Submitted:
02 January 2024
Posted:
04 January 2024
You are already at the latest version
Abstract
Background: The UK dairy sheep industry is relatively small but growing, particularly for cheese and yogurt products. Anecdotally, sheep milk (SM) is better tolerated by humans than cows’ milk and could have environmental as well as health benefits. All milk contains sub-micron particles called extracellular vesicles (EV) which are mainly derived from the mammary epithelium. Physiologically, milk derived EV are thought to aid development of infant immunity and the microbiome but may also have health benefits to adult humans. Methods: Using sequential ultracentrifugation vesicles of <1µm (LEV) followed by <200nm (sEV) were isolated from six individual sheep during mid lactation. RNA was extracted and microRNA analysed by RTqPCR for sequences previously identified in cows’ milk. Human THP-1 monocytes were differentiated into macrophages and incubated with SM-derived LEV and sEV in the presence of pro-inflammatory LPS to measure effects on secretion of the chemokine CCL-2 or in the presence of DMNQ and fluorescent dihydrorhodamine-1,2,3 to measure reactive oxygen species. Results: LEV induced an increase in ROS in both monocytes and macrophages, whilst sEV decreased DMNQ-mediated ROS in macrophages but not monocytes. Interestingly, the LEV did not induce CCL2 release, however they increased LPS-induced CCL2 secretion in monocytes but not macrophages. miR26a, miR92a, miR125b, miR155, miR223, were identified in both sEV and LEV by RT-qPCR and could be responsible for modulation of ROS and CCL2 expression. Conclusions: These findings suggest that sheep milk can influence human monocyte/macrophage responses and so is worthy of further investigation for its potential human and non-human animal health benefits.
Keywords:
sheep milk
; extracellular vesicles
; inflammation
1. Introduction
In 2022 12.4 billion liters of cows’ milk (CM) were produced, accounting for £6.6bn UK revenue [1]. However, recent years have seen a steady decline in the demand for CM, one of the reasons cited being consumer concerns about dietary intolerances [2] as well as welfare and sustainability[3]. Small ruminant milk including sheep milk (SM) may be better tolerated by the human gut due to differences in the whey protein composition and in the lactose content [4]. Nonetheless, SM remains relatively under-researched as a possible alternative to CM products for human consumption.
Extracellular vesicles (EV) are small (<1µm) plasma membrane enclosed particles released from all cells into all bodily fluids so far investigated [5]. They carry cargo which can include proteins, lipids and genetic material, particularly microRNA (miRNA), and can influence recipient cell behavior either by being taken up or by receptor-ligand interactions at the cell surface [6]. EV have recently been classified into large (200-1000nm; LEV; microvesicles) and small (50-150nm; sEV; exosomes). Whilst LEV are mostly released via budding from the cell surface and sEV via an endosomal pathway there is some overlap in size and even expression of characteristic surface protein markers between the two categories [7]. A third class called Apoptotic Bodies can be up to 3µm and are released from apoptosing cells. A fourth class are the milk fat globule (MFG) which are low-density particles released by budding from the plasma membrane of mammary epithelial cells [8] with a third membrane visible under EM. Although the majority of their cargo is lipid they do also contain some cytoplasmic proteins and genetic material [9].
There is continued pressure on the UK livestock industries to produce more output with less input, as well as also minimizing negative impacts across the food system (all the elements, actors and activities involved in producing and consuming food) including the environment, the welfare and health of the food producing animals and consumers all while ensuring prices and accessibility stay at an equitable level [10,11,12]. With the changing policies and subsidies for farmers, factors such as land use, antibiotic usage and environmental damage and animal welfare are becoming more and more important. Sheep can take advantage of lower quality land, for example they can be grazed on upland land not suitable for any other function, enabling less lowland (i.e. more productive land) to be used for dairy cow production; this could allow for more milk production but lessen the overall negative effects of dairy cow milk production.
Milk-derived EV are most likely released from mammary epithelium together with MFG and physiologically are thought to be involved in development of the infant immune system and the infant gut microbiome (and development of the rumen in ruminants) [13,14]. However, there is good evidence that CM-derived EV (CMEV) can survive the acid environment in the human stomach and can influence human cell behavior [15,16]. Cow milk is used for infant human milk supplementation. If human infants do not have access to or cannot tolerate human breast milk or cows’ milk, the option of sheep milk could aid in supplementation and provide them with the beneficial microbial effects seen by cow and goat milk [17]. This could be a more economical option than expensive formula-based products.
Considering the impact that the addition of SM in the dairy industry could have and the potential benefits it could have on human health, we decided to explore this area. We hypothesized that SM-derived EV (SMEV) could have beneficial effects including anti-inflammatory and antioxidant properties and may harbor potentially protective miRNA has been described for CMEV previously [18].
2. Materials and Methods
Materials
All plastics and all tissue culture reagents were from ThermoFisher Scientific (Loughborough, UK). All general laboratory chemicals were from Sigma Aldrich (Merck; Poole, UK).
Milk Collection
Milk was collected from Bevistan Dairy (Bedfordshire, UK) using a standard sample collector during the normal milking parlor routine from six individual sheep at mid-lactation cycle in July 2023. Milk was immediately frozen at –20°C at the dairy and transported frozen to the laboratory at the Royal Veterinary College for EV isolation and further analysis.
Isolation of SM-EV
Milk was thawed quickly in a water bath at 37°C and immediately aliquoted into two 20mL fractions, one of which was refrozen at -80°C for later use. The remaining 20mL fraction was centrifuged at 3000 xg 10 min at 4°C in a swing out benchtop centrifuge to separate the cream and remove any cellular debris. The cream was carefully removed from the top of the samples using a spatula. The supernatant was then decanted into a clean tube being careful not to disturb any pellet containing cellular debris. To remove casein micelles from the remaining milk fraction, glacial acetic acid was added dropwise until the pH reached 4.6 and visible precipitation was seen [19]. The precipitate was then pelleted at 3000 xg 10 min at 4°C in a swing out benchtop centrifuge. Supernatant were carefully decanted into clean Beckman 14mL polyallomer ultracentrifuge tubes (Cat #331374) for further extraction. Supernatants were centrifuged in a Beckman Coulter Optima L-80-XP ultracentrifuge using a SW40Ti rotor at 17,000 xg for 30 minutes, 4°C, without braking, to pellet the large EV population (LEV; microvesicles). SM-LEV pellets were resuspended in 1mL 1xPBS and aliquoted before storage at -80°C for further analysis. Supernatants were decanted into clean ultracentrifuge tubes and centrifuged at 100,000 xg for 90 minutes at 4°C in a SW40Ti rotor without braking to pellet the small EV population (sEV; exosomes). Supernatants were discarded and pellets resuspended in 8mL 1xPBS. This was then carefully layered on top of a 30% sucrose solution and centrifuged at 100,000 xg for 60 min using the SW40Ti rotor, without braking, to further purify sEV. The cloudy layer at the interface of the PBS and sucrose phase containing sEV was removed using a sterile pastette and placed in a clean ultracentrifuge tube, diluted with 1xPBS and centrifuged using the SW40Ti rotor without braking for 60 min 100,000 xg at 4°C. The top 1mL was collected as a negative control and then the remainder of the supernatant was carefully removed and discarded. The remaining pellet was resuspended in 1mL 1xPBS and immediately frozen in aliquots for further analysis.
