Submitted:
29 December 2023
Posted:
03 January 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Coronaviruses are a large family of diverse enveloped RNA viruses found in numerous animal species. They are well-known for their ability to cross species barriers and have been transmitted from bats or intermediate hosts to humans on several occasions. Four of the seven human coronaviruses (hCoVs) are responsible for approximately 20% of common colds (hCoV-229E, -NL63, -OC43, -HKU1). Two others (SARS-CoV-1 and MERS-CoV) cause severe and frequently lethal respiratory syndromes but have only spread to very limited extents in human populations. In contrast, SARS-CoV-2, while exhibiting intermediate pathogenicity, has a profound impact on public health due to its enormous spread. In this review, we discuss which initial features of the SARS-CoV-2 Spike protein and subsequent adaptations to the new human host may have helped this pathogen to cause the COVID-19 pandemic. Our focus is on host forces driving changes in the Spike protein and their consequences for virus infectivity, pathogenicity, immune evasion and resistance to preventive or therapeutic agents. In addition, we discuss the significance and perspectives of broad-spectrum therapeutics and vaccines.
Keywords:
SARS-CoV-2
; Sarbecovirus
; Spike
; mutation
; manifestation
; immune evasion
; zoonoses
Introduction
Coronaviridae represent a large family of diverse enveloped single-strand RNA viruses that received their name from the crown-like appearance of their Spike surface glycoproteins. Bats and rodents are considered the reservoir species of most coronaviruses1. However, coronaviruses are notorious for their ability to cross species barriers2,3. Consequently, they have been detected in many animal species and were successfully transmitted to humans at least seven times4. Four of the seven human coronaviruses (229E, NL63, OC43 and HKU1) cause mild respiratory infections and are responsible for about 20% to 30% of common colds5,6. The remaining three cause severe respiratory diseases, with lethality rates ranging from approximately 1% (SARS-CoV-2) to around 10% (SARS-CoV-1) and 40% (MERS-CoV)7,8. SARS-CoV-2 was most likely transmitted to humans from bats or an intermediate host at the end of 2019 and caused the COVID-19 pandemic6,9–12. To date, SARS-CoV-2 has infected more than 800 million people. Reliable and affordable tests, as well as effective vaccines and therapeutics, have been developed in an amazingly short time. However, emerging new SARS-CoV-2 variants that at least partially escape immunity generated by previous infection or vaccination and/or show increased replication and transmissibility continue to circulate in the human population.
The overall high diversity of different members of the coronavirus family and the ongoing emergence of new SARS-CoV-2 variants through mutation and recombination give the impression that coronaviruses mutate and diversify very rapidly. However, mutation and diversification rates of coronaviruses are actually much lower compared to other RNA viruses, such as human immunodeficiency virus (HIV) or hepatitis C virus (HCV)13,14. The reason for this is that the coronavirus polymerase has a proofreading activity, which rectifies errors during replication—a trait rather rare in the realm of RNA viruses15,16. For SARS-CoV-2 mutation rates of 1.3 × 10-6 per base and infection cycle have been reported14. Thus, coronaviruses are at the low end of the spectrum of 10-4 to 10-6 nucleotide substitutions per replication cycle reported for RNA viruses and approximating those of DNA viruses, which range from 10-6 to 10-8 13. Proof-reading activity also allows coronaviruses to possess one the largest genomes among RNA viruses, approximately 30 kilobases in length and encoding for about 30 proteins17,18.
Mutations, recombination and ongoing replication are prerequisites for virus diversification. Usually, however, only changes providing a selection advantage for viral spread will be enriched and manifested in virus populations19,20. The forces driving fitness advantages vary and change upon zoonotic transmission. For example, it has been reported that SARS-CoV-2 initially evolved alterations increasing its replication and transmission fitness in the new human host19,21. After infection and/or vaccination of large parts of the human population, however, selection pressure to evade humoral immune responses increased and drove the emergence of new SARS-CoV-2 variants19,21,22. In some cases, initial changes allowing humoral immune evasion came at the cost of decreased infectivity but subsequent changes restored viral replication fitness21,23. The ongoing arms race between immune control and viral evasion drives the constant evolution of new SARS-CoV-2 variants.
As of December 2023, more than 13 billion COVID-19 vaccine doses have been administered (WHO COVID-19 dashboard) and most people have some immunity against SARS-CoV-2 through vaccination and/or previous infection. Thus, SARS-CoV-2 lost most of its fright, although it continuous to circulate and evolve within the human population. Thus, new questions arise, such as whether vaccines and therapeutics that are currently available or under development can protect us against newly emerging SARS-CoV-2 variants and future zoonoses of animal coronaviruses. Here, we address some of the factors driving the evolution of SARS-CoV-2 Spike proteins and their consequences for viral fitness and sensitivity to vaccines, neutralizing antibodies (nAbs), and Spike-targeting therapeutics. In addition, we discuss whether broad or even general protection against new SARS-CoV-2 variants and future zoonoses of animal coronaviruses may be feasible.
Early features of the SARS-CoV-2 Spike protein
The Spike proteins of different members of the coronavirus family show high sequence divergence24. SARS-CoV-2 is phylogenetically closely related to some bat CoVs, such as BANAL-20-52 and RaTG13 (Figure 1)9,25,26. The Spike protein of SARS-CoV-2 shows ~96% amino acid sequence homology to its closest bat relatives with most variations being located in the receptor binding domain (RBD) and the N-terminal domain (NTD) (Figure 2). Just like the Spike proteins of SARS-CoV-1 and hCoV-NL63, SARS-CoV-2 utilizes the angiotensin-converting enzyme 2 (ACE2) receptor for infection of human target cells27–29. Thus, ACE2 usage seems helpful but not essential for zoonotic transmission of coronaviruses. The Spike proteins of several coronaviruses detected in bats or intermediate host efficiently use human ACE2 for infection30,31. In comparison, the original RaTG13 Spike is poorly infectious in human cells but a single T403R change allows efficient usage of ACE2 receptors from various species including humans31,32. Most Spike proteins of related bat coronaviruses contain an R or K residue at position 403 of the viral Spike protein and are capable of infecting human cells. Thus, SARS-CoV-2 was most likely able to efficiently use human ACE2 immediately after cross-species transmission. A variety of alternative receptors have been reported to promote ACE2-independent entry into human cells but their relevance for SARS-CoV-2 replication and transmission in vivo is poorly understood33.
The SARS-CoV-2 Spike protein is characterized by two proteolytic cleavage sites: the S1/S2 and the S2' site (Figure 2), which may have played a significant role in SARS-CoV-2 transmission and evolution33. The S1/S2 cleavage site is located at the boundary of the S1/S2 subunits of the Spike protein and distinguishes SARS-CoV-2 from related animal coronaviruses35,36. This polybasic site comprises an insertion (680SPRRAR↓SV687), forming a cleavage motif (RxxR) for furin-like enzymes, enabling proteolytic activation of the Spike protein promoting entry into host cells37. The polybasic site allows Spike cleavage during virus packaging, thereby significantly enhancing viral transmissibility and expanding its tissue tropism36. The furin cleavage site enhances the ability of SARS-CoV-2 to infect certain cell types and induce cell-cell fusion, which may promote efficient viral spread38. Thus, it is thought that this site played a key role in the rapid spread of the COVID-19 pandemic. The origin of the S1/S2 furin cleavage site in SARS-CoV-2 Spike, particularly the question whether it was present before or after zoonotic spillover, has raised significant interest and is still under debate39–41. Some animal coronaviruses carrying a polybasic furin cleavage site in their Spike protein have been reported42. However, furin cleavage sites are not present in pangolin or bat coronaviruses that are closely related to SARS-CoV-240. Thus, it has been suggested that this specific cleavage site developed early in the process of the virus adapting to its human host40, although the possibility that it predisposed an animal virus for efficient zoonotic transmission cannot be excluded43. Several SARS-CoV-2 variants of concern (VOCs) manifested mutations near the S1/S2 furin cleavage site, that altered Spike processing efficiency. For instance, the early Alpha and all subsequent VOCs acquired the P681H mutation increasing cleavage efficiency44. Additionally, the Omicron-specific N679K mutation hampered Spike processing and conferred a shift toward upper airway replication in hamster models45.
The second step of proteolytic activation is mediated by S2' cleavage. It liberates the fusion peptide of the S2 subunit, enabling its insertion into the cellular membrane and subsequent formation of the 6-helix bundle mediating fusion of the viral envelope with the host cell membrane, a step crucial for viral entry into the host cell46,47. Two main types of host cell proteases, Transmembrane Serine Protease 2 (TMPRSS2) and Cathepsins, are involved in this process, and each plays a distinct role depending on the cellular entry pathway of the virus. When SARS-CoV-2 binds to the ACE2 receptor on the host cell surface, TMPRSS2 cleaves the spike protein at the S2' site27. In contrast, Cathepsins become involved when the virus enters cells via the endosomal pathway and fusion is triggered by the acidic environment of the endosome. Notably, it has been reported that the less efficient Spike cleavage of Omicron BA.1 Spike at S1/S2 is associated reduced dependency on TMPRSS2 and a shift towards Cathepsin-dependent endosomal entry. This shift in cellular tropism away from TMPRSS2-expressing cells is largely mediated by a H655Y substitution in Spike and may impact viral pathogenesis48–50.
