also present. Concentrations of EGCG that were used (50μM, 70μM, 100μM) induced the apoptosis of 31%, 40%, and 71% of cells, respectively [17]. However, a chemically modified derivative of EGCG (MST-312) has been found effective as a telomerase inhibitor. With the combination of MST-312 and doxorubicin – a chemotherapy medication used to treat ALL, down-regulation of the hTERT gene was observed. Two pre-B ALL cell lines were put under investigation – NALM-6 and REH. Dosages effective towards the NALM-6 cell line were 2μM of MST-312 and 20nM of doxorubicin, whereas towards the REH line – 4μM of MST-312 and 5nM of doxorubicin [18].
Indole-3-carbinol (I3C) is a polyphenol responsible for the indirect inhibition of telomerase activity. It may be found in cruciferous vegetables: broccoli, cauliflower, Brussels sprouts, cabbage, or kale. In the study performed on the NALM-6 cell line, 60μM of I3C induced levels of the wild-type p53 after 24 and 48 hours. p53 is a well-known telomerase inhibitor, so qRT-PCR confirmed down-regulation of the hTERT gene. That process is crucial for the p53-dependant apoptosis in the leukemic cells [19]. In another study, influence of I3C on the chronic myeloid leukemia cells from the K562 cell line was examined. Caspase activation triggered apoptosis of the cells. Meanwhile, I3C caused down-regulation of c-Myc – a transcription factor that regulates the hTERT gene transcription. That process also inhibited the activity of telomerase. The most effective concentration of I3C was 400μM of the substance after 32 hours, it has to be noted that 200μM had no effect on c-Myc. Expression of c-Myc was measured after 18 and 32 hours, however 32 hours of incubation proved to be more efficient [20].
Butein (3, 4, 2′, 4′- tetrahydroxychalcone) is a polyphenolic substance found in the stem bark of cashews, the heartwood of Dalbergia odonifea, Rhus verniciflua and Caragena jubata. It is believed to have anti-tumour activity. The study performed on the U937, THP-1, HL60 and K562 cell lines not only had pro-apoptotic properties but also inhibited telomerase activity. It was observed that 20 μM of butein down-regulated the hTERT gene by 4 times in the THP-1 cell line after 24 hours. The process was not cell line specific – down-regulation of telomerase activity was caused by suppression of the hTERT gene on the transcriptional level, in a p53-independent manner. In the same study, it was observed that butein inactivates Akt, an up-regulator of telomerase activity, through phosphorylation of hTERT. Levels of p-Akt were decreased in a dose-dependent manner after 24 hours [21].
Gossypol is a polyphenolic aldehyde present in seeds, roots and stems of cotton plants. It has been proven that gossypol induces apoptosis in human prostate cancer. In a study performed on the HL60 cells the IC50 level was established – 4,5 μM. The activity of hTERT mRNA decreased in a time-dependent manner. Expression ratios were 87.06 and 59.12 at 24 and 96 hours respectively [22]. Another study examined the influence of gossypol on U937, Hl60, THP-1 and K562 cells. The activity of telomerase decreased in a dose-dependent manner in all four cell lines. However, 5 μM had no significant impact on HL60 and K562 cells. Whereas, 20 μM suppressed the activity of the enzyme by approximately by 30% in all cell lines, which indicates that the process is not cell-type specific. hTERT mRNA and protein expression levels were also checked in the U937 cells and they were lower due to down-regulation of the hTERT expression. Next, it was observed that gossypol down-regulates c-Myc dependant hTERT expression. Doses larger than 10 μM triggered reduction of c-Myc protein levels in U937 cells. This indicates that gossypol suppresses c-Myc binding on the regulatory regions of hTERT and at the same time attenuates hTERT gene expression. It was also studied that gossypol inactivates Akt, which is responsible for phosphorylation of hTERT, in U937 cells. After adding gossypol p-Akt decrease was noticed in a dose-dependent manner – the most effective being at 20 μM. At the same time, the process of hTERT’s phosphorylation was decreased. Phosphorylation is crucial for the nuclear translocation of hTERT [23].
3.2. Vitamin A
Vitamin A is a vital nutrient that serves several purposes in the human body [24]. It is crucial in embryo development and growth, the formation of organs, immune function, vision, and reproduction [25]. The substances found in food, such as carotenoids, retinol, and retinyl esters undergo sufficient changes in the human body to be transformed into active metabolites, as displayed in Figure 6. Retinoic acid, especially all-trans retinoic acid (ATRA), exhibits anticancer effects by inhibiting telomerase, reducing cellular proliferation, and activating apoptosis, differentiation, and cell growth arrest [26].
Retinoic acids are essential in myeloid differentiation. They regulate the target gene expression through their agonistic nuclear receptors: the retinoic acid receptor (RAR), the retinoid X receptor (RXR), and their isoforms [28]. Retinol is undergoing various changes inside the human body, as shown in Figure 7. Retinol is carried through the organism bound to transthyretin (TTR) and retinol-binding protein (RBP) or RBP alone. After being carried to the cell by transthyretin (TTR) or retinol-binding protein (RBP), retinol crosses the cell membrane and bonds to cellular retinol-binding protein (CRBP). Later, it is oxidized to the retinal by retinol dehydrogenase (RDH), cytosolic alcohol dehydrogenase (ADH), or short-chain dehydrogenase/reductases (SDRs). After that, it is transformed by the aldehyde dehydrogenase (ALDH) enzyme family members into trans-retinoic acid [27].
ATRA either attaches to CRABPs and is transported into the nucleus or is destroyed by ATRA-degrading cytochrome P450 reductases such as CYP26A1, which turns it into inactive metabolites. In the first case, it acts as a ligand and binds to a heterodimer of RXR and RAR. In the regulatory region of target genes, the RAR/RXR heterodimer binds to retinoic acid response elements (RAREs), which results in a conformational change by triggering the release of co-repressors and the engagement of co-activator complexes [27].
Studies using the HL-60 cell line demonstrate that ATRA’s telomerase inhibition appears quite early after its treatment and it is involved in early differentiation. It is thanks to the abrupt reduction of hTERT gene transcription. A marker for granulocytic differentiation of HL-60 cells was discovered due to the observation of the expression of integrin α-M protein (CD11b). Through that, it was found, that down-regulation of telomerase precedes the differentiation of HL-60 cells. These findings imply that ATRA’s antileukemic action may be mediated by its capacity to drive the differentiation of promyelocytic leukemia cells through the down-regulation of the hTERT gene. That gene seems to be the main target of ATRA’s control of cellular differentiation [27]. Apart from ATRA, other substances also influence retinoic acid receptors - the novel retinoids, such as 9cUAB30, induce apoptosis and inhibit telomerase by down-regulating the hTERT gene in HL60 cells [29]. It was also discovered, that recombinant human insulin-like growth factor binding protein 7 (rhIGFBP7) triggers ATRA-driven eradication of leukemia stem/progenitor cells in acute myeloid leukemia (AML) with elevated retinoic acid receptor α gene (RARA) expression. It also decreases growth factor independent 1 transcriptional repressor protein (GFI1) expression, which increases cells’ susceptibility to ATRA. It seems to be a prospective treatment for people with various types of AML [30].
In acute promyelocytic leukemia (APL) occurs chromosomal translocation, which causes the rearrangement of retinoic acid receptor α [28]. It involves promyelocytic leukemia genes and the RARA gene [27]. As a result, myeloid differentiation is stopped at the promyelocytic stage due to the disruption of the RA signal pathway. The pharmacological dosage of ATRA binds to the RARA directly and affects promyelocytic leukemia/retinoic acid receptor α (PML-RARα). It also influences communication with the nuclear receptor–corepressor complex, restoring the wild-type RARα/RXR regulatory pathway. Moreover, it promotes downstream genes’ transcriptional expression [28]. The ATRA’s influence on the DNA is displayed in Figure 8.
When the DNA mutation occurs (1.), the promyelocytic leukemia protein (PML) nuclear bodies are created. While untreated (2a.), it leads to the development of APL. The treatment may consist of a low (2b.) or high (2c.) dose of ATRA, which triggers different outcomes in patients [31].
This mechanism shows its effectiveness with numbers–95% of patients experience complete remission with just ATRA and chemotherapy [27]. The standard treatment schedule consists of an induction period, in which the dosage is 2 times 45mg/m2/day of ATRA and 0,15 mg/kg/day of arsenic trioxide (ATO). It is followed by 4 periods of consolidation, in which ATRA and ATO are also implemented. Patients with low/intermediate risk are treated with a combination of ATRA and ATO or, if ATO is unavailable, ATRA + chemotherapy. For high-risk patients, the treatment is the same, but with added gemtuzumab ozogamicin (GO) or idarubicin (IDA). Although the ATO’s mechanism of action is still not fully known, studies indicate a major synergism between ATO and ATRA in cell differentiation and apoptosis in promyelocytic leukemia [32]. In pediatric APL the standard treatment has ATRA in every stage [33] and the studies demonstrate that ATO may soon be included in the therapy as well [34]. The lack of retinoids prolongs chronic lymphocytic leukemia, and it is suggested to include a retinoic acid-rich diet in the treatment [35].
3.3. Vitamin D
Calcitriol is an active form of vitamin D, that can be synthesized in the human body through a series of changes, as shown in Figure 9. It presents various anti-cancer effects, such as growth inhibition, apoptosis activation, and differentiation of several kinds of malignant cells [36]. Moreover, it sensitizes cancer cells to various treatments and lowers pro-inflammatory cytokine production [37]. Apart from its multiple anticancer properties, it also regulates calcium and phosphate homeostasis, participates in bone mineralization, insulin secretion, and blood pressure regulation [38]. Unfortunately, its application in therapy is limited due to its calcemic properties. This was the reason for the creation of vitamin D3 analogs, such as EB 1089 (1,25 dihydroxy-24,26,27-trishomo-22E,24E-diene-cholecalciferol) - they display improved anti-tumor activity while lowering calcemic complications. They also lower their binding affinity to the vitamin D binding protein (DBP), which results in their easier cellular intake [36].
Vitamin D can be gained by supplementation of specific pills, dietary intake, and exposure to ultraviolet radiation (UVR). UVR mobilizes the conversion of 7-dehydrocholesterol to pre-vitamin D3, which is then converted to cholecalciferol. It’s carried through the bloodstream to the liver via DBP. The next form, calcifediol (25-hydroxyvitamin D) is produced thanks to cytochrome P450 2R1 (CYP2R1) hydroxylation. This form circulates in the blood and when delivered to the kidney, the cytochrome P450 27B1 (CYP27B1) transforms it to calcitriol (1,25- hydroxyvitamin D) [39]. This, now active, form affects a variety of tissues and biological pathways. Calcitriol binds the vitamin D receptor (VDR) and causes a conformational shift in the protein, which allows interaction with the RXR. The VDR-RXR heterodimer then works as a transcription factor by binding vitamin D response elements (VDRE) in DNA, which triggers target gene expression or inhibition [40]. In the case of hTERT, suppression is activated, which causes a lower telomerase level [41].
Cellular studies show that calcitriol regulates both normal and leukemic cell maturation. Configurations of vitamin D derivatives and retinoids display cooperative effects on differentiation in established leukemia cell lines, such as HL-60 and U937. Through ATRA-triggered pathways, vitamin D induces apoptosis in HL-60 cells and influences the expression of apoptosis-related gene products. It also impacts the generation of endogenous nitric oxide by elevating tumor necrosis factor-α protein (TNF-α) level or by the means of a secondary mediator, like the C-type lectin CD23 [42]. The novel vitamin D3 molecule 1,25(OH)2-16-ene-5,6-trans-D3 greatly suppresses cell proliferation by causing a block in the G1-G0 cell cycle, linked with increased expression of numerous cyclin-dependent kinase inhibitors (CDKIs). This process significantly lowers telomerase activity, while having an extremely low calcemic effect [43].
Vitamin D and its analogs have been included in leukemia therapy. It can be used in treating chronic myeloid leukemia (CML), acute lymphoblastic leukemia (ALL), acute myelogenous leukemia (AML) chronic lymphocytic leukemia (CLL), and childhood acute lymphoblastic leukemia (cALL) [23,30,31]. In vivo studies showed decreased levels of this secosteroid circulating in the bloodstream. Supplementing sufficient in CML and ALL or higher in CLL dose of calcitriol has been linked with the arrest of leukemia development [46]. Moreover, in patients experiencing a relapse, vitamin D level was lowered, compared to those, who underwent complete remission [47]. It was also proven, that calcitriol insufficiency is majorly connected with shortened time-to-treatment and overall survival [48].
3.4. Vitamin E
Tocotrienol and tocopherol belong to the vitamin E group. They both comprise four isoforms—alpha, beta, gamma, and delta. Their high levels can be found in some vegetable oil and certain types of seeds, nuts, and grains [49]. Apart from being well-known antioxidants, they display various other properties and act as antitumor, anti-inflammatory, antidiabetic, neuroprotective, and cardioprotective agents [50]. Tocotrienol activities surpass tocopherol’s, because of the former’s more effective incorporation into the cell’s lipid layer. Vitamin E activates the pregnane X receptor (PXR), an orphan nuclear receptor. PXR and RXR can combine into a heterodimer, which then binds to particular elements in gene regions and causes a reaction [51].
Studies in vitro, mostly on K562 cells, show tocotrienol’s involvement in the reduction of telomerase activity. They lower the expression of hTERT and c-myc mRNA through protein kinase C inhibition [5]. They also activate apoptosis by the use of various signaling pathways, including the mitochondrial pathway (intrinsic), the death receptor pathway (extrinsic), or other mechanisms like the endoplasmic reticulum-mediated apoptotic pathway. They upregulate the expression of the Bcl-2 protein, which is vital for an intrinsic pathway. Furthermore, they can activate caspase-3, the release of cytochrome c, and cleavage of PARP-1 to induce cell death [52]. They also activate granulocytic differentiation [53]. Those apoptotic properties are displayed in Figure 10. Tocotrienol and ferulic acid work synergistically and greatly decrease hTERT expression [5].
Vitamin E and its derivatives have been proposed in AML therapy [52]. Patients who were supplemented with vitamin E during the chemotherapy had a great increase in total antioxidant, serum albumin, ceruloplasmin, and immunoglobulin level with a significant reduction in serum ferritin [54]. It can also be considered in treating chronic myeloid leukemia (CML), where it can act as a leukemic cell differentiation inducer [55]. Clinical trials demonstrate positive results in cancer prevention by supplementing tocopherol and catechol-O-methyltransferase (COMT) [56].
3.5. Fatty acids
Dietary lipids compose mainly of fatty acids, which can be saturated or unsaturated. The most known polyunsaturated acids are Ω-3 and Ω-6 fatty acids, due to their role in the human body. They are involved in the creation of cell membranes, regulation of blood pressure, and inflammatory responses. Recent studies also indicate their antitumor properties [57]. They play a vital role in steroid enzyme creation and work as protective agents in cardiovascular disease and dementia [5]. Fatty acids C18-C22 directly affect telomerase activity. Cis-fatty acids surpass trans-fatty acids in effectivity and the inhibitory abilities increase with the number of double bonds, which makes docosahexaenoic acid (DHA) the most effective of them all [58].
Fatty acids inhibit telomerase activity on the transcriptional level. Eicosapentaenoic acid (EPA) shows competitive inhibition toward telomerase substrate primer. This means it can interact with the telomerase primer-binding site. Physiological doses of DHA and EPA (≤50 µM) are shown to inhibit PKC, which causes downregulation of hTERT and c-Myc mRNA, which results in lowered telomerase activity [5]. Mono-unsaturated linear-chain fatty acids in the cis configuration with C4A hydrocarbon chains (i.e. oleic acid) also severely inhibit the activity of telomerase. Oleic acid decreases telomerase activity competitively to the telomerase substrate primer [59]. The use of valproic acid (VPA) alone or combined with bortezomib treatment on HL-60 cells resulted in cyclin D1 and hTERT inhibition, lowered telomerase activity, arrested proliferation, and triggered apoptosis [47,48].
23-Hydroxybetulinic acid is a substance which can be extracted from Pulsatilla chinensis. It is a Chinese medicine herb that can be used in treating malaria and amoebic dysentery. It can also be used as a detoxifying agent. A study on HL-60 shows its apoptotic properties. This acid down-regulates Bcl-2 and lowers telomerase activity [62]. Sodium butyrate also shows anticancer abilities. Studies on U937 and HL-60 cells display inhibition of growth and induction of apoptosis after the use of this substance. The activity of telomerase was dramatically suppressed and the expression of hTERT was gradually down-regulated [50,51].
Clinical trials and treatment focus on fatty acid properties, that are not connected to telomerase activity. Ω-3 fatty acids effectively target the nuclear factor kappa B (NFκB) pathway in chronic lymphocytic leukemia (CLL), which plays a vital role in the control of apoptosis and progression of hematologic malignancies [65]. It was also used to prevent severe neutropenic enterocolitis in patients with AML, and cardiotoxicity in children with ALL, who are treated with doxorubicin [53,54]. Moreover, it was proven to reduce inflammatory risk in patients undergoing chemotherapy, due to hematological malignancies [68]. VPA can be used in AML treatment, where it acts as a histone deacetylase inhibitor and triggers cytotoxicity toward tumors in AML blasts [69]. VPA and low-dose cytosine arabinoside (Ara-C) indicate positive results in the treatment of elderly patients with AML/RAEB, which means it can be considered an alternative treatment for individuals who are unable to receive standard induction therapy [70].
3.6. Other substances
In this section, we have collected studies about the substances that did not fit into the aforementioned categories. There are also few studies about them, but we decided to describe them, as they also seemed to play a role in the inhibition of telomerase in leukemias and, as a result, they may play a major role in future therapy development.
Sulfoquinovosyldiacylglycerol (SQDG) is a substance found in plants and seaweeds. The studies indicate its inhibitory effect on human telomerase [71]. It also targets acute lymphoblastic leukemia (ALL) cells by decreasing the catalytic activity of the topoisomerase I enzyme, activating the p53-dependent apoptotic pathway [72]. Moreover, it promotes ataxia telangiectasia-mutated (ATM) and Rad3-related kinase (ATR), which is an important part of the cell cycle checkpoint machinery, recruitment at chromatin [73]. Those processes stop cells in S-phase [72].
