
Submitted:
28 December 2023
Posted:
29 December 2023
You are already at the latest version
Abstract
Abstract: One of the most significant medical advancements of the 20th century was the discovery of antibiotics, which continue to play a vital tool in the treatment and prevention of diseases in humans and animals. However, the imprudent use of antibiotics in all fields of One-Health and concerns about antibiotic resistance among bacterial pathogens have raised interest in antibiotic use restrictions on a global scale. Despite the failure of conventional antimicrobial agents, only about 15 new antibiotics have been introduced clinically since year 2000 to date. Moreover, there have been reports of resistance to some of these new antibiotics. This has necessitated a need to search for an alternative strategy to combat antimicrobial resistant pathogens. Thus, this review compiles and evaluates the approaches—natural compounds, phage treatment, and nanomaterials—that are being used and/or suggested as the potential substitutes for conventional antibiotics. These strategies include those that focus on the enzymes or proteins, as well as the physiology and metabolism of resistant bacteria
Keywords:
Alternative therapy
; antimicrobial resistance
; natural product/compound
; One-Health
; secondary metabolite
Introduction
Antibiotics are used for a variety of reasons, including the ordinary support of human health and growth, the stimulation of growth in food-producing animals, which is not a therapeutic application—and the prevention of disease, which accounts for a large portion of antibiotic use in both humans and animals (Allen et al., 2014, Ajose et al., 2022a). Antibiotics have traditionally been regarded as the first line of defense in the treatment of bacterial diseases in animal husbandry. They have also prevented innumerable deaths and allowed for the advancement of modern medicine for many years (Czaplewski et al., 2016). However, their misuse and overuse have led to the development of antimicrobial resistance (AMR) in most of pathogenic bacteria. Bacterial resistance development is a persistent phenomenon that frequently renders even last-resort antibiotics ineffective. The lack of progress in the development of new antibacterial medications and the evolution of antibiotic resistance to practically all currently available antibiotics are factors in the global health crisis (Solomon and Oliver, 2014). In recent years, infectious diseases caused by bacteria have become serious and more prevalent in clinical settings, placing a serious economic burden and a threat to public health (de Kraker et al., 2016). According to McLinden et al. (2014), bacterial infections are thought to be responsible for at least half of all the economic costs of all foodborne diseases in the United States, which range from $10 to 83 billion yearly. This report was corroborated by Hoffmann et al. (2015). The spread of genes that confer antibiotic resistance among bacteria is aggravating the problem of antibiotic resistance (Peterson and Kaur, 2018). The acquisition of antibiotic resistance genes (ARGs) by the bacteria, may lead to treatment failure and morbidity and mortality rates in humans (San Millan, 2018).
Antibiotic resistance is a worldwide problem that is causing increasing public health concerns, and it is now recognized as a crucial One-Health issue (Ajose et al., 2022b). When contemplating potential interventions to mitigate or preempt this problem, it is imperative to elucidate the intricate elements that have contributed to the development of bacterial antibiotic resistance. Over 80 years of antibiotic use have put pressure on bacteria to evolve specialized mobile ARGs, especially in pathogenic bacteria. These AMR organisms in environmental "hot spots" translocated the phenomena of resistance to related species (Adhikari et al., 2022, Larsson and Flach, 2022) (Figure 1). Bacteria and their genetic material (DNA and/or plasmids) can be easily transmitted between humans, animals and/or environment (Figure 1). As a result, AMR is dangerous and is characterized by intricate interactions between various microbial populations that affect human, animal, and environmental health (WHO, 2015, Robbins et al., 2016). In light of this, measures made (or not done) to counteract AMR in one division may have an effect on other divisions.
The primary impact of AMR is the large rise in the risk of disease transmission, life-threatening illness, and death as antimicrobials lose their effectiveness and make infections harder to cure. AMR is noteworthy since it appears in all different forms (Murugaiyan et al., 2022). Notably, COVID-19 pandemic has accelerated the spread of MDR bacteria as a result of the overuse and abuse of currently available antibiotics as well as other antibiotic resistance-promoting factors (Manohar et al., 2020, Pelfrene et al., 2021) (Figure 2). Multidrug resistant (MDR) pathogens are becoming a major worldwide health problem because they have an impact on infection identification, treatment, and prevention. However, organisms that are extensively drug-resistant (XDR) and pan-resistant (PDR), which are practically intractable with conventional therapy, are of paramount concern (Murugaiyan et al., 2022). Since even so-called "antibiotics of last resort" are becoming ineffective, new strategies are being applied to prevent and cure MDR, XDR, and PDR infections in light of the impact AMR bacteria have on the world’s health and the need for new antibiotics (Mulani et al., 2019).
Louis Pasteur made groundbreaking discoveries in the fields of immunology, vaccinology, and microbiology that changed clinical science and save millions of deaths. Due to recently developed tools and infection models, our capacity to study infectious disease has experienced significant modifications since the 19th century. We saw a tremendous desire to comprehend the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection and to develop potent disease-prevention vaccines during the COVID-19 pandemic. We need to discover new or alternative strategies to tackle antibiotic-resistant bacterial strains that cause millions of deaths every year with the same haste that we did for the comparatively underappreciated AMR pandemic (Mostowy, 2022). Future advances in disease prevention or treatment will determine how successful humanity remains. The spread of ARGs among pathogenic bacteria raises concerns about the continued effectiveness of the current antibiotic arsenal (Allen et al., 2014).
It might be prudent to think about the possibilities of non-traditional techniques given the emergence of antibiotic resistance and the difficulties in discovering and developing conventional antibiotics, which have resulted in a very short pipeline of new medicines. Utilizing antibiotic alternatives seeks to improve and promote health and reduce antibiotic overuse. Hence, reducing the selective pressure that leads to the establishment and spread of ARGs. Thus, various alternative strategies in the following categories will be fully highlighted in this review:
- a)
- antimicrobial peptides (AMPs);
- b)
- bacteriophages;
- c)
- nanotechnology;
- d)
- ethno-medicine; and
- e)
- probiotics and prebiotics
Mode of Action of Antibiotics and Emergence of Antibiotic Resistance
Antibiotics are categorized into classes based on the class of molecules they include and their targets and principal modes of action (Table 1). Antimicrobial targets include cell membranes, cell walls, protein synthesis, DNA or RNA synthesis, and the creation of biological metabolic compounds.
The overuse and abuse of antibiotics in the human, veterinary, and agricultural sectors has put a lot of pressure on the selection process, leading to an increase in antibiotic resistance in bacteria (Murugaiyan et al., 2022) (Figure 3). This pressure is exerted through a number of mechanisms (Table 1), some of which are either emerging or ancestrally intrinsic to the biology of a pathogen. Some of the mechanisms of bacterial AMR include the following: (1) Enzymatic degradation of antibiotics, such as the bacterial production of beta-lactamases that break down the beta-lactam class of antibiotics; (2) Modification of the antibiotic target, where the target is altered so that the antibiotic can no longer bind to its site of action; (3) Control of drug entry through mutations in bacterial cell wall porin molecules and membrane modifications; (4) The turning on of efflux pump systems, which are able to pump antibiotics outside of the cell before they interact with their targets Figure 4.
Antimicrobial Peptides
Antimicrobial peptides (AMPs) are a class of tiny molecular peptides that are integral to the innate immune response of the host organism (Wang et al., 2019). They have a critical function in defending against a diverse array of pathogens, encompassing bacteria, fungi, parasites, and viruses (Mohammed et al., 2017, Kang et al., 2017, Lei et al., 2019). The use of AMPs is one of the methods for developing new antibiotics (Zharkova et al., 2019, Dijksteel et al., 2021). When compared to conventional antibiotics, AMPs found in nature have equivalent or even better antibacterial action (Tilocca et al., 2019). They are amphipathic in nature, thus thought to make it easier for the negatively charged bacterial membranes to interact electrostatically, which would then disrupt the lipid bilayer structure (Yeaman and Yount, 2003). Their size from 10 to 50 amino acids, and have an overall cationic charge (Sheard et al., 2019). AMPs are present everywhere to include mammals, amphibians, microorganisms and insects and in a variety of natural settings including the oceans (Huan et al., 2020). Through the similar mechanisms of conventional antibiotics, AMPs can target both Gram-positive and Gram-negative bacteria (Sheard et al., 2019, Rima et al., 2021). Once AMPs have penetrated the bacterial cell wall, they continue to act as an antibacterial agent by concentrating on protein biosynthesis, nucleic acids, and/or affecting membrane and cell wall formation (Nguyen et al., 2011, Le et al., 2017, Łoboda et al., 2018).
Antimicrobial Peptide Mode of Action
AMPs are receiving more attention, and new biophysical approaches are continually being developed, although the precise details of their molecular mechanism of action are still not fully understood. In bacteria with negatively charged surface layers made up of lipids like phosphatidylglycerol (PG), cardiolipin (CL), and lipopolysaccharides (LPS), or lipoteichoic acid (LTA) in positively charged surface bacteria, the combination of cationic and hydrophobic residues promotes a strong electrostatic interaction (Molchanova et al., 2017). Additionally, folding of the AMPs caused by hydrophobic contact with bacterial envelopes is what gives these peptides their antibacterial effects. The significant quantity of neutral phosphatidylcholine / cholesterol / sphingomyelin lipids gives mammalian cell membranes a zwitterionic surface, which results in a reduced attraction between cationic peptides and host cells. This is the source of the cell selectivity seen for many AMPs. Since AMPs impair the integrity of bacterial membranes, causing leakage of cell content and subsequent cell death, it is believed that the bacterial membrane is the primary target for AMPs (Torcato et al., 2013, Łojewska and Sakowicz, 2021). Models of AMP disruption of bacterial membranes have been described (Yeaman and Yount, 2003), and they include the carpet model (Shai and Oren, 2001), aggregation (Wu et al., 1999), molecular electroporation (Miteva et al., 1999), toroidal hole development (Hof et al., 2001), sinking raft (Pokorny et al., 2002), barrel-stave model (Huang, 2000), interfacial activity (Wimley, 2010), and lipocentric pore formation (Fuertes et al., 2011). Although the bacterial membrane is the primary target of AMP, intracellular targets including protein synthesis (indolicidin), nucleic acid synthesis (buforin II), RNA synthesis (Bac5 and Bac7), enzymatic activity (pyrrhocoricin), ATP efflux (histatins), or cell wall synthesis (nisin) have also been reported (Nguyen et al., 2011, Pasupuleti et al., 2012) (Figure 5).
Antimicrobial Peptides as Therapeutic Agents
The attributes that make AMPs ideal therapeutic candidates include broad-spectrum antibacterial activity against MDR pathogens, quick onset of killing, and a relatively low chance of resistance development (Sheard et al., 2019, Rima et al., 2021, Łojewska and Sakowicz, 2021). No linear peptides, but several natural AMPs have made it to the market. The peptide antibiotics bacitracin, polymyxin, gramicidin, and tyrothricin were among the first to be used in medicine (Sumi et al., 2015). Bacillus is an example of the bacteria that produce other peptide antibiotics. Tyothricin,the first AMP to be utilized in a clinical setting is composed of two major ingredients namely: tyrocidine and gramicidin. Because of its hemolytic effects, tyrocidine is a combination of cyclic decapeptides that is only used topically (Stevenson, 2009). Also, due to their hemolytic adverse effects, the broad-spectrum cyclodecapeptides tyrocidine A and gramicidin S can only be applied topically (Mösges et al., 2011, Murugaiyan et al., 2022).
The cyclic lipopeptides polymyxin B and polymyxin E (also called colistin) have been used to treat Gram-negative infections, and they are presently the last-resort treatment for MDR Gram-negative infections (Rabanal and Cajal, 2017). Due to their nephro- and neurotoxicity, polymyxins have a limited application; however, by using a prodrug formulation, their acute toxicity is mitigated by the matching sulfomethylated molecule. Nisin, a different naturally occurring (34-residue) AMP derived from Lactococcus lactis, is efficient against Gram-positive bacteria, particularly the pathogens that cause mastitis. Owing to its low toxicity, some countries such as England, Ireland and Denmark have granted the use of nisin as preservative in food industry to prevent proliferation of foodborne pathogens (Deegan et al., 2006). The treatment of infections brought on by MDR Gram-positive organisms includes the use of daptomycin, another cyclic lipopeptide (Humphries et al., 2013).
AMPs are highly effective against vegetative cells. For instance, the EmPis-1L peptide effectively destroys the antibiotic-resistant VBNC state cells of foodborne pathogenic bacteria like Escherichia coli O157 and Vibrio parahaemolyticus OS4 (Hu et al., 2019). Additionally, AMPs are capable of destroying bacterial biofilms structures (Yasir et al., 2018). AMPs LL-37 and α-defensin, salmine, lactoferrin, protamine, casecidin and isracidin, fibrinogen, pleurocidin, ll-AMP1 peptide, α-poly-l-lysine (poly-lys), α-poly-l-arginine (poly-arg) are notable examples of eukaryotic AMPs with therapeutic potential against foodborne pathogens (Wu et al., 2013, Cheng et al., 2015, Wu et al., 2016).
In order to overcome the issue of toxicity in the use of AMPs in clinical practice (human and veterinary sectors), diverse forms of approaches to enhance AMP stability are being considered. These include synthesis of constrained, hybridized, mimetic, immobilized and conjugated AMPs (Vallabani and Singh, 2018, Mitra et al., 2021). Among them, the conjugation of AMPs with conventional antibiotics is showing promise due to the possible synergistic interaction between the two drugs, which enables effective targeting and death of a number of pathogenic-resistant bacteria (Grassi et al., 2017, Li et al., 2018). For instance, when used to treat vancomycin-resistant Enterococci, AMP magainin and vancomycin exhibited very promising outcomes (Arnusch et al., 2012). By combining the cationic AMP ubiquicidin with chloramphenicol via a glutaraldehyde linker, similar outcomes were obtained, demonstrating increased antibiotic activity against E. coli and Staphylococcus aureus and decreased toxicity against human cells (Reinhardt and Neundorf, 2016). It’s interesting to note that promising outcomes have also been obtained by combining AMPs with antibiotics in an effort to improve in situ drug delivery, increase medication selectivity, and decrease toxicity (Riber et al., 2015). This depends on the fundamental mechanism of action of AMPs on the components of bacterial outer walls. Since they cannot enter the microbial cell when the antibiotic’s resistance mechanism involves membrane alteration, AMPs work better in synergy with antibiotics than when used alone. By being able to get beyond this obstacle, AMPs might be able to revive antibiotic activity that has been lost (Sierra et al., 2017).
Recently developed nanocarriers—drug delivery systems based on nanotechnology—could develop into a potent tactic for the effective delivery of AMPs. By shielding peptides from extracellular breakdown by proteases and other peptide-hydrolyzing conditions, AMP administration via nanocarriers may be favorable. Targeted nanocarriers may also help with enhanced medication pharmacokinetic characteristics and target selectivity. Drug delivery mechanisms come in a variety of forms, including liposomes, micelles, polymeric nanoparticles, and dendrimers (Biswaro et al., 2018, Ritsema et al., 2018).
