Submitted:
01 January 2024
Posted:
03 January 2024
You are already at the latest version
Abstract
In the past decade, it has been discovered that intestinal flora can influence various organ-specific cancers by altering the body's energy balance, increasing obesity, synthesizing genetic toxins and small signaling molecules, initiating and modulating immune responses, and metabolizing indigestible dietary components, exogenous substances, and drugs. In this review, we will focus on elucidating the role of gut flora based on its molecular mechanisms and its potential therapeutic implications in head and neck tumors in the near future.
Keywords:
intestinal flora
; head and neck tumors
; tumor immunity
; therapy
1. Introduction
Gut flora is the microbial community in the human gut, consisting of a wide range of microorganisms including bacteria, fungi, archaea and viruses. These microorganisms form complex interactions with the human host. Gut flora plays an important role in human health and disease development [1]. For a long time, it has been believed that the role of intestinal flora is mainly limited to helping food decomposition and absorption, in fact, the role of intestinal flora is not only limited to helping food decomposition and absorption, but also involves many other aspects. In recent years, more and more studies have shown that gut flora is closely related to human health [2]. With increasing understanding, it has been found that the microbiota plays an important role in areas such as pro- or anti-inflammation, and that maintaining a relative balance between them helps to maintain intestinal health and immune homeostasis [3,4]. When the balance of intestinal flora is disturbed, it can negatively affect host metabolism or immune responses and functions, which can lead to the development of immune-related diseases such as inflammatory bowel disease, autoimmune diseases, and even tumors, and H. pylori has been classified as a class I carcinogen, which attacks the gastric mucosal cells by secreting a variety of toxins and enzymes, triggering inflammatory responses and tissue damage, and it may also induce other changes of intestinal microbiota, leading to the development of intestinal microflora [3,4]. flora changes, leading to an imbalance of intestinal flora, which can further exacerbate the occurrence of intestinal diseases [5,6]. In addition, strains of Escherichia coli, Shigella flexneri, and Bacteroides fragilis can produce a toxin known as poly-ADP-ribose, which may lead to inflammatory responses and DNA damage, thereby promoting tumorigenesis [5,6]. damage, which in turn promotes tumorigenesis and development [7,8,9]. Whereas Bifidobacteria and Lactobacilli play an important role in the intestinal flora, they are thought to be associated with reduced tumor incidence. They can promote the balance of beneficial flora in the intestinal tract and inhibit the growth of harmful bacteria, thus maintaining intestinal health, and have a certain immunomodulatory effect, which can inhibit the inflammatory response by regulating the host's immune system, thus contributing to the reduction of the incidence of certain tumors [10]. Immunotherapy for cancer, especially the treatment of immune checkpoint inhibitors (ICHIs), has made some remarkable progress. However, there is significant variation in the response of some patients to immunotherapy, with some patients being ineffective. It was found that this variation may be related to differences in the patients' gut microbiome. By studying the gut microbiome of patients receiving immunotherapy, scientists have found significant differences in the gut microbiomes of patients who are insensitive or ineffective to immunotherapy compared to those who are sensitive to immunotherapy. Several studies have found that the abundance of certain bacterial flora correlates with the efficacy of immunotherapy, and that specific flora may influence the host's immune response and thus the efficacy of immunotherapy [11,12].
The structure and composition of the gut microbiome may have an important impact on the efficacy of immunotherapy. In the future, modulating the gut microbiome, for example by using probiotics or other methods, may help to increase the response rate to immunotherapy and improve the efficacy of immunotherapy. This also provides new ideas and possibilities for individualized medicine and precision medicine. Of course, more in-depth studies in this field are still needed to fully understand the complex relationship between the gut microbiome and immunotherapy. Therefore, the importance of microbiota in tumor development, diagnosis and treatment needs to be clarified. This article reviews the molecular mechanisms of gut flora and their role in the pathogenesis and progression of various head and neck tumors, and discusses their potential as therapeutic targets.
2. History of Intestinal Flora
In 1683, Anton van Leeuwenhoek observed microorganisms in the gastrointestinal tract through a microscope in the 17th century, which can be considered as one of the earliest perceptions of intestinal flora.Subsequent studies gradually revealed the importance of intestinal flora for human health [13].In 1908 Ilya Mechnikov proposed that lactobacilli are beneficial to health and that human health can be improved by altering intestinal flora. This is considered to be the starting point of intestinal flora research [14].In 1981, several studies quantitatively analyzed the early development of intestinal flora [15]. The influence of gut flora on immune development can be traced back as far as the 1990s, where attention began to be paid to the influence of gut flora on immune development. It was found that a large number of microorganisms coexist in the human body, including bacteria and fungi [16,17].In 1996, Wilson and Blitchington gained insight into the gut flora by sequencing human feces using 16S rRNA technology. [In 2001, Joshua Lederberg coined the term microbiome to emphasize that commensal microorganisms form a whole with humans and play a protective role in our health [19].In 2003, viral macro genome analysis from human feces revealed that viruses and fungi are also important members of microorganisms that have an impact on human health Since then, studies have demonstrated that other microorganisms are equally important for human health [20].In 2007, Eline Klaassens et al. used a macro-proteomics approach to analyze non-cultured fecal flora, providing insights beyond the taxonomic identification of flora [21]. In recent years, through the Human Gut Macrogenomics and Human Microbiome projects, we have gained a deeper understanding of the structure and function of the human microbiota, as well as its relationship to disease. Also, potential molecular mechanisms of gut flora-brain interactions have been revealed. These findings provide a solid foundation for further exploration of the role and applications of gut flora. The study of gut flora will help us to better understand the relationship between human health and microorganisms and provide guidance for the development of relevant treatments and interventions [22,23]. (Figure 1)
3. Mechanisms of Intestinal Flora in Disease
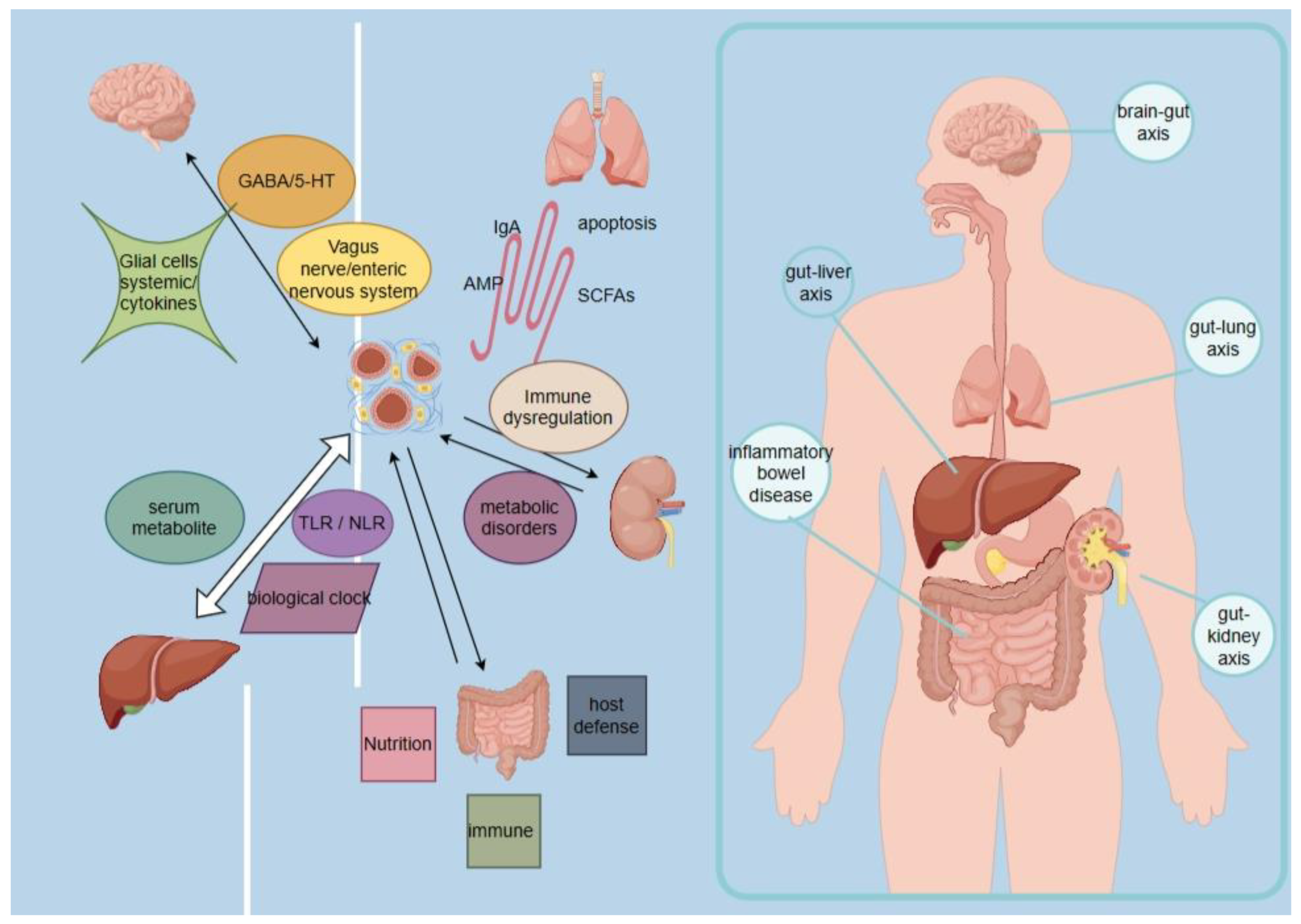
3.1. Gut Microbiota Regulates Brain Function through the "Gut-Brain Axis"
Recent studies have found a complex interaction between the gut flora and the brain. This two-way communication can be carried out through the neural, immune and endocrine systems and other pathways and media. First, gut flora can directly affect brain function through neural pathways. Microorganisms in the gut can produce and release a variety of chemicals, such as neurotransmitters and metabolites, which can enter the circulation, cross the blood-brain barrier, and affect brain function and behavioral performance. For example, the products of certain probiotics can modulate the levels of neurotransmitters, such as gamma-aminobutyric acid (GABA), which is important for the regulation of mood and anxiety. Second, there is a close connection between gut flora and the immune system. The gut is one of the largest immune organs in the body, and gut flora can interact with intestinal immune cells to regulate the immune response. Some studies have found that poor gut flora composition may be associated with autoimmune diseases such as leukemia and rheumatoid arthritis. In addition, gut flora can influence brain function through the endocrine system. Microorganisms in the gut can influence the synthesis and metabolism of a variety of endocrine substances, such as serotonin, dopamine, and hormones, which are important for mood, cognition, and behavior.