Genistein is an isoflavone found in such plants as lupin, fava beans, soybeans and psoralea. In the study performed on the cells from the HL-60 cell line, both cell viability and down-regulation of the hTERT gene were noticed. Induction of apoptosis increased by 4,25-fold, with IC50 of 50μM. The activity of the hTERT decreased in a dose- and time-dependant manner. Compared to the control group, it has decreased the most 5,16-fold at 24 hours [11].
Sulforaphane (SFN) is a chemical compound belonging to the isothiocyanate group. It is found naturally in cruciferous vegetables, such as broccoli or Brussel sprouts. In the study performed by H. Shang et al. on the cells from the HL-60 cell line 8mM of SFN after 48 hours of application down-regulated hTERT gene expression 2,1-fold, which was noted by cDNA microarray analysis [74].
Melatonin is a hormone widely known for its control of the sleep-wake cycle [75]. It is synthesized in a human pineal gland, but can also be found in a variety of foods, such as eggs, fish, and nuts [76]. Apart from its wide range of bioactive properties, including anti-oxidant and anti-inflammatory qualities, it also displays anti-tumor effects. Studies on cell lines RS4-11 (MLL-AF4+ B-ALL), Nalm-6 (non-MLL-r B-ALL) and MOLM-13 (MLL-AF9+ AML) show the melatonin-induced suppression of mixed lineage leukemia (MLL). This substance inhibits hTERT expression by invalidating the binding activity of RBFOX3 to the hTERT promoter, which ceases cell proliferation. Moreover, it blocks NF-κB nuclear translocation and inhibits NF-B binding to the COX-2 promoter, causing lower COX-2 production. Clinical data showed that melatonin displays an anti-leukemic effect in primary MLL-r leukemia blasts ex vivo. In an MLL-r leukemia xenograft mouse model, animals treated with melatonin had a greater decrease in leukemic burden than the control group [75].
Crocin is a water-soluble carotenoid pigment, which can be found in the stigma of crocuses (Crocus sativus) or gardenias. It has four chemical analogs, but due to its pharmacological effects and the amount found in saffron, crocin 1 (alpha-crocin; crocetin digentiobiose ester) has been studied the most. It has multiple properties and participates in the creation of adenosine triphosphate (ATP), signal transduction, and redox homeostasis. Its direct connection with lowered telomerase activity was observed on HepG2 cells, where hTERT expression was halted. Its anti-leukemic properties were studied on cell lines HL-60 and resulted in inhibited proliferation and induced apoptosis. Crocin treatment of HL-60 xenografted mice reduces tumor weight and size, as well as Bcl-2 expression, while increasing Bax expression [77]. Crocin also induces apoptosis and stops cell growth in Jurkat (human T-cell leukemia cell line), which can be an indication of its therapeutic properties in treating T-lineage acute lymphoblastic leukemia (T-ALL) [78].
Withaferin-A (Wi-A) is a steroidal lactone that can be found in Withania somnifera (Ashwagandha). The direct effect of this substance on telomerase is yet to be established, but it has been reported, that it has an antitumor and growth inhibitory effect. Moreover, this substance impairs metastasis and angiogenesis. A study on U937 cells proves that Wi-A induces apoptosis through the activation of caspase-3, Akt signal pathway, and NF-KB activity inhibition [79]. Another study on JFCF-6B (TEP – cells with telomerase) and JFCF-1L (ALT – cells without telomerase) showed, that Wi-A works on both TEP and ALT. It affects p53 activation, induction of reactive oxygen species (ROS), induction of DNA damage, and cytoskeletal dysfunction in both cases. In TEP cells, it also affects telomerase and causes growth arrest and in ALT cells result in apoptosis. This study was not conducted on the leukemia cell line, but it describes how Wi-A influences the ALT mechanism, which is observed in leukemia and may be prospective study material [80].
Diosgenin is a natural steroidal sapogenin found in the seeds of fenugreek (Trigonella foenum-graecum) and the roots of wild yam (Dioscorea villosa) [81]. It has multiple purposes and can be used in treating diabetes, asthma, arthritis, cancer, and cardiovascular disease [82]. A study on chronic myeloid leukemia (CML) cell lines K562 and BaF3-WT display strong anti-cancer properties. It generates ROS, which has a cytotoxic effect on CML cells and induces autophagy, which functions as a cytoprotective [83]. The direct connection between telomerase and diosgenin was found during research on rat C6 and human T98G glioblastoma cell lines – the study showed downregulation of TERT expression [81].
Berberine is an alkaloid, that can be found in barberries (Berberis vulgaris), Oregon grape (Mahonia aquifolium), tree turmeric (Berberis aristate), and goldenseal (Hydrastis canadensis) [84]. It can be used in treating various ailments, such as cancer, cardiovascular, digestive, neurological, and metabolic diseases. Moreover, it inhibits toxins and bacteria, such as Helicobacter pylori, regulates lipid metabolism, and reduces hypertension [85]. A study on HL-60 cells showed berberine-induced apoptosis and significant inhibition of telomerase activity [86]. Another study showed, that berberine and its derivatives, such as coralyne, interact with G-quadruplex DNA structure (guanine-rich DNA sequence) at telomeres [87].
Apigenin (4′,5,7-trihydroxyflavone) is a flavonoid, that can be found in vegetables, like onions and parsley, fruits, like oranges, herbs, like basil, and oregano, and plant-based beverages, like wine and tea. It has antioxidant, inflammatory, and autoimmune properties ad can be used in therapy for neurogenerative diseases and several types of cancer [88]. A study on human leukemia cells U937, HL-60, and THP-1showed apigenin-induced toxicity. This substance activates the caspase pathway and through caspase-dependent apoptosis and suppresses hTERT expression, it lowers the level of telomerase activity. Moreover, it increases the intracellular level of ROS [89].
Cordycepin (3′-deoxyadenosine) is an adenosine nucleoside derivative, which can be found in the fungus Cordyceps militaris. It has a wide therapeutic potential and can be used as an antioxidant, anti-cancer, anti-diabetic, antiviral, inflammatory, hepato-protective, anti-inflammatory, anti-aging, and anti-arthritic drug [90]. A study on human leukemia cells THP-1 and H937showed that cordycepin-induced apoptosis influences telomerase, causing its deactivation and downregulation of both hTERT, and its transcription factors - c-Myc and Sp1. Moreover, cordycepin inhibits the phosphoinositide-3-kinase (PI3K)/protein kinase B (Akt) pathway, which lowers the phosphorylation and translocation of hTERT [91].
Tanshinone IIA (Tan IIA) is a substance found in Salvia miltiorrhiza. It has a long history of usage in traditional Chinese medicine and is now used as a pharmacologically active lipophilic agent. It can be used in the treatment of cancer, obesity, diabetes, cardiovascular, neurogenerative, and cerebrovascular diseases [92]. A study on K562 and HL-60 cells display strongly reduced cell proliferation and lowered activity of telomerase with Tan IIA-induced apoptosis [93].
Tan IIA is not the only compound, that can be extracted from Salvia miltiorrhiza - the other is tanshinone I (Tan I). It is an abietane diterpenoid, that can be used as an anticoronaviral agent. It has therapeutic properties used in treating myeloid leukemia, osteoporosis, left ventricular hypertrophy, hearing loss, hypertension, and atherosclerosis [94]. A study on monocytic leukemia cells U937, SHI 1 and THP-1 showed Tan I-induced apoptosis by activation of the caspase-3 pathway and slowed down cell growth. This substance lowers the activity of telomerase and down-regulates the expression of hTERT in a dose-dependent manner [95].
Rapamycin is a substance, that was first isolated from the soil of Rapa Nui, the South Pacific island [96]. It is produced by the Streptomyces hygroscopius bacteria [97]. Rapamycin has been shown to significantly slow the aging process, and extend a lifespan. Moreover, it alleviates age-related disorders, efficiently prolonging a health span. It methodically eases aging in numerous organ systems, such as endocrine, digestive, neurological, circulatory, urinary, motor, respiratory, immunological, integumentary, and reproductive systems [96]. A study on the T-cell leukemia cell line Jurkat has been conducted to test rapamycin’s ability for growth suppression. This substance lowers telomerase activity and reduces hTERT expression. Moreover, it halts the cell at the G1 phase [98].
Caffeic acid undecyl ester (CAUE) can be naturally found in Daphne oleoides [99]. It displays multiple bioactivities, such as anti-inflammatory, antioxidant, and anti-leukemic properties [100]. A study on B cell precursor leukemia NALM-6 cells show, that CAUE selectively impairs DNA synthesis and lowers the telomerase activity via suppression of hTERT protein. The higher concentration of CAUE, the better inhibiting properties toward hTERT it displayed [101].
β-Lapachone is a substance found in the heartwood of the lapacho tree (Handroanthus impetiginosus). It has various biological activities, such as anti-cancer, antiviral, anti-parasitic, anti-inflammatory, antiobesity, neuroprotective, nephroprotective, antioxidant, and wound healing effects [102]. A study on human leukemia cells Hl-60, K562, U937, and THP-1 confirmed direct LAPA-induced cytotoxicity. It activates the subsequent cleavage of poly (ADP-ribose) polymerase and caspase-3. The cell death was connected to lowered telomerase activity and downregulation of hTERT expression [103].
Ginsenoside Rg1 (Rg1) is a bioactive substance, that can be found in the roots of ginseng (Panax ginseng), and turned white or red ginseng [90,91]. Rg1 has a hepatoprotective effect, promotes cerebral angiogenesis and lymphatic drainage, improves chronic inflammatory arthritis, and prevents PTSD-like behaviors [92,93,94,95]. A study on CD34+CD38− leukemic stem cells (LSC) showed, that Rg1 downregulates the expression of hTERT and upregulates p16INK4a (protein, that slows down the cell cycle) expression, which causes senescence of LSC [104]. Another study, using Korean red ginseng (KRG) on U937 cells showed its anti-cancer properties. KRG has been found to induce apoptosis and down-regulate hTERT expression in a concentration-dependent manner [105].
Bengalin is a protein isolated from the venom of an Indian black scorpion (Deccanometrus bengalensis). This substance has antileukemic and antiosteoporosis properties [96,97]. In a study on U973 cells, Bengalis display antiproliferative and apoptogenic effects selectively towards leukemic cells. This protein lowers telomerase activity, damages the nuclei, arrests the cell cycle at the G1 phase, and causes early apoptosis, mainly through the inhibition of heat shock proteins and induction of the mitochondrial pathway [112].
Ascorbic acid, widely known as vitamin C (VC), is being supplemented by many people every day. It can be found in fruits and vegetables and plays an important role in the human system. This well-known antioxidant helps with neuronal differentiation, reparations of tissue, the formation of collagen, and the activity of the immune system [99,100,101]. Perfluorooctanesulfonate (PFOS) is a global pollutant, which can cause toxic effects on various cells, including blood cells, which can result in leukemia. A bioinformatics study on PFOS-associated leukemia showed that VC targets hTERT, amongst other core proteins, which would suggest that it could lower the telomerase activity in vitro and in vivo [116]. A study using CD34+, HL-60, and U937 cell lines shows, that a high dose of vitamin C stops cells’ proliferation in a concentration-dependent manner. With higher VC concentration, a greater rate of apoptosis was detected [117].
Telomestatin is a new inhibitor of telomerase, that can be extracted from Streptomyces anulatus. It has great inhibition properties and affects telomerase without influencing polymerases and reverse transcriptases. This substance also promotes the shortening of telomeres [118]. A study on U937 cells showed activation of the telomestatin-induced caspase-3 pathway and apoptosis, due to telomerase inhibition. The growth of cells was also slowed in U937 cells in xenograft mouse mode, thanks to decreased telomerase level. Moreover, telomestatin suppresses the activity of the telomerase in BCR-ABL-positive leukemic cell lines in a reproducible manner [119].
Silymarin is a substance extracted from milk thistle (Silybum marianum). It is composed of flavonolignans, like silybin. This substance act as an antioxidant and have immunomodulatory, anti-cancerous, anti-inflammatory, and hepatoprotective effects [120]. A study on K562 cells was conducted in order to establish the antiproliferative effect of silymarin. This substance lowers the activity of telomerase and suppresses cell growth. The lowered telomerase level was connected with silymarin-induced apoptosis [121].
Tianshengyuan-1 (TSY-1) is a Chinese herbal medicine, which is a liquid mixture of various Chinese herbs. It has been used in treating bone marrow-connected diseases, such as anaplastic anemia [122]. It also affects the activity of telomerase in hematopoietic cells. In a study on mononuclear cells from normal peripheral blood (PBMC), HL-60, and CD34+ cell lines, the mechanism of the TSY-1 effect on telomerase was investigated. In PBMC and CD34+ stem cells, firstly low telomerase level increased and in HL-60 initially, high telomerase activity decreased. Those processes happen in a dose-dependent manner. TSY-1 effects on the activity of telomerase are related to cell senescence. TSY-1 induces hypomethylation of the core promoter of TERT in HL-60 cells and hypermethylation of the core promoter of TERT in PBMC and CD34+ [123].
Wogonin is one of a few active compounds found in Scutellaria baicalensis’ roots. It has been used in Chinese herbal medicine and is being tested in clinical trials now. This substance displays various bioactivities and can be used as an anti-inflammation, anti-cancer, antibacterial, antiviral, neuroprotective, and chondroprotective agent [110,111]. A study on HL-60 showed, that wogonin inhibits cell growth mostly via suppression of telomerase through inhibition of c-Myc and promotion of Bax/Bcl-2 apoptosis. Moreover, wogonin causes DNA fragmentation, increases caspase-3 activity, and inhibits hTERT [126].
Interferon alpha (IFN-α) is a protein made in the human body against viral infections, mainly by plasmacytoid dendritic cells (pDCs). It has been used in the treatment of myeloproliferative neoplasms (MPNs) and has been shown to induce hematological, histopathological, and molecular responses [127]. Although IFN-α has various impacts on stem cells and immunology, the mechanism of its anti-leukemic activity is still unknown. Numerous clinical trials have shown that chronic myeloid leukemia treatment with IFN- α had a higher number of remissive patients than traditional chemotherapy did. From the 1980s till 2001, it has been the first line of treatment for CML—right now the combination of IFN-α [EB1] and other drugs is being tested [128]. It was also tested as maintenance therapy on patients with favorable-risk AML and showed positive results [129]. IFN-α has also been shown to amplify graft-versus-leukemia (GVL) response in the case of allogeneic hematopoietic cell transplantation (HCT) for high-risk AML, which prevents leukemia relapse [130].
Angelica sinensis polysaccharide (ASP) is a water-soluble substance that can be found in Angelica sinensis and is used in traditional Chinese medicine. It has multiple purposes, such as promoting immunity, hematopoietic activity, and liver protection, and can be used as an antitumor, antioxidant, anti-inflammatory, antiviral, and anti-aging agent. Moreover, it can be used as a drug carrier [131]. A study shows, that ASP efficiently suppresses the proliferation of AML CD34+CD38− cells in a dose-dependent manner while preserving progenitor and normal hematopoietic stem cells at physiologically attainable doses. Moreover, it exhibits cytotoxic properties on AML K562 cells. ASP represses telomerase activity and upregulates p53, p21, p16, and Rb genes, which causes cell senescence [132].
Agaricus blazei Murill (AbM) is an edible mushroom found in Brazil. It exhibits various effects, such as antitumor, anti-inflammatory, anti-allergic, anti-HIV, hepatoprotective, and antioxidant [119,120,121]. An RNA-protein complex, FA-2-b-ß fraction, can be isolated from AbM. The antileukemic and proapoptotic effects of this substance were tested on HL-60 cells. FA-2-b-ß fraction causes a decrease in telomerase activity and induces caspase-3 gene expression, which may lead to apoptosis [136].
Platycodin D (PD) is a triterpenoid saponin, that can be found in Platycodon grandiflorus. It exhibits various bioactive properties and can be used in treating brain ischemia, cardiomegaly, cholestasis, esophagitis, fibrosis, hepatitis C, liver cirrhosis, obesity, osteoporosis, pneumonia, and some kinds of neoplasm [137]. A study on THP-1, K562, and U937, cells was conducted to test PD’s anticancer effects. PD has a direct cytotoxic influence on telomerase through a decrease in hTERT expression. It reduces the level of c-Myc and Sp1 proteins as well as DNA binding actions. Moreover, it suppresses activation, which reduces the phosphorylation and nuclear translocation of hTERT. Those findings reveal, that PD causes hTERT posttranscriptional and translational inhibition of telomerase activity [138].
Pectenotoxin-2 (PTX-2) is a natural toxin that can be isolated from marine sponges (Dinophysis species). They can affect actin cytoskeletons in vivo and in vitro. They display cytotoxicity toward various cancer cells, such as lung, colon, and breast cancer cells. A study on U937, HL-60, and THP-1 cells showed, that PTX-2 decreases the viability of cells and inhibits telomerase activity via suppression of hTERT expression. It reduces Sp1 and c-Myc gene expression and lowers the binding activity of their proteins to the hTERT regulatory regions. It also reduces phosphorylation and nuclear translocation of hTERT via attenuating the phosphorylation of Akt. This concludes, that PTX-2 suppresses telomerase activity through posttranscriptional and translational inhibition of hTERT [139].
Camptothecin (CPT) is a cytotoxic alkaloid that can be extracted from Camptotheca acuminata. It is known for having broad antiproliferative and anticancer properties due to its ability to inhibit DNA topoisomerase 1. Due to its instability, low solubility, and tumor cells’ developed resistance, a few derivatives were created. There are currently three CPT-inspired pharmaceuticals - topotecan, irinotecan, and belotecan – that are used in the treatment of various types of cancer [140]. They are silicon-containing agents with enhanced lipophilicity, which makes them more bioactive than their carbon analog [141]. In order to analyze its prospective use in leukemia treatment, a study on HL-60 human leukemia cells was conducted. This study shows, that CPT decreases telomerase activity in a time-dependent way, whilst inducing apoptosis. It also gradually reduces the expression of Bcl-2, but it has no impact on TERT expression [142].
Diterpene triepoxide (triptonide) is a substance, that can be acquired from a Tripterygium wilfordii Hook F (TwHF). It is used as a Chinese medicinal herb as an immunosuppressive agent and can be used in treating rheumatoid arthritis, psoriasis, and systemic lupus erythematosus [143]. It can act as a reversible non-hormonal contraceptive for male mice and non-human primates [144]. It has anti-cancer activities and is researched as a potential treatment for various neoplasms, like lung, breast, and thyroid cancer [131,132,133]. To test its anti-leukemic properties a study on AML cell lines was conducted. It was found, that triptonide induces absolute senescence in U937 and HL-60 cells, as well as inhibits growth and colony formation. In a mouse xenograft model, this substance almost completely reduces human leukemia cell tumorigenicity with no evident toxicity. It causes apoptosis by down-regulating the transcription of TERT and oncogenic c-Myc. At the same time, it promotes the transcription of senescence-promoting genes, such as p16 and p21, as well as pro-apoptotic genes [148].
