
Submitted:
27 December 2023
Posted:
27 December 2023
You are already at the latest version
Abstract
Olive trees have a unique reproductive pattern marked by biennial fruiting. This study delves into the repercussions of alternate fruit bearing on the water relations of olive trees and the associated ecophysiological mechanisms. The experiment spanned two consecutive years: the "ON" year, characterized by a high crop load, and the "OFF" year, marked by minimal fruit production. Key ecophysiological parameters, including sap flow, stomatal conductance, and photosynthetic rate, were monitored in both years. Pre-dawn water potential was measured using continuous stem psychrometers and the pressure chamber technique. Biochemically, non-structural carbohydrate concentrations (starch, sucrose, and mannitol) and the carbon-stable isotope ratio (δ13C) in olive leaves were examined. Results unveiled heightened leaf gas exchange during the "ON" year, leading to increased water consumption and photosynthetic rates. This escalated water usage during the "ON" year significantly reduced leaf water potential. Sucrose and starch concentrations were notably elevated in the "ON" year, with no significant impact on mannitol concentration. Regarding the carbon stable isotope ratio, leaves from the "OFF" year exhibited significantly higher δ13C abundance, indicative of heightened resistance to the CO2 pathway from the atmosphere to carboxylation sites compared to the "ON" year plants.
Keywords:
sink-source effect
; alternate bearing
; thermocouple psychrometry
; non-structural carbohydrates
; carbon stable isotope ratio
1. Introduction
Olive (Olea europaea L.) is one of the most important fruit crops worldwide, with a significant contribution to the economy of many Mediterranean countries [1]. Olive trees have a unique reproductive habit, with alternate bearing being a common phenomenon [2]. Alternate bearing refers to the tendency of a fruit tree to produce a full crop load in one year (ON year) followed by a low crop load in the following year (OFF year), which can significantly affect tree productivity. The vegetative development is restricted during the “ON” year, which impacts flower induction and, consequently, the following year’s production. The competitive dominance of growing fruits on shoot meristems regarding the photosynthates is responsible for this decrease in vegetative development [3] generally reported as the “sink-source effect.”
The sink-source theory is a widely accepted conceptual framework for understanding the relationship between fruit production and plant growth [4]. According to this theory, the plant is divided into two major components: the source, which refers to the leaves and other photosynthetic organs that produce carbohydrates through photosynthesis, and the sink, which refers to the fruit and other non-photosynthetic organs, such as roots, that consume carbohydrates for growth and development. The balance between source and sink determines the rate of photosynthesis and the allocation of resources within the plant. Particularly, heavy crop loads are reported to increase stomatal conductance and, therefore, the photosynthetic rate in various species like apples [5,6,7], avocados [8], peaches [9,10], citrus [11,12], grapevines [13,14,15], and olives [16,17,18,19]. Two primary mechanisms are reported to be involved in the downregulation of photosynthesis as a result of alternations in the sink–source balance; one is by decreasing Rubisco content or its activation state [20], and the other is by reducing stomatal and mesophyll conductance, which could lead in a reduction in CO2 concentration in the sites of carboxylation, potentially as a result of the accumulation of carbohydrates [21]. Indeed, Bustan et al. [22] found that during summer, when there was a high demand for carbohydrates for fruit growth and oil production, the stored non-structural carbohydrates in olives during the “ON” years decreased. Moreover, Haouari et al. [16] reported an accumulation of leaf soluble sugars and starch concentrations after severe fruit thinning in Olea europaea cv. Besbassi. Mainly, sucrose and mannitol are considered the dominant soluble sugars and, along with starch, consist of the most abundant carbohydrates in the leaves of olive trees [22,23]. Additionally, to the abundance of these two sugars, they have also been deemed the primary translocated sugars in the leaves and branches of olive trees [24]. On the other hand, alterations in stomatal conductance are known to be closely related to plant water status. In particular, high crop load is reported to result in lower stem water potential values in several species, such as apples [25,26], nectarines [27], and peaches [28]. In contrast, other researchers reported no effect on plums [29] and peaches [30]. However, regarding olive trees, not only little research has been conducted on the relationship between fruit load and tree water status, but also contradictory results have been reported. Specifically, while Naor et al. [31] and Bustan et al. [32] found that high values of sink-to-source ratio led to a decrease in stem water potential, Gucci er al [33] reported no such effect. On the other hand, Trentacoste et al. [34] observed a significant effect when Ψstem dropped below −1.4MPa. This contradiction in reported results can be partly attributed to the well-known methodological difficulties in obtaining continuous water potential data using the pressure chamber technique [35,36]. However, implementing recently developed and automated technologies such as thermocouple psychrometry allows, after proper calibration, the collection of robust data sets thus, facilitating continuous monitoring of plant water status.
Thus, the aim of this study is to evaluate the effects of alternate fruit bearing in olive trees’ physiological and hydrodynamic parameters as well as to identify the ecological significance and mechanisms underlying alternate bearing impact in olive trees.
2. Materials and Methods
2.1. Experimental orchard
The study was carried out in a commercial olive orchard in Aitoloakarnania prefecture, Western Greece, which was planted with the “Kalamon” olive variety (Olea europea cv. “Kalamon”). The region has a typical Mediterranean climate with mild, rainy winters and hot, dry summers. The trees were cultivated in heavy clay soil with adequate depth and were irrigated using a sprinkler system, with one sprinkler per tree, providing a flow rate of 320 liters per hour, according to the irrigation strategy commonly used by the farmers of this area. The same agricultural practices, including fertilization and plant protection, were followed during both study years. Additionally, the trees were not pruned from the beginning of the growing season in 2021 until the period prior to the harvest of the 2022 season.
2.2. Olive bearing cycle
The study was carried out for two consecutive years, with 2021 designated as the “OFF” year, characterized by low fruit production, while the other year (2022) was designated as the “ON” year, with high yields. During the “ON” year, the olive orchard produced a typical full crop load for the specific cultivar and orchard of 20.4 tons per hectare. On the other hand, during the “OFF” year, a low yield of 3.43 tons per hectare was recorded, indicating low production.
2.3. Monitoring of soil water content and meteorological parameters
Throughout the experiment, a weather station permanently installed in the field was used to continuously monitor various microclimatic parameters. Hourly measurements of solar radiation (Wh/m2), wind speed at 2 meters height (km/h), rainfall (mm), air relative humidity (%), and air temperature (°C) were collected and used to calculate the potential evapotranspiration (ETo) according to the Penman-Monteith equation [37].
Additionally, soil water content was continuously measured using capacitance sensors (EnviroSCAN, Sentek Sensor Technologies, Stepney, Australia) at a depth of 100cm. To ensure accurate measurements, the sensors were calibrated according to the manufacturer’s instructions and installed in a manner that covered the most absorption root area [38,39,40] with measurements taken at 10cm intervals.
2.4. Measurement of ecophysiological and hydrodynamic parameters
Stem water potential (Ψstem) values were obtained every 30 minutes using thermocouple psychrometers (PSY1, ICT International, Pty., Australia) installed on one main branch in two trees. The psychrometers’ calibration and installation followed the manufacturer’s recommended procedure. Initially, to calibrate the stem psychrometers, a series of standard sodium chloride (NaCl) solutions with known molality ranging from 0.1M to 1.0M were prepared. These concentrations correspond to the range of water potentials that plants typically experience, according to Lang [41]. Regarding the installation, a flat spot was initially engraved on the branch allowing the proper contact of the thermocouples with the sample surface. Following this, they were fully sealed with silicon grease and insulated with successive layering of foam materials and reflective coating to avoid thermal gradients due to incident sunlight.
Moreover, since there are not adequate results in the literature concerning the accuracy of this methodology in olive trees, concurrent measurements of water potential were also conducted during pre-dawn with a pressure chamber bomb (SKPM 1400/80, Skye Instruments, Powys, UK). For this measurement, five apical stems per tree with ca. five leaf pairs per stem were used [42].
Sap flow estimations were made using the heat ratio methodology (HRM-SFM1 Sap Flow Meter, ICT International, Armidale NSW, Australia). This technique was preferred because of its precision in measuring low sap velocity and its’ ability to provide robust measurements in olive tree water relations studies [43]. Briefly, this methodology is used to measure sap flow velocity by estimating the ratio of temperature increase following the emission of a short heat pulse [44]. Specifically, four sets of sensors were installed at an azimuthal angle of 90° and 50 cm above the soil surface on two selected trees, located in the center of the experimental orchard, during both study years. Using more sensors per tree instead of more sample trees with less sensors per tree was considered as the most appropriate method for obtaining reliable results since the azimuthal variability of sap flow is known to be considerably high in mature olive trees [45] and thus it must be thoroughly assessed by a sufficient number of sensors [46] in order to increase the accuracy of the results. On the other hand, sap flow values appeared not to be significantly different between plants in the same orchard (data not shown) a fact that is consistent with the results of Moreno et al. [47], Fuentes et al. [48], López-Bernal et al. [49], and Hernandez-Santana et al. [50], who also perform similar experiments using the same number of sample trees.
Prior to installation to identify the active xylem in the sapwood area, a sapwood core sample was extracted from the trunks of both trees using a tree-coring tool. Subsequently, methyl orange dye was used to differentiate the sapwood from the heartwood by applying it to the extracted sample with a micropipette. The active conducting xylem was then isolated, and its depth was measured using calipers. The sap flow measurements were used to estimate tree water consumption, and to compare it between the two study years, the ratio of daily sap flow to daily potential evapotranspiration (hereafter normalized sap flow- SFnorm) was calculated.
Maximum values of both stomatal conductance and net photosynthetic rate were measured on ten healthy, fully matured exterior leaves using an open gas analyzer system (LCPro+, ADC, Bioscientific Ltd., Hoddesdon, UK) under light-saturated conditions (i.e., photosynthetically active radiation at leaf surface higher that 1200 μmol/m2s) from 9:00 am to 11:00 am, at the same days when water potential was measured with the pressure chamber bomb.
2.5. Leaf area measurements
Leaf area index (LAI) was measured using the LAI-2000 Plant Canopy Analyzer (LI-COR Biosciences, Lincoln, NE, USA) as described by Villalobos et al. [51] on the same days as water potential and stomatal conductance measurements [52]. LAImax was measured at 50-80 cm from the trunk and LAImin at the center of the canopy since all plants had an open-shape canopy through pruning. LAIavg was calculated by integrating plant ground cover (GC) using the following equation [53]:
2.6. Measurement of soluble sugars and starch concentration
Analytical measurements regarding the concentration of non-structural carbohydrates in olive tree leaves were conducted using a Dionex P680 high-performance liquid chromatography (HPLC) system (Dionex Corporation, CA, USA) [54] on ten fully mature leaves (five from each tree) which were collected on DOY 210, 224, 243, and 264. The samples were immediately frozen in liquid nitrogen, kept at −80°C, and subjected to freeze-drying. After recording their weight, the samples were grounded into a fine powder with a mill (pulverisette 11, Fritsch GmbH 93 Milling and Sizing). Soluble sugars were extracted from 100 mg of fine leaf powder with 5 ml of ethanol (80%). The tubes were maintained in a heating block at 80°C for 1 hour and periodically shacked gently. Then they were centrifuged, and supernatants were used to perform the measurements in the mobile phase, which consisted of degassed, distilled, deionized water at a 0.6 mL/min flow rate. This process was repeated twice. Glucose extraction from starch was used to measure starch concentration according to the amyloglucosidase digestion (A-3042) procedure described by Schaffer et al., 1991. [55], mannitol, and starch concentrations were all expressed in mg/gr of dry weight.