Bacteriophages
Bacteriophages (phages) are perhaps among the oldest and most prevalent biological entities that can infect and replicate within bacteria. As a result, they are crucial to maintaining the balance of an environment where bacteria are present (Jaglan et al., 2022). There is now curiosity in utilizing phage as a biocontrol weapon because of the numerous alluring properties these natural antibacterial agents possess (Sillankorva et al., 2012). These include, low toxicity to humans, selectivity by acting only on their host without harming the natural microflora, relatively easy to propagate, cost-effective and sustainable solution to the control of pathogens (Shousha et al., 2015, Korf et al., 2019, Montso et al., 2019). In addition to being absorbed, they have also been separated from various human body parts, demonstrating their close ties to humans (Petsong et al., 2019, Fiscarelli et al., 2021). In the days before antibiotics, phages were utilized to combat infectious diseases agents soon after their discovery. However, their usage for therapeutic purposes was discontinued in the Western world after the development of antibiotics. The lack of additional therapeutic options and the ineffectiveness of traditional antibiotics in treating and managing harmful bacterial strains have recently accelerated the revival of phage therapy (Jaglan et al., 2022). In previous study, Montso et al. (2021) demonstrated that lytic phages effectively reduced E. coli O177 cells under artificial rumen fermentation conditions, and could therefore be used as a biocontrol strategy in live cattle to reduce meat and milk contamination in slaughterhouses and milking parlors, respectively.
Mechanism of Action of Bacteriophages
Once they have injected their DNA, phages employ the machinery of the host to replicate and interpret the information needed to generate viral progeny (in the case of lytic phages) and enzymes like spannin, holin, and endolysin (Briers, 2019, Gondil et al., 2020, Abeysekera et al., 2022). Caudoviricetes utilise holin-endolysin or pinholin single arrest release (SAR) mechanisms to break the peptidoglycan link of the cell wall in Gram-negative hosts. This causes host lysis and the release of viral offspring during the lytic cycle (Abdelrahman et al., 2021). Conversely, in lysogenic or temperate phages, the viral genome fuses with the host genome to form a prophage and replicates alongside it over a long period of time (Nanda et al., 2015, Pinto et al., 2020). Prophage induction, or its excision from the genome, can happen in response to cellular internal or external stimuli such as antibiotics, heat, low nutrient condition and ultraviolet (UV) light (Nanda et al., 2015). As a result, vulnerable bacteria are lysed and offspring phages are released, which may then go on to lyse or lysogenize further susceptible bacteria.
Application of Bacteriophages as Bio-control Agents
Phages have been adopted in a variety of sectors including medicine and agriculture. Phages have been employed to eliminate spoilage organisms in a variety of foods (Table 2) in both clinical and laboratory settings, reducing food loss and the costs that come with it. Bacteria can contaminate food ingredients before, during, or after manufacture. Any of these stages can be stopped or reduced by using certain phages. As a result, phages have lately proven to have a wide range of applications for improving safety in a number of food products (Table 2).
Phage therapy has been successfully used in clinical settings to treat severe or persistent bacterial infections (Abedon, 2019). Phage therapy for the treatment of persistent infections has primarily been used in Georgia, Poland, and Russia (Nikolich and Filippov, 2020, Barron, 2022). The type of infection, the type of phage or phage-based products, the dose of phage, and the method of delivery all affect how well the therapy works. It was reported that the Shanghai Institute of Phages in China administered clinical phage therapy to patients with MDR illnesses (Wu et al., 2021).
Application of Bacteriophages as Bio-sensor
Beyond the therapeutic application of phages, genetically engineered phages can be utilized to identify individual bacteria even in a combination of many different bacterial species; bio-sensors. Typically, sensors have two or more useful parts. The first step is some sort of target recognition aspect. Second, there must be some sort of reporting transducing mechanism for when the recognition element finds that target. Phages function something like sensors (or, more exactly, a biosensor). The target, a host bacteria, is located by the proteins that bind to receptors. The reporter is the phage’s genome, which produces other phages once it enters the target cell and may take the form of a plaque. Phages are extremely particular in identifying their target in this way, which can be a significant advantage in a biosensor (Gibb et al., 2021). Several methods use the typical phage recognition components to identify bacteria while modifying the reporter to increase the sensitivity or speed of the detection. Many detection methods merely depend on phage reproduction to signal the presence of the matching host cells (Ahovan et al., 2020, Meile et al., 2020).
Regulations for the Application of Phage Therapy
Phage therapy utilization in treatment regimens is not governed by any particular, global standards. Different standards for employing phage treatment in clinical cases have been developed by regulatory agencies around the world. Patients are treated in the United States under the FDA’s emergency investigational new drug (eIND) approval, whereas Australia has created a specific access program and Belgium is using a magistral phage method (Donovan, 2017, Jarow et al., 2017, Pirnay et al., 2018, Djebara et al., 2019). The widespread use of phage therapy has been hampered by the inconsistent restrictions in different nations. However, the first phage therapy facility in the United States, UC San Diego Center for Innovative Phage Application and Therapeutics, has just been founded, and numerous patients have successfully undergone phage therapy with no negative effects reported (Aslam et al., 2020).
A phage-based anti-E. coli O157:H7 product called EcoShieldTM was developed for use in red meat (Retrieved from the Intralytix website at http://www.intralytix.com/index.php?page=prod&id=2, October 2022). A different product called "Secure Shield E1" from the German company FINK TEC GmbH is intended to be used on the surfaces of beef carcasses. Each product has received FDA approval. Recent approval for the use of phage mixtures from OmniLytics (Sandy, UT), Micreos’ PhageGuard E., and Intralytix’s EcoShield PXTM to prevent shiga toxin producing E. coli (STEC) infection on a variety of food products was granted (Retrieved from https://www.fda.gov/food/generally-recognized-safegras/gras-notice-inventory, October 2022). The FDA approved the aforementioned phage products as generally recognized as safe (GRAS) food additives.
Other countries have also approved the use of phage to guarantee food safety. The National Food Service of the Israeli Ministry of Health approved all phages that had already been approved by the FDA to be used for analogous purposes in 2014 under the regulations titled "Guidelines: use of bacteriophages (bacteria-killing viruses) in food (Brüssow, 2019)." In Canada, phage-based products such as PhageGuard LTM (formerly ListexTM), ListShieldTM, SalmoFreshTM, and EcoShieldTM have been authorized as aids in food processing (Brüssow, 2019). For some ready-to-eat (RTE) products, the Food Standards Australia New Zealand in Australia and New Zealand approved the use of phage-based treatments to control Salmonella and Listeria monocytogenes as processing aids. In Europe, the situation is somewhat different. Despite previous attempts to approve phage-based products, none of them have yet to display the “CE mark”. Despite the regulatory barriers, ListexTM P100 has petitioned for clearance to be used as a decontamination agent to lower the level of L. monocytogenes in food, raw seafood (EFSA, 2012), and RTE food items (EFSA, 2016). To stop Listeria from developing in many food products, several food companies are already employing this phage product. In the absence of a regulatory framework from the European Union (EU), the European Court of Justice announced a court order in October 2019 permitting food producers to continue using phages to prevent Listeria on all RTE meals. European, American, Japanese, and other regulatory organizations acquired unified standards for the development of pharmaceuticals through the International Conference on Harmonization (ICH) (Jaglan et al., 2022).
Nanotechnology
Nanotechnology-based methodologies are employed across a range of disciplines, including engineering, material sciences, biology, and medicine. Nanomaterials are often defined as substances having sizes between 1 and 100 nm. To help provide distinctive features for a wide range of functions, they come in a variety of forms and sizes. When restricted to a relatively small scale, the material’s characteristics experience considerable modifications. Nanomaterials are typically made of metals, metal oxides, or composites made of carbon and emulsions (Mubeen et al., 2021). A paradigm change in antibacterial medicine has been brought about by the use of nanoscale materials as antibacterial agents. High surface area to volume ratio nanomaterials are being researched for the creation of potent bactericidal medications because they can enhance interactions with the target agents. They demonstrate efficacy against bacteria that have already developed resistance through the use of mechanisms that are fundamentally different from the modes of action of conventional antibiotics. They also target a number of biomolecules, posing a threat to the emergence of resistant strains (Adhikari et al., 2022).
Nanomaterials have been found to have increased membrane permeability, the capacity to function as efflux pump inhibitors, and the potential for a variety of antibacterial actions, making them less likely to produce bacterial resistance than conventional antibiotics (Rudramurthy et al., 2016, Slavin et al., 2017). In addition to providing important pharmacological advantages such as improved drug solubility and half-life, extended and stimuli-responsive drug release, site-targeted administration, and combination treatment, nanoparticles (NPs) are valuable as antimicrobial vector (Colilla and Vallet-Regí, 2020, Spirescu et al., 2021).
Classification of Nanomaterials and their Properties
Numerous nanostructures, including liposomes, NPs, and dendrimers, have demonstrated their capacity to increase the effectiveness of antibiotic treatments and fight infectious diseases (Lee et al., 2019, Singh et al., 2019). Nanomaterials have been investigated in vitro and in vivo to control and combat bacterial infection, including antibacterial polypeptides, noble metal NPs, nanocomposites, semiconductor NPs, polymeric nanostructures, and carbon-based nanomaterials (CNMs) (Yeon et al., 2019, Mba and Nweze, 2021, Mubeen et al., 2021). We have also discussed extensively on the various forms of synthesis, characterization, probable mode of action, application and/or antibacterial activities and advantages of nanotechnology over antibiotics in our previous works (Ajose et al., 2022b, Abolarinwa et al., 2022)
Since severe membrane damage would require extensive regeneration of membrane components, which would be physically taxing for target cells, membrane-acting NPs are anticipated to be highly alluring and detrimental to the establishment of resistance. Modern physicochemical polymeric NPs have been proven to be a breakthrough treatment for bacterial diseases in humans (Spirescu et al., 2021). In comparison to currently available substances, this family of nanocarriers have a number of benefits including being proven to be safe, biodegradable, biocompatible, swiftly eliminated, and non-toxic to tissues and organs with superior pharmacokinetics properties. Additionally, this kind of therapy represents a breakthrough due to its ability to target a particular organ, reduce the adverse effects of numerous antibiotics, and gradually accumulate in the area that is diseased thus, more effective than conventional antibiotics for a longer period of time, which is crucial for a long-lasting, sustained therapeutic impact. The antibacterial potential of many inorganic (metal) NPs, including gold (Au), silver (Ag), and others, has been demonstrated (Gharpure et al., 2020, Sánchez-López et al., 2020). With good thermal stability, low toxicity, and antibacterial qualities, Ag is a noble metal. It has been shown that polyurethane and plastic catheters with AgNP surface functionalization are effective at preventing the growth of biofilms on a variety of harmful bacteria (Prasher et al., 2018).
Due to their distinctive physicochemical characteristics, CNMs have garnered general acceptance in the scientific community, but their clinical efficacy has not yet been established. These materials’ insoluble nature, which makes it difficult for them to enter biological systems, is overcome by carbon nanostructures with programmable morphologies. Due to their capacity to eliminate harmful bacteria and stop their adhesion and biofilm formation, carbon NPs in general are promising antibacterial candidates with a range of biological applications (Xin et al., 2019, Azizi-Lalabadi et al., 2020). In bacteria, the adsorption of positive ions such as Ag+, Cu2+, and Zn2+ onto the bacterial cell membrane can lead to the breakdown of the cell wall and the creation of pits (Gupta et al., 2019a). A breakthrough in this kind of therapy is also represented by the targeting of a particular organ, the reduction of many antibiotics’ side effects, and the sustained accumulation in the infected area over time. Au, Ag, and other inorganic NPs have demonstrated antibacterial properties (Gharpure et al., 2020, Sánchez-López et al., 2020). A noble metal with good thermal stability, low toxicity, and antibacterial qualities is Ag. AgNP surface functionalization of polyurethane and plastic catheters was shown to be effective at preventing the growth of biofilms from a variety of pathogenic bacteria (Prasher et al., 2018).
Applications of Nanotechnology
Although NPs are among the most effective weapons against MDR bacteria, the pharmaceutical industry still faces challenges in translating NP-based therapies into everyday use because nanotechnology hasn’t yet permeated clinical practice. However, Martinez et al. (2020) demonstrated the capacity of photoactive metallated porphyrin-doped conjugated polymer NPs to eliminate pathogenic bacterial strains, such as antibiotic-resistant bacteria of the Enterococcus faecium, S. aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp. (ESKAPE) pathogens group. Methicillin-resistant S. aureus (MRSA) can cause major infections with high morbidity and death in both the community and the hospital because it has evolved resistance to nearly all antibiotics. Functional nanomaterials and NPs can kill MRSA and be used in anti-MRSA therapy (Gao et al., 2021).
Some NPs are in clinical trials or have already received FDA approval for use in humans, and numerous proof-of-concept studies using nanomaterials in cell culture and small animal models are now underway (Anselmo and Mitragotri, 2016, Ventola, 2017). According to Gupta et al. (2019b), the biofilms of S. aureus and P. aeruginosa isolated from food were found to be susceptible to a concentration of 62.5 mg/mL of AgNP, but doses of 125 and 250 mg/mL of the same NP inhibited biofilms by 85 and 90%, respectively. In a another study, the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of green synthesized AgNPs against MDR P. aeruginosa (MDR-PA) were reported as 3.88 ± 0.13 and 7.77 ± 0.25 g/mL and 7.77 ± 0.25 and 31.08 ± 1.01 g/mL against MRSA, respectively (Ansari et al., 2021).
Ethno-Medicine
The Effect of Antibiotic Resistance on Sustainable Development Goals
The best plan currently available for creating a better world for people and the planet by the year 2030 is the set of 17 United Nations Sustainable Development Goals (UN SDGs). The UN SDGs, which were adopted by all United Nations Member States in 2015, are a call to action for all nations; rich, middle-income, and poor, to advance prosperity while preserving the environment. The fight against AMR and the SDGs go hand in hand. AMR has an impact on achieving a number of the 17 SDGs, especially SDG 3: "Good health and well-being."
AMR is now officially mentioned under SDG 3, and fulfilling many SDGs also depends on properly addressing AMR. First, with the use of efficient antibiotics, epidemics of infectious diseases including human immunodeficiency virus (HIV), gonorrhea, and tuberculosis can be controlled as well as maternal, neonatal, and pediatric fatalities can be avoided (SDG 3). Second, antibiotics are crucial for human health as well as the production of food and animals as well as for livelihoods (SDG 8, 2, 1). Third, improperly handled medical waste and wastewater can introduce antibiotic-resistant bacteria into soils, drinking water, and groundwater (SDG 6, 12). Finally, collaboration is necessary to combat AMR. The Tripartite, a group made up of the World Health Organization (WHO), the Food and Agriculture Organization (FAO), and the World Organization for Animal Health (OIE), has chosen to address AMR using the One Health strategy (SDG 17) (CDC, 2022, WHO, 2022).