Constunolide is a sesquiterpene lactone present in the stem bark of Magnolia sieboldii and it exhibits anti-carcinogenic, anti-viral, anti-inflammatory, and anti-fungal properties. In the study performed on the NALM-6 cells, 10 μM of constunolide were used, which caused suppression of telomerase activity to 94%, 89%, 21% and 2% after 1, 2, 4 and 6 hours, respectively. Moreover, hTERT mRNA and hTERT protein expression decreased. In the case of mRNA – to 96, 69, 21, 20% after 1, 2, 4, 6 hours respectively, and when it came to the protein – to 97, 69, 21, 20% after similar times. The effect of caspase inhibitors was also examined during the study. It was concluded that the caspase inhibitors blocked apoptotic death of the cells (however, not the caspase 9 inhibitor) [149].
Salvicine is a structurally modified derivative of Salvia prionitis that has anti-tumour activities. In the study, telomerase activity was measured in the HL60 cells after adding 10 μM of salvicine. Telomerase activity was lowered to 77.9, 51.3, 36.2% after 2, 4, 6 hours, respectively. Meanwhile, the enzyme activity was also measured after 4 hours of applying 2.5, 5, 10 and 20 μM of the substance, which caused a decrease to 76.3, 60.1, 40.9, 32.2% of activity, respectively [150].
Patensin is a substance derived from Pulsatilla patens var. multifida and is used as a treatment of amoebic, bacterial diseases, as well as cancers. It causes growth arrest and apoptotic cell death. In the study performed on the HL60 cell line, telomerase activity was measured by the TRAP assay. 100 μM of patensin was applied and after 3, 6, 12 and 24 hours, telomerase activity was down-regulated to 62.98, 44.23, 32.32, 23.30% respectively [151].
Dideoxypetrosynol A is a polyacetylene present in the marine sponge Petrosia sp. It has shown cytotoxicity towards several human cancer cell lines. In the study performed by K. Mandel et al. on the U937 cell line, different concentrations of dideoxypetrosynol A were added and the cells were incubated for 48 hours. Down-regulation of hTERT mRNA was observed at 0,6 μg/ml, however, there was no effect on hTR, TEP-1 and c-myc mRNA [152].
(Z)-stellettic acid C is an acetylenic acid present in the Stelletta sp. sponges. It has shown selective cytotoxicity toward human cancer cell lines. In the study on U937 cells, telomerase activity was inhibited in a concentration-dependent manner and was the most effective at 30 μg/ml, however, there was no change in TEP-1 and hTR mRNA expression. Similarly, (Z)-stellettic acid C had no effect on the crucial transcription factors for the regulation of transcription of hTERT – Sp-1 and c-Myc [153].
Trichostatin A is an organic substance with a histone deacetylase activity. In the study performed on the U937 cells different doses of the substance were added and the cells were incubated for 48 hours. The hTERT mRNA expression was inhibited and was the most effective with the concentration of 75 nM, whereas TEP-1 and hTR mRNA expression remained unchanged [154].
Homoharringtonine (HHT) is a substance derived from the Chinese evergreen Cephalotaxus harringtonia and exhibits anti-tumour activity. W. Xie et al. performed a study on the HL60 cells in order to examine inhibition of telomerase activity after adding HHT. Different doses (0-500 μg/ml) were added, and the cells were incubated for 48 hours. The activity decreased from 1.056 ± 0.107 to 0.067 ± 0,023. Inhibition of telomerase proceeded in a time- and dose-dependent manner [155].
Baicalin is a flavonoid substance isolated from Scutellaria baicalensis. It has hepatoprotective, anti-bacterial and anti-inflammatory properties. In a study carried out on the HL60 cells, it was concluded that 20 μg/ml of baicalin used for different durations of time down-regulate c-Myc mRNA and protein levels. As a result hTERT mRNA expression was also decreased as it is a transcriptional target of c-Myc [156].
Manisa propolis is a wax-like substance collected by honeybees. It is believed to have ant-microbial, anti-inflammatory anti-oxidant and anti-tumoral properties. O. Cogulu et al. examined the impact of propolis on four childhood leukemia cases – three of which were acute lymphoid leukemia and one was chronic myeloid leukemia. At 24 hours after applying 60 ng/ml of propolis, the expression level of hTER decreased [157]. In another study performed on the T-cell ALL cells (CCFR-CEM) 0,03 μg/ml were added to cells, which resulted in a decrease in the hTERT ratio by 60 and 93% at 24 and 72 hours respectively [158].
Gymnodinium sp. A3 (GA3) is a marine microalga, that was first isolated from the Seto Inland Sea. When this dinoflagellate grows in saltwater, it produces extracellular acidic GA3 polysaccharide, which is a D-galactan sulfate associated with L(+)-lactic acid. This substance demonstrated significant cytotoxicity to various human leukemic cell lines [159]. A study on K562 cells displays GA3 polysaccharides’ ability to lower telomerase activity [160]. Moreover, it inhibits topoisomerase-I and topoisomerase-II resulting in apoptosis [159].
Oridonin is a natural terpenoid, that can be isolated from herbs like Isodon rubescens and Rabdosia rubescens, which is used in traditional Chinese medicine. It has a variety of bioactive properties, such as anti-cancer, cardioprotective, anti-inflammatory, and hepatorenal activities [161]. A study on HL-60 cells shows, that oridonin-induced apoptosis, decrease in telomerase activity, and inhibited cell growth are time- and dose-dependent [162]. Moreover, in a study on K52 cells, telomerase activity and Bcl-2 expression were downregulated, whilst Bax expression was upregulated during the time of apoptosis [163,164]. These results provide a reason for clinical trials of oridonin treatment of acute leukemia [162]. This substance shows its effectiveness on the HPB-ALL cell line. It causes downregulation of Bcl-2 and Bcl-XL, activation of caspase-3, and finally, apoptosis and necrosis [164].
Gold is a natural element that has been used for treating human pathologies for a long time. It has a wide spectrum of properties and can be used both in diagnosis and treatment. It is proven to work on treating degenerative and infectious diseases, as well as rheumatoid arthritis and cancer [165]. Gold nanoparticles (AuNPs) are mainly used in early neoplasm detection and diagnosis. Moreover, they are also able to selectively target the cells and deliver specific pharmaceuticals to a distinct location, which makes them a very worthy research material [166]. Furthermore, in order to test AuNPs’ cytotoxicity, research on K562 cell lines was conducted, in which the cells were exposed to the nanoparticles for three days straight. The outcome indicates that, although certain nanoparticle precursors may be toxic, the AuNPs themselves do not necessarily harm biological function and were concluded to not cause acute cytotoxicity [167]. Another study, using AuNPs synthesized using leaf extract of Camellia sinensis shows a great increase in the anti-inflammatory cytokines and the lymphocyte in the leukemic mouse model. Moreover, it displays a decrease in the pro-inflammatory cytokines, and the total white blood cells count. It is also suggested as a chemotherapeutic drug for AML treatment [168]. The low level of cytotoxicity makes it a great potential target for future studies and hopefully results in further research of AuNPs as a way of delivering telomerase-inhibiting substances into specific cells.
Ultrasonography (USG) is a diagnostic tool that is non-invasive, safe, and inexpensive. Due to its easy use, it has been spreading among doctors as an everyday use object. It enables physicians to see patients’ organs in real-time and in the least invasive way possible. Nowadays, scientists discover new ways of using USG as a therapeutic appliance because of its ability to induce chemical, thermal, and mechanical effects in the human body [169]. A study on human K562 and U937 cell lines and primary T cells show very promising effects of low-intensity pulsed ultrasound. It was not connected with telomerase activity directly, but it showed a decrease in Bcl2 and a great increase in apoptotic cells. This technique was able to selectively attack leukemic cells and save more than 80% of T cells - because of that, it may soon become a new target therapy [170].

Table 1.
Substances used in this review.
| Substance | Natural occurance | Cell line | Dose applied | Durance | |
|---|---|---|---|---|---|
| Curcumin | Turmeric | HL-60 | 1, 10, 50 μM | 24 h | [13] |
| HL-60 | 10, 15, 20, 40 μM | 24 and 48 h | [14] | ||
| K-562 | 1, 10, 50 μM | 6, 16, 24, 48 h | [15] | ||
| Epigallocatechin-3-gallate (EGCG) | Green tea | HL-60 | 50 μM | 3, 6, 9, 12 days | [16] |
| Jurkat | 30, 50, 70, 85, 100 μM | 24, 48, 72 h | [17] | ||
| Indole-3-carbinol | Broccoli, cauliflower, brussels sprouts, cabbage, kale | NALM-6 | 20, 30, 40, 50, 60 μM | 24, 48 h | [19] |
| K-562 | 100, 200, 400 μM | 24, 48 h | [20] | ||
| Butein | Stem bark of cashews, the heartwood of Dalbergia odonifea, Rhus verniciflua, and Caragena jubata | U937, THP-1, HL60, K562 | 5, 10, 15, 20, 30 μM | 24 h | [21] |
| Gossypol | Seeds, roots and stems of cotton plants | HL-60 | 1, 2, 4.5, 6, 10 μM | 24, 48, 72, 96 h | [22] |
| U937, HL60, THP-1, K562 | 5, 10, 20 μM | 24 h | [23] | ||
| Retinoic acid | Yellow, green, red, and leafy vegetables, yellow fruits | HL-60 | 1 μM | 12 days | [27] |
| 242 cases of APL | 60 mg/kg/day | 2 years | [171] | ||
| 124 cases of cAPL | 25mg/m2/day | 5 years | [172] | ||
| Eμ-TCL1 mice | 1 μM | 24, 48 h | [35] | ||
| 9cUAB30 | Novel retinoid | HL-60d | 5 μM | 12 days | [29] |
| Growth factor binding protein 7 | Human insulin recombinant | HL60, KG1a, THP1, HEK293T | 100 μg/mL, 300 μg/mL, 10 mg/kg, 12 mg/kg | 48, 72, 120 h, 7, 14 days, 16 weeks | [30] |
| Vitamin D | Produced in the human body form 7-dehydrocholesterol; fish oil, egg yolk | HL-60 | 10-8M | 3 days | [173] |
| U937 | 10-9-10-7M | 48, 72 h | [174] | ||
| 26 cases of AML | 1 μg/day | 2 x 5 weeks | [175] | ||
| 17 cases of elderly AML | 100,000 IU/week | 6 months | [176] | ||
| EB 1089 | Vitamin D3 analog | HL-60, U937 | 5x10-10M | 96 h | [42] |
| 1,25(OH)2-16-ene-5,6-trans-D3 | Vitamin D3 analog | HL-60 | 10-7M | 4 days | [43] |
| Vitamin E | Vegetable oil, seeds, nuts, grains | U937, KG-1 | 10-50 μM | 24 h | [52] |
| K562 | 100 μM | 48 h | [177] | ||
| 25 cases of AML | 400 IU/day | 30 days | [54] | ||
| 2396 study/2235 control | 600 IU/day | 10 years | [56] | ||
| Valproic acid | Valeriana officinalis | HL-60, HL-60A | 0.5, 1, 2, 4, 8 mM | 24, 48, 72 h | [60] |
| 18 + 114 patients of AML | 400 mg/day + from day 3 60 - 150 mg/L | 2 x 21 days, 3 x 21 days | [69] | ||
| 31 cases of elderly AML/RAEB | 5 mg/kg daily | 2-23 months | [70] | ||
| 23-Hydroxybetulinic acid | Pulsatilla chinensis | HL-60 | 1 - 1000 μM | 3, 6, 12, 24 h | [62] |
| Ω-3 fatty acids (DHA, EPA) | Seafood, especially fatty fish, nuts, seeds, plant oils | 16 cases of Rai Stage 0-1 CLL | 1. month: 3 x 1250 mg/day 2. month: 6 x 1250 mg/day 3. – 12. month: 9 x 1250 mg/day |
12 months | [65] |
| 14 cases of AML | 100 mL/day | 2 years | [67] | ||
| 60 cases of cALL (30 study/30 control) | 1000 mg/day | 6 months | [66] | ||
| SQDG | Azadirachta indica | MOLT-4 | 5, 10, 15, 20, 25 μM | 2, 4, 8, 12, 16, 20, 24, 48, 72 h | [72] |
| Genistein | Lupin, fava beans, soybeans, psoralea | HL-60 | 50 μM | 24, 48, 72 h | [11] |
| Sulforaphane | Cruciferous vegetables | HL-60 | 6, 7, 8, 9, 10 μM | 24, 48 h | [74] |
| Melatonin | Synthesized in a human pineal gland, eggs, fish, nuts | RS4-11 (MLL-AF4+ B-ALL), MOLM-13 (MLL-AF9+ AML), Nalm-6 (non MLL-r B-ALL) | 1, 2, 3 mM | 24, 48, 72 h | [75] |
| Crocin | Crocus sativus | Jurkat | 0.625, 1.25, 2.5, 5, 10 mg/mL | 12, 24, 26, 48 h | [78] |
| Withaferin-A | Withania somnifera | U937 | 0.25, 0.5, 1, 2 μM | 12, 24 h | [79] |
| Diosgenin | Trigonella foenum-graecum | K562 | 5, 10, 15, 20, 30 μM | 48 h | [83] |
| Berberine | Berberis vulgaris, Mahonia aquifolium, Berberis aristate, Hydrastis canadensis | HL-60 | 5, 10, 15, 25, 50 μM | 24, 48, 72, 96 h | [86] |
| Apigenin | Basil, oregano, onion, parsley, wine, tea, beer | HL-60, U937, THP-1 | 25, 50, 75, 100 μM | 24 h | [89] |
| Cordycepin | Cordyceps militaris | THP-1, H937 | 10, 20, 30 μM | 24 h | [91] |
| Tanshinone IIA | Salvia miltiorrhiza | HL-60, K562 | 0.5 μg/mL | 5, 6 days | [93] |
| Tanshinone I | Salvia miltiorrhiza | U937, THP-1, SHI 1 | 10, 20, 30, 40, 50 μM | 24, 48, 72 h | [95] |
| Rapamycin | Streptomyces hygroscopius bacteria | Jurkat | 1, 10, 100 nM | 8, 16, 24, 32, 72 h | [98] |
| Caffeic acid undecyl ester | Daphne oleoides | NALM-6 | 0.1, 0.3, 0.6, 1, 3 μM | 4, 18 h | [101] |
| NALM-6 | 0.1, 0.3, 0.6, 1μM | 6, 12, 24, 72 h | [100] | ||
| β-Lapachone | Handroanthus impetiginosus | U937, K562, HL60, THP-1 | 1, 2, 3, 4 μM | 24 h | [103] |
| Ginsenoside Rg1 | Panax ginseng | CD34+CD38- LSCs | 20, 40, 80 μM | 24, 48, 72 h | [104] |
| U937 | 0.2, 0.4, 0.6, 0.8, 1, 1.2, 1.4, 1.6, 0.8, 2 mg/mL | 24, 48, 72 h | [105] | ||
| Bengalin | Venom of Deccanometrus bengalensis | U937, K562 | 0.3, 0.75, 1.5, 1.85, 2.05, 2.23, 3, 3.7, 4.1, 4.5, 6 μg/mL | 24, 48 h | [112] |
| U937 | 1.8, 3.7 μg/mL | 12, 24 h | [110] | ||
| Ascorbic acid | Citrus fruits, green and leafy vegetables | CD34+, HL-60, U937 | 8, 20 mM | 24 h | [117] |
| Telomestatin | Streptomyces anulatus | U937 | 1, 2, 5, 10 μM | 48 h, 0 – 60 days | [119] |
| Silymarin | Silybum marianum | K562 | 10, 25, 50, 75, 100 μg/mL | 24, 48, 72 h | [121] |
| Tianshengyuan-1 | A mixture of various Chinese herbs | HL-60, PBMCs, CD34+ HSCs | 31.2, 62.5 μg/mL | 24 h | [123] |
| Wogonin | Scutellaria baicalensis | HL-60 | 0.5, 1, 2, 3 mg/mL 10, 25, 50, 75, 100 μM | 24 h | [126] |
| Interferon alpha | Synthesized in the human organism by plasmacytoid dendritic cells | 42 study/42 control of AML | 3 x week 3 mln IU | 12 – 18 months | [129] |
| 36 | -1. before HCT, 14., 28., 42. (+/- 7) days 45, 90, 180 μg | 24 months | [130] | ||
| Angelica sinensis polysaccharide | Angelica sinensis | K562, CD34+CD38− | 20, 40, 80 μg/mL | 48 h | [132] |
| FA-2-b-ß fraction (an RNA-protein complex) | Agaricus blazei Murill | HL-60 | 5, 10, 20, 40, 80 μg/mL | 24, 48, 72, 96 h | [136] |
| Platycodin D | Platycodon grandiflorus | U937, THP-1, K562 | 5, 10, 15, 20 μM | 48 h | [138] |
| Pectenotoxin-2 | Dinophysis species | U937, THP-1, HL-60 | 2, 4, 6, 8, 10 ng/mL | 72 h | [139] |
| Camptothecin | Camptotheca acuminata | HL-60 | 1 mg/L | 2, 4, 6 h | [142] |
| Triptonide | Tripterygium wilfordii Hook F | U937, HL-60 | 0-10, 15, 20 nM 2, 4 mg/kg |
72 h, 3, 6, 9, 14, 21 days | [148] |
| Constunolide | Stem bark of Magnolia sieboldii | NALM-6 | 2, 6, 8, 10 μM | 1, 2, 4, 6 h | [149] |
| Salvicine | Salvia prionitis (modified) | HL-60 | 2.5, 5, 10, 20 μM | 2, 4, 6 h | [150] |
| Patensin | Pulsatilla patens var. multifida | HL-60 | 0.001, 0.01, 0.1, 1, 10 mmol/L | 3, 6, 12, 24 h | [151] |
| Dideoxypetrosynol A | Marine sponge Petrosia sp. | U-937 | 0.2, 0.4, 0.6, 0.8, 1 μg/mL | 48 h | [152] |
| (Z)-stellettic acid C | Stelletta sp. sponges | U-937 | 5, 10, 20, 30, 40 μg/mL | 48 h | [153] |
| Trichostatin A | Culture broth of Streptomyces platensis | U-937 | 15, 30, 45, 60, 75 nM | 48 h | [154] |
| Homoharringtonine | Chinese evergreen Cephalotaxus harringtonia | HL-60 | 5, 10, 50, 100, 500 μg/L | 3, 6, 12, 24, 48 h | [155] |
| Baicalin | Scutellaria baicalensis | HL-60 | HL-60 5, 10, 20, 40, 80 μg/mL | 12, 24, 48 h | [156] |
| Manisa propolis | Substance collected by honeybees | 4 childhood leukemia cases (3 ALL, 1 CML) | 15, 30, 60 ng/mL | 24, 48, 72 h | [157] |
| CCFR-CEM | 0.03 μg/mL | 24, 48, 72, 96 h | [158] | ||
| D-galactan sulfate | Gymnodinium sp. A3 | K562 | 0.003, 0.01, 0.03, 0.1, 0.3, 1 μg/mL | 15, 45 min | [159] |
| Oridonin | Isodon rubescens, Rabdosia rubescens | HL-60 | 4, 8, 16, 24 μM | 12, 36, 48, 60, 72 h | [162] |
| HPB-ALL | 16, 24, 32, 40, 56 μM | 24, 48, 72 h | [164] | ||
| Gold nanoparticles | Gold | HL-60 | 1-1000 μg/mL, 2500 mg/kg | 28 days | [168] |
| Low-intensity pulsed ultrasound | Ultrasound | K562, U937 | 0.3, 0.5, 0.67 MHz | 60 s cycle 1, 10, 20, 30 ms pulse duration 72 h 2 min cycle 1, 10, 20, 30, 100 ms pulse duration 2 days |
[170] |
4. Conclusions
The inhibitory activity towards telomerase of natural substances is gaining more attention from researchers. Experimental studies carried out in vitro and in vivo indicate their significant potential to reduce the development of leukemias. Moreover, the results presented in the paper indicate that natural substances can exert their anti-leukemic effects in three ways. Not only do they arrest the growth of the cells, but they also induce apoptosis and differentiation of the cells. Nowadays, there are too few research papers to decide whether or not they are, indeed, sufficient in reducing the development of leukemias and in treating them. Additionally, it should be mentioned that apart from the benefits, there may also be several limitations associated with the use of natural substances, such as:
- Their ambiguous effect on chemoprevention;
- The lack of data indicating the optimal and toxic doses;
- The lack of data regarding their potential side effects;
- The lack of data evaluating their pharmacodynamic properties.