Some studies have shown that disturbances in gut flora are associated with the onset and progression of several brain disorders. For example, disorders such as stroke, epilepsy, autism, and Alzheimer's disease have been associated with abnormalities in the gut flora [24,25]. There are three main pathways of interaction between the gut microbiota and the brain axis: 1. Bacteria can act on the neuroendocrine system to regulate the concentrations of Y-aminobutyric acid (GABA) and pentraxin (5-HT), as well as other neurotransmitters; 2. Bacteria and their metabolites can also act on the vagus and enteric nervous systems to affect the brain and behavior; 3. The last one is the immune system, where microglia and systemic cytokines have key mediating roles. These pathways may be involved in the pathogenesis of neurodegenerative and other neurologically related diseases [26,27,28,29]. An in-depth study of these pathways will help us to better understand the complex relationship between the gut microbiota and the nervous system and provide new ideas and approaches for future disease treatment and prevention.
3.2. Intestinal Flora Regulates Liver Function through the "Intestinal-Liver Axis"
The "gut-liver axis" refers to the bi-directional communication between the liver and the intestine via the bile ducts, portal vein and the body circulation. There is a close interaction between the liver and the intestine and the liver, with liver products influencing the composition and barrier integrity of the intestinal flora, and intestinal factors regulating the synthesis of bile acids and glucolipid metabolism in the liver. In the liver and gut, pro-inflammatory changes play an important role in mediating the development of diseases that may lead to liver fibrosis, cirrhosis, and hepatocellular carcinoma. For example, patients with alcoholic and nonalcoholic fatty liver disease often exhibit features such as dysbiosis of the intestinal flora, increased permeability of the intestinal wall, and altered levels of bile acids, ethanol, and choline metabolites [30,31,32]. An in-depth study of the gut-liver axis interactions is important for our understanding of the mechanisms of liver disease onset and progression. Further exploration of this area will provide useful guidance for the development of new therapeutic strategies and preventive measures.
There are three main interaction pathways between the gut microbiota and the liver axis: 1). Dysbiosis of the gut microbiota may be associated with abnormal accumulation of serum metabolites; 2). Gut barrier disruption and induction of inflammatory responses in the TLR / NLR pathway; 3). The biological clock affects the gut microbiota mainly through diet, sleep, and light/dark cycles, and the gut flora regulates the expression of genes related to the biological clock, which in turn affects metabolism and immunity. First, dysbiosis of the gut microbiota may be associated with abnormal accumulation of serum metabolites; when the gut microbiota is dysbiotic, metabolite production and clearance may be compromised, leading to abnormal accumulation of metabolites in the serum, which in turn has an impact on liver function. Secondly, gut barrier disruption and inflammatory response of Toll-like receptor (TLR) / Nod-like receptor (NLR) pathway are also important pathways between the gut microbiota and the liver axis. An imbalance of gut microbes may lead to gut barrier disruption, allowing harmful substances to enter the blood circulation through the intestinal mucosa, stimulating inflammatory response of the TLR / NLR pathway, which may in turn lead to inflammation and injury of the liver. Finally, the biological clock plays an important role between the gut microbiota and the hepatic axis. The biological clock influences the composition and function of the gut microbiota by modulating lifestyle factors such as diet, sleep, and the light/dark cycle, and at the same time, the gut microbiota is able to regulate the expression of bioclimate-associated genes in the host, which in turn affects the host's metabolism and immunity [33,34,35,36]. The study of these interaction pathways provides important clues to our understanding of the relationship between the gut microbiota and liver health, which can help to deeply explore the pathogenesis of liver diseases and provide new ideas and strategies for corresponding treatment and prevention.
3.3. Gut flora Regulates Functions Related to Lung Diseases and Respiratory Infections through the Gut-Lung Axis (GLA)
Gut flora regulates lung disease and respiratory infection-related functions through the "gut-lung axis" (GLA). In fact, the interaction between gut flora and the lungs has been shown to play an important role in a variety of diseases, including asthma, chronic obstructive pulmonary disease (COPD), pneumonia and lung cancer. Several studies have shown that dysbiosis of the gut flora is strongly associated with the onset and progression of asthma. For example, some studies have shown that a decrease in Bifidobacterium spp. bacteria and an increase in Clostridium spp. bacteria in the gut are associated with asthma early in life. In addition, partial removal of gut flora with antibiotics can affect lung disease and allergic inflammation in mice. These findings suggest that dysregulation of gut flora may be an important factor in the development of asthma. In addition, interactions between gut flora and the lungs have been linked to diseases such as COPD, pneumonia, and lung cancer. For example, some studies have shown that the use of neomycin to remove sensitive gut bacteria leads to an increased susceptibility of the lungs to influenza virus infection. In addition, some studies have found that lipopolysaccharides from Streptococcus aureus have anti-inflammatory effects and significantly ameliorate COPD by acting as antagonists of the Toll-like receptor 4 signaling pathway [37,38,39]. The mucosal immune system (MIS) between the gut microbiota and the pulmonary axis has been identified as a key communication framework for mitigating interactions between the immune system and the microbiota, which includes three main interaction pathways, 1). Stimulation of immune system cells by the microbial community, which stimulates gut-lung interactions through the synthesis of specific secreted IgA and antimicrobial peptides (AMPs); 2). Stimulation of immune system cells through the production of certain microbial metabolites, such as tryptophan catabolic metabolites and short-chain fatty acids (SCFAs).3). Lung microbiota influences intestinal immunity, with Staphylococcus aureus or Pseudomonas aeruginosa pneumonia inducing intestinal sepsis and apoptotic processes, Specifically, the MIS involves a variety of cell types and molecular signaling pathways, including substances such as T cells, B cells, macrophages, dendritic cells, and antimicrobial peptides in specific pathways. These cells and molecules interact with each other through multiple pathways, thereby facilitating communication and coordination between the gut microbiota and the pulmonary axis. In particular, the gut microbiota stimulates gut-lung interactions by synthesizing specific secretory IgA and antimicrobial peptides (AMP), which in turn have a stimulatory effect on immune system cells. In addition, the gut microbiota can stimulate immune system cells by producing certain microbial metabolites, such as tryptophan catabolic metabolites and short-chain fatty acids (SCFAs). These metabolites affect the function and expression of immune cells and may influence the level of inflammation in the gut and lungs. Additionally, the lung microbiota can influence intestinal immunity. Several studies have shown that lung infections and inflammation are closely linked to alterations in the gut microbiota, e.g., Staphylococcus aureus or Pseudomonas aeruginosa pneumonia can induce intestinal sepsis and apoptotic processes. This interaction is achieved through multiple molecular signaling pathways, including cytokines, hormones, neuromodulation and metabolites in the gut and lungs [40,41,42,43,44].