The recently introduced SDG indicator on AMR intends to lower the percentage of bloodstream infections brought on by E. coli and S. aureus that are resistant to 3rd-generation cephalosporin drugs and methicillin, respectively. The ability to treat infections with currently available antibiotic will ultimately be preserved by the effective management of these two categories of antibiotic resistant pathogens while new preventative and treatment approaches are developed (WHO, 2022).
Plants Secondary Metabolites: Key Drivers of Pharmacological Actions of Medicinal Plants
Natural and/or organic by-products continue to be crucial to the success of drug development. Therefore, biological variety provides a never-ending supply of novel chemical entities (NCEs) with the potential to be therapeutic leads. These NCEs are made from substances that plants produce to protect them from herbivores and diseases or to entice pollinators. The vast majority of chemical compounds that are produced by plants, known as secondary metabolites, are those that are indirectly linked with development and growth (Harborne, 1999). Indicators of the potency of natural items as modern medications include the effectiveness of secondary metabolites in plants (Jahn et al., 2012). The biological effects of secondary metabolites have been shown to vary, providing the scientific underpinning for the use of herbs in traditional medicine in many ancient communities. They are able to shield plants from pathogens because they are antibiotic, antifungal, and antiviral in nature. They also have antibiofilm or anti-quorum sensing function (Adhikari et al., 2022). According to their chemical structures, plant secondary metabolites are divided into a variety of classes which include: phenolics (tannins, coumarins, flavonoids), alkaloids (ergots, imidazoles, pyridines, purines, quinolines), saponins (steroidal or triterpenoidal saponins), terpenes (mono-, sesqui-, sester-, hemi-, di-terpenes), lipids (fixed oil, waxes, essential oil) and carbohydrates (mono-,di-,oligo- and polysaccharides) (Bennetts et al., 1946).
Ethno-Medicine as an Alternative to Antibiotic
Since the dawn of time, people have had close relationships with plants for use as food, fuel, medicine, and clothing. The use of medicinal plants is common in both industrialized and developing nations, and it has a long history even before antibiotics were produced (Alli and Mangamoori, 2016, Anju et al., 2022). Currently, as a result of the development and the effects of resistance to antibiotics, millions of people in developing nations throughout the world use medicinal plants and other natural goods as a way to treat infectious diseases. The market for the use of natural products for therapeutic purposes is expanding at a rapid rate. Ethno-medicine is applied in various sectors of life including medicinal, pharmaceutical and agricultural. Plants are sleeping giants of the modern pharmaceuticals. Hence, they are being explored as alternative antimicrobials. Several researchers have reported the activities of medicinal plants in the various sectors (Ajose and Okozi, 2017, McGaw et al., 2020, Mwinga et al., 2022, Ajose et al., 2022a), usage and antimicrobial form of secondary metabolites (Table 3).
Globally, the plant kingdom offers a variety of species that are used as therapies for different illnesses (Brusotti et al., 2014). According to World Health Organization (WHO) reports, a significant portion of the global population uses traditional medicines, including those that use plant decoctions or functional chemicals (WHO, 1991, WHO, 2013). The most crucial need for using an ethno-pharmacological approach is knowledge of the plant parts previously employed as remedies. The two most well-known traditional therapies for treating various illnesses are Chinese herbal medicine and Ayurveda, but in areas where there are no texts to consult, ethno-botanical surveys are the only method to learn about the traditional use of medicinal plants.
Probiotics and Prebiotics
For today’s health-conscious population, food serves as more than just a source of energy; they also look for food components and nutrients that can improve health benefits or avoid chronic diseases (Webb, 2011). The emphasis of nutritional biology is therefore on "functional foods," which largely consist of probiotics, prebiotics, and synbiotics. Additionally, functional foods give the body the necessary amounts of lipids, proteins, carbohydrates, and vitamins (Cencic and Chingwaru, 2010). Prebiotic and probiotic techniques involve the use of microbial food supplements that benefit the host by enhancing the balance of intestinal microbes. Prebiotics and probiotics are preferable to antibiotics for treatment since they are safe for long-term usage and do not induce allergies or negative effects (Ghosh et al., 2019).
Applications and Mechanism of Action of Probiotics
Dietary fibers known as prebiotics are frequently combined with probiotics to increase their viability. The most common prebiotics include fructo- and galacto-oligosaccharides (FOS and GOS), xylo- and oligosaccharides (XOS), inulin, and fructans. Synbiotics is a common word for the combination. Therefore, synbiotics are a carefully chosen combination of probiotics and prebiotics that aid in the development of a balanced microbiota in the intestine of the host. It is crucial to reach agreement on the makeup and functions of probiotics, prebiotics, and synbiotics in promoting human health. Some of the probiotics drugs marketed worldwide include enteral capsules (Vanhee et al., 2010), acidolac (Zawistowska-Rojek et al., 2016), biogermin (Celandroni et al., 2019), entromax (Kesavelu et al., 2020), bifilac GG (Kesavelu et al., 2020), and gutpro (Kesavelu et al., 2020). Probiotics have been found to have other immunomodulatory activities that are advantageous in conditions like allergies, cancer, obesity and type 2 diabetes, aging, fatigue, autism, osteoporosis, and others, in addition to limiting the growth of drug-resistant bacteria (Harish and Varghese, 2006 ).
Probiotic effects vary based on the strain combination employed (Prado et al., 2015). According to recent studies, using probiotics can help prevent or treat yeast infections that develop as a result of antibiotic use, abnormalities of the lower intestine’s epithelium caused by Clostridium difficile, and antibiotic-associated diarrhea (AAD) in humans (Ferreira et al., 2017, Adhikari et al., 2022), as antimicrobial agents against foodborne pathogens (Li et al., 2014), reduction of cross-contamination and dissemination of infections (Crouzet et al., 2015), as feed additives in agricultural production (Frizzo et al., 2018, Mohamed et al., 2019, Ringø, 2020), and combating biofilm formation of pathogenic E. coli (Fang et al., 2018). de Melo Pereira et al. (2018) and (Tang et al., 2022) reported that several probiotic bacteria are known to produce a variety of antimicrobial substances, organic acids, and bacteriocins that can prevent the growth of bacteria that are resistant to multiple drugs.
Forest ecosystem research revealed that fungal and bacterial communities can respond to environmental changes in accordance with their host trees (Park et al., 2020). Other research has shown that microbial and biochemical indicators of soil health can be used to evaluate the ecological risk of soil. These findings demonstrated that soil respiration can be utilized to estimate soil ecological conditions and microbial activity (Niemeyer et al., 2012).
In environmental health, probiotics play an essential role as agents of remediation, in assisting the host to adapt to environmental changes. Certain genera also function as bioremediation or decomposition agents of hazardous substances (Helmy et al., 2019), such as a bacterial consortium (Xanthomonadaceae, Brachybacterium sp., Bhargavaea sp., Gordonia sp., Thalassospira sp., Pseudomonas sp., Dietzia sp., Mesorhizobium sp., Cytophaga sp. Conventional remediation strategies for the majority of types of environmental contamination are not only costly but also ineffective, particularly at low concentrations of contaminants (Goyal et al., 2019). Probiotics-assisted remediation has emerged as an inexpensive and straightforward alternative.
The theory behind the use of probiotics is that once the microbial flora in the gut has returned to equilibrium, the commensal bacteria can outgrow and actively exclude harmful strains. These commensal bacteria foster disease resistance either directly, through interactions with other bacteria, or indirectly, by triggering host immune systems (Isolauri et al., 2002, Łojewska and Sakowicz, 2021). However, probiotics also carry the danger of introducing and/or transmitting AMR characteristics via a variety of routes and may also cause non-genetically determined resistance in the endogenous microflora (phenotypic susceptibility due to the probiotic strain) (Palma et al., 2020). To establish uniform procedures for evaluating the safety of probiotics before approving their use in clinical practice, several research groups and regulatory organizations are concentrating on resolving the issue of resistance (Ouwehand et al., 2016).
Established Probiotics Risk Assessment Protocol
Probiotics may be clinically effective, but it is still important to guarantee the safety of the involved microbe. It is preferable to talk about probiotic safety generally since it affects both people and farm animals. More significant than the source of the isolate are the specific properties of a probiotic. It is essential that a probiotic agent’s potential endures for a long time at the site of action (Alayande et al., 2020). This implies that enough consideration must be given to the examination of a particular strain’s risk factors. Due to their extensive history of usage as probiotics, the majority of probiotic strains have earned the designation "Generally Recognized as Safe" (GRAS) (Plessas et al., 2017). However, not all innovative probiotics may be expected to have the same safety record as the traditional strains. The synthesis of toxic biochemical, virulence factors, transferrable ARGs, and hemolytic potential are only a few of the unfavorable traits that probiotics may have (Lee et al., 2017, Ahmad et al., 2022). For instance, it has been underlined that there may be a risk of horizontal antibiotic-resistant gene (ARG) transfer from one probiotic strain to other bacteria in the host’s gastrointestinal environment (Plessas et al., 2017). Along with this, there have also been a small number of reports of infectious disorders such endocarditis, bacteremia, pneumoniae, meningitis, and septic arthritis linked to specific Lactobacillus and Enterococcus strains, which have largely affected immunocompromised people (Vankerckhoven et al., 2008).
Although it is a member of the lactic acid bacteria, Enterococcus was listed as one of the bacteria with a high potential for virulence features in a joint study of the Food and Agriculture Organization/World Health Organization (FAO/WHO) Expert Consultation in Rome (FAO/WHO, 2006). Due to the fact that the Enterococcus genus frequently exhibits a high amount of vancomycin-resistant genes, its use as a probiotic has been discouraged.
Other potential risk factors that need to be highlighted include the potential to foster harmful metabolic effects, excessive immune stimulation, and toxigenicity of the particular strain, level of purity of the product, colonization and genetic stability of the strain over time (Suresh et al., 2013). Additionally, the FAO working group study (FAO/WHO, 2002) advises that prospective probiotic strains be examined for virulence in animal models with weakened immune systems and potential negative effects on end users. In addition, Kim et al. (2018) suggest the following guidelines for the safety assessment and regulation of probiotics, which have been approved by the European Union Scientific Committee on Animal Nutrition: taxonomical definition of the strains, collection of substantial information revealing data such as history of use, industrial applications, ecological niche, human intervention, exclusion of pathogenicity, and description of end users.
Conclusion and Future Perspective
Although the use of antibiotics in controlling bacterial infections is still necessary, it is critical to drastically reduce their use in all facets of One-Health by using alternate strategies to limit the pathogens. Since antibiotic resistance is mostly brought about by the abuse of antibiotics, antibiotic-free techniques are a viable new approach to deal with the current antibacterial dilemma. Hence, a range of approaches are required for both microbial illness prevention and treatment. The use of an effective combination of living and nonliving structures from natural resources is one of the promising "one health" approaches for addressing the disruption of human, animal, plant, and environmental health. From preclinical optimization to phase 3 studies, academic researchers and the pharmaceutical industries have successfully produced a diverse portfolio of potential antibiotic alternatives projects. Some approaches, such as nanomaterials, phage therapy, natural compounds, and probiotics, have been prioritized for more thorough review. These approaches have shown potential in studies, which are still in the early stages.
Further insight into how the alternative measures function to fight antibiotic resistant pathogens will come from a complete comprehension of the equilibrium between the opposing mechanisms of action and resistance. These discoveries may offer fresh approaches or models from which new drugs can be created to cure or prevent illnesses, particularly those brought on by microorganisms resistant to common antibiotics. In order to reduce pathogen resistance to host defenses, restore or amp up the actions of conventional antibiotics against drug-resistant infections and target strategic microbial structures or functions, pharmacologic compounds may be produced and identified. These viewpoints suggest that there may be numerous undiscovered or underappreciated mysteries about the alternative approaches mechanisms of action and resistance.
Author Contributions
Conceptualization, D.J.A.; methodology, D.J.A. and C.N.A.; validation, D.J.A. P.K.M. and C.N.A.; investigation, D.J.A.; writing—original draft preparation, D.J.A.; writing—review and editing, D.J.A. P.K.M. and C.N.A.; supervision, C.N.A.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the North-West University and the Department of Microbiology, School of Biological Sciences.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to appreciate the Department of Microbiology, School of Biological Sciences, North-West University, Mafikeng, South Africa.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- ABDELRAHMAN, F., EASWARAN, M., DARAMOLA, O. I., RAGAB, S., LYNCH, S., ODUSELU, T. J., KHAN, F. M., AYOBAMI, A., ADNAN, F. & TORRENTS, E. Phage-encoded endolysins. Antibiotics 2021, 10, 124.
- ABEDON, S. T.Use of phage therapy to treat long-standing, persistent, or chronic bacterial infections. Advanced Drug Delivery Reviews 2019, 145, 18–39. [CrossRef] [PubMed]
- ABEYSEKERA, G. S., LOVE, M. J., MANNERS, S. H., BILLINGTON, C. & DOBSON, R. C. Bacteriophage-encoded lethal membrane disruptors: Advances in understanding and potential applications. Frontiers in Microbiology 2022, 13, 1044143.
- ABOLARINWA, T. O., AJOSE, D. J., OLUWARINDE, B. O., FRI, J., MONTSO, K. P., FAYEMI, O. E., AREMU, A. O. & ATEBA, C. N. Plant-derived Nanoparticles as Alternative Therapy against diarrhoeal pathogens in the era of antimicrobial resistance: A review. Frontiers in Microbiology 2022, 4991.
- ADHIKARI, M. D., SAHA, T. & TIWARY, B. K. 2022. Quest for alternatives to antibiotics: An Urgent Need of the Twenty-First Century. Alternatives to Antibiotics. Springer.
- AHMAD, R., YU, Y.-H., HSIAO, F. S.-H., DYBUS, A., ALI, I., HSU, H.-C. & CHENG, Y.-H. Probiotics as a friendly antibiotic alternative: assessment of their effects on the health and productive performance of poultry. Fermentation 2022, 8, 672.
- AHOVAN, Z., HASHEMI, A., DE PLANO, L., GHOLIPOURMALEKABADI, M. & SEIFALIAN, A. Bacteriophage based biosensors: trends, outcomes and challenges. Nanomaterials 2020, 10.
- AJOSE, D. J., OLUWARINDE, B. O., ABOLARINWA, T. O., MONTSO, P. K., FAYEMI, O. E., AREMU, A. O. & ATEBA, C. N. Combating bovine mastitis in the dairy sector in an era of antimicrobial resistance: ethnoveterinary medicinal option as a viable alternative approach. Frontiers in Veterinary Science 2022a, 10, 287.
