Submitted:
25 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Jamoytius kerwoodi, is a primitive, eel-like jawless vertebrate found uniquely in an Early Silurian (Llandovery epoch; 444-433 Ma) horizon near Lesmahagow, Scotland. This species is a rare component of a low-diversity dominantly nektonic detritus and herbivorous fauna living over an anoxic bottom and is found at the transition from a marine-influenced, probably brackish-water, deep-water basin to a shallower-water, less saline and likely freshwater basin. In the absence of true teeth, Jamoytius was probably a detritivore or herbivore feeding on Ceratiocaris. Jamoytius may be a direct ancestor of living lampreys, especially as their ectoparasitic mode of life might have evolved from ancestral microphagous filterers or herbivores.

Keywords:
Jamoytius
; lamprey
; herbivore
; detritivore
; Silurian
1. Introduction
Jamoytius kerwoodi White was a primitive, eel-like jawless fish that lived in the Llandovery epoch (444-433 Ma) of the Early Silurian period (White, 1946) (Figure 1). The fossil is preserved as rare carbonized films on bedding planes in one laminated siltstone horizon in the bank of the Logan water in the Lesmahagow inlier of Lanarkshire, SW Scotland (Ritchie, 1968a). It was once considered the most primitive known vertebrate (White, 1946), but with additional studies, its affinities are now debatable (Janvier, 1996, 2008; Sansom et al., 2010; Keating and Donaghue, 2016; Chevrinais et al., 2018; Miyashita et al., 2019, 2021). Because the interpretations of the even such exceptionally preserved soft-bodied fossils is difficult, observed features can be interpreted in different ways (Reeves and Sansom, 2023) (Figure 2). Various cladistic analyses of Jamoytius with other jawless vertebrates, using different character codings, give divergent results (Janvier 1981; Forey, 1984, 1995; Forey and Janvier 1993; Turner, 2004; Miyashiro et al., 2019; Reeves et al., 2023). Choice of the in-group taxa affects its placement (Donoghue et al., 2003; Gess et al. 2006; Reves et al., 2023). The position of Jamoytius on cladograms has consequently not stabilized, though it often appears as a sister taxon to the lampreys, the anaspids, or euphaneropids (Janvier, 1981; 2008; Ritchie, 1984; Janvier and Arsenault, 2007; Donoghue and Keating, 2014; Reeves et al., 2023) (Figure 3).
As a sister taxon to the lampreys, Jamoytius has been compared with parasitic lampreys which attack fish. But, only 18 of the 38 known species of lamprey, are carnivorous. Non-carnivorous lampreys are smaller (less than 40cm long) than the parasitic sea lampreys (35-120cm long) (Potter et al., 2015). The size and anatomy of Jamoytius is thus more compatible with non-carnivorous lampreys, though Priscomyzon riniensis, the oldest undoubted lamprey, from the Devonian (419-359 Ma) of South Africa, is also very small (Gess et al., 2006). The fossil evidence for early evolution of lampreys is scanty because only four undoubted Palaeozoic parasitic lamprey species have been recorded, the above Devonian Priscomyzon, and three from the Carboniferous (359-299 Ma). These lack, however, the specialised, heavily toothed discs with plate-like laminae present in modern lampreys, and it is possible that they were grazers, scraping algae off surfaces (Wu et al., 2023). Furthermore, their post-Paleozoic fossil record of lampreys is equally bad. Based on both morphological and molecular evidence, Brownstein and Near (2023) estimated that 90% of living lamprey clades originated only since the late Cretaceous. As reconstructed by Reeves and Sansom (2023), carnivorous lampreys evolved from non-carnivorous early Paleozoic forms and then radiated from the late Cretaceous times (~100Ma) and especially from Miocene times (~25 Ma) onwards into many both carnivorous and non-carnivorous forms.
As a sister taxon to the anaspids, Jamoytius resembles the genera, Loganiella, Birkenia, and, especially, Lasanius (Blom and Märss, 2010) (Figure 2B and Figure 4). Loganiella also occurs in the Jamoytius bed, while Birkenia and Lasanius occur in slightly younger fish beds in the Lesmahagow succession (Lovelock, 1998; Dineley, 1999).
As a sister taxon to the Upper Devonian, Euphanerops longaevus, (though many of the structures observed on the available fossils remain unexplained) (Janvier and Arsenault, 2007) (Figure 6 A, B), Jamoytius was originally classed with Euphanerops in the Jamoytiiformes (Tarlo 1967) (Figure 3C). Several other euphaneropids have now been recognized: one, Ciderius cooperi van der Brugghen from the fish beds above the Jamoytius bed at Lesmahagow (van der Brugghen, 2015). These are similar to Jamoytius, both in anatomy and possibly mode of life (Figure 6C).
The mode of life of Jamoytius kerwoodi is thus unresolved; even its life orientation is still not certain (Sansom et al., 2010). In this paper I am not particularly concerned with its affinities, but with its mode of life as inferred from its anatomy (which bears, of course, on its affinities), adaptative morphology and palaeoenvironment based on the sedimentology of the enclosing strata and the life styles of it and its associated biota.
Figure 1.
Jamoytius reconstructions: A) with ventral ‘lamprey’ mouth (wuht permission from Nobu Tamura); B) with terminal suspension/detritus feeding mouth (Lingham-Soliar, 2014, fig. 2.11.
Figure 1.
Jamoytius reconstructions: A) with ventral ‘lamprey’ mouth (wuht permission from Nobu Tamura); B) with terminal suspension/detritus feeding mouth (Lingham-Soliar, 2014, fig. 2.11.
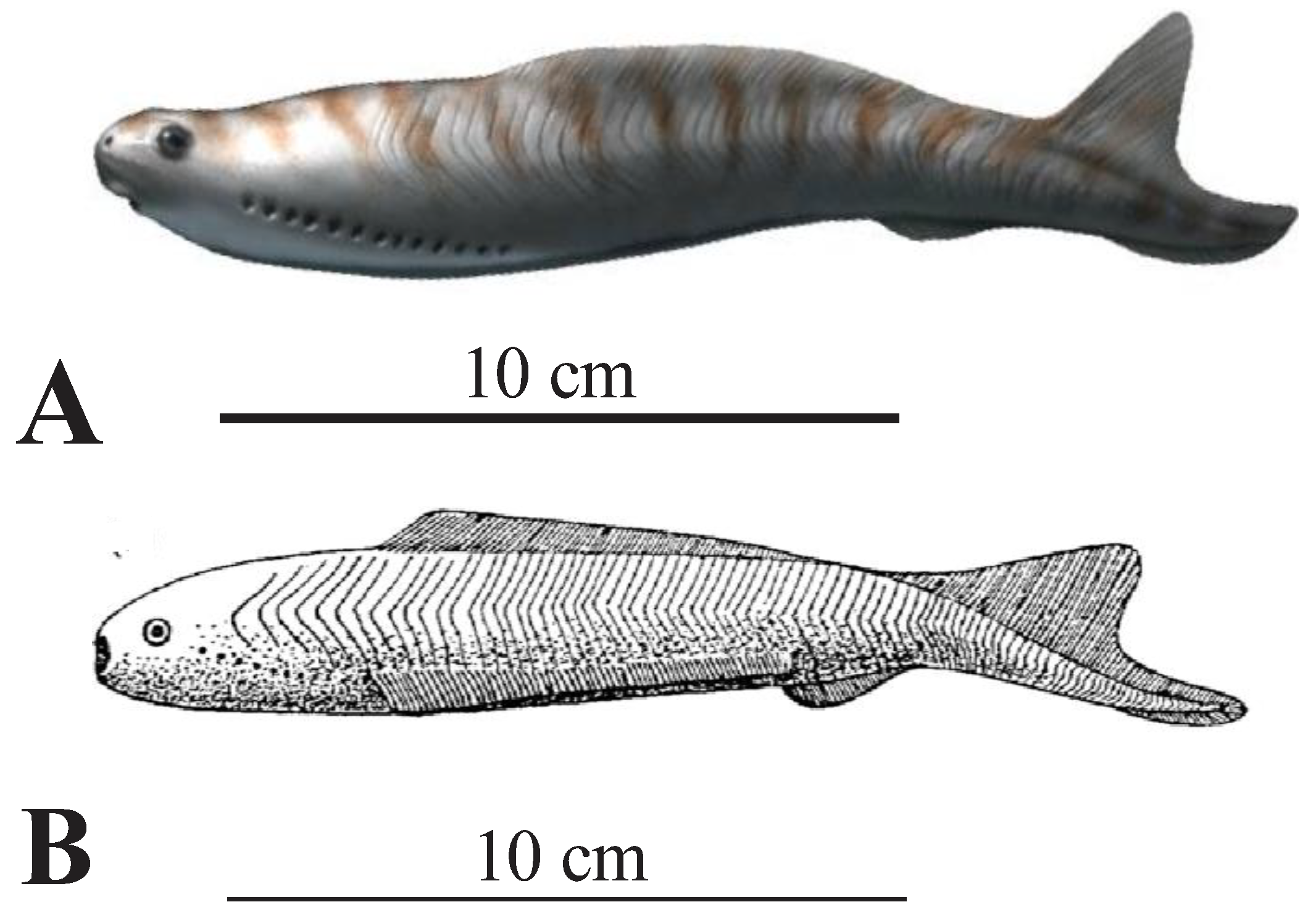
Figure 2.