Both, the S1/S2 and S2’ cleavage sites of the Spike protein play a crucial role in the ability of SARS-CoV-2 to infect human cells and have been a focal point in understanding the virus's transmission dynamics and pathogenicity. The evolution of these sites in different VOCs may have significant implications for transmissibility, disease severity, and vaccine efficacy. Thus, continued monitoring and research into these mutations are essential for managing the pandemic and developing effective countermeasures.
In addition to using ACE2 and cellular protease, the Spike protein of SARS-CoV-2 has been shown to hijack usually antiviral IFITM2 and IFITM3 proteins for efficient infection51–54. Furthermore, it has been reported that the SARS-CoV-2 Spike counteracts the restriction factor tetherin that otherwise inhibits the release of viral particles55. However, if these mechanisms are conserved in the coronavirus family and to which extent they contributed to the efficient spread of SARS-CoV-2 remains to be determined.
Initial human adaptation of SARS-CoV-2 Spike proteins
Common cold coronaviruses, which circulate in the human population, are highly divergent from SARS-CoV-2. In contrast, SARS-CoV-1 Spike proteins share about 76% homology with that of SARS-CoV-2 and might induce cross-neutralizing antibodies56. However, SARS-CoV-1 infected only ~8.000 individuals and has fortunately disappeared. Thus, SARS-CoV-2 essentially hit an immunologically naïve population and initial adaptations to humans manifested in changes that increased viral infectivity and transmission. For example, mutation of D614G in the receptor-binding domain (RBD) of the Spike became prevalent during the first few months of the pandemic, indicating a significant selective advantage for the spread of SARS-CoV-2 in human populations57–60. It has been reported that D614G alters Spike configuration, and enhances viral replication in human cells, as well as in the human respiratory tract, hence increasing transmission rates but not pathogenicity59–63. Another mutation that emerged relatively rapidly is N501Y thought to increase the binding affinity of the Spike protein to the ACE2 receptor64,65. Additionally, evolution of SARS-CoV-2 codon usage and the slower-than-expected acquisition of mutations, hinting at a purifying selection during the initial phase of the pandemic, provide insight into the virus's genomic adaptation strategies66,67. Alterations in the RBD of the Spike protein, which increased the affinity for the human ACE2 receptor were suggested to represent adaptive steps critical for efficient human-to-human transmission of SARS-CoV-268,69. Since its emergence, thousands of mutations have been observed, with novel variations continuously emerging as the virus replicates and spreads across the human population70.
Evasion of adaptive immunity
After SARS-CoV-2 infection and/or vaccination of significant parts of the human population, selection pressures shaping the evolution of Spike shifted from alterations increasing viral infection and replication fitness to mutations allowing evasion of humoral immune responses19,21. The initial variants of concern (VOCs), named Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), and Delta (B.1.617.2) all emerged independently. Each of these variants contained about 6 to 8 changes in the Spike protein, most of them in the RBD (Figure 2), promoting mainly immune evasion. However, changes affecting ACE2 binding and increasing fusogenicity have also been reported21,71. This has changed with the emergence of the Omicron VOCs that contained a strikingly high number of changes, especially in the viral Spike protein, compared to all previous SARS-CoV-2 variants72. The initial Omicron BA.1 VOC outcompeted the previously dominating Delta VOC at enormous speed, although it displayed low infectivity and replication fitness in many cell culture systems and animal models49,73–75. It was itself outcompeted by BA.2 which differs by ~20 amino acid changes in Spike from BA.1. All subsequent and current VOCs originated from BA.2 and contain further amino acid changes in their Spike proteins. Accumulating evidence suggests that initial mutations facilitated viral immune evasion at the cost of reduced infectivity and subsequent changes restored infectiousness and replication fitness23,76. New variants of Omicron are constantly emerging showing mutations in the RBD and the N-terminal domain (NTD) of Spike allowing immune evasion77–79 (Figure 2) leading to the simultaneous emergence of sub-variants, each characterized by mutations that converge on several hotspots of their RBDs. Specific strains like BQ.1.1.10, BA.4.6.3, XBB, and CH.1.1 were identified as highly antibody-evasive80,81. This phenomenon of convergent evolution is driven in part by the humoral immune pressure, which promotes the evolution of the virus in a way that helps it evade nAbs82. Just like common cold coronaviruses, infection with one strain of SARS-CoV-2 does not efficiently protect against infection with another strain or newly emerging variants. For example, the XBB lineages that currently dominate the pandemic are largely resistant against neutralization by humoral immune responses induced by infection with earlier SARS-CoV-2 variants (including BA.1, BA.2 and BA4/5) or vaccination83–86. The SARS-CoV-2 variant of interest BA.2.86 that was first isolated in July 2023, has 36 amino acid substitutions compared to XBB.1.5, many of them located in key antigenic sites of the Spike protein. The proportion of BA.2.86 and closely related descendent lineages, such as JN.1 characterized by an additional substitution of L455S in the Spike protein, is currently steadily increasing indicating high transmission fitness and efficient humoral immune evasion87. However, T cell responses may remain effective and prevent severe disease in most cases88,89. Altogether, SARS-CoV-2 continues to infect humans but in relatively stable numbers and has transitioned from the pandemic to the endemic phase90,91.
Broadly acting vaccines or therapeutics targeting the SARS-CoV-2 Spike protein
The evolution of new SARS-CoV-2 variants also conferred resistance to therapeutic antibodies and vaccines. For example, neutralizing antibodies (nAbs) against SARS-CoV-2 initially inhibited a variety of virus strains and showed great promise in treating and preventing infections92. Initially developed nAbs target epitopes in the RBD that overlap the ACE2 receptor-binding site (RBS), thus sterically hindering binding of the Spike glycoprotein to its receptor. However, emerging SARS-CoV-2 variants show resistance to essentially all first generation FDA approved monoclonal antibodies (Figure 3)93,94. Specifically, mutations of E484A/K and Q493R in the RBD of the Spike protein rendered SARS-CoV-2 resistant to Bamlanivimab and N440K and G446S to Imdevimab76,95. The Omicron variants have shown an impressive capability to evade even those nAbs that target more conserved domains in the Spike RBD region96,97. However, while coronavirus Spike proteins are highly variable and tolerate numerous changes in their RBD some structural and functional features are highly conserved, offering perspectives for broad-spectrum antiviral agents.
Broadly neutralizing antibodies (bnAbs) designed to target highly conserved regions of the S2 region of the Spike protein including the fusion peptide and Stem-helix regions combine the potential for pan coronavirus activity with a high genetic barrier to evasion 98. S2 stem helix binding bnAbs, isolated from SARS-CoV-2 recovered-vaccinated donors, showed broad cross protection against SARS-CoV-1, SARS-CoV-2, and MERS-CoV in mouse mdels98. Abs targeting the SARS-CoV-2 Spike fusion peptide not only reduced viral fusion, but additionally impaired proteolytic maturation of the Spike glycoprotein99. Thus, bnAbs targeting conserved domains in the S2 region of the viral Spike protein are promising candidates for next-generation pan-coronavirus vaccine development.
Peptide inhibitors are increasingly recognized in antiviral drug development due to their high specificity and biocompatibility. Similar to pan-coronavirus bnAbs, targeting conserved sequence and fusion mechanisms of the SARS-CoV-2 Spike protein S2 domain allows for the development of broadly, pan-coronavirus antiviral peptides. ACE2-mimicking, H1 derived peptides, are designed to mimic the key functional elements of the Spike protein - ACE2 interaction100–102. These peptides aim to competitively inhibit the binding of the virus to its ACE2 receptor, thereby blocking viral entry into host cells. An approach based on the principle of molecular mimicry, where the peptides structurally resemble the ACE2 interface that interacts with the spike protein of the virus100. Receptor usage by the SARS-CoV-2 VOCs is consistent, and although other receptors for SARS-CoV-2 have been identified, a switch in main receptor preference appears improbable. Thus, peptides mimicking ACE2 may effectively inhibit SARS-CoV-2 infectivity and offer a promising direction for developing long-lasting anti-SARS-CoV-2 treatments.
Just like many other enveloped viruses, such as HIV-1, entry of SARS-CoV-2 requires insertion of a fusion peptide into the cellular membrane and subsequent formation of a six-helix bundle between heptad repeat regions (HR1 and HR2) that pull the viral and cellular membranes together to mediate fusion. For instance, the peptide EK1 interacts with the highly conserved HR2 domain of the S2 subunit of the Spike protein (Figure 2), thereby preventing the interaction between HR1 and HR2 and hence virus-cell fusion103. Interestingly, EK1 and its optimized derivative (EK1C4) not only display broad or even pan activity against coronaviruses but even show cross-activity against HIV104. Mutation of N969K in Omicon Spike proteins induces substantial changes in the structure of the HR2 backbone in the HR1/HR2 post-fusion bundle. Nonetheless, EK1 and EK1C4 inhibit membrane fusion mediated by Spike proteins of Omicron subvariants105. In addition, inhibitors of the proteases activating coronavirus Spike proteins, i.e. furin, TMPRSS2 and Cathepsins may offer perspectives for broad-based inhibitors106–108.