Flow Cytometry
5µL of final preparation of SM-LEV or SM-sEV from each milk sample was diluted with 45µL 1xPBS. 1µL Vesidye-600 (Vesiculab, Nottingham UK; reconstituted according to the manufacturer’s instructions) was added to each and incubated at 37°C for 1h before further dilution to 500µL with 1xPBS. 10µL Enumeration beads (Coulter) were added to each sample before analysis using a BD FACS CANTO, calibrated using CST beads (both BD, Witney, UK). EV were gated on size using 1µm NIST beads (ThermoFisher) and the population <1µM was then gated on Vesidye positivity (see Figure 1 for representative PercP-Cy5.5 / SSC plot). The total number of EV were calculated using the equation: EV/µL = 20*[events of interest]*[dilution factor] / [enumeration bead count] as described previously [20].
Determination of Protein Concentration of EV
10µL 5xRIPA buffer was added to 40µL of final preparation of SM-LEV or SM-SEV from each milk sample. After vortexing to mix, the samples were sonicated for 15 min at 30°C and immediately analysed using Pierce micro-BCA assay kit (ThermoFisher) according to the manufacturer’s instructions.
RNA Isolation and miRNA Analysis
For RNA isolation from SM-SEV, 500µL of the final preparations from each milk sample were used to isolate total RNA using the exoEasy total RNA kit (Qiagen) according to the manufacturer’s instructions. For RNA isolation from SM-LEV, 500µL of the final preparations were re-centrifuged at 17,000 xg, 15 min, 4°C in a bench top microfuge. The resulting pellet was dissolved in 700µL Qiazol (Qiagen) and the rest of the protocol for the exoEasy total RNA kit was followed. RNA was quantified using a DeNovix DS-11 nanodrop DeNovix DS-11 (DeNovix, Wilmington, DE, USA). Individual miRNAs (miR10b, miR26a, miR27a, miR92a, miR125b, miR126, miR155, and miR223) were reverse transcribed using the TaqMan® microRNA RT kit (Cat #4366596, ABI. Thermofisher. UK) and the associated miRNA-specific inventoried predesigned primers (TaqMan® micro-RNA assay kit, #4427975. ABI). Total RNA was normalized at a concentration of 2ng/μl and 5 μl of RNA were added to the reaction mix containing 0.15 μl 100mM dNTP, 1 μl Reverse Transcriptase enzyme (50U/μl), 1.5 μl 10 × RT buffer, 0.19 μl RNase inhibitor (20U/μl), 3 μl 5× RT specific-primer and 4.16 μl DEPC-treated water to obtain a final volume of 15 μl. RT reaction conditions were as follows: primer annealing for 30 min at 16°C, extension for 30 min at 42°C and final incubation for 5 min at 85°C to stop the reaction. All RT-qPCRs were carried out in triplicate with TaqMan™ Universal Master Mix II, no UNG (Cat# 4440040) and were performed on the Bio-Rad CFX384 Real Time PCR detection cycler system with the following conditions: an initial inactivation step at 95°C for 10 min, followed by 40 cycles consisting of denaturing at 95°C for 10 s, annealing and elongation at 60°C for 60s.
Cell Culture
THP-1 human monocytes were from ATCC. They were cultured at 37°C in RPMI, 10% FCS, with 0.05mM β-mercaptoethanol. For differentiation, THP-1 were incubated with 5ng/mL PMA for 72h at 37°C.
Reactive Oxygen Species (ROS) Assays
THP-1 monocytes were pelleted, washed and resuspended at 2x106 cells per mL in RPMI without serum. They were incubated with 5µM dihydrorhodamine 1,2,3 for 10 min in the dark, before seeding at 10,000 cells per well in 96 well plates. SMEV were added at a final dilution of 1:100. 2,3-Dimethoxy-1,4-naphthoquinone (DMNQ; AbCam Cambridge, UK), a well described stimulus for ROS generation in monocytes [21] was added at a final concentration of 250µM. Cells were incubated for 24h at 37°C and plates were read in a fluorescent plate reader (Tecan Cyto 400, Reading, UK) at the start and end of the experiment. All treatments were added in triplicate. For THP-1 macrophages, they were plated at 100,000 cells per well in full medium together with PMA at 5ng/mL, incubated for 72h at 37°C. The medium was removed together with non-adherent cells. The adherent population were washed twice with serum free RPMI to remove bovine-derived EV from the serum. Dihydrorhodamine 1,2,3 was added at a final concentration of 5µM and the remainder of the procedure was as outlined above.
Measurement of CCL2 Secretion
THP-1 monocytes were pelleted, washed and resuspended at 2x106 cells per mL in RPMI without serum. They were seeded at 25,000 cells per well in 24 well plates and LPS O55:B5 was added to some wells at a final concentration of 1µg/mL. SMEV were added at a final dilution of 1:100. Cells were incubated for 24h before supernatants were collected and cells pelleted by centrifugation at 5,000 xg for 10 minutes at 4°C. Supernatants were carefully decanted and stored at -80°C followed by analysis for CCL2 using eBioscience Ready-Set-Go! ELISA (Invitrogen, ThermoFisher) according to the manufacturer’s instructions. All treatments were added in triplicate. For THP-1 macrophages, they were plated at 250,000 cells per well in full medium together with 5ng/mL PMA and incubated for 72h at 37°C. Medium was removed together with non-adherent cells. The adherent population were washed twice with serum free RPMI to remove bovine-derived EV from the serum. Cells were incubated in serum free RPMI with LPS (1µg/mL) and SMEV at a final dilution of 1:100 for 24h at 37°C. Supernatants were harvested as above and subjected to the same ELISA assays.
Statistical Analysis
All statistical analysis was carried out using Prism 10 (GraphPad, CA). Data were tested for normality using Shapiro-Wilke test and One way ANOVA followed by Fishers LSD for ROS assays and ELISA. ROS assays were carried out using the six sheep EV samples in triplicate, CCL2 ELISAs were carried out using the six sheep EV samples in duplicate. p<0.05 was considered statistically significant (*). ** p<0.01, *** p<0.001.
3. Results
3.1. Characterisation of Sheep Milk Derived LEV and sEV
As shown in Table 1, we identified small and large EV by flow cytometry using a lipid permeable dye, and were able to detect measureable total protein and total RNA in small and large EV from each sample.