Although the interactions between the gut flora and the lungs still need to be studied in depth, this field has provided new perspectives and strategies for our understanding of the pathogenesis of respiratory diseases and is expected to provide new ideas for the treatment and prevention of related diseases.
3.4. Gut Flora Regulates Renal Function via the Gut-Renal Axis
Imbalances in the gut microbiota have been shown to be associated with the development of hypertension and kidney disease. When the gut microbiota is imbalanced, it may lead to physiological changes such as immune dysregulation, metabolic disorders, and sympathetic activation, which in turn may affect the health status of the host. The concept that the gut is connected to the kidneys through the formation of a "gut-kidney axis" suggests that the state of the gut microbiota can influence kidney function and disease state. Studies have shown that alterations in the gut microbiota may contribute to pathophysiologic processes such as glomerular disease and chronic kidney disease. Therefore, maintaining a balanced gut microbiota is important for the prevention and treatment of hypertension and renal diseases [45,46]. The gut-kidney axis can be divided into three pathways: 1). Metabolism-dependent, the metabolism-dependent pathway is mainly mediated by metabolites produced by the gut microbiota that have the ability to modulate host physiological functions; 2). Immune pathways, components of the immune system that play a key role in the communication between the gut and the kidney; 3). Crosstalk between metabolism-dependent and immune pathways, specifically, metabolism-dependent pathways are those in which the gut microbiota regulates renal function and metabolic status through the production of metabolites. The gut microbiota can influence host metabolic processes through metabolites such as short-chain fatty acids (SCFA), medium-chain fatty acids, bile acids, and amino acid metabolites. These metabolites can reach the kidneys through the blood circulation, thus affecting kidney function and metabolic status. For example, SCFA can inhibit the inflammatory response and the production of fibrinogen activator, thereby reducing the risk of kidney disease. The immune pathway primarily involves interactions between the immune system and the inflammatory response between the gut and the kidney. Both the gut and the kidney are rich in immune cells and immune molecules, including T cells, B cells, macrophages, and dendritic cells. These immune cells and molecules can interact with each other through the circulation or the lymphatic system to influence the state of health of the gut and kidneys. For example, imbalances in the gut microbiota may lead to rupture of the intestinal mucosal barrier and aberrant activation of the immune system, which can lead to renal inflammation and fibrosis. Crosstalk between metabolic dependence and immune pathways is also an important mechanism, meaning that metabolites can influence the function of the immune system, which in turn can regulate the production and breakdown of metabolites. For example, the immune system can influence the production and utilization of metabolites by regulating the composition of the gut microbiota., [47,48,49,50,51,52,53,54]. In summary, the gut-kidney axis is a complex physiological pathway that interacts between the gut and kidney through metabolic-dependent and immune pathways, and there is synergy between these two pathways.
3.5. Dysbiosis of Intestinal Flora Affects Inflammatory Bowel Disease
A variety of microorganisms are involved in the development and progression of inflammatory bowel disease. Inflammatory bowel disease is an abnormal immune response disease against one's own normal intestinal flora, and the benefits of the intestinal microbiota to host physiology can be categorized into three groups, 1). Nutrition: human commensal bacteria, such as Bifidobacterium bifidum, synthesize and provide vitamins, such as vitamin K and water-soluble B vitamins, and intestinal bacteria also provide short-chain fatty acids (SCFA; C2-C6) through fermentation of resistant starch or indigestible carbohydrates (dietary fibers) [55,56]; 2). Immune development: the gut microbiota plays an important role in the development of the host immune system, which in turn shapes the structure and function of the gut microbiota [57,58]; 3). Host defense: the gut microbiota contributes to host defense against pathogens. Animals under sterile conditions are susceptible to infection by enteric pathogens. Abnormalities in the mucosal immune system may contribute to this susceptibility. Another mechanism for targeting pathogens is to prevent pathogen colonization by competitively colonizing the commensal microbiota, thereby disrupting the physical and nutritional ecological niche defects of the gastrointestinal tract. Inflammation, with the participation of the intestinal flora, initiates unstoppable episodes of alternating episodes and remissions of intestinal natural and acquired immune responses, leading to pathological changes such as damage to the intestinal mucosal barrier, long-lasting ulcers, and inflammatory hyperplasia [59,60]. (Figure 2)
4. The Role of Gut Flora in Tumors
Gut microbial homeostasis is closely related to human disease and health.The human gastrointestinal tract contains a rich microbial community with a concentration of about 100 trillion microorganisms[61], and recent studies have indeed demonstrated that gut microbiota play an important role in carcinogenesis. The following are some of the main mechanisms by which microbiota are involved in carcinogenesis:, 1. Influence on host cell proliferation and death: certain gut microbes can produce metabolites, such as short-chain fatty acids (SCFA), which can influence the proliferation and apoptosis of host intestinal cells. Excessive proliferation and apoptosis may lead to cell mutation and precancerous lesion formation; 2. Alteration of immune system activity: the gut microbiota can regulate the activity of the host immune system. Certain microbial strains can cause activation of the immune system and release inflammatory factors, leading to intestinal inflammation and injury. A prolonged inflammatory state may increase the risk of tumorigenesis; 3. Influence host metabolism: the microbiota is involved in regulating host metabolic processes. Certain microorganisms can break down specific dietary components and produce harmful metabolites, such as nitrites. These harmful metabolites may be associated with the development of intestinal cancers [62]. For example, dysbiosis of the gut microbiota, which impairs the intestinal barrier function, activates oncogenic MAPK / ERK signaling in the colonic epithelium to play a pro-tumorigenic role in colorectal cancer [63], In addition, disruption of the balance of the intestinal flora can regulate tryptophan metabolism to promote hepatocellular carcinomas [64], and there are differences in the composition of the gut microbes between patients with lung cancer and healthy populations, and metabolites of the aminocarbamoyl- tRNA biosynthesis pathway also seems to be closely associated with lung cancer progression [65]. This evidence suggests that the gut flora plays an important role in tumorigenesis and progression, and it is important to note that the role of microbiota in carcinogenesis is complex and diverse, and there are still many unanswered questions. Further studies can help us better understand the relationship between microbes and cancer and provide new strategies for cancer prevention and treatment. In the next section, we will focus on their roles and mechanisms in several common head and neck squamous cell carcinomas (HNSCC).
4.1. Oral Cancer (OC) and Intestinal Flora
Oral cancer is a common malignant tumor of the head and neck with a high recurrence rate and poor prognosis, with a 5-year survival rate of <50% [66,67].In recent years, more and more studies have shown that there is a strong association between the oral microbiota and oral cancer. In addition, the oral and intestinal microbiomes have been recognized as potential biomarkers of cancer and may influence the response to anticancer therapies. Several studies have found that disturbances in the oral and gut microbiota are associated with the development and progression of oral cancer. For example, patients with periodontitis have an increased susceptibility to non-infectious chronic inflammatory diseases driven by multiple microbe-induced immune mechanisms. In addition, radiotherapy may lead to shifts in the composition of the flora in oropharyngeal swabs from patients with oropharyngeal squamous cell carcinoma, which may have prognostic and therapeutic implications [68,69]. The oral and intestinal microbiome has emerged as a potential biomarker of cancer, and the microbiome residing in the oral gastrointestinal tract has been implicated in carcinogenesis and modulating the response to anticancer therapies in many tumor types, as shown in a prospective study. Radiotherapy leads to a shift toward gut-like microbiome composition in oropharyngeal swabs of HPV+ oropharyngeal squamous cell carcinoma, a finding that may have prognostic and therapeutic implications [70]. The oral microbiome is connected to the lung and gut microbiomes in a complex manner and is able to regulate inflammatory signals through the coordinated crosstalk that occurs between different species of resident bacteria that promote TLR-2-mediated T-cell activation through their most representative strains, maintaining the oral microenvironment on leading to tumorigenesis [71]. A recent study examined the association between the community composition and structure of the colonic adherent microbiota and oral health, using the number of tooth loss and gingival disease as markers of oral health in the population, and concluded that oral health is associated with alterations in the community composition and structure of adherent enteric bacteria in the colon [72]. And we have already mentioned the close relationship between gut flora and several cancers, based on this, it is interesting to imply that disturbances in gut flora also influence the development and progression of oral cancer, just more experimental validation is needed to prove our conjecture.