2.7. Measurement of carbon stable isotope ratio
The contribution of different CO2 resistances from the atmosphere to the sites of carboxylation in chloroplasts was evaluated by determining the carbon stable isotope ratio in the leaves of both treatments using an Elementar Isoprime 100 isotope ratio mass spectrometer (IRMS) (IsoPrime Ltd., Cheadle Hulme, UK) coupled to an elemental analyzer (Elementar Vario Isotope EL Cube, Elementar Analysensystem GmbH, Hanau, Germany). Measurements were performed in twenty (20) leaves per treatment which were collected ten (10) days before harvest. The preparation of the samples in the laboratory followed the following procedure: at first, samples were oven-dried at 90°C for 68h. Then, they were grounded into a fine powder using a mill (pulverisette 11, Fritsch GmbH 93 Milling and Sizing) and were stored in falcon tubes placed in glass desiccators until IRMS analysis. Before analysis, samples were oven-dried again at 90°C for 48h. The results of the isotope ratio analysis were expressed in permille (‰) using the delta “δ” notation which was calculated according to the following equation:
where Rsample and Rstandard are the 13C/12C ratio of the sample and standard (V-PDB) respectively. The results were normalized to VPDB using ΙAΕA-600 (Caffeine, IAEA, Vienna, Austria), with assigned carbon isotope delta values and standard uncertainties (δ13CV-PDB=-27.77‰ ± 0.043‰) [56].
2.8. Statistical analysis
Differences in all parameters between the “ON” and “OFF” study years were assessed using a T-test at a 95% confidence level after testing for normal distribution. The standard error (SE) was used to measure statistical spread. Statistical analysis was performed with SPSS v.27. The values of sap flow are represented as means of eight measurements (n=8), i.e., four measurements per plant from two selected trees. Regarding the measurements of maximum photosynthetic rate (A) and stomatal conductance (gs), the values presented are the average of ten measurements (five measurements per tree). Water potential values measured with the pressure chamber bomb at pre-dawn represent the mean of ten measurements (five per tree), and the measurements from the psychrometry are the mean of two values (one measurement per tree).
3. Results
3.1. Environmental parameters and tree growth pattern
The environmental conditions were assessed by comparing potential evapotranspiration (ETo) during both study years. Results indicated that average ETo during the growing season of the “OFF” year (2021) exhibited significantly higher values (5.22±0.13 mm/day) compared to that during the “ON” year (4.83±0.12 mm/day) (Figure 1, Table 1). As far as leaf area index (LAI) is concerned there were no significant differences between the two study years (Table 1) a fact that can be attributed to the local farmer’s cultivation practice to avoid tree pruning after an “OFF” year in order to maintain annual shoots for fruit-bearing in the following year.
3.2. Soil moisture content and sap flow dynamics
Changes in soil moisture dynamics are shown in Figure 2. Two irrigation events (one per year) at DOY 205 of the “ON” year and DOY 210 of the “OFF” year are evident, which lasted for almost 4 hours each and applied a total of 26.44mm of water. During the growing season, no rainfall was recorded during the “ON” year, while during the “OFF” year, one rainfall of a magnitude of 3mm occurred and is considered insignificant since it did not affect soil moisture content.
During the major part of the growing season (DOY 214 to 270), olive plants exhibited significantly higher Normalized sap flow (SFnorm) values in the “ON” year compared to those in the “OFF” year. Despite these differences in SFnorm no significant differences in soil moisture content were recorded between the two studied years (Figure 3B).
3.3. Assessment of plant water status
Hydrodynamic measurements conducted either using the pressure chamber technique or the psychrometer resulted in almost similar values of pre-dawn leaf water potential values during the “ON” or the “OFF” year.
Figure 4.
Comparison of pre-dawn water potential (ΨPD) measurements obtained with the pressure chamber and the psychrometer in 14 samples. Significances: ns, no significant differences (p<0.05).
Figure 4.
Comparison of pre-dawn water potential (ΨPD) measurements obtained with the pressure chamber and the psychrometer in 14 samples. Significances: ns, no significant differences (p<0.05).
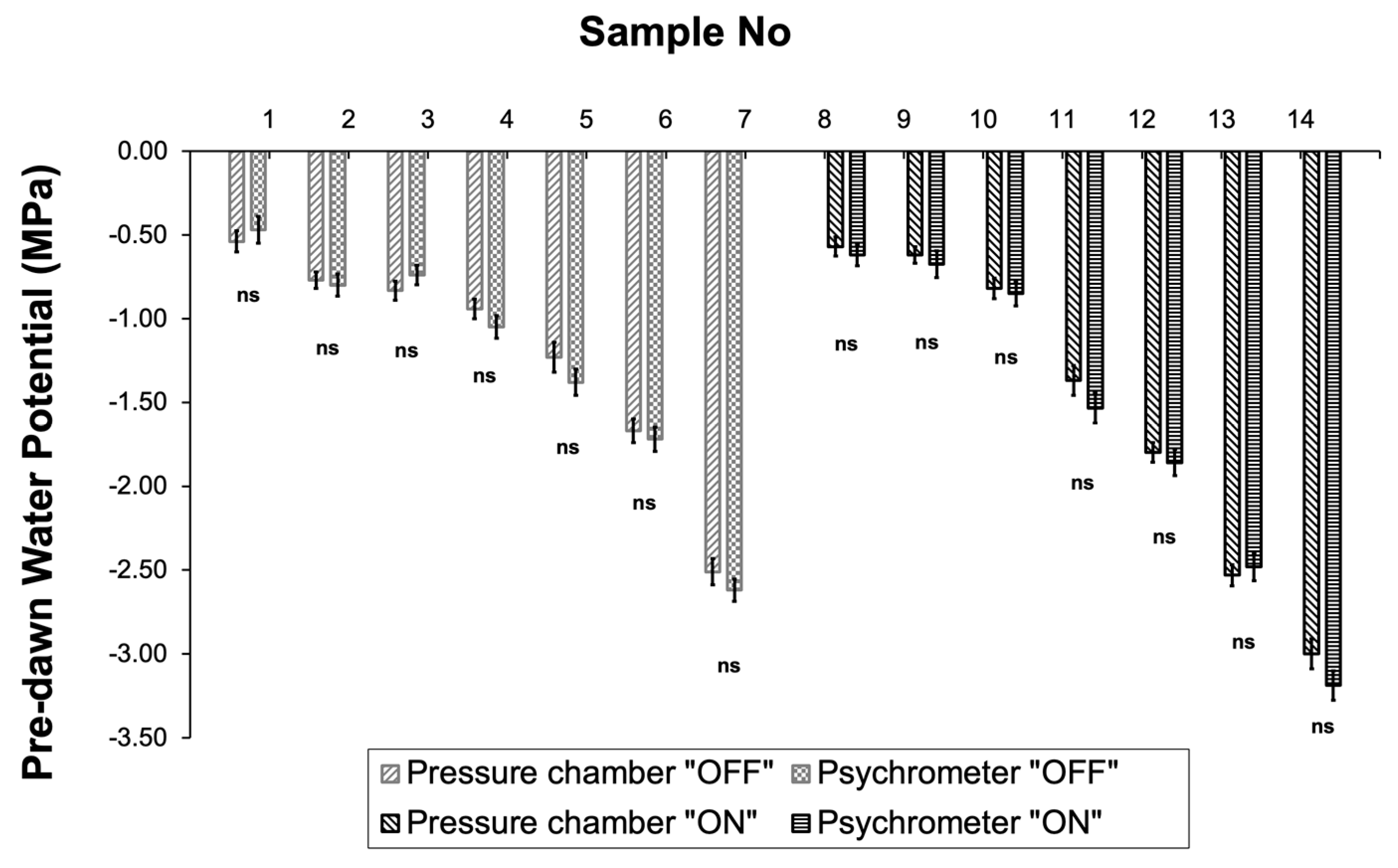
Measurements of ΨPD obtained with the psychrometers were then used to elucidate the relationship between plant water status and daily sap flow (Figure 5). Results indicated significantly higher tree water losses during the “ON” year compared to the “OFF” year at the same plant water status conditions, which could be attributed to the higher stomatal conductance (Figure 6). Indeed, olive plants exhibited higher values of maximum stomatal conductance in the “ON” years under similar plant water status conditions.
In addition, this differentiation in water consumption seemed to affect tree water status, with ΨPD being significantly higher during the “OFF” year compared to the “ON” year (Figure 7). ΨPD values during the “ON” year ranged from −0.785 MPa (measured after irrigation) to −4.32MPa at the end of the drying cycle, while the corresponding range during the “OFF” year was from −0.45MPa to −3.11MPa respectively.
A similar pattern to the stomatal conductance was also observed concerning the photosynthetic rate. The comparison between the two experimental years revealed that the photosynthetic rate of trees in the “OFF” year was significantly lower than in the “ON” year, irrespective of the plant water status conditions (Figure 8).
The concentration of sucrose and starch varied significantly, while the concentration of mannitol showed no significant difference between the two study years (Figure 9). During the “ON” year, the concentration of mannitol decreased at the end of September, while it remained nearly constant in the “OFF” year. Starch was the most abundant among the three measured carbohydrates, with an average concentration of ca. 41% and ca. 68% higher than mannitol and sucrose, respectively.
Carbon stable isotope ratio also exhibited a statistically significant difference between the two study years (Figure 10). During the “OFF” year leaves fixed a significantly higher amount of 13C compared to the “ON” year.