3.2. Monocyte and Macrophage ROS Production Induced by Sheep Milk Derived LEV and sEV
We hypothesised that SMEV could have an effect on leukocyte oxidant stress both in the presence or absence of other stimuli. Equal volumes of isolated LEV or sEV were added to THP-1 monocytes or differentiated macrophages that had been pre-loaded with dihydrorhodamine-1,2,3 fluorescent label and incubated for 24h in the presence or absence of DMNQ, which is known to induce oxidant stress [21]. There was a significant increase in ROS in cells treated with LEV, but not with sEV in both monocytes (Figure 2A) and macrophages (Figure 2B). Neither LEV or sEV affected the DMNQ response in monocytes. sEV significantly downmodulated the DMNQ response in macrophages.
3.3. Secretion of CCL2 Chemokine Induced by Sheep Milk Derived LEV and sEV
Since SMEV were able to modulate ROS in monocytes and macrophages and these can be an indication of wider pro-inflammatory gene expression we next measured secretion of CCL2 chemokine after 24 incubation with either LEV or sEV (Figure 3). Incubation with LEV or sEV alone did not induce CCL2 secretion, however in monocytes (Figure 3A) there was a significant decrease in CCL2 secretion compared to LPS alone when cells were incubated with LPS and LEV or sEV. This was not replicated in macrophages (Figure 3B).
3.4. miRNA Contents of Sheep Milk Derived LEV and sEV
In order to determine whether there could be an influence of miRNA cargo in the increase in ROS production or modulation of CCL2 production we measured miRNA in LEV and sEV from individual SM samples. We measured eight different miRNA based on previous reports of their presence in either SM or CM and reporting in other settings of their effects on inflammatory pathways. We were able to measure expression in five of the eight miRNA tested (miR26, 92a, 125b, 155 and 223). Expression levels of individual miRNA was consistent between LEV and sEV (Figure 4). There was no detectable expression of miR10b, miR27a, or miR126 in either sEV or LEV (data not shown). There was no detectable expression of any mir in water controls (data not shown).
4. Discussion
As the market in the UK for sheep milk and dairy products increases, it is important to gain a greater understanding of the constitutive components that make up this complex highly nutritious biofluid. There is an increasing awareness that milk contains not only macronutrients which are regularly tested and reported on in commercial settings, but also includes the potential for transfer of genetic material from sheep to human. Secretion of extracellular vesicles by mammary epithelial cells into milk is increasingly thought to function physiologically to aid development of the infant immune system and gut [13], starting with intake of colostrum and continuing throughout lactation. Here, we have demonstrated that it is possible to isolate small and large EV-like particles from raw sheep milk. We have then demonstrated a functional effect on human inflammatory pathways in vitro; we believe this is the first such publication to do so. Intriguingly, we observed an increase in ROS when LEV were applied to both THP-1 monocytes and macrophages, and this was additive when cells were treated with DMNQ, a known pro-oxidant compound. In contrast, sEV did not induce any ROS and inhibited DMNQ-stimulated ROS, suggesting that there may be differences in cargo, cell signaling efficiency or rates of cellular uptake for the two species of EV.
ROS production and oxidative stress is often associated directly with pro-inflammatory pathways and can lead to activation of secretion of cytokines and chemokines, so it is important to understand if intake of milk-derived EV contribute to, or can alleviate, oxidative stress, especially in human individuals with underlying chronic gastrointestinal inflammation such as ulcerative colitis and Crohn’s Disease [22]. CCL2 is a key chemokine that drives recruitment of monocytes to inflamed endothelium. It is secreted by endothelial cells, monocytes and macrophages in large amounts in several highly prevalent human chronic inflammatory conditions including inflammatory bowel disease [23], rheumatoid arthritis [24], type 2 diabetes, insulin resistance and metabolic syndrome [25], chronic kidney disease [26], atherosclerosis [27] and endothelial dysfunction [28]. Here, we have found that though SM-LEV increased ROS production in monocytes (Figure 2A) and macrophages (Figure 2B), they also significantly inhibited CCL2 production in monocytes (Figure 3A). SM-sEV significantly inhibited ROS in macrophages (Figure 2B), but not monocytes (Figure 2A), though they did significantly inhibit CCL2 production in monocytes (Figure 3A). Since CCL2 has such a large impact on many different chronic inflammatory disorders, non-pharmacologic approaches to lessen its production and therefore reduce monocyte chemotaxis through simple dietary intervention could be a useful strategy. Based on these promising data, if these potential health benefits are mirrored in vivo, encouraging consumption of sheep milk as part of a healthy diet might meet with a positive consumer response.
We investigated the RNA cargo of the isolated SM sEV and LEV and were able to identify several microRNA species that have previously been shown to be associated with chronic inflammation, either exacerbating or demonstrating regulation of immune cell function, activation and differentiation. The most highly expressed (lowest Ct; Figure 4) in our panel was miR26a, which has previously been shown to be elevated in raw milk and to be relatively consistently expressed across lactation [29], more recently as one of the most abundant miRNA in cows’ milk [30] and in sheep milk EV [31]. In a separate study, miR26a has been shown to target IFNb and IL6 genes, which was postulated to be positively related to asthma reduction in infants that drink raw milk [32].
MiR92a was the next best expressed (Figure 4). MiR92a has been identified in a bovine study of mastitis [33]. Functionally, it has been associated with STAT3, CTLA4 and TLR1 regulation, genes that have been implicated as key players in chronic inflammatory immune responses. MiR125b has also been shown to regulate STAT3 [34] and Luoreng et al [35] found that MiR125b is a negative regulator of mastitis, with its expression significantly downregulated during LPS stimulation of bovine mammary epithelial cells in vitro. Mastitis is a key long standing endemic disease in sheep farming as well as in the dairy cattle industry [36], that provides a constant source of strain on already scarce resources such as time and money [37]. Although prevalence of the different forms of mastitis are scarce for meat sheep in the UK, let alone dairy sheep, it is thought that the incidence acute mastitis is 0-6.6% of meat sheep per flock [38] and chronic mastitis affected up to half a flock at a time [39]. Since we were able to identify expression in all six samples from healthy dairy sheep, we suggest that miR125b may hold promise as useful biomarker of ovine mammary health and worthy of further investigation. We detected low levels of MiR155 in both LEV and sEV tested, which may represent a species difference between small and large ruminants since Izumi et al [40] reported the presence of miR155 in bovine colostrum but not mature milk. MiR155 has been known for a long time to be a key miRNA in the regulation of both innate and adaptive immune responses [41]. For example, it has been shown to modulate differentiation of T and B cells as well as being a positive regulator for regulatory T cell differentiation [41]. Specifically, it has been implicated in expression of STAT3, CTLA4, as well as VCAM-1 and eNOS, suggesting that its presence could influence cytokine and chemokine expression in activated endothelial cells as well as contributing to leukocyte inflammatory responses. A link has recently been established between miR155 expression and CCL2 in RAW264 murine macrophages in vitro and in a rodent model of sepsis [42]. This is intriguing and worthy of further study, given our finding that both large and small SMEV included a cargo of miR155 but reduced rather than exacerbated LPS driven CCL2 in human THP1 cells.