4.2. Nasopharyngeal Carcinoma (NPC) and Intestinal flora
Nasopharyngeal Carcinoma (NPC, NPC) is one of the most common malignant tumors in East and Southeast Asia. Since early symptoms of nasopharyngeal carcinoma are not obvious and difficult to be detected, most of the patients are already in advanced stages when they arrive at the hospital. Advanced NPC is difficult to treat and has a poor prognosis, with nearly 80% of NPC patients being diagnosed at an advanced stage [73], and an imbalance of intestinal flora is thought to be associated with multiple organ malignancies. The current study suggests that disruption of the intestinal flora is associated with nasopharyngeal cancer, which was found to be associated with an imbalance in the structure of the intestinal flora, and patients in the familial nasopharyngeal cancer group and sporadic nasopharyngeal cancer group showed significant changes in the structure of the intestinal flora compared with healthy controls. In the familial nasopharyngeal cancer group, there was a significant increase in Trichoderma, Citrobacter spp, Verotrichum spp and Prevotella spp, and a significant decrease in Mucor and Roseola spp, while in the sporadic nasopharyngeal cancer group, there was a significant increase in Clostridium rayonii, Chromobacterium finely, B. microtus, and Bifidobacterium klebsiensis spp and a significant decrease in Bifidobacterium adolescentum. And a study found that 5-HT was overexpressed in human NPC samples, C. ramosum is an anaerobic bacterium found mainly in the intestinal tract, and its metabolites can stimulate the secretion of 5-HT from ECs, which can promote plasma levels of 5-HT potentially facilitating the progression of nasopharyngeal cancer. This suggests that elevated 5-HT is associated with a significant increase in opportunistic pathogens and a significant decrease in probiotics [74,75,76,77]. The results of a randomized clinical trial showed that probiotics combined with radiotherapy for the treatment of nasopharyngeal carcinoma patients undergoing concurrent radiotherapy significantly enhanced host immunity by altering the intestinal flora [78], and that interactions between the host immune system and the microbiome may underlie the role of the microbiome in cancer therapy [79]. As the relationship between the two continues to be studied, alterations in gut flora may not only serve as an early noninvasive diagnosis of nasopharyngeal carcinoma, but may also play a special role in future nasopharyngeal carcinoma treatment in order to change the prognosis of the patients and achieve a more satisfactory outcome.
4.3. Thyroid Cancer (TC) and Intestinal Flora
Thyroid cancer (TC) is the most common endocrine malignancy and its incidence is still increasing worldwide [80]. Emerging studies have provided a preliminary understanding of the thyroid-gut axis, suggesting that intestinal flora and their metabolites may act directly or indirectly on the thyroid by influencing intestinal micronutrient uptake, iodothyronine conversion and storage, and immunomodulation, providing new insights into the pathogenesis of thyroid disease and clinical management strategies [81]. The thyroid gland requires iodine as a supplemental micronutrient for the synthesis of thyroid hormones, and trillions of microorganisms in the gut play a key role in regulating iodine metabolism, and studies have shown that a reduced gut microbiota may affect radioactive iodine uptake and thyroid function[82]. Thyroid hormone homeostasis is closely linked to the effect of gut microbiota on iodothyronine metabolism, and the dose of levothyroxine required to maintain stable TSH levels is related to the microbiota[83,84]. The genus of altered gut microbiota in the TC group was found to be significantly associated with lipid metabolites and lipid levels, and serum metabolomics analyses of patients with distant metastases of thyroid cancer also showed that the interaction between diet and gut microbiota may play an important role in tumor aggressiveness, with the microbiota in the thyroid cancer group having a higher abundance and diversity than that in the healthy control group and altered in relation to serum lipid metabolites [85,86,87]. The correlation between thyroid cancer, thyroid nodules and microbiota was investigated by high-throughput sequencing comparing the microbial structural characteristics of 36 patients with thyroid cancer, 72 patients with thyroid nodules, and 73 healthy controls, which showed that thyroid cancer and thyroid nodules were strongly associated with altered microbiota [88].SCFAs are thought to have a regulatory role in the immune microenvironment, and these studies have found that reduced abundance of SCFAs-producing bacteria such as Spirochaetaceae and Aeromonas butyricola in thyroid cancer patients [89,90]. Cholesterol can increase the aggressiveness of TC, with significant changes in the abundance and composition of gut microbiota and metabolites in the TC group, and these genera have been significantly associated with disturbances in lipid metabolism in TC patients [87,91]. Further understanding of the characterization of gut microecology in TC will help to identify risk factors affecting the development and progression of TC.
4.4. Glioma and Intestinal Flora
Maturation and development of the human central nervous system (CNS) is regulated by intrinsic and extrinsic factors. Studies, mainly from germ-free or broad-spectrum antibiotic-treated animals, have shown that specific microbiota can influence CNS physiology and neurochemistry [92], that the gut-brain axis is used to define the relationship between the microbiota and its interactions with the brain, leading to changes in the state of the CNS, and that ecological dysregulation of microbial species may induce atypical immune signaling, imbalance of homeostasis in the host, and even CNS system disease progression [93,94]. Recent studies have shown that the microbiome affects microglia properties and functions [95]. Glioblastoma is one of the most malignant tumors with a particularly high mortality rate[96]. Therefore, new therapeutic agents and approaches are needed to combat this deadly disease. Recent studies have demonstrated the potential role of the microbiome in immuno-oncology[97], and in the analysis of fecal samples from patients with metastatic melanoma, Bifidobacterium longum, Corinobacterium perfringens, and Enterococcus faecalis were found to be in increased abundance in subjects responding to PD-1 inhibition with therapeutic antibodies, suggesting that certain microbial taxa in the intestinal tract may provide supportive roles to augment the effects of PD-1 blockade[98]. Glioma development leads to dysregulation of gut ecology in a mouse model, with increased abundance of the warty microbial phylum and the genus Akkermansia following glioma growth, and fecal samples were collected from healthy controls before and after radiochemotherapy, as well as from patients with gliomas at the time of diagnosis. Compared to healthy controls, mice and glioma patients showed significant differences in beta diversity, the Fusobacterium thicketi / Fusobacterium anisopliae (F / B) ratio, and an increase in the warty microbial phylum and Ackermansia spp. [99]. A meta-analysis revealed an association between human cytomegalovirus (HCMV) and glioma the mechanism of which may be related to the stress response of glioma cells triggered by HCMV through the down-regulation of specific miRNAs, as well as polyomaviral and adenoviral infections have been associated with the development of gliomas[100,101,102]. Typical pathogens Enterobacteriaceae are overrepresented in meningiomas and can also inhibit SCFA-producing bacteria and lead to ecological dysregulation of the immune and intestinal environments, and the genus Akkermansia can induce inflammatory responses, neurotoxicity, and blood-brain barrier disruption in the microenvironment of gliomas through its ability to degrade the intestinal mucosal layer Therefore, ecological dysregulation of the intestinal microbiota is more severe in patients with neurogliomas,. While both Lactobacillus spp. and Agathobacterium spp. may have antitumor activity through butyric acid production, which prevents pathogen invasion, slows tumor progression, and modulates immune responses in the CNS, in addition, Bifidobacterium spp. belongs to the probiotic family, which contributes to the dynamic balance of immunity, neurohormones, and metabolism [103]. Early in life, the gut flora is balanced and healthy, but with various treatments or conditions, the gut flora is compromised, which promotes glioma progression. As new evidence emerges, gut flora can even be used as a biomarker to predict prognosis and response to treatment in glioma patients, however more research is needed to determine the detailed function of gut microbes on glioma treatment.