The initial vaccines developed against SARS-CoV-2 were designed based on the Hu-1 variant, the earliest strain of the virus109–111. These vaccines, including mRNA-based vaccines, like BNT162b2 (Pfizer-BioNTech) or mRNA-1273 (Moderna), vector-based vaccines like Vaxzevria (Astra Zeneka), or inactivated virus vaccines like CoronaVac (Sinovac) were highly effective in reducing COVID-19 pathogenicity and mortality112,113. In response to the highly immune evasive SARS-CoV-2 Omicron BA.4/BA.5 and XBB variants, adapted vaccines, like the bivalent Spikevax (Moderna) or the Omicron XBB1.5 adapted variants of BNT162b2, were designed to better match circulating strains114. Authorized for use, these vaccines aim to offer broader protection against COVID-19, preventing hospitalization and death due to infection. First studies demonstrate the enhanced effectiveness of these bivalent boosters against currently circulating SARS-CoV-2 variants compared to the original monovalent vaccinations114. Several strategies have been pursued to induce broad protection against diverse coronaviruses. For example, a trivalent sortase-conjugate nanoparticle vaccine that contained RBDs from SARS-CoV-2, RsSHC014 (a bat coronavirus), and MERS-CoV elicited neutralizing antibody responses against these viruses115. Another approach is based on mRNA vaccines designed to express chimeric Spike proteins aiming to elicit protection against a range of Sarbecoviruses, including SARS-CoV, SARS-CoV-2, and various bat coronaviruses116. However, neither previous coronavirus infections nor current vaccines confer long-term protection against all newly emerging SARS-CoV-2 variants. Thus, whether and how long-lasting and broad protection e.g. by alternative vaccination strategies can be achieved remains to be determined.
Conclusions and future perspectives
Coronaviruses have already been successfully transmitted from animals to humans on at least seven independent occasions. Furthermore, exposure to diverse Sarbecoviruses has been identified among high-risk human communities, providing epidemiological and immunological evidence that zoonotic spillover is continuously occurring117. Thus, broadly and long-lasting therapeutic and preventive agents are highly desirable. Human immune responses and newly emerging SARS-CoV-2 variants are in an arms race. Notably, even the most recent and divergent SARS-CoV-2 variants show only 3.3% (43 mutations in Omicron XBB1.5) to 3.5% (45 mutations in Omicron EG.5.1) amino acid diversity from the Spike proteins of the early virus strains. This is sufficient to confer efficient resistance against humoral immune responses induced by previous infection or vaccination. In comparison, bat coronaviruses and SARS-CoV-1 showing up to 30% amino acid diversity in Spike from SARS-CoV-2, are efficiently neutralized31,56,118,119. This illustrates the enormous power of the selection pressures in humans driving those changes in viral Spike proteins that allow efficient immune escape. On the positive side, this suggests that immunity induced by SARS-CoV-2 infections and vaccination might confer significant protection against newly emerging animal coronaviruses. However, it will be important to monitor if constant chasing of newly emerging SARS-CoV-2 variants by regular vaccine adaptation of may lead to immunogens showing increasing diversity from the initial zoonotic pathogen. This raises the possibility that future vaccines could be effective against new SARS-CoV-2 variants but might lose activity against animal coronaviruses, thereby showing reduced potential in preventing future pandemics. Altogether, immunogens conferring broad protective responses are highly desirable and may also help to design effective preventive strategies against other viral pathogens.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
F.Z. and F.K. wrote the manuscript. F.Z. and C.J. drafted the figures and F.K. modified and revised them. T.J. provided resources and supervised structural analyses.
Acknowledgments
We thank Konstantin Sparrer, Dre van der Merwe and Dorota Kmiec for critical reading of the article and helpful discussions. This work was supported by the Deutsche Forschungsgemeinschaft (CRC 1279). We apologize to all authors whose interesting studies could not be cited due to space limitations.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Banerjee, A., Kulcsar, K., Misra, V., Frieman, M., and Mossman, K. (2019). Bats and Coronaviruses. Viruses 11. [CrossRef]
- Mahdy, M.A.A., Younis, W., and Ewaida, Z. (2020). An Overview of SARS-CoV-2 and Animal Infection. Frontiers in Veterinary Science 7. [CrossRef]
- Dhama, K., Patel, S.K., Sharun, K., Pathak, M., Tiwari, R., Yatoo, M.I., Malik, Y.S., Sah, R., Rabaan, A.A., Panwar, P.K., et al. (2020). SARS-CoV-2 jumping the species barrier: Zoonotic lessons from SARS, MERS and recent advances to combat this pandemic virus. Travel Med Infect Dis 37. 101830. [CrossRef]
- Ye, Z.W., Yuan, S., Yuen, K.S., Fung, S.Y., Chan, C.P., and Jin, D.Y. (2020). Zoonotic origins of human coronaviruses. Int J Biol Sci. 16. [CrossRef]
- Corman, V.M., Muth, D., Niemeyer, D., and Drosten, C. (2018). Hosts and Sources of Endemic Human Coronaviruses. In Advances in Virus Research 100. [CrossRef]
- Cui, J., Li, F., and Shi, Z.L. (2019). Origin and evolution of pathogenic coronaviruses Nature Reviews Microbiology 17. [CrossRef]
- Ksiazek, T.G., Erdman, D., Goldsmith, C.S., Zaki, S.R., Peret, T., Emery, S., Tong, S., Urbani, C., Comer, J.A., Lim, W., et al. (2003). A novel coronavirus associated with severe acute respiratory syndrome. New England Journal of Medicine 348. 1953–1966. [CrossRef]
- Bermingham, A., Chand, M.A., Brown, C.S., Aarons, E., Tong, C., Langrish, C., Hoschler, K., Brown, K., Galiano, M., Myers, R., et al. (2012). Severe respiratory illness caused by a novel coronavirus, in a patient transferred to the United Kingdom from the Middle East, September 2012. Eurosurveillance 17. [CrossRef]
- Zhou, P., Yang, X.L., Wang, X.G., Hu, B., Zhang, L., Zhang, W., Si, H.R., Zhu, Y., Li, B., Huang, C.L., et al. (2020). A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579. [CrossRef]
- Boni, M.F., Lemey, P., Jiang, X., Lam, T.T.Y., Perry, B.W., Castoe, T.A., Rambaut, A., and Robertson, D.L. (2020). Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nature Microbiology 5. [CrossRef]
- Worobey, M., Levy, J.I., Malpica Serrano, L., Crits-Christoph, A., Pekar, J.E., Goldstein, S.A., Rasmussen, A.L., Kraemer, M.U.G., Newman, C., Koopmans, M.P.G., et al. (2022). The Huanan Seafood Wholesale Market in Wuhan was the early epicenter of the COVID-19 pandemic. Science 377. [CrossRef]
- Hao, X., Cheng, S., Wu, D., Wu, T., Lin, X., and Wang, C. (2020). Reconstruction of the full transmission dynamics of COVID-19 in Wuhan. Nature 584. [CrossRef]
- Sanjuán, R., Nebot, M.R., Chirico, N., Mansky, L.M., and Belshaw, R. (2010). Viral mutation rates. J Virol 84. [CrossRef]
- De Maio, N., Walker, C.R., Turakhia, Y., Lanfear, R., Corbett-Detig, R., and Goldman, N. (2021). Mutation Rates and Selection on Synonymous Mutations in SARS-CoV-2. Genome Biol Evol 13. [CrossRef]
- Moeller, N.H., Shi, K., Demir, Ö., Belica, C., Banerjee, S., Yin, L., Durfee, C., Amaro, R.E., and Aihara, H. (2022). Structure and dynamics of SARS-CoV-2 proofreading exoribonuclease ExoN. Proceedings of the National Academy of Sciences 119. [CrossRef]
- Eckerle, L.D., Becker, M.M., Halpin, R.A., Li, K., Venter, E., Lu, X., Scherbakova, S., Graham, R.L., Baric, R.S., Stockwell, T.B., et al. (2010). Infidelity of SARS-CoV Nsp14-Exonuclease Mutant Virus Replication Is Revealed by Complete Genome Sequencing. PLOS Pathogens 6. [CrossRef]
- Cao, C., Cai, Z., Xiao, X., Rao, J., Chen, J., Hu, N., Yang, M., Xing, X., Wang, Y., Li, M., et al. (2021). The architecture of the SARS-CoV-2 RNA genome inside virion. Nat Commun 12. [CrossRef]
- Jones, A.N., Mourão, A., Czarna, A., Matsuda, A., Fino, R., Pyrc, K., Sattler, M., and Popowicz, G.M. (2022). Characterization of SARS-CoV-2 replication complex elongation and proofreading activity. Sci Rep 12. [CrossRef]
- Markov, P.V., Ghafari, M., Beer, M., Lythgoe, K., Simmonds, P., Stilianakis, N.I., and Katzourakis, A. (2023). The evolution of SARS-CoV-2. Nat Rev Microbiol, 21. [CrossRef]
- Focosi, D., and Maggi, F. (2022). Recombination in Coronaviruses, with a Focus on SARS-CoV-2. Viruses 14. [CrossRef]