Jamoytius fossil and inferred features: A) with the conflicting interpretations of White (1946), in bold, and Ritchie (1960, 1963, 1968, 1984) in plain italics (modified from Sansom et al. 2010, Plate 1); B) body parts and topological interpretation of the holotype (NHM P11284a) (from Sansom et al., 2010, text fig. 5).
Figure 2.
Jamoytius fossil and inferred features: A) with the conflicting interpretations of White (1946), in bold, and Ritchie (1960, 1963, 1968, 1984) in plain italics (modified from Sansom et al. 2010, Plate 1); B) body parts and topological interpretation of the holotype (NHM P11284a) (from Sansom et al., 2010, text fig. 5).
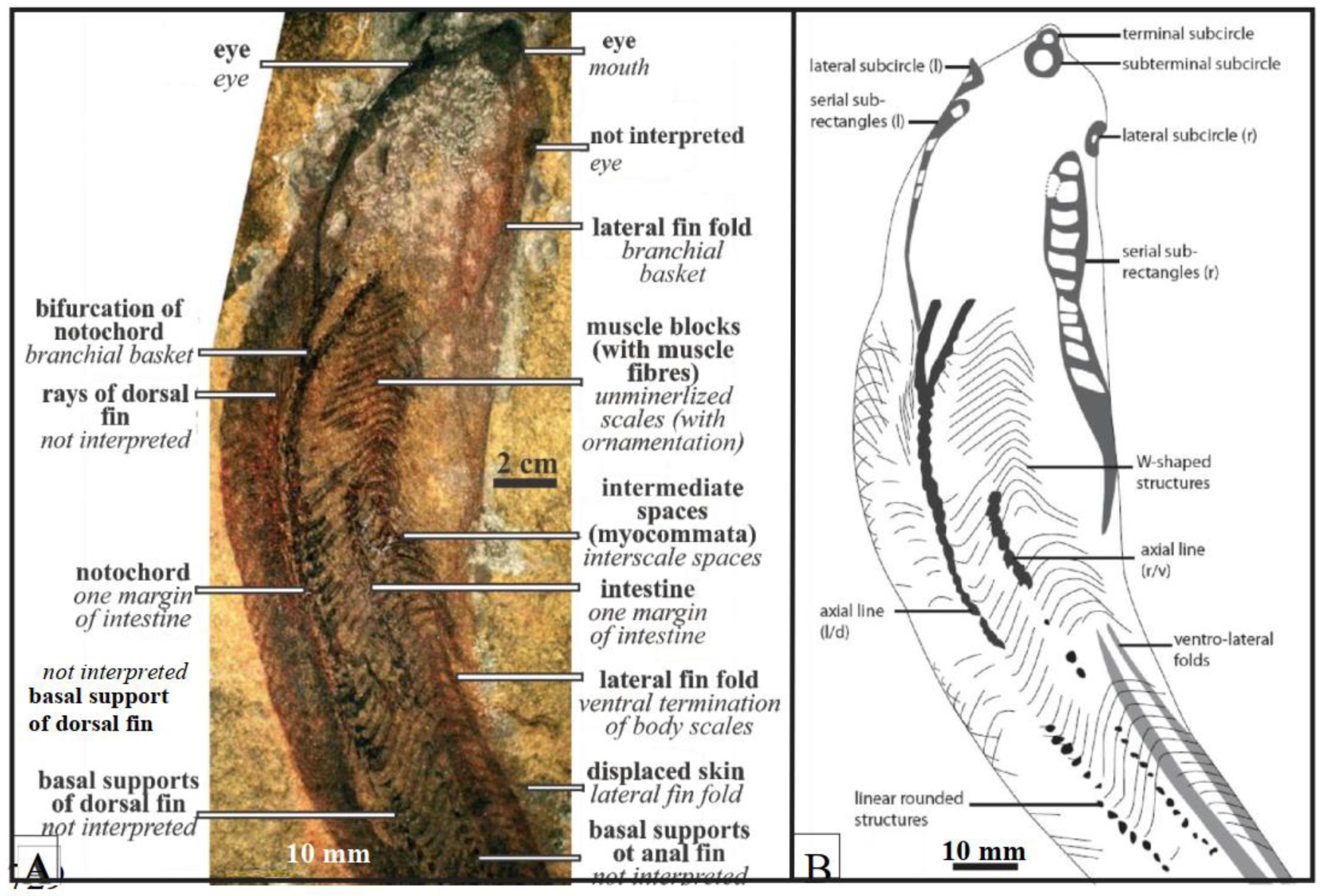
Figure 3.
Examples of cladistic analyses showing three interpretations for Jamoytius (from Sansom et al., 2010).
Figure 3.
Examples of cladistic analyses showing three interpretations for Jamoytius (from Sansom et al., 2010).