In the present study we saw the most variation in expression of miR223 between individual sheep. MiR223 has previously been shown to be a negative regulator of neutrophil activation [41] and has recently been associated with mammary inflammation [43,44]. This could be a useful miRNA to follow up as a potential biomarker of early mastitis or other inflammatory conditions that might impact dairy sheep production or their health and welfare.
MiR10b, miR27a and miR126 have all previously been reported to be present in EV from cows and sheep [29,31], however we were not able to detect these species in the samples. This may be due to differences in processing or the phase of lactation at which our samples were taken.
Sheep milk consumption is gaining popularity and therefore its production is becoming commercially more viable for farmers in the UK. From a One Health and One Welfare perspective, if sheep milk contains beneficial anti-inflammatory properties for human infants this may also provide an advantage for the sheep offspring. Thereby, possibly providing lambs with an advantage over calves in being able to better ward off diseases and syndromes that are of greater prevalence in farmed settings. It will also be important in the sheep meat industry to ascertain whether these benefits also impact dietary phospholipids in the lamb meat that would then be eaten by human consumers [45]. Delving more into these interesting questions would possibly unlock hidden advantages of the sheep industry for both the health of human and non-human animals. This represents an important moment to ensure that small ruminant health and welfare is maintained as the industry grows. There are also many questions that should be addressed that relate to the sustainability of sheep dairy enterprises, including the land use for ovine vs bovine systems, water and land contamination from ovine waste, production of greenhouse gases (methane) and use possible increased use of medicines such as antimicrobials. Whilst these are clearly beyond the scope of this small molecular study, there could be a benefit to following up with an interrogation of ovine excreta to determine whether similar miRNA are present, what effect they might have on the microbiome, whether there are ovine miRNA present that can influence soil and aquatic micro-organisms and therefore influence wider environmental questions. Additionally, it would be interesting to ascertain if the health and welfare of the sheep impact these properties, for example if sheep have higher welfare and/or health status does this positively impact the number or cargo of the LEV and or sEV in the milk. This type of One Health-One Welfare (where the health and welfare of humans, non-human animals and the ecosystem are considered to be interdependent) investigation could move us more towards more win-win-win actions for more sustainable farming [46].
This study represents a first step towards understanding the potential benefits of human ingestion of sheep milk, particularly for those individuals who are not able to tolerate cows’ milk. Our approach to interrogating the molecular profile of the mammary epithelial cell via analysis of secreted EV could also be useful to understand more about potential health and welfare of dairy sheep (and could be applied to other small ruminant dairy species). In particular, the finding that it is possible to measure presence of different miRNA species could allow insight into the changes that take place, either physiologically during progression of lactation, or during mastitis. There is also potential to develop a biomarker panel based on the presence of miRNA that are up or down regulated in mastitis or that change at different stages of lactation.
5. Conclusions
In this study we have shown that both small and large EV particles can be obtained from raw sheep milk; these particles are able to influence human immune cell behaviour, in vitro, including reactive oxygen species production and secretion of CCL2 chemokine. We have demonstrated that it is possible to isolate sufficient miRNA cargo from individual SM-derived EV to test by qRT-PCR. Our findings broadly suggest that human intake of sheep milk may be beneficial for individuals with chronic inflammatory conditions. However, further work is required to fully interrogate the miRNA cargo and further explore the downstream effects that these particles have on human inflammation. The benefits of this work could impact not only human health and welfare but has the potential to also investigate benefits to livestock and environmental health and welfare and sustainable farming.
Author Contributions
Conceptualization, C.L.; methodology, C.L., B.S., R.P.; writing—original draft preparation, C.L., B.S., L.W., R.P.; writing—review and editing, C.L., B.S., L.W., R.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
This study did not require ethical approval. Milk was collected as part of routine flock management.
Data Availability Statement
raw data is available upon request.
Acknowledgments
We thank Valley View Farm, Carlton Bedfordshire, UK for providing milk samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- AHDB. Dairy Market Dashboard https://ahdb.org.uk/dairy-market-dashboard. (accessed on 29th December 2023).
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N.W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa, M. Invited review: Milk lactose-Current status and future challenges in dairy cattle. J Dairy Sci 2019, 102, 5883–5898. [Google Scholar] [CrossRef]
- Mee, J.F.; Boyle, L.A. Assessing whether dairy cow welfare is "better" in pasture-based than in confinement-based management systems. N Z Vet J 2020, 68, 168–177. [Google Scholar] [CrossRef]
- Roy, D.; Ye, A.; Moughan, P.J.; Singh, H. Composition, Structure, and Digestive Dynamics of Milk From Different Species-A Review. Front Nutr 2020, 7, 577759. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc Natl Acad Sci U S A 2012, 109, E2110–2116. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Simpson, R.J.; Ji, H.; Aikawa, E.; Altevogt, P.; Askenase, P.; Bond, V.C.; Borràs, F.E.; Breakefield, X.; Budnik, V.; et al. Vesiclepedia: a compendium for extracellular vesicles with continuous community annotation. PLoS Biol 2012, 10, e1001450. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Chen, M.; Li, N.; Han, R.; Yang, Y.; Zheng, N.; Zhao, S.; Zhang, Y. Bioactive Functions of Lipids in the Milk Fat Globule Membrane: A Comprehensive Review. Foods 2023, 12. [Google Scholar] [CrossRef]
