
Submitted:
26 December 2023
Posted:
27 December 2023
You are already at the latest version
Abstract
Yersiniosis is a common zoonotic enteral disease among humans, which has been linked to pigs and contaminated food, especially pork. Studies on yersiniosis in pets are very scarce. In this study, we performed pheno- and genotypic characterization of 50 Yersinia strains isolated from pets in Finland between 2012 and 2023. Y. enterocolitica 4/O:3/ST135, the most common type in human yersiniosis, was also the most common Y. enterocolitica type (68%) found in clinical faecal samples in our study. Also, human pathogenic Y. enterocolitica 2/O:9/ST139 and Y. pseudotuberculosis O:1/ST9 and O:1/ST42 strains carrying all essential pathogenic genes were identified. Three Y. enterocolitica 4/O:3/ST9 strains were multi-drug resistant and two of them were highly related showing one allelic difference (AD) with core-genome multi-locus sequence typing. Non-pathogenic, genotypically highly diverse Y. enterocolitica 1A strains, showing more than 1000 ADs and missing the essential virulence genes, were also recognized in dogs and cats. Our study demonstrates that pets can excrete human pathogenic Y. enterocolitica and Y. pseudotuberculosis in their faeces and may serve as an infection source for human yersiniosis, especially in families with small children in close contact with their pets.
Keywords:
Yersinia enterocolitica
; Yersinia pseudotuberculosis
; pets
; virulence
; resistance
; characteristics
1. Introduction
Yersinia are gram-negative rods quite recently reclassified into the Yersiniaceae family within the Enterobacterales order [1]. Two species, Yersinia enterocolitica and Y. pseudotuberculosis, are enteropathogens causing enteric yersiniosis in humans and animals [2]. Yersiniosis was the third most reported enteric disease in 2021 within the EU, and the highest detection rate was reported in under five-year-old children [3]. The highest notification rates were reported in Denmark (12.7/100,000 inhabitants) and Finland (7.4/100,000). The infections were primarily caused by Y. enterocolitica and only rarely by Y. pseudotuberculosis. Symptoms range from mild diarrhoea to systemic infection. Typically, the infection is an uncomplicated enteric disease with diarrhoea and abdominal pain and only in some cases do extraintestinal complications occur [2].
Y. enterocolitica is divided into six biotypes (1A, 1B, 2–5) based on biochemical characteristics and into numerous serotypes based on its O antigen. However, only certain biotype and serotype combinations have been associated with human and animal infections. The most common type reported in the EU is bioserotype 4/O:3, followed by 2/O:9 [3]. Biotype 1A strains, regarded as non-pathogenic, are very commonly found especially in non-human samples. They are missing the virulence plasmid (pYV) and essential chromosomal genes needed for pathogenesis. Correctly identified Y. pseudotuberculosis are regarded as pathogenic, carrying virulence genes both on the pYV plasmid and in the chromosome. Y. enterocolitica and Y. pseudotuberculosis strains found in the EU are usually sensitive to most antimicrobials used for human and animal treatment [4,5]. However, Y. enterocolitica has intrinsic resistance to ampicillin due to the blaA gene in its chromosome.
Several possible transmission routes occur for Y. enterocolitica and Y. pseudotuberculosis. Occupational contact with pigs and pork consumption are significantly associated with sporadic Y. enterocolitica infections [6,7]. The consumption of raw or undercooked pork and drinking untreated water are particularly important risk factors. Dogs and cats are potential sources of human yersiniosis through close contact, especially for young children [8]. Pathogenic Y. enterocolitica may be transmitted to dogs and cats via raw pork [9]. However, yersiniosis in dogs and cats has seldom been reported. In an earlier study conducted in Germany, Y. enterocolitica was isolated from 4.6% and 0.3% of dog and cat faecal samples, respectively [10].
To obtain more information concerning Yersinia strains isolated from dogs and cats in Finland, we characterized the strains pheno- and genotypically. We studied the presence of various virulence and resistance genes and the sensitivity to clinically important antimicrobials among pet Yersinia strains to attain more information of their possible relevance to public health. We additionally studied the genetic relationship between the strains using core genome multi-locus sequence typing (cgMLST) to obtain new information on genotypes circulating among dogs and cats.
2. Materials and Methods
2.1. Yersinia enterocolitica and Yersinia pseudotuberculosis strains
We characterized 50 Yersinia strains isolated from pets between 2012 and 2023 at the Veterinary Teaching Hospital of the University of Helsinki (Figure 1). Most (92%, 46/50) strains were from dogs and the rest (8%, 4/50) from cats. In total, 38 (76%) Y. enterocolitica strains (35 from dogs and 3 from cats) and 12 (24%) Y. pseudotuberculosis strains (11 from dogs and one from a cat) were characterized pheno- and genotypically. They were isolated from clinical specimens, mostly (88%, 44/50) from the faeces of patients. Six strains were isolated from extra-intestinal sites: from an abscess (n=2), urine (n=2), blood (n=1), and a wound (n=1).