- Carabelli, A.M., Peacock, T.P., Thorne, L.G., Harvey, W.T., Hughes, J., de Silva, T.I., Peacock, S.J., Barclay, W.S., de Silva, T.I., Towers, G.J., et al. (2023). SARS-CoV-2 variant biology: immune escape, transmission and fitness. Nat Rev Microbiol 21. [CrossRef]
- Chakraborty, C., Sharma, A.R., Bhattacharya, M., and Lee, S.-S. (2022). A Detailed Overview of Immune Escape, Antibody Escape, Partial Vaccine Escape of SARS-CoV-2 and Their Emerging Variants With Escape Mutations. Front Immunol 13. [CrossRef]
- Pastorio, C., Noettger, S., Nchioua, R., Zech, F., Sparrer, K.M.J., and Kirchhoff, F. (2023). Impact of mutations defining SARS-CoV-2 Omicron subvariants BA.2.12.1 and BA.4/5 on Spike function and neutralization. iScience 26. [CrossRef]
- Zmasek, C.M., Lefkowitz, E.J., Niewiadomska, A., and Scheuermann, R.H. (2022). Genomic evolution of the Coronaviridae family. Virology 570. [CrossRef]
- Temmam, S., Vongphayloth, K., Baquero, E., Munier, S., Bonomi, M., Regnault, B., Douangboubpha, B., Karami, Y., Chrétien, D., Sanamxay, D., et al. (2022). Bat coronaviruses related to SARS-CoV-2 and infectious for human cells. Nature 604. [CrossRef]
- Ou, X., Liu, Y., Lei, X., Li, P., Mi, D., Ren, L., Guo, L., Guo, R., Chen, T., Hu, J., et al. (2020). Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun 11. [CrossRef]
- Hoffmann, M., Kleine-Weber, H., Schroeder, S., Krüger, N., Herrler, T., Erichsen, S., Schiergens, T.S., Herrler, G., Wu, N.-H., Nitsche, A., et al. (2020). SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 181. [CrossRef]
- Hofmann, H., Pyrc, K., van der Hoek, L., Geier, M., Berkhout, B., and Pöhlmann, S. (2005). Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci U S A 102. [CrossRef]
- Letko, M., Marzi, A., and Munster, V. (2020). Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nature Microbiology 5. [CrossRef]
- Zheng, M., Zhao, X., Zheng, S., Chen, D., Du, P., Li, X., Jiang, D., Guo, J.-T., Zeng, H., and Lin, H. (2020). Bat SARS-Like WIV1 coronavirus uses the ACE2 of multiple animal species as receptor and evades IFITM3 restriction via TMPRSS2 activation of membrane fusion. Emerging Microbes & Infections 9. [CrossRef]
- Wang, Q., Noettger, S., Xie, Q., Pastorio, C., Seidel, A., Müller, J.A., Jung, C., Jacob, T., Sparrer, K.M.J., Zech, F., et al. (2023). Determinants of species-specific utilization of ACE2 by human and animal coronaviruses. Commun Biol 6. [CrossRef]
- Zech, F., Schniertshauer, D., Jung, C., Herrmann, A., Cordsmeier, A., Xie, Q., Nchioua, R., Prelli Bozzo, C., Volcic, M., Koepke, L., et al. (2021). Spike residue 403 affects binding of coronavirus spikes to human ACE2. Nat Commun 12. [CrossRef]
- Lim, S., Zhang, M., and Chang, T.L. (2022). ACE2-Independent Alternative Receptors for SARS-CoV-2. Viruses 14. 2535. [CrossRef]
- Sasaki, M., Toba, S., Itakura, Y., Chambaro, H.M., Kishimoto, M., Tabata, K., Intaruck, K., Uemura, K., Sanaki, T., Sato, A., et al. (2021). SARS-CoV-2 Bearing a Mutation at the S1/S2 Cleavage Site Exhibits Attenuated Virulence and Confers Protective Immunity. mBio 12. e0141521. [CrossRef]
- Whittaker, G.R. (2021). SARS-CoV-2 spike and its adaptable furin cleavage site. The Lancet Microbe 2. [CrossRef]
- Peacock, T.P., Goldhill, D.H., Zhou, J., Baillon, L., Frise, R., Swann, O.C., Kugathasan, R., Penn, R., Brown, J.C., Sanchez-David, R.Y., et al. (2021). The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nat Microbiol 6. [CrossRef]
- Örd, M., Faustova, I., and Loog, M. (2020). The sequence at Spike S1/S2 site enables cleavage by furin and phospho-regulation in SARS-CoV2 but not in SARS-CoV1 or MERS-CoV. Sci Rep 10. [CrossRef]
- Chan, Y.A., and Zhan, S.H. (2022). The Emergence of the Spike Furin Cleavage Site in SARS-CoV-2. Mol Biol Evol 39. [CrossRef]
- Coutard, B., Valle, C., de Lamballerie, X., Canard, B., Seidah, N.G., and Decroly, E. (2020). The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral Res 176. [CrossRef]
- Andersen, K.G., Rambaut, A., Lipkin, W.I., Holmes, E.C., and Garry, R.F. (2020). The proximal origin of SARS-CoV-2. Nature Medicine 26. [CrossRef]
- Zhang, T., Wu, Q., and Zhang, Z. (2020). Probable Pangolin Origin of SARS-CoV-2 Associated with the COVID-19 Outbreak. Current Biology 30. [CrossRef]
- Xiong, Q., Cao, L., Ma, C., Tortorici, M.A., Liu, C., Si, J., Liu, P., Gu, M., Walls, A.C., Wang, C., et al. (2022). Close relatives of MERS-CoV in bats use ACE2 as their functional receptors. Nature 612. [CrossRef]
- Stout, A.E., Millet, J.K., Stanhope, M.J., and Whittaker, G.R. (2021). Furin cleavage sites in the spike proteins of bat and rodent coronaviruses: Implications for virus evolution and zoonotic transfer from rodent species. One Health 13. [CrossRef]
- Lubinski, B., Fernandes, M.H.V., Frazier, L., Tang, T., Daniel, S., Diel, D.G., Jaimes, J.A., and Whittaker, G.R. (2021). Functional evaluation of the P681H mutation on the proteolytic activation of the SARS-CoV-2 variant B.1.1.7 (Alpha) spike. iScience 25. [CrossRef]
- Vu, M.N., Alvarado, R.E., Morris, D.R., Lokugamage, K.G., Zhou, Y., Morgan, A.L., Estes, L.K., McLeland, A.M., Schindewolf, C., Plante, J.A., et al. (2023). Loss-of-function mutation in Omicron variants reduces spike protein expression and attenuates SARS-CoV-2 infection. bioRxiv, 2023.04.17.536926. [CrossRef]
- Schaefer, S.L., Jung, H., and Hummer, G. (2021). Binding of SARS-CoV-2 Fusion Peptide to Host Endosome and Plasma Membrane. J Phys Chem B 125. [CrossRef]
- Yu, S., Zheng, X., Zhou, B., Li, J., Chen, M., Deng, R., Wong, G., Lavillette, D., and Meng, G. (2022). SARS-CoV-2 spike engagement of ACE2 primes S2’ site cleavage and fusion initiation. Proc Natl Acad Sci U S A 119. [CrossRef]
- Hu, B., Chan, J.F.-W., Liu, H., Liu, Y., Chai, Y., Shi, J., Shuai, H., Hou, Y., Huang, X., Yuen, T.T.-T., et al. (2022). Spike mutations contributing to the altered entry preference of SARS-CoV-2 omicron BA.1 and BA.2. Emerging Microbes & Infections 11. [CrossRef]
- Meng, B., Abdullahi, A., Ferreira, I.A.T.M., Goonawardane, N., Saito, A., Kimura, I., Yamasoba, D., Gerber, P.P., Fatihi, S., Rathore, S., et al. (2022). Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 603. [CrossRef]
- Qu, P., Evans, J.P., Kurhade, C., Zeng, C., Zheng, Y.-M., Xu, K., Shi, P.-Y., Xie, X., and Liu, S.-L. (2023). Determinants and Mechanisms of the Low Fusogenicity and High Dependence on Endosomal Entry of Omicron Subvariants. mBio 14. [CrossRef]
- Nchioua, R., Schundner, A., Kmiec, D., Prelli Bozzo, C., Zech, F., Koepke, L., Graf, A., Krebs, S., Blum, H., Frick, M., et al. (2022). SARS-CoV-2 Variants of Concern Hijack IFITM2 for Efficient Replication in Human Lung Cells. J Virol 96. [CrossRef]
- Prelli Bozzo, C., Nchioua, R., Volcic, M., Koepke, L., Krüger, J., Schütz, D., Heller, S., Stürzel, C.M., Kmiec, D., Conzelmann, C., et al. (2021). IFITM proteins promote SARS-CoV-2 infection and are targets for virus inhibition in vitro. Nat Commun 12. [CrossRef]
- Xie, Q., Bozzo, C.P., Eiben, L., Noettger, S., Kmiec, D., Nchioua, R., Niemeyer, D., Volcic, M., Lee, J.-H., Zech, F., et al. (2023). Endogenous IFITMs boost SARS-coronavirus 1 and 2 replication whereas overexpression inhibits infection by relocalizing ACE2. iScience 26. [CrossRef]
- Basile, A., Zannella, C., De Marco, M., Sanna, G., Franci, G., Galdiero, M., Manzin, A., De Laurenzi, V., Chetta, M., Rosati, A., et al. (2023). Spike-mediated viral membrane fusion is inhibited by a specific anti-IFITM2 monoclonal antibody. Antiviral Res 211. [CrossRef]
- Stewart, H., Palmulli, R., Johansen, K.H., McGovern, N., Shehata, O.M., Carnell, G.W., Jackson, H.K., Lee, J.S., Brown, J.C., Burgoyne, T., et al. (2023). Tetherin antagonism by SARS-CoV-2 ORF3a and spike protein enhances virus release. EMBO Rep. [CrossRef]
- Dangi, T., Palacio, N., Sanchez, S., Park, M., Class, J., Visvabharathy, L., Ciucci, T., Koralnik, I.J., Richner, J.M., and Penaloza-MacMaster, P. (2021). Cross-protective immunity following coronavirus vaccination and coronavirus infection. J Clin Invest. [CrossRef]
- Weissman, D., Alameh, M.G., de Silva, T., Collini, P., Hornsby, H., Brown, R., LaBranche, C.C., Edwards, R.J., Sutherland, L., Santra, S., et al. (2021). D614G Spike Mutation Increases SARS CoV-2 Susceptibility to Neutralization. Cell Host and Microbe 29. [CrossRef]
- Zhang, L., Jackson, C.B., Mou, H., Ojha, A., Peng H., Quinlan B.D., Rangarajan, E.S., Pan A., Vanderheiden A., Suthar M.S., Li W., Izard, T., Rader C., Farzan, M., and Choe, H. (2020). SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat Commun. 11. 10.1038/s41467-020-19808-4.