- Pawlowski, K.; Lago-Novais, D.; Bevilacqua, C.; Mobuchon, L.; Crapart, N.; Faulconnier, Y.; Boby, C.; Carvalho, G.; Martin, P.; Leroux, C. Different miRNA contents between mammary epithelial cells and milk fat globules: a random or a targeted process? Mol Biol Rep 2020, 47, 8259–8264. [Google Scholar] [CrossRef]
- Boulton, A.C.; Rushton, J.; Wathes, C.M. Past trends and future challenges for a sustainable UK dairy industry. 2011, 172, 1–7. [Google Scholar]
- Shortall, O. Cows eat grass, don't they? Contrasting sociotechnical imaginaries of the role of grazing in the UK and Irish diary sectors. Journal of Rural Studies 2019, 72, 45–57. [Google Scholar] [CrossRef]
- Trewern, J.; Chenoweth, J.; Christie, I.; Keller, E.; Halevy, S. Are UK retailers well placed to deliver ‘less and better’meat and dairy to consumers? Sustainable Production and Consumption 2021, 28, 154–163. [Google Scholar] [CrossRef]
- Zempleni, J.; Aguilar-Lozano, A.; Sadri, M.; Sukreet, S.; Manca, S.; Wu, D.; Zhou, F.; Mutai, E. Biological Activities of Extracellular Vesicles and Their Cargos from Bovine and Human Milk in Humans and Implications for Infants. J Nutr 2017, 147, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; You, L.; Zhang, Z.; Cui, X.; Zhong, H.; Sun, X.; Ji, C.; Chi, X. Biological Properties of Milk-Derived Extracellular Vesicles and Their Physiological Functions in Infant. Front Cell Dev Biol 2021, 9, 693534. [Google Scholar] [CrossRef] [PubMed]
- Benmoussa, A.; Lee, C.H.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk microRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J Nutr 2016, 146, 2206–2215. [Google Scholar] [CrossRef]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. J Nutr 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Gallier, S.; Van den Abbeele, P.; Prosser, C. Comparison of the Bifidogenic Effects of Goat and Cow Milk-Based Infant Formulas to Human Breast Milk in an. Front Nutr 2020, 7, 608495. [Google Scholar] [CrossRef] [PubMed]
- Arntz, O.J.; Pieters, B.C.; Oliveira, M.C.; Broeren, M.G.; Bennink, M.B.; de Vries, M.; van Lent, P.L.; Koenders, M.I.; van den Berg, W.B.; van der Kraan, P.M.; et al. Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models. Mol Nutr Food Res 2015, 59, 1701–1712. [Google Scholar] [CrossRef]
- Mukhopadhya, A.; Santoro, J.; O'Driscoll, L. Extracellular vesicle separation from milk and infant milk formula using acid precipitation and ultracentrifugation. STAR Protoc 2021, 2, 100821. [Google Scholar] [CrossRef]
- Heinrich, L.F.; Andersen, D.K.; Cleasby, M.E.; Lawson, C. Long-term high fat feeding of rats results in increased numbers of circulating microvesicles with pro-inflammatory effects on endothelial cells. Br J Nutr 2015, 113, 1704–1711. [Google Scholar] [CrossRef]
- Namgaladze, D.; Preiss, S.; Dröse, S.; Brandt, U.; Brüne, B. Phospholipase A2-modified low density lipoprotein induces mitochondrial uncoupling and lowers reactive oxygen species in phagocytes. Atherosclerosis 2010, 208, 142–147. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Heilmann, R.M.; Paital, B.; Patel, A.; Yadav, V.K.; Wong, D.; Jergens, A.E. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front Endocrinol (Lausanne) 2023, 14, 1217165. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.P.; Singh, N.P.; Murphy, E.A.; Price, R.L.; Fayad, R.; Nagarkatti, M.; Nagarkatti, P.S. Chemokine and cytokine levels in inflammatory bowel disease patients. Cytokine 2016, 77, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Moadab, F.; Khorramdelazad, H.; Abbasifard, M. Role of CCL2/CCR2 axis in the immunopathogenesis of rheumatoid arthritis: Latest evidence and therapeutic approaches. Life Sci 2021, 269, 119034. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Kaminga, A.C.; Wen, S.W.; Liu, A. Chemokines in Prediabetes and Type 2 Diabetes: A Meta-Analysis. Front Immunol 2021, 12, 622438. [Google Scholar] [CrossRef] [PubMed]
- Puthumana, J.; Thiessen-Philbrook, H.; Xu, L.; Coca, S.G.; Garg, A.X.; Himmelfarb, J.; Bhatraju, P.K.; Ikizler, T.A.; Siew, E.D.; Ware, L.B.; et al. Biomarkers of inflammation and repair in kidney disease progression. J Clin Invest 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Georgakis, M.K.; Bernhagen, J.; Heitman, L.H.; Weber, C.; Dichgans, M. Targeting the CCL2-CCR2 axis for atheroprotection. Eur Heart J 2022, 43, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Martynowicz, H.; Janus, A.; Nowacki, D.; Mazur, G. The role of chemokines in hypertension. Adv Clin Exp Med 2014, 23, 319–325. [Google Scholar] [CrossRef]
- Chen, X.; Gao, C.; Li, H.; Huang, L.; Sun, Q.; Dong, Y.; Tian, C.; Gao, S.; Dong, H.; Guan, D.; et al. Identification and characterization of microRNAs in raw milk during different periods of lactation, commercial fluid, and powdered milk products. Cell Res 2010, 20, 1128–1137. [Google Scholar] [CrossRef]
- Li, W.; Li, W.; Wang, X.; Zhang, H.; Wang, L.; Gao, T. Comparison of miRNA profiles in milk-derived extracellular vesicles and bovine mammary glands. International Dairy Journal 2022, 134. [Google Scholar] [CrossRef]
- Quan, S.; Nan, X.; Wang, K.; Jiang, L.; Yao, J.; Xiong, B. Characterization of Sheep Milk Extracellular Vesicle-miRNA by Sequencing and Comparison with Cow Milk. Animals (Basel) 2020, 10. [Google Scholar] [CrossRef]
- Kirchner, B.; Pfaffl, M.W.; Dumpler, J.; von Mutius, E.; Ege, M.J. microRNA in native and processed cow's milk and its implication for the farm milk effect on asthma. J Allergy Clin Immunol 2016, 137, 1893–1895. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.C.; Fujikawa, T.; Maemura, T.; Ando, T.; Kitahara, G.; Endo, Y.; Yamato, O.; Koiwa, M.; Kubota, C.; Miura, N. Inflammation-related microRNA expression level in the bovine milk is affected by mastitis. PLoS One 2017, 12, e0177182. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ou-Yang, W.X.; Zhang, H.; Jiang, T.; Tang, L.; Tan, Y.F.; Luo, H.Y.; Xiao, Z.H.; Li, S.J. MiR-125b enhances autophagic flux to improve septic cardiomyopathy via targeting STAT3/HMGB1. Exp Cell Res 2021, 409, 112842. [Google Scholar] [CrossRef] [PubMed]
- Luoreng, Z.M.; Wei, D.W.; Wang, X.P. MiR-125b regulates inflammation in bovine mammary epithelial cells by targeting the NKIRAS2 gene. Vet Res 2021, 52, 122. [Google Scholar] [CrossRef] [PubMed]
- AHDB. https://ruminanthw.org.uk/wp-content/uploads/2021/05/Cattle-and-sheep-HW-priorities-survey-May-2021-FINAL.pdf. (accessed on 29th December 2023).