2.2. Phenotypic characterization of Yersinia strains
Yersinia strains isolated between 2012 and 2017 were identified using API 20E (BioMerieux, France) and, from 2018 onwards, with MALDI-TOF MS. In total, 38 Y. enterocolitica strains were biotyped based on tween esterase activity, indole production, and xylose and trehalose fermentation. All 50 strains were serotyped with commercial antisera (Sifin, Berlin, Germany): 38 Y. enterocolitica strains against O:3 and O:9 and 12 Y. pseudotuberculosis strains against O:1. Phenotypical susceptibility to antimicrobials was studied by a minimum inhibitory concentration (MIC) method (Senititre, EUVSEC3, Thermo Fisher Diagnostic, Vantaa, Finland), which included 15 antimicrobial agents: amikacin, ampicillin, azithromycin, cefotaxime, ceftazidime, chloramphenicol, ciprofloxacin, colistin, gentamicin, meropenem, nalidixic acid, sulfamethoxazole, tetracycline, tigecycline, and trimethoprim. The plates were incubated at 30 °C for 24 h and the MIC values were read by the Sensititre Vizion instrument (Thermo Fisher Diagnostic). Susceptibility thresholds were interpreted in accordance with EUCAST https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_13.1_Breakpoint_Tables.pdf).
2.3. Genotypic characterization of Yersinia strains
Identification and pathogenicity of Yersinia strains were confirmed by PCR (Table 1). For whole-genome sequencing (WGS), the DNA was extracted using QIAcube Connect instrument (Qiagen, Hilden, Germany) with a DNeasy Blood & Tissue Kit (Qiagen, Valencia, CA, US). Yersinia cells for DNA extraction were harvested from 2 mL of tryptic soya broth (Oxoid, Basingstoke, UK) after overnight incubation for 20–22 h at 30 °C. DNA quality (A260/A280 ratio) was measured by a NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, US) and quantity with a Qubit 2.0 fluorometer (Invitrogen, Life Technologies, Carlsbad, CA, US). WGS was performed using Illumina NovaSeq 6000 (Novogene, Cambridge, UK) with 2 x 150 bp read length and targeted genomic coverage of 100 x. The reads were assembled de novo by a Unicycler v0.4.8 assembler available on the PATRIC 3.6.12 platform (https://www.bv-brc.org/app/Assembly). The sequence types (ST) were defined with 7-gene MLST based on the Achtman scheme [11] and the core genome sequence types (CTs) using the CGE (Center for Genomic Epidemiology) platform (https://cge.food.dtu.dk/services/MLST/). Ad hoc cgMLST targeting 3839 genes of Y. enterocolitica and 2886 genes of Y. pseudotuberculosis was performed with Ridom SeqSphere+ software v7.7.5 (Ridom GmbH, Muenster, Germany). The minimum spanning tree (MST), representing pairwise allele distances ignoring missing values of 26 Y. enterocolitica 4/O:3 strains, was used to visualize ADs between the strains. Antimicrobial resistance genes were confirmed by the Ridom software and ResFinder v4.4.1 on the CGE platform.
3. Results
Phenotypic species identification was based on API20E or MALDI-TOF MS. The identification rate (ID%) of API 20E varied between 88.4% and 99.8% for Y. enterocolitica and between 98.6% and 99.9% for Y. pseudotuberculosis. The identification scores of MALDI-TOF MS varied from 2.00 to 2.44 for Y. enterocolitica and from 2.00 to 2.40 for Y. pseudotuberculosis, respectively. All strains had a score over 2.00, indicating probable species identification. Most Y. enterocolitica strains (26/38, 68%) belonged to bioserotype 4/O:3, and four (11%) strains belonged to bioserotype 2/O:9; both types are regarded as pathogenic. Non-pathogenic biotype 1A strains were identified in 8 (21%) cases (Table 2). All 12 Y. pseudotuberculosis strains belonged to serotype O:1. In six (12%) animals, the infection was extra-intestinal. Y. pseudotuberculosis O:1 was identified in five of the six extra-intestinal infections and Y. enterocolitica 1A in one abscess from a dog (Table 2).
All Y. enterocolitica strains belonging to bioserotypes 4/O:3 and 2/O:9 carried the chromosomal ail, inv, and ystA genes, and most (97%, 29/30) of the strains also carried the genes virF and yadA located on the virulence plasmid pYV (Table 3). All 4/O:3 strains belonged to the sequence type ST135 and all 2/O:9 strains to ST139. One 1A strain carried the ail gene associated with virulence. The ystB gene, which is often found in non-pathogenic Yersinia strains, was detected in five 1A strains; however, ystA associated with Yersinia pathogenicity was not detected. All eight 1A strains were also negative for virF and yadA genes located on the pYV plasmid. Each also belonged to an individual sequence type (Table 3). All Y. pseudotuberculosis O:1 strains carried the same virulence genes (ail, inv, irp, and psaA) located in the chromosome. Three (25%) strains missed the virF and yadA genes located on the pYV plasmid (Table 3). Two sequence types (ST9 and ST42) were identified among the Y. pseudotuberculosis O:1 strains.
Most (92%; 35/38) Y. enterocolitica strains were susceptible to 13 out of 15 antimicrobials included in the EUVSEC3 kit. Only intrinsic resistance to ampicillin and streptogramin was observed (Table 4). Three strains expressed high MIC values to sulfamethoxazole, two of which carried the sul1 gene and one carried the sul2 gene. Two strains carried the catA1 gene and were resistant to chloramphenicol, and one strain was resistant to tetracycline and carried the tet(A) gene. Additionally, three strains carried genes coding for streptomycin resistance (Table 4). Y. pseudotuberculosis strains were susceptible to most (14/15) of the tested antimicrobials, only intrinsic resistance to colistin was observed. Three Y. enterocolitica 4/O:3 strains were multi-drug resistant (MDR): two MDR strains showed resistance to chloramphenicol (catA1), streptomycin (aadA12), and sulfamethoxazole (sul1), and one MDR strain showed resistance to streptomycin (aph), sulfamethoxazole (sul2), and tetracycline (tetA).