- Plante, J.A., Liu, Y., Liu, J., Xia, H., Johnson, B.A., Lokugamage, K.G., Zhang, X., Muruato, A.E., Zou, J., Fontes-Garfias, C.R., et al. (2021). Spike mutation D614G alters SARS-CoV-2 fitness. Nature 592. [CrossRef]
- Daniloski, Z., Jordan, T.X., Ilmain, J.K., Guo, X., Bhabha, G., TenOever, B.R., and Sanjana, N.E. (2021). The Spike D614G mutation increases SARS-CoV-2 infection of multiple human cell types. eLife 10. [CrossRef]
- Bhattacharya, M., Chatterjee, S., Sharma, A.R., Agoramoorthy, G., and Chakraborty, C. (2021). D614G mutation and SARS-CoV-2: impact on S-protein structure, function, infectivity, and immunity. Appl Microbiol Biotechnol 105. [CrossRef]
- Yurkovetskiy, L., Wang, X., Pascal, K.E., Tomkins-Tinch, C., Nyalile, T.P., Wang, Y., Baum, A., Diehl, W.E., Dauphin, A., Carbone, C., et al. (2020). Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 183. [CrossRef]
- Korber, B., Fischer, W.M., Gnanakaran, S., Yoon, H., Theiler, J., Abfalterer, W., Hengartner, N., Giorgi, E.E., Bhattacharya, T., Foley, B., et al. (2020). Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 182. [CrossRef]
- Martin, D.P., Weaver, S., Tegally, H., San, J.E., Shank, S.D., Wilkinson, E., Lucaci, A.G., Giandhari, J., Naidoo, S., Pillay, Y., et al. (2021). The emergence and ongoing convergent evolution of the SARS-CoV-2 N501Y lineages. Cell 184. [CrossRef]
- Tian, F., Tong, B., Sun, L., Shi, S., Zheng, B., Wang, Z., Dong, X., and Zheng, P. (2021). N501Y mutation of spike protein in SARS-CoV-2 strengthens its binding to receptor ACE2. eLife 10. [CrossRef]
- Wright, E.S., Lakdawala, S.S., and Cooper, V.S. (2020). SARS-CoV-2 genome evolution exposes early human adaptations. bioRxiv. [CrossRef]
- Posani, E., Dilucca, M., Forcelloni, S., Pavlopoulou, A., Georgakilas, A.G., and Giansanti, A. (2022). Temporal evolution and adaptation of SARS-CoV-2 codon usage. Frontiers in Bioscience-Landmark 27. [CrossRef]
- Choe, H., and Farzan, M. (2021). How SARS-CoV-2 first adapted in humans: An early spike protein mutation promotes transmission and will shape the next vaccines. Science 372. [CrossRef]
- Tosta, E. (2021). The adaptation of SARS-CoV-2 to humans. Memórias do Instituto Oswaldo Cruz 116. [CrossRef]
- Cosar, B., Karagulleoglu, Z.Y., Unal, S., Ince, A.T., Uncuoglu, D.B., Tuncer, G., Kilinc, B.R., Ozkan, Y.E., Ozkoc, H.C., Demir, I.N., et al. (2022). SARS-CoV-2 Mutations and their Viral Variants. Cytokine & growth factor reviews 63. [CrossRef]
- Tao, K., Tzou, P.L., Nouhin, J., Gupta, R.K., de Oliveira, T., Kosakovsky Pond, S.L., Fera, D., and Shafer, R.W. (2021). The biological and clinical significance of emerging SARS-CoV-2 variants. Nat Rev Genet 22. [CrossRef]
- Jung, C., Kmiec, D., Koepke, L., Zech, F., Jacob, T., Sparrer, K.M.J., and Kirchhoff, F. (2022). Omicron: what makes the latest SARS-CoV-2 variant of concern so concerning? J Virol. [CrossRef]
- Nchioua, R., Diofano, F., Noettger, S., von Maltitz, P., Stenger, S., Zech, F., Münch, J., Sparrer, K.M.J., Just, S., and Kirchhoff, F. (2022). Strong attenuation of SARS-CoV-2 Omicron BA.1 and increased replication of the BA.5 subvariant in human cardiomyocytes. Sig Transduct Target Ther 7. [CrossRef]
- van Doremalen, N., Singh, M., Saturday, T.A., Yinda, C.K., Perez-Perez, L., Bohler, W.F., Weishampel, Z.A., Lewis, M., Schulz, J.E., Williamson, B.N., et al. (2022). SARS-CoV-2 Omicron BA.1 and BA.2 are attenuated in rhesus macaques as compared to Delta. bioRxiv. [CrossRef]
- Shuai, H., Chan, J.F.-W., Hu, B., Chai, Y., Yoon, C., Liu, H., Liu, Y., Shi, J., Zhu, T., Hu, J.-C., et al. (2023). The viral fitness and intrinsic pathogenicity of dominant SARS-CoV-2 Omicron sublineages BA.1, BA.2, and BA.5. eBioMedicine 95. [CrossRef]
- Pastorio, C., Zech, F., Noettger, S., Jung, C., Jacob, T., Sanderson, T., Sparrer, K.M.J., and Kirchhoff, F. (2022). Determinants of Spike infectivity, processing, and neutralization in SARS-CoV-2 Omicron subvariants BA.1 and BA.2. Cell Host & Microbe 30. [CrossRef]
- Hachmann, N.P., Miller, J., Collier, A.Y., Ventura, J.D., Yu, J., Rowe, M., Bondzie, E.A., Powers, O., Surve, N., Hall, K., et al. (2022). Neutralization Escape by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, and BA.5. New England Journal of Medicine 387. [CrossRef]
- Wang, Q., Guo, Y., Iketani, S., Nair, M.S., Li, Z., Mohri, H., Wang, M., Yu, J., Bowen, A.D., Chang, J.Y., et al. (2022). Antibody evasion by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4 and BA.5. Nature 608. [CrossRef]
- Wang, Q., Guo, Y., Iketani, S., Nair, M.S., Li, Z., Mohri, H., Wang, M., Yu, J., Bowen, A.D., Chang, J.Y., et al. (2022). Antibody evasion by SARS-CoV-2 Omicron subvariants BA.2.12.1, BA.4 and BA.5. Nature 608. [CrossRef]
- Park, Y.-J., Pinto, D., Walls, A.C., Liu, Z., De Marco, A., Benigni, F., Zatta, F., Silacci-Fregni, C., Bassi, J., Sprouse, K.R., et al. (2022). Imprinted antibody responses against SARS-CoV-2 Omicron sublineages. Science 378. [CrossRef]
- Cao, Y., Jian, F., Wang, J., Yu, Y., Song, W., Yisimayi, A., Wang, J., An, R., Chen, X., Zhang, N., et al. (2023). Imprinted SARS-CoV-2 humoral immunity induces convergent Omicron RBD evolution. Nature 614. [CrossRef]
- Ito, J., Suzuki, R., Uriu, K., Itakura, Y., Zahradnik, J., Kimura, K.T., Deguchi, S., Wang, L., Lytras, S., Tamura, T., et al. (2023). Convergent evolution of SARS-CoV-2 Omicron subvariants leading to the emergence of BQ.1.1 variant. Nature Communications 2023 14. [CrossRef]
- Ao, D., He, X., Hong, W., and Wei, X. (2023). The rapid rise of SARS-CoV-2 Omicron subvariants with immune evasion properties: XBB.1.5 and BQ.1.1 subvariants. MedComm 4. [CrossRef]