- AJOSE, D. J., ABOLARINWA, T. O., OLUWARINDE, B. O., MONTSO, P. K., FAYEMI, O. E., AREMU, A. O. & ATEBA, C. N. Application of plant-derived nanoparticles (PDNP) in food-producing animals as a bio-control agent against antimicrobial-resistant pathogens. Biomedicines, 2022b, 2426.
- AJOSE, D. J. & OKOZI, M. O. Antibacterial activity of Ocimum gratissimum L. against some selected human bacterial pathogens. Journal of Pharmaceutical Research International 2017, 20, 1–8.
- AKMAL, M., RAHIMI-MIDANI, A., HAFEEZ-UR-REHMAN, M., HUSSAIN, A. & CHOI, T.-J. Isolation, characterization, and application of a bacteriophage infecting the fish pathogen Aeromonas hydrophila. Pathogens 2020, 9, 215.
- ALAYANDE, K. A., AIYEGORO, O. A. & ATEBA, C. N. Probiotics in animal husbandry: Applicability and associated risk factors. Sustainability 2020, 12, 1087.
- ALLEN, H. K., TRACHSEL, J., LOOFT, T. & CASEY, T. A. Finding alternatives to antibiotics. Annals of the New York Academy of Sciences 2014, 1323, 91–100.
- ALLI, K. & MANGAMOORI, L. N. Phytochemical compound identification and evaluation of antimicrobial activity of Eugenia Bracteata Roxb. International Journal of Biotechnology and Biochemistry 2016, 12, 73–83.
- ANBALAHAN, N. Pharmacological activity of mucilage isolated from medicinal plants. International Journal of Applied and Pure Science and Agriculture 2017, 3, 98–113.
- ANJU, T., RAI, N. K. S. & KUMAR, A. Sauropus androgynus (L.) Merr.: a multipurpose plant with multiple uses in traditional ethnic culinary and ethnomedicinal preparations. Journal of Ethnic Foods 2022, 9, 1–29.
- ANSARI, M. A., KALAM, A., AL-SEHEMI, A. G., ALOMARY, M. N., ALYAHYA, S., AZIZ, M. K., SRIVASTAVA, S., ALGHAMDI, S., AKHTAR, S. & ALMALKI, H. D. Counteraction of biofilm formation and antimicrobial potential of Terminalia catappa functionalized silver nanoparticles against Candida albicans and multidrug-resistant Gram-negative and Gram-positive bacteria. Antibiotics 2021, 10, 725.
- ANSELMO, A. C. & MITRAGOTRI, S. Nanoparticles in the clinic. Bioengineering & Translational Medicine 2016, 1, 10–29.
- ARNUSCH, C. J., PIETERS, R. J. & BREUKINK, E. Enhanced membrane pore formation through high-affinity targeted antimicrobial peptides. PLoS One 2012, 7, e39768.
- ASIF, H., AKRAM, M., SAEED, T., KHAN, M., AKHTAR, N., REHMAN, R., SHAH, S., AHMED, K. & SHAHEEN, G. Carbohydrates. Intern. Res. J. Biochem. Bioinformat 2011, 1, 1–5.
- ASLAM, S., LAMPLEY, E., WOOTEN, D., KARRIS, M., BENSON, C., STRATHDEE, S. & SCHOOLEY, R. T. Lessons learned from the first 10 consecutive cases of intravenous bacteriophage therapy to treat multidrug-resistant bacterial infections at a single center in the United States. Open forum infectious diseases, 2020. Oxford University Press US, ofaa389.
- AZIZI-LALABADI, M., HASHEMI, H., FENG, J. & JAFARI, S. M. Carbon nanomaterials against pathogens; the antimicrobial activity of carbon nanotubes, graphene/graphene oxide, fullerenes, and their nanocomposites. Advances in Colloid and Interface Science 2020, 284, 102250.
- BARRON, M. 2022. Phage therapy: past, present and future. American Society for Microbiology.
- BENNETTS, H., UNDERWOOD, E. & SHIER, F. L. A specific breeding problem of sheep on subterranean clover pastures in western Australia. Veterinary Journal 1946, 102, 348–352.
- BISWARO, L. S., DA COSTA SOUSA, M. G., REZENDE, T. M., DIAS, S. C. & FRANCO, O. L. Antimicrobial peptides and nanotechnology, recent advances and challenges. Frontiers in Microbiology 2018, 9, 855.
- BRIERS, Y. Phage lytic enzymes. Viruses 2019, 11, 113. [CrossRef] [PubMed]
- BRUSOTTI, G., CESARI, I., DENTAMARO, A., CACCIALANZA, G. & MASSOLINI, G. Isolation and characterization of bioactive compounds from plant resources: the role of analysis in the ethnopharmacological approach. Journal of Pharmaceutical and Biomedical Analysis 2014, 87, 218–228.
- BRÜSSOW, H. Hurdles for phage therapy to become a reality—an editorial comment. Viruses 2019, 11(6), 557. [CrossRef]
- CDC. 2022. ANTIMICROBIAL-RESISTANCE THREATENS SUCCESSFUL ACHIEVEMENT OF THE SDG TARGET [Online]. Available: https://www.cdc.gov/drugresistance/pdf/amr-sdg-pamphlet.pdf [Accessed December 26, 2022].
- CELANDRONI, F., VECCHIONE, A., CARA, A., MAZZANTINI, D., LUPETTI, A. & GHELARDI, E. Identification of Bacillus species: implication on the quality of probiotic formulations. PLoS One 2019, 14, e0217021.
- CENCIC, A. & CHINGWARU, W. The role of functional foods, nutraceuticals, and food supplements in intestinal health. Nutrients 2010, 2, 611–625.
- CHENG, C., ARRITT, F. & STEVENSON, C. Controlling Listeria monocytogenes in cold smoked salmon with the antimicrobial peptide salmine. Journal of Food Science 2015, 80, M1314–M1318.
- COFFEY, B., RIVAS, L., DUFFY, G., COFFEY, A., ROSS, R. P. & MCAULIFFE, O. Assessment of Escherichia coli O157: H7-specific bacteriophages e11/2 and e4/1c in model broth and hide environments. International Journal of Food Microbiology 2011, 147, 188–194.
- COLILLA, M. & VALLET-REGÍ, M. Targeted stimuli-responsive mesoporous silica nanoparticles for bacterial infection treatment. International Journal of Molecular Sciences 2020, 21, 8605.
- CORREIA, S., POETA, P., HÉBRAUD, M., CAPELO, J. L. & IGREJAS, G. Mechanisms of quinolone action and resistance: where do we stand? Journal of Medical Microbiology 2017, 66, 551–559.
- CROUZET, L., RIGOTTIER-GOIS, L. & SERROR, P. Potential use of probiotic and commensal bacteria as non-antibiotic strategies against vancomycin-resistant Enterococci. FEMS Microbiology Letters 2015, 362, fnv012.
- CULIOLI, G., MATHE, C., ARCHIER, P. & VIEILLESCAZES, C. A lupane triterpene from frankincense (Boswellia sp., Burseraceae). Phytochemistry 2003, 62, 537–541.
- CZAPLEWSKI, L., BAX, R., CLOKIE, M., DAWSON, M., FAIRHEAD, H., FISCHETTI, V. A., FOSTER, S., GILMORE, B. F., HANCOCK, R. E. & HARPER, D. Alternatives to antibiotics—a pipeline portfolio review. The Lancet Infectious Diseases 2016, 16, 239–251.
- DAVIS, J. L. Pharmacologic principles. Equine Internal Medicine 2018, 4, 79–137.
- DE KRAKER, M. E., STEWARDSON, A. J. & HARBARTH, S. Will 10 million people die a year due to antimicrobial resistance by 2050? PLoS Medicine 2016, 13, e1002184.
- DE MELO PEREIRA, G. V., DE OLIVEIRA COELHO, B., JÚNIOR, A. I. M., THOMAZ-SOCCOL, V. & SOCCOL, C. R. How to select a probiotic? A review and update of methods and criteria. Biotechnology Advances 2018, 36, 2060–2076.
- DEEGAN, L. H., COTTER, P. D., HILL, C. & ROSS, P. Bacteriocins: biological tools for bio-preservation and shelf-life extension. International Dairy Journal 2006, 16, 1058–1071.
- DIJKSTEEL, G. S., ULRICH, M. M., MIDDELKOOP, E. & BOEKEMA, B. K. Lessons learned from clinical trials using antimicrobial peptides (AMPs). Frontiers in Microbiology 2021, 12, 616979.
- DJEBARA, S., MAUSSEN, C., DE VOS, D., MERABISHVILI, M., DAMANET, B., PANG, K. W., DE LEENHEER, P., STRACHINARU, I., SOENTJENS, P. & PIRNAY, J.-P. Processing phage therapy requests in a Brussels military hospital: Lessons identified. Viruses 2019, 11, 265.
- DONOVAN, P. Access to unregistered drugs in Australia. Australian Prescriber 2017, 40, 194. [CrossRef]
- DOWLING, A., O’DWYER, J. & ADLEY, C. Antibiotics: mode of action and mechanisms of resistance. Antimicrobial Research: Novel Bioknowledge and Educational Programs 2017, 1, 536–545.
- EFSA, E. P. O. B. H. Scientific Opinion on the evaluation of the safety and efficacy of Listex™ P100 for the removal of Listeria monocytogenes surface contamination of raw fish. EFSA Journal 2012, 10, 2615.
- EFSA, E. P. O. B. H. Evaluation of the safety and efficacy of Listex™ P100 for reduction of pathogens on different ready-to-eat (RTE) food products. EFSA Journal 2016, 14, e04565.
- FAHY, E., SUBRAMANIAM, S., MURPHY, R. C., NISHIJIMA, M., RAETZ, C. R., SHIMIZU, T., SPENER, F., VAN MEER, G., WAKELAM, M. J. & DENNIS, E. A. Update of the LIPID MAPS comprehensive classification system for lipids1. Journal of Lipid Research 2009, 50, S9–S14.
- FANG, K., JIN, X. & HONG, S. H. Probiotic Escherichia coli inhibits biofilm formation of pathogenic E. coli via extracellular activity of DegP. Scientific Reports 2018, 8, 1–12.
- FAO/WHO 2002. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations: Rome, Italy, 30 April and 1 May 2002.
- FAO/WHO. 2006. Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live. In Health and Nutrition Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria; FAO Food and Nutrition Paper 85; FAO: Rome, Italy, 2006 [Online]. Available: https://www.fao.org/3/a0512e/a0512e.pdf [Accessed December 27, 2022].
- FERREIRA, A., PEREIRA-MANFRO, W. & ROSA, A. D. P. Diarrheagenic Escherichia coli and probiotic activity against foodborne pathogens: a brief review. Gastroenterol Hepatol Open Access 2017, 7, 00248.
- FISCARELLI, E. V., ROSSITTO, M., ROSATI, P., ESSA, N., CROCETTA, V., DI GIULIO, A., LUPETTI, V., DI BONAVENTURA, G. & POMPILIO, A. In vitro newly isolated environmental phage activity against biofilms preformed by Pseudomonas aeruginosa from patients with cystic fibrosis. Microorganisms 2021, 9, 478.
- FRIZZO, L. S., SIGNORINI, M. L. & ROSMINI, M. R. 2018. Probiotics and prebiotics for the health of cattle. Probiotics and prebiotics in animal health and food safety. Springer.
- FUERTES, G., GIMÉNEZ, D., ESTEBAN-MARTÍN, S., SÁNCHEZ-MUNOZ, O. L. & SALGADO, J. A lipocentric view of peptide-induced pores. European Biophysics Journal 2011, 40, 399–415.
- GAO, Y., CHEN, Y., CAO, Y., MO, A. & PENG, Q. Potentials of nanotechnology in treatment of methicillin-resistant Staphylococcus aureus. European Journal of Medicinal Chemistry 2021, 213, 113056.
- GHARPURE, S., AKASH, A. & ANKAMWAR, B. A review on antimicrobial properties of metal nanoparticles. Journal of Nanoscience and Nanotechnology 2020, 20, 3303–3339.
- GHOSH, C., SARKAR, P., ISSA, R. & HALDAR, J. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends in Microbiology 2019, 27, 323–338.
- GIBB, B., HYMAN, P. & SCHNEIDER, C. L. The many applications of engineered bacteriophages—An overview. Pharmaceuticals 2021, 14, 634.
- GOŁAWSKA, S., SPRAWKA, I., ŁUKASIK, I. & GOŁAWSKI, A. Are naringenin and quercetin useful chemicals in pest-management strategies? Journal of pest science 2014, 87, 173–180.
- GONDIL, V. S., HARJAI, K. & CHHIBBER, S. Endolysins as emerging alternative therapeutic agents to counter drug-resistant infections. International Journal of Antimicrobial Agents 2020, 55, 105844.
- GOYAL, P., BELAPURKAR, P. & KAR, A. A review on in vitro and in vivo bioremediation potential of environmental and probiotic species of Bacillus and other probiotic microorganisms for two heavy metals, cadmium and nickel. Biosciences Biotechnology Research Asia 2019, 16, 01–13.
- GRASSI, L., MAISETTA, G., ESIN, S. & BATONI, G. Combination strategies to enhance the efficacy of antimicrobial peptides against bacterial biofilms. Frontiers in Microbiology 2017, 8, 2409.
- GUPTA, A., MUMTAZ, S., LI, C.-H., HUSSAIN, I. & ROTELLO, V. M. Combatting antibiotic-resistant bacteria using nanomaterials. Chemical Society Reviews 2019a, 48, 415–427.
- GUPTA, K., SINGH, S. P., MANHAR, A. K., SAIKIA, D., NAMSA, N. D., KONWAR, B. K. & MANDAL, M. Inhibition of Staphylococcus aureus and Pseudomonas aeruginosa biofilm and virulence by active fraction of Syzygium cumini (L.) Skeels leaf extract: in-vitro and in silico studies. Indian Journal of Microbiology, 2019b, 59, 13–21.
- HARBORNE, J. B. Classes and functions of secondary products from plants. Chemicals from Plants 1999, 26, 1–25.
- HARISH, K. & VARGHESE, T. Probiotics in humans–evidence based review. Calicut Med J, 2006, 4, e3.
- HELMY, Q., KARDENA, E. & GUSTIANI, S. 2019. Probiotics and bioremediation. Microorganisms. IntechOpen.
- HOF, W. V. T., VEERMAN, E. C., HELMERHORST, E. J. & AMERONGEN, A. V. N. Antimicrobial peptides: properties and applicability. Biological Chemistry 2001, 382, 597–619.
- HOFFMANN, D. 2003. Medical herbalism: the science and practice of herbal medicine, Simon and Schuster.
- HOFFMANN, S. A., MACULLOCH, B. & BATZ, M. 2015. Economic burden of major foodborne illnesses acquired in the United States. United States Department of Agriculture, Economic Research Service.
