Submitted:
02 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
Seeds play a vital role in the perpetuation of plant species, both in natural environments and agriculture. However, they often face challenges from biotic stresses, such as seedborne pathogenic fungi. The transgenerational transmission of these seedborne fungi, along with their dissemination during seed commercialization, can contribute to the emergence of global epidemic diseases, resulting in substantial economic losses. One such pathogenic fungus is Alternaria brassicicola, responsible for causing black spot disease in Brassicaceae. This necrotrophic seedborne fungus negatively impacts plant development, consequently affecting crop production and seed quality. To gain a deeper understanding of the interaction between seeds and necrotrophic fungi, the Arabidopsis thaliana/Alternaria brassicicola model pathosystem has been extensively studied. Recent transcriptomic and genetic studies revealed a distinctive immune response of seeds against this pathogen, divergent from the immune response described in the plant vegetative phase. In germinating seeds and seedlings, the necrotrophic fungus seems to induce susceptible response [1]. It was observed at the RNA level that to the detriment of jasmonic acid pathway, Alternaria induced the salicylic acid and glucosinolate (GSL) pathways. Genetic evidence identified that seedling necrosis, potentially beneficial to the growth of the necrotrophic agent, is hypersensitive response mediated by GSL pathway. Transmission of necrotrophic fungal from seed to seedling unveils novel regulations of the immune response, that would illustrate a trade-off between seedling establishment and fungal infection. This knowledge is promising to sustainably ensure the sanitary quality of crops.
Keywords:
seedborne pathogens
; Alternaria brassicicola
; seed defense
; transcriptomics
; Arabidopsis.
1. Introduction
Seeds are the fundamental reproductive structure that play a crucial role in the propagation and survival of flowering plants and gymnosperms, including many economically important species [2,3,4]. Following seed development, the mature seed is metabolically equipped to optimize germination and seedling establishment after its dissemination in a stressful environment [2] . Overall seed tissues that generally comprises the embryo (sporophyte), and reserve tissues (cotyledons, endosperm or perisperm) surrounded by covering tissue, known as the testa contribute to this transition [2,3]. Seed germination is controlled by complex regulation integrating environmental signals such as light, temperature, nitrate sources, and endogenous hormonal signals such as the well described abscisic acid and gibberellin pathways [2,4,5]. Germination begins with water imbibition that induces a burst of respiratory and energetic metabolism, an activation of translation from stored RNA and RNA neo-syntheses. Environmental factors will interfere with hormonal metabolism and seed responsiveness controlling embryo growth potential and radicle protrusion through weakened endosperm and testa rupture. Hydrolysis and mobilization of storage nutriment are concomitant with this step of germination completion to support the transition to an autotrophic development phase signaling seedling establishment [2,6].
In addition to its physical and chemical attributes, the seed also hosts a microbial diversity, called microbiota, which can influence its ecological and biological functions. This seed-associated microbiota is acquired during various stages of the seed’s life cycle [7,8]. The community of seed-borne microorganisms, including bacteria, fungi, and others, that can colonize the surface of the seed, are referred to as the epiphytic microbiota. While microorganisms colonizing internal tissues such as the embryo and endosperm are known as the endophytic microbiota. The microbiota can be transmitted from the mother plant to the seedling. This phenomenon is known as vertical transmission and provides a way for the microbiome to spread through seed germination between plants [7,8]. The beneficial microbiota contributes significantly to seed nutrient uptake, disease resistance, and adaptation to environmental changes [8]. Moreover, some seed-borne microbial agents are pathogenic and can afflict seed viability and germination vigor. Their transmission to seedlings is a major cause of disease outbreaks and crop yield loss [9,10]. This is particularly critical in the case of endophytic localization of pathogens because seed treatments are not effective. Moreover, the increasingly restrictive ban on the use of pesticides makes it necessary to seek alternative solutions to control the sanitary quality of the seed [9,10,11,12].
A wide variety of plant microorganisms, including bacteria, nematodes, viruses, and fungi are described as plant pathogenic agents [13,14] (Figure 1a). The phytopathogenic fungi strongly affects agricultural production [12,15]. According to their nutritional strategies and host interactions, fungi can be classified as biotrophs, necrotrophs, and hemibiotrophs [16]. Biotrophs rely on living host cells for nutrition, necrotrophs use hydrolytic enzymes and toxins to induce host cell death and subsequently feed on dead tissues, while hemibiotrophs alternate between biotrophic and necrotrophic phases [16,17,18]. Plants have developed complex defense responses to protect against these pathogens. Defense mechanisms differ according to pathogen lifestyles [13,14]. They involve different physical barriers such as cell wall appositions, hormonal signaling and antimicrobial metabolites such as phytoalexins or reactive oxygen species (ROS) and proteins [13,14,19,20] (Figure 1b).
Recent studies using the pathosystem Arabidopsis thaliana/Alternaria brassicicola [21] have described non-canonical defense responses in the germinating seeds 1 compared to the established model describing defense mechanism against a necrotrophic agent [20,22], most of which are based on the physiological model of rosette leaf inoculation [23]. This review addresses the specific features of germinating seeds in terms of their responses to necrotrophic attacks [1]. This new knowledge will be placed in the context of general concepts on plant resistance mechanisms to biotic aggression and the particularities of necrotrophic interaction that is documented by the Arabidopsis thaliana/Alternaria brassicicola pathosystem. These studies, usually carried out on plant phylosphere tissues during growth, will be compared with existing knowledge regarding seed defense against seedborne fungal pathogens [21,24,25] and with the recent molecular mechanisms working during the germination stage, with a view to developing new crop protection strategies
2. Plant immune response
2.1. A general model for plant pathogen interaction
To safeguard themselves against phytopathogens attack, plants have developed a sophisticated immune system. This can be classified as either innate immunity or induced immunity [13,14,26]. In host plants, the innate immune response is the first line of defense. Upon contact with external pathogens, the plant utilizes specialized Pathogen Recognition Receptors (PRRs) to identify specific molecules from the infectious agent, known as Pathogen/Microbe Associated Molecular Patterns (P/MAMPs), as well as damage-associated molecular patterns (DAMPs). This process initiates a swift response designed to impede the pathogen's proliferation, referred to as P/MAMP-triggered Immunity (PTI) [13,14]. However, if the pathogen manages to evade PTI through the deployment of virulent effectors molecules, an induced immune response is activated. This response involves the recognition of pathogen’s effector molecules by plant resistance proteins, such as nucleotide-binding site leucine-rich repeat (NBS-LRR) proteins. Subsequently the Effector-Triggered Immunity (ETI) mechanism is activated to effectively detect and target the pathogen [13,18,19].