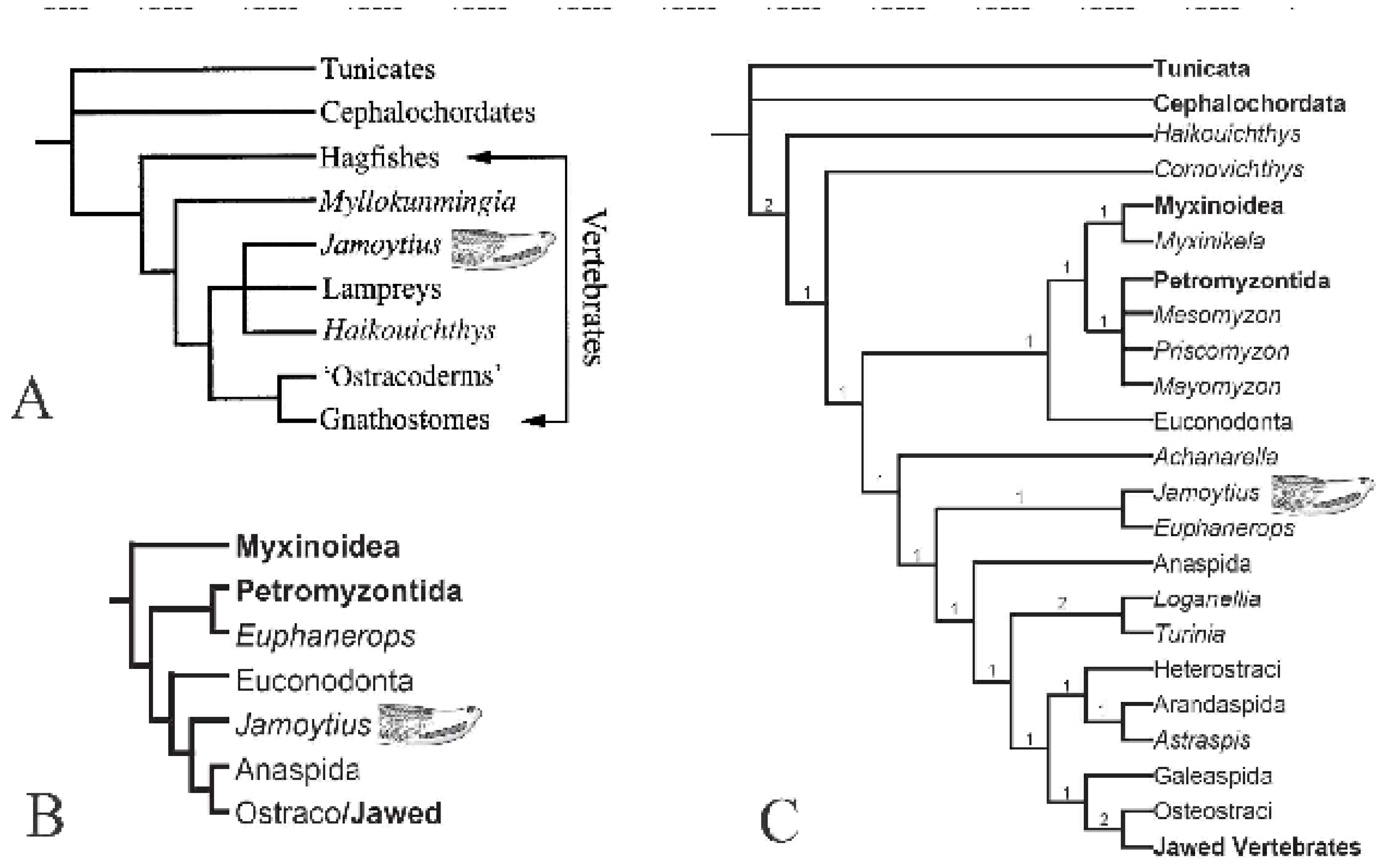
Figure 4.
Living and oldest fossil (Devonian) lampreys. A) parasitic sea lamprey (Petromyzon marinus Linnaeus), 35-60 cm long; B) American brook lamprey (Lethenteron appendix DeKay) 15-25cm long (A&B courtesy of North Carolina Wildlife Resources Commission; C) Devonian parasitic fossil lamprey (Priscomyzon riniensis Gess et al., ~5 cm long (public domain).
Figure 4.
Living and oldest fossil (Devonian) lampreys. A) parasitic sea lamprey (Petromyzon marinus Linnaeus), 35-60 cm long; B) American brook lamprey (Lethenteron appendix DeKay) 15-25cm long (A&B courtesy of North Carolina Wildlife Resources Commission; C) Devonian parasitic fossil lamprey (Priscomyzon riniensis Gess et al., ~5 cm long (public domain).

Figure 5.
Agnathan reconstructions: A) Thelodont Loganellia scotica (permission of Opal Raptor); B) Anaspid Birkenia (permission of Highlander Fossils, www.highlanderfossils.com; C) Anaspid? Lasanius (permission of Rob Van Assen, Museon Omniversum, Den Haag).
Figure 5.
Agnathan reconstructions: A) Thelodont Loganellia scotica (permission of Opal Raptor); B) Anaspid Birkenia (permission of Highlander Fossils, www.highlanderfossils.com; C) Anaspid? Lasanius (permission of Rob Van Assen, Museon Omniversum, Den Haag).
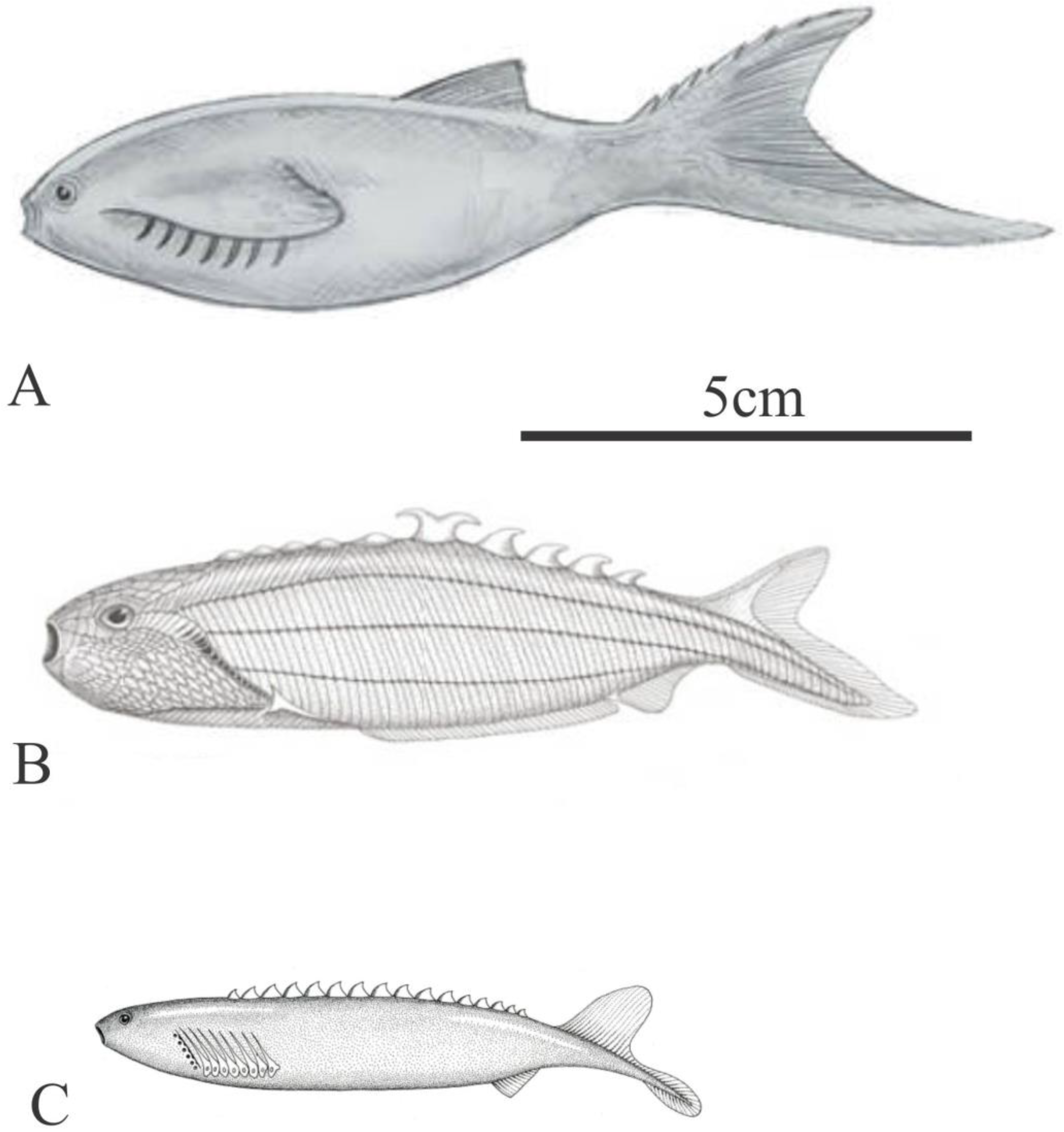
Figure 6.