4. Discussion
Changes in environmental parameters indicated that the “OFF” year (from DOY 204 to DOY 278) was significantly drier compared to the corresponding period of the “ON” year (Figure 1, Table 1). Thus, it was expected that tree water consumption should be higher during the “OFF” year, leading to a higher tree water consumption rate [57]. However, this was not evident in our results. The comparison of the SFnorm, which by definition integrates the effect of the prevailing climatic conditions, clearly shows that the presence of fruits, which are considered the most potent sinks [58] is the preponderant factor responsible for the significantly higher tree water consumption (Figure 3A). This is also supported by the fact that even under similar soil moisture conditions, SFnorm was significantly higher during the “ON” year (Figure 3B). A reduction in water consumption during the “OFF” year in olive trees is also reported by [59] who attributed the differences in water consumption to the substantial changes induced by different fruit loads to tree canopy size. However, in our study, the measurements of LAI indicated that canopy size remained relatively constant between “ON” and “OFF” years, providing evidence that the sink-source effect was the determinant factor for differences in plant water consumption. Similar results regarding the sink-source effect on tree water consumption were reported in other studies related to olives [32], avocados [8], grapevine [15,60] and coffee [61], as a result of the significant increase of stomatal conductance of the heavy-fruiting trees. On the other hand, higher water consumption measured in the “ON” year trees could explain the significantly lower values of ΨPD. This is also confirmed by previous findings that highlight the effect of the sink: source ratio on water potential in apples [62,63], nectarines [27], and peaches [64]. In contrast, Gucci et al. [33] reported no effect of the “sink- source” ratio on ΨPD. However, in our results, the acquisition of a substantial amount of water potential data using the psychrometry method that have been previously verified for their accuracy, provides a more detailed understanding of the dynamic of leaf water potential changes in response to differences in fruit load. According to this data, ΨPD during the “ON” year dropped significantly to -4.32 MPa, indicating a moderate-to-high plant stress level [65]. Despite the above-mentioned low ΨPD values, olive trees are well known for their ability not only to withstand such conditions [66] but also preserving leaves’ physiological performance, including the production of assimilates [67,68].
The downregulation of photosynthesis, which is usually occurred under low fruit-bearing conditions, have been attributed to the rapid accumulation of photosynthesis-end products such as starch and soluble sugars in the leaves caused by the imbalance between the carbon assimilation rate and the carbon export rate to sinks [10,22,69,70]. According to Körner [21] the accumulation of carbohydrates in leaves is part of a “feedback” mechanism resulting in a significant decrease in photosynthetic rate. In this frame, the significant accumulation of carbohydrates, namely starch and sucrose, during the “OFF” year, which was evident in our results, seemed to contribute to the downregulation of photosynthesis. Our results demonstrate that starch was the carbohydrate with the greater concentration, followed by mannitol and sucrose. Furthermore, the concentration of starch and sucrose exhibited significant variations between the two study years in response to different crop loads, in accordance with the findings from other studies [17,22]. Conversely, no significant differences between treatments were observed regarding the mannitol concentration. A possible explanation might be based on the well-known positive role of this specific carbohydrate on olive responses to abiotic stresses [71] mainly on the drought tolerance of this species [24]. In the present study, the non-significant differentiation between the study years might be attributed to the fact that during the “ON” year, the trees experienced more intense water stress conditions. Given the significant osmoregulatory role of this molecule [72], it seems possible that olive trees in the “ON” year actively accumulate mannitol in leaf cells, thus increasing their concentration. As a result, the concentration of mannitol during the “ON” year increased, reaching almost similar values to those of the “OFF” year in which the high accumulation of this carbohydrate occurred due to the absence of strong sinks.
Despite numerous studies, the mechanisms and interspecific differences of the downregulation of photosynthesis in relation to changes in fruit load have not yet been fully understood [20,73] However, most studies suggest that when non-structural carbohydrates accumulate in source leaves, a downregulation of photosynthesis could result not only from decreases in the content and activity of Rubisco [74] but also from anatomical factors, such as an increase in cell wall thickness [75,76] which in turn could affect mesophyll conductance leading to reduced chloroplast CO2 concentrations. In this frame, stable carbon isotope analysis could be a useful tool to evaluate possible alterations induced by different fruit loads to the CO2 resistances (stomatal and mesophyll) from the atmosphere to the sites of carboxylation. Under optimal conditions, C3 plants discriminate against the heavier 13C atom [77,78] leading to lower values of δ13C. In our results, the significant increase in δ13C values observed during the “OFF” year indicates a reduction in carbon isotope discrimination due to significant alterations in CO2 conductances to the sites of carboxylation. Similar results were also reported in coffee plants [61], Phaseolus vulgaris plants [74], and apple trees [79], providing evidence that alterations in both stomatal and mesophyll conductances could contribute to photosynthesis downregulation in response to changes in olive fruit load.
5. Conclusions
Alternate bearing significantly affects the physiological parameters of olive trees, resulting in a substantial decrease in plant water usage during periods of lower fruit load due to the sink-source effect. Consequently, plant water status is affected, leading to significantly lower water potential values during the “ON” year. The present study provides evidence that the reduction in photosynthesis in the absence of fruits was caused by the concurrent effect of carbohydrate accumulation in the leaves and the stomatal closure, leading to increased total resistances in the CO2 pathway from the atmosphere to the chloroplasts’ carboxylation sites. Given the substantial influence of alternate bearing on hydrodynamic and ecophysiological parameters, particularly on tree water consumption, data obtained could be very useful for optimization of irrigation scheduling in olive orchards.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, E.K., A.Z. and A.P.; methodology, E.K., A.Z. and A.P.; validation, E.K., A.Z. and A.P.; curation, E.K., A.Z.; writing—original draft preparation, E.K., A.Z.; writing—review and editing, E.K., A.Z., A.P.; supervision, A.P.;. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Data Availability Statement
Data are available upon communication with corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Steduto, P.; Hsiao, T.C.; Fereres, E.; Raes, D. Crop Yield Response to Water; FAO, 2012; ISBN 9789251072745.
- Famiani, F.; Farinelli, D.; Gardi, T.; Rosati, A. The Cost of Flowering in Olive (Olea Europaea L.). Sci Hortic 2019, 252, 268–273. [CrossRef]
- Smith, H.M.; Samach, A. Constraints to Obtaining Consistent Annual Yields in Perennial Tree Crops. I: Heavy Fruit Load Dominates over Vegetative Growth. Plant Science 2013, 207, 158–167. [CrossRef]
- Kozlowski, T.; Pallardy, S. Chapter 5: Photosynthesis. In Physiology of Woody Plants; Academic Press: San Diego, California, USA, 1997; pp. 87–133 ISBN 0-12-424162-X.
- Baïram, E.; leMorvan, C.; Delaire, M.; Buck-Sorlin, G. Fruit and Leaf Response to Different Source–Sink Ratios in Apple, at the Scale of the Fruit-Bearing Branch. Front Plant Sci 2019, 10. [CrossRef]
- Fan, P.G.; Li, L.S.; Duan, W.; Li, W.D.; Li, S.H. Photosynthesis of Young Apple Trees in Response to Low Sink Demand under Different Air Temperatures. Tree Physiol 2010, 30, 313–325. [CrossRef]
- Wünsche, J.N.; Greer, D.H.; Laing, W.A.; Palmer, J.W. Physiological and Biochemical Leaf and Tree Responses to Crop Load in Apple. Tree Physiol 2005, 25, 1253–1263. [CrossRef]
- Silber, A.; Israeli, Y.; Levi, M.; Keinan, A.; Chudi, G.; Golan, A.; Noy, M.; Levkovitch, I.; Narkis, K.; Naor, A.; et al. The Roles of Fruit Sink in the Regulation of Gas Exchange and Water Uptake: A Case Study for Avocado. Agric Water Manag 2013, 116, 21–28. [CrossRef]
- Andrade, D.; Covarrubias, M.P.; Benedetto, G.; Pereira, E.G.; Almeida, A.M. Differential Source–Sink Manipulation Affects Leaf Carbohydrate and Photosynthesis of Early- and Late-Harvest Nectarine Varieties. Theor Exp Plant Physiol 2019, 31, 341–356. [CrossRef]
- Wei, D.; Li, S.H.; Fan, P.G.; Wang, L.J.; Li, W.D.; Yan, S.T. Photosynthetic Response to Low Sink Demand after Fruit Removal in Relation to Photoinhibition and Photoprotection in Peach Trees. Tree Physiol 2008, 28, 123–132. [CrossRef]
- Nebauer, S.G.; Renau-Morata, B.; Guardiola, J.L.; Molina, R.V.; Pereira, J. Photosynthesis Down-Regulation Precedes Carbohydrate Accumulation under Sink Limitation in Citrus. Tree Physiol 2011, 31, 169–177. [CrossRef]
- Syvertsen, J.P.; Goñi, C.; Otero, A. Fruit Load and Canopy Shading Affect Leaf Characteristics and Net Gas Exchange of ‘Spring’ Navel Orange Trees. Tree Physiol 2003, 23, 899–906. [CrossRef]
- Greer, D.H. Short-Term Temperature Dependency of the Photosynthetic and PSII Photochemical Responses to Photon Flux Density of Leaves of Vitis Vinifera Cv. Shiraz Vines Grown in Field Conditions with and without Fruit. Functional Plant Biology 2019, 46, 634–648. [CrossRef]
- Greer, D.H. Changes in the Temperature-Dependency of the Photosynthetic Response to Chloroplast CO2 Concentrations of Outdoor-Grown Vitis Vinifera Cv. Shiraz Vines with a Mid-Season Crop Removal. Environ Exp Bot 2020, 169, 103914. [CrossRef]
- Naor, A.; Gal, Y.; Bravdo, B. Crop Load Affects Assimilation Rate, Stomatal Conductance, Stem Water Potential and Water Relations of Field-Grown Sauvignon Blanc Grapevines; 1997; Vol. 48;
- Haouari, A.; Van Labeke, M.-C.; Steppe, K.; Mariem, F. Ben; Braham, M.; Chaieb, M. Fruit Thinning Affects Photosynthetic Activity, Carbohydrate Levels, and Shoot and Fruit Development of Olive Trees Grown under Semiarid Conditions. Functional Plant Biology 2013, 40, 1179–1186. [CrossRef]
- Perez-Arcoiza, A.; Diaz-Espejo, A.; Fernandez-Torres, R.; Perez-Romero, L.F.; Hernandez-Santana, V. Dual Effect of the Presence of Fruits on Leaf Gas Exchange and Water Relations of Olive Trees. Tree Physiol 2023, 43, 277–287. [CrossRef]
- Proietti, P.; Nasini, L.; Famiani, F. Effect of Different Leaf-to-Fruit Ratios on Photosynthesis and Fruit Growth in Olive (Olea Europaea L.); 2006; Vol. 44. [CrossRef]
- Rosati, A.; Paoletti, A.; Al Hariri, R.; Morelli, A.; Famiani, F. Resource Investments in Reproductive Growth Proportionately Limit Investments in Whole-Tree Vegetative Growth in Young Olive Trees with Varying Crop Loads. Tree Physiol 2018, 38, 1267–1277. [CrossRef]
- Kasai, M. Regulation of Leaf Photosynthetic Rate Correlating with Leaf Carbohydrate Status and Activation State of Rubisco under a Variety of Photosynthetic Source/Sink Balances. Physiol Plant 2008, 134, 216–226. [CrossRef]
- Körner, C. Carbon Limitation in Trees. Journal of Ecology 2003, 91, 4–17. [CrossRef]
- Bustan, A.; Avni, A.; Lavee, S.; Zipori, I.; Yeselson, Y.; Schaffer, A.A.; Riov, J.; Dag, A. Role of Carbohydrate Reserves in Yield Production of Intensively Cultivated Oil Olive (Olea Europaea L.) Trees. Tree Physiol 2011, 31, 519–530. [CrossRef]
- Gómez-González, S.; Ruiz-Jiménez, J.; Priego-Capote, F.; Luque de Castro, M.D. Qualitative and Quantitative Sugar Profiling in Olive Fruits, Leaves, and Stems by Gas Chromatography−Tandem Mass Spectrometry (GC-MS/MS) after Ultrasound-Assisted Leaching. J Agric Food Chem 2010, 58, 12292–12299. [CrossRef]
- Connor, D.J.; Fereres, E. The Physiology of Adaptation and Yield Expression in Olive. In Horticultural Reviews; 2005; Vol. 31, pp. 155–229 ISBN 9780470650882.