In total, 17 sequence types (CTs) based on cgMLST were identified among 26 Y. enterocolitica bioserotype 4/O:3/ST135 strains (Table 5). The ADs varied between 1 and 189. Y. enterocolitica bioserotype 2/O:9/ST139 strains three CTs, which differed from each other with 2 to 37 ADs. All eight non-pathogenic Y. enterocolitica 1A strains differed from each other with more than 1600 ADs (Table 5). Y. pseudotuberculosis O:1/ST9 strains differed from each other with 20 to 93 ADs and ST42 strains with one to 78 ADs.
Two clusters including two highly related strains in both clusters were observed among Y. enterocolitica 4/O:3/ST135 strains (Figure 2). Two MDR Y. enterocolitica 4/O:3/ST135 strains belonging to CT69 were highly related with one allelic mismatch, and these were found in dogs from the same litter. Additionally, two strains belonging to CT4483 showing only one AD were isolated from dog faeces sampled in 2019 and 2020. All other strains differed from each other by more than 10 ADs, which was the maximum AD for a cluster definition. Two strains with CT782 among the four bioserotype 2/O:9/ST139 strains formed a small highly related cluster with only 2 ADs between the strains (Table 5). These strains were isolated in 2018 and 2021. The strains belonging non-pathogenic biotype 1A were very heterogenous compared to the pathogenic 4/O:3 and 2/O:9 strains.
4. Discussion
Y. enterocolitica of bioserotype 4/O:3 was the most common yersinia found in dog faeces in our study. We detected the most important chromosomal virulence genes (ail and ystA) in all bioserotype 4/O:3 strains. The virulence plasmid (pYV), which can easily be lost during culturing, was missing in only one strain. This shows that the bioserotype 4/O:3 strains we identified in dogs and cats can be regarded as pathogenic, capable of causing yersinosis. However, canine yersiniosis has barely been investigated. In an earlier study, Fenwick et al. [23] demonstrated that asymptomatic dogs can excrete Y. enterocolitica 4/O:3 in their faeces for weeks. Stamm et al. [10] found ail-positive Y. enterocolitica most frequently in under one-year-old dogs. They also reported bioserotype 4/O:3 to be the most common type in dog faeces, which is in accordance with our study. Bioserotype 4/O:3 has frequently been found in fattening pigs at slaughter, and raw or undercooked pork has been shown to be the main infection source of this type [6]. Raw pork has been demonstrated to be a possible infection source of yersiniosis in dogs and cats [9]. Unfortunately, no information about possible raw feeding was available in our study.
We found Y. enterocolitica bioserotype 2/O:9 in some dogs, which is the second most reported bioserotype in humans in the EU, including Finland [3]. All these strains can be regarded as pathogenic because they carried all the studied essential virulence genes, including genes located on the pYV plasmid. Stamm et al. [10] also reported bioserotype 2/O:9 to be the second most frequently identified type in dog faecal samples. Bioserotype 2/O:9 has sporadically been found in ruminants and wild animals in the EU, including Finland [24,25]; however, the infection sources and transmission routes remain unclear. Our study shows that dogs should be regarded as one source of bioserotype 2/O:9 infections among humans with close dog contact. Also, cats cannot be excluded, but the number of cat strains was very small in our study.
We also identified Y. enterocolitica biotype 1A in dogs and cats, which is regarded as a non-pathogenic type. In one dog faecal strain, we detected the ail gene, which is typically found only in pathogenic strains. However, all strains were ystA- and pYV-negative, which are important virulence factors needed for the pathogenesis of yersiniosis. Stamm et al. [10] also found biotype 1A in some dog faecal samples, and all strains were ail- and pYV-negative. The significance of 1A strains in diarrhoea remains unclear. Interestingly, we identified one biotype 1A strain from an abscess. Biotype 1A is widely distributed in the environment and has been isolated from various sources; thus, cross-contamination cannot be excluded. Biotype 1A strains have also been found in the faecal samples of humans with diarrhoea, especially in Finland [26]. These strains are speculated to have unknown virulence mechanisms and should therefore be regarded as potentially pathogenic. One discussed virulence gene found only in non-pathogenic 1A strains is ystB, which we detected in some of the 1A strains in our study. However, more research is needed concerning the pathogenicity of biotype 1A strains.
Y. pseudotuberculosis O:1 strains were found in many faecal and extra-intestinal pet samples in our study. Serotype O:1 is the most common serotype reported in human and non-human samples worldwide [2]. Serotype O:1 has been identified in several foodborne outbreaks in Finland; however, the transmission routes remain unclear [27,28]. Wild animals, especially rodents and birds, are regarded as the most important reservoirs of Y. pseudotuberculosis [2]. Contaminated fresh produce and raw milk have been identified as the most frequent vehicles in human outbreaks. Outdoor cats can be infected, e.g., by eating birds and rats or by drinking raw milk. However, further studies are needed to clarify the possible infection sources and transmission routes for pets. In our study, Y. pseudotuberculosis O:1 was identified in most extra-intestinal samples.
We only identified ST135 among pathogenic Y. enterocolitica 4/O:3 strains and ST139 among pathogenic 2/O:9 strains using WGS. These sequence types were heterogeneous, showing numerous ADs and several CTs by cgMLST. However, they were more homogeneous compared to the non-pathogenic 1A strains, which all belonged to individual STs and CTs with more than 1600 ADs. This high heterogeneity among the 1A strains can be explained by their large environmental distribution, which makes it challenging to interpret their true relevance in the pathogenesis of yersiniosis. We additionally identified two sequence types, ST9 and ST42, among pathogenic Y. pseudotuberculosis strains, which clearly differed from each other by cgMLST. Both sequence types have been reported in humans and animals in Finland in previous studies [27,28]. Only three small clusters were observed among Y. enterocolitica strains. However, most Y. enterocolitica and Y. pseudotuberculosis strains differed with more than 10 ADs, which was the cut-off for a cluster. This indicates that the yersiniosis cases in our study were sporadic, with no common sources. This was expected because the number of strains was small, and the strains were collected over quite a long period. The next step is to design a prospective epidemiological study to compare pet strains with human and food strains using WGS and to collect comprehensive historical data from pets with yersinosis.