Once plant immunity is triggered, physical barriers are reinforced [13,20,22] such as callose deposition or cuticular wax accumulation and a complex network orchestrated by the phytohormones salicylic acid, jasmonic acid, and ethylene is coming into play [16,19,27]. These signaling pathways lead to an oxidative burst [16,19], to the production of antimicrobial peptides (AMPs) [28] and the biosynthesis of secondary metabolites such as indoles derived metabolites, including phytoalexins [19,29] (Figure 1b). The salicylic acid (SA) pathway has been associated with plant response to biotrophic microorganisms [16,30] (Figure 1b). The accumulation of SA induces ROS production, molecules that usually trigger the hypersensitive response (HR), a type of programmed cell death targeting infected cells, thus limiting the spread of pathogens [19,30,31]. At this stage when a plant successfully overcomes an infection, it could develop systemic acquired resistance (SAR) that makes the plant ready to induce resistance mechanism in case of pathogenic attack. This phenomenon works like a memory, providing heightened resistance to subsequent attacks and ensuring extended and more robust protection against phytopathogens [32,33].
Other hormones such as ethylene, jasmonic acid, and secondary metabolites such as camalexin, brassinin, and glucosinolates (GSL) are described to be produced in response to infection by necrotrophic and hemibiotrophic pathogens [18,27,29,34] (Figure 1b).
To gain a comprehensive understanding of plant defense mechanisms against microbial pathogens, the research emphasis has predominantly centered on plant susceptibility [13,14]. However, the intricate defense mechanisms implicated on a non-host plant remain unknown until now [26,35]. The crucial distinction in these mechanisms lies in the specificity of resistance, where host resistance is specifically tailored to particular pathogens, while non-host resistances (NHR) provide a more generalized defense. Although they share common mechanisms such as the activation of signaling pathways and the production of antimicrobial compounds, host resistance is highly adaptive, providing specific immune responses, while NHR act as broader defenses against various threats [35,36,37]. NHR comes down to a capability to recognize diverse microbial agents as non-hosts, granting plants with broad and persistent resistance against a wide range of pathogens [26,35]. This phenomenon is believed to originate from a coevolutionary dynamic between plants and pathogens [14]. Plant immune resistance is influenced by a combination of abiotic factors, pathogen characteristics, host physiology, and plant tissues under attack. Certain studies proposed this event arises either due to the inability of pathogens to suppress the PTI [14], or because one of the plant's NB-LRR receptors recognizes the effector molecules by the pathogen, thus triggering ETI [14,26,36,37]. However, detailed NHR mechanisms are still not fully understood. Consequently, it is imperative to conduct further research to elucidate the molecular foundations on plant resistance and achieve a more comprehensive grasp of plant defense mechanisms [35].
2.2. Susceptible response and plant tolerance
Furthermore, the endophytic condition assumed by specific pathogenic agents in non-host plants engenders considerable scientific interest, given its potential association with the elicitation of susceptible responses (SR) [26]. This scenario is marked by the host's constrained capacity to counteract infection, fostering an environment conducive to pathogen proliferation and establishment inside the plant.
A pathogen strategy encompasses the manipulation of plant defenses and metabolic processes to its advantage. The pathogen must employ factors such as effectors, phytohormones and phytotoxins, to activate the so-called SR genes within the plant. This enables the pathogen to control both the plant's defense mechanisms (physical and physiological) and metabolic processes such the reallocation of energy-rich compounds and the regulation of metal equilibrium, for its own benefit. As example for phytopathogenic fungi that have evolved several strategies to better face plant immunity, their ability to recognize plant-derived factors like chemical signals in the cuticle, leaf topology, and trichomes to facilitate their colonization of the host [20]. Moreover, some seed borne necrotrophic fungi such as Botrytis cinerea (B. cinerea) possess sophisticated systems to evade host recognition, hijack or disrupt plant defense signaling, and disable defense mechanisms by secreting proteinaceous effectors, and sRNA effectors for promoting their own benefit (Table 1) [38,39].
Identifying genes that facilitate pathogen proliferation contributing to disease development is crucial for implementing genetic strategies focused on suppressing symptom-associated genes. Leading to plants better adapted to infections. Tolerant plants would be able to maintain their productivity levels, remain symptom-free, and potentially even benefit from the presence of endogenous pathogens microorganisms [40]. Nevertheless, it is important to acknowledge that immunity defense mechanisms remain effective, and their efficacy may vary depending on the plant and pathogen species, as well as other environmental factors.
3. Black spot disease in Brassicaceae
The Brassicaceae family (Cruciferae), comprise approximately 4,140 species, including the model species Arabidopsis thaliana (Arabidopsis) and economically important brassica vegetables such as Brassica oleracea (broccoli, cauliflower, kale, brussels sprouts, and cabbage), Brassica rapa (vegetables, oilseeds, and forage), Brassica juncea (vegetables and mustard seeds), and Brassica napus (oilseeds) [46,47]. Brassicaceae species are susceptible to the seed-borne necrotrophic fungi Alternaria brassicicola (Berk.) Sacc., (referred to as Alternaria hereinafter) the causal agent of black spot disease. This seedborne pathogen belongs to the phylum Ascomycota, class Dothideomycetes, order Pleosporales, and family Pleosporaceae [48,49]. Alternaria is a common genus that occurs frequently as a saprotroph of decaying organic matter, but few species are plant pathogens. Diseases caused by Alternaria species are widespread, and can affect other crops such as tomatoes, apples, carrots, and potatoes [50,51]. The economic impact on agricultural production is significant, as it affects the plant at all stages of development, from seed germination to post-harvest [21,49,52].
The infection process begins with fungal penetration into the plant through wounds or stomata [20]. Once inside the plant, the fungus secretes secondary metabolites, toxins [53,54], and a series of hydrolytic enzymes such as proteases, lipases, and Cell Wall-Degrading Enzymes (CWDEs) leading to the death of plant tissues and facilitating the entry of the fungus into plant cells [48,50,55]. In leaf tissue, disease symptoms include leaf spots, yellow chlorotic halos, necrotic lesions, and leaf size reduction, resulting in a decrease of leaf photosynthetic capacity. In seedlings, a symptom known as damping off occurs, and is characterized by the rotting of the plants stem base and roots, wilting, and causing the death of young plants as a consequence of the infection [48,50,55].