Moreover, most epidemiological studies assess the overall impact of fruit and vegetable consumption on the development of various types of cancer. Plants are the source of many phytochemicals, and for this reason, it is not always possible to clearly determine which chemical compound has a significant anticancer effect. Nevertheless, it seems that phytochemicals have a significant preventive as well as therapeutic potential regarding leukemias. For this reason, the consumption of larger quantities of vegetables, fruit, and whole-grain products should be recommended for the prevention of various leukemias.
Author Contributions
Conceptualization, E.B., K.M.,M.W.; methodology, E.B., K.M.; formal analysis, E.B., A.C.; investigation, E.B., K.M.; data curation, E.B., K.M.; writing—original draft preparation, E.B., K.M.; writing—review and editing, E.B., K.M., M.W.; visualization, E.B., K.M.; supervision, M.W., A.C.; project administration, M.W., A.C.; funding acquisition, M.W, A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zvereva, M.I.; Shcherbakova, D.M.; Dontsova, O.A. Telomerase: Structure, Functions, and Activity Regulation. Biochemistry (Moscow) 2010, 75, 1563–1583. [Google Scholar] [CrossRef]
- Wang, Y.; Sušac, L.; Feigon, J. Structural Biology of Telomerase. Cold Spring Harb Perspect Biol 2019, 11. [Google Scholar] [CrossRef]
- Dratwa, M.; Wysoczańska, B.; Łacina, P.; Kubik, T.; Bogunia-Kubik, K. TERT—Regulation and Roles in Cancer Formation. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Sandin, S.; Rhodes, D. Telomerase Structure. Curr Opin Struct Biol 2014, 25, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Goel, H.; Kumar, R.; Tanwar, P.; Upadhyay, T.K.; Khan, F.; Pandey, P.; Kang, S.; Moon, M.; Choi, J.; Choi, M.; Park, M.N.; Kim, B.; Saeed, M. Unraveling the therapeutic potential of natural products in the prevention and treatment of leukemia. Biomed Pharmacother 2023, 160, 114351. [Google Scholar] [CrossRef]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.C.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific Association of Human Telomerase Activity with Immortal Cells and Cancers. Science (1979) 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Trybek, T.; Kowalik, A.; Góźdź, S.; Kowalska, A. Telomeres and Telomerase in Oncogenesis (Review). Oncol Lett 2020, 20, 1015–1027. [Google Scholar] [CrossRef]
- Cesare, A.J.; Reddel, R.R. Alternative Lengthening of Telomeres: Models, Mechanisms and Implications. Nat Rev Genet 2010, 11, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Telomerase Inhibitors from Natural Products and Their Anticancer Potential. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Wei, Z.-Z.; Qin, Q.-P.; Chen, J.-N.; Chen, Z.-F. Oxoisoaporphine as Potent Telomerase Inhibitor. Molecules 2016, 21, 1534. [Google Scholar] [CrossRef]
- Balci Okcanoglu, T.; Avci, C.B.; Yılmaz Süslüer, S.; Kayabasi, C.; Saydam, G.; Gunduz, C. Genistein-Induced Apoptosis Affects Human Telomerase Reverse Transcriptase Activity in Acute Promyelocytic Leukemia. Cyprus Journal of Medical Sciences 2020, 5, 153–156. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee (nee Chakraborty), S.; Ghosh, U.; Bhattacharyya, N.P.; Bhattacharya, R.K.; Dey, S.; Roy, M. Curcumin-Induced Apoptosis in Human Leukemia Cell HL-60 Is Associated with Inhibition of Telomerase Activity. Mol Cell Biochem 2007, 297, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Dikmen, M.; Canturk, Z.; Ozturk, Y.; Tunali, Y. Investigation of the Apoptotic Effect of Curcumin in Human Leukemia HL-60 Cells by Using Flow Cytometry. Cancer Biother Radiopharm 2010, 25, 749–755. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Ghosh, U.; Bhattacharyya, N.P.; Bhattacharya, R.K.; Roy, M. Inhibition of Telomerase Activity and Induction of Apoptosis by Curcumin in K-562 Cells. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2006, 596, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Berletch, J.B.; Liu, C.; Love, W.K.; Andrews, L.G.; Katiyar, S.K.; Tollefsbol, T.O. Epigenetic and Genetic Mechanisms Contribute to Telomerase Inhibition by EGCG. J Cell Biochem 2008, 103, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi-Pirbaluti, M.; Pourgheysari, B.; Shirzad, H.; Motaghi, E.; Askarian Dehkordi, N.; Surani, Z.; Shirzad, M.; Beshkar, P.; Pirayesh, A. Effect of Epigallocatechin-3-Gallate (EGCG) on Cell Proliferation Inhibition and Apoptosis Induction in Lymphoblastic Leukemia Cell Line. Journal of HerbMed Pharmacology 2015, 65–68. [Google Scholar]
- Ghasemimehr, N.; Farsinejad, A.; Mirzaee Khalilabadi, R.; Yazdani, Z.; Fatemi, A. The Telomerase Inhibitor MST-312 Synergistically Enhances the Apoptotic Effect of Doxorubicin in Pre-B Acute Lymphoblastic Leukemia Cells. Biomedicine & Pharmacotherapy 2018, 106, 1742–1750. [Google Scholar] [CrossRef]
- Safa, M.; Tavasoli, B.; Manafi, R.; Kiani, F.; Kashiri, M.; Ebrahimi, S.; Kazemi, A. Indole-3-Carbinol Suppresses NF-ΚB Activity and Stimulates the P53 Pathway in Pre-B Acute Lymphoblastic Leukemia Cells. Tumor Biology 2015, 36, 3919–3930. [Google Scholar] [CrossRef]
- Safa, M.; Jafari, L.; Alikarami, F.; Manafi Shabestari, R.; Kazemi, A. Indole-3-Carbinol Induces Apoptosis of Chronic Myelogenous Leukemia Cells through Suppression of STAT5 and Akt Signaling Pathways. Tumor Biology 2017, 39, 101042831770576. [Google Scholar] [CrossRef]
- Moon, D.-O.; Kim, M.-O.; Lee, J.-D.; Choi, Y.H.; Kim, G.-Y. Butein Suppresses C-Myc-Dependent Transcription and Akt-Dependent Phosphorylation of HTERT in Human Leukemia Cells. Cancer Lett 2009, 286, 172–179. [Google Scholar] [CrossRef]
- Sahin, F.; Avci, C.B.; Gunduz, C.; Sezgin, C.; Simsir, I.Y.; Saydam, G. Gossypol Exerts Its Cytotoxic Effect on HL-60 Leukemic Cell Line via Decreasing Activity of Protein Phosphatase 2A and Interacting with Human Telomerase Reverse Transcriptase Activity. Hematology 2010, 15, 144–150. [Google Scholar] [CrossRef]
- Ali, J.H.; Walter, M. Combining old and new concepts in targeting telomerase for cancer therapy: transient, immediate, complete and combinatory attack (TICCA). Cancer Cell Int 2023, 23, 197. [Google Scholar] [CrossRef] [PubMed]
- Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. [Google Scholar] [CrossRef]
- Wolf, G. The Discovery of the Visual Function of Vitamin A. J Nutr 2001, 131, 1647–1650. [Google Scholar] [CrossRef]
- Chang, Q.; Chen, Z.; You, J.; McNutt, M.A.; Zhang, T.; Han, Z.; Zhang, X.; Gong, E.; Gu, J. All-Trans-Retinoic Acid Induces Cell Growth Arrest in a Human Medulloblastoma Cell Line. J Neurooncol 2007, 84, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Berletch, J.B.; Green, J.G.; Pate, M.S.; Andrews, L.G.; Tollefsbol, T.O. Telomerase Inhibition by Retinoids Precedes Cytodifferentiation of Leukemia Cells and May Contribute to Terminal Differentiation. Mol Cancer Ther 2004, 3, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-W.; Gu, J.; Wang, Z.-Y.; Chen, S.-J.; Chen, Z. Mechanisms Ofall-Trans Retinoic Acid-Induced Differentiation of Acute Promyelocytic Leukemia Cells. J Biosci 2000, 25, 275–284. [Google Scholar] [CrossRef]
- Love, W.K.; Tyson DeAngelis, J.; Berletch, J.B.; Phipps, S.M.O.; Andrews, L.G.; Brouillette, W.J.; Muccio, D.D.; Tollefsbol, T.O. The Novel Retinoid, 9cUAB30, Inhibits Telomerase and Induces Apoptosis in HL60 Cells. Transl Oncol 2008, 1, 148–152. [Google Scholar] [CrossRef]
- van Gils, N.; Verhagen, H.J.M.P.; Rutten, A.; Menezes, R.X.; Tsui, M.L.; Vermue, E.; Dekens, E.; Brocco, F.; Denkers, F.; Kessler, F.L.; et al. IGFBP7 Activates Retinoid Acid-Induced Responses in Acute Myeloid Leukemia Stem and Progenitor Cells. Blood Adv 2020, 4, 6368–6383. [Google Scholar] [CrossRef]
- de Thé, H. Differentiation Therapy Revisited. Nat Rev Cancer 2018, 2, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Kantarjian, H.; Ravandi, F. Acute Promyelocytic Leukemia Current Treatment Algorithms. Blood Cancer J 2021, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Masetti, R.; Vendemini, F.; Zama, D.; Biagi, C.; Gasperini, P.; Pession, A. All-Trans Retinoic Acid in the Treatment of Pediatric Acute Promyelocytic Leukemia. Expert Rev Anticancer Ther 2012, 12, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Kutny, M.A.; Todd, ; Alonzo, A.; Abla, O.; Et Al Key Points, ; Assessment of Arsenic Trioxide and All-Trans Retinoic Acid for the Treatment of Pediatric Acute Promyelocytic Leukemia A Report From the Children’s Oncology Group AAML1331 Trial; 2021.
- Farinello, D.; Wozińska, M.; Lenti, E.; Genovese, L.; Bianchessi, S.; Migliori, E.; Sacchetti, N.; di Lillo, A.; Bertilaccio, M.T.S.; de Lalla, C.; et al. A Retinoic Acid-Dependent Stroma-Leukemia Crosstalk Promotes Chronic Lymphocytic Leukemia Progression. Nat Commun 2018, 9, 1787. [Google Scholar] [CrossRef] [PubMed]
- Luong, Q.T.; Koeffler, H.P. Vitamin D Compounds in Leukemia. J Steroid Biochem Mol Biol 2005, 97, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kulling, P.M.; Olson, K.C.; Olson, T.L.; Feith, D.J.; Loughran, T.P. Vitamin D in Hematological Disorders and Malignancies. Eur J Haematol 2017, 98, 187–197. [Google Scholar] [CrossRef]
- Holick, M.F. Vitamin D: Importance in the Prevention of Cancers, Type 1 Diabetes, Heart Disease, and Osteoporosis. Am J Clin Nutr 2004, 79, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol Rev 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Trump, D.L.; Deeb, K.K.; Johnson, C.S. Vitamin D: Considerations in the Continued Development as an Agent for Cancer Prevention and Therapy. Cancer J 2010, 16, 1–9. [Google Scholar] [CrossRef]
- Kasiappan, R.; Shen, Z.; Tse, A.K.-W.; Jinwal, U.; Tang, J.; Lungchukiet, P.; Sun, Y.; Kruk, P.; Nicosia, S. v.; Zhang, X.; et al. 1,25-Dihydroxyvitamin D3 Suppresses Telomerase Expression and Human Cancer Growth through MicroRNA-498. Journal of Biological Chemistry 2012, 287, 41297–41309. [Google Scholar] [CrossRef]
- James, S.; Williams, M.; Newland, A.; Colston, K. Leukemia Cell Differentiation: Cellular and Molecular Interactions of Retinoids and Vitamin D. General Pharmacology: The Vascular System 1999, 32, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Hisatake, J.; Kubota, T.; Hisatake, Y.; Uskokovic, M.; Tomoyasu, S.; Koeffler, H.P. 5,6-Trans-16-Ene-Vitamin D3: A New Class of Potent Inhibitors of Proliferation of Prostate, Breast, and Myeloid Leukemic Cells. Cancer Res 1999, 59, 4023–4029. [Google Scholar] [PubMed]
- Delvin, E.; Alos, N.; Rauch, F.; Marcil, V.; Morel, S.; Boisvert, M.; Lecours, M.-A.; Laverdière, C.; Sinnett, D.; Krajinovic, M.; et al. Vitamin D Nutritional Status and Bone Turnover Markers in Childhood Acute Lymphoblastic Leukemia Survivors: A PETALE Study. Clinical Nutrition 2019, 38, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Xu, Y.; de Necochea-Campion, R.; Baylink, D.J.; Payne, K.J.; Tang, X.; Ratanatharathorn, C.; Ji, Y.; Mirshahidi, S.; Chen, C.-S. Application of Vitamin D and Vitamin D Analogs in Acute Myelogenous Leukemia. Exp Hematol 2017, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Muindi, J.R.; Tan, W.; Hu, Q.; Wang, D.; Liu, S.; Wilding, G.E.; Ford, L.A.; Sait, S.N.J.; Block, A.W.; et al. Low 25(OH) Vitamin D3 Levels Are Associated with Adverse Outcome in Newly Diagnosed, Intensively Treated Adult Acute Myeloid Leukemia. Cancer 2014, 120, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Thomas, X.; Chelghoum, Y.; Fanari, N.; Cannas, G. Serum 25-Hydroxyvitamin D Levels Are Associated with Prognosis in Hematological Malignancies. Hematology 2011, 16, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Shanafelt, T.D.; Drake, M.T.; Maurer, M.J.; Allmer, C.; Rabe, K.G.; Slager, S.L.; Weiner, G.J.; Call, T.G.; Link, B.K.; Zent, C.S.; et al. Vitamin D Insufficiency and Prognosis in Chronic Lymphocytic Leukemia. Blood 2011, 117, 1492–1498. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienols: Vitamin E beyond tocopherols. Life Sci 2006, 18, 2088–2098. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the Vitamin E of the 21st Century: Its Potential against Cancer and Other Chronic Diseases. Biochem Pharmacol 2010, 80, 1613–1631. [Google Scholar] [CrossRef]
- Traber, M.G. Vitamin E, Nuclear Receptors and Xenobiotic Metabolism. Arch Biochem Biophys 2004, 423, 6–11. [Google Scholar] [CrossRef]
- Ghanem; Zouein; Mohamad; Hodroj; Haykal; Abou Najem; Naim; Rizk The Vitamin E Derivative Gamma Tocotrienol Promotes Anti-Tumor Effects in Acute Myeloid Leukemia Cell Lines. Nutrients 2019, 11, 2808. [CrossRef] [PubMed]
- Shvachko, L.P.; Zavelevich, M.P.; Gluzman, D.F.; Kravchuk, I. v; Telegeev, G.D. Vitamin Е Activates Expression of С/EBP Alpha Transcription Factor and G-CSF Receptor in Leukemic K562 Cells. Exp Oncol 2018, 40, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, K.N.; Sheikha, A.; Thanoon, I.A.-J.; Hussein, F.N. Effect of Vitamin E on Chemotherapy Induced Oxidative Stress and Immunoglobulin Levels in Patients with Acute Myeloid Leukemia. Tikrit Medical Journal 2008, 14, 146–151. [Google Scholar]
- Telegeev, G.D.; Gluzman, D.F.; Zavelevich, M.P.; Shvachko, L.P. Aberrant Expression of Placental-like Alkaline Phosphatase in Chronic Myeloid Leukemia Cells in Vitro and Its Modulation by Vitamin E. Exp Oncol 2020, 42. [Google Scholar] [CrossRef]
- Hall, K.T.; Buring, J.E.; Mukamal, K.J.; Vinayaga Moorthy, M.; Wayne, P.M.; Kaptchuk, T.J.; Battinelli, E.M.; Ridker, P.M.; Sesso, H.D.; Weinstein, S.J.; et al. COMT and Alpha-Tocopherol Effects in Cancer Prevention: Gene-Supplement Interactions in Two Randomized Clinical Trials. JNCI: Journal of the National Cancer Institute 2019, 111, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.; Theobald, H.E. The Health Effects of Dietary Unsaturated Fatty Acids. Nutr Bull 2006, 31, 178–224. [Google Scholar] [CrossRef]
- Eitsuka, T.; Nakagawa, K.; Suzuki, T.; Miyazawa, T. Polyunsaturated Fatty Acids Inhibit Telomerase Activity in DLD-1 Human Colorectal Adenocarcinoma Cells: A Dual Mechanism Approach. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2005, 1737, 1–10. [Google Scholar] [CrossRef]