- Hoffmann, M., Arora, P., Nehlmeier, I., Kempf, A., Cossmann, A., Schulz, S.R., Morillas Ramos, G., Manthey, L.A., Jäck, H.-M., Behrens, G.M.N., et al. (2023). Profound neutralization evasion and augmented host cell entry are hallmarks of the fast-spreading SARS-CoV-2 lineage XBB.1.5. Cell Mol Immunol 20. [CrossRef]
- Wang, Q., Iketani, S., Li, Z., Liu, L., Guo, Y., Huang, Y., Bowen, A.D., Liu, M., Wang, M., Yu, J., et al. (2023). Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell 186. [CrossRef]
- Yue, C., Song, W., Wang, L., Jian, F., Chen, X., Gao, F., Shen, Z., Wang, Y., Wang, X., and Cao, Y. (2023). ACE2 binding and antibody evasion in enhanced transmissibility of XBB.1.5. The Lancet Infectious Diseases 23. [CrossRef]
- Yang, S., Yu, Y., Xu, Y., Jian, F., Song, W., Yisimayi, A., Wang, P., Wang, J., Liu, J., Yu, L., et al. (2023). Fast evolution of SARS-CoV-2 BA.2·86 to JN.1 under heavy immune pressure. The Lancet Infectious Diseases. [CrossRef]
- Belik, M., Liedes, O., Vara, S., Haveri, A., Pöysti, S., Kolehmainen, P., Maljanen, S., Huttunen, M., Reinholm, A., Lundberg, R., et al. (2023). Persistent T cell-mediated immune responses against Omicron variants after the third COVID-19 mRNA vaccine dose. Front Immunol 14. [CrossRef]
- Arieta, C.M., Xie, Y.J., Rothenberg, D.A., Diao, H., Harjanto, D., Meda, S., Marquart, K., Koenitzer, B., Sciuto, T.E., Lobo, A., et al. (2023). The T-cell-directed vaccine BNT162b4 encoding conserved non-spike antigens protects animals from severe SARS-CoV-2 infection. Cell 186. [CrossRef]
- Cohen, L.E., Spiro, D.J., and Viboud, C. (2022). Projecting the SARS-CoV-2 transition from pandemicity to endemicity: Epidemiological and immunological considerations. PLOS Pathogens 18. [CrossRef]
- Biancolella, M., Colona, V.L., Mehrian-Shai, R., Watt, J.L., Luzzatto, L., Novelli, G., and Reichardt, J.K.V. (2022). COVID-19 2022 update: transition of the pandemic to the endemic phase. Human Genomics 16. [CrossRef]
- Saunders, K.O., Lee, E., Parks, R., Martinez, D.R., Li, D., Chen, H., Edwards, R.J., Gobeil, S., Barr, M., Mansouri, K., et al. (2021). Neutralizing antibody vaccine for pandemic and pre-emergent coronaviruses. Nature 594. [CrossRef]
- Planas, D., Saunders, N., Maes, P., Guivel-Benhassine, F., Planchais, C., Buchrieser, J., Bolland, W.-H., Porrot, F., Staropoli, I., Lemoine, F., et al. (2021). Considerable escape of SARS-CoV-2 variant Omicron to antibody neutralization. Nature 602. [CrossRef]
- Imai, M., Ito, M., Kiso, M., Yamayoshi, S., Uraki, R., Fukushi, S., Watanabe, S., Suzuki, T., Maeda, K., Sakai-Tagawa, Y., Iwatsuki-Horimoto, K., Halfmann, P.J., Kawaoka, Y. (2022). Efficacy of Antiviral Agents against Omicron Subvariants BQ.1.1 and XBB. N Engl J Med. 388. [CrossRef]
- Wang, P., Nair, M.S., Liu, L., Iketani, S., Luo, Y., Guo, Y., Wang, M., Yu, J., Zhang, B., Kwong, P.D., et al. (2021). Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature 593. [CrossRef]
- Cao, Y., Wang, J., Jian, F., Xiao, T., Song, W., Yisimayi, A., Huang, W., Li, Q., Wang, P., An, R., et al. (2022). Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 602. [CrossRef]
- Liu, L., Iketani, S., Guo, Y., Chan, J.F.-W., Wang, M., Liu, L., Luo, Y., Chu, H., Huang, Y., Nair, M.S., et al. (2022). Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature 602. [CrossRef]
- Zhou, P., Song, G., Liu, H., Yuan, M., He, W., Beutler, N., Zhu, X., Tse, L.V., Martinez, D.R., Schäfer, A., et al. (2023). Broadly neutralizing anti-S2 antibodies protect against all three human betacoronaviruses that cause deadly disease. Immunity 56. [CrossRef]
- Chen, Y., Zhao, X., Zhou, H., Zhu, H., Jiang, S., and Wang, P. (2023). Broadly neutralizing antibodies to SARS-CoV-2 and other human coronaviruses. Nat Rev Immunol 23. [CrossRef]
- Karoyan, P., Vieillard, V., Gómez-Morales, L., Odile, E., Guihot, A., Luyt, C.-E., Denis, A., Grondin, P., and Lequin, O. (2021). Human ACE2 peptide-mimics block SARS-CoV-2 pulmonary cells infection. Commun Biol 4. [CrossRef]
- Larue, R.C., Xing, E., Kenney, A.D., Zhang, Y., Tuazon, J.A., Li, J., Yount, J.S., Li, P.-K., and Sharma, A. (2021). Rationally Designed ACE2-Derived Peptides Inhibit SARS-CoV-2. Bioconjug Chem 32. [CrossRef]
- Adhikary, P., Kandel, S., Mamani, U.-F., Mustafa, B., Hao, S., Qiu, J., Fetse, J., Liu, Y., Ibrahim, N.M., Li, Y., et al. (2021). Discovery of Small Anti-ACE2 Peptides to Inhibit SARS-CoV-2 Infectivity. Adv Ther (Weinh) 4. [CrossRef]
- Xia, S., Lan, Q., Zhu, Y., Wang, C., Xu, W., Li, Y., Wang, L., Jiao, F., Zhou, J., Hua, C., et al. (2021). Structural and functional basis for pan-CoV fusion inhibitors against SARS-CoV-2 and its variants with preclinical evaluation. Sig Transduct Target Ther 6. [CrossRef]
- Yu, D., Zhu, Y., Yan, H., Wu, T., Chong, H., and He, Y. (2021). Pan-coronavirus fusion inhibitors possess potent inhibitory activity against HIV-1, HIV-2, and simian immunodeficiency virus. Emerg Microbes Infect 10. [CrossRef]
- Xia, S., Wang, L., Jiao, F., Yu, X., Xu, W., Huang, Z., Li, X., Wang, Q., Zhu, Y., Man, Q., et al. (2023). SARS-CoV-2 Omicron subvariants exhibit distinct fusogenicity, but similar sensitivity, to pan-CoV fusion inhibitors. Emerg Microbes Infect 12. [CrossRef]
- Cheng, Y.-W., Chao, T.-L., Li, C.-L., Chiu, M.-F., Kao, H.-C., Wang, S.-H., Pang, Y.-H., Lin, C.-H., Tsai, Y.-M., Lee, W.-H., et al. (2020). Furin Inhibitors Block SARS-CoV-2 Spike Protein Cleavage to Suppress Virus Production and Cytopathic Effects. Cell Rep 33. [CrossRef]