A) Euphanerops longaevus reconstruction (Upper Devonian, Canada) (from Phillipe Janvier in Parc Nationale de Miguasha 2003); B) Euphanerops longaevus as a swimming nektonic detritivore/ herbivore (with permission from Nobu Tamura); C) various euphaneropds; Ciuderius couperi, (Lower Silurian), Achanarella trewinii, Cornovichthys blaaeweni (Middle Devonian), Endiolepis aneri (Upper Devonian) (with permission from Nobu Tamura).
Figure 6.
A) Euphanerops longaevus reconstruction (Upper Devonian, Canada) (from Phillipe Janvier in Parc Nationale de Miguasha 2003); B) Euphanerops longaevus as a swimming nektonic detritivore/ herbivore (with permission from Nobu Tamura); C) various euphaneropds; Ciuderius couperi, (Lower Silurian), Achanarella trewinii, Cornovichthys blaaeweni (Middle Devonian), Endiolepis aneri (Upper Devonian) (with permission from Nobu Tamura).

Anatomy
Because the preservation of soft-bodied organism like Jamoytius is so variable, and because there are often so few fossils of them preserved, then even their basic anatomy is subject to different interpretations, leading to radically different reconstructions and affinities (Reeves and Sansom, 2023).
Jamoytius had an elongated body, ranging in size from 14-18cm long by 3-4 cm wide, a cartilaginous skeleton, a branchial basket resembling that of the cyclostomes, and weakly mineralized scales (Sansom et al., 2010). Earlier reconstructions show side-fins running the length of its body, but these are now interpreted as artifacts formed as a corpse was squashed post-burial. A ring-like stain, interpreted as cartilage, encircles the very small ‘mouth’ (less than one centimetre in diameter) (Ritchie, 1968a, 1984), which together suggested that it was an ancestral parasitic lamprey (Ritchie 1960, 1968a, 1984; Mallatt, 1984). Jamoytius, however, apparently had no true teeth or teeth-like structures, in its ‘mouth’ (Sansom et al., 2010), If Jamoytius had rasping keratin teeth like living parasitic lampreys, as Stensiö (1958) inferred for Norwegian anapsids, then these should probably be preserved carbonized, as is much of the rest of the animal (Figure 1A). The controversy about whether this ‘mouth’ was anterior terminal, or subterminal ventral, seems to be resolved in favour of the latter (Sansom et al., 2010). Towards the anterior end, many specimens preserve a pair of linear features composed of serially repeated, contiguous, sub-rectangular shapes, interpreted as branchial openings (Sansom et al., 2010).
The anterior of Jamoytius has room for a piston-like tongue comparable with living parasitic lampreys (Mallat, 2023). In living parasitic lampreys, this holds the biting and cutting plates used to parasitize fish, which are not present in Jamoytius. On the other hand, such plates would not be required to eat soft vegetation, which is a possibility considering the holes in associated Ceratiocaris (see Paleoecology section), and Jamoytius does not have the lamprey lips used for suction (Richardson et al., 2010).
Most specimens do not preserve the posterior portion of Jamoytius, and where they do, it is too faint to be seen clearly (Sansom et al., 2010). So, the inferred hypocercal tail is reconstructed only by analogy with other near-contemporary anaspids, like Birkenia and Lasanius (Blom, 2012; Reeves et al., 2023).
Mode of life of Jamoytius
Jamoytius has been compared with parasitic lampreys which attack fish (Figure 2A and Figure 3). But, only 18 of the 38 known species of lamprey, are carnivorous. The ancestral crown lamprey was probably a freshwater nonparasitic species, some of which evolved into parasites (Brownstein and Near, 2023). Living non-parasitic lampreys are smaller (less than 40cm long) than parasitic sea lampreys (35-120cm long), and all inhabit freshwater (Potter et al., 2015). The non-carnivorous lampreys do not eat at all, since they have a nonfunctional intestine, only live for four to six months on the energy stored when young; as a result, they typically have small mouths and poorly-developed teeth, useless for attaching to a host, and die after spawning. (Cochran, 2008). For example, Lethenteron appendix, the American brook lamprey, has small larvae (1-2cm long) that feed on algae and detritus for between three and seven years, before metamorphosing into sexually mature adults (15-25cm long) (Fuller and Neilson, 2015). The size and anatomy of Jamoytius is more compatible with non-carnivorous living lampreys, though the Devonian undoubted parasitic lamprey, Priscomyzon riniensis, is also very small (Figure 4C).
The comparable agnathans, have terminal anterior mouths which do not appear to be protrusible (Figure 5). Such mouths are often found among omnivorous mid-water feeders, which eating anything available, by grabbing bits of food as they move (Oleh, 2018).
2. Palaeoenvironments and paleoecology.
2.1. Sediments
The Jamoytius-bearing horizon is one of several eurypterid- and fish-bearing units in the Silurian (Llandoverian; 444-433 Ma) of the Lesmahagow inlier in the Midland Valley of Scotland (Ritchie, 1960,1968a; Clarkson and Harper, 2016)). It is exposed at Birk Knowes in a cliff next to the Logan Water (NS737346) (Figure 7A) near the top of the Patrick Burn Formation, an over 500-metre-thick section of alternating grey sandstones, siltstones, and mudstones (Jennings, 1961; Lovelock, 1998) (Figure 7B). The sediments of the Patrick Burn Formation change gradually upwards from deeper water interbedded mudstone/ turbidite sandstone facies into shallower water interbedded mudstone/cross-bedded and laminated sandstone facies. (Jennings, 1961; Lovelock, 1998) This is accompanied by changes in the taxonomy and ecology of the fossil biotas from marine to freshwater (Jennings, 1961; Ritchie,1963; Lovelock, 1998) (Figure 7B).
Throughout the Patrick Burn Formation, and in the Jamoytius bed itself, there is a complete absence of burrowing organisms and there are no tracks or trails on the bedding plane surfaces (Lovelock, 1998) The undisturbed nature of the sediments, together with abundant pyrite and organic matter, indicates anaerobic bottom conditions in very quiet water subject to periodic underflows (Ritchie, 1963); in keeping with the abundance of Ceratiocaris - most modern Phyllocarids prefer low energy conditions and are tolerant of low oxygen concentrations (Rolfe & Beckett, 1984). Geochemical evidence indicates a gradual salinity drop through the upper part of the Priesthill Group, which contains all the fish beds (Jennings, 1961).
The Jamoytius bed itself is about 10 metres thick and consists of alternating fine-grained thin sandstones, deposited by turbidity currents, and varved siltstones in which the varves consist of a lower siltstone sharply overlain by an organic-rich muddy layer: the latter enclose most of the fossils (Jennings, 1961; Ritchie, 1963) (Figure 8A, B). The bounding sandstones are low-angle tabular cross-bedded fine-grained mature muddy quartz- and felspathic-sandstones (Jennings, 1961), deposited by sand waves in shallow water by storms (Harms et al., 1975; Lovelock, 1998) (Figure 8C, D). The deeper water Jamoytius bed with its turbidite sandstones and laminated siltstone contrast markedly with the enclosing shallow water cross-bedded fine sandstones and testify to great fluctuations in water levels and climate at millennial scales typical of marginal marine and semi-arid lake basins such as those of Lake Cariaco in northern Venezuela and Lake Chad in the central Sahara (González et al., 2008; Armitage et al., 2015).