- Liu, W.H.; Hsu, C.Y.; Yung, B.Y.M. Nucleophosmin/B23 Regulates the Susceptibility of Human Leukemia HL-60 Cells to Sodium Butyrate-Induced Apoptosis and Inhibition of Telomerase Activity. Int J Cancer 1999, 83, 765–771. [Google Scholar] [CrossRef]
- Nie, D.; Huang, K.; Yin, S.; Li, Y.; Xie, S.; Ma, L.; Wang, X.; Wu, Y.; Xiao, J. Synergistic/Additive Interaction of Valproic Acid with Bortezomib on Proliferation and Apoptosis of Acute Myeloid Leukemia Cells. Leuk Lymphoma 2012, 53, 2487–2495. [Google Scholar] [CrossRef]
- Li, Y.-Q.; Yin, S.-M.; Feng, S.-Q.; Nie, D.-N.; Xie, S.-F.; Ma, L.-P.; Wang, X.-J.; Wu, Y.-D. Effect of Valproic Acid on Apoptosis of Leukemia HL-60 Cells and Expression of h-Tert Gene. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2010, 18, 1445–1450. [Google Scholar]
- Ji, Z.-N.; Ye, W.-C.; Liu, G.-G.; Hsiao, W.L.W. 23-Hydroxybetulinic Acid-Mediated Apoptosis Is Accompanied by Decreases in Bcl-2 Expression and Telomerase Activity in HL-60 Cells. Life Sci 2002, 72, 1–9. [Google Scholar] [CrossRef]
- ODA, M.; UENO, T.; KASAI, N.; TAKAHASHI, H.; YOSHIDA, H.; SUGAWARA, F.; SAKAGUCHI, K.; HAYASHI, H.; MIZUSHINA, Y. Inhibition of Telomerase by Linear-Chain Fatty Acids: A Structural Analysis. Biochemical Journal 2002, 367, 329–334. [Google Scholar] [CrossRef]
- Choi, Y.H. Apoptosis of U937 Human Leukemic Cells by Sodium Butyrate Is Associated with Inhibition of Telomerase Activity. Int J Oncol 2006, 29, 1207–1213. [Google Scholar] [CrossRef] [PubMed]
- Fahrmann, J.F.; Ballester, O.F.; Ballester, G.; Witte, T.R.; Salazar, A.J.; Kordusky, B.; Cowen, K.G.; Ion, G.; Primerano, D.A.; Boskovic, G.; et al. Inhibition of Nuclear Factor Kappa B Activation in Early-Stage Chronic Lymphocytic Leukemia by Omega-3 Fatty Acids. Cancer Invest 2013, 31, 24–38. [Google Scholar] [CrossRef] [PubMed]
- el Amrousy, D.; El-Afify, D.; Khedr, R.; Ibrahim, A.M. Omega 3 Fatty Acids Can Reduce Early Doxorubicin-induced Cardiotoxicity in Children with Acute Lymphoblastic Leukemia. Pediatr Blood Cancer 2022, 69. [Google Scholar] [CrossRef]
- Bükki, J.; Stanga, Z.; Tellez, F.B.; Duclos, K.; Kolev, M.; Krähenmann, P.; Pabst, T.; Iff, S.; Jüni, P. Omega-3 Poly-Unsaturated Fatty Acids for the Prevention of Severe Neutropenic Enterocolitis in Patients with Acute Myeloid Leukemia. Nutr Cancer 2013, 65, 834–842. [Google Scholar] [CrossRef]
- Chagas, T.R.; Borges, D.S.; de Oliveira, P.F.; Mocellin, M.C.; Barbosa, A.M.; Camargo, C.Q.; del Moral, J.Â.G.; Poli, A.; Calder, P.C.; Trindade, E.B.S.M.; et al. Oral Fish Oil Positively Influences Nutritional-Inflammatory Risk in Patients with Haematological Malignancies during Chemotherapy with an Impact on Long-Term Survival: A Randomised Clinical Trial. Journal of Human Nutrition and Dietetics 2017, 30, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Rücker, F.G.; Lang, K.M.; Fütterer, M.; Komarica, V.; Schmid, M.; Döhner, H.; Schlenk, R.F.; Döhner, K.; Knudsen, S.; Bullinger, L. Molecular Dissection of Valproic Acid Effects in Acute Myeloid Leukemia Identifies Predictive Networks. Epigenetics 2016, 11, 517–525. [Google Scholar] [CrossRef]
- Corsetti, M.T.; Salvi, F.; Perticone, S.; Baraldi, A.; de Paoli, L.; Gatto, S.; Pietrasanta, D.; Pini, M.; Primon, V.; Zallio, F.; et al. Hematologic Improvement and Response in Elderly AML/RAEB Patients Treated with Valproic Acid and Low-Dose Ara-C. Leuk Res 2011, 35, 991–997. [Google Scholar] [CrossRef]
- Eitsuka, T.; Nakagawa, K.; Igarashi, M.; Miyazawa, T. Telomerase Inhibition by Sulfoquinovosyldiacylglycerol from Edible Purple Laver (Porphyra Yezoensis). Cancer Lett 2004, 212, 15–20. [Google Scholar] [CrossRef]
- Jain, C.K.; Pradhan, B.S.; Banerjee, S.; Mondal, N.B.; Majumder, S.S.; Bhattacharyya, M.; Chakrabarti, S.; Roychoudhury, S.; Majumder, H.K. Sulfonoquinovosyl Diacylglyceride Selectively Targets Acute Lymphoblastic Leukemia Cells and Exerts Potent Anti-Leukemic Effects in Vivo. Sci Rep 2015, 5, 12082. [Google Scholar] [CrossRef]
- Dart, D.A.; Adams, K.E.; Akerman, I.; Lakin, N.D. Recruitment of the Cell Cycle Checkpoint Kinase ATR to Chromatin during S-Phase. Journal of Biological Chemistry 2004, 279, 16433–16440. [Google Scholar] [CrossRef]
- Shang, H.-S.; Shih, Y.-L.; Lee, C.-H.; Hsueh, S.-C.; Liu, J.-Y.; Liao, N.-C.; Chen, Y.-L.; Huang, Y.-P.; Lu, H.-F.; Chung, J.-G. Sulforaphane-Induced Apoptosis in Human Leukemia HL-60 Cells through Extrinsic and Intrinsic Signal Pathways and Altering Associated Genes Expression Assayed by CDNA Microarray. Environ Toxicol 2017, 32, 311–328. [Google Scholar] [CrossRef]
- Tang, Y.-L.; Sun, X.; Huang, L.-B.; Liu, X.-J.; Qin, G.; Wang, L.-N.; Zhang, X.-L.; Ke, Z.-Y.; Luo, J.-S.; Liang, C.; et al. Melatonin Inhibits MLL-Rearranged Leukemia via RBFOX3/HTERT and NF-ΚB/COX-2 Signaling Pathways. Cancer Lett 2019, 443, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.-Y.; Xu, D.-P.; Li, H.-B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. [Google Scholar] [CrossRef]
- Veisi, A.; Akbari, G.; Mard, S.A.; Badfar, G.; Zarezade, V.; Mirshekar, M.A. Role of Crocin in Several Cancer Cell Lines: An Updated Review. Iran J Basic Med Sci 2020, 23, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, Z.; Wang, L.; Wang, L.-Z.; Zang, C.; Sun, L.-R. The Effect and Mechanisms of Proliferative Inhibition of Crocin on Human Leukaemia Jurkat Cells. West Indian Medical Journal 2016. [Google Scholar] [CrossRef]
- Oh, J.H.; Lee, T.-J.; Kim, S.H.; Choi, Y.H.; Lee, S.H.; Lee, J.M.; Kim, Y.-H.; Park, J.-W.; Kwon, T.K. Induction of Apoptosis by Withaferin A in Human Leukemia U937 Cells through Down-Regulation of Akt Phosphorylation. Apoptosis 2008, 13, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Katiyar, S.P.; Sundar, D.; Kaul, Z.; Miyako, E.; Zhang, Z.; Kaul, S.C.; Reddel, R.R.; Wadhwa, R. Withaferin-A Kills Cancer Cells with and without Telomerase: Chemical, Computational and Experimental Evidences. Cell Death Dis 2017, 8, e2755–e2755. [Google Scholar] [CrossRef]
- Khathayer, F.; Ray, S.K. Diosgenin as a Novel Alternative Therapy for Inhibition of Growth, Invasion, and Angiogenesis Abilities of Different Glioblastoma Cell Lines. Neurochem Res 2020, 45, 2336–2351. [Google Scholar] [CrossRef]
- Semwal, P.; Painuli, S.; Abu-Izneid, T.; Rauf, A.; Sharma, A.; Daştan, S.D.; Kumar, M.; Alshehri, M.M.; Taheri, Y.; Das, R.; et al. Diosgenin: An Updated Pharmacological Review and Therapeutic Perspectives. Oxid Med Cell Longev 2022, 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Fan, J.; Wang, Q.; Ju, D.; Feng, M.; Li, J.; Guan, Z.; An, D.; Wang, X.; Ye, L. Diosgenin Induces ROS-Dependent Autophagy and Cytotoxicity via MTOR Signaling Pathway in Chronic Myeloid Leukemia Cells. Phytomedicine 2016, 23, 243–252. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.; Wang, B.; Zhang, N. A review on analytical methods for natural berberine alkaloids. J Sep Sci 2019, 9, 1794–1815. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Hao, J.; Fan, D. Biological Properties and Clinical Applications of Berberine. Front Med 2020, 14, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.L.; Hsu, C.Y.; Liu, W.H.; Yung, B.Y.M. Berberine-Induced Apoptosis of Human Leukemia HL-60 Cells Is Associated with down-Regulation of Nucleophosmin/B23 and Telomerase Activity. Int J Cancer 1999, 81, 923–929. [Google Scholar] [CrossRef]
- Franceschin, M.; Rossetti, L.; D’Ambrosio, A.; Schirripa, S.; Bianco, A.; Ortaggi, G.; Savino, M.; Schultes, C.; Neidle, S. Natural and Synthetic G-Quadruplex Interactive Berberine Derivatives. Bioorg Med Chem Lett 2006, 16, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int J Mol Sci 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Jayasooriya, R.G.P.T.; Kang, S.H.; Kang, C.H.; Choi, Y.H.; Moon, D.O.; Hyun, J.W.; Chang, W.Y.; Kim, G.Y. Apigenin Decreases Cell Viability and Telomerase Activity in Human Leukemia Cell Lines. Food and Chemical Toxicology 2012, 50, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for Health and Wellbeing: A Potent Bioactive Metabolite of an Entomopathogenic Medicinal Fungus Cordyceps with Its Nutraceutical and Therapeutic Potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef]
- Jang, K.J.; Kwon, G.S.; Jeong, J.W.; Kim, C.H.; Yoon, H.M.; Kim, G.Y.; Shim, J.H.; Moon, S.K.; Kim, W.J.; Choi, Y.H. Cordyceptin Induces Apoptosis through Repressing HTERT Expression and Inducing Extranuclear Export of HTERT. J Biosci Bioeng 2015, 119, 351–357. [Google Scholar] [CrossRef]
- Ansari, M.A.; Khan, F.B.; Safdari, H.A.; Almatroudi, A.; Alzohairy, M.A.; Safdari, M.; Amirizadeh, M.; Rehman, S.; Equbal, M.J.; Hoque, M. Prospective Therapeutic Potential of Tanshinone IIA: An Updated Overview. Pharmacol Res 2021, 164, 105364. [Google Scholar] [CrossRef] [PubMed]
- Chiu, T.L.; Su, C.C. Tanshinone IIA induces apoptosis in human lung cancer A549 cells through the induction of reactive oxygen species and decreasing the mitochondrial membrane potential. Int J Mol Med 2010, 25, 231–236. [Google Scholar] [PubMed]
- Huang, X.; Jin, L.; Deng, H.; Wu, D.; Shen, Q. K.; Quan, Z. S.; Zhang, C. H.; & Guo, H. Y. (2022). Research and Development of Natural Product Tanshinone I: Pharmacology, Total Synthesis, and Structure Modifications. Front Pharmacol 2022, 13, 920411. [Google Scholar] [CrossRef]
- Liu, X.D.; Fan, R.F.; Zhang, Y.; Yang, H.Z.; Fang, Z.G.; Guan, W.B.; Lin, D.J.; Xiao, R.Z.; Huang, R.W.; Huang, H.Q.; et al. Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro. Int J Mol Sci 2010, 11, 2267–2280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Wang, S. The Role of Rapamycin in Healthspan Extension via the Delay of Organ Aging. Ageing Res Rev 2021, 70, 101376. [Google Scholar] [CrossRef]
- Vézina, C.; Kudelski, A.; Sehgal, S.N. Rapamycin (AY-22,989), a new antifungal antibiotic. I. Taxonomy of the producing streptomycete and isolation of the active principle. J Antibiot (Tokyo) 1975, 10, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.M.; Zhou, Q.; Xu, Y.; Lai, X.Y.; Huang, H. Antiproliferative Effect of Rapamycin on Human T-Cell Leukemia Cell Line Jurkat by Cell Cycle Arrest and Telomerase Inhibition. Acta Pharmacol Sin 2008, 29, 481–488. [Google Scholar] [CrossRef]
- Tomizawa, A.; Kanno, S.I.; Osanai, Y.; Yomogida, S.; Ishikawa, M. Cytotoxic effects of caffeic acid undecyl ester are involved in the inhibition of telomerase activity in NALM-6 human B-cell leukemia cells. Oncol Lett 2013, 4, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, A.; Kanno, S.I.; Osanai, Y.; Goto, A.; Sato, C.; Yomogida, S.; Ishikawa, M. Induction of apoptosis by a potent caffeic acid derivative, caffeic acid undecyl ester, is mediated by mitochondrial damage in NALM-6 human B cell leukemia cells. Oncol Rep 2013, 2, 425–429. [Google Scholar] [CrossRef]
- Feriotto, G.; Tagliati, F.; Giriolo, R.; Casciano, F.; Tabolacci, C.; Beninati, S.; Khan, M.T.H.; Mischiati, C. Caffeic Acid Enhances the Anti-Leukemic Effect of Imatinib on Chronic Myeloid Leukemia Cells and Triggers Apoptosis in Cells Sensitive and Resistant to Imatinib. Int. J. Mol. Sci. 2021, 22, 1644. [Google Scholar] [CrossRef]
- Gomes, C.L.; de Albuquerque Wanderley Sales, V.; Gomes de Melo, C.; Ferreira da Silva, R.M.; Vicente Nishimura, R.H.; Rolim, L.A.; Rolim Neto, P.J. Beta-Lapachone: Natural Occurrence, Physicochemical Properties, Biological Activities, Toxicity and Synthesis. Phytochemistry 2021, 186, 112713. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O.; Kang, C.-H.; Kim, M.-O.; Jeon, Y.-J.; Lee, J.-D.; Choi, Y.H.; Kim, G.-Y. β-Lapachone (LAPA) Decreases Cell Viability and Telomerase Activity in Leukemia Cells: Suppression of Telomerase Activity by LAPA. J Med Food 2010, 13, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wang, X.; Zhou, Y.; Wang, Y.; Ding, J. Ginsenoside Rg1 Induces Senescence of Leukemic Stem Cells by Upregulating P16INK4a and Downregulating HTERT Expression. Advances in Clinical and Experimental Medicine 2021, 30, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Park, C.; Kim, S.H.; Hossain, M.A.; Kim, M.Y.; Chung, H.Y.; Son, W.S.; Kim, G.-Y.; Choi, Y.H.; Kim, N.D. Korean Red Ginseng Extract Induces Apoptosis and Decreases Telomerase Activity in Human Leukemia Cells. J Ethnopharmacol 2009, 121, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chu, S.; Zhang, Z.; Chen, N. Hepataprotective Effects of Ginsenoside Rg1 – A Review. J Ethnopharmacol 2017, 206, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, X.; Liu, X.; Zhang, C.; Shang, W.; Xue, J.; Chen, R.; Xing, Y.; Song, D.; Xu, R. Ginsenoside Rg1 Promotes Cerebral Angiogenesis via the PI3K/Akt/MTOR Signaling Pathway in Ischemic Mice. Eur J Pharmacol 2019, 856, 172418. [Google Scholar] [CrossRef]
- Hou, T.; Liu, Y.; Wang, X.; Jiao, D.; Xu, H.; Shi, Q.; Wang, Y.; Li, W.; Wu, T.; Liang, Q. Ginsenoside Rg1 Promotes Lymphatic Drainage and Improves Chronic Inflammatory Arthritis. J Musculoskelet Neuronal Interact 2020, 20, 526–534. [Google Scholar] [PubMed]
- Zhang, Z.; Song, Z.; Shen, F.; Xie, P.; Wang, J.; Zhu, A.; Zhu, G. Ginsenoside Rg1 Prevents PTSD-Like Behaviors in Mice Through Promoting Synaptic Proteins, Reducing Kir4.1 and TNF-α in the Hippocampus. Mol Neurobiol 2021, 58, 1550–1563. [Google Scholar] [CrossRef]
- das Gupta, S.; Halder, B.; Gomes, A.; Gomes, A. Bengalin Initiates Autophagic Cell Death through ERK–MAPK Pathway Following Suppression of Apoptosis in Human Leukemic U937 Cells. Life Sci 2013, 93, 271–276. [Google Scholar] [CrossRef]
- Haldar, S.; das Gupta, S.; Gomes, A.; Giri, B.; Dasgupta, S.C.; Biswas, A.; Mishra, R.; Gomes, A. A High Molecular Weight Protein Bengalin from the Indian Black Scorpion (Heterometrus Bengalensis C.L. Koch) Venom Having Antiosteoporosis Activity in Female Albino Rats. Toxicon 2010, 55, 455–461. [Google Scholar] [CrossRef]
- Gupta, S. das; Gomes, A.; Debnath, A.; Saha, A.; Gomes, A. Apoptosis Induction in Human Leukemic Cells by a Novel Protein Bengalin, Isolated from Indian Black Scorpion Venom: Through Mitochondrial Pathway and Inhibition of Heat Shock Proteins. Chem Biol Interact 2010, 183, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press: Washington, D.C., 2000; Vol. chapter 3, ISBN 978-0-309-06935-9.