Alternaria is transmitted to the seed and considered a seedborne pathogen. At the seed level, Alternaria infection can adversely affect seed quality, germination rates, and crop yield. Several factors influence disease development and severity, including moisture, low light intensity, temperature, plant age, and conidia density [49,52]. The impact of infection on agricultural production and the search for solutions to reduce its harmful effects motivate the study of the mechanisms of interaction between plants of agronomic interest and Alternaria.
4. Seed defense mechanisms and interaction with the necrotrophic seedborne fungi Alternaria brassicicola
4.1. A silent interaction with the seed
Mechanisms of transgenerational transmission of pathogens are poorly documented because few infection symptoms can be detected on seeds. Many seedborne pathogens are considered non-hosts during seed development [56,57]. A striking example is the bacterial pathogen Xanthomonas campestris pv. campestris, known for attacking cruciferous plants [56]. In beans, this pathogen is transmitted from seed to seedling. Interestingly, its growth in these plants is like how it behaves in plants contaminated with X. citri pv. phaseoli var. fuscans (a bean-related pathogen). This does not significantly affect seed germination or seedling development, and it does not cause visible disease symptoms. Additionally, the gene (T3SS) that usually defends plants against this pathogen does not seem to be necessary in these non-host plants [56].
4.2. Interaction with the necrotrophic seedborne fungi Alternaria brassicicola
Because seeds contribute to reproductive success, it is hypothesis that seed protection mechanisms would be part of their adaptive traits [24]. At the time of dissemination, the mature seed is already equipped with protection systems [5,24] that could contribute to non-host resistance. This could be illustrated by the accumulation in the mature seed of secondary metabolites such as phenolic compounds [25] and by the development of physical barriers such as the seed coat [5,24]. Also, pending seed germination and seedling establishment, these properties enable seeds to maintain their survival when challenged by abiotic and biotic pressures in their environment [8,24,25,58,59].
The endosperm acts not only in the control of seed germination but also as a protective barrier during germination [5,60]. Genes encoding detoxifying enzymes, such as glutathione S-transferases and peroxidases, play a crucial role in protecting embryo against ROS that could result from pathogen interaction during seed germination. Also, genes related to plant-fungus interactions and hormonal metabolism hold crucial roles in enhancing immunity and regulating plant growth. A transcriptomic study showed an over-accumulation of genes responsible for synthesizing salicylic acid (SA), indole-3-acetic acid (IAA), and amino synthetases (GH3) [60]. At the transcriptome level, the activation of secondary metabolites and defense response in the endosperm illustrates the activation of defense pathways during germination [60]. It is quite remarkable that certain mechanisms in seeds seem to differ from those in plants. Nevertheless, there are also instances of similar defense mechanisms being observed. For example, this includes plant defense through polyphenol oxidase (PPO), a group of enzymes that catalyze the oxidation of hydroxy phenols, yielding products with antimicrobial properties. In dormant wild oat (Avena fatua L.) seeds attacked by Fusarium avenaceum trigger post-translational activation of the PPO, demonstrating the seed ability to induce enzymatic biochemical defenses against pathogenic fungi. This PPO activity was also found to be induced in non-living caryopsis cover tissues, such as lemma and pale [25].
The mature seed has developed resistance properties to face biotic and abiotic stress after dispersal. However, the immune responses of seeds to biotic interactions during germination have been little studied, although this step is crucial to control seed-borne pathogen transmission to the new plant [21,24]. The model pathosystem Arabidopsis thaliana/Alternaria brassicicola has been recently used to describe gene regulation induced by the necrotrophic interaction during germination and seedling establishment [1,21,61].
RNAseq data from Alternaria-infected and healthy Arabidopsis seeds were compared at different time points of the germination kinetics until the stage of seedling establishment. Although a transcriptomic study carried out on inoculated rosette leaves 23 highlighted an induction by Alternaria of the jasmonate pathway and camalexin metabolism (Figure 1b), the response of the germinating seed rather reflected an activation of the SA and ET pathways to the detriment of the jasmonate pathway [1]. It is documented that this type of unexpected response could constitute a diversion towards inappropriate defense pathways of the plant by the fungus to induce a SR favoring its development [38,39]. Additionally, genes relate to response to hypoxia and indole-derived metabolites such as GSL were induced by Alternaria during germination [1]. Gene ontology analysis showed a stronger and more significant association among genes related to the response to hypoxia at an early development stage. Their fold enrichment decreased as seedling establishment progressed (3 Days After Sowing (DAS): 7.08, 6 DAS: 4.29 and 10 DAS: 3.07) [1]. This response is surprising, but it illustrates a competition for oxygen which benefits the physiology of the early imbibed seed. Seed germination is well adapted to low oxygen levels [62] while these low oxygen levels are limiting the growth of the fungus. Either the induction at the RNA level of indole GSL metabolism that was observed in Arabidopsis seeds [1] is well described [63].
Phenotyping analyses of mutant seeds deficient for the identified pathways showed that the mutants deficient for ethylene response ein2 and etr1 exhibited a lower rate of necrosis, respectively 21.9% and 55.4% lower than the WT control. Also, GSL-deficient mutants (qk0 and gtr1gtr2) exhibited a lower rate of necrosis, respectively 65.9% and 73.6% lower than those in the control group. Noticeably, the GSL-deficient mutants displayed also a reduced Alternaria overgrowth [1], indicating that the lack of necrosis, usually mediated by Alternaria-induced GSL pathway in WT, would be limiting for the colonization by the necrotrophic fungus. In this context, the induction of GSL metabolism could also contribute to the SR of the infected seed. The identified changes in gene regulation induced by Alternaria in germinating seed provide an original working model illustrating a simultaneous existence of SR and competition strategy (Figure 2).
This study reinforces the hypothesis that the interaction of the seed with the pathogen differs from the models described for whole plants. It is notable that the transcriptome of infected tissues differed drastically between 6-day-old seedlings and 10-day-old seedlings [1,61]. As an additional illustration, necrosis symptoms (Figure 3) in newly germinated seedlings (4 days after sowing) show much more delineated patterns than in older seedlings (after 10 days), where necrosis is more diffuse and widespread in the infected tissue.