5. Gut Flora in Head and Neck Tumors
Radiation and chemotherapy for cancer may lead to decreased immunity in cancer patients, and research evidence suggests that the gut microbiota can modulate anticancer immune responses and attenuate toxic side effects associated with cancer treatment [104,105,106,107]. The main treatment for nasopharyngeal carcinoma is radiation therapy, but radiation therapy can cause a variety of side effects and can also alter the composition of the intestinal flora. Macrosequence sequencing analysis of intestinal flora data revealed that raw wheat and turmeric dispersions in combination with irradiation can enhance the effectiveness of tumor therapy. Radiation therapy reduced the total number and diversity of intestinal flora in nude mice and altered the structure of the flora. In contrast, raw wheat and turmeric san both protected the diversity of the flora and partially restored the imbalance of the flora caused by irradiation [108]. A randomized double-blind trial found that probiotic combinations significantly enhanced patients' immune responses and reduced the severity of oral mucositis (OM) in nasopharyngeal carcinoma patients undergoing simultaneous radiotherapy by altering the gut microbiota [109], while a phase II randomized clinical trial also concluded that, in a rat model, probiotic mixtures ameliorated the severity of OM, reduced inflammatory responses, induced apoptosis and intestinal permeability, and normalize the structure of the gut microbiota. In conclusion, a modified probiotic cocktail significantly reduced the severity of OM by enhancing the immune response and altering the structure of the gut microbiota in nasopharyngeal cancer patients [110]. A recent study found that in 131I-treated postoperative TC patients, the most notable differences were a decrease in Thickettsia and an increase in Bifidobacterium anisoplia, resulting in a significant decrease in the ratio of Thickettsia to Bifidobacterium anisoplia after treatment, and that G-Dorea and g-Bifidobacterium may be potential targets for clinical interventions to improve the 131I in TC patients in the postoperative period [111]. Another randomized clinical trial also found that probiotics significantly restored gut and oral microbial diversity and reduced the incidence of complications in patients after thyroid cancer resection [112]. Neuroglioma is a life-threatening malignant tumor, and traditional radiotherapy is not very effective. More and more studies have shown that microorganisms and their derivatives can be used as cancer therapies, which mainly include three kinds: 1. microbial derivatives used in glioma therapy, bacteria can produce active substances such as bacteriocins and antimicrobial peptides, which are used in the treatment of gliomas through different mechanisms [113,114,115]; 2. phages can target gliomas for drug delivery, and phages have a strong affinity for the VAV3 protein GICP (glioma-initiating cell peptide), which has a strong affinity for VAV3 protein, and phages can be used to precisely deliver drugs to gliomas [116]; 3. Microbial targeting of gliomas, where many bacteria can cross the blood-brain barrier to enter the center through a unique mechanism, which lays the groundwork for bacterial entry into targeted gliomas, and where viruses have their own mechanisms or use immune cells as carriers, which, when administered systemically, can cross the blood-brain barrier and enter gliomas. administered, can cross the blood-brain barrier to reach the glioma site [117,118]. The metabolite of gut bacterial flora, CK (a metabolite of ginsenoside), reduces the migration and invasive capacity of glioma cells in vitro by inhibiting downstream SDF-1 and CXCR4 signaling [119,120], and alterations in the gut microbiota caused by antibiotic treatment induced early damage to NK cells and induced changes in microglia phenotype, suggesting that altered gut-immune-brain communication may contribute to the induction of a tumor-tolerant CNS microenvironment, which facilitates tumor development [121]. Recent studies have demonstrated that the gut microbiota can be involved in glioma development and individualized efficacy through immunomodulation, and have identified gut bacteria as predictive biomarkers as well as clinical therapeutic targets [122].
6. Summary and Prospects
The intestinal flora currently plays a role in the development of diseases mainly by the pathways of intestinal-brain axis, intestinal-lung axis, intestinal-hepatic axis and intestinal-renal axis, which mainly regulate the mechanisms of neurological, endocrine and immune aspects, and how it affects the occurrence and progression of tumors is an interesting question.
Cancer is very harmful to human health, and there are limited ways to treat cancer. At present, the clinical treatment for head and neck cancer is relatively single, synchronous radiotherapy combined with immunotherapy or targeted therapy is a common pathway for clinical treatment, but the toxic side effects are large, the study found that the intestinal flora can regulate tumor immunity and may be involved in the progression of the disease, so there is a huge potential in the desensitization of tumors. The microbiome may play an important role in the tumor microenvironment, and if the intestinal flora can be used to stimulate stronger immune function in the body, this could be another major advance in tumor desensitization therapy. With the further understanding of the relationship between intestinal bacteria and immunotherapy, radiotherapy, etc., we have learned that some intestinal genera can also reduce the serious complications brought by radiotherapy, and how to regulate the intestinal flora seems to have become a new topic and direction to study the future anti-tumor therapy.
It has the potential to lead a new era in the treatment of cancer and other diseases, but we should not exaggerate the importance of intestinal flora in cancer treatment. At present, despite these new discoveries about intestinal flora, there are still very few studies on its role in diseases, especially on the role played by intestinal flora in head and neck tumors, which is still limited, and more experiments and clinical trials are needed to explore its practical value and clinical applications to support our conclusions.
Author Contributions
All authors actively participated to the review design, preparation, and organization. They also extensively discussed about the topics presented, giving a positive criticism to the work.,All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China (No. 82060556);Science and Technology Fund Project of Guizhou Provincial Health Commission [gzwkj2023-118]
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021, 19, 55-71. [CrossRef]
- Fujisaka S, Avila-Pacheco J, Soto M, et al. Diet, Genetics, and the Gut Microbiome Drive Dynamic Changes in Plasma Metabolites. Cell Rep. 2018, 22, 3072-3086. [CrossRef]
- Brown RL, Larkinson M, Clarke TB. Immunological design of commensal communities to treat intestinal infection and inflammation. PLoS Pathog. 2021, 17, e1009191. [CrossRef] [PubMed]
- Caballero S, Pamer EG. Microbiota-mediated inflammation and antimicrobial defense in the intestine. Annu Rev Immunol. 2015, 33: 227-56. [CrossRef]
- Fessler J, Matson V, Gajewski TF. Exploring the emerging role of the microbiome in cancer immunotherapy. J Immunother Cancer. 2019, 7, 108. [CrossRef] [PubMed]
- Hatakeyama M. Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer. Proc Jpn Acad Ser B Phys Biol Sci. 2017, 93, 196-219. [CrossRef]
- Goodwin AC, Destefano Shields CE, Wu S, et al. Polyamine catabolism contributes to enterotoxigenic Bacteroides fragilis-induced colon tumorigenesis. Proc Natl Acad Sci U S A. 2011, 108, 15354-9. [CrossRef]
- Bergounioux J, Elisee R, Prunier AL, et al. Calpain activation by the Shigella flexneri effector VirA regulates key steps in the formation and life of the bacterium's epithelial niche. Cell Host Microbe. 2012, 11, 240-52. [CrossRef]
- Chaturvedi R, Asim M, Romero-Gallo J, et al. Spermine oxidase mediates the gastric cancer risk associated with Helicobacter pylori CagA. Gastroenterology. 2011, 141, 1696-708.e1-2. [CrossRef]
- Kuugbee ED, Shang X, Gamallat Y, et al. Structural Change in Microbiota by a Probiotic Cocktail Enhances the Gut Barrier and Reduces Cancer via TLR2 Signaling in a Rat Model of Colon Cancer. Dig Dis Sci. 2016, 61, 2908-2920. [CrossRef]
- Johnson DB, Frampton GM, Rioth MJ, et al. Targeted Next Generation Sequencing Identifies Markers of Response to PD-1 Blockade. Cancer Immunol Res. 2016, 4, 959-967. [CrossRef]
- Sivan A, Corrales L, Hubert N, et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015, 350, 1084-9. [CrossRef] [PubMed]
- Porter, J.R. Antony van Leeuwenhoek: tercentenary of his discovery of bacteria. Bacteriol Rev. 1976, 40, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Majem LS, Marcos AMR, Guardia JAM, Batrina JA, Marcos A, Anta RMO. Alimentos Funcionales. Probióticos. 2002.
- Bourdelles F, Avril JL, Ghnassia JC. [Quantitative study of the faecal flora of breast- or bottle-fed neonates (author's transl)]. Archives franaises de pédiatrie. 1981, 38, 35-39.
- Ursell LK, Haiser HJ, Van Treuren W, et al. The intestinal metabolome: an intersection between microbiota and host. Gastroenterology. 2014, 146, 1470-6. [CrossRef] [PubMed]
- Brown AL, Tucker B, Baker LR, Raine AE. Seizures related to blood transfusion and erythropoietin treatment in patients undergoing dialysis. Bmj British Medical Journal. 1989, 299, 1258-1259. [CrossRef]
- Wilson KH, Blitchington RB. Human colonic biota studied by ribosomal DNA sequence analysis. Appl Environ Microbiol. 1996, 62, 2273-2278. [CrossRef]
- Lederberg, J. Infectious History. SCIENCE. 2000, 288, 287–293. [Google Scholar] [CrossRef]
- Breitbart M, Hewson I, Felts B, et al. Bacteriophages, transposons, and plasmids: metagenomic analyses of an uncultured viral community from human feces. JOURNAL OF BACTERIOLOGY. 2003, 185, 6220-6223. [CrossRef]
- Klaassens ES, De Vos WM, Vaughan EE. Metaproteomics Approach To Study the Functionality of the Microbiota in the Human Infant Gastrointestinal Tract. Applied & Environmental Microbiology. 2007, 73, 1388-92. [CrossRef]
- Heijtz RD, Wang S, Anuar F, et al. Normal gut microbiota modulates brain development and behavior. Proceedings of the National Academy of Sciences. 2011, 108, 3047-3052. [CrossRef]
- Sebastián Domingo JJ, Sánchez Sánchez C. From the intestinal flora to the microbiome. Rev Esp Enferm Dig. 2018, 110, 51-56. [CrossRef]
- Cryan JF, O'Riordan KJ, Cowan C, Sandhu KV, Dinan TG. The Microbiota-Gut-Brain Axis. PHYSIOLOGICAL REVIEWS. 2019, 99, 1877-2013.