- HU, B., PAN, Y., LI, Z., YUAN, W. & DENG, L. EmPis-1L, an effective antimicrobial peptide against the antibiotic-resistant VBNC state cells of pathogenic bacteria. Probiotics and Antimicrobial Proteins 2019, 11, 667–675.
- HUAN, Y., KONG, Q., MOU, H. & YI, H. Antimicrobial peptides: classification, design, application and research progress in multiple fields. Frontiers in Microbiology 2020, 2559.
- HUANG, H. W. Action of antimicrobial peptides: two-state model. Biochemistry 2000, 39, 8347–8352. [CrossRef] [PubMed]
- HUDSON, J., BILLINGTON, C., PREMARATNE, A. & ON, S. L. Inactivation of Escherichia coli O157: H7 using ultraviolet light-treated bacteriophages. Food Science and Technology International 2016, 22, 3–9.
- HUMPHRIES, R. M., POLLETT, S. & SAKOULAS, G. A current perspective on daptomycin for the clinical microbiologist. Clinical Microbiology Reviews 2013, 26, 759–780.
- HUSSEIN, R. A. & EL-ANSSARY, A. A. Plants secondary metabolites: the key drivers of the pharmacological actions of medicinal plants. Herb. Med 2019, 1.
- IBRAHIM, R. A., CRYER, T. L., LAFI, S. Q., BASHA, E.-A., GOOD, L. & TARAZI, Y. H. Identification of Escherichia coli from broiler chickens in Jordan, their antimicrobial resistance, gene characterization and the associated risk factors. BMC Veterinary Research 2019, 15, 1–16.
- ISOLAURI, E., KIRJAVAINEN, P. & SALMINEN, S. Probiotics: a role in the treatment of intestinal infection and inflammation? Gut 2002, 50, iii54–iii59.
- JAGLAN, A. B., ANAND, T., VERMA, R., VASHISTH, M., VIRMANI, N., BERA, B. C., VAID, R. K. & TRIPATHI, B. N. Tracking the phage trends: A comprehensive review of applications in therapy and food production. Frontiers in Microbiology 2022, 13.
- JAHN, S., SEIWERT, B., KRETZING, S., ABRAHAM, G., REGENTHAL, R. & KARST, U. Metabolic studies of the Amaryllidaceous alkaloids galantamine and lycorine based on electrochemical simulation in addition to in vivo and in vitro models. Analytica Chimica Acta 2012, 756, 60–72.
- JAROW, J. P., LURIE, P., IKENBERRY, S. C. & LEMERY, S. Overview of FDA’s expanded access program for investigational drugs. Therapeutic Innovation & Regulatory Science 2017, 51, 177–179.
- JEPSON, R. G. Cranberries for preventing urinary tract infections. Sao Paulo Medical Journal 2013, 131, 363–363. [CrossRef] [PubMed]
- JIANG, H., CHENG, H., LIANG, Y., YU, S., YU, T., FANG, J. & ZHU, C. Diverse mobile genetic elements and conjugal transferability of sulfonamide resistance genes (sul1, sul2, and sul3) in Escherichia coli isolates from Penaeus vannamei and pork from large markets in Zhejiang, China. Frontiers in Microbiology 2019, 10, 1787.
- KANG, H.-K., KIM, C., SEO, C. H. & PARK, Y. The therapeutic applications of antimicrobial peptides (AMPs): a patent review. Journal of Microbiology 2017, 55, 1–12.
- KESAVELU, D., ROHIT, A., KARUNASAGAR, I. & KARUNASAGAR, I. Composition and laboratory correlation of commercial probiotics in India. Cureus 2020, 12, e11334.
- KIM, M. J., KU, S., KIM, S. Y., LEE, H. H., JIN, H., KANG, S., LI, R., JOHNSTON, T. V., PARK, M. S. & JI, G. E. Safety evaluations of Bifidobacterium bifidum BGN4 and Bifidobacterium longum BORI. International Journal of Molecular Sciences 2018, 19, 1422.
- KORF, I. H., MEIER-KOLTHOFF, J. P., ADRIAENSSENS, E. M., KROPINSKI, A. M., NIMTZ, M., ROHDE, M., VAN RAAIJ, M. J. & WITTMANN, J. Still something to discover: Novel insights into Escherichia coli phage diversity and taxonomy. Viruses 2019, 11, 454.
- LARSSON, D. & FLACH, C.-F. Antibiotic resistance in the environment. Nature Reviews Microbiology 2022, 20, 257–269.
- LE, C.-F., FANG, C.-M. & SEKARAN, S. D. Intracellular targeting mechanisms by antimicrobial peptides. Antimicrobial Agents and Chemotherapy 2017, 61, e02340–16.
- LE, T. S., SOUTHGATE, P. C., O’CONNOR, W., POOLE, S. & KURTBӦKE, D. I. Bacteriophages as biological control agents of enteric bacteria contaminating edible oysters. Current Microbiology 2018, 75, 611–619.
- LEE, N.-Y., KO, W.-C. & HSUEH, P.-R. Nanoparticles in the treatment of infections caused by multidrug-resistant organisms. Frontiers in Pharmacology 2019, 10, 1153.
- LEE, S., LEE, J., JIN, Y.-I., JEONG, J.-C., CHANG, Y. H., LEE, Y., JEONG, Y. & KIM, M. Probiotic characteristics of Bacillus strains isolated from Korean traditional soy sauce. LWT - Food Science and Technology 2017, 79, 518–524.
- LEI, J., SUN, L., HUANG, S., ZHU, C., LI, P., HE, J., MACKEY, V., COY, D. H. & HE, Q. The antimicrobial peptides and their potential clinical applications. American Journal of Translational Research 2019, 11, 3919.
- LI, W., O’BRIEN-SIMPSON, N. M., HOLDEN, J. A., OTVOS, L., REYNOLDS, E. C., SEPAROVIC, F., HOSSAIN, M. A. & WADE, J. D. Covalent conjugation of cationic antimicrobial peptides with a β-lactam antibiotic core. Peptide Science 2018, 110, e24059.
- LI, W., TAILHADES, J., O’BRIEN-SIMPSON, N. M., SEPAROVIC, F., OTVOS, L., HOSSAIN, M. A. & WADE, J. D. Proline-rich antimicrobial peptides: potential therapeutics against antibiotic-resistant bacteria. Amino acids 2014, 46, 2287–2294.
- LI, Z., LI, X., ZHANG, J., WANG, X., WANG, L., CAO, Z. & XU, Y. Use of phages to control Vibrio splendidus infection in the juvenile sea cucumber Apostichopus japonicus. Fish & Shellfish Immunology 2016, 54, 302–311.
- ŁOBODA, D., KOZŁOWSKI, H. & ROWIŃSKA-ŻYREK, M. Antimicrobial peptide–metal ion interactions–a potential way of activity enhancement. New Journal of Chemistry 2018, 42, 7560–7568.
- ŁOJEWSKA, E. & SAKOWICZ, T. An Alternative to Antibiotics: Selected Methods to Combat Zoonotic Foodborne Bacterial Infections. Current Microbiology 2021, 78, 4037–4049.
- LUO, X., LIAO, G., LIU, C., JIANG, X., LIN, M., ZHAO, C., TAO, J. & HUANG, Z. Characterization of bacteriophage HN 48 and its protective effects in Nile tilapia Oreochromis niloticus against Streptococcus agalactiae infections. Journal of Fish Diseases 2018, 41, 1477–1484.
- MANOHAR, P., LOH, B. & LEPTIHN, S. Will the overuse of antibiotics during the Coronavirus pandemic accelerate antimicrobial resistance of bacteria? Infectious Microbes and Diseases 2020, 2, 87–88.
- MARTINEZ, S. R., IBARRA, L. E., PONZIO, R. A., FORCONE, M. V., WENDEL, A. B., CHESTA, C. A., SPESIA, M. B. & PALACIOS, R. E. Photodynamic inactivation of ESKAPE group bacterial pathogens in planktonic and biofilm cultures using metallated porphyrin-doped conjugated polymer nanoparticles. ACS Infectious Diseases 2020, 6, 2202–2213.
- MASOTTI, V., JUTEAU, F., BESSIÈRE, J. M. & VIANO, J. Seasonal and phenological variations of the essential oil from the narrow endemic species Artemisia molinieri and its biological activities. Journal of agricultural and food chemistry 2003, 51, 7115–7121.
- MBA, I. E. & NWEZE, E. I. Nanoparticles as therapeutic options for treating multidrug-resistant bacteria: Research progress, challenges, and prospects. World Journal of Microbiology and Biotechnology 2021, 37, 1–30.
- MCGAW, L. J., FAMUYIDE, I. M., KHUNOANA, E. T. & AREMU, A. O. Ethnoveterinary botanical medicine in South Africa: A review of research from the last decade (2009 to 2019). Journal of Ethnopharmacology 2020, 257, 112864.
- MCLINDEN, T., SARGEANT, J. M., THOMAS, M. K., PAPADOPOULOS, A. & FAZIL, A. Component costs of foodborne illness: a scoping review. BMC Public Health 2014, 14, 1–12.
- MEILE, S., KILCHER, S., LOESSNER, M. J. & DUNNE, M. Reporter phage-based detection of bacterial pathogens: Design guidelines and recent developments. Viruses 2020, 12, 944.
- MITEVA, M., ANDERSSON, M., KARSHIKOFF, A. & OTTING, G. Molecular electroporation: a unifying concept for the description of membrane pore formation by antibacterial peptides, exemplified with NK-lysin. FEBS Letters 1999, 462, 155–158.
- MITRA, D., KANG, E.-T. & NEOH, K. G. Polymer-based coatings with integrated antifouling and bactericidal properties for targeted biomedical applications. ACS Applied Polymer Materials 2021, 3, 2233–2263.
- MOHAMED, F. M., THABET, M. H. & ALI, M. F. The use of probiotics to enhance immunity of broiler chicken against some intestinal infection pathogens. SVU-International Journal of Veterinary Sciences 2019, 2, 1–19.
- MOHAMMED, I., SAID, D. G. & DUA, H. S. Human antimicrobial peptides in ocular surface defense. Progress in Retinal and Eye Research 2017, 61, 1–22.
- MOLCHANOVA, N., HANSEN, P. R. & FRANZYK, H. Advances in development of antimicrobial peptidomimetics as potential drugs. Molecules 2017, 22, 1430.
- MONTANHER, A. B., ZUCOLOTTO, S. M., SCHENKEL, E. P. & FRÖDE, T. S. Evidence of anti-inflammatory effects of Passiflora edulis in an inflammation model. Journal of Ethnopharmacology 2007, 109, 281–288.
- MONTSO, P. K., MLAMBO, V. & ATEBA, C. N. Characterization of lytic bacteriophages infecting multidrug-resistant shiga toxigenic atypical Escherichia coli O177 strains isolated from cattle feces. Frontiers in Public Health 2019, 7, 355.
- MONTSO, P. K., MNISI, C. M., ATEBA, C. N. & MLAMBO, V. An assessment of the viability of lytic phages and their potency against multidrug resistant Escherichia coli O177 strains under simulated rumen fermentation conditions. Antibiotics 2021, 10, 265.
- MÖSGES, R., BAUES, C., SCHRÖDER, T. & SAHIN, K. Acute bacterial otitis externa: efficacy and safety of topical treatment with an antibiotic ear drop formulation in comparison to glycerol treatment. Current Medical Research and Opinion 2011, 27, 871–878.
- MOSTOWY, S. Louis Pasteur continues to shape the future of microbiology. Disease Models and Mechanisms 2022, 15, dmm050011. [CrossRef] [PubMed]
- MUBEEN, B., ANSAR, A. N., RASOOL, R., ULLAH, I., IMAM, S. S., ALSHEHRI, S., GHONEIM, M. M., ALZAREA, S. I., NADEEM, M. S. & KAZMI, I. Nanotechnology as a novel approach in combating microbes providing an alternative to antibiotics. Antibiotics 2021, 10, 1473.
- MULANI, M. S., KAMBLE, E. E., KUMKAR, S. N., TAWRE, M. S. & PARDESI, K. R. Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: a review. Frontiers in Microbiology 2019, 10, 539.
- MURUGAIYAN, J., KUMAR, P. A., RAO, G. S., ISKANDAR, K., HAWSER, S., HAYS, J. P., MOHSEN, Y., ADUKKADUKKAM, S., AWUAH, W. A. & JOSE, R. A. M. Progress in alternative strategies to combat antimicrobial resistance: Focus on antibiotics. Antibiotics 2022, 11, 200.
- MWINGA, J. L., OTANG-MBENG, W., KUBHEKA, B. P. & AREMU, A. O. Ethnobotanical survey of plants used by subsistence farmers in mitigating cabbage and spinach diseases in OR Tambo Municipality, South Africa. Plants 2022, 11, 3215.
- NANDA, A. M., THORMANN, K. & FRUNZKE, J. Impact of spontaneous prophage induction on the fitness of bacterial populations and host-microbe interactions. Journal of Bacteriology 2015, 197, 410–419.
- NGUYEN, F., STAROSTA, A. L., ARENZ, S., SOHMEN, D., DÖNHÖFER, A. & WILSON, D. N. Tetracycline antibiotics and resistance mechanisms. Biological Chemistry 2014, 395, 559–575.
- NGUYEN, L. T., HANEY, E. F. & VOGEL, H. J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends in Biotechnology 2011, 29, 464–472.
- NIEMEYER, J. C., LOLATA, G. B., DE CARVALHO, G. M., DA SILVA, E. M., SOUSA, J. P. & NOGUEIRA, M. A. Microbial indicators of soil health as tools for ecological risk assessment of a metal contaminated site in Brazil. Applied Soil Ecology 2012, 59, 96–105.
- NIKOLICH, M. P. & FILIPPOV, A. A. Bacteriophage therapy: developments and directions. Antibiotics 2020, 9, 135.
- OUWEHAND, A. C., FORSSTEN, S., HIBBERD, A. A., LYRA, A. & STAHL, B. Probiotic approach to prevent antibiotic resistance. Annals of Medicine 2016, 48, 246–255.
- PALMA, E., TILOCCA, B. & RONCADA, P. Antimicrobial resistance in veterinary medicine: An overview. International Journal of Molecular Sciences 2020, 21, 1914.
- PARK, K. H., OH, S.-Y., YOO, S., FONG, J. J., KIM, C. S., JO, J. W. & LIM, Y. W. Influence of season and soil properties on fungal communities of neighboring climax forests (Carpinus cordata and Fraxinus rhynchophylla). Frontiers in Microbiology 2020, 2643.
- PASUPULETI, M., SCHMIDTCHEN, A. & MALMSTEN, M. Antimicrobial peptides: key components of the innate immune system. Critical Reviews in Biotechnology 2012, 32, 143–171.