We hypothesize that at the germinative stage, the young seedling reacts to infection with a HR whose profile is generally well circumscribed. The fact that GLS-deficient mutants did not show any necrosis symptoms at the young seedling stage (4 days after sowing) suggests that the observed HR would be mediated by activation of the plant's GSL pathway, whereas after 6 and 10 days of development, new necrosis were observed, including in the GSL mutants. The diffused appearance of the latter would result from an effect of the fungus as it penetrates the plant tissue.
5. Future perspectives
Despite significant advancements, there remain substantial gaps in our understanding of seed immunity. Recent molecular and genetic analyses of Arabidopsis interactions with Alternaria during germination and seedling emergence have highlighted a particular defense mechanism and adaptive strategies employed when competitiveness is crucial for the survival of both the fungus and the germinating seedling [1]. The Arabidopsis thaliana /Alternaria brassicicola pathosystem represents a valuable model for unraveling the intricate molecular dynamics of their interaction [1,21]. Particularly, the contribution of the GSL metabolism to necrosis symptoms and Alternaria colonization in germinating seeds [1] need to be described in more detail and could provide strategic lever to control seed tolerance to necrotrophic attack and manage transgenerational pathogen propagation.
Arabidopsis is a widely studied model species which presents advantages of a wide diversity of well-characterized genetic, genomic, and phenotypic resources [64,65,66]. Although it lacks direct economic relevance, its close phylogenetic relationship (genetic and molecular similarities) with brassica species [67] allows to transfer knowledge acquired about seed fungal interaction in brassica crops. The leveraging Arabidopsis gene expression profiles, including transcriptomic analyses of deficient mutants in GSL, and in camalexin, along in a context of seed germination infected by Alternaria through Weighted Gene Correlation Network Analysis (WGCNA) approach [68], can unveil co-expression networks that group genes with similar expression patterns and phenotypic contribution. WGCNA approach would be helpful to identify master regulatory genes controlling seed tolerance and to provide candidate genes to search orthologs that contribute to seed resistance in brassica crops, such as Brassica oleracea.
Investigation of seed defense mechanisms against pathogenic fungi has become essential in the pursuit of strategies to ensure seed health and mitigate extensive losses in agricultural fields. Comparative genomics will provide deeper insights into the plant-pathogen interactions. This knowledge is needed to develop genetic resistance or to better manage diseases in brassica crops.
Author Contributions
Conceptualization, P.G., T.A., A.V-P., and A.R.; methodology, P.G., T.A., A.V-P., S.A., A.R., C.R-P., and M.O-C.; validation, P.G. and T.A.; formal analysis, M.O-C., S.A., and C.R-P.; investigation, P.G., A.R., C.R-P., and M.O-C.; writing—original draft preparation, M.O-C.; writing—review and editing, P.G., T.A., and M.O-C.; visualization, P.G., T.A., and A.V-P.; supervision, P.G., T.A., and A.V-P.; project administration, P.G.; funding acquisition, P.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Programme Prioritaire de Recherche (PPR) de l’ANR, Cultiver et Protéger Autrement (CPA), SUCSEED (https://anr.fr/ProjetIA-20-PCPA-0009).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Acknowledgments
We would like to thank the FUNGISEM team for their support in this investigation and thank Luz Marina Melgarejo, Adriana Tofiño, and Nubia Velazquez for their productive discussion about this project. We would like to express our gratitude to Jerome Verdier for his invaluable assistance in conducting experimental analyses and interpreting data. We thank Fabienne Simonneau of IMAC imaging plateau of the SFR 4207 QUASAV.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Ortega-Cuadros, M.; De Souza, T.L.; Berruyer, R.; Aligon, S.; Pelletier, S.; Renou, J.-P.; Arias, T.; Campion, C.; Guillemette, T.; Verdier, J.; Grappin, P. Seed Transmission of Pathogens: Non-Canonical Immune Response in Arabidopsis Germinating Seeds Compared to Early Seedlings against the Necrotrophic Fungus Alternaria Brassicicola. Plants 2022, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd Edition; Springer New York, 2013; Vol. 978146144 6934.
- Gupta, S.; Van Staden, J.; Doležal, K. An Understanding of the Role of Seed Physiology for Better Crop Productivity and Food Security. Plant Growth Regul 2022, 97, 171–173. [Google Scholar] [CrossRef]
- Nonogaki, H. Seed Biology Updates - Highlights and New Discoveries in Seed Dormancy and Germination Research. Front Plant Sci 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H. Seed Germination and Dormancy: The Classic Story, New Puzzles, and Evolution. J Integr Plant Biol 2019, 61, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H. Seed Dormancy and Germination—Emerging Mechanisms and New Hypotheses. Front Plant Sci 2014, 5, 233. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.B. The Seed Microbiome: Origins, Interactions, and Impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- War, A.F.; Bashir, I.; Reshi, Z.A.; Kardol, P.; Rashid, I. Insights into the Seed Microbiome and Its Ecological Significance in Plant Life. Microbiol Res 2023, 269, 127318. [Google Scholar] [CrossRef]
- Buitink, J.; Leprince, O. Advances in Seed Science and Technology for More Sustainable Crop Production, 1st ed.; Burleigh Dodds Science Publishing, 2022.
- Dell’Olmo, E.; Tiberini, A.; Sigillo, L. Leguminous Seedborne Pathogens: Seed Health and Sustainable Crop Management. Plants 2023, 12, 2040. [Google Scholar] [CrossRef]
- Cram, M.M.; Fraedrich, S.W. Seed Diseases and Seedborne Pathogens of North America. Tree Planters’ Notes 2010, 53. [Google Scholar]
- Kumar, R.; Gupta, A. Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management; 2020. [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant–Pathogen Interactions. Nat Rev Genet 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Harper, A.L.; Trick, M.; Higgins, J.; Fraser, F.; Clissold, L.; Wells, R.; Hattori, C.; Werner, P.; Bancroft, I.; Gullino, M.L.; Munkvold, G. Global Perspectives on the Health of Seeds and Plant Propagation Material; Springer Netherlands: Dordrecht, 2014; Vol. Plant Path.