Nearly all Y. enterocolitica strains were susceptible to most of the clinically important microbial agents. Intrinsic ampicillin resistance, which was observed among Y. enterocolitica strains, is linked to the blaA gene located in their chromosome [4]. Also, all Y. pseudotuberculosis strains were very susceptible to most of the studied antimicrobials, and only high MIC values to colistin were observed. Colistin resistance is expected in Y. pseudotuberculosis without the colistin resistance gene and is most likely due to chromosomal mutations conferring the intrinsic resistance [29]. MDR Y. enterocolitica strains have rarely been found in Europe except in Spain and Italy [5]. We could identify two MDR Y. enterocolitica 4/O:3/ST135/CT69 strains from dogs that carried resistance genes against chloramphenicol (catA1), streptomycin (aadA1), and sulfonamides (sul1). These genes have recently been reported in Y. enterocolitica 4/O:3 outbreak strains in Sweden [30]. The two strains in our study were highly related, and they originated from two dogs from the same litter, indicating a common infection source. Unfortunately, this is the only historical information we have, and raw feeding cannot be excluded. The third MDR strain (Y. enterocolitica 4/O:3/ST135/CT1010) was resistant to sulfonamides (sul2), streptomycin (aph3-ib and aph6-id), and tetracycline (tetA), which are antimicrobials commonly used to treat animals.
Most (84%) of the Yersinia strains originated from dog faeces, and only three (6%) strains were from cat faeces. One reason may be that cat faecal samples were more rarely investigated than dog samples or that cats more rarely suffer from diarrhoea. We know that raw feeding of pets is a risk factor for developing diarrhoea, and that Y. enterocolitica 4/O:3 is associated with raw pork [9,31]. We also know that young dogs are more prone to diarrhoea caused by Y. enterocolitica. To obtain more information about the incidence of enteric yersinosis in pets and to investigate any possible associations between animal age, feeding and yersiniosis in dogs and cats, a prospective study should be conducted.
5. Conclusions
We showed that enteropathogenic Yersinia can be present in clinical faecal samples of dogs and cats. Therefore, yersiniosis should be included in the differential diagnosis of enteritis in pets. The most frequently identified Y. enterocolitica type was bioserotype 4/O:3, which is also the most common type among human yersiniosis. A possible link between human and pet yersiniosis needs to be studied further. According to the WGS analyses used, Yersinia-positive cases in our study were sporadic without a clear relationship with each other. However, yersiniosis should be kept in mind, especially when diarrhoea occurs in a litter fed with raw meat.
Author Contributions
Conceptualisation: all authors; Investigation: MFA, TG, VH and VJ; Methodology; MFA, MR, AH and RLN; Software: TG, VH and VJ; Visualisation: MFA and VH; Validation: all authors; Writing-original draft preparation: MFA; Writing-review and editing: all authors; Resources: MFA, TG and AH; Project administration and funding acquisition: MFA. All authors have read and agreed to be published version of the manuscript.
Funding
This study was partly funded by the Finnish Foundation of Veterinary Research. Proofreading was provided by the University of Helsinki.
Data Availability Statement
The data analysed during the study are available from the corresponding author on reasonable request.
Acknowledgments
Maria Stark and Kirsi Ristkari are gratefully acknowledged for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-Based Phylogeny and Taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales Ord. Nov. Divided into the Families Enterobacteriaceae, Erwiniaceae Fam. Nov., Pectobacteriaceae Fam. Nov., Yersiniaceae Fam. Nov., Hafniaceae Fam. Nov., Morganellaceae Fam. Nov., and Budviciaceae Fam. Nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson-Ahomaa, M. Enteropathogenic Yersinia Spp. In Zoonoses: Infections Affecting Humans and Animals; Sing, A., Ed.; Springer International Publishing: Cham, 2023; pp. 329–353. ISBN 978-3-031-27164-9. [Google Scholar]