- Travica, N.; Ried, K.; Sali, A.; Scholey, A.; Hudson, I.; Pipingas, A. Vitamin C Status and Cognitive Function: A Systematic Review. Nutrients 2017, 9, 960. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guo, C.; Li, Y.; Liang, X.; Su, M. Functional Benefit and Molecular Mechanism of Vitamin C against Perfluorooctanesulfonate-Associated Leukemia. Chemosphere 2021, 263. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-J.; Zou, X.-L.; Zhao, Z.-Y.; Wang, J.; Yang, Z.; Ni, X.; Wei, J. Effect of High Dose Vitamin C on Proliferation and Apoptosis of Acute Myeloid Leukemia Cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2020, 28, 833–841. [Google Scholar]
- Shin-ya, K. Telomerase Inhibitor, Telomestatin, a Specific Mechanism to Interact with Telomere Structure. Nihon Rinsho 2004, 62, 1277–1282. [Google Scholar] [PubMed]
- Tauchi, T.; Shin-ya, K.; Sashida, G.; Sumi, M.; Okabe, S.; Ohyashiki, J.H.; Ohyashiki, K. Telomerase Inhibition with a Novel G-Quadruplex-Interactive Agent, Telomestatin: In Vitro and in Vivo Studies in Acute Leukemia. Oncogene 2006, 25, 5719–5725. [Google Scholar] [CrossRef]
- Chambers, C.S.; Holečková, V.; Petrásková, L.; Biedermann, D.; Valentová, K.; Buchta, M.; Křen, V. The Silymarin Composition… and Why Does It Matter??? Food Research International 2017, 100, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Faezizadeh, Z.; Mesbah-Namin, S.A.; Allameh, A. The effect of silymarin on telomerase activity in the human leukemia cell line K562. Planta Med 2012, 9, 899–902. [Google Scholar] [CrossRef]
- Lu, S.; Qin, X.; Yuan, S.; Li, Y.; Wang, L.; Jin, Y.; Zeng, G.; Yen, L.; Hu, J.; Dang, T.; et al. Effect of Tianshengyuan-1 (TSY-1) on Telomerase Activity and Hematopoietic Recovery - in Vitro, Ex Vivo, and in Vivo Studies. Int J Clin Exp Med 2014, 7, 597–606. [Google Scholar]
- Yu, W.; Qin, X.; Jin, Y.; Li, Y.; Santiskulvong, C.; Vu, V.; Zeng, G.; Zhang, Z.; Chow, M.; Rao, J. Tianshengyuan-1 (TSY-1) Regulates Cellular Telomerase Activity by Methylation of TERT Promoter. Oncotarget 2017, 8, 7977–7988. [Google Scholar] [CrossRef] [PubMed]
- Huynh, D.L.; Ngau, T.H.; Nguyen, N.H.; Tran, G.-B.; Nguyen, C.T. Potential Therapeutic and Pharmacological Effects of Wogonin: An Updated Review. Mol Biol Rep 2020, 47, 9779–9789. [Google Scholar] [CrossRef]
- Khan, N.M.; Haseeb, A.; Ansari, M.Y.; Devarapalli, P.; Haynie, S.; Haqqi, T.M. Wogonin, a Plant Derived Small Molecule, Exerts Potent Anti-Inflammatory and Chondroprotective Effects through the Activation of ROS/ERK/Nrf2 Signaling Pathways in Human Osteoarthritis Chondrocytes. Free Radic Biol Med 2017, 106, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Wang, C.Y.; Yang, R.C.; Chu, C.J.; Wu, H.T.; Pang, J.H.S. Wogonin, an Active Compound in Scutellaria Baicalensis, Induces Apoptosis and Reduces Telomerase Activity in the HL-60 Leukemia Cells. Phytomedicine 2010, 17, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Kiladjian, J.-J.; Giraudier, S.; Cassinat, B. Interferon-Alpha for the Therapy of Myeloproliferative Neoplasms: Targeting the Malignant Clone. Leukemia 2016, 30, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Simonsson, B.; Hjorth-Hansen, H.; Weis Bjerrum, O.; Porkka, K. Interferon Alpha for Treatment of Chronic Myeloid Leukemia. Curr Drug Targets 2011, 12, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liu, X.-H.; Kong, J.; Wang, J.; Jia, J.-S.; Lu, S.-Y.; Gong, L.-Z.; Zhao, X.-S.; Jiang, Q.; Chang, Y.-J.; et al. Interferon-α as Maintenance Therapy Can Significantly Reduce Relapse in Patients with Favorable-Risk Acute Myeloid Leukemia. Leuk Lymphoma 2021, 62, 2949–2956. [Google Scholar] [CrossRef] [PubMed]
- Magenau, J.M.; Peltier, D.; Riwes, M.; Pawarode, A.; Parkin, B.; Braun, T.; Anand, S.; Ghosh, M.; Maciejewski, J.; Yanik, G.; et al. Type 1 Interferon to Prevent Leukemia Relapse after Allogeneic Transplantation. Blood Adv 2021, 5, 5047–5056. [Google Scholar] [CrossRef] [PubMed]
- Nai, J.; Zhang, C.; Shao, H.; Li, B.; Li, H.; Gao, L.; Dai, M.; Zhu, L.; Sheng, H. Extraction, Structure, Pharmacological Activities and Drug Carrier Applications of Angelica Sinensis Polysaccharide. Int J Biol Macromol 2021, 183, 2337–2353. [Google Scholar] [CrossRef]
- Liu, J.; Xu, C.Y.; Cai, S.Z.; Zhou, Y.; Li, J.; Jiang, R.; Wang, Y.P. Senescence Effects of Angelica Sinensis Polysaccharides on Human Acute Myelogenous Leukemia Stem and Progenitor Cells. Asian Pacific Journal of Cancer Prevention 2013, 14, 6549–6556. [Google Scholar] [CrossRef]
- Hetland, G.; Tangen, J.-M.; Mahmood, F.; Mirlashari, M.R.; Nissen-Meyer, L.S.H.; Nentwich, I.; Therkelsen, S.P.; Tjønnfjord, G.E.; Johnson, E. Antitumor, Anti-Inflammatory and Antiallergic Effects of Agaricus Blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium Erinaceus and Grifola Frondosa: A Review of Preclinical and Clinical Studies. Nutrients 2020, 12, 1339. [Google Scholar] [CrossRef]
- Al-Dbass, A.M.; Al- Daihan, S.K.; Bhat, R.S. Agaricus Blazei Murill as an Efficient Hepatoprotective and Antioxidant Agent against CCl4-Induced Liver Injury in Rats. Saudi J Biol Sci 2012, 19, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tian, N.; Wang, H.; Yan, H. Chemically Sulfated Polysaccharides from Agaricus Blazei Murill: Synthesis, Characterization and Anti-HIV Activity. Chem Biodivers 2021, 18. [Google Scholar] [CrossRef]
- Gao, L.; Sun, Y.; Chen, C.; Xi, Y.; Wang, J.; Wang, Z. Primary mechanism of apoptosis induction in a leukemia cell line by fraction FA-2-b-ss prepared from the mushroom Agaricus blazei Murill. Braz J Med Biol Res 2007, 40, 1545–1555. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.O.; Moon, D.O.; Choi, Y.H.; Lee, J.D.; Kim, N.D.; Kim, G.Y. Platycodin D induces mitotic arrest in vitro, leading to endoreduplication, inhibition of proliferation and apoptosis in leukemia cells. Int J Cancer 2008, 122, 2674–2681. [Google Scholar] [CrossRef]
- Kim, M.O.; Moon, D.O.; Choi, Y.H.; Shin, D.Y.; Kang, H.S.; Choi, B.T.; Lee, J.D.; Li, W.; Kim, G.Y. Platycodin D Induces Apoptosis and Decreases Telomerase Activity in Human Leukemia Cells. Cancer Lett 2008, 261, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.O.; Moon, D.O.; Kang, S.H.; Heo, M.S.; Choi, Y.H.; Jung, J.H.; Lee, J.D.; Kim, G.Y. Pectenotoxin-2 Represses Telomerase Activity in Human Leukemia Cells through Suppression of HTERT Gene Expression and Akt-Dependent HTERT Phosphorylation. FEBS Lett 2008, 582, 3263–3269. [Google Scholar] [CrossRef]
- Khaiwa, N.; Maarouf, N.R.; Darwish, M.H.; Alhamad, D.W.M.; Sebastian, A.; Hamad, M.; Omar, H.A.; Orive, G.; Al-Tel, T.H. Camptothecin’s Journey from Discovery to WHO Essential Medicine: Fifty Years of Promise. Eur J Med Chem 2021, 223, 113639. [Google Scholar] [CrossRef]
- Lazareva, N.F.; Baryshok, V.P.; Lazarev, I.M. Silicon-Containing Analogs of Camptothecin as Anticancer Agents. Arch Pharm (Weinheim) 2018, 351, 1700297. [Google Scholar] [CrossRef]
- Jiang, J.F.; Liu, W.J.; Ding, J. Regulation of Telomerase Activity in Camptothecin-Induced Apoptosis of Human Leukemia HL-60 Cells. Acta Pharmacol Sin 2000, 21, 759–764. [Google Scholar]
- Bao, J.; Dai, S.-M. A Chinese Herb Tripterygium Wilfordii Hook F in the Treatment of Rheumatoid Arthritis: Mechanism, Efficacy, and Safety. Rheumatol Int 2011, 31, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Qin, W.; Zheng, H.; Schegg, K.; Han, L.; Liu, X.; Wang, Y.; Wang, Z.; McSwiggin, H.; Peng, H.; et al. Triptonide Is a Reversible Non-Hormonal Male Contraceptive Agent in Mice and Non-Human Primates. Nat Commun 2021, 12, 1253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Tan, S.; Yu, D.; Zhao, Z.; Zhang, B.; Zhang, P.; Lv, C.; Zhou, Q.; Cao, Z. Triptonide Inhibits Lung Cancer Cell Tumorigenicity by Selectively Attenuating the Shh-Gli1 Signaling Pathway. Toxicol Appl Pharmacol 2019, 365, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Meng, M.; Liu, Y.; Qi, J.; Zhao, Z.; Qiao, Y.; Hu, Y.; Lu, W.; Zhou, Z.; Xu, P.; et al. Triptonide Effectively Inhibits Triple-Negative Breast Cancer Metastasis through Concurrent Degradation of Twist1 and Notch1 Oncoproteins. Breast Cancer Research 2021, 23, 116. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, X.; Jin, R.; Xu, F.; Ding, J.; Zhang, L.; Huang, Z. Triptonide Inhibits Metastasis Potential of Thyroid Cancer Cells via Astrocyte Elevated Gene-1. Transl Cancer Res 2020, 9, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Meng, M.; Zheng, N.; Cao, Z.; Yang, P.; Xi, X.; Zhou, Q. Targeting of Multiple Senescence-Promoting Genes and Signaling Pathways by Triptonide Induces Complete Senescence of Acute Myeloid Leukemia Cells. Biochem Pharmacol 2017, 126, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Kitajima, Y.; Kakuta, M.; Osanai, Y.; Kurauchi, K.; Ujibe, M.; Ishikawa, M. Costunolide-Induced Apoptosis Is Caused by Receptor-Mediated Pathway and Inhibition of Telomerase Activity in NALM-6 Cells. Biol Pharm Bull 2008, 31, 1024–1028. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-J.; Jiang, J.-F.; Xiao, D.; Ding, J. Down-Regulation of Telomerase Activity via Protein Phosphatase 2A Activation in Salvicine-Induced Human Leukemia HL-60 Cell Apoptosis. Biochem Pharmacol 2002, 64, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.N.; Ye, W.C.; Rui, J.; Wang, L.; Liu, G.Q. Patensin-Induced Apoptosis Is Accompanied by Decreased Bcl-2 Expression and Telomerase Activity in HL-60 Cells. J Asian Nat Prod Res 2004, 6, 107–114. [Google Scholar] [CrossRef]
- Park, C.; Jung, J.H.; Kim, N.D.; Choi, Y.H. Inhibition of Cyclooxygenase-2 and Telomerase Activities in Human Leukemia Cells by Dideoxypetrosynol A, a Polyacetylene from the Marine Sponge Petrosia Sp. Int J Oncol 2007, 30, 291–298. [Google Scholar] [CrossRef]
- Park, C.; Kim, G.Y.; Kim, W. il; Hong, S.H.; Park, D. il; Kim, N.D.; Bae, S.J.; Jung, J.H.; Choi, Y.H. Induction of Apoptosis by (Z)-Stellettic Acid C, an Acetylenic Acid from the Sponge Stelletta Sp., Is Associated with Inhibition of Telomerase Activity in Human Leukemic U937 Cells. Chemotherapy 2007, 53, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.J.; Lee, S.J.; Choi, B.T.; Park, Y.-M.; Choi, Y.H. Induction of Apoptosis and Inhibition of Telomerase Activity by Trichostatin A, a Histone Deacetylase Inhibitor, in Human Leukemic U937 Cells. Exp Mol Pathol 2007, 82, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-Z.; Lin, M.-F.; Huang, H.; Cai, Z. Homoharringtonine-Induced Apoptosis of Human Leukemia HL-60 Cells Is Associated with Down-Regulation of Telomerase. Am J Chin Med (Gard City N Y) 2006, 34, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, Z.; Tian, J.; Wang, H.; Song, G.; Guo, Q.; Tian, J.; Han, Y.; Liao, Q.; Liu, G.; et al. The Downregulation of C-Myc and Its Target Gene HTERT Is Associated with the Antiproliferative Effects of Baicalin on HL-60 Cells. Oncol Lett 2017. [Google Scholar] [CrossRef] [PubMed]
- Cogulu, O.; Biray, C.; Gunduz, C.; Karaca, E.; Aksoylar, S.; Sorkun, K.; Salih, B.; Ozkinay, F. Effects of Manisa Propolis on Telomerase Activity in Leukemia Cells Obtained from the Bone Marrow of Leukemia Patients. Int J Food Sci Nutr 2009, 60, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Gunduz, C.; Biray, C.; Kosova, B.; Yilmaz, B.; Eroglu, Z.; Şahin, F.; Omay, S.B.; Cogulu, O. Evaluation of Manisa Propolis Effect on Leukemia Cell Line by Telomerase Activity. Leuk Res 2005, 29, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, K.; Yamada, T.; Sumida, T.; Hamakawa, H.; Kuwabara, H.; Matsuda, M.; Muramatsu, Y.; Kose, H.; Matsumoto, K.; Sasaki, Y.; et al. Induction of Apoptosis and Inhibition of DNA Topoisomerase-I in K-562 Cells by a Marine Microalgal Polysaccharide. Life Sci 2000, 66, PL227–PL231. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, K.; Sumida, T.; Hamakawa, H.; Yamada, T.; Matsumoto, K.; Matsuda, M.; Oda, H.; Miyake, H.; Tashiro, S.; Okutani, K. Inhibitory Effect of a Marine Microalgal Polysaccharide on the Telomerase Activity in K562 Cells.
- Li, X.; Zhang, C.-T.; Ma, W.; Xie, X.; Huang, Q. Oridonin: A Review of Its Pharmacology, Pharmacokinetics and Toxicity. Front Pharmacol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-J.; Wu, X.-Y.; Lul, H.-L.; Pan, X.-L.; Peng, J.; Huang, R.-W. Anti-Proliferation Effect of Oridonin on HL-60 Cells and Its Mechanism. Chin Med Sci J 2004, 19, 134–137. [Google Scholar]
- Liu, J.J.; Huang, R.W.; Lin, D.J.; Wu, X.Y.; Peng, J.; Pan, X.L.; Song, Y.Q.; Lin, Q.; Hou, M.; Wang, D.N.; et al. Oridonin-Induced Apoptosis in Leukemia K562 Cells and Its Mechanism. Neoplasma 2005, 52, 225–230. [Google Scholar]
- Liu, J.-J.; Huang, R.-W.; Lin, D.-J.; Wu, X.-Y.; Peng, J.; Pan, X.-L.; Lin, Q.; Hou, M.; Zhang, M.-H.; Chen, F. Antiproliferation Effects of Oridonin on HPB-ALL Cells and Its Mechanisms of Action. Am J Hematol 2006, 81, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Faa, G.; Gerosa, C.; Fanni, D.; Lachowicz, J.I.; Nurchi, V.M. Gold - Old Drug with New Potentials. Curr Med Chem 2018, 25, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Pandit, S.; Mokkapati, V.R.S.S.; Garg, A.; Ravikumar, V.; Mijakovic, I. Gold Nanoparticles in Diagnostics and Therapeutics for Human Cancer. Int J Mol Sci 2018, 19, 1979. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.E.; Mwamuka, J.; Gole, A.; Murphy, C.J.; Wyatt, M.D. Gold Nanoparticles Are Taken Up by Human Cells but Do Not Cause Acute Cytotoxicity. Small 2005, 1, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chinnathambi, A.; Nasif, O.; Alharbi, S.A. Green Synthesis and Chemical Characterization of a Novel Anti-Human Pancreatic Cancer Supplement by Silver Nanoparticles Containing Zingiber Officinale Leaf Aqueous Extract. Arabian Journal of Chemistry 2021, 14, 103081. [Google Scholar] [CrossRef]
- Phenix, C.P.; Togtema, M.; Pichardo, S.; Zehbe, I.; Curiel, L. High Intensity Focused Ultrasound Technology, Its Scope and Applications in Therapy and Drug Delivery. J Pharm Pharm Sci 2014, 17, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Mittelstein, D.R.; Ye, J.; Schibber, E.F.; Roychoudhury, A.; Martinez, L.T.; Fekrazad, M.H.; Ortiz, M.; Lee, P.P.; Shapiro, M.G.; Gharib, M. Selective Ablation of Cancer Cells with Low Intensity Pulsed Ultrasound. Appl Phys Lett 2020, 116. [Google Scholar] [CrossRef]
- Zhu, H.-H.; Wu, D.-P.; Jin, J.; Li, J.-Y.; Ma, J.; Wang, J.-X.; Jiang, H.; Chen, S.-J.; Huang, X.-J. Oral Tetra-Arsenic Tetra-Sulfide Formula Versus Intravenous Arsenic Trioxide As First-Line Treatment of Acute Promyelocytic Leukemia: A Multicenter Randomized Controlled Trial. Journal of Clinical Oncology 2013, 31, 4215–4221. [Google Scholar] [CrossRef]
- Testi, A.M. GIMEMA-AIEOPAIDA Protocol for the Treatment of Newly Diagnosed Acute Promyelocytic Leukemia (APL) in Children. Blood 2005, 106, 447–453. [Google Scholar] [CrossRef]
- Seol, J.G.; Kim, E.S.; Park, W.H.; Jung, C.W.; Kim, B.K.; Lee, Y.Y. Telomerase Activity in Acute Myelogenous Leukaemia: Clinical and Biological Implications. Br J Haematol 1998, 100, 156–165. [Google Scholar] [CrossRef]
- Hewison, M.; Barker, S.; Brennan, A.; Nathan, J.; Katz, D.R.; O’Riordan, J.L. Autocrine Regulation of 1,25-Dihydroxycholecalciferol Metabolism in Myelomonocytic Cells. Immunology 1989, 68, 247–252. [Google Scholar] [PubMed]
- Ferrero, D.; Campa, E.; Dellacasa, C.; Campana, S.; Foli, C.; Boccadoro, M. Differentiating Agents + Low-Dose Chemotherapy in the Management of Old/Poor Prognosis Patients with Acute Myeloid Leukemia or Myelodysplastic Syndrome. Haematologica 2004, 89, 619–620. [Google Scholar] [PubMed]
- Paubelle, E.; Zylbersztejn, F.; Alkhaeir, S.; Suarez, F.; Callens, C.; Dussiot, M.; Isnard, F.; Rubio, M.-T.; Damaj, G.; Gorin, N.-C.; et al. Deferasirox and Vitamin D Improves Overall Survival in Elderly Patients with Acute Myeloid Leukemia after Demethylating Agents Failure. PLoS One 2013, 8, e65998. [Google Scholar] [CrossRef] [PubMed]
- Shvachko, L.; Zavelevich, M.; Gluzman, D.; Telegeev, G. (2021) Vitamin E in Chronic Myeloid Leukemia (CML) Prevention. Vitamin E in Health and Disease - Interactions, Diseases and Health Aspects. IntechOpen. [CrossRef]
Figure 1.