- Mpelasoka, B.; Behboudian, M.H.; Mills, T. Water Relations, Photosynthesis, Growth, Yield and Fruit Size of ‘Braeburn’ Apple: Responses to Deficit Irrigation and to Crop Load. J Hortic Sci Biotechnol 2001, 76, 150–156. [CrossRef]
- Naor, A.; Naschitz, S.; Peres, M.; Gal, Y. Responses of Apple Fruit Size to Tree Water Status and Crop Load. Tree Physiol 2008, 28, 1255–1261. [CrossRef]
- Naor, A.; Hupert, H.; Greenblat, Y.; Peres, M.; Kaufman, A.; Klein, I. The Response of Nectarine Fruit Size and Midday Stem Water Potential to Irrigation Level in Stage III and Crop Load. Journal of the American Society for Horticultural Science 2001, 126, 140–143. [CrossRef]
- Marsal, J.; Girona, J. Relationship between Leaf Water Potential and Gas Exchange Activity at Different Phenological Stages and Fruit Loads in Peach Trees. Journal of the American Society for Horticultural Science 1997, 122, 415–421. [CrossRef]
- Intrigliolo, D.S.; Castel, J.R. Crop Load Affects Maximum Daily Trunk Shrinkage of Plum Trees. Tree Physiol 2007, 27, 89–96. [CrossRef]
- Conejero, W.; Ortuño, M.F.; Mellisho, C.D.; Torrecillas, A. Influence of Crop Load on Maximum Daily Trunk Shrinkage Reference Equations for Irrigation Scheduling of Early Maturing Peach Trees. Agric Water Manag 2010, 97, 333–338. [CrossRef]
- Naor, A.; Schneider, D.; Ben-Gal, A.; Zipori, I.; Dag, A.; Kerem, Z.; Birger, R.; Peres, M.; Gal, Y. The Effects of Crop Load and Irrigation Rate in the Oil Accumulation Stage on Oil Yield and Water Relations of “Koroneiki” Olives. Irrig Sci 2013, 31, 781–791. [CrossRef]
- Bustan, A.; Dag, A.; Yermiyahu, U.; Erel, R.; Presnov, E.; Agam, N.; Kool, D.; Iwema, J.; Zipori, I.; Ben-Gal, A. Fruit Load Governs Transpiration of Olive Trees. Tree Physiol 2016, 36, 380–391. [CrossRef]
- Gucci, R.; Lodolini, E.; Rapoport, H.F. Productivity of Olive Trees with Different Water Status and Crop Load. Journal of Horticultural Science and Biotechnology 2007, 82, 648–656. [CrossRef]
- Trentacoste, E.R.; Sadras, V.O.; Puertas, C.M. Effects of the Source:Sink Ratio on the Phenotypic Plasticity of Stem Water Potential in Olive (Olea Europaea L.). J Exp Bot 2011, 62, 3535–3543. [CrossRef]
- Fernández, J.E. Understanding Olive Adaptation to Abiotic Stresses as a Tool to Increase Crop Performance. Environ Exp Bot 2014, 103, 158–179. [CrossRef]
- Fernández, J.E.; Diaz-espejo, A.; Romero, R.; Hernandez-santana, V.; García, J.M.; Padilla-díaz, C.M.; Cuevas, M. V Precision Irrigation in Olive (Olea Europaea L.) Tree Orchards. In Water scaricity and Sustainable Agriculture in Semiarid environment; Garcia Tejero, I., Duaran Zuazo, V., Eds.; AcademicPress, 2018; pp. 179–217 ISBN 9780128131640.
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. FAO Irrigation and Drainage Paper No. 56 - Crop Evapotranspiration; 1998;
- Kokkotos, E.; Zotos, A.; Patakas, A. Evaluation of Water Stress Coefficient Ks in Different Olive Orchards. Agronomy 2020, 10, 11. [CrossRef]
- Rallo, G.; Baiamonte, G.; Manzano Juárez, J.; Provenzano, G. Improvement of FAO-56 Model to Estimate Transpiration Fluxes of Drought Tolerant Crops under Soil Water Deficit: Application for Olive Groves. Journal of Irrigation and Drainage Engineering 2014, 140. [CrossRef]
- Xiloyannis, C.; Montanaro, G.; Dichio, B. Irrigation in Mediterranean Fruit Tree Orchards. In Irrigation systems and practices in challenging environments; Lee, T.S., Ed.; InTech, 2012 ISBN 978-95351-0420-9.
- Lang, A.R.G. Osmotic Coefficients and Water Potentials of Sodium Chloride Solutions from 0 to 40°C. Aust J Chem 1967, 20, 2017–2023. [CrossRef]
- Ahumada-Orellana, L.; Ortega-Farías, S.; Poblete-Echeverría, C.; Searles, P.S. Estimation of Stomatal Conductance and Stem Water Potential Threshold Values for Water Stress in Olive Trees (Cv. Arbequina). Irrig Sci 2019, 37, 461–467. [CrossRef]
- Fernandez, J.E.; Palomo, M.J.; Diaz-Espejo, A.; Clothier, B.E.; Green, S.R.; Giron, I.F.; Moreno, F. Heat-Pulse Measurements of Sap Flow in Olives for Automating Irrigation: Tests, Root Ow and Diagnostics of Water Stress. Agric Water Manag 2001, 51, 99–123. [CrossRef]
- Burgess, S.S.O.; Adams, M.A.; Turner, N.C.; Beverly, C.R.; Ong, C.K.; Khan, A.A.H.; Bleby, T.M. An Improved Heat Pulse Method to Measure Low and Reverse Rates of Sap Flow in Woody Plants. Tree Physiol 2001, 21, 589–598. [CrossRef]
- Nadezhdina, N.; Nadezhdin, V.; Ferreira, M.I.; Pitacco, A. Variability with Xylem Depth in Sap Flow in Trunks and Branches of Mature Olive Trees. Tree Physiol 2007, 27, 105–113. [CrossRef]
- López-Bernal, Á.; Alcántara, E.; Testi, L.; Villalobos, F.J. Spatial Sap Flow and Xylem Anatomical Characteristics in Olive Trees under Different Irrigation Regimes. Tree Physiol 2010, 30, 1536–1544. [CrossRef]
- Moreno, F.; Fernández, J.E.; Clothier, B.E.; Green, S.R. Transpiration and Root Water Uptake by Olive Trees. Plant Soil 1996, 184, 85–96. [CrossRef]
- Fuentes, S.; Mahadevan, M.; Bonada, M.; Skewes, M.A.; Cox, J.W. Night-Time Sap Flow Is Parabolically Linked to Midday Water Potential for Field-Grown Almond Trees. Irrig Sci 2013, 31, 1265–1276. [CrossRef]
- López-Bernal, Á.; García-Tejera, O.; Vega, V.A.; Hidalgo, J.C.; Testi, L.; Orgaz, F.; Villalobos, F.J. Using Sap Flow Measurements to Estimate Net Assimilation in Olive Trees under Different Irrigation Regimes. Irrig Sci 2015, 33, 357–366. [CrossRef]
- Hernandez-Santana, V.; Fernández, J.E.; Rodriguez-Dominguez, C.M.; Romero, R.; Diaz-Espejo, A. The Dynamics of Radial Sap Flux Density Reflects Changes in Stomatal Conductance in Response to Soil and Air Water Deficit. Agric For Meteorol 2016, 218–219, 92–101. [CrossRef]
- Villalobos, F.J.; Orgaz, F.; Mateos, L. Non-Destructive Measurement of Leaf Area in Olive (Olea Europaea L.) Trees Using a Gap Inversion Method. Agric For Meteorol 1995, 73, 29–42. [CrossRef]
- Kokkotos, E.; Zotos, A.; Tsirogiannis, G.; Patakas, A. Prediction of Olive Tree Water Requirements under Limited Soil Water Availability, Based on Sap Flow Estimations. Agronomy 2021, 11. [CrossRef]
- Diaz-Espejo, A.; Buckley, T.N.; Sperry, J.S.; Cuevas, M. v; de Cires, A.; Elsayed-Farag, S.; Martin-Palomo, M.J.; Muriel, J.L.; Perez-Martin, A.; Rodriguez-Dominguez, C.M.; et al. Steps toward an Improvement in Process-Based Models of Water Use by Fruit Trees: A Case Study in Olive. Agric Water Manag 2012, 114, 37–49. [CrossRef]
- Ma, B.; Chen, J.; Zheng, H.; Fang, T.; Ogutu, C.; Li, S.; Han, Y.; Wu, B. Comparative Assessment of Sugar and Malic Acid Composition in Cultivated and Wild Apples. Food Chem 2015, 172, 86–91. [CrossRef]
- Schaffer, A.A.; Nerson, H.; Zamski, E. Premature Leaf Chlorosis in Cucumber Associated with High Starch Accumulation. J Plant Physiol 1991, 138, 186–190. [CrossRef]
- Thomatou, A.A.; Psarra, E.; Mazarakioti, E.C.; Katerinopoulou, K.; Tsirogiannis, G.; Zotos, A.; Kontogeorgos, A.; Patakas, A.; Ladavos, A. Stable Isotope Analysis for the Discrimination of the Geographical Origin of Greek Bottarga ‘Avgotaracho Messolongiou’: A Preliminary Research. Foods 2022, 11. [CrossRef]
- Miserere, A.; Searles, P.S.; Manchó, G.; Maseda, P.H.; Rousseaux, M.C. Sap Flow Responses to Warming and Fruit Load in Young Olive Trees. Front Plant Sci 2019, 10, 1–13. [CrossRef]
- Blanke, M.M. Regulatory Mechanisms in Source Sink Relationships in Plants-a Review. In Proceedings of the International Symposium on Source-Sink Relationships in Plants; Ron’zhina, E.S., Blanke, M., Eds.; ISHS, Acta Horticulturae 835, 2007; pp. 13–20.