Both the Jamoytius bed, and higher fish-bearing beds at Lesmahagow, have alternations of fine silt-clay couplets (varves) which contain the biota, and olive-grey massive mudstones which are barren (Zolitschka, et al., 2015). In the Jamoytius horizon, itself, the alternations of laminated siltstones and graded quartz sandstones are almost identical to those of Cariaco basin in Venezuela (Athearn,1965; Hughen et al., 1996). Water circulation inside this partially isolated basin is restricted, which, combined with high annual primary productivity, causes it to be permanently anoxic at depth, where alternating light and dark coloured varved sediments correspond respectively to the winter-spring dry season and the summer-fall rainy season (Muller-Karger et al 2001).
The tectonic situation of the Midland basin Silurian inliers is also comparable with that of the Cariaco and related basins along the northern Venezuelan coast. Both are complex basins between ocean and continent, which evolved through time from marine to freshwater continental conditions due to shortening and inversion caused by major strike-slip faulting, occupying about he same ~ 100 Ma time period, Cretaceous to Recent for Venezuela, and Silurian through Devonian for the Midland Valley (Dewey and Strachan, 2003; James, 1990).
Figure 7.
A) Birk Knowes exposure of Jamoytius beds. B) Section of Priesthill Group with fossil associations, ranges and inferred salinity (modified from Lovelock, 1998).
Figure 7.
A) Birk Knowes exposure of Jamoytius beds. B) Section of Priesthill Group with fossil associations, ranges and inferred salinity (modified from Lovelock, 1998).
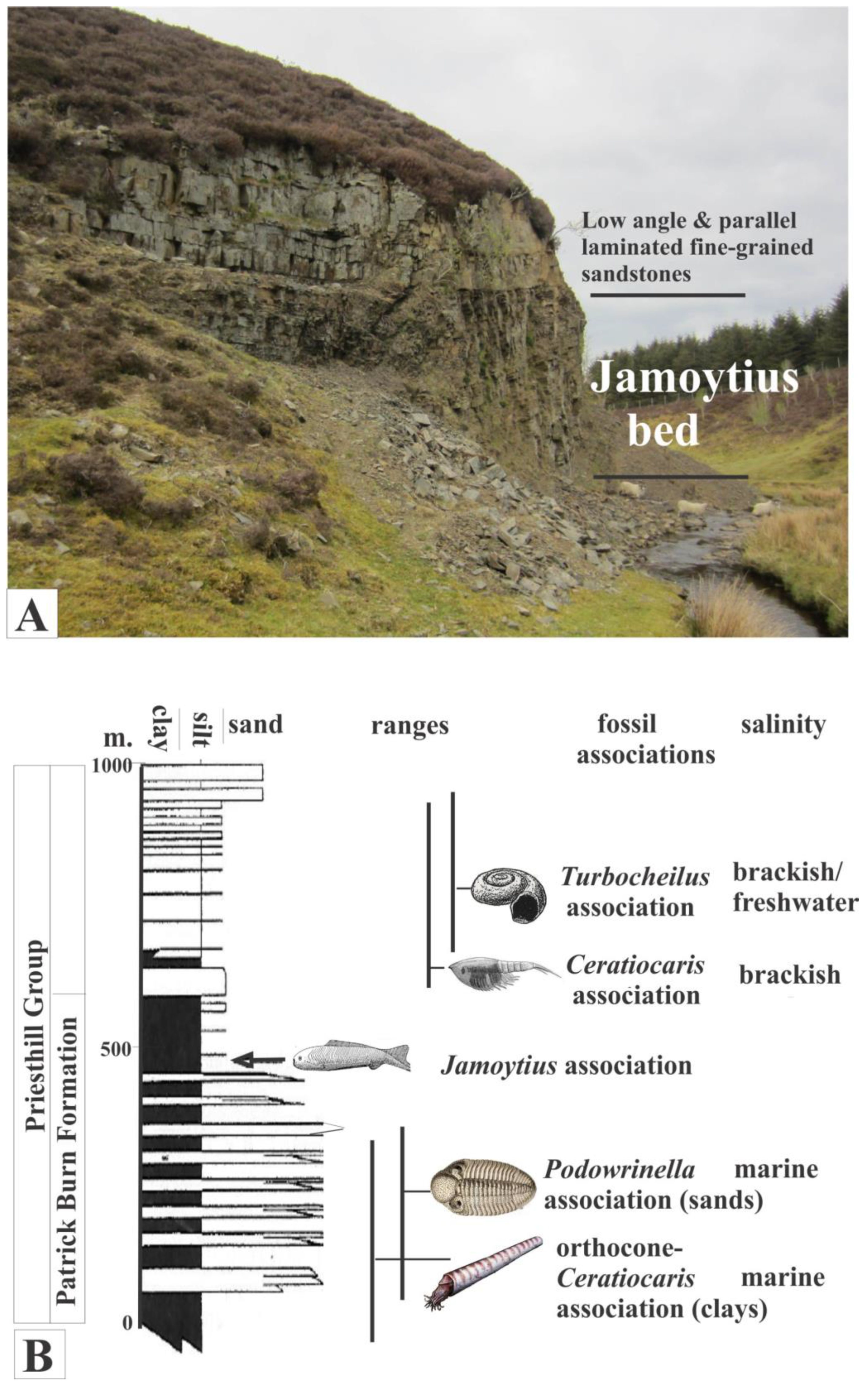
Figure 8.
Sediments, A) View of part of Jamoytius bed showing varved siltstones alternating with graded fine-grained sandstones; B) photomicrograph of Jamoytius bed varved siltstone showing lower silt overlain by organic-rich silt cap; C) view of outcrop of fine-grained quartz-sandstones overlying Jamoytius bed; D) Photomicrograph of fine-grained sandstone from C, dominated by subangular quartz grains with rare plagioclase felspar, in carbonate cement.
Figure 8.
Sediments, A) View of part of Jamoytius bed showing varved siltstones alternating with graded fine-grained sandstones; B) photomicrograph of Jamoytius bed varved siltstone showing lower silt overlain by organic-rich silt cap; C) view of outcrop of fine-grained quartz-sandstones overlying Jamoytius bed; D) Photomicrograph of fine-grained sandstone from C, dominated by subangular quartz grains with rare plagioclase felspar, in carbonate cement.

Palaeoecology
Though the lower Partick Burn Formation has transported marine, or brackish water, fossils in turbidite sandstone, which shows source connections with the sea, the lack of normal marine planktonic organisms above these basal beds is clear evidence that the oceanic connection was tenuous at best (Lovelock, 1998).
The Jamoytius bed lies above the Podowrinella (sands) and orthocone-Ceratiocaris (clays) biofacies, between shallow water unfossiliferous sandstones (Figure 6) (Lovelock, 1998). The Podowrinella biofacies is in turbidite sandstones and has been transported from shallower water. It has benthonic scavengers (4 trilobite species, 1 ostracod), attached filter feeders (3 brachiopods, 1 bivalve, crinoid ossicles, bryozoa), herbivores (1 gastropod), free living filter feeders (Tentaculites, Cornulitids). This fauna suggests living conditions in shallow turbulent marine, possibly slightly brackish, water (Lovelock, 1968). The orthocone-Ceratiocaris biofacies is in the clays and has only the podshrimp, Ceratiocaris papilio, rare orthocones and the occasional patch of thelodont scales. The orthocones are upright in the sediment and have floated in and settled with decomposition gas in chambers holding them upright as they settled though the water. The Ceratiocaris and thelodonts, in the absence of marine fossils in situ, indicate brackish to freshwater environments (Lovelock, 1998).