- EFSA; ECDC The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [CrossRef]
- Bonke, R.; Wacheck, S.; Stüber, E.; Meyer, C.; Märtlbauer, E.; Fredriksson-Ahomaa, M. Antimicrobial Susceptibility and Distribution of β-Lactamase A (BlaA) and β-Lactamase B (BlaB) Genes in Enteropathogenic Yersinia Species. Microb. Drug Resist. 2011, 17, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, J.; Ortiz-Martínez, P.; Keto-Timonen, R.; Joutsen, S.; Fredriksson-Ahomaa, M.; Korkeala, H. Prudent Antimicrobial Use Is Essential to Prevent the Emergence of Antimicrobial Resistance in Yersinia Enterocolitica 4/O: 3 Strains in Pigs. Front. Microbiol. 2022, 13, 841841. [Google Scholar] [CrossRef] [PubMed]
- Guillier, L.; Fravalo, P.; Leclercq, A.; Thébaut, A.; Kooh, P.; Cadavez, V.; Gonzales-Barron, U. Risk Factors for Sporadic Yersinia Enterocolitica Infections: A Systematic Review and Meta-Analysis. Microb. Risk Anal. 2020, 100141. [Google Scholar] [CrossRef]
- Rosner, B.; Stark, K.; Höhle, M.; Werber, D. Risk Factors for Sporadic Yersinia Enterocolitica Infections, Germany 2009–2010. Epidemiol. Infect. 2012, 140, 1738–1747. [Google Scholar] [CrossRef]
- Boqvist, S.; Pettersson, H.; Svensson, Å.; Andersson, Y. Sources of Sporadic Yersinia Enterocolitica Infection in Children in Sweden, 2004: A Case-Control Study. Epidemiol. Infect. 2009, 137, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson-Ahomaa, M.; Korte, T.; Korkeala, H. Transmission of Yersinia Enterocolitica 4/O:3 to Pets via Contaminated Pork. Lett. Appl. Microbiol. 2001, 32, 375–378. [Google Scholar] [CrossRef]
- Stamm, I.; Hailer, M.; Depner, B.; Kopp, P.A.; Rau, J. Yersinia Enterocolitica in Diagnostic Fecal Samples from European Dogs and Cats: Identification by Fourier Transform Infrared Spectroscopy and Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2013, 51, 887–893. [Google Scholar] [CrossRef]
- Laukkanen-Ninios, R.; Didelot, X.; Jolley, K.A.; Morelli, G.; Sangal, V.; Kristo, P.; Brehony, C.; Imori, P.F.; Fukushima, H.; Siitonen, A. Population Structure of the Yersinia Pseudotuberculosis Complex According to Multilocus Sequence Typing. Environ. Microbiol. 2011, 13, 3114–3127. [Google Scholar] [CrossRef]
- Sen, K. Rapid Identification of Yersinia Enterocolitica in Blood by the 5′ Nuclease PCR Assay. J. Clin. Microbiol. 2000, 38, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, H.; Hensel, A.; Aleksic, S.; Meyer, H. Identification of Yersinia Enterocolitica within the Genus Yersinia. Syst. Appl. Microbiol. 2000, 23, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.N.; Rasmussen, O.F.; Andersen, J.K.; Olsen, J.E. Specific Detection of Pathogenic Yersinia Enterocolitica by Two-Step PCR Using Hot-Start and DMSO. Mol. Cell. Probes 1994, 8, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Inoue, M.; Mori, T.; Itoh, K.; Arakawa, E.; Watanabe, H. Detection and Identification of Yersinia Pseudotuberculosis and Pathogenic Yersinia Enterocolitica by an Improved Polymerase Chain Reaction Method. J. Clin. Microbiol. 1992, 30, 2484–2486. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, S.T.; Nilsson, C.; Hallanvuo, S.; Lindblad, M. Real-Time PCR Method for Detection of Pathogenic Yersinia Enterocolitica in Food. Appl. Environ. Microbiol. 2008, 74, 6060–6067. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, S.T.; Nilsson, C.; Hallanvuo, S. TaqMan-Based Real-Time PCR Method for Detection of Yersinia Pseudotuberculosis in Food. Appl. Environ. Microbiol. 2008, 74, 6465–6469. [Google Scholar] [CrossRef]