- Müller, P., Maus, H., Hammerschmidt, S.J., Knaff, P., Mailänder, V., Schirmeister, T., and Kersten, C. (2021). Interfering with Host Proteases in SARS-CoV-2 Entry as a Promising Therapeutic Strategy. Curr Med Chem. [CrossRef]
- Wettstein, L., Immenschuh, P., Weil, T., Conzelmann, C., Almeida-Hernández, Y., Hoffmann, M., Kempf, A., Nehlmeier, I., Lotke, R., Petersen, M., et al. (2023). Native and activated antithrombin inhibits TMPRSS2 activity and SARS-CoV-2 infection. J Med Virol 95. [CrossRef]
- Polack, F.P., Thomas, S.J., Kitchin, N., Absalon, J., Gurtman, A., Lockhart, S., Perez, J.L., Pérez Marc, G., Moreira, E.D., Zerbini, C., et al. (2020). Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med 383. [CrossRef]
- Folegatti, P.M., Ewer, K.J., Aley, P.K., Angus, B., Becker, S., Belij-Rammerstorfer, S., Bellamy, D., Bibi, S., Bittaye, M., Clutterbuck, E.A., et al. (2020). Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: a preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 396. [CrossRef]
- Xia, S., Zhang, Y., Wang, Y., Wang, H., Yang, Y., Gao, G.F., Tan, W., Wu, G., Xu, M., Lou, Z., et al. (2021). Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: a randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect Dis 21. [CrossRef]
- Voysey, M., Costa Clemens, S.A., Madhi, S.A., Weckx, L.Y., Folegatti, P.M., Aley, P.K., Angus, B., Baillie, V.L., Barnabas, S.L., Bhorat, Q.E., et al. (2021). Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: a pooled analysis of four randomised trials. Lancet 397. [CrossRef]
- Lopez Bernal, J., Andrews, N., Gower, C., Gallagher, E., Simmons, R., Thelwall, S., Stowe, J., Tessier, E., Groves, N., Dabrera, G., et al. (2021). Effectiveness of Covid-19 Vaccines against the B.1.617.2 (Delta) Variant. N Engl J Med 385. [CrossRef]
- Tseng, H.F., Ackerson, B.K., Sy, L.S., Tubert, J.E., Luo, Y., Qiu, S., Lee, G.S., Bruxvoort, K.J., Ku, J.H., Florea, A., et al. (2023). mRNA-1273 bivalent (original and Omicron) COVID-19 vaccine effectiveness against COVID-19 outcomes in the United States. Nat Commun 14. [CrossRef]
- Martinez, D.R., fer, A.S., Gavitt, T.D., Mallory, M.L., Lee, E., Catanzaro, N.J., Chen, H., Gully, K., Scobey, T., Brown, A., et al. (2023) Vaccine-mediated protection against merbecovirus and sarbecovirus challenge in mice. bioRxiv. [CrossRef]
- Martinez, D.R., Schäfer, A., Gavitt, T.D., Mallory, M.L., Lee, E., Catanzaro, N.J., Chen, H., Gully, K., Scobey, T., Korategere, P., et al. (2023). Vaccine-mediated protection against Merbecovirus and Sarbecovirus challenge in mice. Cell reports 42. [CrossRef]
- Evans, T.S., Tan, C.W., Aung, O., Phyu, S., Lin, H., Coffey, L.L., Toe, A.T., Aung, P., Aung, T.H., Aung, N.T., et al. (2023). Exposure to diverse sarbecoviruses indicates frequent zoonotic spillover in human communities interacting with wildlife. International Journal of Infectious Diseases 131. [CrossRef]
- Cantoni, D., Mayora-Neto, M., Thakur, N., Elrefaey, A.M.E., Newman, J., Vishwanath, S., Nadesalingam, A., Chan, A., Smith, P., Castillo-Olivares, J., et al. (2022). Pseudotyped Bat Coronavirus RaTG13 is efficiently neutralised by convalescent sera from SARS-CoV-2 infected patients. Commun Biol 5. [CrossRef]
- Lawrenz, J., Xie, Q., Zech, F., Weil, T., Seidel, A., Krnavek, D., van der Hoek, L., Münch, J., Müller, J.A., and Kirchhoff, F. (2022). Severe Acute Respiratory Syndrome Coronavirus 2 Vaccination Boosts Neutralizing Activity Against Seasonal Human Coronaviruses. Clinical Infectious Diseases 75. [CrossRef]
Figure 1.
Phylogenetic relationship between human coronavirus (left), animal Sarbecovirus (middle) and human SARS-CoV-2 VOC Spike (right) amino acid sequences from the indicated viral strains or species. Host species are indicated by silhouettes: bat (blue), pangolin (turquoise), civet (orange) and human (red). Sequence identifiers are provided in supplementary Table S1.
Figure 1.
Phylogenetic relationship between human coronavirus (left), animal Sarbecovirus (middle) and human SARS-CoV-2 VOC Spike (right) amino acid sequences from the indicated viral strains or species. Host species are indicated by silhouettes: bat (blue), pangolin (turquoise), civet (orange) and human (red). Sequence identifiers are provided in supplementary Table S1.
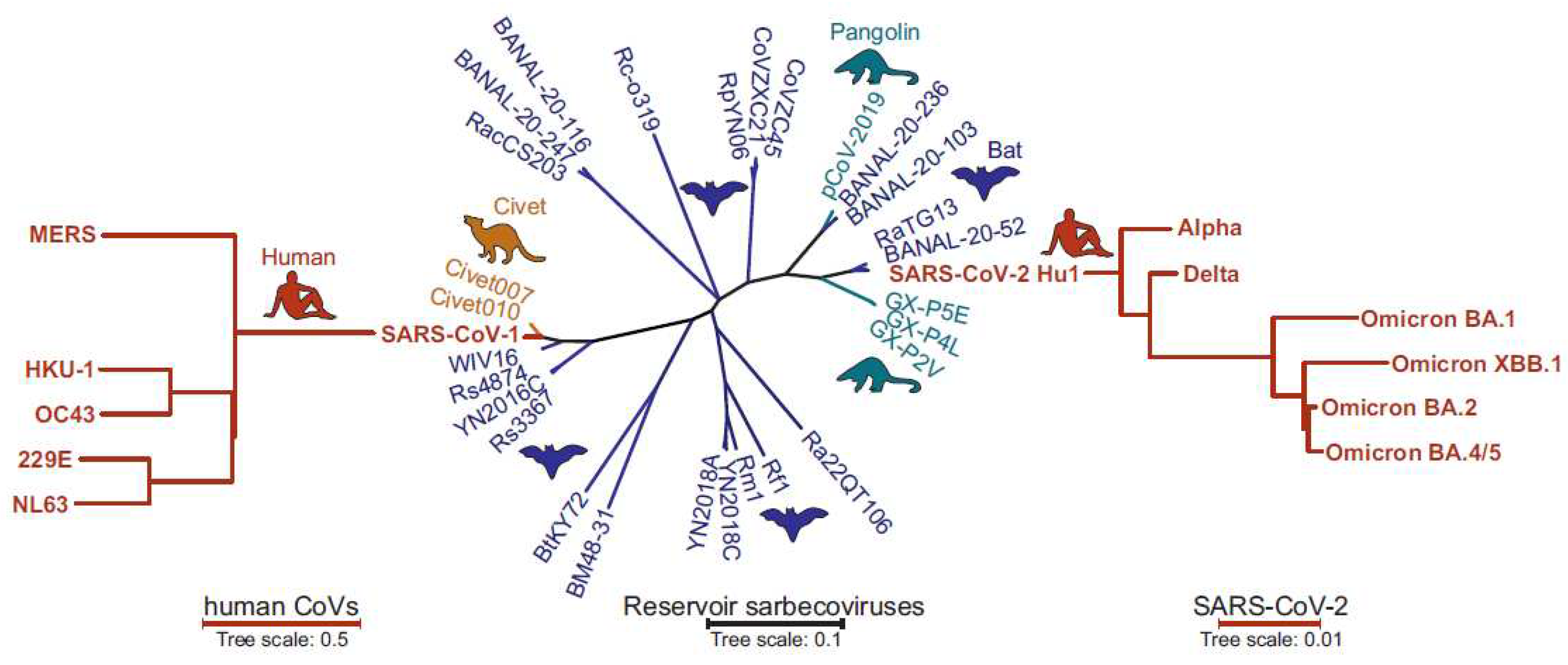
Figure 2.
Conservation and mutation mapping of SARS-CoV-2 VOCs and Sarbecovirus Spike proteins. (A) Conservation of 29 Sarbecovirus spike proteins with regions indicated as NTD (N-terminal domain, orange), RBD (receptor-binding domain, orange), RBM (receptor-binding motif, dark orange), S1/S2 and S2' (protease cleavage sites, black arrows), FP (fusion peptide, blue) and HR1/2 (heptad repeat 1/2, dark blue). Positions of mutations of selected bat Sarbecoviruses (gray) and SARS-CoV-2 VOCs (red) are indicated as circles. (B) Overview of the SARS-CoV-2 Spike structure (PDB: 7KNB) and localization of amino acid changes in the indicated SARS-CoV-2 VOCs. Color coding according to domains as indicated.
Figure 2.
Conservation and mutation mapping of SARS-CoV-2 VOCs and Sarbecovirus Spike proteins. (A) Conservation of 29 Sarbecovirus spike proteins with regions indicated as NTD (N-terminal domain, orange), RBD (receptor-binding domain, orange), RBM (receptor-binding motif, dark orange), S1/S2 and S2' (protease cleavage sites, black arrows), FP (fusion peptide, blue) and HR1/2 (heptad repeat 1/2, dark blue). Positions of mutations of selected bat Sarbecoviruses (gray) and SARS-CoV-2 VOCs (red) are indicated as circles. (B) Overview of the SARS-CoV-2 Spike structure (PDB: 7KNB) and localization of amino acid changes in the indicated SARS-CoV-2 VOCs. Color coding according to domains as indicated.