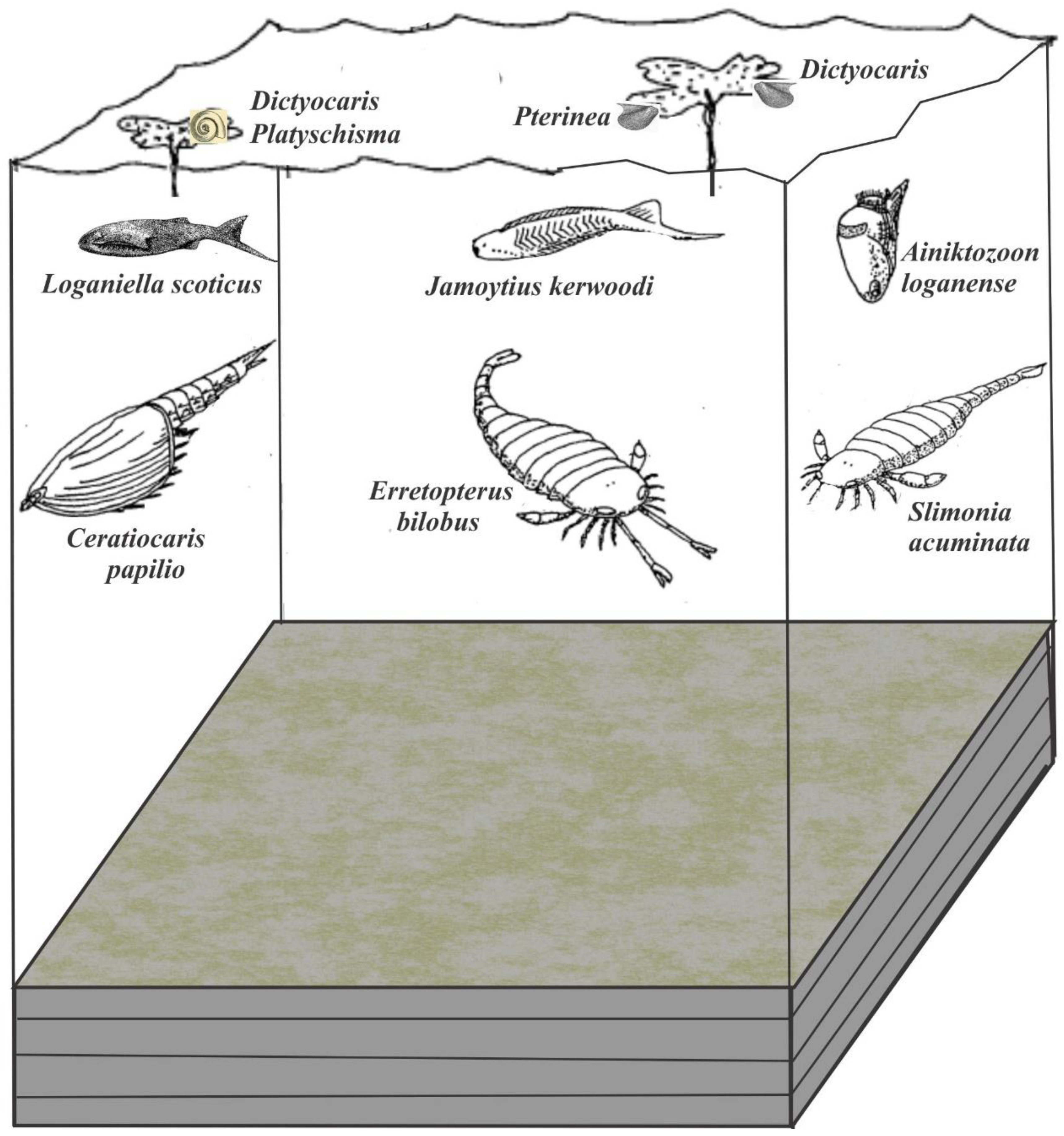
For the fossil biota of the Jamoytius bed, I use the list of Lovelock (1998), which list only those fossils from the actual laminated siltstones. Peach & Horne (1899) believed the Birk Knowes outcrop to be equivalent to those at Shank Castle, which was later shown to be incorrect (Jennings, 1961). Unfortunately, this mis-correlation has led to confusion over the attribution of some fossils to the Jamoytius bed (Lovelock, 1998, p.166-7), which error passed through successive editions of 'The Geology of Scotland' (Walton & Oliver, 1991). The single example of the blind “horseshoe crab”, Cyamocephalus loganensis Currie 1927, is a museum specimen, attributed to the Jamoytius bed only on similar lithology (Currie, 1927; Anderson, 1999) and though Hunter (1884) never recorded from where he got his single specimen of the scorpion Palaeophonus caledonicus (though this might be a plant – Ritchie, 1963), yet Peach and Horne (1899, p.574) attributed it to the Jamoytius horizon.
The actual fossil biota of the Jamoytius bearing laminated siltstone is dominated by the crustacean Ceratiocaris papilio, accompanied by the thelodont, Loganiella scotica, the enigmatic thylacocephalan crustacean? Anitkozoon loganenses (van der Brugghen et al., 1997), Dictyocaris slimoni (most likely a plant thallus, Ritchie, 1963), and disc- and stem-shaped plants. Other members are rare to very rare. Rare members are the eurypterids Slimonia acuminata, Jamoytius kerwoodi itself and the molluscs. Very rare members are the eurypterids, Erretopterus bilobus, Hughmilleria sp., the ostracod, Beyrichia sp. (one specimen), and the problematica, Taitia catena and Striatuncus scoticus (Ritchie, 1963) (Table 1).
Ceratiocaris, the pod shrimp, is up to 30 cm long, and is the most abundant fossil. Most shrimps are opportunistic omnivores that will eat plants, organic detritus, and any living or dead organism that does not eat them first (Albertoni et al., 2003; Walker, 2009).
Loganellia scoticus is up to 30 cm long, and was originally reconstructed as a bottom detritus feeder with heterocercal tail (Traquair, 1899): but it more likely lived as a nektonic feeder with hypocercal tail, as supposed for the anaspid Birkenia, especially considering the anoxic bottom over which it lived (Ritchie, 1963; Turner, 1982, 1999). Indirect evidence comes from fossil scroll coprolites assigned to the anaspids Birkenia and Loganiella, which occur in post-Llandoverian varved siltstones in northern Ireland, and are ascribed to detritus feeders (Gilmore, 1992).
Ainiktozoon, is about 12 cm long, and though originally described as an early chordate (Scourfield, 1937) is now more plausibly an arthropod, more precisely a thylacocephalan crustacean (van der Brugghen et al., 1997). Its mode of life is unknown, though its abundance suggests an herbivore or detritus feeder rather than a carnivore as postulated for other thylacocephalans (Haug et al., 2014).
Other arthropods, which are sometimes attributed to the Jamoytius bed, come from the overlying Kip Burn Formation, now mostly covered by the waters of the Logan reservoir, including the millipede Archidesmus loganensis Peach, 1899 (Peach and Horne, 1899, p. 573).
Dictyocaris slimoni, forms large carbonaceous sheets up to 30 cm in diameter, commonly pierced by variably sized circular holes up to 5 mm across, with raised rims (Störmer, 1935; van der Brugghen, 1995) (Figure 9). The holes are the same size as the mouth of Jamoytius, and thus could be parasitic injuries (Ritchie, 1963). But the abundant, often articulated, Ceratiocaris and eurypterid exoskeletons also associated with Jamoytius show no such holes; and Dictyocaris is never found even partially articulated, despite its association with these articulated arthropods. The large number of holes on the illustrated specimen also seems too many to be the results of parasitism, considering the size of Jamoytius (14-18cm). Dictyocaris thus is likely a plant (Ritchie, 1963).