- Martín-Vertedor, A.I.; Rodríguez, J.M.P.; Losada, H.P.; Castiel, E.F. Interactive Responses to Water Deficits and Crop Load in Olive (Olea Europaea L., Cv. Morisca). II: Water Use, Fruit and Oil Yield. Agric Water Manag 2011, 98, 950–958. [CrossRef]
- Sandras, V.O.; Collins, M.; Soar, C.J. Modelling Variety-Dependent Dynamics of Soluble Solids and Water in Berries of Vitis Vinifera. Aust J Grape Wine Res 2008, 14, 250–259. [CrossRef]
- Damatta, F.M.; Cunha, R.L.; Antunes, W.C.; Martins, S.C. V; Araujo, W.L.; Fernie, A.R.; Moraes, G.A.B.K.; Damatta, F.M. In Field-Grown Coffee Trees Source-Sink Manipulation Alters Photosynthetic Rates, Independently of Carbon Metabolism, via Alterations in Stomatal Function. 2008.
- Girona, J.; Behboudian, M.H.; Mata, M.; Del Campo, J.; Marsal, J. Exploring Six Reduced Irrigation Options under Water Shortage for ‘Golden Smoothee’ Apple: Responses of Yield Components over Three Years. Agric Water Manag 2010, 98, 370–375. [CrossRef]
- Naschitz, S.; Naor, A.; Genish, S.; Wolf, S.; Goldschmidt, E.E. Internal Management of Non-Structural Carbohydrate Resources in Apple Leaves and Branch Wood under a Broad Range of Sink and Source Manipulations. Tree Physiol 2010, 30, 715–727. [CrossRef]
- Lopez, G.; Mata, M.; Arbones, A.; Solans, J.R.; Girona, J.; Marsal, J. Mitigation of Effects of Extreme Drought during Stage III of Peach Fruit Development by Summer Pruning and Fruit Thinning. Tree Physiol 2006, 26, 469–477. [CrossRef]
- Dichio, B.; Xiloyannis, C.; Angelopoulos, K.; Nuzzo, V.; Bufo, S.A.; Celano, G. Drought-Induced Variations of Water Relations Parameters in Olea Europaea. Plant Soil 2003, 257, 381–389. [CrossRef]
- Moriana, A.; Orgaz, F.; Pastor, M.; Fereres, E. Yield Responses of a Mature Olive Orchard to Water Deficits. Journal of the American Society for Horticultural Science jashs 2003, 128, 425–431. [CrossRef]
- Angelopoulos, K.; Dichio, B.; Xiloyannis, C. Inhibition of Photosynthesis in Olive Trees (Olea Europaea L.) during Water Stress and Rewatering. 1996, 47, 1093–1100.
- Naor, A. Irrigation Scheduling and Evaluation of Tree Water Status in Deciduous Orchards. In Horticultural Reviews; 2006; pp. 111–165 ISBN 9780470767986.
- Drossopoulos, J.B.; Niavis, C.A. Seasonal Changes of the Metabolites in the Leaves, Bark and Xylem Tissues of Olive Tree (Olea Europaea. L) II. Carbohydrates. Ann Bot 1988, 62, 321–327. [CrossRef]
- Li, W.D.; Li, S.H.; Yang, S.H.; Yang, J.M.; Zheng, X.B.; Li, X.D.; Yao, H.M. Photosynthesis in Response to Sink-Source Manipulations during Different Phenological Stages of Fruit Development in Peach Trees: Regulation by Stomatal Aperture and Leaf Temperature. J Hortic Sci Biotechnol 2005, 80, 481–487. [CrossRef]
- Smirnoff, N. Plant Resistance to Environmental Stress. Curr Opin Biotechnol 1998, 9, 214–219. [CrossRef]
- Stoop, J.M.H.; Williamson, J.D.; Mason Pharr, D. Mannitol Metabolism in Plants: A Method for Coping with Stress. Trends Plant Sci 1996, 1, 139–144. [CrossRef]
- Ainsworth, E.A.; Rogers, A.; Nelson, R.; Long, S.P. Testing the “Source–Sink” Hypothesis of down-Regulation of Photosynthesis in Elevated [CO2] in the Field with Single Gene Substitutions in Glycine Max. Agric For Meteorol 2004, 122, 85–94. [CrossRef]
- Sugiura, D.; Betsuyaku, E.; Terashima, I. Interspecific Differences in How Sink-Source Imbalance Causes Photosynthetic Downregulation among Three Legume Species. Ann Bot 2019, 123, 715–726. [CrossRef]
- Sugiura, D.; Betsuyaku, E.; Terashima, I. Manipulation of the Hypocotyl Sink Activity by Reciprocal Grafting of Two Raphanus Sativus Varieties: Its Effects on Morphological and Physiological Traits of Source Leaves and Whole-Plant Growth. Plant Cell Environ 2015, 38, 2629–2640. [CrossRef]
- Sugiura, D.; Watanabe, C.K.A.; Betsuyaku, E.; Terashima, I. Sink–Source Balance and Down-Regulation of Photosynthesis in Raphanus Sativus: Effects of Grafting, N and CO2. Plant Cell Physiol 2017, 58, 2043–2056. [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the Relationship Between Carbon Isotope Discrimination and the Intercellular Carbon Dioxide Concentration in Leaves. Functional Plant Biology 1982, 9, 121–137. [CrossRef]
- Evans, J.R.; Caemmerer, S. V; Setchell, B.A.; Hudson, G.S. The Relationship Between CO2 Transfer Conductance and Leaf Anatomy in Transgenic Tobacco With a Reduced Content of Rubisco. Functional Plant Biology 1994, 21, 475–495. [CrossRef]
- Ding, N.; Chen, Q.; Zhu, Z.; Peng, L.; Ge, S.; Jiang, Y. Effects of Crop Load on Distribution and Utilization of 13C and 15N and Fruit Quality for Dwarf Apple Trees. Sci Rep 2017, 7, 14172. [CrossRef]
Figure 1.
Comparative assessment of ETo during the two study years. (DOY 155= 4th of June).
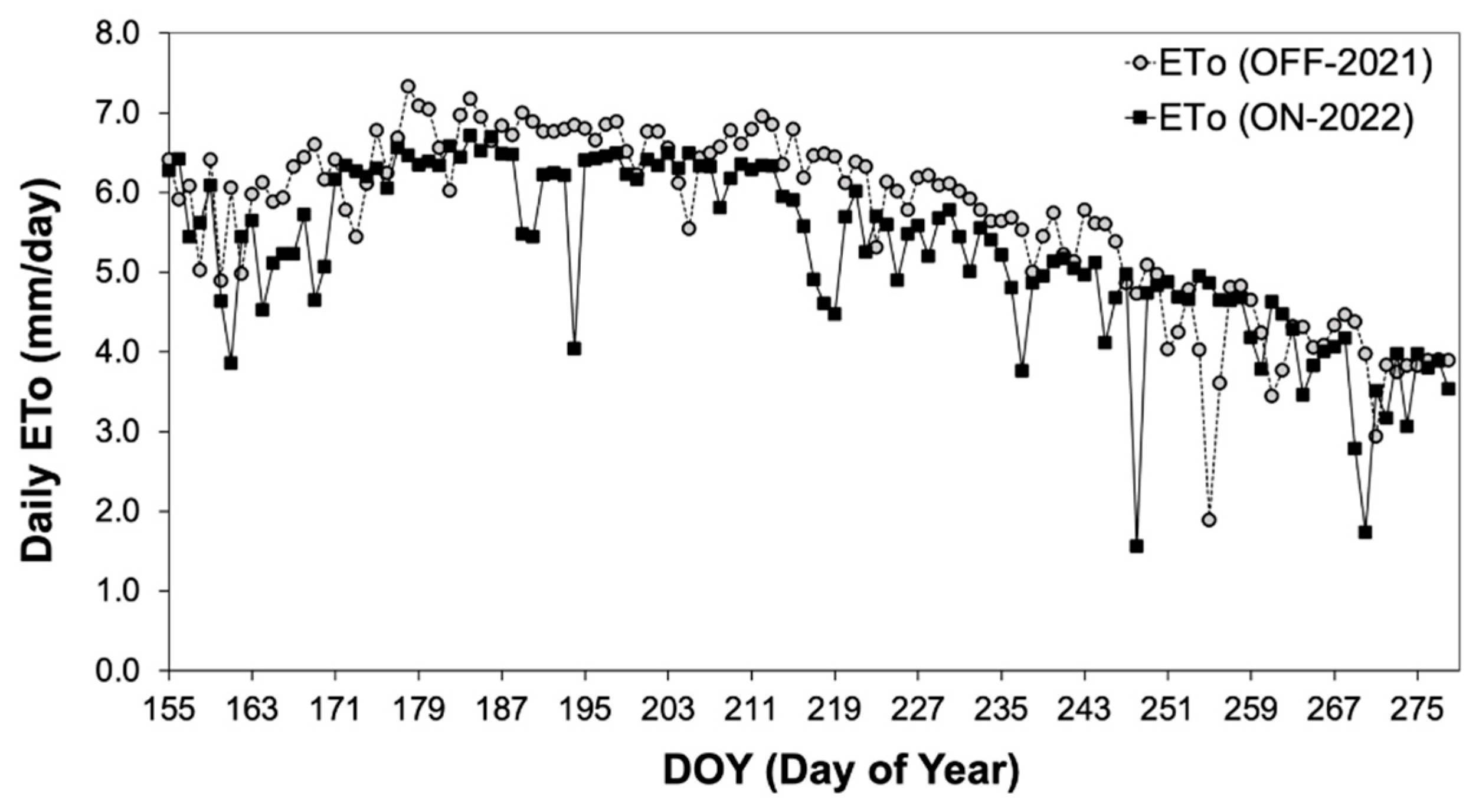
Figure 2.
Soil moisture dynamics during the study period (DOY 204= 23rd of July).