- Fang P, Kazmi SA, Jameson KG, Hsiao EY. The Microbiome as a Modifier of Neurodegenerative Disease Risk. Cell Host & Microbe. 2020. [CrossRef]
- Morais LH, Henry IV, Mazmanian SK. The gut microbiota–brain axis in behaviour and brain disorders. NATURE REVIEWS MICROBIOLOGY. [CrossRef]
- Fluckiger A, Daillère R, Sassi M, Sixt BS, Zitvogel L. Cross-reactivity between tumor MHC class I-restricted antigens and an enterococcal bacteriophage. SCIENCE. 2020, 369, 936-942. [CrossRef]
- Abbott, A. Are infections seeding some cases of Alzheimer's disease. NATURE. 2020, 587, 22–25. [Google Scholar] [CrossRef]
- Farhangi MA, Vajdi M. Gut microbiota–associated trimethylamine N ﹐xide and increased cardiometabolic risk in adults: a systematic review and dose-response meta-analysis. NUTRITION REVIEWS. 2020. [CrossRef]
- Wang T, Rong X, Zhao C. Circadian Rhythms Coordinated With Gut Microbiota Partially Account for Individual Differences in Hepatitis B-Related Cirrhosis. Front Cell Infect Microbiol. 2022, 12: 936815. [CrossRef]
- Oikonomou T, Papatheodoridis GV, Samarkos M, Goulis I, Cholongitas E. Clinical impact of microbiome in patients with decompensated cirrhosis. WORLD JOURNAL OF GASTROENTEROLOGY. 2018. [CrossRef] [PubMed]
- Bauer KC, Littlejohn PT, Ayala V, Creus-Cuadros A, Finlay BB. Nonalcoholic Fatty Liver Disease and the Gut-Liver Axis: Exploring an Undernutrition Perspective. Gastroenterology. 2022, 162, 1858-1875.e2. [CrossRef] [PubMed]
- Lifeng L, Yunhai Y, Yunyun L, Junwei Q, Xuejing L, Gengyun Z. A Comprehensive Genome Survey Provides Novel Insights into Bile Salt Hydrolase (BSH) in Lactobacillaceae. MOLECULES. 2018, 23, 1157.
- Arab JP, Martin-Mateos RM, Shah VH. Gut–liver axis, cirrhosis and portal hypertension: the chicken and the egg. Hepatology International. 2017. [CrossRef]
- Lucie, Etienne-Mesmin, Andrew, et al. Microbiota-liver axis in hepatic disease. Hepatology Official Journal of the American Association for the Study of Liver Diseases. 2014. [CrossRef]
- John II, Behrendt CL, Ruhn KA, et al. The microbiota coordinates diurnal rhythms in innate immunity with the circadian clock. CELL. 2021, (184-16). [CrossRef]
- Mjösberg J, Rao A. Lung inflammation originating in the gut. Science. 2018, 359, 36-37. [CrossRef]
- Lai HC, Lin TL, Chen TW, et al. Gut microbiota modulates COPD pathogenesis: role of anti-inflammatory Parabacteroides goldsteinii lipopolysaccharide. Gut. 2022, 71, 309-321. [CrossRef]
- Yun KY, Zuo T, Lui CY, Zhang F, Ng SC. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. GUT. 70, 698-706.
- Gill N, Wlodarska M, Finlay BB. The future of mucosal immunology: studying an integrated system-wide organ. Nat Immunol. 2010, 11, 558-60.
- Anand S, Mande SS. Diet, Microbiota and Gut-Lung Connection. Front Microbiol. 2018, 9: 2147. [CrossRef]
- McGhee JR, Fujihashi K. Inside the mucosal immune system. PLoS Biol. 2012, 10, e1001397. [CrossRef] [PubMed]
- Dang AT, Marsland BJ. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol. 2019, 12, 843-850. [CrossRef] [PubMed]
- Perrone EE, Jung E, Breed E, et al. Mechanisms of methicillin-resistant Staphylococcus aureus pneumonia-induced intestinal epithelial apoptosis. Shock. 2012, 38, 68-75. [CrossRef]
- Yang T, Richards EM, Pepine CJ, Raizada MK. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat Rev Nephrol. 2018, 14, 442-456. [CrossRef] [PubMed]
- Wang X, Yang S, Li S, et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut. 2020, 69, 2131-2142. [CrossRef] [PubMed]
- Sirich TL, Plummer NS, Gardner CD, Hostetter TH, Meyer TW. Effect of Increasing Dietary Fiber on Plasma Levels of Colon-Derived Solutes in Hemodialysis Patients. Clinical Journal of the American Society of Nephrology. 2014, 9(9). [CrossRef]
- Plummer NS, Holmes S, Meyer TW, et al. Colonic contribution to uremic solutes. 2013. [CrossRef] [PubMed]
- Yan J, Herzog JW, Tsang K, et al. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc Natl Acad Sci U S A. 2016, 113, E7554-E7563. [CrossRef] [PubMed]
- Hahm E, Wei C, Fernandez I, et al. Bone marrow-derived immature myeloid cells are a main source of circulating suPAR contributing to proteinuric kidney disease. Nat Med. 2017, 23, 100-106. [CrossRef] [PubMed]
- Schumacher M, Wanner C, Beyersmann J. Soluble Urokinase Receptor and Chronic Kidney Disease. NEW ENGLAND JOURNAL OF MEDICINE. 2016, 374, 890-891.
- Sirich TL, Aronov PA, Plummer NS, Hostetter TH, Meyer TW. Numerous protein-bound solutes are cleared by the kidney with high efficiency. Kidney Int. 2013, 84, 585-90. [CrossRef]
- Ito S, Yoshida M. Protein-bound uremic toxins: new culprits of cardiovascular events in chronic kidney disease patients. Toxins (Basel). 2014, 6, 665-78. [CrossRef] [PubMed]
- Koppe L, Pillon NJ, Vella RE, et al. p-Cresyl sulfate promotes insulin resistance associated with CKD. J Am Soc Nephrol. 2013, 24, 88-99. [CrossRef] [PubMed]
- Marchesi JR, Adams DH, Fava F, Hermes G, Hart A. The gut microbiota and host health. 2015. [CrossRef] [PubMed]
- LeBlanc JG, Laiño JE, del Valle MJ, et al. B-group vitamin production by lactic acid bacteria--current knowledge and potential applications. J Appl Microbiol. 2011, 111, 1297-309. [CrossRef] [PubMed]
- Gomaa EZ. Human gut microbiota/microbiome in health and diseases: a review. Antonie Van Leeuwenhoek. 2020, 113, 2019-2040. [CrossRef] [PubMed]
- Kamada N, Núñez G. Regulation of the immune system by the resident intestinal bacteria. Gastroenterology. 2014, 146, 1477-88. [CrossRef]
- Sekirov I, Russell SL, Antunes L, Finlay BB. Gut microbiota in health and disease. PHYSIOLOGICAL REVIEWS. 2010, 90, 859-904. [CrossRef]
- Kamada N, Kim YG, Sham HP, et al. Regulated Virulence Controls the Ability of a Pathogen to Compete with the Gut Microbiota. SCIENCE. 2012, 336, 1325-9. [CrossRef] [PubMed]
- Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. NATURE. 2006, 444, 1022-1023. [CrossRef]
- Chen Y, Zhou J, Wang L. Role and Mechanism of Gut Microbiota in Human Disease. Front Cell Infect Microbiol. 2021, 11: 625913. [CrossRef]
- Bai X, Wei H, Liu W, et alBai X, Wei H, Liu W, et al. Cigarette smoke promotes colorectal cancer through modulation of gut microbiota and related metabolites. Gut. 2022, 71, 2439-2450. [CrossRef]
- Chen W, Wen L, Bao Y, et al. Gut flora disequilibrium promotes the initiation of liver cancer by modulating tryptophan metabolism and up-regulating SREBP2. Proc Natl Acad Sci U S A. 2022, 119, e2203894119. [CrossRef]
- Chen S, Gui R, Zhou XH, et al. Combined Microbiome and Metabolome Analysis Reveals a Novel Interplay Between Intestinal Flora and Serum Metabolites in Lung Cancer. Front Cell Infect Microbiol. 2022, 12: 885093. [CrossRef]
- Rodríguez-Molinero J, Migueláñez-Medrán B, Puente-Gutiérrez C, et al. Association between Oral Cancer and Diet: An Update. Nutrients. 2021, 13(4). [CrossRef]
- Sklenicka S, Gardiner S, Dierks EJ, Potter BE, Bell RB. Survival analysis and risk factors for recurrence in oral squamous cell carcinoma: does surgical salvage affect outcome. J Oral Maxillofac Surg. 2010, 68, 1270-5. [CrossRef] [PubMed]
- Perera M, Al-Hebshi NN, Speicher DJ, Perera I, Johnson NW. Emerging role of bacteria in oral carcinogenesis: a review with special reference to perio-pathogenic bacteria. J Oral Microbiol. 2016, 8: 32762. [CrossRef]
- Hajishengallis G, Chavakis T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat Rev Immunol. 2021, 21, 426-440. [CrossRef] [PubMed]
- Oliva M, Schneeberger P, Rey V, et al. Transitions in oral and gut microbiome of HPV+ oropharyngeal squamous cell carcinoma following definitive chemoradiotherapy (ROMA LA-OPSCC study). Br J Cancer. 2021, 124, 1543-1551. [CrossRef] [PubMed]
- Nocini R, Muzio LL, Gibellini D, et al. Oral microbiota in oropharyngeal cancers: Friend or foe. Front Oncol. 2022, 12: 948068. [CrossRef]
- Xu AA, Hoffman K, Gurwara S, et al. Oral Health and the Altered Colonic Mucosa-Associated Gut Microbiota. Dig Dis Sci. 2021, 66, 2981-2991. [CrossRef] [PubMed]
- He YQ, Wang TM, Ji M, et al. A polygenic risk score for nasopharyngeal carcinoma shows potential for risk stratification and personalized screening. Nat Commun. 2022, 13, 1966. [CrossRef] [PubMed]
- Peters M, Meijer C, Fehrmann R, et al. Serotonin and Dopamine Receptor Expression in Solid Tumours Including Rare Cancers. Pathol Oncol Res. 2020, 26, 1539-1547. [CrossRef] [PubMed]
- Senda S, Fujiyama Y, Ushijima T, et al. Clostridium ramosum, an IgA protease-producing species and its ecology in the human intestinal tract. Microbiol Immunol. 1985, 29, 1019-28.
- Mandić AD, Woting A, Jaenicke T, et al. Clostridium ramosum regulates enterochromaffin cell development and serotonin release. Sci Rep. 2019, 9, 1177. [CrossRef]
- Jiang H, Li J, Zhang B, et al. Intestinal Flora Disruption and Novel Biomarkers Associated With Nasopharyngeal Carcinoma. Front Oncol. 2019, 9: 1346. [CrossRef]
- Guo H, Chou WC, Lai Y, et al. Multi-omics analyses of radiation survivors identify radioprotective microbes and metabolites. Science. 2020, 370(6516). [CrossRef]
- Yu ZK, Xie RL, You R, et al. The role of the bacterial microbiome in the treatment of cancer. BMC Cancer. 2021, 21, 934. [CrossRef]
- Gu Y, Yu Y, Ai L, et al. Association of the ATM gene polymorphisms with papillary thyroid cancer. Endocrine. 2014, 45, 454-61. [CrossRef] [PubMed]
- Lerner A, Jeremias P, Matthias T. Gut-thyroid axis and celiac disease. Endocr Connect. 2017, 6, R52-R58. [CrossRef] [PubMed]
- Vought RL, Brown FA, Sibinovic KH, McDaniel EG. Effect of changing intestinal bacterial flora on thyroid function in the rat. Horm Metab Res. 1972, 4, 43-7. [CrossRef] [PubMed]
- Yao Z, Zhao M, Gong Y, et al. Relation of Gut Microbes and L-Thyroxine Through Altered Thyroxine Metabolism in Subclinical Hypothyroidism Subjects. Front Cell Infect Microbiol. 2020, 10: 495. [CrossRef]
- DiStefano JJ 3rd, de Luze A, Nguyen TT. Binding and degradation of 3,5,3'-triiodothyronine and thyroxine by rat intestinal bacteria. Am J Physiol. 1993, 264(6 Pt 1): E966-72. [CrossRef]
- Shen CT, Zhang Y, Liu YM, et al. A distinct serum metabolic signature of distant metastatic papillary thyroid carcinoma. Clin Endocrinol (Oxf). 2017, 87, 844-852. [CrossRef] [PubMed]
- Feng J, Zhao F, Sun J, et al. Alterations in the gut microbiota and metabolite profiles of thyroid carcinoma patients. Int J Cancer. 2019, 144, 2728-2745. [CrossRef]
- Lu G, Yu X, Jiang W, et al. Alterations of Gut Microbiome and Metabolite Profiles Associated With Anabatic Lipid Dysmetabolism in Thyroid Cancer. Front Endocrinol (Lausanne). 2022, 13: 893164. [CrossRef]
- Yano Y, Matsui T, Uno H, Hirai F, Futami K, Iwashita A. Risks and clinical features of colorectal cancer complicating Crohn's disease in Japanese patients. J Gastroenterol Hepatol. 2008, 23, 1683-8. [CrossRef] [PubMed]
- Eckburg PB, Bik EM, Bernstein CN, et al. Diversity of the human intestinal microbial flora. Science. 2005, 308, 1635-8. [CrossRef] [PubMed]
- Aghajani MJ, Cooper A, McGuire H, et al. Pembrolizumab for anaplastic thyroid cancer: a case study. Cancer Immunol Immunother. 2019, 68, 1921-1934. [CrossRef] [PubMed]
- Asghari A, Umetani M. Obesity and Cancer: 27-Hydroxycholesterol, the Missing Link. Int J Mol Sci. 2020, 21(14). [CrossRef]
- Smith, PA. The tantalizing links between gut microbes and the brain. Nature. 2015, 526, 312–4. [Google Scholar] [CrossRef]