Mechanism of telomerase activity at the end of the replication cycle. Column a) During the replication, the lagging strand is created based on the leading strand, that was present in the nucleus. The new strand is complementary to the old one but has its length shortened, which is the reason for cell aging. Elongation of the lagging strand is an extra step that requires telomerase and does not happen in every human cell. Column b) 1. The 3’ end of the G-overhang positions on the RNA template in TER via base pair formation at the active site of TERT. 2. The addition of nucleotides creates the telomeric DNA repeat. 3. The telomerase translocates to resume telomeric repeat synthesis. 4. The elongated DNA strands are ready [4].
Figure 1.
Mechanism of telomerase activity at the end of the replication cycle. Column a) During the replication, the lagging strand is created based on the leading strand, that was present in the nucleus. The new strand is complementary to the old one but has its length shortened, which is the reason for cell aging. Elongation of the lagging strand is an extra step that requires telomerase and does not happen in every human cell. Column b) 1. The 3’ end of the G-overhang positions on the RNA template in TER via base pair formation at the active site of TERT. 2. The addition of nucleotides creates the telomeric DNA repeat. 3. The telomerase translocates to resume telomeric repeat synthesis. 4. The elongated DNA strands are ready [4].
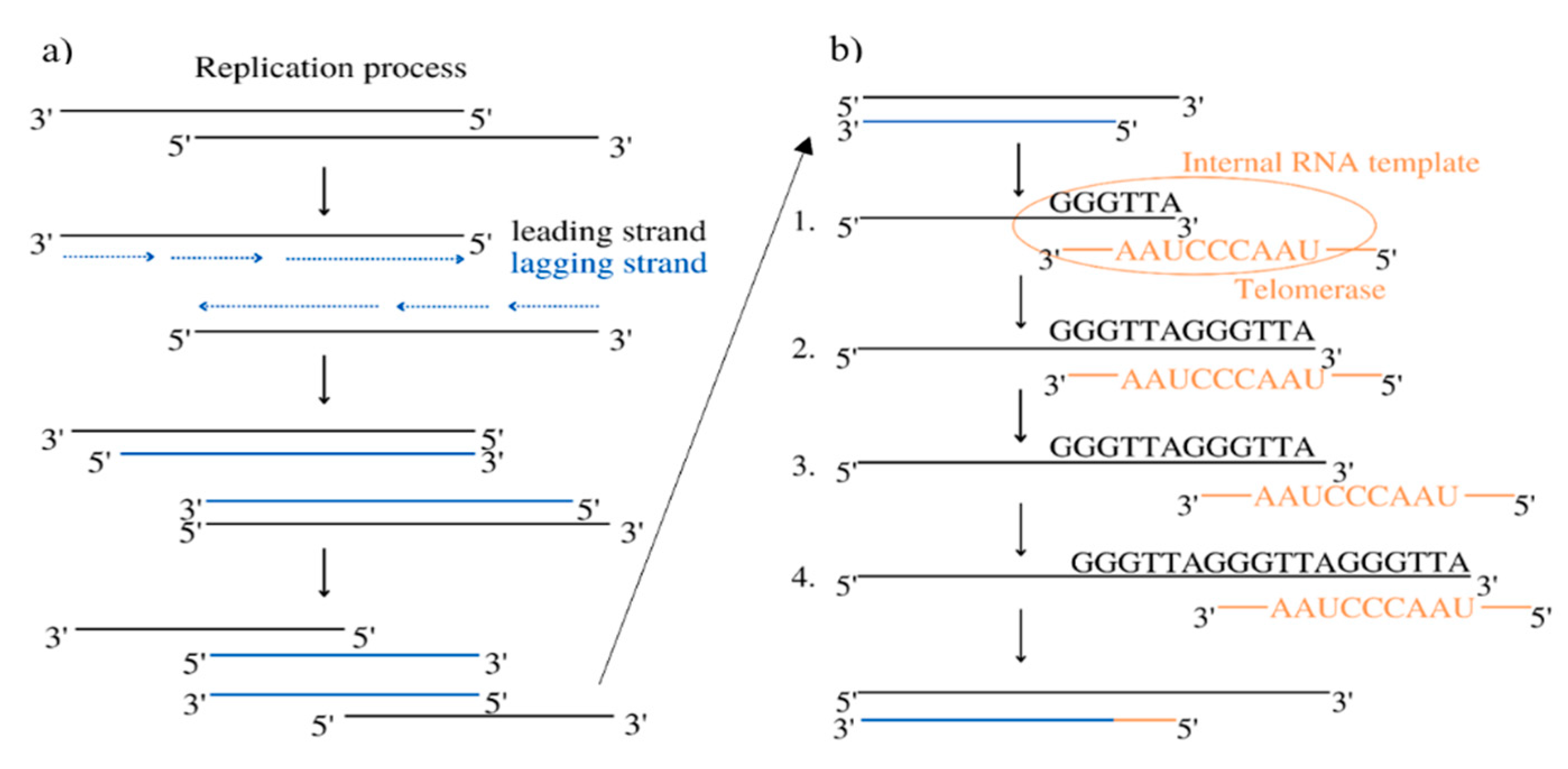
Figure 2.
Substances listed in the article except for bengalin, tianshengyuan-1, Angelica sinensis polysaccharide, RNA-protein complex extracted from Agaricus blazei Murill, patensin, manisa propolis, and polysaccharide from Gymnodinium sp. A3. (All substances have been downloaded from pubchem.ncbi.nlm.nih.gov).
Figure 2.
Substances listed in the article except for bengalin, tianshengyuan-1, Angelica sinensis polysaccharide, RNA-protein complex extracted from Agaricus blazei Murill, patensin, manisa propolis, and polysaccharide from Gymnodinium sp. A3. (All substances have been downloaded from pubchem.ncbi.nlm.nih.gov).
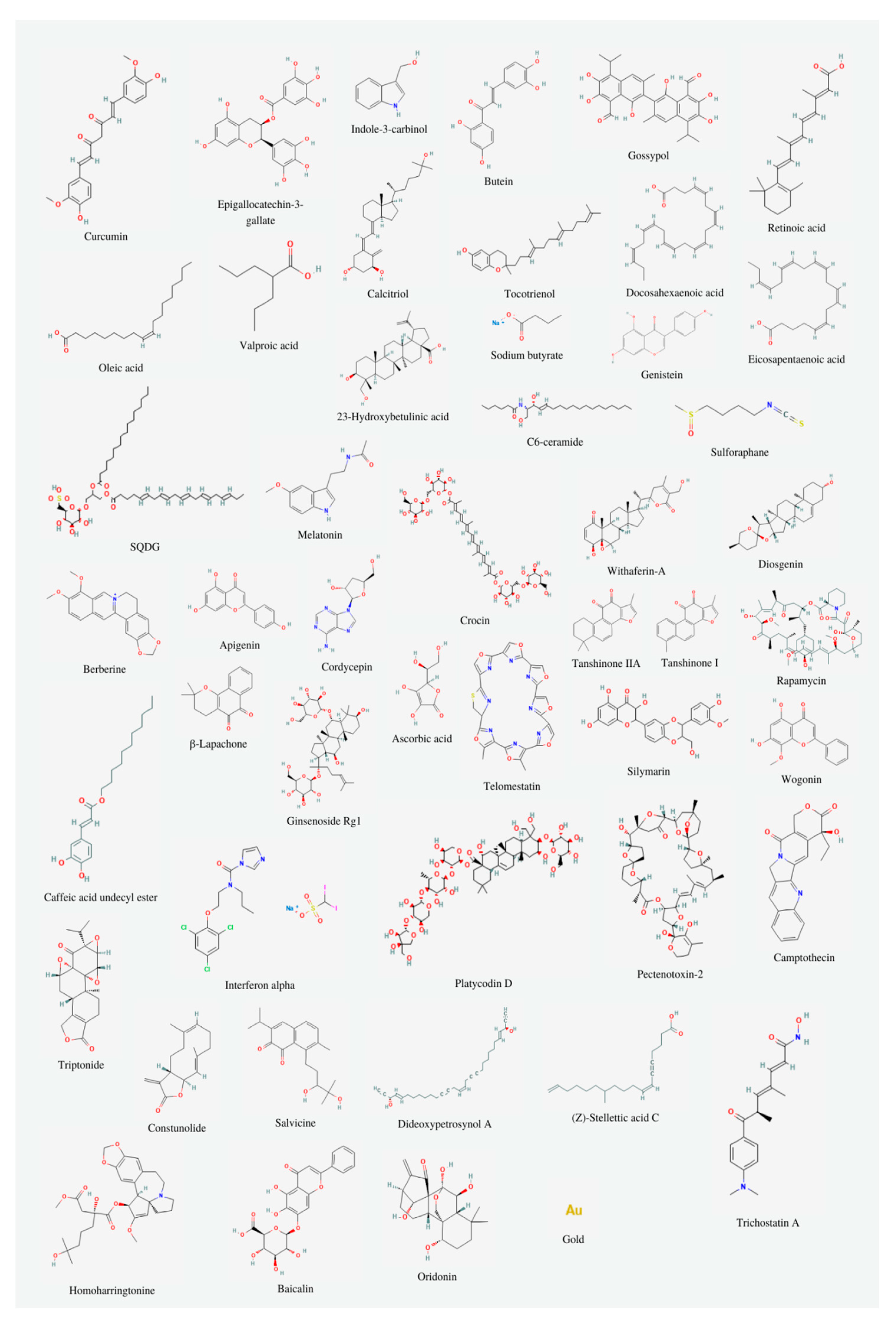
Figure 3.
The influence of natural telomerase inhibitors on leukemia cells.
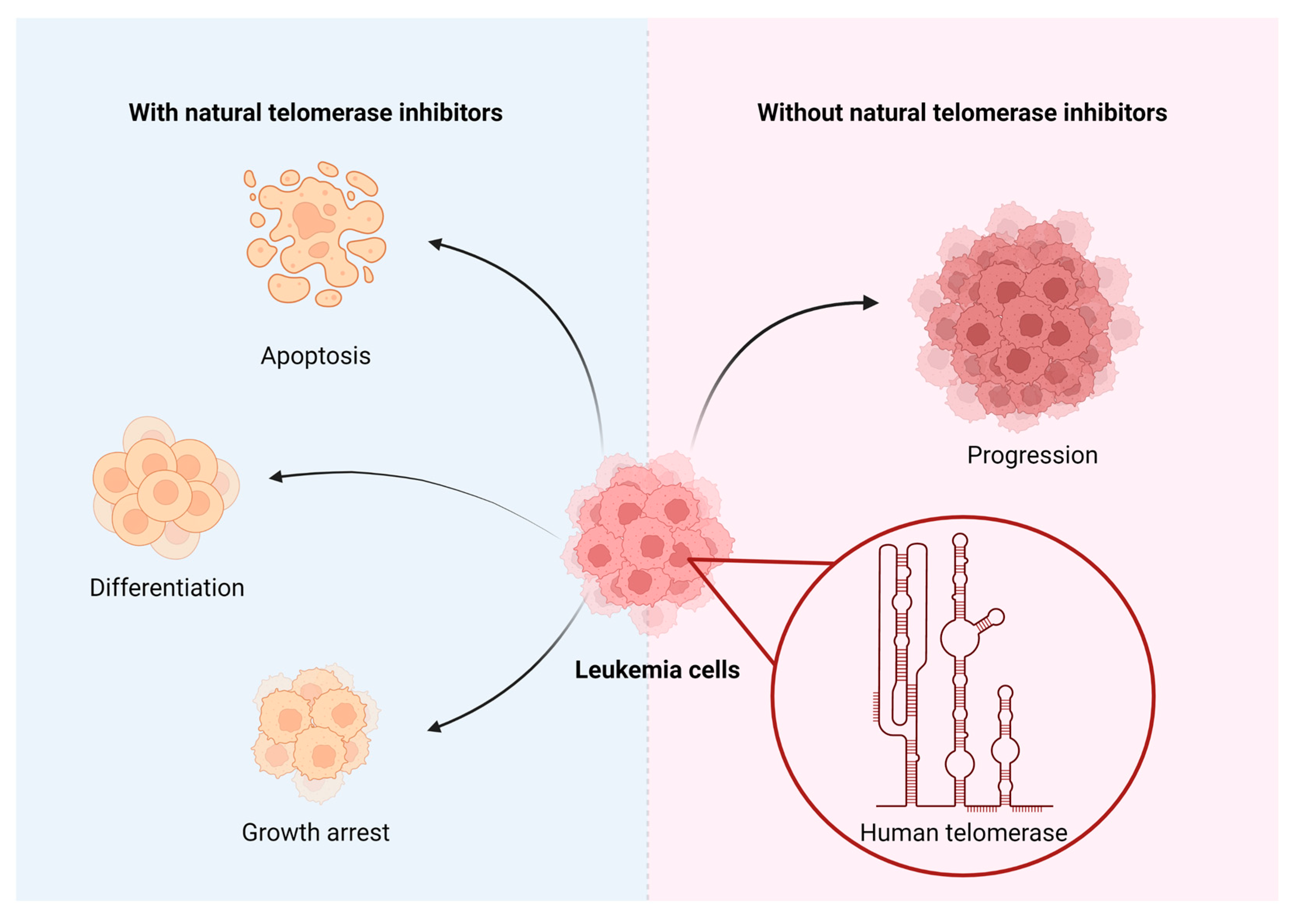
Figure 4.
Literature research diagram.