- Whatford, L.; van Winden, S.; Häsler, B. A systematic literature review on the economic impact of endemic disease in UK sheep and cattle using a One Health conceptualisation. Prev Vet Med 2022, 209, 105756. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.M.; Willis, Z.N.; Blakeley, M.; Lovatt, F.; Purdy, K.J.; Green, L.E. Bacterial species and their associations with acute and chronic mastitis in suckler ewes. J Dairy Sci 2015, 98, 7025–7033. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.; Smith, E.M.; Green, L.E. A longitudinal study of factors associated with acute and chronic mastitis and their impact on lamb growth rate in 10 suckler sheep flocks in Great Britain. Prev Vet Med 2016, 127, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Izumi, H.; Kosaka, N.; Shimizu, T.; Sekine, K.; Ochiya, T.; Takase, M. Bovine milk contains microRNA and messenger RNA that are stable under degradative conditions. J Dairy Sci 2012, 95, 4831–4841. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, M.A. microRNAs and the immune response. Trends Immunol 2008, 29, 343–351. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, Y.; Fan, Q.; Li, Z.; Qi, X.; Chen, F.; Xu, L.; Zhou, S.; Chen, X.; Li, Y.; et al. MiR-155 promotes acute organ injury in LPS-induced endotoxemic mice by enhancing CCL-2 expression in macrophages. Shock 2023. [Google Scholar] [CrossRef]
- Zhou, M.; Barkema, H.W.; Gao, J.; Yang, J.; Wang, Y.; Kastelic, J.P.; Khan, S.; Liu, G.; Han, B. MicroRNA miR-223 modulates NLRP3 and Keap1, mitigating lipopolysaccharide-induced inflammation and oxidative stress in bovine mammary epithelial cells and murine mammary glands. Vet Res 2023, 54, 78. [Google Scholar] [CrossRef]
- Li, Y.X.; Jiao, P.; Wang, X.P.; Wang, J.P.; Feng, F.; Bao, B.W.; Dong, Y.W.; Luoreng, Z.M.; Wei, D.W. RNA-seq reveals the role of miR-223 in alleviating inflammation of bovine mammary epithelial cells. Res Vet Sci 2023, 159, 257–266. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of Animal and Marine Origin: Structure, Function, and Anti-Inflammatory Properties. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Scherer, L.; Behrens, P.; Tukker, A. Opportunity for a dietary win-win-win in nutrition, environment, and animal welfare. One Earth 2019, 1, 349–360. [Google Scholar] [CrossRef]
Figure 1.
Representative flow cytometry to determine Vesidye positive EV in each sample. A small EV, total events; B small EV Vesidye positive (gate P1); C large EV total events; D Vesidye positive (gate P1).
Figure 1.
Representative flow cytometry to determine Vesidye positive EV in each sample. A small EV, total events; B small EV Vesidye positive (gate P1); C large EV total events; D Vesidye positive (gate P1).
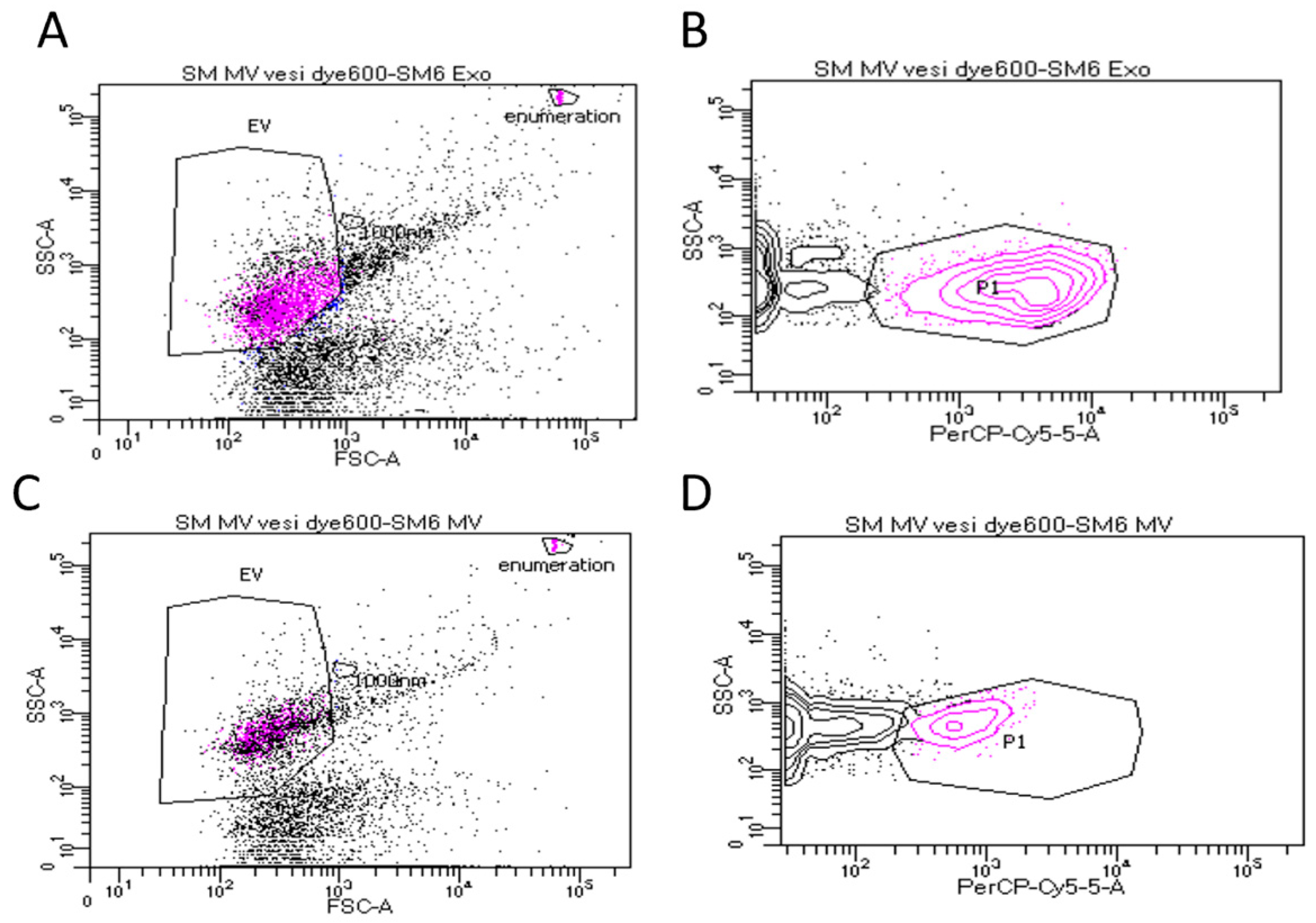
Figure 2.
Measurement of ROS in THP-1 monocytes and macrophages after incubation with SMEV. THP-1 monocytes (A) or PMA- differentiated THP-1 macrophages (B) were pre-loaded with dihydrorhodamine -1,2,3 and then incubated for 24h in the presence or absence of DMNQ. All wells in triplicate. n=6 LEV and sEV samples from individual sheep. One way ANOVA followed by Fishers LSD. * p<0.05, ** p<0.01, *p<0.001.
Figure 2.
Measurement of ROS in THP-1 monocytes and macrophages after incubation with SMEV. THP-1 monocytes (A) or PMA- differentiated THP-1 macrophages (B) were pre-loaded with dihydrorhodamine -1,2,3 and then incubated for 24h in the presence or absence of DMNQ. All wells in triplicate. n=6 LEV and sEV samples from individual sheep. One way ANOVA followed by Fishers LSD. * p<0.05, ** p<0.01, *p<0.001.