The morphologies of the rare eurypterids in the Jamoytius bed are more easily interpreted as those of nektonic scavengers, as they have none of the specialized adaptations for catching prey found in higher fish beds at Lesmahagow, such as the large spiny grasping arms of the mixoperid Lanarkopterus dolichoschelus (Ritchie, 1968b; Schmidt et al., 2022) (Figure 10A). For example, Erettopterus with its small pincers and compound eyes was probably a predator/scavenger with high visual acuity, but it was not as highly specialized or active as other eurypterids (Plotnik, 1999). Similarly, Ainiktozoon had likely neither the speed, nor the appendages, to catch a fast-moving Loganiella (Figure 10B)
Of the rare molluscs and the small orthocone cephalopod recorded by Ritchie (1963) in the Jamoytius horizon, the bivalve Pterinea is a byssally attaches suspension feeder. Small clusters of Pterinea occur with carbonaceous patches which may be floating plants to which they attached (Ritchie, 1963, p. 149); such Pteriniids often attach to floating vegetation (Stanley, 1972), The low-spired gastropod Platyschisma is most likely a grazing or scavenging form (Linsley, 1977). The small orthocones are not part of the Jamoytius bed biota, and were transported in from another shallower water environment, as they occur in the turbidite sandstones interbedded with the laminated siltstones that have the vertebrates and the eurypterids (personal observations in 2022).
The Jamoytius association is thus dominated by supposed omnivores and herbivore/ detritus feeders, with primary production represented by phytoplankton and land plant spores and? algal thalluses (?Dictyocaris) that contribute the dark laminae within the siltstone beds (Figure 8B). There is no evidence for large scale transportation of the fossils after death: they simply settled into the anoxic bottom.
Figure 9.
Dictyocaris: A) thallus with holes; B) drawing showing holes of varying sizes and raised rims (both from van der Brugghen, 1995).
Figure 9.
Dictyocaris: A) thallus with holes; B) drawing showing holes of varying sizes and raised rims (both from van der Brugghen, 1995).
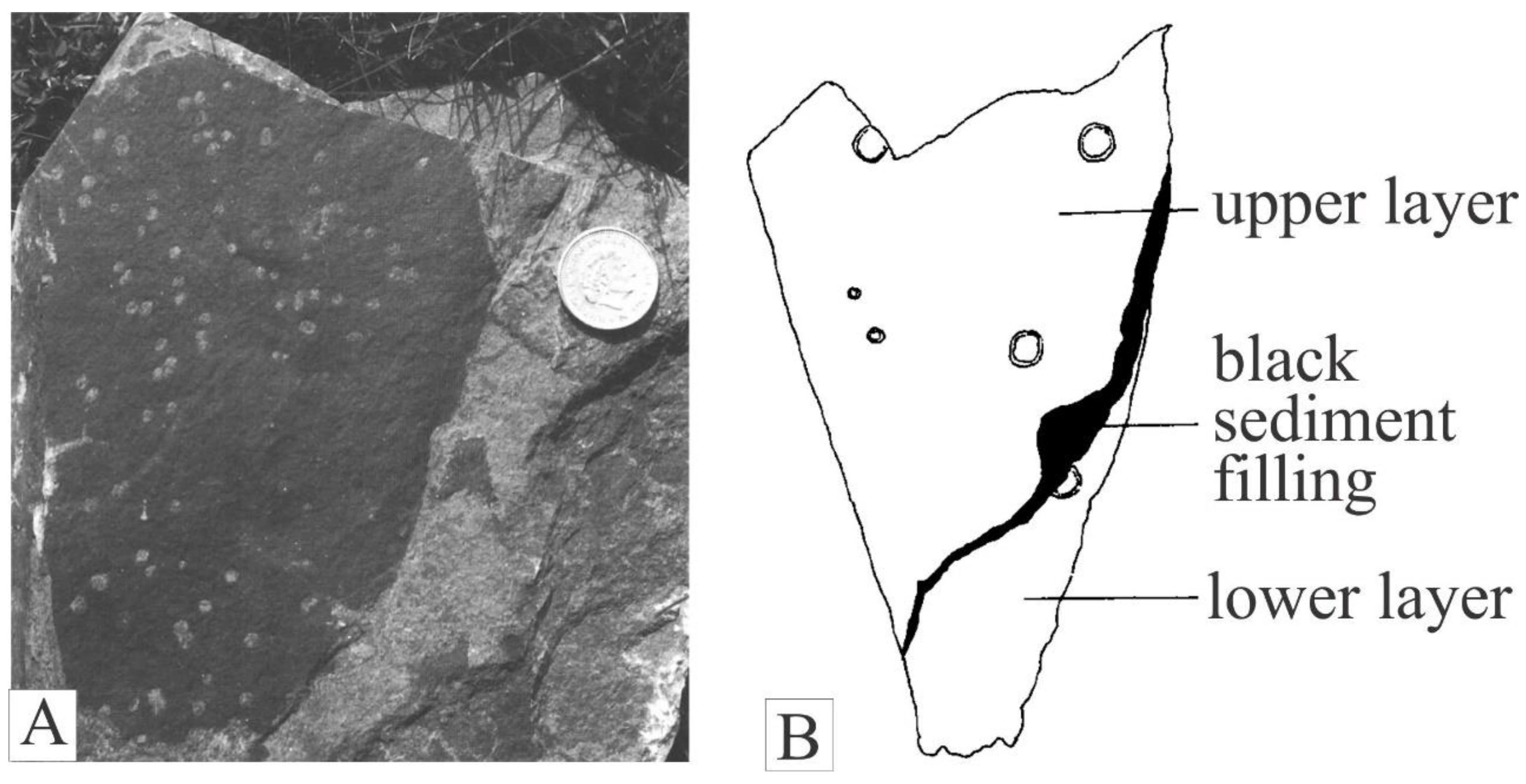
Figure 10.
A) Reconstruction of Lanarkopterus dolichelus Ritchie 1968 (after Ritchie, 1968b); B) Ainitkozoon chasing a Loganiella (with permission from Nobu Tamura).
Figure 10.
A) Reconstruction of Lanarkopterus dolichelus Ritchie 1968 (after Ritchie, 1968b); B) Ainitkozoon chasing a Loganiella (with permission from Nobu Tamura).