- Sharon G, Sampson TR, Geschwind DH, Mazmanian SK. The Central Nervous System and the Gut Microbiome. Cell. 2016, 167, 915-932. [CrossRef]
- Cussotto S, Sandhu KV, Dinan TG, Cryan JF. The neuroendocrinology of the microbiota-gut-brain axis: a behavioural perspective. FRONTIERS IN NEUROENDOCRINOLOGY. 2018 : S0091302218300396. [CrossRef]
- Erny D, Hrabě de Angelis AL, Jaitin D, et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci. 2015, 18, 965-77. [CrossRef]
- Davis, ME. Glioblastoma: Overview of Disease and Treatment. Clin J Oncol Nurs. 2016, 20(5 Suppl): S2-8. [CrossRef]
- Routy B, Gopalakrishnan V, Daillère R, Zitvogel L, Wargo JA, Kroemer G. The gut microbiota influences anticancer immunosurveillance and general health. Nat Rev Clin Oncol. 2018, 15, 382-396. [CrossRef] [PubMed]
- Matson V, Fessler J, Bao R, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. 2018, 359, 104-108. [CrossRef] [PubMed]
- Patrizz A, Dono A, Zorofchian S, et al. Glioma and temozolomide induced alterations in gut microbiome. Sci Rep. 2020, 10, 21002. [CrossRef] [PubMed]
- Farias K, Moreli ML, Floriano VG, da Costa VG. Evidence based on a meta-analysis of human cytomegalovirus infection in glioma. Arch Virol. 2019, 164, 1249-1257. [CrossRef] [PubMed]
- Hu M, Yu B, Zhang B, et al. Human Cytomegalovirus Infection Activates Glioma Activating Transcription Factor 5 via microRNA in a Stress-Induced Manner. ACS Chem Neurosci. 2021, 12, 3947-3956. [CrossRef] [PubMed]
- Egan KM, Kim Y, Bender N, et al. Prospective investigation of polyomavirus infection and the risk of adult glioma. Sci Rep. 2021, 11, 9642. [CrossRef]
- Jiang H, Zeng W, Zhang X, Pei Y, Zhang H, Li Y. The role of gut microbiota in patients with benign and malignant brain tumors: a pilot study. Bioengineered. 2022, 13, 7847-7859. [CrossRef] [PubMed]
- Roy S, Trinchieri G. Microbiota: a key orchestrator of cancer therapy. Nat Rev Cancer. 2017, 17, 271-285. [CrossRef] [PubMed]
- Lakritz JR, Poutahidis T, Levkovich T, et al. Beneficial bacteria stimulate host immune cells to counteract dietary and genetic predisposition to mammary cancer in mice. Int J Cancer. 2014, 135, 529-40. [CrossRef]
- Vétizou M, Pitt JM, Daillère R, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015, 350, 1079-84. [CrossRef] [PubMed]
- Daillère R, Vétizou M, Waldschmitt N, et al. Enterococcus hirae and Barnesiella intestinihominis Facilitate Cyclophosphamide-Induced Therapeutic Immunomodulatory Effects. Immunity. 2016, 45, 931-943. [CrossRef] [PubMed]
- Yang JB, Zhu DQ, Shao M, et al. Effects of Shengmai Jianghuang San on intestinal flora in nude mice with radio resistant cells of nasopharyngeal carcinoma. Zhongguo Zhong Yao Za Zhi. 2019, 44, 553-558. [CrossRef]
- Jiang C, Wang H, Xia C, et al. A randomized, double-blind, placebo-controlled trial of probiotics to reduce the severity of oral mucositis induced by chemoradiotherapy for patients with nasopharyngeal carcinoma. Cancer. 2019, 125, 1081-1090. [CrossRef] [PubMed]
- Xia C, Jiang C, Li W, et al. A Phase II Randomized Clinical Trial and Mechanistic Studies Using Improved Probiotics to Prevent Oral Mucositis Induced by Concurrent Radiotherapy and Chemotherapy in Nasopharyngeal Carcinoma. Front Immunol. 2021, 12: 618150. [CrossRef]
- Zheng L, Zhang L, Tang L, et al. Gut microbiota is associated with response to (131)I therapy in patients with papillary thyroid carcinoma. Eur J Nucl Med Mol Imaging. 2023, 50, 1453-1465. [CrossRef] [PubMed]
- Lin B, Zhao F, Liu Y, et al. Randomized Clinical Trial: Probiotics Alleviated Oral-Gut Microbiota Dysbiosis and Thyroid Hormone Withdrawal-Related Complications in Thyroid Cancer Patients Before Radioiodine Therapy Following Thyroidectomy. Front Endocrinol (Lausanne). 2022, 13: 834674. [CrossRef]
- Cheng SY, Chen NF, Kuo HM, et al. Prodigiosin stimulates endoplasmic reticulum stress and induces autophagic cell death in glioblastoma cells. Apoptosis. 2018, 23(5-6): 314-328. [CrossRef]
- Song D, Liang H, Qu B, et al. Moxidectin inhibits glioma cell viability by inducing G0/G1 cell cycle arrest and apoptosis. Oncol Rep. 2018, 40, 1348-1358. [CrossRef] [PubMed]
- Pang Z, Gu MD, Tang T. Pseudomonas aeruginosa in Cancer Therapy: Current Knowledge, Challenges and Future Perspectives. Front Oncol. 2022, 12: 891187. [CrossRef]
- Zhang M, Lu W. Enhanced glioma-targeting and stability of (L)GICP peptide coupled with stabilized peptide (D)A7R. Acta Pharm Sin B. 2018, 8, 106-115. [CrossRef]
- Al-Obaidi M, Desa M. Mechanisms of Blood Brain Barrier Disruption by Different Types of Bacteria, and Bacterial-Host Interactions Facilitate the Bacterial Pathogen Invading the Brain. Cell Mol Neurobiol. 2018, 38, 1349-1368. [CrossRef]
- Suryawanshi YR, Schulze AJ. Oncolytic Viruses for Malignant Glioma: On the Verge of Success. Viruses. 2021, 13(7). [CrossRef]
- Kim H, Roh HS, Kim JE, Park SD, Park WH, Moon JY. Compound K attenuates stromal cell-derived growth factor 1 (SDF-1)-induced migration of C6 glioma cells. Nutr Res Pract. 2016, 10, 259-64. [CrossRef]
- Hung AL, Garzon-Muvdi T, Lim M. Biomarkers and Immunotherapeutic Targets in Glioblastoma. World Neurosurg. 2017, 102: 494-506. [CrossRef]
- D'Alessandro G, Antonangeli F, Marrocco F, et al. Gut microbiota alterations affect glioma growth and innate immune cells involved in tumor immunosurveillance in mice. Eur J Immunol. 2020, 50, 705-711. [CrossRef]
- Hou X, Du H, Deng Y, et al. Gut microbiota mediated the individualized efficacy of Temozolomide via immunomodulation in glioma. J Transl Med. 2023, 21, 198. [CrossRef] [PubMed]
Figure 1.
History of the development of gut flora.
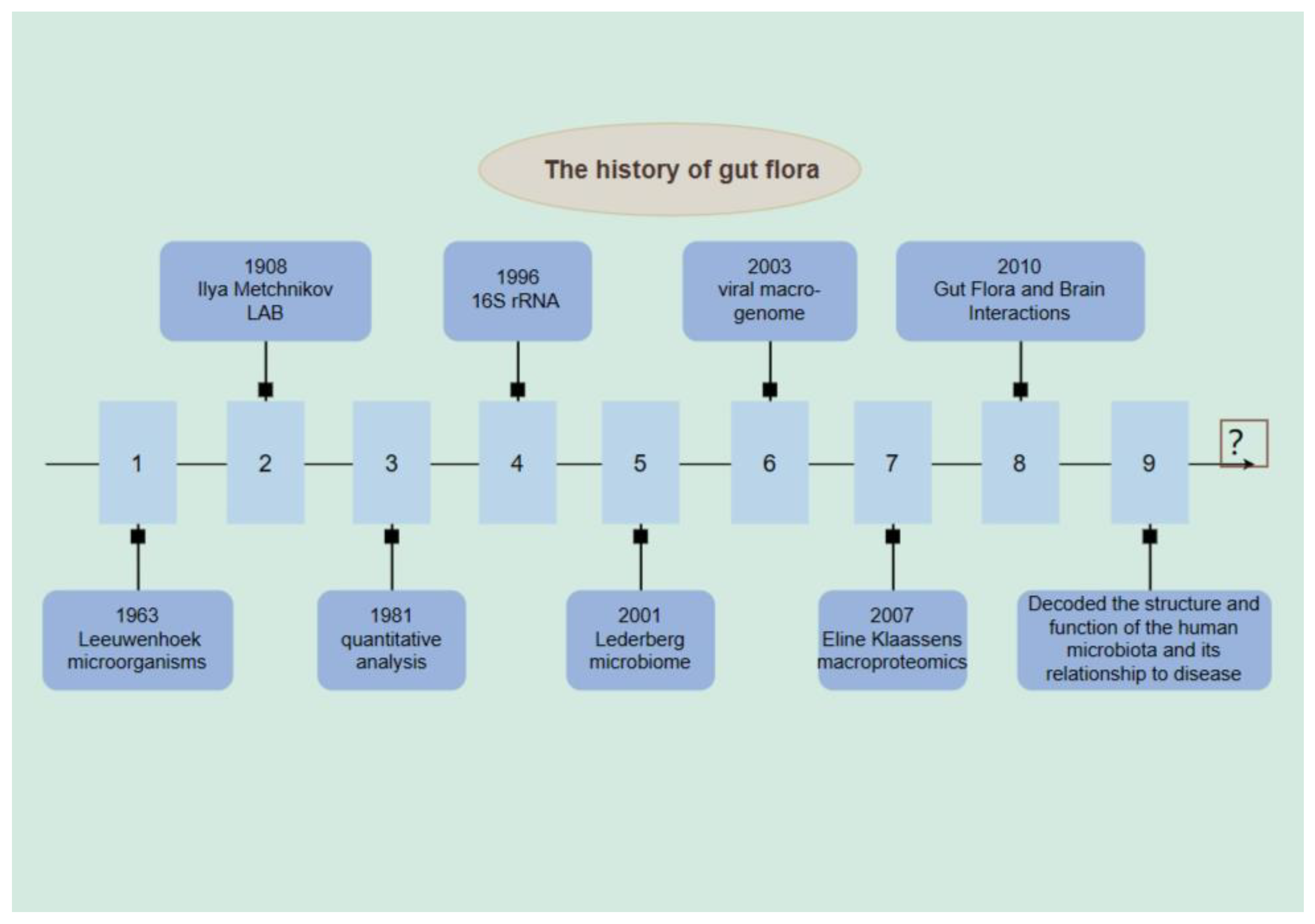
Figure 2.
Mechanisms of intestinal flora in disease.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