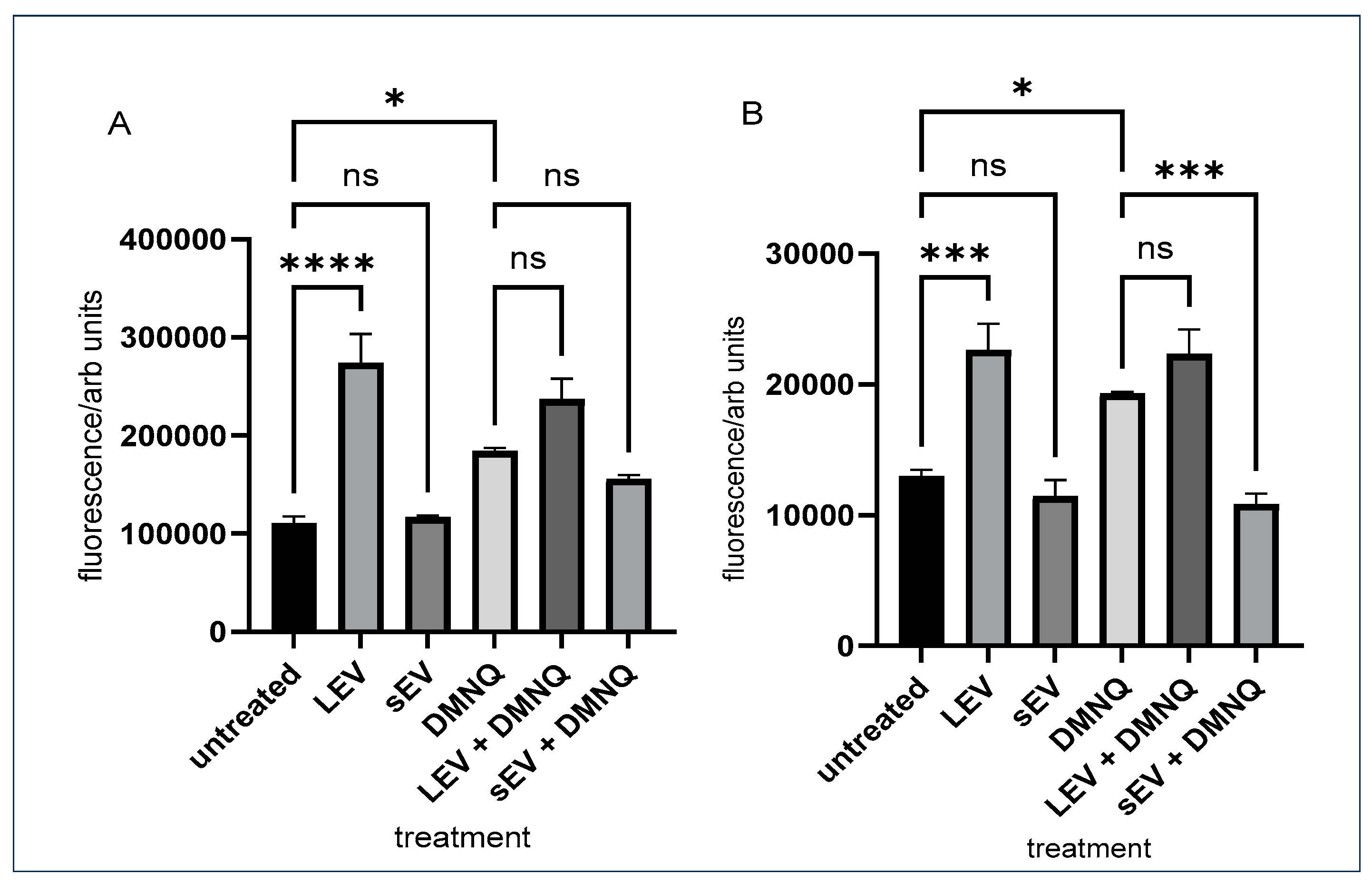
Figure 3.
SMEV reduces CCL2 secreted from THP-1 monocytes but not macrophages. THP-1 monocytes (A) or PMA- differentiated THP-1 macrophages (B) were incubated for 24h in the presence or absence of LPS and SMEV. All wells in triplicate. n=6 LEV and sEV samples from individual sheep. One way ANOVA followed by uncorrected Fishers LSD. * p<0.05, ** p<0.01, *p<0.001.
Figure 3.
SMEV reduces CCL2 secreted from THP-1 monocytes but not macrophages. THP-1 monocytes (A) or PMA- differentiated THP-1 macrophages (B) were incubated for 24h in the presence or absence of LPS and SMEV. All wells in triplicate. n=6 LEV and sEV samples from individual sheep. One way ANOVA followed by uncorrected Fishers LSD. * p<0.05, ** p<0.01, *p<0.001.
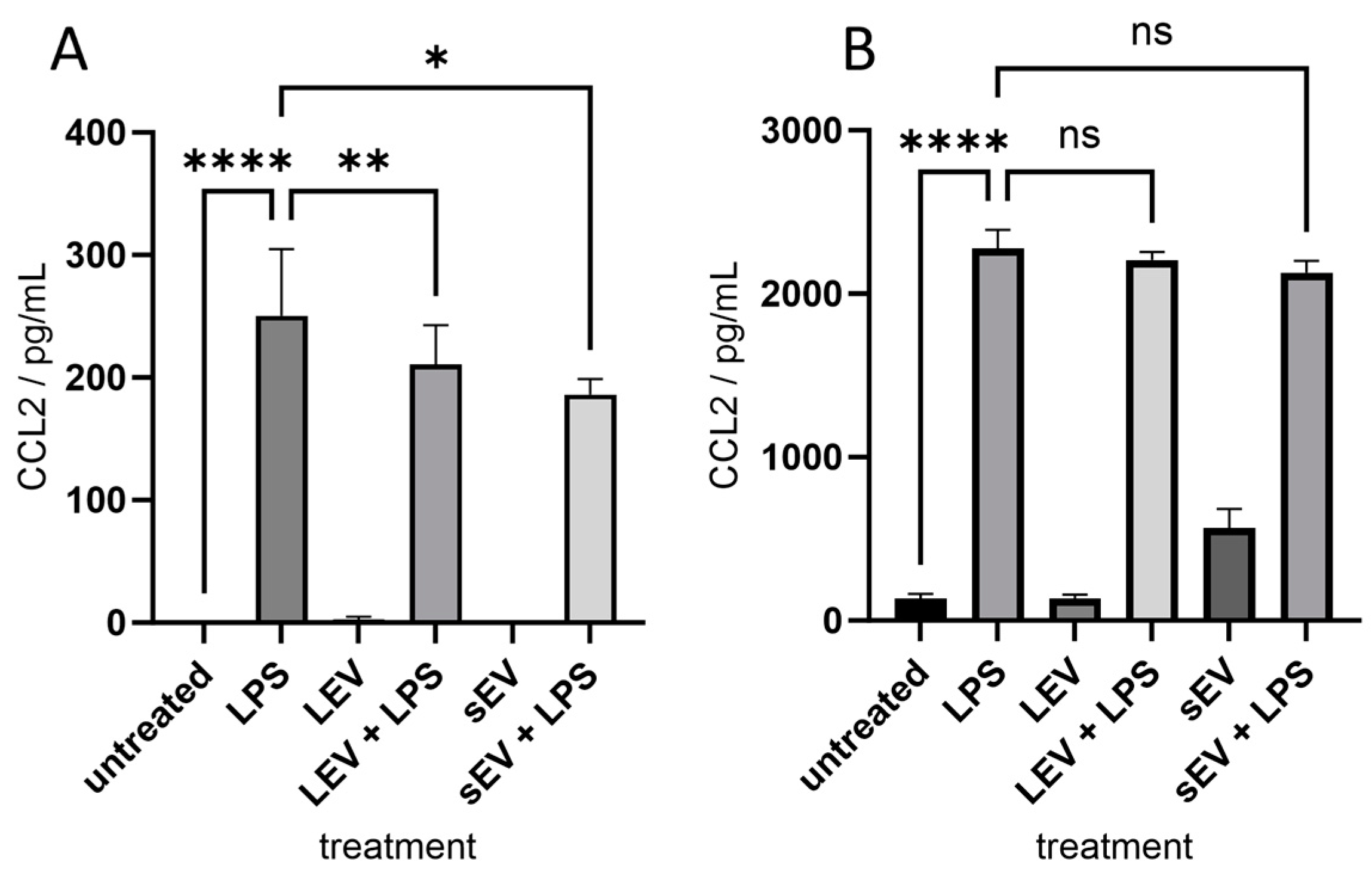
Figure 4.