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. https://doi.org/10.1146/annurev.phyto.43.040204.135923 2005, 43, 205–227. [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition Acquisition Strategies during Fungal Infection of Plants. FEMS Microbiol Lett 2007, 266, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ghozlan, M.H.; EL-Argawy, E.; Tokgöz, S.; Lakshman, D.K.; Mitra, A.; Ghozlan, M.H.; EL-Argawy, E.; Tokgöz, S.; Lakshman, D.K.; Mitra, A. Plant Defense against Necrotrophic Pathogens. Am J Plant Sci 2020, 11, 2122–2138. [Google Scholar] [CrossRef]
- Bolton, M.D. Primary Metabolism and Plant Defense—Fuel for the Fire. https://doi.org/10.1094/MPMI-22-5-0487 2009, 22, 487–497. [CrossRef]
- Łaźniewska, J.; Macioszek, V.K.; Kononowicz, A.K. Plant-Fungus Interface: The Role of Surface Structures in Plant Resistance and Susceptibility to Pathogenic Fungi. Physiol Mol Plant Pathol 2012, 78, 24–30. [Google Scholar] [CrossRef]
- Pochon, S.; Terrasson, E.; Guillemette, T.; Iacomi-Vasilescu, B.; Georgeault, S.; Juchaux, M.; Berruyer, R.; Debeaujon, I.; Simoneau, P.; Campion, C. The Arabidopsis Thaliana-Alternaria Brassicicola Pathosystem: A Model Interaction for Investigating Seed Transmission of Necrotrophic Fungi. Plant Methods 2012, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Łaźniewska, J.; Macioszek, V.K.; Lawrence, C.B.; Kononowicz, A.K. Fight to the Death: Arabidopsis Thaliana Defense Response to Fungal Necrotrophic Pathogens. Acta Physiol Plant 2010, 32, 1–10. [Google Scholar] [CrossRef]
- van Wees, S.C.M.; Chang, H.; Zhu, T.; Glazebrook, J. Characterization of the Early Response of Arabidopsis to Alternaria Brassicicola Infection Using Expression Profiling. Plant Physiol 2003, 132, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Dalling, J.W.; Davis, A.S.; Arnold, A.E.; Sarmiento, C.; Zalamea, P.-C. Extending Plant Defense Theory to Seeds. Annu Rev Ecol Evol Syst 2020, 51, 123–141. [Google Scholar] [CrossRef]
- Fuerst, E.P.; Okubara, P.A.; Anderson, J.V.; Morris, C.F. Polyphenol Oxidase as a Biochemical Seed Defense Mechanism. Front Plant Sci 2014, 5, 117840. [Google Scholar] [CrossRef]
- Panstruga, R.; Moscou, M.J. What Is the Molecular Basis of Nonhost Resistance? Molecular Plant-Microbe Interactions, 2020. [Google Scholar] [CrossRef]
- Macioszek, V.K.; Jęcz, T.; Ciereszko, I.; Kononowicz, A.K. Jasmonic Acid as a Mediator in Plant Response to Necrotrophic Fungi. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant Antimicrobial Peptides: Structures, Functions, and Applications. Botanical Studies. 2021. [CrossRef] [PubMed]
- Stotz, H.U.; Sawada, Y.; Shimada, Y.; Hirai, M.Y.; Sasaki, E.; Krischke, M.; Brown, P.D.; Saito, K.; Kamiya, Y. Role of Camalexin, Indole Glucosinolates, and Side Chain Modification of Glucosinolate-Derived Isothiocyanates in Defense of Arabidopsis against Sclerotinia Sclerotiorum. Plant Journal 2011, 67, 81–93. [Google Scholar] [CrossRef] [PubMed]
- L. Schlaich, N. Arabidopsis Thaliana- The Model Plant to Study Host-Pathogen Interactions. Curr Drug Targets 2011, 12, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Balint-Kurti, P. The Plant Hypersensitive Response: Concepts, Control and Consequences. Mol Plant Pathol 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [PubMed]
- Ádám, A.L.; Nagy, Z.; Kátay, G.; Mergenthaler, E.; Viczián, O. Signals of Systemic Immunity in Plants: Progress and Open Questions. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U. Systemic Acquired Resistance. Plant Signal Behav 2006, 1, 179–184. [Google Scholar] [CrossRef]
- Sellam, A.; Iacomi-Vasilescu, B.; Hudhomme, P.; Simoneau, P. In Vitro Antifungal Activity of Brassinin, Camalexin and Two Isothiocyanates against the Crucifer Pathogens Alternaria Brassicicola and Alternaria Brassicae. Plant Pathol 2007, 56, 296–301. [Google Scholar] [CrossRef]
- Gill, U.S.; Lee, S.; Mysore, K.S. Host versus Nonhost Resistance: Distinct Wars with Similar Arsenals. Phytopathology 2015, 105, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Nürnberger, T.; Lipka, V. Non-Host Resistance in Plants: New Insights into an Old Phenomenon. Molecular Plant Pathology. 2005. [CrossRef]
- Schulze-Lefert, P.; Panstruga, R. A Molecular Evolutionary Concept Connecting Nonhost Resistance, Pathogen Host Range, and Pathogen Speciation. Trends in Plant Science. 2011. [CrossRef] [PubMed]
- Patkar, R.N.; Naqvi, N.I. Fungal Manipulation of Hormone-Regulated Plant Defense. PLoS Pathog 2017, 13, e1006334. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Smith, D.L.; Kabbage, M.; Roth, M.G. Effectors of Plant Necrotrophic Fungi. Front Plant Sci 2021, 12, 995. [Google Scholar] [CrossRef] [PubMed]
- Gorshkov, V.; Tsers, I. Plant Susceptible Responses: The Underestimated Side of Plant–Pathogen Interactions. Biological Reviews 2022, 97, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Frías, M.; González, C.; Brito, N. BcSpl1, a Cerato-Platanin Family Protein, Contributes to Botrytis Cinerea Virulence and Elicits the Hypersensitive Response in the Host. New Phytologist 2011, 192, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Qiu, D.; Zeng, H.; Guo, L.; Yang, X. BcGs1, a Glycoprotein from Botrytis Cinerea, Elicits Defence Response and Improves Disease Resistance in Host Plants. Biochem Biophys Res Commun 2015, 457, 627–634. [Google Scholar] [CrossRef] [PubMed]
- El-Oirdi, M.; El-Rahman, T.A.; Rigano, L.; El-Hadrami, A.; Rodriguez, M.C.; Daayf, F.; Vojnov, A.; Bouarab, K. Botrytis Cinerea Manipulates the Antagonistic Effects between Immune Pathways to Promote Disease Development in Tomato. Plant Cell 2011, 23, 2405. [Google Scholar] [CrossRef] [PubMed]
- González-Lamothe, R.; El Oirdi, M.; Brisson, N.; Bouarab, K. The Conjugated Auxin Indole-3-Acetic Acid–Aspartic Acid Promotes Plant Disease Development. Plant Cell 2012, 24, 762. [Google Scholar] [CrossRef] [PubMed]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H. Da; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science (1979) 2013, 342. [Google Scholar] [CrossRef]