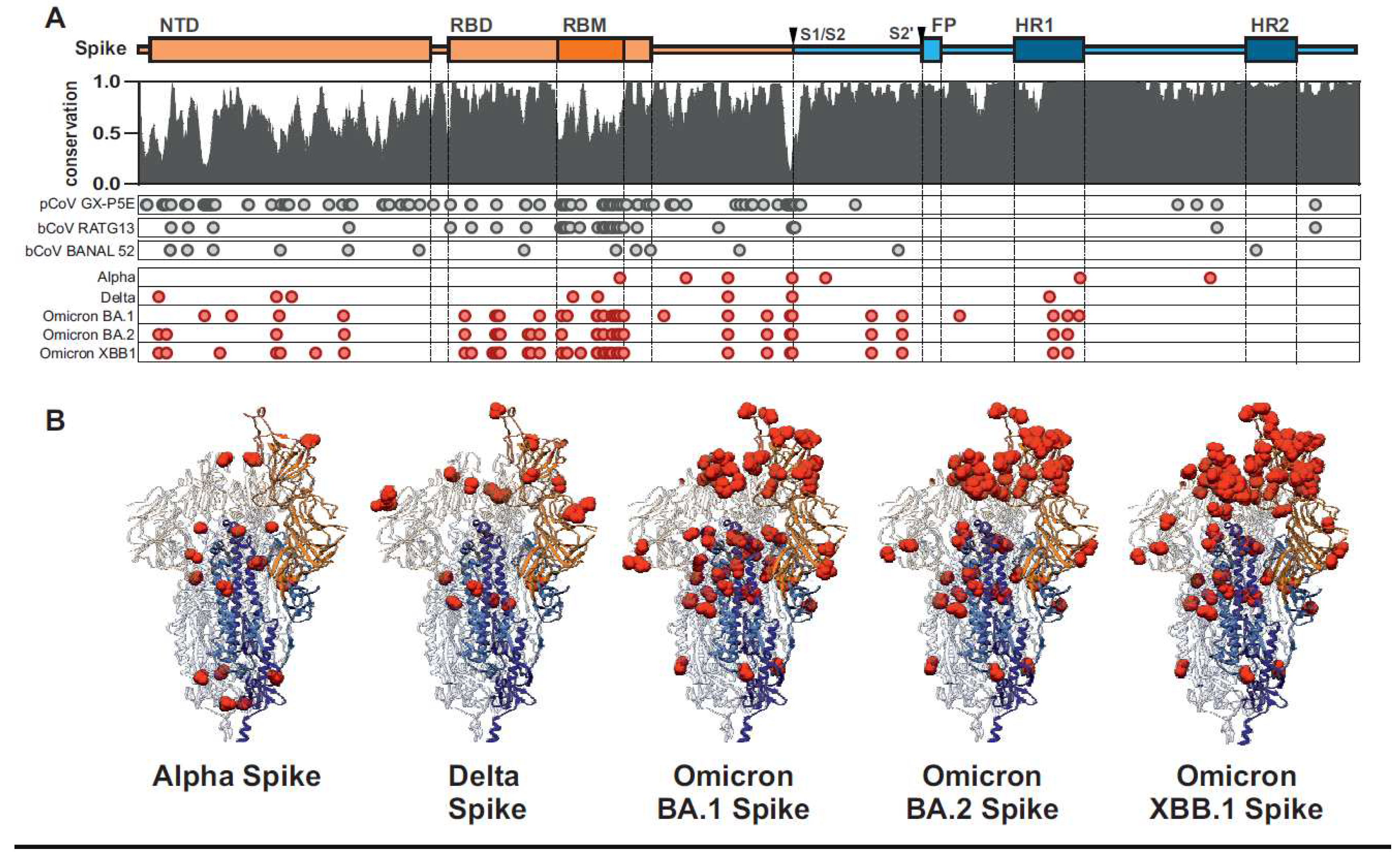
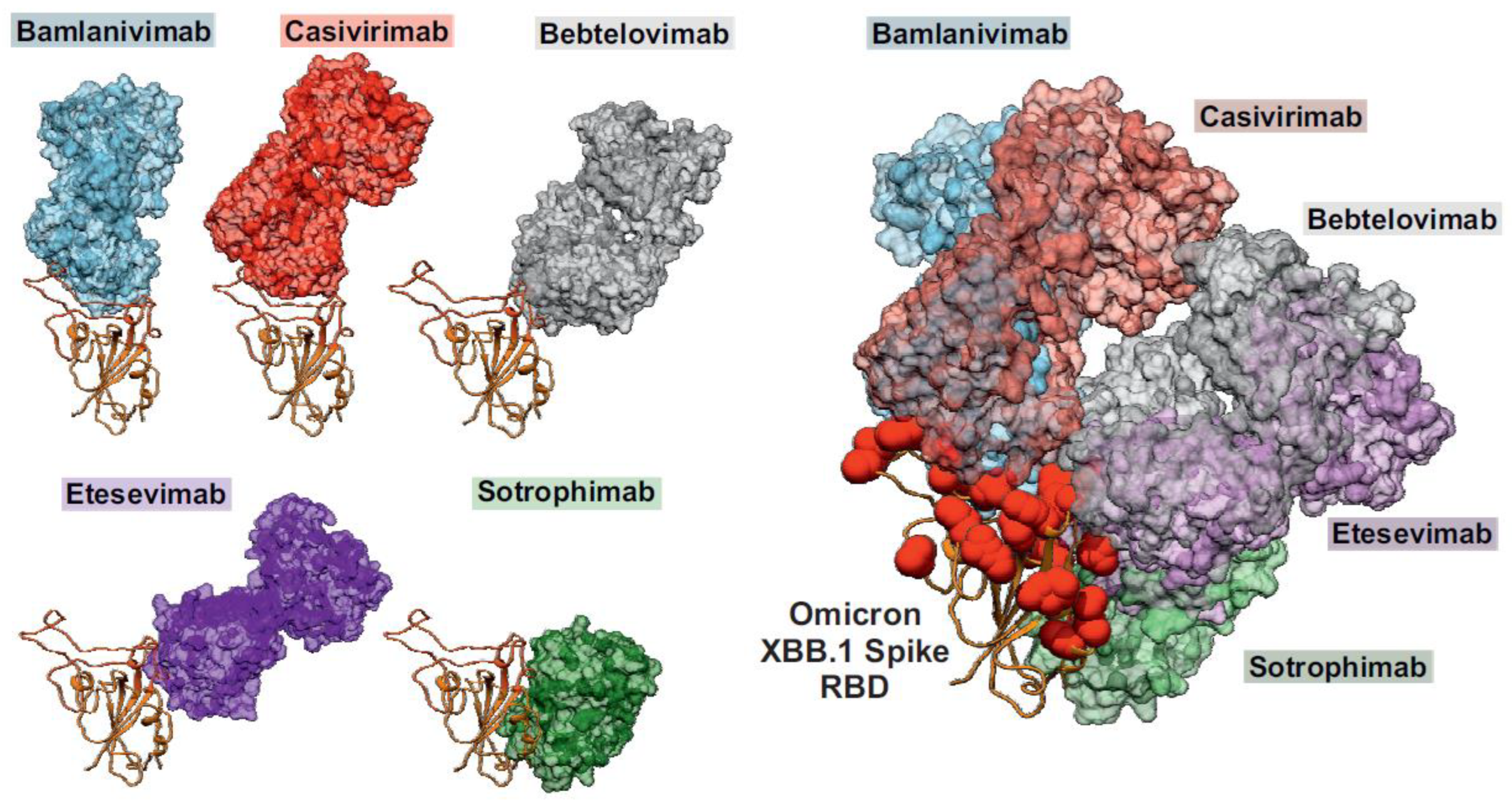
Figure 3.
Illustration of binding sites of nAbs to the Spike RBD. The RBD of the SARS-CoV-2 Spike protein (PDB: 7KNB, yellow ribbon) and interacting therapeutic monoclonal antibodies are indicated: Bamlanivimab (blue, PDB: 7KMG), Casirivimab (red, PDB: 7M42), Bebtelovimab (gray, PDB: 7MMO), Etesevimab (purple, PDB: 7F7E) and Sotrovimab (green, PDB: 7TLY). The positions of RBD-specific mutations in SARS-CoV-2 Omicron XBB.1 are highlighted by red spheres.
Figure 3.
Illustration of binding sites of nAbs to the Spike RBD. The RBD of the SARS-CoV-2 Spike protein (PDB: 7KNB, yellow ribbon) and interacting therapeutic monoclonal antibodies are indicated: Bamlanivimab (blue, PDB: 7KMG), Casirivimab (red, PDB: 7M42), Bebtelovimab (gray, PDB: 7MMO), Etesevimab (purple, PDB: 7F7E) and Sotrovimab (green, PDB: 7TLY). The positions of RBD-specific mutations in SARS-CoV-2 Omicron XBB.1 are highlighted by red spheres.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