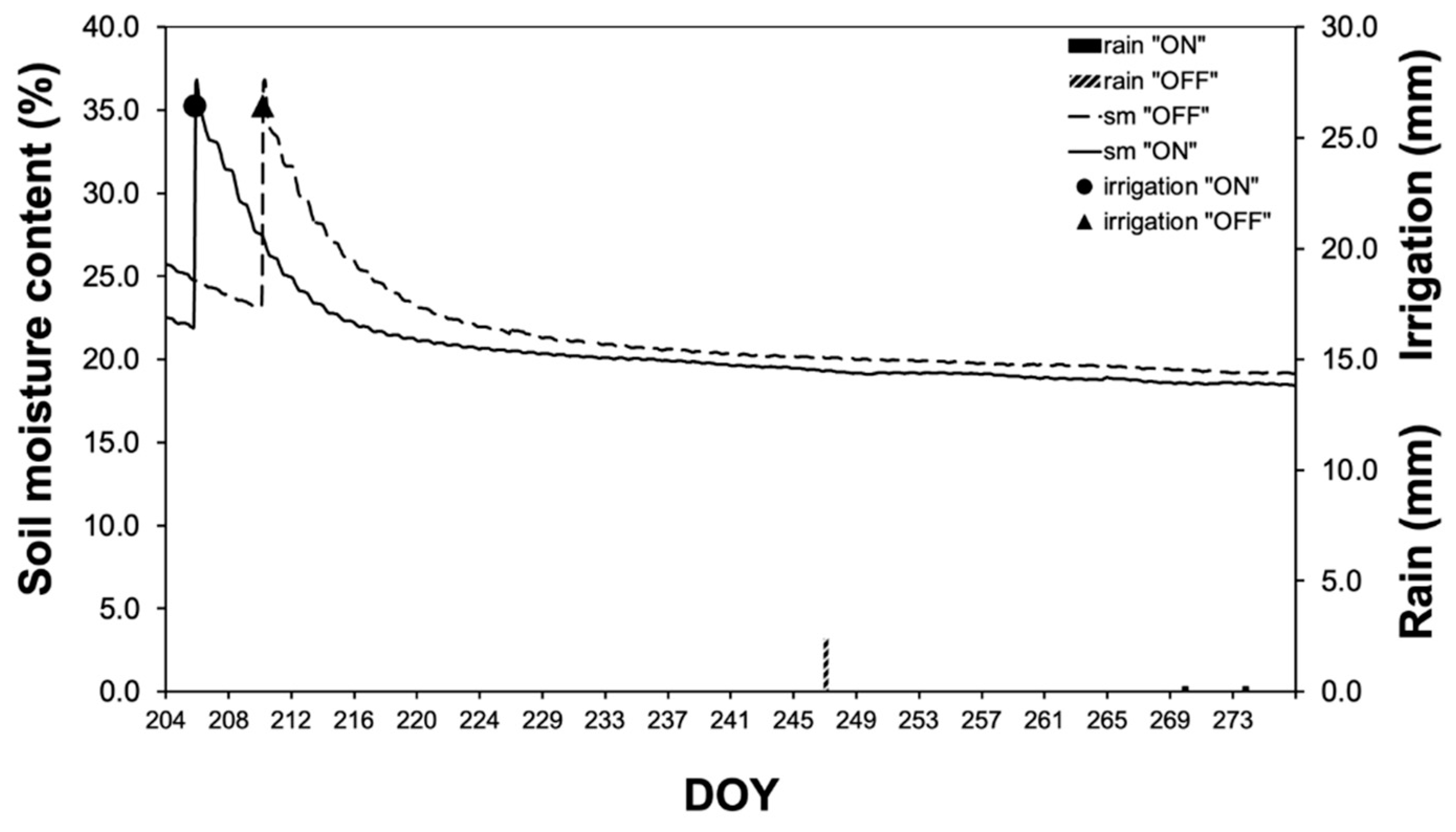
Figure 3.
Changes in SFnorm (A) and SFnorm in relation to the daily average soil moisture content (s.m.) (B) during the experimental period.
Figure 3.
Changes in SFnorm (A) and SFnorm in relation to the daily average soil moisture content (s.m.) (B) during the experimental period.
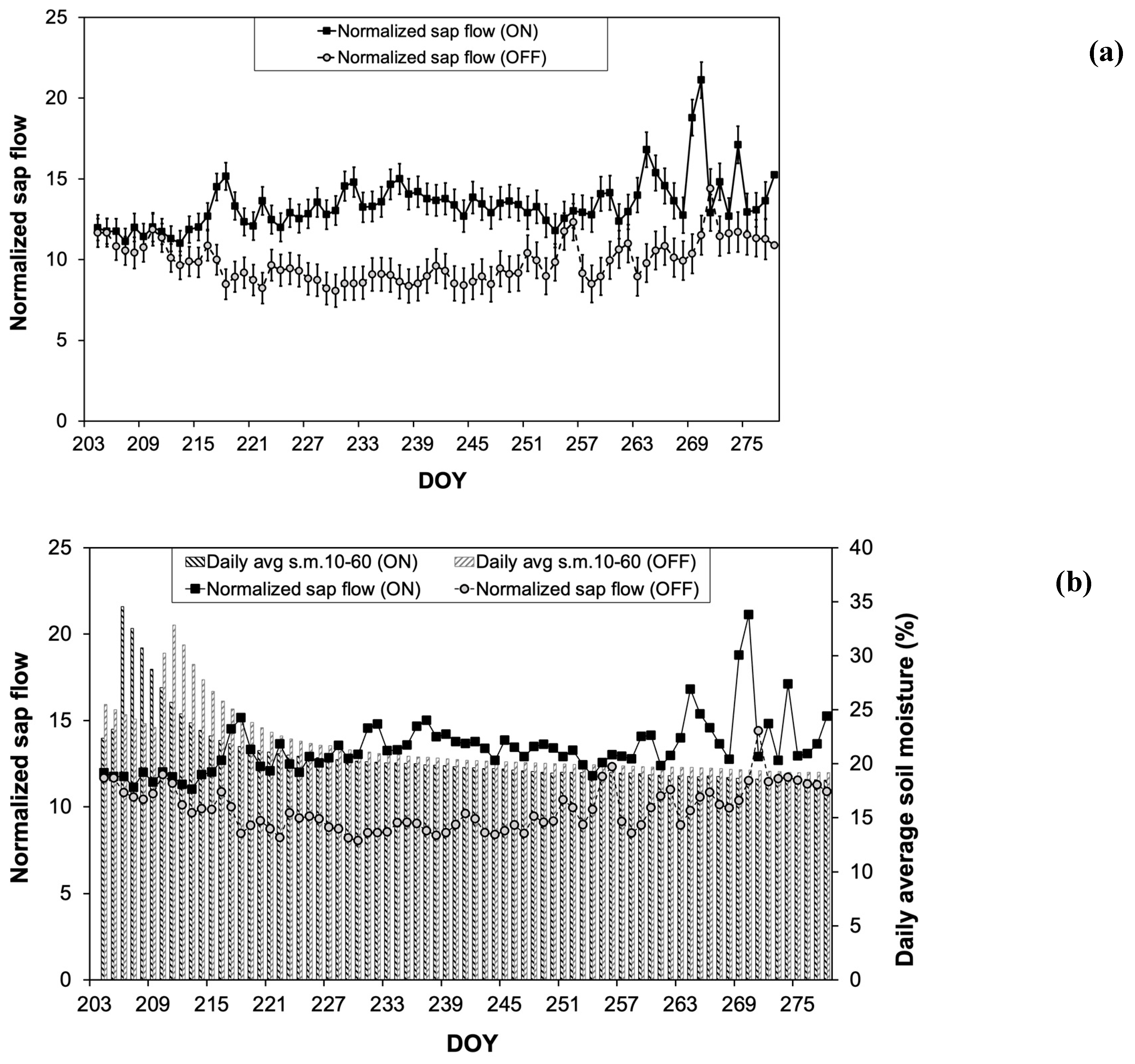
Figure 5.
Relationship between daily water consumption (liters/day) and plant water status during “ON” and “OFF” years.
Figure 5.
Relationship between daily water consumption (liters/day) and plant water status during “ON” and “OFF” years.
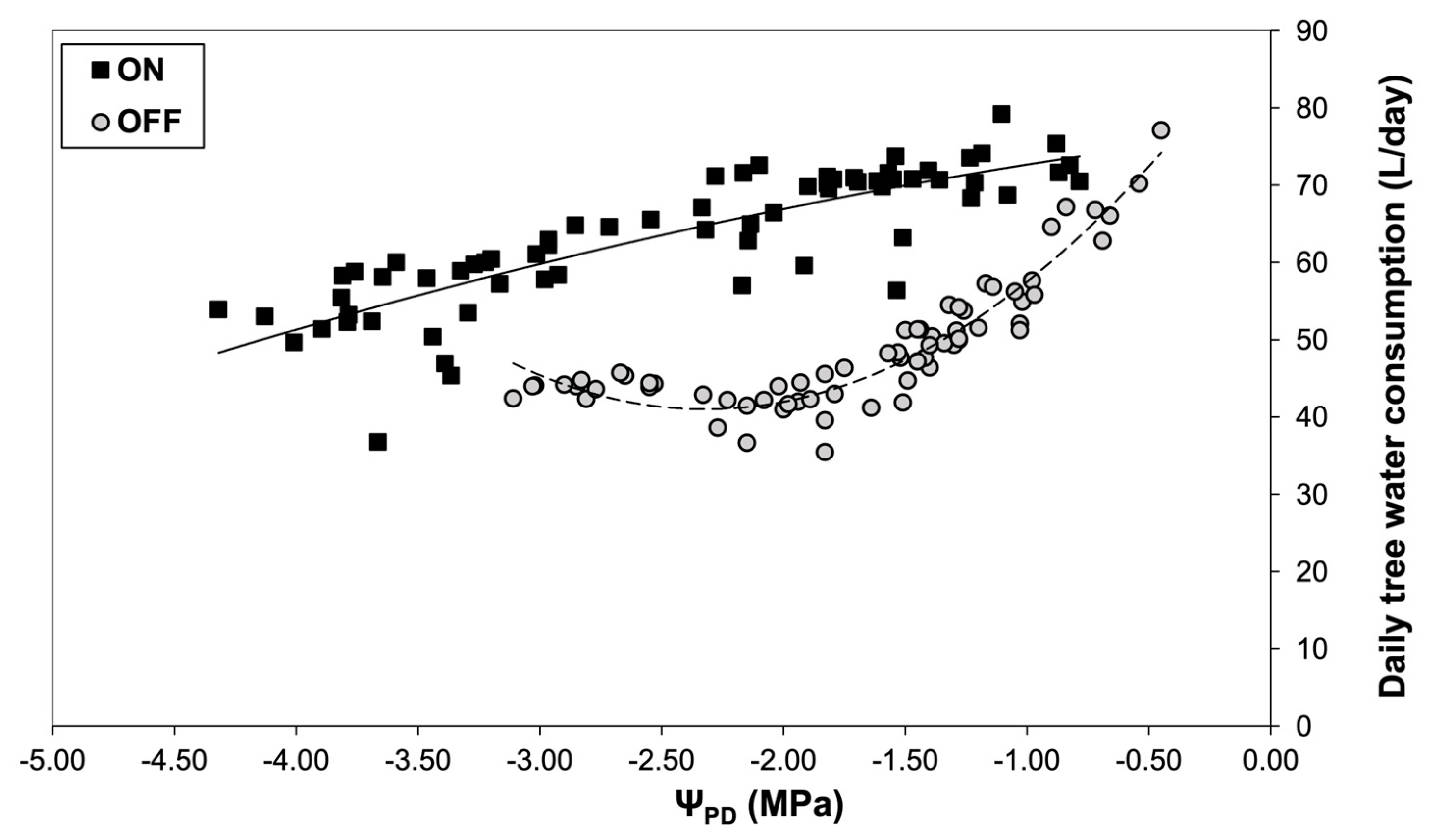
Figure 6.