- German, D.A.; Hendriks, K.P.; Koch, M.A.; Lens, F.; Lysak, M.A.; Bailey, C.D.; Mummenhoff, K.; Al-Shehbaz, I.A. An Updated Classification of the Brassicaceae (Cruciferae). PhytoKeys 2023, 220, 127. [CrossRef]
- Rakow, G. Species Origin and Economic Importance of Brassica. 2004, 3–11. [CrossRef]
- Cho, Y. How the Necrotrophic Fungus Alternaria Brassicicola Kills Plant Cells Remains an Enigma. Eukaryot Cell 2015, 14, 335–344. [Google Scholar] [CrossRef]
- Dharmendra, K.; Neelam, M.; Yashwant, K.B.; Ajay, K.; Kamlesh, K.; Kalpana, S.; Gireesh, C.; Chanda, K.; Sushil, K.S.; Raj, K.M.; Adesh, K. Alternaria Blight of Oilseed Brassicas: A Comprehensive Review. Afr J Microbiol Res 2014, 8, 2816–2829. [Google Scholar] [CrossRef]
- Ali, S.; Tyagi, A.; Rajarammohan, S.; Mir, Z.A.; Bae, H. Revisiting Alternaria-Host Interactions: New Insights on Its Pathogenesis, Defense Mechanisms and Control Strategies. Sci Hortic 2023, 322, 112424. [Google Scholar] [CrossRef]
- Mamgain, A.; Roychowdhury, R.; Tah, J. Alternaria Pathogenicity and Its Strategic Controls. Research & Reviews: Journal of Biology 2013, 1, 1–9. [Google Scholar]
- Nowicki, M.; Nowakowska, M.; Niezgoda, A.; Kozik, E. Alternaria Black Spot of Crucifers: Symptoms, Importance of Disease, and Perspectives of Resistance Breeding. Journal of Fruit and Ornamental Plant Research 2012, 76, 5–19. [Google Scholar] [CrossRef]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria Toxins: Potential Virulence Factors and Genes Related to Pathogenesis. Frontiers in Microbiology. 2017. [CrossRef]
- Pedras, M.S.C.; Chumala, P.B.; Jin, W.; Islam, M.S.; Hauck, D.W. The Phytopathogenic Fungus Alternaria Brassicicola: Phytotoxin Production and Phytoalexin Elicitation. Phytochemistry 2009, 70. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Mitchell, T.K.; Craven, K.D.; Cho, Y.; Cramer, R.A.; Kim, K.H. At Death’s Door: Alternaria Pathogenicity Mechanisms. Plant Pathol J (Faisalabad) 2008, 24, 101–111. [Google Scholar] [CrossRef]
- Darrasse, A.; Darsonval, A.; Boureau, T.; Brisset, M.N.; Durand, K.; Jacques, M.A. Transmission of Plant-Pathogenic Bacteria by Nonhost Seeds without Induction of an Associated Defense Reaction at Emergence. Appl Environ Microbiol 2010, 76, 6787–6796. [Google Scholar] [CrossRef]
- Darsonval, A.; Darrasse, A.; Meyer, D.; Demarty, M.; Durand, K.; Bureau, C.; Manceau, C.; Jacques, M.A. The Type III Secretion System of Xanthomonas Fuscans Subsp. Fuscans Is Involved in the Phyllosphere Colonization Process and in Transmission to Seeds of Susceptible Beans. Appl Environ Microbiol 2008, 74. [Google Scholar] [CrossRef]
- Debeaujon, I.; Léon-Kloosterziel, K.M.; Koornneef, M. Influence of the Testa on Seed Dormancy, Germination, and Longevity in Arabidopsis. Plant Physiol 2000, 122. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The Role of the Testa during Development and in Establishment of Dormancy of the Legume Seed. Frontiers in Plant Science. 2014. [CrossRef]
- Endo, A.; Tatematsu, K.; Hanada, K.; Duermeyer, L.; Okamoto, M.; Yonekura-Sakakibara, K.; Saito, K.; Toyoda, T.; Kawakami, N.; Kamiya, Y.; Seki, M.; Nambara, E. Tissue-Specific Transcriptome Analysis Reveals Cell Wall Metabolism, Flavonol Biosynthesis and Defense Responses Are Activated in the Endosperm of Germinating Arabidopsis Thaliana Seeds. Plant Cell Physiol 2012, 53, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Cuadros, M.; Chir, L.; Aligon, S.; Arias, T.; Verdier, J.; Grappin, P. Dual-Transcriptomic Datasets Evaluating the Effect of the Necrotrophic Fungus Alternaria Brassicicola on Arabidopsis Germinating Seeds. Data Brief 2022, 44, 108530. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Lee, Y.H. Hypoxia: A Double-Edged Sword During Fungal Pathogenesis? Front Microbiol 2020, 11, 1920. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Miao, H.; Chen, L.; Wang, M.; Xia, C.; Zeng, W.; Sun, B.; Zhang, F.; Zhang, S.; Li, C.; Wang, Q. WRKY33-Mediated Indolic Glucosinolate Metabolic Pathway Confers Resistance against Alternaria Brassicicola in Arabidopsis and Brassica Crops. J Integr Plant Biol 2022, 64. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.W.; Hancock, A.M.; Huang, Y.S.; Toomajian, C.; Atwell, S.; Auton, A.; Muliyati, N.W.; Platt, A.; Sperone, F.G.; Vilhjálmsson, B.J.; Nordborg, M.; Borevitz, J.O.; Bergelson, J. Genome-Wide Patterns of Genetic Variation in Worldwide Arabidopsis Thaliana Accessions from the RegMap Panel. Nat Genet 2012, 44, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Alonso-Blanco, C.; Vreugdenhil, D. NATURALLY OCCURRING GENETIC VARIATION IN ARABIDOPSIS THALIANA. Annu Rev Plant Biol 2004, 55, 141–172. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.; Koornneef, M. A Fortunate Choice: The History of Arabidopsis as a Model Plant. Nat Rev Genet 2002, 3, 883–889. [Google Scholar] [CrossRef]
- Nikolov, L.A.; Shushkov, P.; Nevado, B.; Gan, X.; Al-Shehbaz, I.A.; Filatov, D.; Bailey, C.D.; Tsiantis, M. Resolving the Backbone of the Brassicaceae Phylogeny for Investigating Trait Diversity. New Phytologist 2019, 222, 1638–1651. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinformatics 2008, 9. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
General model of plant defense signaling and immune responses. (a) Overview of the plant's signal detection mechanisms involving interactions with herbivorous, beneficial, and pathogenic microorganisms. (b) The plant immune response against necrotrophs and biotrophs involves diverse defenses, including physical barriers, hormonal signaling, and antimicrobial metabolites. Swift responses, initiated by Pathogen Recognition Receptors (PRRs), progress from P/MAMP-triggered Immunity (PTI) to Effector-Triggered Immunity (ETI) upon evasion by pathogens.