- PATEL, P. H. & HASHMI, M. F. 2021. Macrolides. StatPearls StatPearls Publishing, Tampa, FL, USA.
- PELCZAR, M., CHAN, E. & KRIEG, N. Control of microorganisms, the control of microorganisms by physical agents. Microbiology 1988, 469, 509.
- PELFRENE, E., BOTGROS, R. & CAVALERI, M. Antimicrobial multidrug resistance in the era of COVID-19: a forgotten plight? Antimicrobial Resistance & Infection Control 2021, 10, 1–6.
- PETERSON, E. & KAUR, P. Antibiotic resistance mechanisms in bacteria: relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Frontiers in Microbiology 2018, 9, 2928.
- PETSONG, K., BENJAKUL, S., CHATURONGAKUL, S., SWITT, A. I. M. & VONGKAMJAN, K. Lysis profiles of Salmonella phages on Salmonella isolates from various sources and efficiency of a phage cocktail against S. enteritidis and S. typhimurium. Microorganisms 2019, 7, 100.
- PINTO, G., ALMEIDA, C. & AZEREDO, J. Bacteriophages to control shiga toxin-producing E. coli–safety and regulatory challenges. Critical Reviews in Biotechnology 2020, 40, 1081–1097.
- PIRNAY, J.-P., VERBEKEN, G., CEYSSENS, P.-J., HUYS, I., DE VOS, D., AMELOOT, C. & FAUCONNIER, A. The magistral phage. Viruses 2018, 10, 64.
- PLESSAS, S., NOUSKA, C., KARAPETSAS, A., KAZAKOS, S., ALEXOPOULOS, A., MANTZOURANI, I., CHONDROU, P., FOURNOMITI, M., GALANIS, A. & BEZIRTZOGLOU, E. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from feta-type cheese. Food Chemistry 2017, 226, 102–108.
- POKORNY, A., BIRKBECK, T. H. & ALMEIDA, P. F. Mechanism and kinetics of δ-lysin interaction with phospholipid vesicles. Biochemistry 2002, 41, 11044–11056.
- PRADO, M., BLANDÓN, L., VANDENBERGHE, L., RODRIGUES, C., CASTRO, G., THOMAZ-SOCCOL, V. & SOCCOL, C. Milk kefir: composition, microbial cultures, biological activities, and related products. Frontiers in Microbiology 2015, 30, 1177.
- PRASHER, P., SINGH, M. & MUDILA, H. Silver nanoparticles as antimicrobial therapeutics: current perspectives and future challenges. Biotech 2018, 8, 1–23.
- RABANAL, F. & CAJAL, Y. Recent advances and perspectives in the design and development of polymyxins. Natural Product Reports 2017, 34, 886–908.
- RAMIREZ, K., CAZAREZ-MONTOYA, C., LOPEZ-MORENO, H. S. & CASTRO-DEL CAMPO, N. Bacteriophage cocktail for biocontrol of Escherichia coli O157: H7: Stability and potential allergenicity study. PLoS One 2018, 13, e0195023.
- REINHARDT, A. & NEUNDORF, I. Design and application of antimicrobial peptide conjugates. International Journal of Molecular Sciences 2016, 17, 701.
- RIBER, C. F., SMITH, A. A. & ZELIKIN, A. N. Self-Immolative linkers literally bridge disulfide chemistry and the realm of Thiol-free drugs. Advanced Healthcare Materials 2015, 4, 1887–1890.
- RIMA, M., RIMA, M., FAJLOUN, Z., SABATIER, J.-M., BECHINGER, B. & NAAS, T. Antimicrobial peptides: A potent alternative to antibiotics. Antibiotics 2021, 10, 1095.
- RINGØ, E. Probiotics in shellfish aquaculture. Aquaculture and Fisheries 2020, 5, 1–27. [CrossRef]
- RITSEMA, J. A., DER WEIDE, H. V., TE WELSCHER, Y. M., GOESSENS, W. H., VAN NOSTRUM, C. F., STORM, G., BAKKER-WOUDENBERG, I. A. & HAYS, J. P. Antibiotic-nanomedicines: facing the challenge of effective treatment of antibiotic-resistant respiratory tract infections. Future Microbiology 2018, 13, 1683–1692.
- ROBBINS, T., BU, D. & CARRIQUE-MAS, J. F evre. EM, Gilbert, M., Grace, D., Hay, SI, Jiwakanon, J., Kakkar, M., Kariuki, S., Laxminarayan, R., Lubroth, J., Magnusson, U., Thi Ngoc, P., van Bockel, TP and Woolhouse. MEJ 2016, 1–3.
- RUDRAMURTHY, G. R., SWAMY, M. K., SINNIAH, U. R. & GHASEMZADEH, A. Nanoparticles: alternatives against drug-resistant pathogenic microbes. Molecules 2016, 21, 836.
- SAN MILLAN, A. Evolution of plasmid-mediated antibiotic resistance in the clinical context. Trends in Microbiology 2018, 26, 978–985. [CrossRef]
- SÁNCHEZ-LÓPEZ, E., GOMES, D., ESTERUELAS, G., BONILLA, L., LOPEZ-MACHADO, A. L., GALINDO, R., CANO, A., ESPINA, M., ETTCHETO, M. & CAMINS, A. Metal-based nanoparticles as antimicrobial agents: an overview. Nanomaterials 2020, 10, 292.
- SEIGLER, D. S. 1998. Indole Alkaloids. Plant Secondary Metabolism. Springer.
- SHAI, Y. & OREN, Z. From “carpet” mechanism to de-novo designed diastereomeric cell-selective antimicrobial peptides. Peptides 2001, 22, 1629–1641.
- SHEARD, D. E., O’BRIEN-SIMPSON, N. M., WADE, J. D. & SEPAROVIC, F. Combating bacterial resistance by combination of antibiotics with antimicrobial peptides. Pure and Applied Chemistry 2019, 91, 199–209.
- SHIN, S. W., SHIN, M. K., JUNG, M., BELAYNEHE, K. M. & YOO, H. S. Prevalence of antimicrobial resistance and transfer of tetracycline resistance genes in Escherichia coli isolates from beef cattle. Applied and Environmental Microbiology 2015, 81, 5560–5566.
- SHOUSHA, A., AWAIWANONT, N., SOFKA, D., SMULDERS, F. J., PAULSEN, P., SZOSTAK, M. P., HUMPHREY, T. & HILBERT, F. Bacteriophages isolated from chicken meat and the horizontal transfer of antimicrobial resistance genes. Applied and Environmental Microbiology 2015, 81, 4600–4606.
- SIERRA, J. M., FUSTÉ, E., RABANAL, F., VINUESA, T. & VIÑAS, M. An overview of antimicrobial peptides and the latest advances in their development. Expert Opinion on Biological Therapy 2017, 17, 663–676.
- SILLANKORVA, S. M., OLIVEIRA, H. & AZEREDO, J. Bacteriophages and their role in food safety. Bacteriophages and their role in food safety. International Journal of Microbiology 2012, 2012.
- SINGH, A. P., BISWAS, A., SHUKLA, A. & MAITI, P. Targeted therapy in chronic diseases using nanomaterial-based drug delivery vehicles. Signal Transduction and Targeted Therapy 2019, 4, 1–21.
- SLAVIN, Y. N., ASNIS, J., HÄFELI, U. O. & BACH, H. Metal nanoparticles: understanding the mechanisms behind antibacterial activity. Journal of Nanobiotechnology 2017, 15, 1–20.
- SOLOMON, S. L. & OLIVER, K. B. Antibiotic resistance threats in the United States: stepping back from the brink. American Family Physician 2014, 89, 938–941.
- SPILLER, F., ALVES, M. K., VIEIRA, S. M., CARVALHO, T. A., LEITE, C. E., LUNARDELLI, A., POLONI, J. A., CUNHA, F. Q. & DE OLIVEIRA, J. R. Anti-inflammatory effects of red pepper (Capsicum baccatum) on carrageenan-and antigen-induced inflammation. Journal of Pharmacy and Pharmacology 2008, 60, 473–478.
- SPIRESCU, V. A., CHIRCOV, C., GRUMEZESCU, A. M. & ANDRONESCU, E. Polymeric nanoparticles for antimicrobial therapies: An up-to-date overview. Polymers 2021, 13, 724.
- STEVENSON, C. L. Advances in peptide pharmaceuticals. Current Pharmaceutical Biotechnology 2009, 10, 122–137. [CrossRef] [PubMed]
- SUBRAMANIAM, S., FAHY, E., GUPTA, S., SUD, M., BYRNES, R. W., COTTER, D., DINASARAPU, A. R. & MAURYA, M. R. Bioinformatics and systems biology of the lipidome. Chemical reviews 2011, 111, 6452–6490.
- SUMI, C. D., YANG, B. W., YEO, I.-C. & HAHM, Y. T. Antimicrobial peptides of the genus Bacillus: a new era for antibiotics. Canadian Journal of Microbiology 2015, 61, 93–103.
- SURESH, K., KRISHNAPPA, S. & BHARDWAJ, P. Safety concerns of Probiotic use: a review. Safety 2013, 34, 40.
- TANG, R., YU, H., QI, M., YUAN, X., RUAN, Z., HU, C., XIAO, M., XUE, Y., YAO, Y. & LIU, Q. Biotransformation of citrus fruits phenolic profiles by mixed probiotics in vitro anaerobic fermentation. LWT 2022, 160, 113087.
- TILOCCA, B., BALMAS, V., HASSAN, Z. U., JAOUA, S. & MIGHELI, Q. A proteomic investigation of Aspergillus carbonarius exposed to yeast volatilome or to its major component 2-phenylethanol reveals major shifts in fungal metabolism. International Journal of Food Microbiology 2019, 306, 108265.
- TOMAT, D., CASABONNE, C., AQUILI, V., BALAGUÉ, C. & QUIBERONI, A. Evaluation of a novel cocktail of six lytic bacteriophages against shiga toxin-producing Escherichia coli in broth, milk and meat. Food Microbiology 2018, 76, 434–442.
- TOOKE, C. L., HINCHLIFFE, P., BRAGGINTON, E. C., COLENSO, C. K., HIRVONEN, V. H., TAKEBAYASHI, Y. & SPENCER, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. Journal of Molecular Biology 2019, 431, 3472–3500.
- TORCATO, I. M., HUANG, Y.-H., FRANQUELIM, H. G., GASPAR, D., CRAIK, D. J., CASTANHO, M. A. & HENRIQUES, S. T. Design and characterization of novel antimicrobial peptides, R-BP100 and RW-BP100, with activity against Gram-negative and Gram-positive bacteria. Biochimica et Biophysica Acta (BBA)-Biomembranes 2013, 1828, 944–955.
- VALLABANI, N. & SINGH, S. Recent advances and future prospects of iron oxide nanoparticles in biomedicine and diagnostics. 3 Biotech 2018, 8, 1–23.
- VANHEE, L. M., GOEMÉ, F., NELIS, H. J. & COENYE, T. Quality control of fifteen probiotic products containing Saccharomyces boulardii. Journal of Applied Microbiology 2010, 109, 1745–1752.
- VANKERCKHOVEN, V., HUYS, G., VANCANNEYT, M., VAEL, C., KLARE, I., ROMOND, M.-B., ENTENZA, J. M., MOREILLON, P., WIND, R. D., KNOL, J., WIERTZ, E., POT, B., VAUGHAN, E. E., KAHLMETER, G. & GOOSSENS, H. Biosafety assessment of probiotics used for human consumption: recommendations from the EU-PROSAFE project. Trends in Food Science & Technology 2008, 19, 102–114.
- VENTOLA, C. L. Progress in nanomedicine: approved and investigational nanodrugs. Pharmacy and Therapeutics 2017, 42, 742.
- VIAZIS, S., AKHTAR, M., FEIRTAG, J. & DIEZ-GONZALEZ, F. Reduction of Escherichia coli O157: H7 viability on hard surfaces by treatment with a bacteriophage mixture. International Journal of Food Microbiology 2011, 145, 37–42.
- VIAZIS, S. & DIEZ-GONZALEZ, F. Enterohemorrhagic Escherichia coli: the twentieth century’s emerging foodborne pathogen: a review. Advances in Agronomy 2011, 111, 1–50.
- WANG, J., DOU, X., SONG, J., LYU, Y., ZHU, X., XU, L., LI, W. & SHAN, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Medicinal Research Reviews 2019, 39, 831–859.
- WEBB, G. P. An overview of dietary supplements and functional foods. Dietary Supplements and Functional Foods 2011, 1–51.
- WENDLANDT, S., FEßLER, A. T., MONECKE, S., EHRICHT, R., SCHWARZ, S. & KADLEC, K. The diversity of antimicrobial resistance genes among staphylococci of animal origin. International Journal of Medical Microbiology 2013, 303, 338–349.
- WHO. 1991. Programme on Traditional Medicine. Guidelines for the assessment of herbal medicines. World Health Organization. [Online]. Available: https://apps.who.int/iris/handle/10665/58865 [Accessed December 26, 2022].
- WHO. 2013. WHO traditional medicine strategy 2014–2023. Altern Integr Med: 1-78 [Online]. Available: http://www.who.int/medicines/publications/traditional/trm_strategy14_23/en/ [Accessed December 26, 2022].
- WHO 2015. Global Action Plan on Antimicrobial Resistance. WHO, Geneva, Switzerland.
- WHO. 2022. The fight against Antimicrobial Resistance is closely linked to the Sustainable Development Goals. [Online]. Available: https://apps.who.int/iris/bitstream/handle/10665/337519/WHO-EURO-2020-1634-41385-56394-eng.pdf [Accessed December 26, 2022].
- WIMLEY, W. C. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chemical Biology 2010, 5, 905–917. [CrossRef] [PubMed]
- WU, J., HU, Y., DU, C., PIAO, J., YANG, L. & YANG, X. The effect of recombinant human lactoferrin from the milk of transgenic cows on Salmonella enterica serovar typhimurium infection in mice. Food & Function 2016, 7, 308–314.
- WU, M., MAIER, E., BENZ, R. & HANCOCK, R. E. Mechanism of interaction of different classes of cationic antimicrobial peptides with planar bilayers and with the cytoplasmic membrane of Escherichia coli. Biochemistry 1999, 38, 7235–7242.
- WU, N., DAI, J., GUO, M., LI, J., ZHOU, X., LI, F., GAO, Y., QU, H., LU, H. & JIN, J. Pre-optimized phage therapy on secondary Acinetobacter baumannii infection in four critical COVID-19 patients. Emerging Microbes & Infections 2021, 10, 612–618.
- WU, W.-H., DI, R. & MATTHEWS, K. R. Activity of the plant-derived peptide Ib-AMP1 and the control of enteric foodborne pathogens. Food Control 2013, 33, 142–147.