Stomatal conductance (gs) during two different fruiting years in relation to plant water status.
Figure 6.
Stomatal conductance (gs) during two different fruiting years in relation to plant water status.
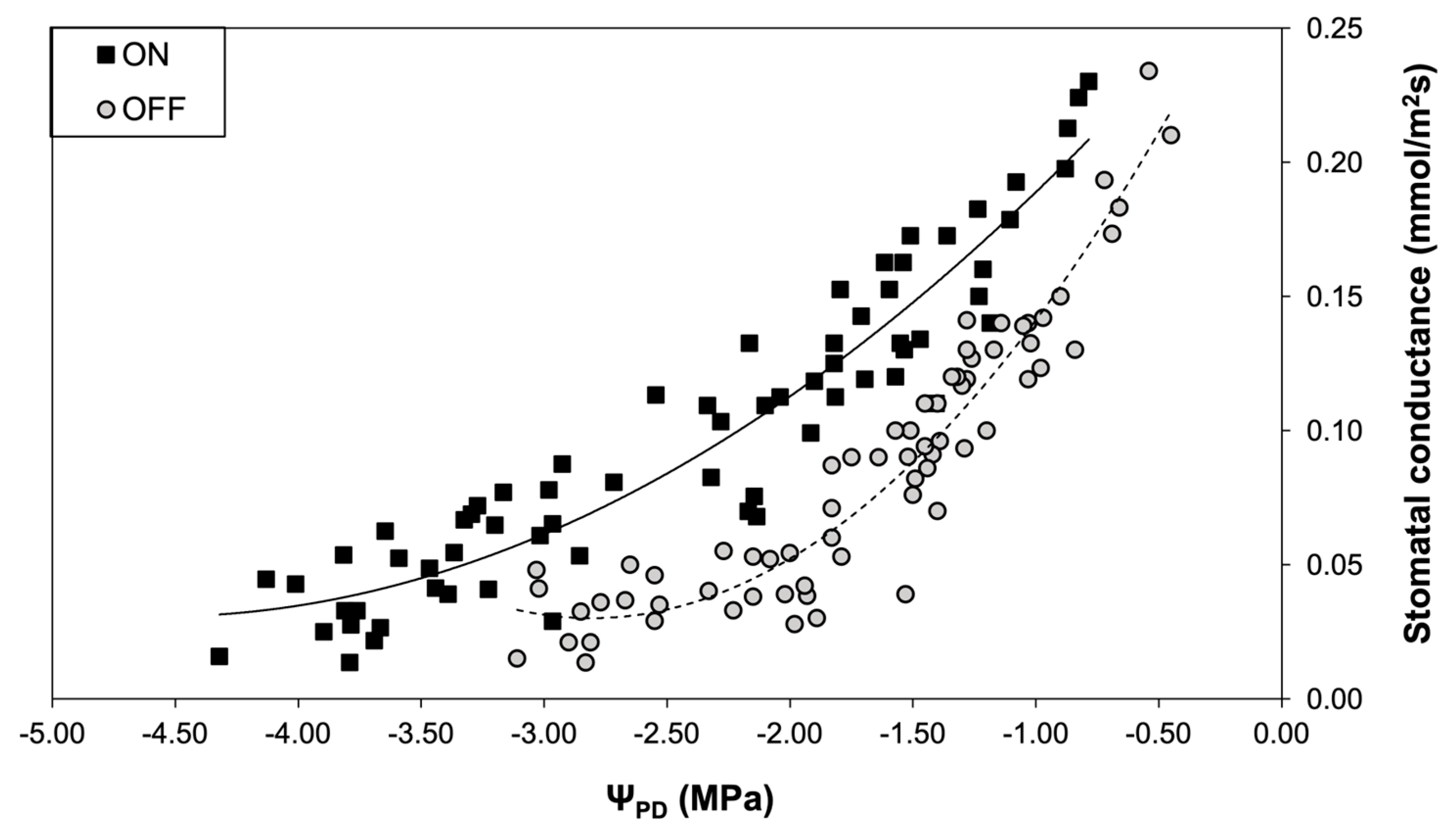
Figure 7.
Comparative assessment of ΨPD measured with thermocouple psychrometers during the two study years. Significances: *, statistically significant differences (p<0.05).
Figure 7.
Comparative assessment of ΨPD measured with thermocouple psychrometers during the two study years. Significances: *, statistically significant differences (p<0.05).
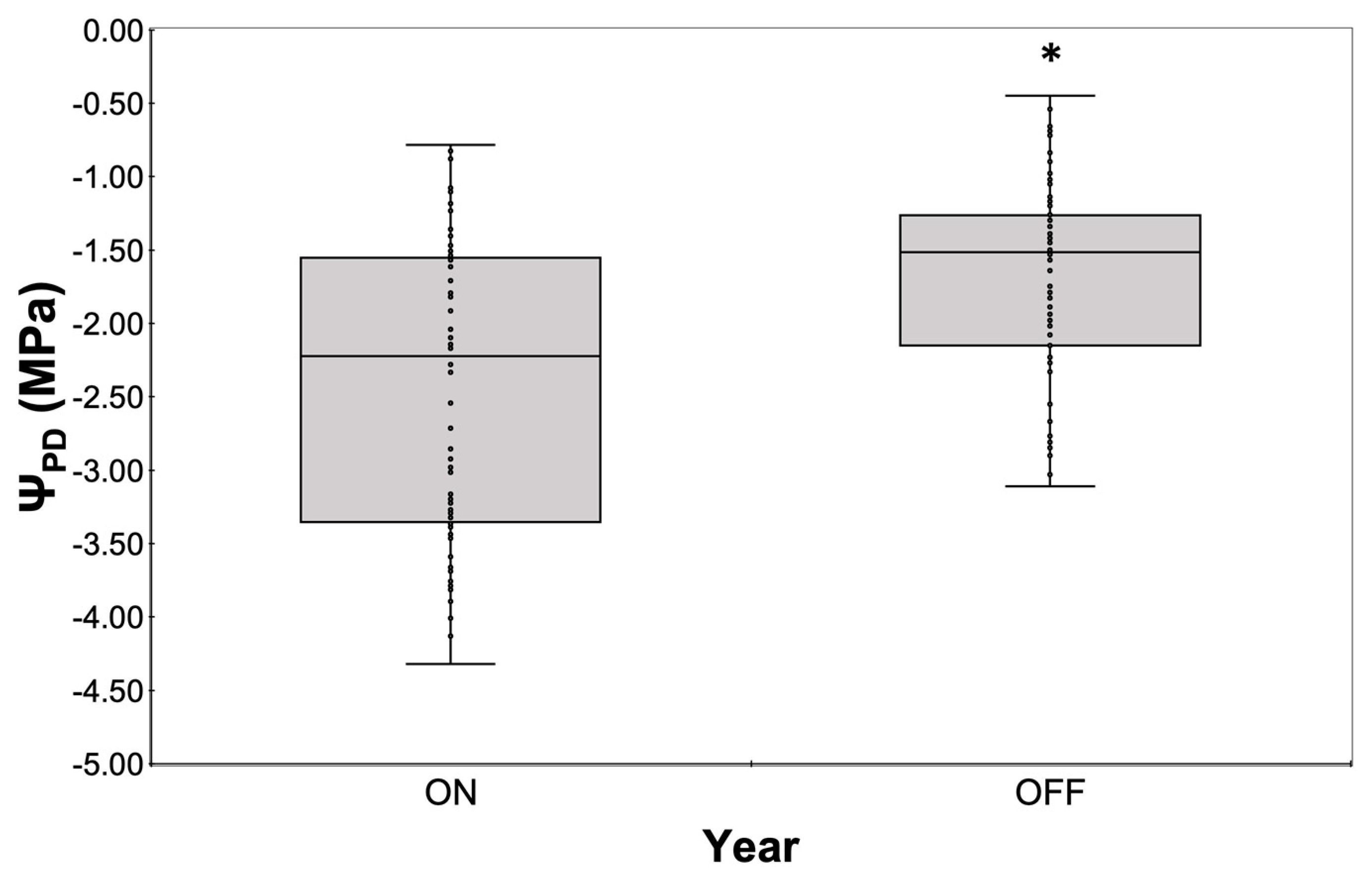
Figure 8.
Correlation between plant water status and photosynthetic rate during “ON” and “OFF” years.
Figure 8.
Correlation between plant water status and photosynthetic rate during “ON” and “OFF” years.
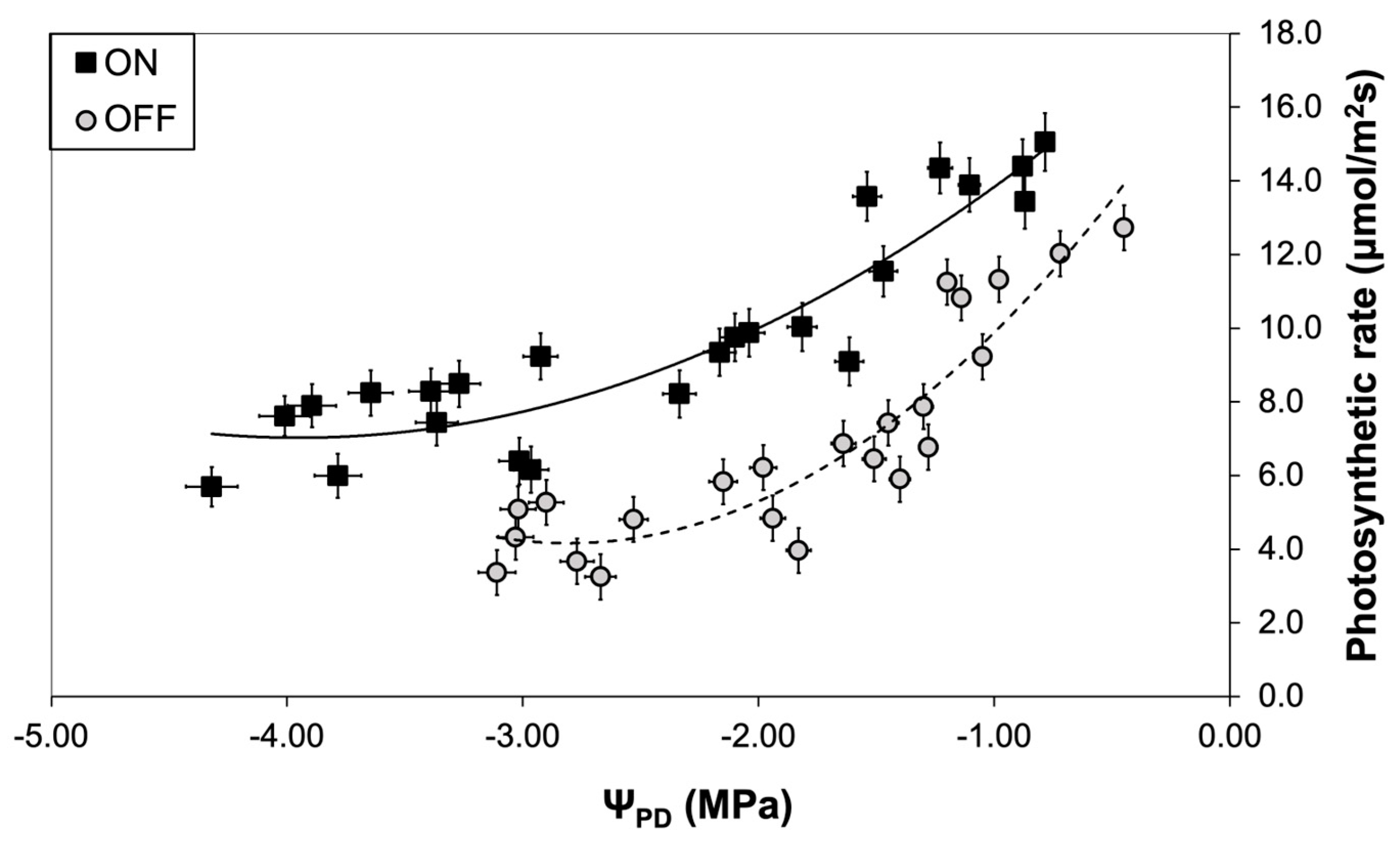
Figure 9.