Figure 1.
General model of plant defense signaling and immune responses. (a) Overview of the plant's signal detection mechanisms involving interactions with herbivorous, beneficial, and pathogenic microorganisms. (b) The plant immune response against necrotrophs and biotrophs involves diverse defenses, including physical barriers, hormonal signaling, and antimicrobial metabolites. Swift responses, initiated by Pathogen Recognition Receptors (PRRs), progress from P/MAMP-triggered Immunity (PTI) to Effector-Triggered Immunity (ETI) upon evasion by pathogens.
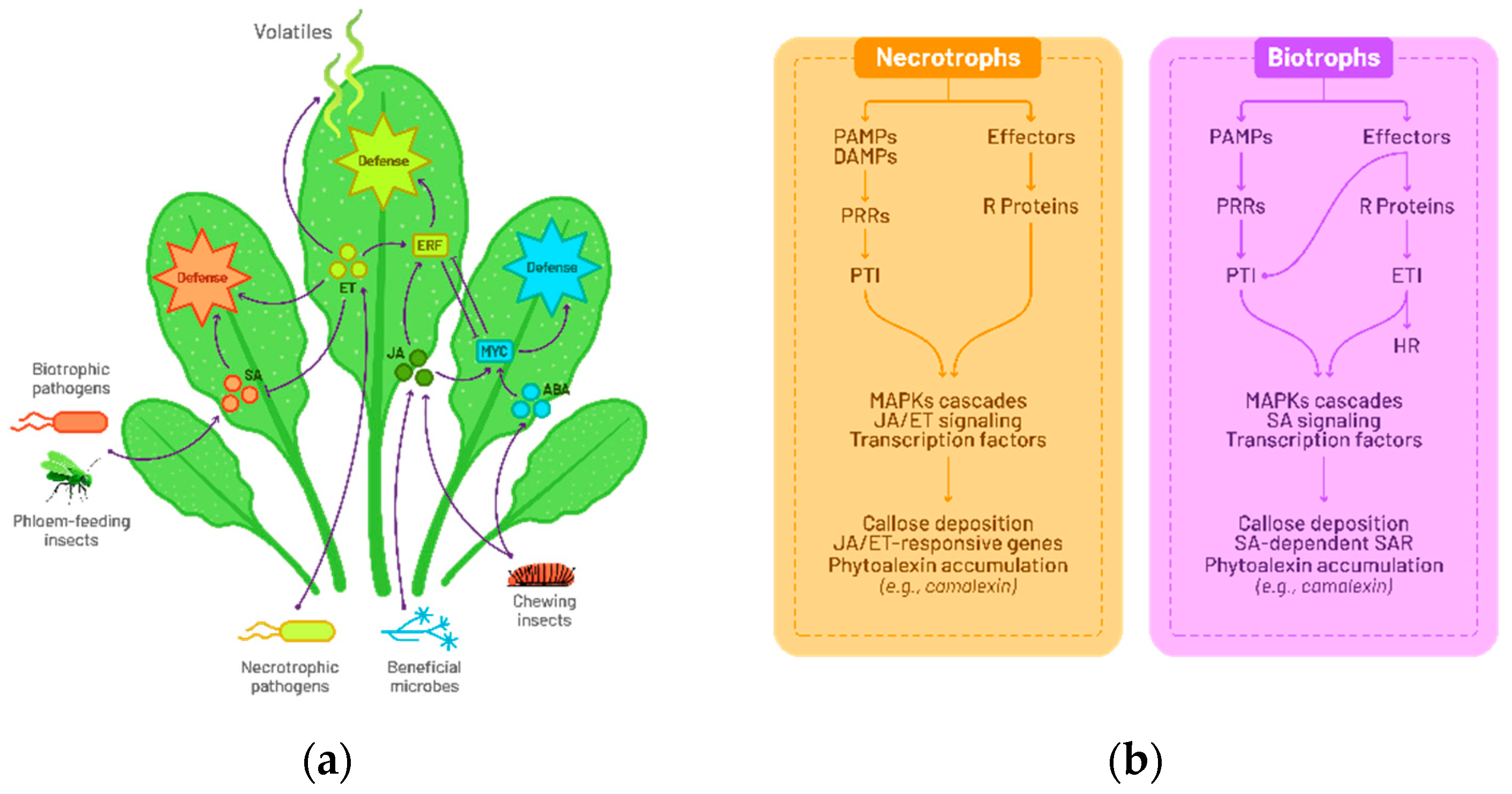
Figure 2.
Working model illustrating a Susceptible Response (SR) to Alternaria in germinating seeds. Proposed regulations have been provided by transcriptomic and mutant analyses [1]. Alternaria induced at the RNA level salicylic acid (SA) pathway, indole-glucosinolate (GSL) metabolism as well as ethylene (ET) signalling and the response to hypoxia. SA pathway is usually induced to the detriment to jasmonic acid (JA) response that has not been found repressed in the transcriptomic data. Mutant analyses illustrated the contribution of GSL and of response to ET in the occurrence of necrosis symptoms that are needed for Alternaria development. Response to hypoxia have been also documented to induce ET signalling.
Figure 2.
Working model illustrating a Susceptible Response (SR) to Alternaria in germinating seeds. Proposed regulations have been provided by transcriptomic and mutant analyses [1]. Alternaria induced at the RNA level salicylic acid (SA) pathway, indole-glucosinolate (GSL) metabolism as well as ethylene (ET) signalling and the response to hypoxia. SA pathway is usually induced to the detriment to jasmonic acid (JA) response that has not been found repressed in the transcriptomic data. Mutant analyses illustrated the contribution of GSL and of response to ET in the occurrence of necrosis symptoms that are needed for Alternaria development. Response to hypoxia have been also documented to induce ET signalling.