- XIN, Q., SHAH, H., NAWAZ, A., XIE, W., AKRAM, M. Z., BATOOL, A., TIAN, L., JAN, S. U., BODDULA, R. & GUO, B. Antibacterial carbon-based nanomaterials. Advanced Materials 2019, 31, 1804838.
- XU, L., ZHAO, X.-Y., WU, Y.-L. & ZHANG, W. The study on biological and pharmacological activity of coumarins. 2015 Asia-Pacific Energy Equipment Engineering Research Conference, 2015. Atlantis Press, 135-138.
- YARNELL, E. Botanical medicines for the urinary tract. World Journal of Urology 2002, 20, 285–293. [CrossRef] [PubMed]
- YASIR, M., WILLCOX, M. D. P. & DUTTA, D. Action of antimicrobial peptides against bacterial biofilms. Materials 2018, 11, 2468.
- YEAMAN, M. R. & YOUNT, N. Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacological Reviews 2003, 55, 27–55.
- YEON, K.-M., YOU, J., ADHIKARI, M. D., HONG, S.-G., LEE, I., KIM, H. S., KIM, L. N., NAM, J., KWON, S.-J. & KIM, M. I. Enzyme-immobilized chitosan nanoparticles as environmentally friendly and highly effective antimicrobial agents. Biomacromolecules 2019, 20, 2477–2485.
- ZAWISTOWSKA-ROJEK, A., ZAREBA, T., MRÓWKA, A. & TYSKI, S. Assessment of the microbiological status of probiotic products. Polish Journal of Microbiology 2016, 65, 97–104.
- ZHARKOVA, M. S., ORLOV, D. S., GOLUBEVA, O. Y., CHAKCHIR, O. B., ELISEEV, I. E., GRINCHUK, T. M. & SHAMOVA, O. V. Application of antimicrobial peptides of the innate immune system in combination with conventional antibiotics—a novel way to combat antibiotic resistance? Frontiers in Cellular and Infection Microbiology 2019, 9, 128.
Figure 1.
Transmission routes of AMR bacteria between human, animal and the environment Adapted from: (Ajose et al., 2022b).
Figure 1.
Transmission routes of AMR bacteria between human, animal and the environment Adapted from: (Ajose et al., 2022b).
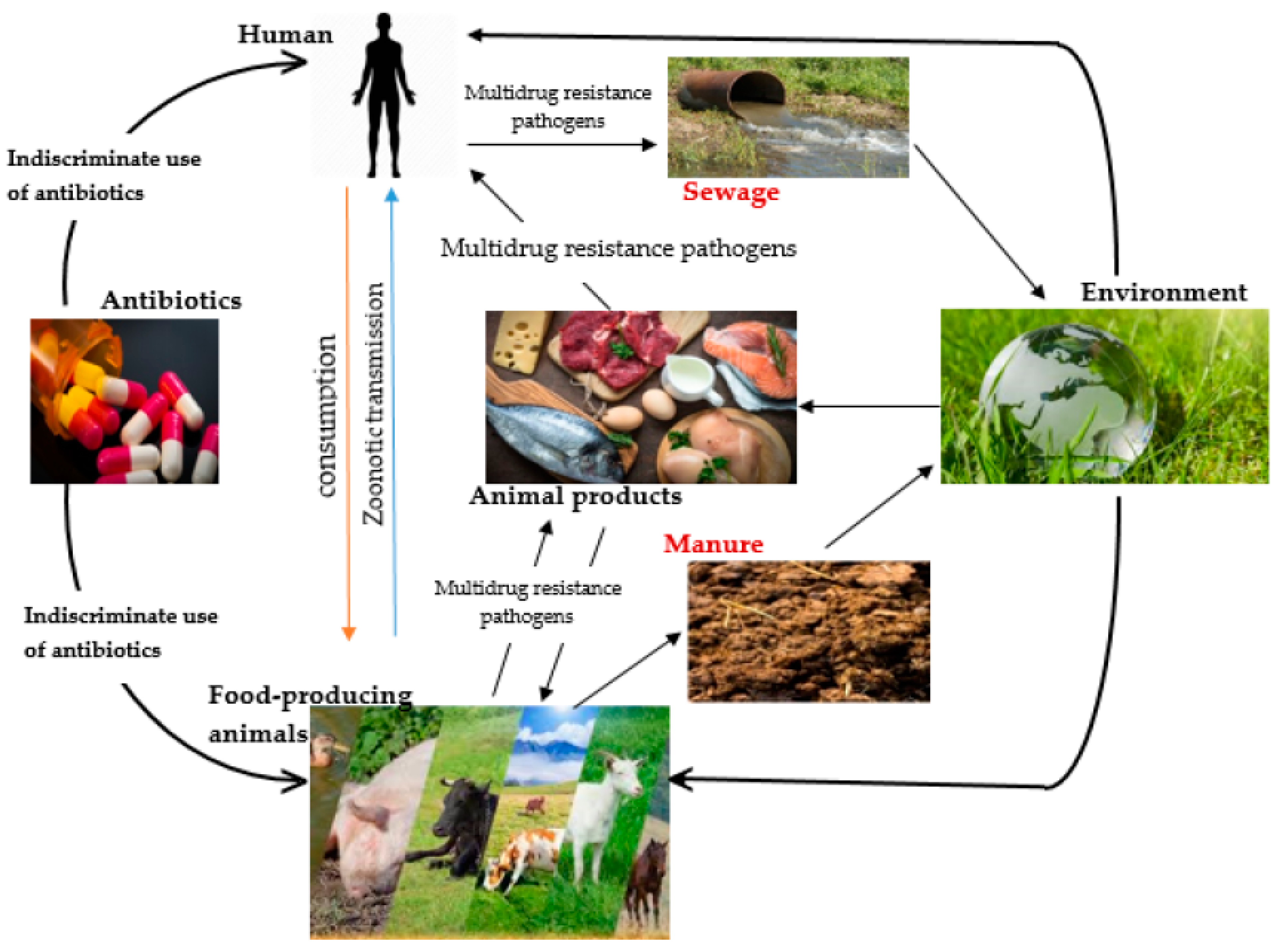
Figure 2.
Variables that contribute to antibiotic resistance.
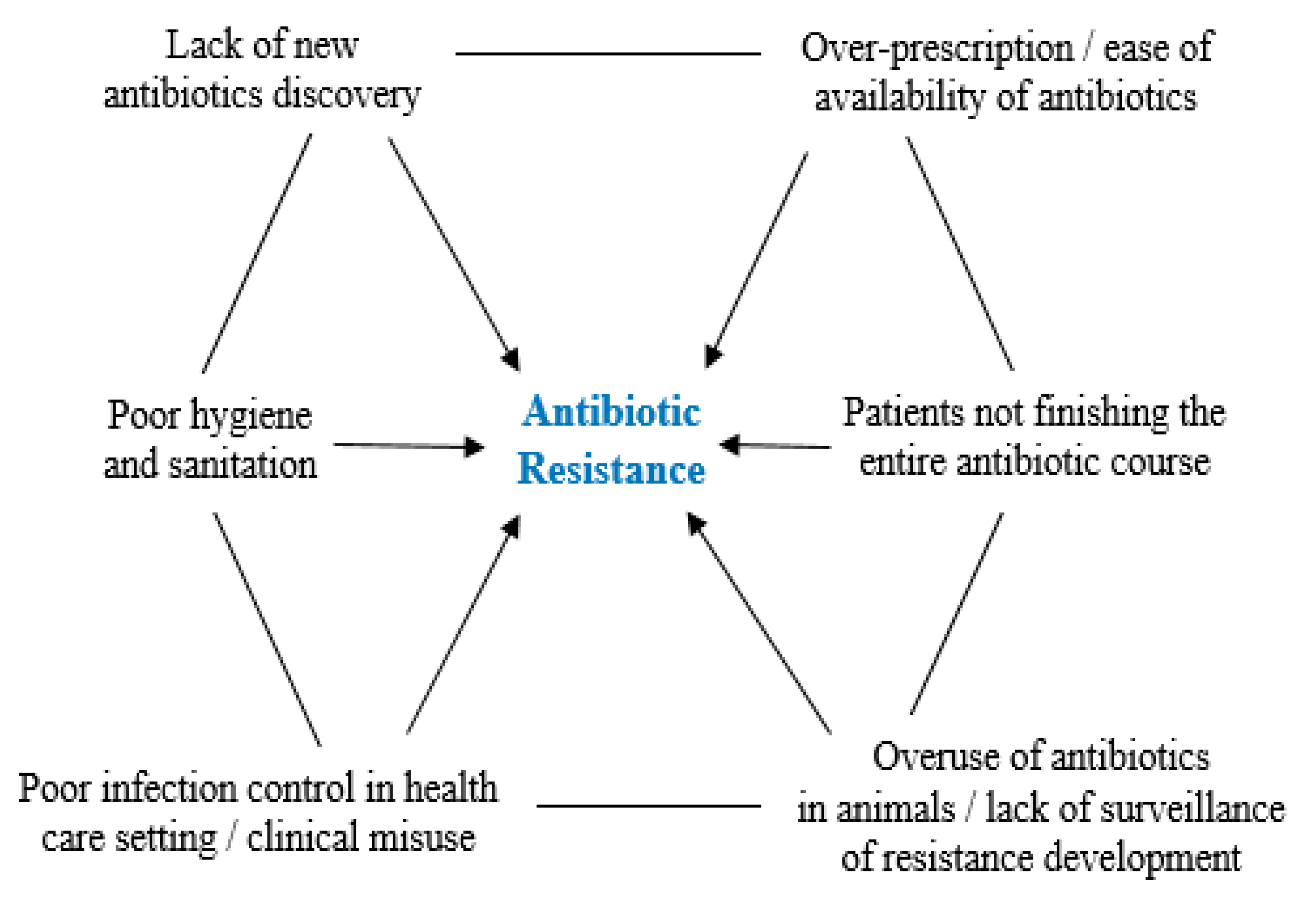
Figure 3.
Timeline showing the discovery of antibiotics and the rise of antibiotic resistance over the course of eight decades. XDR – Extensive-drug resistant, PDR – Pan-drug resistant.
Figure 3.
Timeline showing the discovery of antibiotics and the rise of antibiotic resistance over the course of eight decades. XDR – Extensive-drug resistant, PDR – Pan-drug resistant.
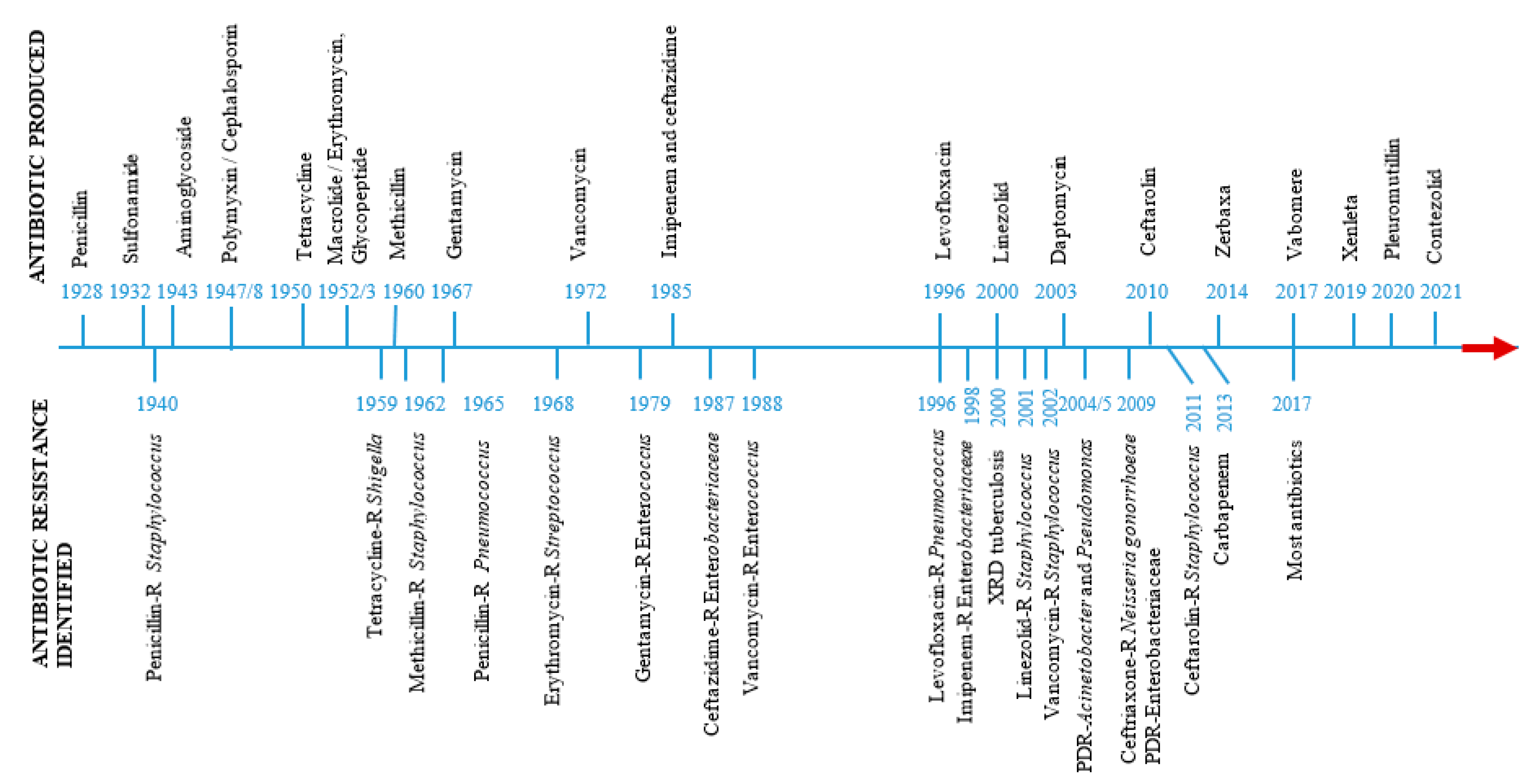
Figure 4.
Mechanisms of antimicrobial resistance in bacteria cells.