Sucrose, mannitol, and starch concentration in olive leaves during “ON” and “OFF” years. Significances: *, statistically significant differences (p<0.05), ns: no significant differences (p<0.05).
Figure 9.
Sucrose, mannitol, and starch concentration in olive leaves during “ON” and “OFF” years. Significances: *, statistically significant differences (p<0.05), ns: no significant differences (p<0.05).
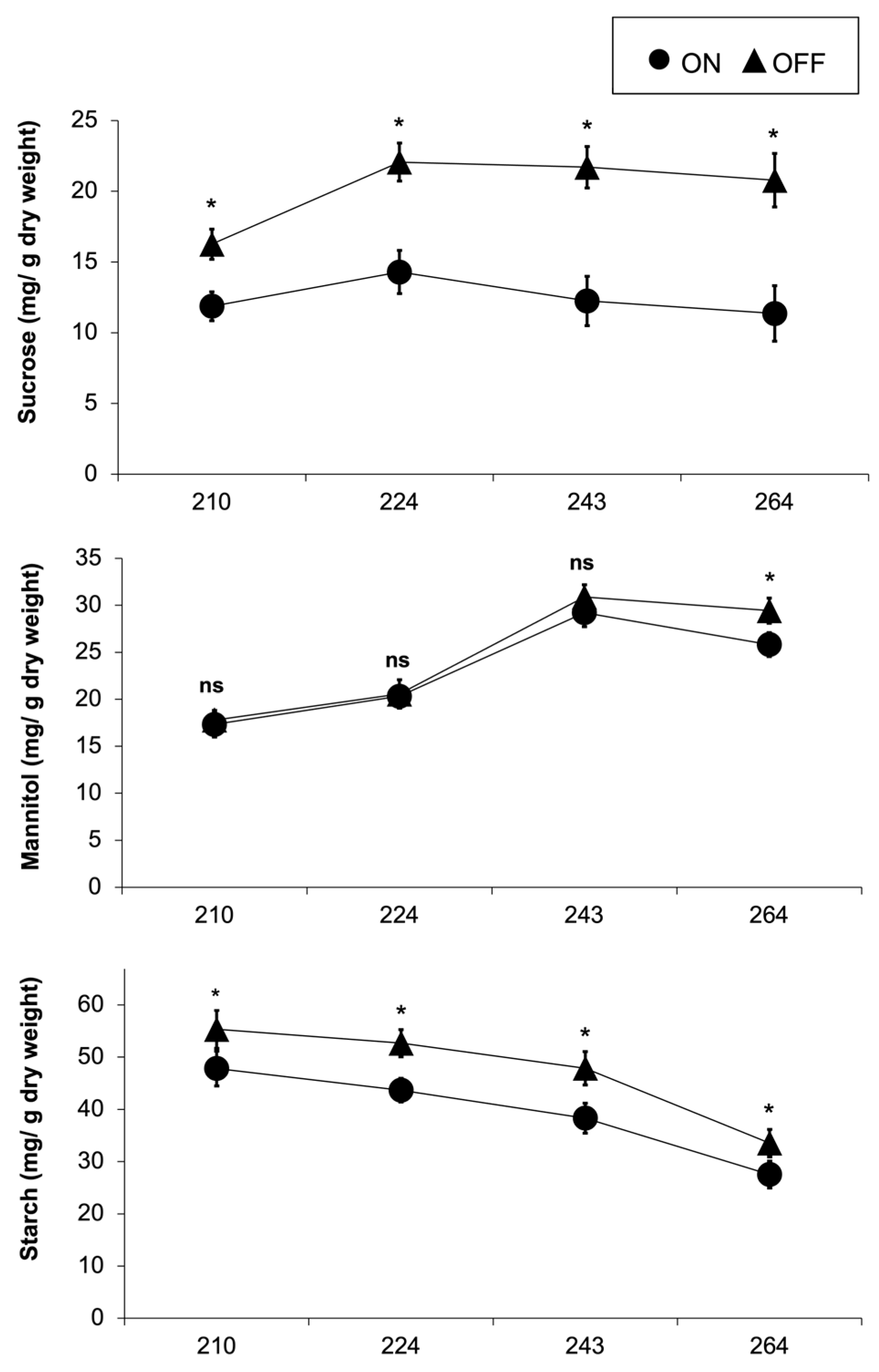
Figure 10.
Stable carbon isotope ratio (δ13C) during the two study years. Significances: *, statistically significant differences (p<0.05).
Figure 10.
Stable carbon isotope ratio (δ13C) during the two study years. Significances: *, statistically significant differences (p<0.05).
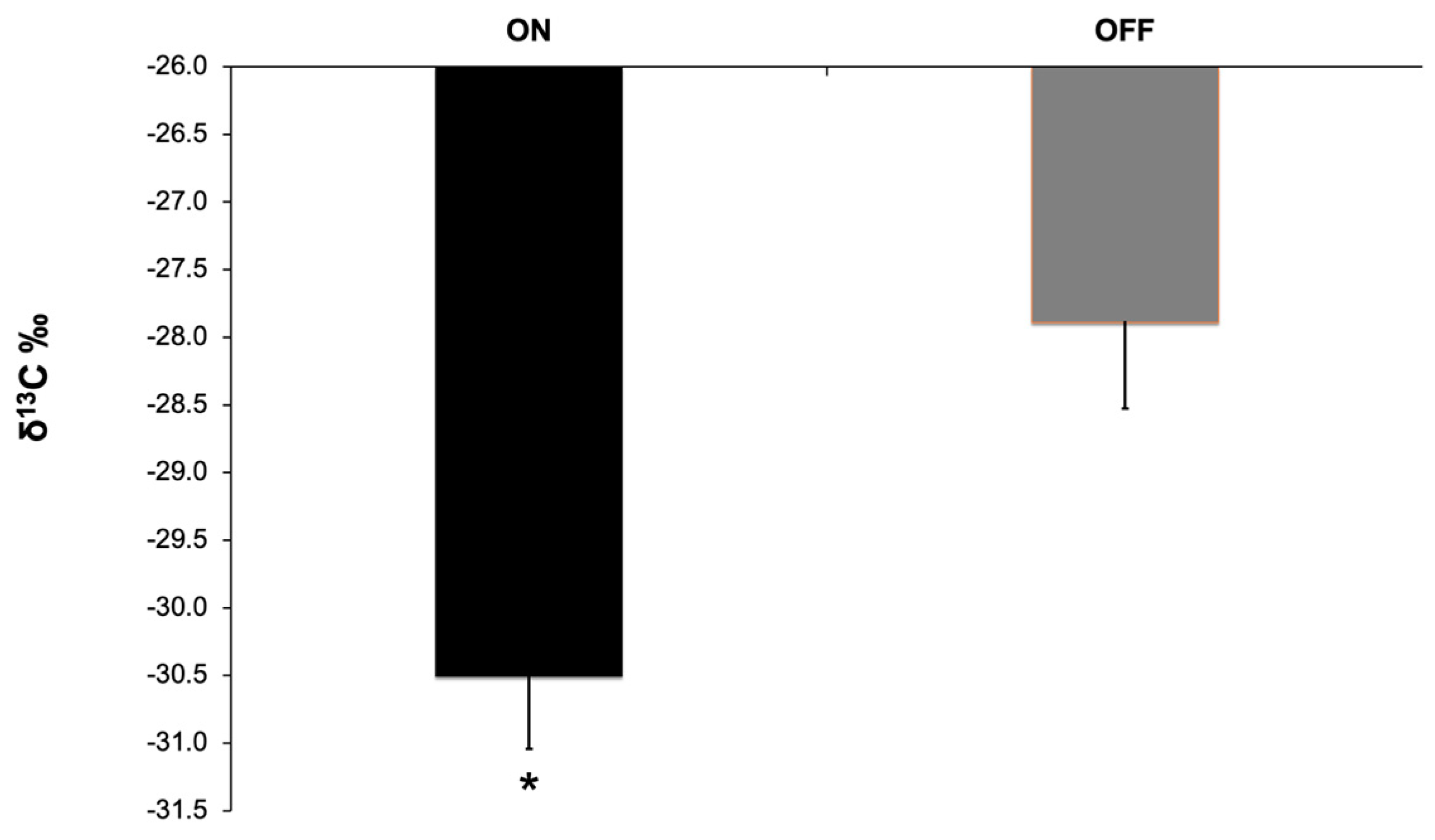

Table 1.
Fruit production, leaf area index (LAI), and average ETo during the study years 2021 (“OFF”) and 2022 (“ON”) years.
Table 1.
Fruit production, leaf area index (LAI), and average ETo during the study years 2021 (“OFF”) and 2022 (“ON”) years.
| Experimental year | Fruit production (kg/hectare) | LAI | Average ETo (mm) from DOY 204 to DOY 278 |
|---|---|---|---|
| 2021 | 3430±158 | 2.80 | 5.22±0.13 |
| 2022 | 20400±325* | 2.82ns | 4.83±0.12* |
Significances: *, statistically significant differences (p<0.05), ns: no significant differences (p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