- Thoerner, P.; Bin Kingombe, C.; Bogli-Stuber, K.; Bissig-Choisat, B.; Wassenaar, T.; Frey, J.; Jemmi, T. PCR Detection of Virulence Genes in Yersinia Enterocolitica and Yersinia Pseudotuberculosis and Investigation of Virulence Gene Distribution. Appl. Environ. Microbiol. 2003, 69, 1810–1816. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, H.; Tsunomori, Y.; Seki, R. Duplex Real-Time SYBR Green PCR Assays for Detection of 17 Species of Food-or Waterborne Pathogens in Stools. J. Clin. Microbiol. 2003, 41, 5134–5146. [Google Scholar] [CrossRef] [PubMed]
- Wren, B.; Tabaqchali, S. Detection of Pathogenic Yersinia Eriterocolitica by the Polymerase Chain Reaction. The Lancet 1990, 336, 693. [Google Scholar] [CrossRef]
- Weynants, V.; Jadot, V.; Denoel, P.A.; Tibor, A.; Letesson, J.-J. Detection of Yersinia Enterocolitica Serogroup O: 3 by a PCR Method. J. Clin. Microbiol. 1996, 34, 1224–1227. [Google Scholar] [CrossRef]
- Jacobsen, N.; Bogdanovich, T.; Skurnik, M.; Lübeck, P.; Ahrens, P.; Hoorfar, J. A Real-Time PCR Assay for the Specific Identification of Serotype O: 9 of Yersinia Enterocolitica. J. Microbiol. Methods 2005, 63, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, S.G.; Madie, P.; Wilks, C.R. Duration of Carriage and Transmission of Yersinia Enterocolitica Biotype 4, Serotype 0:3 in Dogs. Epidemiol. Infect. 1994, 113, 471–477. [Google Scholar] [CrossRef]
- Joutsen, S.; Laukkanen-Ninios, R.; Henttonen, H.; Niemimaa, J.; Voutilainen, L.; Kallio, E.R.; Helle, H.; Korkeala, H.; Fredriksson-Ahomaa, M. Yersinia Spp. in Wild Rodents and Shrews in Finland. Vector-Borne Zoonotic Dis. 2017, 17, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Joutsen, S.; Eklund, K.-M.; Laukkanen-Ninios, R.; Stephan, R.; Fredriksson-Ahomaa, M. Sheep Carrying Pathogenic Yersinia Enterocolitica Bioserotypes 2/O:9 and 5/O:3 in the Feces at Slaughter. Vet. Microbiol. 2016, 197, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Sihvonen, L.M.; Jalkanen, K.; Huovinen, E.; Toivonen, S.; Corander, J.; Kuusi, M.; Skurnik, M.; Siitonen, A.; Haukka, K. Clinical Isolates of Yersinia Enterocolitica Biotype 1A Represent Two Phylogenetic Lineages with Differing Pathogenicity-Related Properties. BMC Microbiol. 2012, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Blomvall, L.; Pelkola, K.; Lienemann, T.; Lehtoniemi, S.; Pohjola, L.; Fredriksson-Ahomaa, M. Osteomyelitis in a Slaughter Turkey Flock Caused by Yersinia Pseudotuberculosis Sequence Type ST42. Vet. Microbiol. 2022, 269, 109424. [Google Scholar] [CrossRef] [PubMed]
- Castro, H.; Jaakkonen, A.; Hakakorpi, A.; Hakkinen, M.; Isidro, J.; Korkeala, H.; Lindström, M.; Hallanvuo, S. Genomic Epidemiology and Phenotyping Reveal On-Farm Persistence and Cold Adaptation of Raw Milk Outbreak-Associated Yersinia Pseudotuberculosis. Front. Microbiol. 2019, 10, 1049. [Google Scholar] [CrossRef] [PubMed]
- Cabanel, N.; Galimand, M.; Bouchier, C.; Chesnokova, M.; Klimov, V.; Carniel, E. Molecular Bases for Multidrug Resistance in Yersinia Pseudotuberculosis. Int. J. Med. Microbiol. 2017, 307, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, P.A.; Tano, E.; Jernberg, C.; Hickman, R.A.; Guy, L.; Järhult, J.D.; Wang, H. Molecular Characterization of Multidrug-Resistant Yersinia Enterocolitica from Foodborne Outbreaks in Sweden. Front. Microbiol. 2021, 12, 664665. [Google Scholar] [CrossRef]
- Fredriksson-Ahomaa, M.; Heikkilä, T.; Pernu, N.; Kovanen, S.; Hielm-Björkman, A.; Kivistö, R. Raw Meat-Based Diets in Dogs and Cats. Vet. Sci. 2017, 4, 33. [Google Scholar] [CrossRef]
Figure 1.
Number of Y. enterocolitica (Ye) and Y. pseudotuberculosis (Yp) strains from pets isolated between 2012 and 2023 in Finland. *Distribution of four cat strains
Figure 1.
Number of Y. enterocolitica (Ye) and Y. pseudotuberculosis (Yp) strains from pets isolated between 2012 and 2023 in Finland. *Distribution of four cat strains
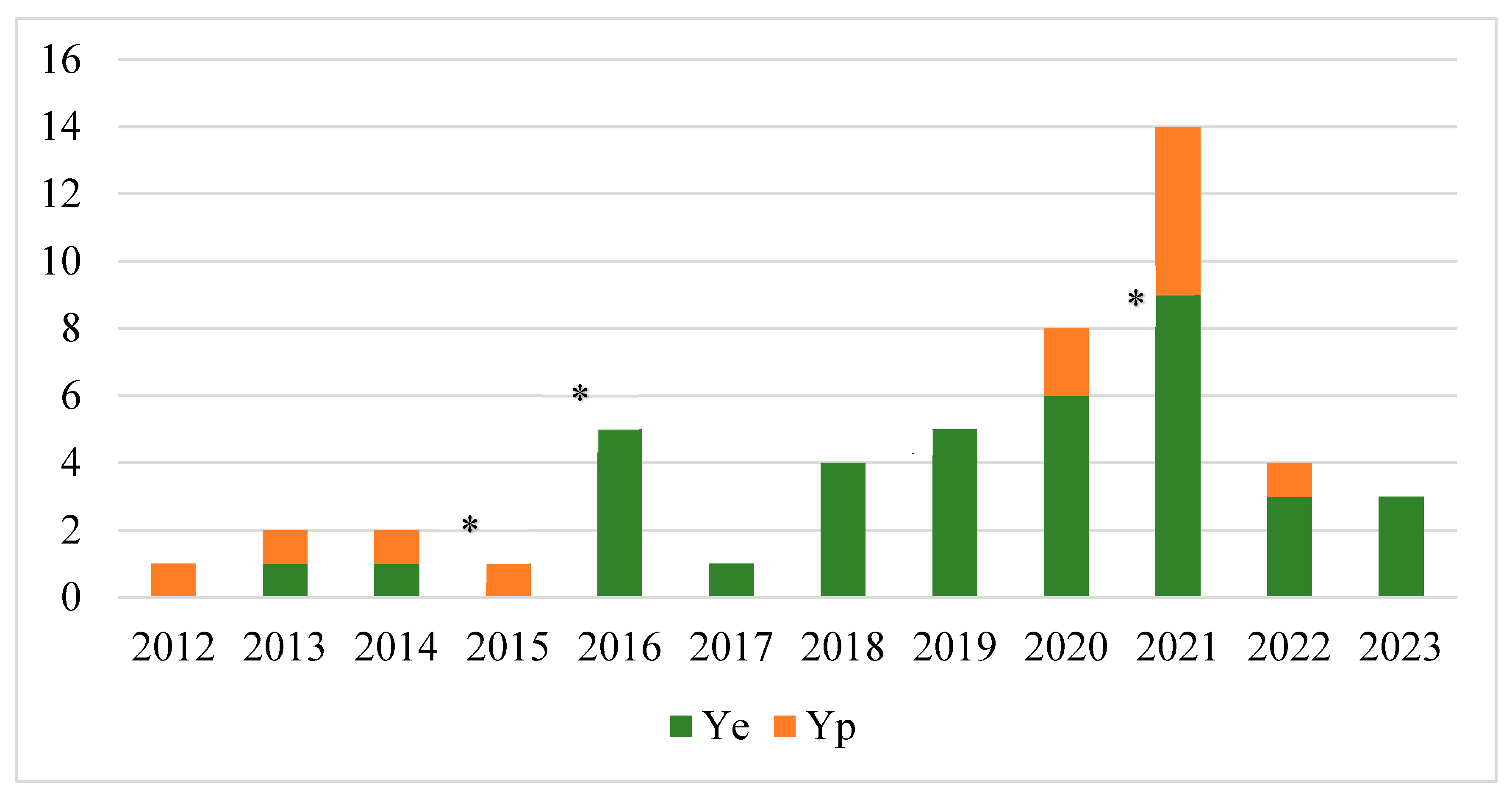
Figure 2.