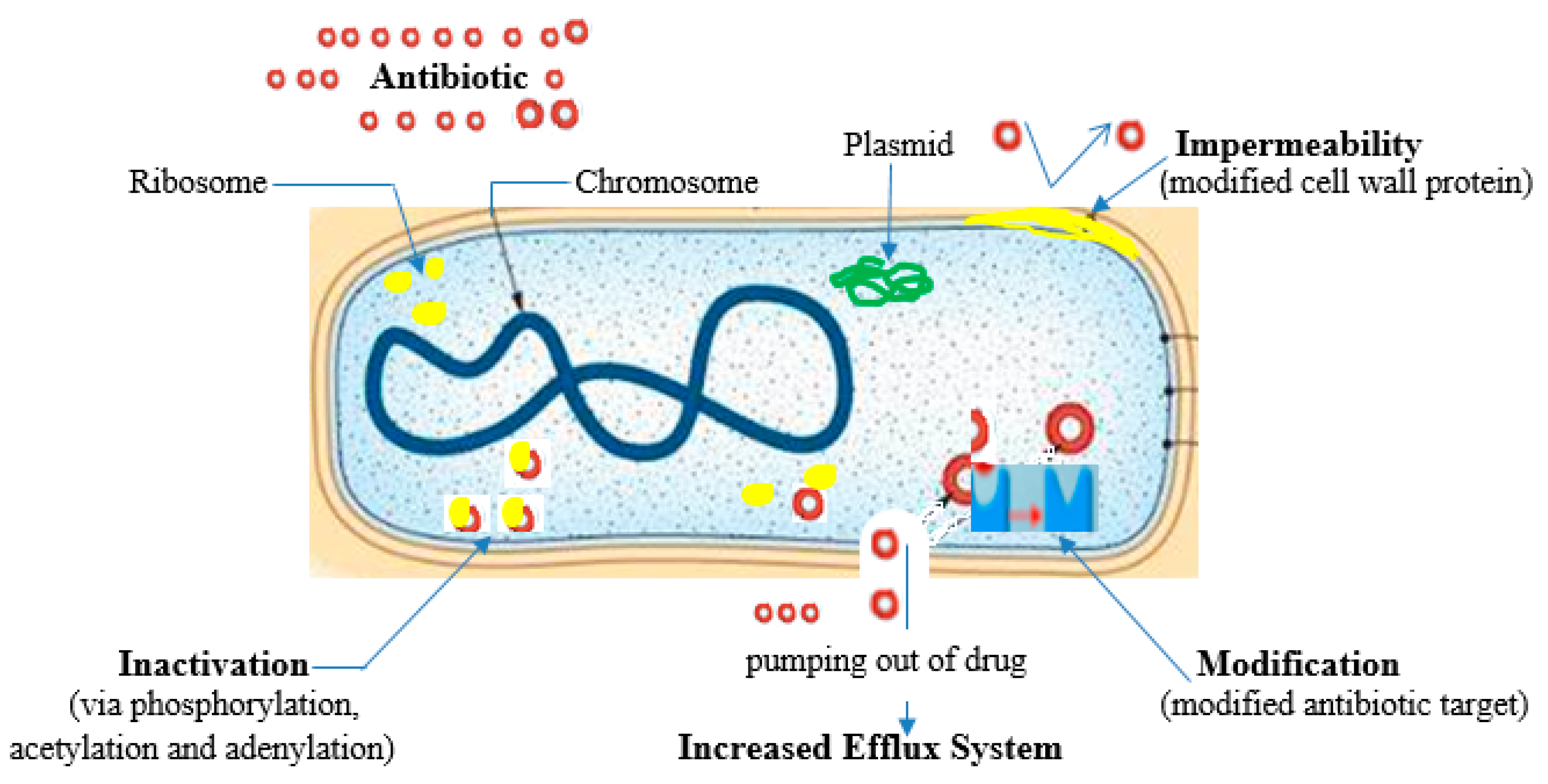
Figure 5.
Antimicrobial peptides mechanisms of action.


Table 1.
Mode of action of antibiotics and mechanisms of bacterial antibiotic resistance.
| Antibiotic family | Mode of action | Mechanism of resistance | Reference |
| β-lactams β-lactamase inhibitors Fluoroquinolones Macrolides, Lincosamides and Streptogamin (MLS) Aminoglycosides Tetracycline Sulfonamides (Folate pathway inhibitors) |
Cell wall synthesis inhibitors. Binds trans peptidase also known as penicillin binding proteins (PBPs) that help form peptidoglycan Inactivates the enzyme; beta-lactamase Hydrolysis of the beta-lactam ring Binds DNA-gyrase or topoisomerase II and topoisomerase IV; enzymes needed for supercoiling, replication and separation of circular bacterial DNA. Binds the bacterial 50S ribosomal subunits; inhibit protein synthesis Bind to the bacterial 30S ribosomal subunit thus inhibit bacterial protein synthesis Bind reversibly to the 30S ribosomal subunit as such blocks the binding of the aminoacyl-tRNA to the acceptor site on the mRNA-ribosome complex Inhibit the bacterial enzyme dihydropteroate synthetase (DPS) in the folic acid pathway, thereby blocking bacterial nucleic acid synthesis |
Beta-lactamase production primarily - bla genes Expression of alternative PBPs Production of extended spectrum beta-lactamases (ESBLs) Target modification Decreased membrane permeability Efflux pumps Target site modification Active drug efflux Target site modification (via the action of 16S rRNA methyltransferases (RMTs)) Enzymatic Drug Modification (adenylation, acetylation and phosphorylation), Efflux systems Efflux systems, Target modification, Inactivating enzymes, Ribosomal protection Excessive bacterial production of dihydrofolate reductase (DHFR) Reduction in the ability of the drug to penetrate the bacterial cell wall Production of altered forms of the dihydropteroate synthetase (DPS) enzyme with a lower affinity for sulfonamides Hyperproduction of para-amino benzoic acid (PABA), which overcomes the competitive substitution of the sulfonamides |
(Dowling et al., 2017, Tooke et al., 2019, Ibrahim et al., 2019) (Correia et al., 2017) (Patel and Hashmi, 2021) (Wendlandt et al., 2013) (Nguyen et al., 2014, Shin et al., 2015) (Davis, 2018, Jiang et al., 2019) |
Adapted from: (Ajose et al., 2022b).
Table 2.
Bacteriophages as bio-control agents against food-borne (food products and food contact surface) pathogens.
Table 2.
Bacteriophages as bio-control agents against food-borne (food products and food contact surface) pathogens.
| Phages | Sample | Technique | Outcome | Reference |
| BEC8 cocktail (38, 39, 41, CEV2, AR1, 42, ECA1, and ECB7) A cocktail composed by the phages e11/2 and e4/1c BEC8 cocktail Cocktail compose by phages DT1 to DT6 FAHEc1 phiEco1, phiEco2, phiEco3, phiEco5, phiEco6 and phiS1 Phages phiJLA23, phiKP26, phiC119 and phiE142 AKH-2 PVS-1, and PVS-2, PVS-3 HN48 |
Spinach leaves and romaine lettuce Cattle hide Sterilized hard surfaces (stainless steel chips, ceramic tile chips, and high density polyethylene chips - HDPEC). Milk and meat UHT milk; Ready-to-eat meat; Raw beef Oyster Tomatoes Misgurnus anguillicaudatus Apostichopus japonicas (Sea cucumber) Oreochromis niloticus (Nile tilapia) |
Following a one-hour drying period in a biosafety enclosure, the leaves were spot-inoculated with bacteria. On top of the previously inoculated leaf, BEC8, Trans-cinnamaldehyde, or TSB was administered. Without dehydrating the bacterial inoculum, positive controls were generated by combining it with BEC8 or TC. The phage cocktail was introduced into the organism using a portable spray container. Negative control: An absence of wash treatment was observed. The semiconductor was spot-treated with bacteria before being dried in a biosafety cabinet. Prior to inoculation, the chip surface was treated with BEC8 or TSB. MOIs of 1, 10, and 100 were utilised. To generate positive controls, the bacterial inoculum was combined with BEC8 or TC without the process of dehydrating Sterile, commercially available milk that had been reconstituted with CaCl2 was used to inoculate one bacterial strain per batch. One portion of each batch was subjected to a phage cocktail, while the other was set aside as a control. 0.4 cm thick, 1 cm2 portions of meat were spot-treated with bacterial strains and left to adhere for 10 minutes at room temperature. Following this, a phage cocktail was introduced into every meat piece. In order to establish controls, TMG buffer was added. Before being applied to food products, phage FAHEc1 was exposed to ultraviolet radiation; phages are capable of lysing bacterial cells, even if they lose viability. Phages and E. coli O157:H7 were utilised to inoculate UHT milk. Inoculating raw beef at 37 °C simulated phage application immediately prior to slathering in carcasses At 37 °C, bacteria that had been grown overnight were introduced to the oysters and allowed to adhere for one hour. After adding phage suspension, the oyster meat was incubated at 3 °C for two days, followed by two hours at 37 °C A mixture of phages comprising 109 PFU/mL of each phage. In addition, microencapsulated phages were generated by combining a polymer mixture comprising 30% phage cocktail, 60% SM Buffer, and 10% solids (modified starch and maltodextrin). The tomato plants were categorised into three groups: the first group received E. coli O157:H7 inoculation, the second group received a microencapsulated cocktail phage inoculated with the bacterial host, and the third group served as a control without any inoculation Loach immersed against Aeromonas hydrophilia Individual phage or cocktail supplementation of the diet to combat Vibrio splendidus Containment of Streptococcus agalactiae by means of phage preparation introduced to the tank |
Cell counts were reduced by both BEC8 and Trans-cinnamaldehyde at the various MOIs and temperatures. The effect of the BEC8 cocktail on both liquid and desiccated cells was identical. An augmentation of the antimicrobial effect was observed upon the combination of both agents. After one hour of application to the cattle hide, phage cocktail demonstrated enhanced efficacy. The degree of bacterial eradication was equivalent to that obtained by washing the sample with water alone, when the sampling was conducted immediately following phage application Phage cocktail exhibited superior performance rating in inactivating the bacterial mixture across a range of conditions, including low to high MOIs, low to high temperatures, and shorter to extended periods of exposure. Both under arid and liquid conditions, bacterial levels could be regulated by phages. Phage-insensitive variants were not identified Phage cocktail could detectably reduce the quantity of various E. coli isolates tested at 4 °C. A decrease in value was observed at higher temperatures (25 and 37 °C), but it persisted only during the initial hours of incubation. Phage cocktails induce greater E. coli reduction in meat at elevated temperatures. A decline in cell count was observed exclusively with an increased phage concentration, encompassing both UV-treated and untreated phages. Untreated phages generally produce superior outcomes in milk. Consistency in observations was maintained for the control in RTE meat. The utilisation of UV-treated phages resulted in a more pronounced reduction of the host in uncooked beef Attenuating all bacterial genotypes is possible with a high concentration of phages. When bacteria are present singly or in combination, a reduction is observed The concentrations of E. coli O157:H7 in tomatoes encapsulated with microencapsulated phages were substantially reduced after 24 hours at 4 °C, in comparison to the control group that did not receive the phage cocktail. The observed differences persisted for a duration of five days. Free phages are less stable in the presence of stress factors than microencapsulated phages. Phage-treated loach exhibited a higher survival rate In contrast to the single phage and control groups, the phage cocktail-treated group exhibited an 82% survival rate 60% greater survival rate than the control group |
Viazis and Diez-Gonzalez, 2011 Coffey et al., 2011 Viazis et al., 2011 Tomat et al., 2018 Hudson et al., 2016 Le et al., 2018 Ramirez et al., 2018 Akmal et al., 2020 Li et al., 2016 Luo et al., 2018 |
Table 3.
An overview of medicinal plants secondary metabolites.
| Secondary metabolite (SM) | Characteristics | Sub-category of SM | Uses | References |
|
Phenols |
Probably constitute the largest group of plant SMs, They share the presence of one or more phenol groups as a common feature and range from simple structures with one aromatic ring to highly complex polymeric substances Widespread in plants and contribute significantly to the color, taste and flavor of many herbs, foods and drinks |
Quercetin | Anti-inflammatory | (Goławska et al., 2014, Hussein and El-Anssary, 2019) |
| Flavonoids | Antioxidant, anti-inflammatory and anti-allergic effects, anti-tumor | (Montanher et al., 2007, Serafin et al., 2009) | ||
| Gallic acid, Phenol | Antibacterial, antiviral, antifungal, anti-inflammatory, antitumor, anti-anaphylactic, antiseptic, anti-mutagenic, choleretic and bronchodilatory actions | (Pelczar et al., 1988, Yarnell, 2002, Spiller et al., 2008) | ||
| Tannin | Anti-diarrhea, antidote, antiseptic | (Jepson et al., 2013) | ||
| Coumarin | Anti-inflammatory, anticoagulant, anticancer and anti-Alzheimer’s | (Xu et al., 2015) | ||
|
Alkaloids |
Organic compounds with at least one nitrogen atom in a heterocyclic ring. Except for the fact that they are all nitrogen-containing compounds, no general definition fits all alkaloids. Many are toxic to animals to cause death if eaten |
Aromatics, Carbolines, Ergots, Imidazoles, Pyridines, Purines, Quinolines, Piperidines, etc. |
Analgesia, local anesthesia, cardiac stimulation, respiratory stimulation and relaxation, vasoconstriction muscle relaxation and toxicity, as well as antineoplastic, hypertensive and hypotensive properties. antibacterial, antifungal, antiviral |
(Seigler, 1998, Hoffmann, 2003, Hussein and El-Anssary, 2019) |
|
Saponins |
This hydrophobic-hydrophilic asymmetry means that these compounds have the ability to lower surface tension and are soap-like. They form foam in aqueous solutions and cause hemolysis of blood erythrocytes in vitro. |
Pentoses, Hexoses, or Uronic acids |
Antitumor, piscicidal, molluscicidal, spermicidal, sedative, expectorant, anti-inflammatory and analgesic properties. | Rehab 2018 (Hussein and El-Anssary, 2019) |
|
Terpenes |
Most diverse group of plant SMs All forms are derived chemically from 5-carbon isoprene units assembled in different ways Classified according to the number of isoprene units in the molecule |
Monoterpenes, Sesquiterpenes, Sesterterpenes, Hemiterpenes, Diterpenes Triterpenes |
Anti-hemorrhagic, analgesic, antibacterial, antifungal, anti-inflammatory, antineoplastic and antiprotozoal activities, anti-rheumatics |
(Hoffmann, 2003, Culioli et al., 2003) |
|
Lipids |
Major structural components of all biological membranes Source of energy reservoirs and fuel for cellular activities in addition to being vitamins and hormones Although lipids are primary metabolites, recent studies revealed pharmacological activities to members of this class of SMs |
Fixed oils, Waxes, Essential oils, Sterols, Fat-soluble vitamins (such as vitamins A, D, E and K), Phospholipids and others |
Anti-inflammatory, anti-aging, wound healing activities, antiseptic, antimicrobial, analgesic, sedative, spasmolytic and locally anesthetic remedies. They are also used as fragrances in embalmment, as sunscreens, moisturizer and in food preservation |
(Masotti et al., 2003, Fahy et al., 2009, Subramaniam et al., 2011) |
|
Carbohydrates |
Starter for all SMs and animal biochemical. Although carbohydrates are primary metabolites, they are incorporated in plenty of SMs through glycosidation linkages. Polymers of simple sugars and uronic acids produce mucilage and gums |
Monosaccharides, Disaccharides, Oligosaccharides and Polysaccharides | Demulcent, emollient |
(Asif et al., 2011, Anbalahan, 2017) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