Figure 3.
Necrosis profiles at 4, 6 and 10 days after sowing (DAS). Microscopic view (objective X 20) of necrotic symptoms on Alternaria-infected Arabidopsis cotyledons in visible and fluorescent light. The emission corresponding to the necrotic area, shown in red (false color), is revealed using a long-pass GFP filter. At the early germinative station stage (4 DAS), necrosis occurs in well-defined areas, whereas at later stages (6 and 10 DAS), necrosis is more diffuse. GSL mutants show no necrosis at 3 DAS. Scale bar represents 50 µm.
Figure 3.
Necrosis profiles at 4, 6 and 10 days after sowing (DAS). Microscopic view (objective X 20) of necrotic symptoms on Alternaria-infected Arabidopsis cotyledons in visible and fluorescent light. The emission corresponding to the necrotic area, shown in red (false color), is revealed using a long-pass GFP filter. At the early germinative station stage (4 DAS), necrosis occurs in well-defined areas, whereas at later stages (6 and 10 DAS), necrosis is more diffuse. GSL mutants show no necrosis at 3 DAS. Scale bar represents 50 µm.
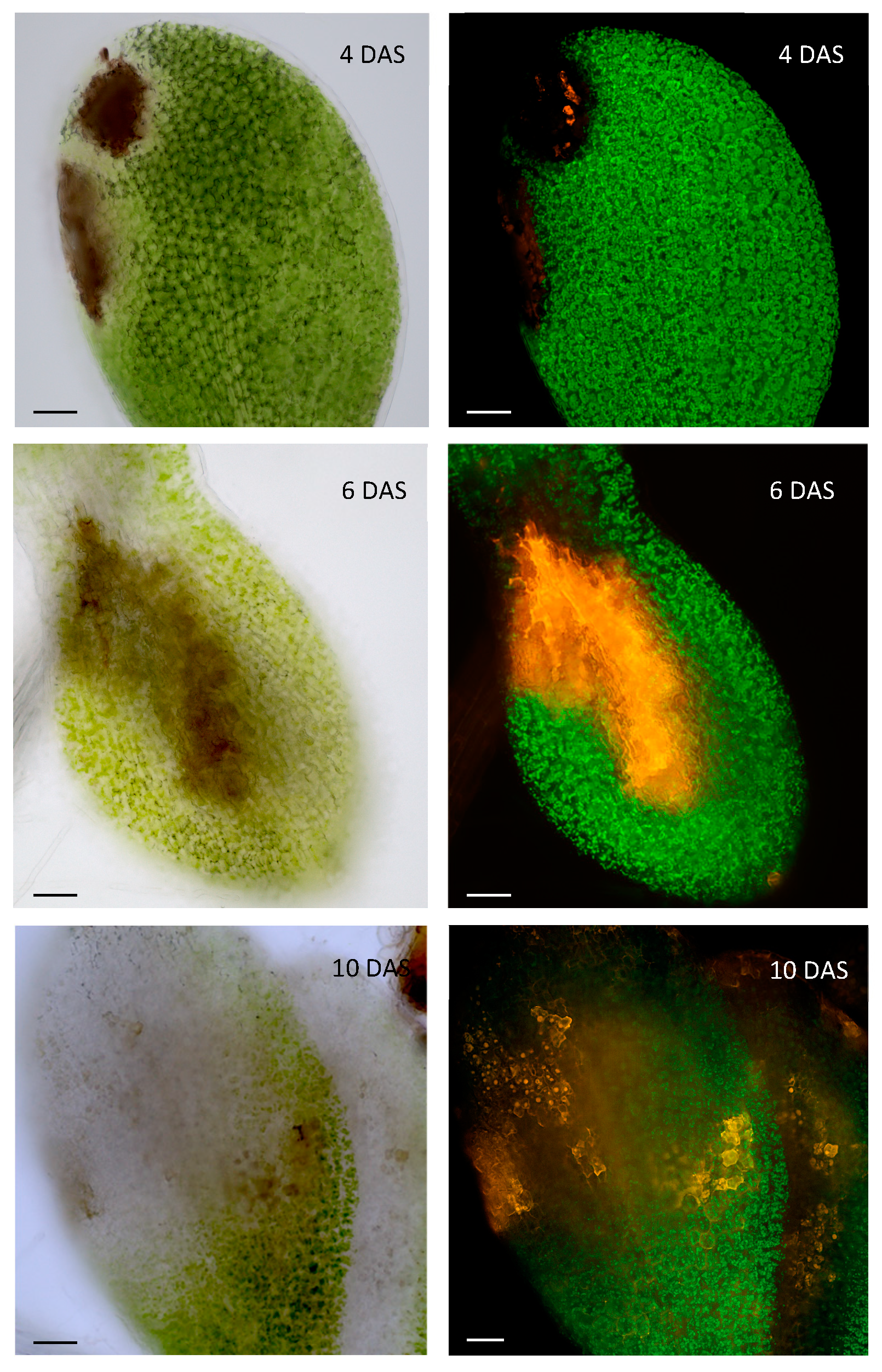

Table 1.
Botrytis cinerea strategies to hijack the plant immune response.
| Host | Manipulate mechanism | Elicitors or effectors | Reference |
|---|---|---|---|
| Tomato, Tobacco, and Arabidopsis | Programmed cell death (PCD) | Cerato-platanin family proteins (CPPs) – BcSpl Necrosis-inducing proteins (NIPs) - BcGs1. |
Frías et al. [41] Zhang et al. [42] |
| Tomato | SA pathway | Exopolysaccharide (EPS) - β-(1,3)(1,6)-d-glucan | El Oirdi M et al. [43] |
| Arabidopsis | Auxin pathway | Auxin Indole-3-Acetic Acid–Aspartic Acid -IAA-Aspartate | González-Lamothe R et al. [44] |
| Tomato and Arabidopsis | Peroxiredoxin (oxidative stress-related gene), mitogen activated protein kinases (MPK1, MPK2, MPKKK4), and a cell wall-associated kinase (WAK) | sRNAs: Bc-siR3.1 Bc-siR3.2 Bc-siR5 |
Weiberg et al. [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