Minimum spanning tree (MST) of 26 Yersinia enterocolitica 4/O:3 strains based on cgMLST (3839 targets). Number of allelic differences (ADs) between the strains are indicated on the connecting lines. Nodes are numbered with sequence types (CTs) according to the Table 5. Clusters are shaded in grey and a cluster distance threshold of maximum 10 ADs was used.
Figure 2.
Minimum spanning tree (MST) of 26 Yersinia enterocolitica 4/O:3 strains based on cgMLST (3839 targets). Number of allelic differences (ADs) between the strains are indicated on the connecting lines. Nodes are numbered with sequence types (CTs) according to the Table 5. Clusters are shaded in grey and a cluster distance threshold of maximum 10 ADs was used.
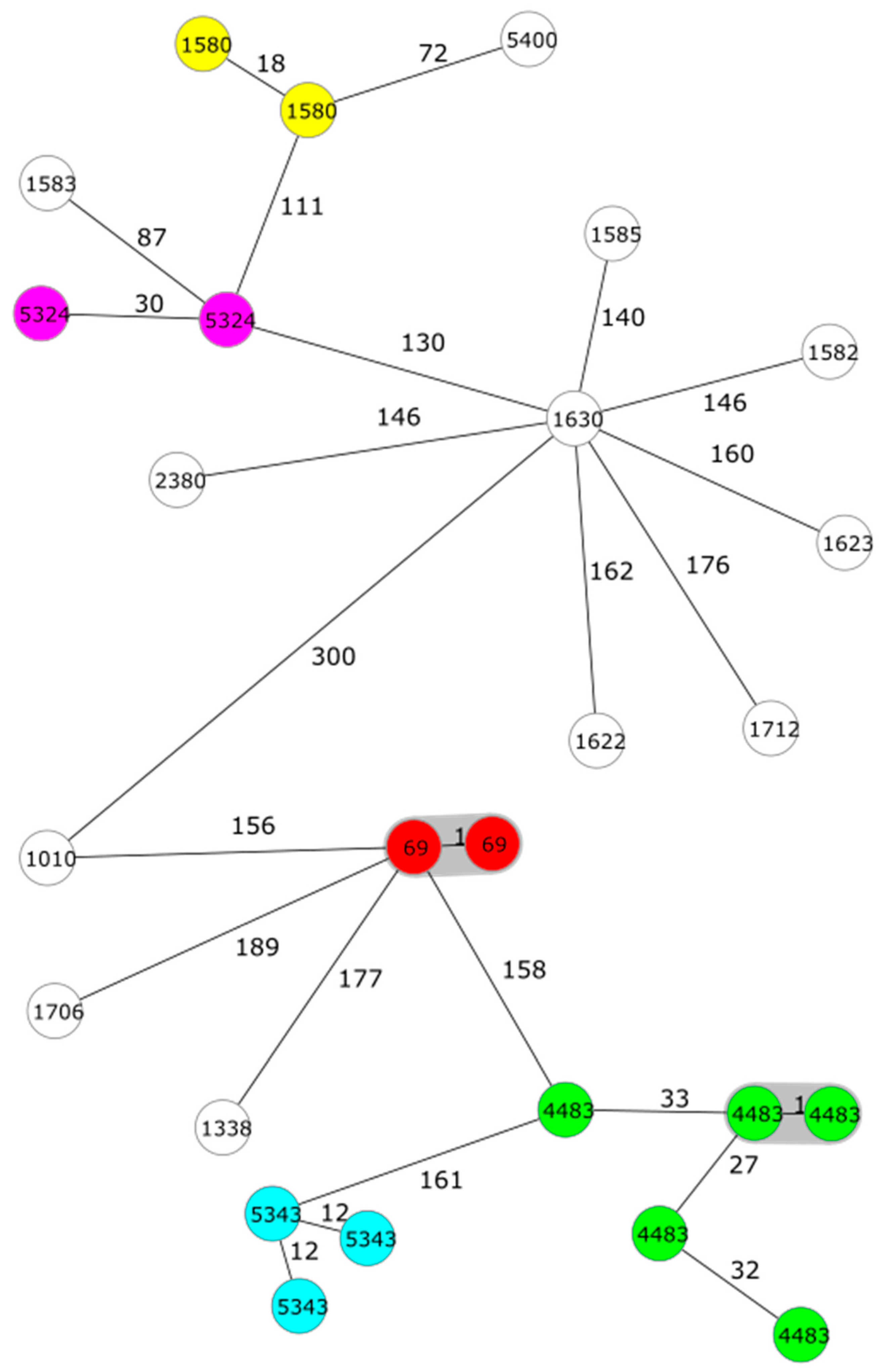

Table 1.
Target genes used for identification and characterization of Y. enterocolitica (Ye) and Y. pseudotuberculosis (Yp) strains.
Table 1.