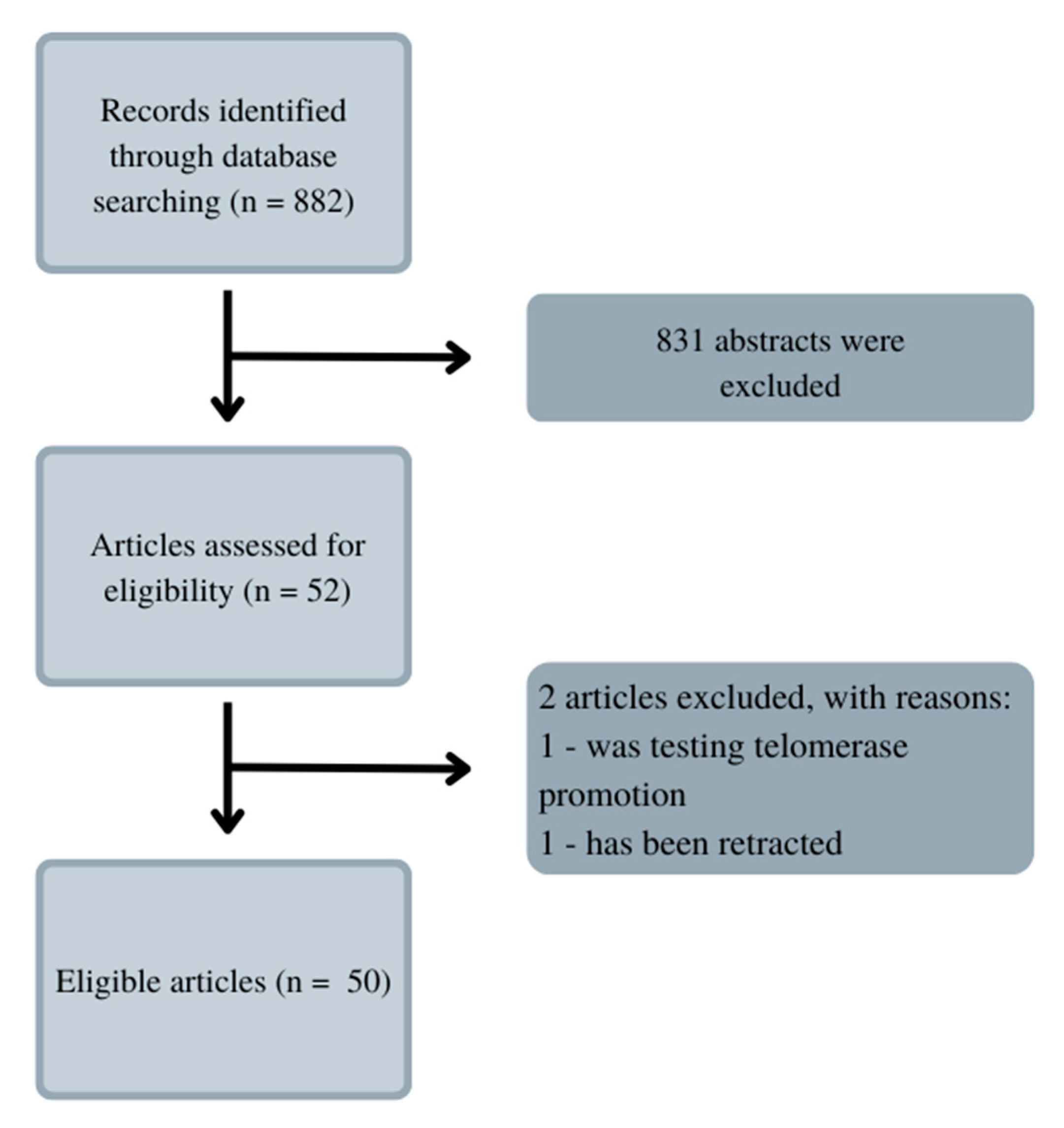
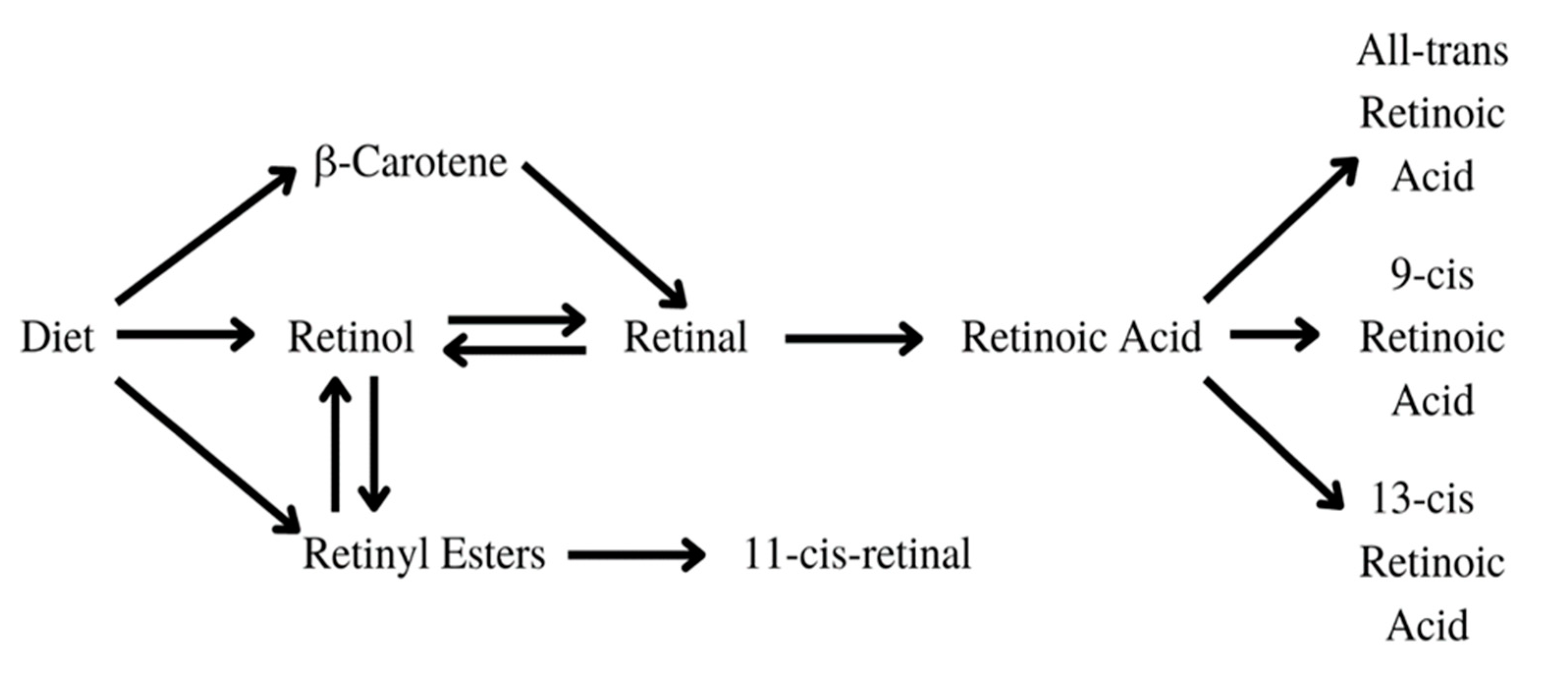
Figure 7.
Transformation of retinol to ATRA, which leads to cellular changes.
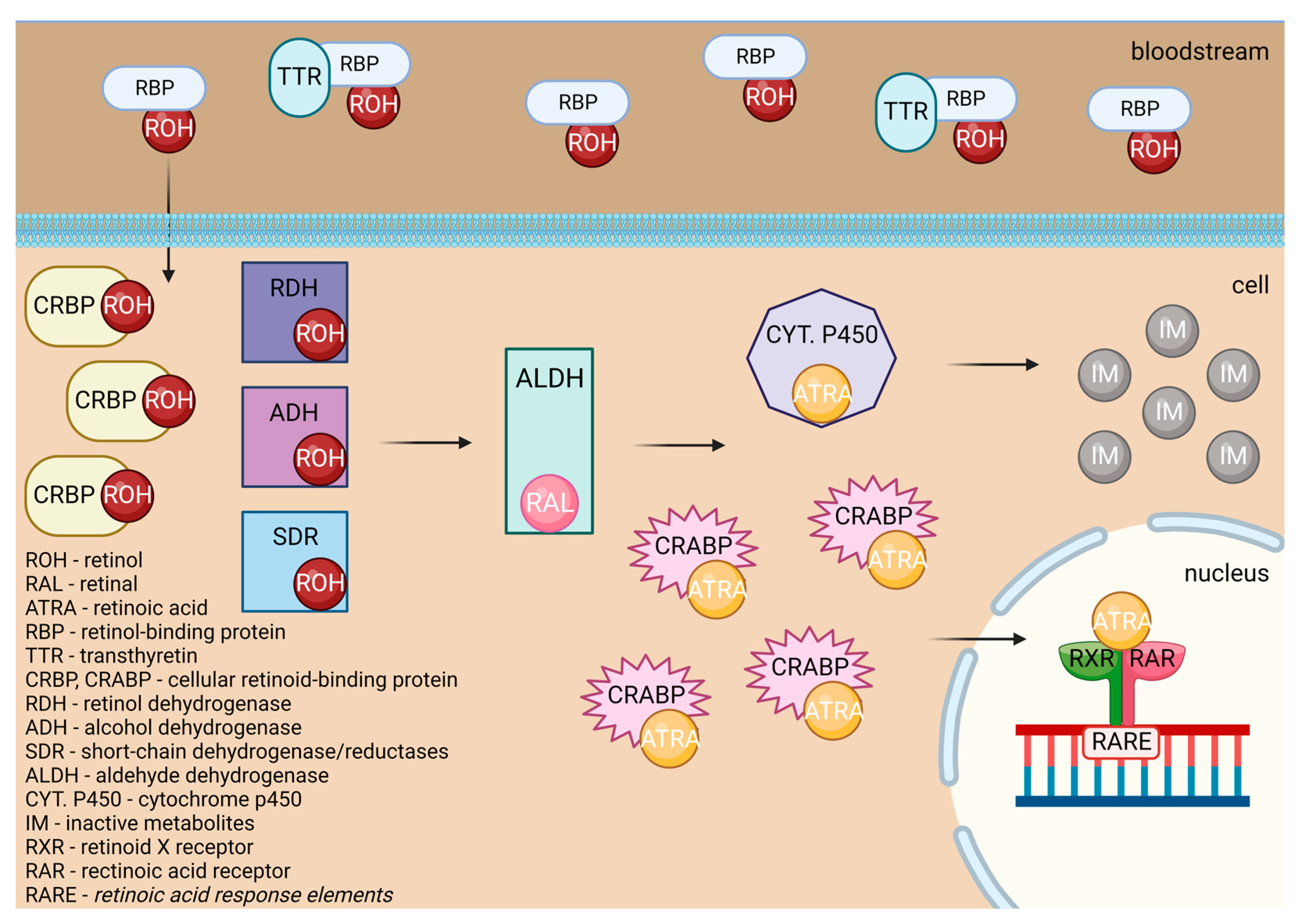
Figure 8.
The influence of ATRA on DNA, which is used whilst treating APL patients. 1. Patient’s DNA. 2a. Treatment without ATRA. 2b. Treatment with ATRA. 2c. Treatment with a big dosage of ATRA.
Figure 8.
The influence of ATRA on DNA, which is used whilst treating APL patients. 1. Patient’s DNA. 2a. Treatment without ATRA. 2b. Treatment with ATRA. 2c. Treatment with a big dosage of ATRA.
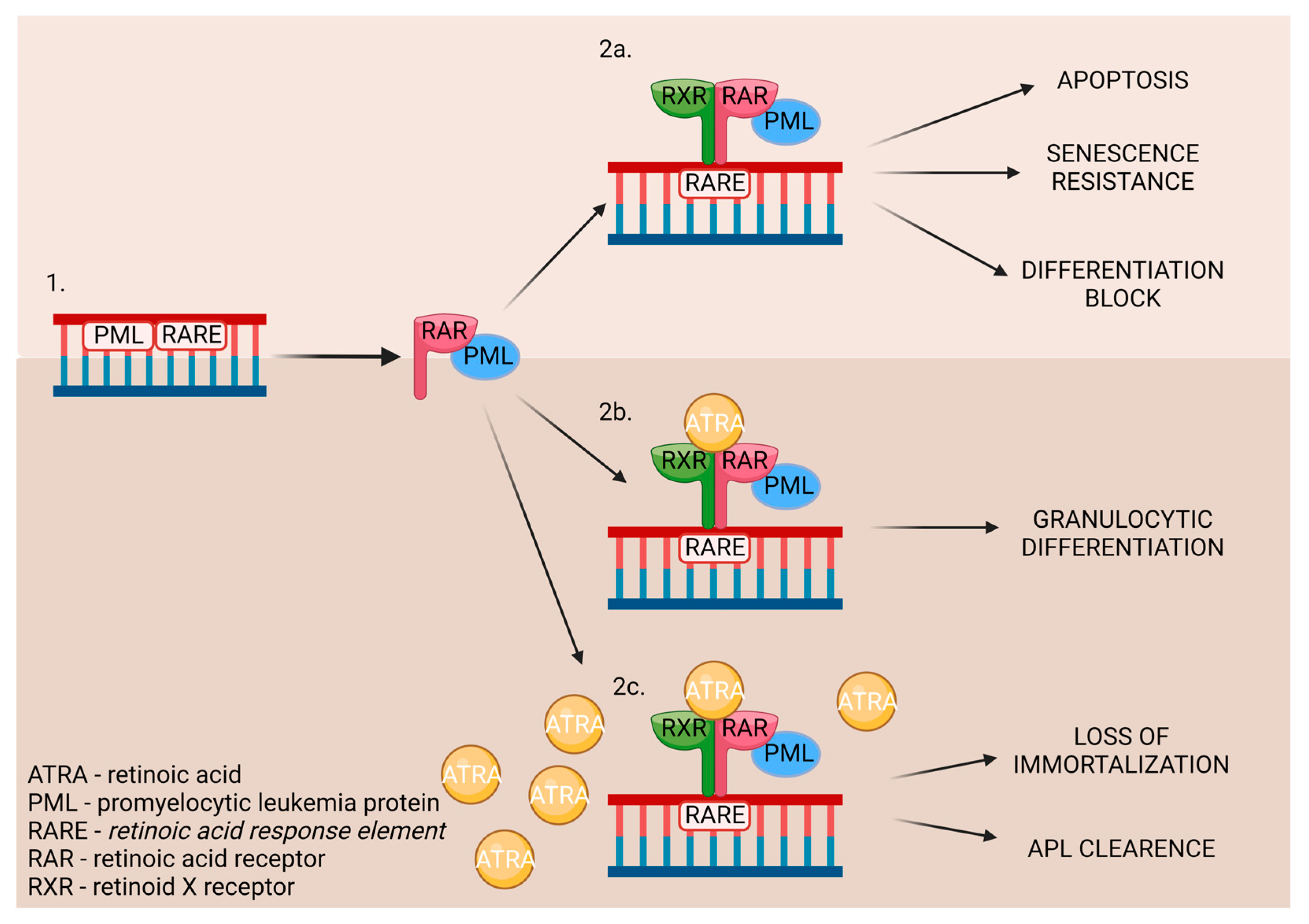
Figure 9.
Transformation of vitamin D [37].
Figure 9.
Transformation of vitamin D [37].
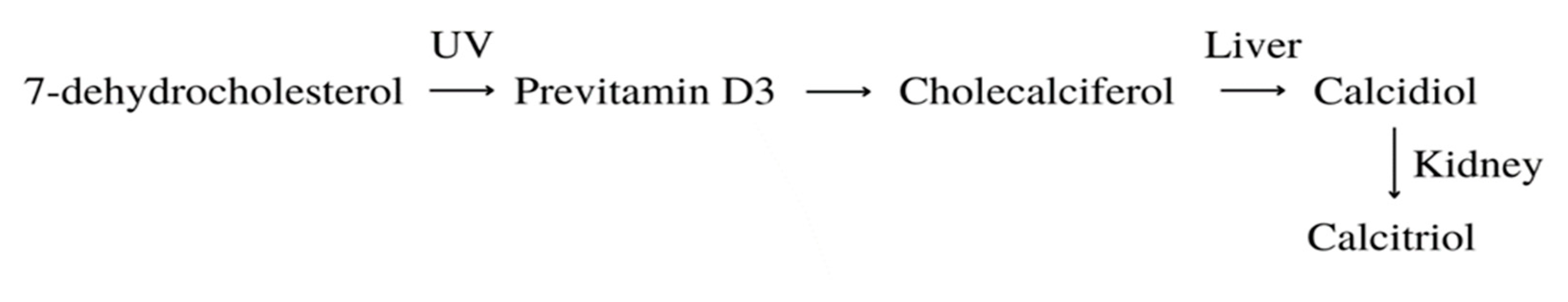
Figure 10.
The influence of tocotrienol on DNA, through various pathways, leads to apoptosis.
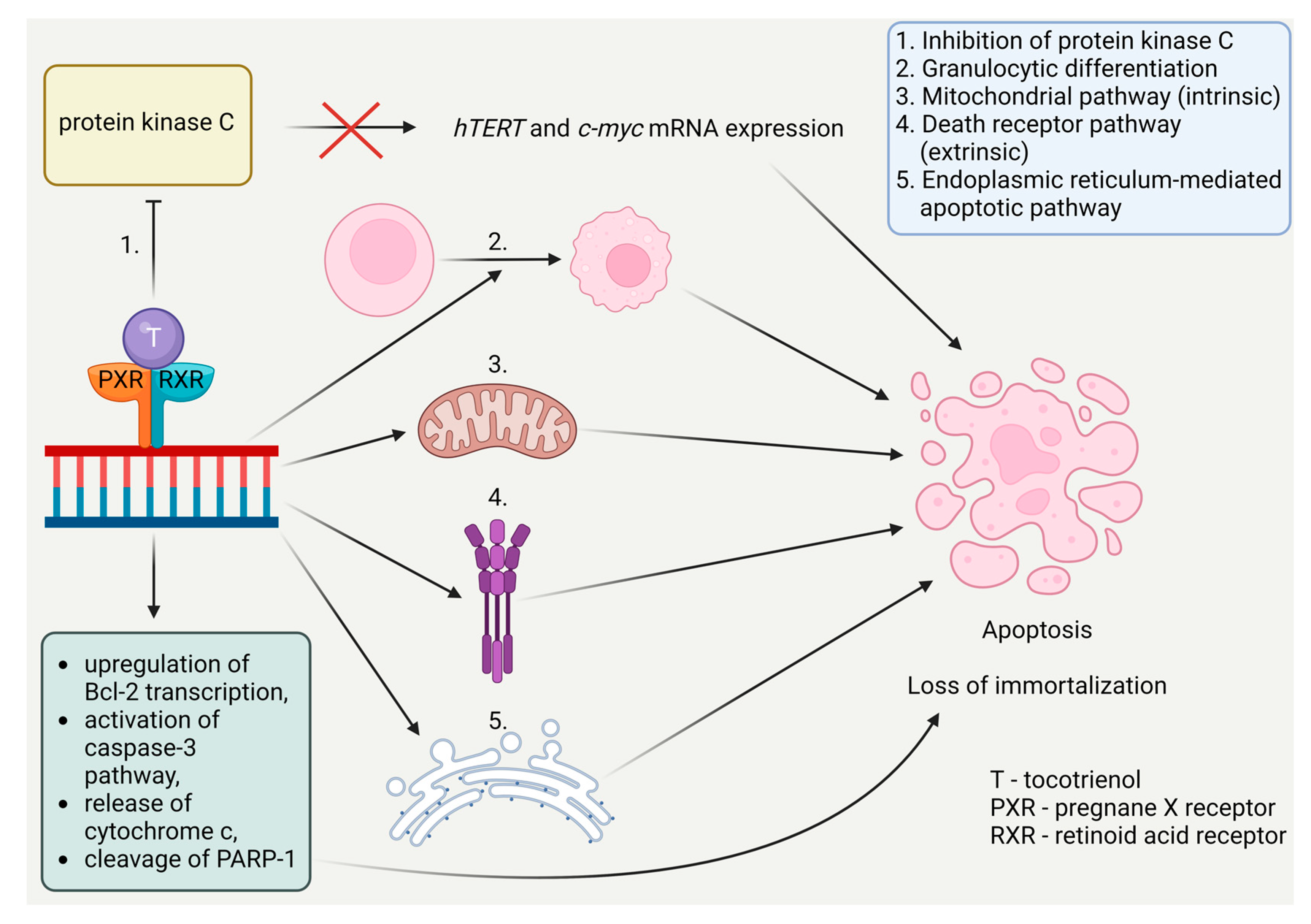
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