Cycle threshold measurements of miRNA in SM-EV. Total RNA was isolated from LEV (A) and sEV (B), converted to cDNA and qRT-PCR was performed for miR26a, miR92a, miR125b, miR155, miR223. Water controls were carried out in triplicate (no signal detected for any sample – data not shown). All samples were in triplicate. Individual Ct n=6 LEV and sEV samples from individual sheep.
Figure 4.
Cycle threshold measurements of miRNA in SM-EV. Total RNA was isolated from LEV (A) and sEV (B), converted to cDNA and qRT-PCR was performed for miR26a, miR92a, miR125b, miR155, miR223. Water controls were carried out in triplicate (no signal detected for any sample – data not shown). All samples were in triplicate. Individual Ct n=6 LEV and sEV samples from individual sheep.
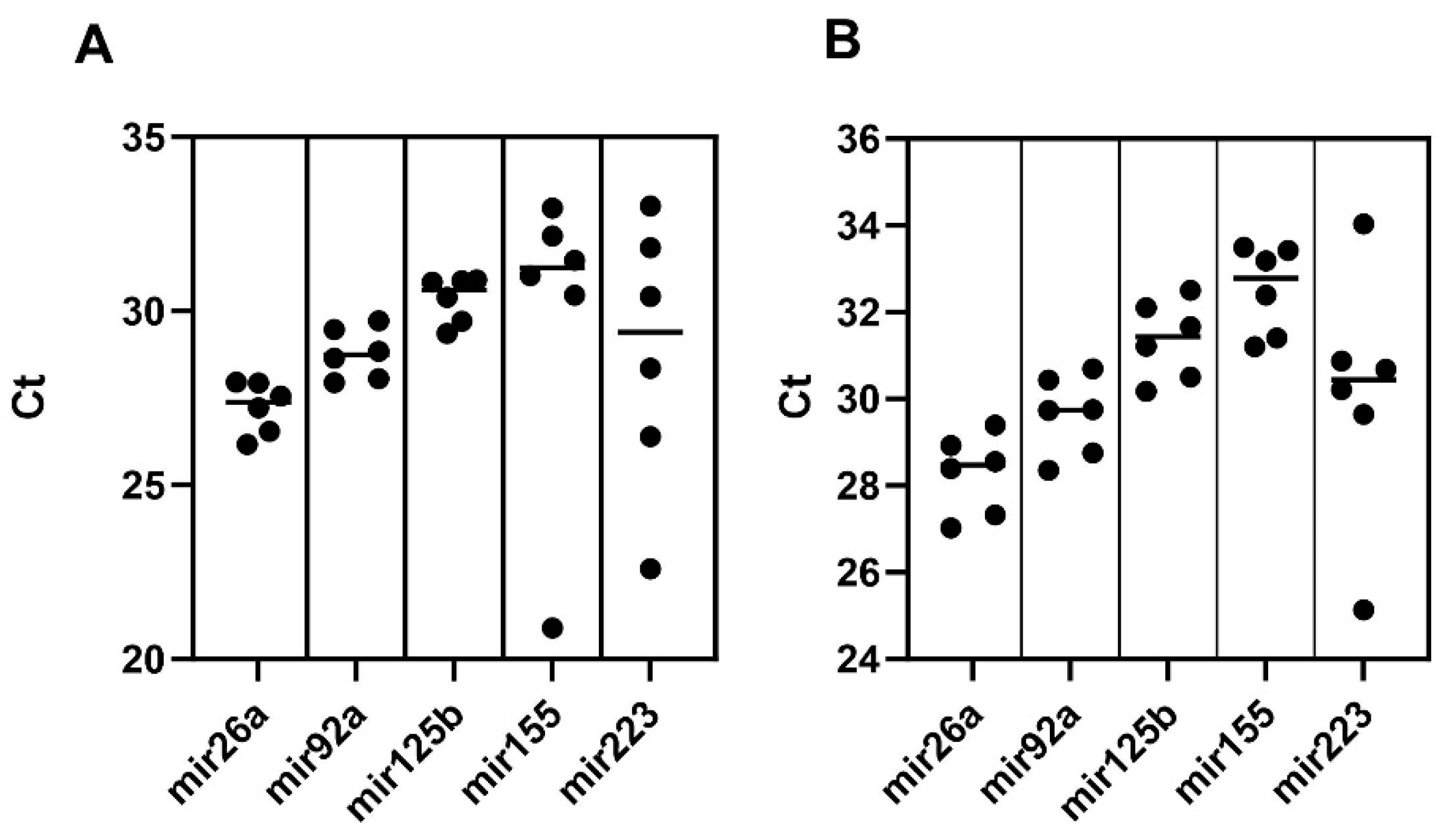

Table 1.
characteristics of individual sheep milk derived EV samples.
| small EV | large EV | |||||
|---|---|---|---|---|---|---|
| total RNA (ng/µl) | total protein (µg/µl) | VesiDye positive EV/µl | total RNA (ng/µl) | total protein (µg/µl) | VesiDye positive EV/µl | |
| SMEV1 | 22.1 | 145.58 | 18411.52 | 59.053 | 1675.15 | 90541.44 |
| SMEV2 | 52.0 | 140.23 | 26384.11 | 159.075 | 1915.58 | 39303.23 |
| SMEV3 | 23.0 | 153.25 | 36564.71 | 79.977 | 1915.85 | 17200.00 |
| SMEV4 | 100.3 | 127.4 | 32495.47 | 92.803 | 1414.13 | 134833.3 |
| SMEV5 | 49.4 | 197.95 | 23083.8 | 54.648 | 1810.05 | 56615.38 |
| SMEV6 | 39.0 | 136.35 | 30837.61 | 54.485 | 1618.4 | 18077.92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