Target genes used for identification and characterization of Y. enterocolitica (Ye) and Y. pseudotuberculosis (Yp) strains.
| Identification | Target | Gene | Ye | Yp | Reference |
|---|---|---|---|---|---|
| Species | 16SrRNA | rrn | x | x | [12] |
| rrn | x | [13] | |||
| Pathogenicity | Chromosome | inv | x | [14] | |
| inv | x | [15] | |||
| ail | x | [16] | |||
| ail | x | [17] | |||
| ystA | x | [18] | |||
| ystB | x | [18] | |||
| Plasmid | yadA | x | x | [19] | |
| virF | [20] | ||||
| Serotype | O:3 | rfbC | [21] | ||
| O:9 | per | [22] |
Table 2.
Phenotypic characteristics of 38 Y. enterocolitica (Ye) and 12 Y. pseudotuberculosis (Yp) strains from dogs and cats.
Table 2.
Phenotypic characteristics of 38 Y. enterocolitica (Ye) and 12 Y. pseudotuberculosis (Yp) strains from dogs and cats.
| Animal | Numberof strains | Sampletype | Ye | Yp | |
| Biotype | Serotype | Serotype | |||
| Dog | 6 | Faeces (5), abscess (1) | 1A | NTa | |
| 4 | Faeces | 2 | O:9 | ||
| 25 | Faeces | 4 | O:3 | ||
| 11 | Faeces (7), urine (2),abscess (1), blood (1) | O:1 | |||
| Cat | 2 | Faeces | 1A | NT | |
| 1 | Faeces | 4 | O:3 | ||
| 1 | Wound | O:1 | |||
a not typed.
Table 3.
Virulence genes present in 38 Y. enterocolitica (Ye) and 12 Y. pseudotuberculosis (Yp) strains belonging to different sequence types.
Table 3.
Virulence genes present in 38 Y. enterocolitica (Ye) and 12 Y. pseudotuberculosis (Yp) strains belonging to different sequence types.
| Sp. | Type | No. ofstrains | Virulence gene | Sequencetypea | ||||||
| ail | inv | irp | psaA | ystA | ystB | virF/yadA | ||||
| Ye | 1A | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 388 |
| 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 138 | ||
| 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 290, 738 | ||
| 4 | 0 | 4 | 0 | 0 | 0 | 4 | 0 | 148, 179, 365,479/551 | ||
| 2/O:9 | 4 | 4 | 4 | 0 | 4 | 4 | 0 | 4 | 139 | |
| 4/O:3 | 26 | 26 | 26 | 0 | 26 | 26 | 0 | 25 | 135 | |
| Yp | O:1 | 3 | 3 | 3 | 3 | 3 | 0 | 0 | 2 | 9 |
| 9 | 9 | 9 | 9 | 9 | 0 | 0 | 7 | 42 | ||
a 7-gene multi-locus sequence typing (MLST) based on the Achtman scheme.
Table 4.
The minimum inhibitory concentration (MIC) values and antimicrobial resistance (AMR) genes among Yersinia strains isolated from dogs and cats.
Table 4.
The minimum inhibitory concentration (MIC) values and antimicrobial resistance (AMR) genes among Yersinia strains isolated from dogs and cats.
| Antimicrobial agent | Numberof strains | MIC (µg/mL) | AMR genepresent | |
| Breakpoint S ≤ | Observedvalue | |||
| Y. enterocolitica (n=38) | ||||
| Ampicillin | 38 | 8 | ≥32a | blaA |
| Chloramphenicol | 2 | 8 | >64 | catA1 |
| Streptomycin | 2 | NTb | aadA12 | |
| 1 | NT | aph(3)-lb, aph(6)-ld | ||
| Streptogramin | 38 | NT | vat(F) | |
| Sulfamethoxazole | 2 | NAc | >512 | sul1 |
| 1 | NA | >512 | sul2 | |
| Tetracycline | 1 | 4 | >32 | tet(A) |
| Y. pseudotuberculosis (n=12) | ||||
| Colistin | 12 | 2 | >16a | 0 |
a intrinsic resistance; b not tested; c not available.
Table 5.
Sequence types and allelic differences (ADs) of Y. enterocolitica (Ye) and Y. pseudotuberculosis (Yp) strains based on 7-gene multi-locus sequence typing (MLST) (ST) and core genome MLST (CT).
Table 5.
Sequence types and allelic differences (ADs) of Y. enterocolitica (Ye) and Y. pseudotuberculosis (Yp) strains based on 7-gene multi-locus sequence typing (MLST) (ST) and core genome MLST (CT).
| Species | Type | ST | CTs (number of strains) | ADs |
|---|---|---|---|---|
| Ye | 4/O:3 | 135 | 69 (2), 1580 (2), 4483 (5), 5324 (2), 5343 (3) | 1–50 |
| 1583, 5400 | 51–100 | |||
| 1010, 1338, 1582, 1585, 1622, 1623, 1630, 1706, 1712, 2380 | 101–189 | |||
| 2/O:9 | 139 | 510 (2), 904, 5351 | 2-37 | |
| 1A | 138 | 5575 | 2025 | |
| 148 | 1401 | 1661 | ||
| 179 | 1295 | 1661 | ||
| 290 | 1379 | 2030 | ||
| 365 | 2282 | 2067 | ||
| 388 | 1710 | 2164 | ||
| 479/551 | 5505 | 2650 | ||
| 738 | 5332 | 2025 | ||
| Yp | O:1 | 9 | 782 (2), 806 | 20–93 |
| 42 | 816, 1728, 3900 (2), 5560 (3), 5610 (2) | 1–78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