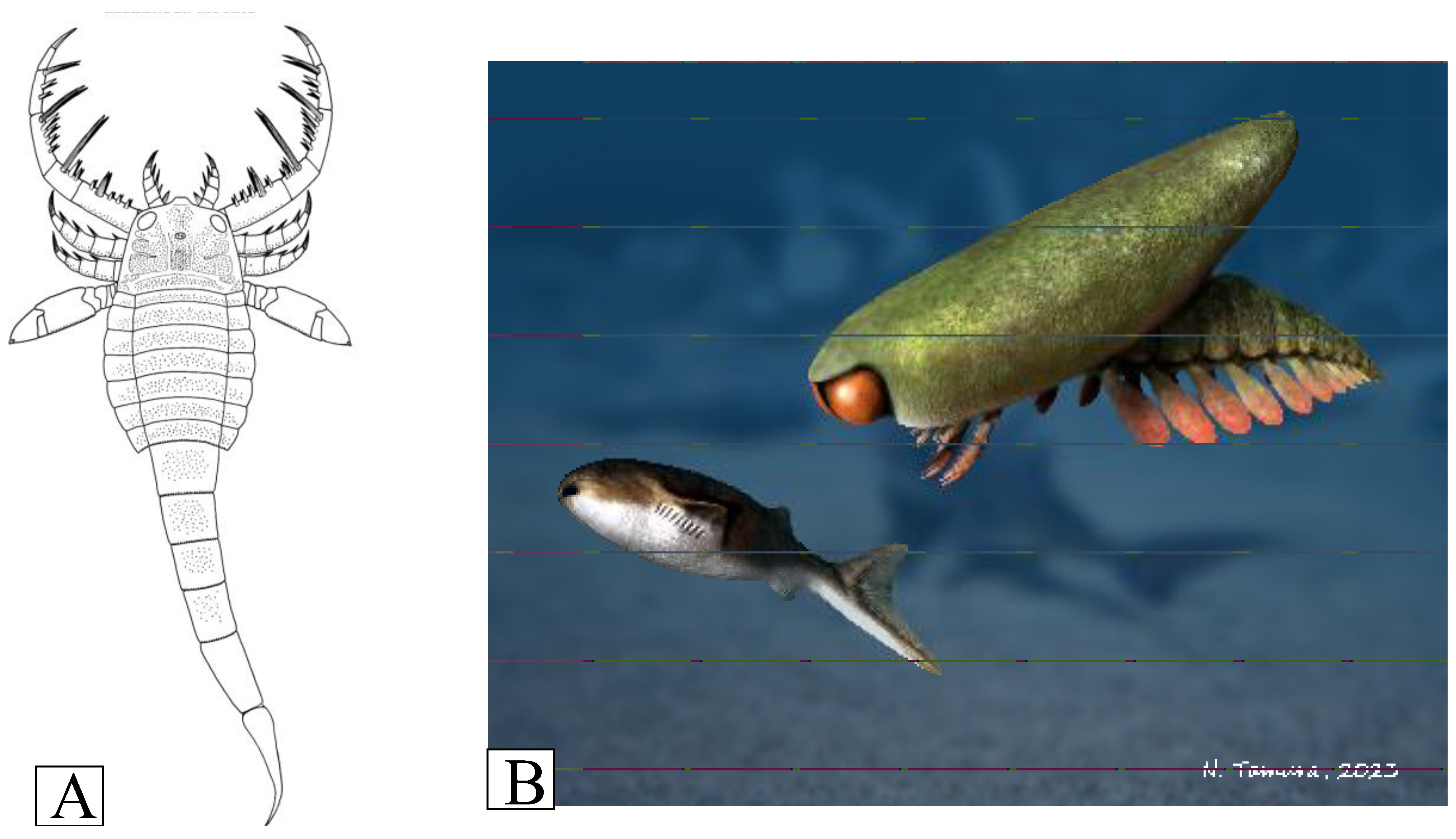
Figure 11.
Palaeoecological sketch of the Jamoytius association living over an anoxic bottom (modified from Lovelock, 1998, fig. 4.3).
Figure 11.
Palaeoecological sketch of the Jamoytius association living over an anoxic bottom (modified from Lovelock, 1998, fig. 4.3).
3. Discussion
The Jamoytius reconstructions with a terminal mouth suggest a filter-feeder or a detritus-feeder, analogous to larval lampreys (Denison, 1961) (Figure 1 C, E) and possibly to the Loganelliform thelodonts with which it is associated: the latter are interpreted as pelagic slow swimmers in open water (Ferrón and Botella, 2017). Larval lampreys can feed on highly concentrated food suspensions so thick that they border on organic deposits (Mallat, 1984, 1985). Jamoytius, however, lacks any obvious adaptations to suspension feeding (Lammons, 2009), and the more likely anterior ventral position of the mouth indicates particulate feeding or grazing (Oleh, 2018).
A bottom detritus feeding life style proposed by Parrington (1958) is unlikely given the anoxic bottom inferred from sedimentology, and the hypocercal tail which would give lift to a fish whose morphology also suggests an active lifestyle (Kermack, 1943). Jamoytius, in fact, like other euphaneropids, resembles elongate arrow-like bony fish, like pike (Esox spp.) and barracuda (Sphyraena spp.), with posterior dorsal and anal fins, which assist the tail in bursts of rapid acceleration, but are inefficient at steady swimming (Fletcher et al., 2014). Ritchie (1968) considered that its highly developed metamerism (a linear series of body segments fundamentally similar in structure) and large eyes of Jamoytius indicated a very active, fast-swimming vertebrate.
Sedimentological and palaeoecological characteristics of the associated biota indicate that Jamoytius lived in a brackish water environment in which the bottom waters and sediments were anoxic, and inhospitable to benthos and a predominantly planktonic and nektonic biota lived only in the overlying oxic waters. A benthonic mode of life for any of the Jamoytius association organisms is unlikely. Jamoytius and its likely euphaneropid sisters, despite their possibly autapomorphic, elongated branchial basket, could be plausible stem lampreys (Janvier, 2008). The inferred herbivorous life style of Jamoytius may not be too far apart from that of living secondarily non-parasitic lampreys. Petromyzids may initially have evolved as microphagous filterers or herbivorous organisms and only later developed ectoparasitic modes of life (Strahan, 1963; Mallatt, 1984, 2023).
4. Conclusions
Jamoytius is associated with a low-diversity dominantly nektonic detritus and herbivorous fauna living over an anoxic bottom, at the transition from a marine-influenced, probably brackish-water, deep-water basin to a shallower-water, less saline and likely freshwater basin. Jamoytius was likely either a free-living surface feeding detritivore/herbivore. The paleoenvironment in which it lived compares well with that of living non-parasitic lampreys. Though its immediate descendants seem to be parasitic, this ectoparasitic mode of life may have evolved from ancestral microphagous filterers or herbivores.
5. Acknowledgements
This study is part of an ongoing study on the palaeoenvironments and ages of the Silurian inliers of the Midland Valley of Scotland. I thank my colleagues, Elizabeth Catlos and Hector Garza for help during joint field work, and Susan Turner for a detailed review. This paper is dedicated to Alex Ritchie, who sadly died on 16 November, 2023 at the age of 88.
References
- Albertoni, E.F., C. Palma-Silva & F.A. Esteves. 2003. Overlap of dietary niche and electivity of three shrimp species (Crustacea, Decapoda) in a tropical coastal lagoon (Rio de Janeiro, Brazil). Revista Brasileira de Zoologia 20, 135-140. [CrossRef]
- Anderson, L.I., 1999. A new specimen of the Silurian synziphosurine arthropod Cyamocephalus. Proceedings of the Geologist’s Association 110 (3), 211-216. [CrossRef]
- Armitage, S.J., Bristow, C./S., Drake, N.A., 2015. West African monsoon dynamics inferred from abrupt fluctuations of Lake Mega-Chad. Proceedings of the National Academy of Science 112(28), 8543-8548. [CrossRef]
- Athearn, W.D., 1965. Sediment Cores from the Cariaco Trench, Venezuela. Woods Hole Oceanographic Institution unpublished Technical Report 65-37, 20p.
- Blom, 2012. New Birkeniid anaspid from the Lower Devonian of Scotland and its phylogenetic implications. Palaeontology 55 (30, 641–652. [CrossRef]
- Blom, H., Märss, T., 2010. The interrelationships and evolutionary history of anaspid fishes. In: D.K. Elliott, J.G. Maisey, X. Yu, and D.Miao (eds.), Morphology, Phylogeny, and Paleobiogeography of Fossil Fishes, 45–58. Verlag Dr. F. Pfeil, Munich.
- Brownstein, C.D., Near, T.J., 2023. Phylogenetics and the Cenozoic Radiation of lampreys. Current Biology 33 (2), 397-404. [CrossRef]
- Chevrinais, M., Johanson, Z., Trinajstic, K., Long, J., Morel, C., Renaud, C., Cloutier, R., 2018. Evolution of vertebrate postcranial complexity: Axial skeleton regionalization and paired appendages in a Devonian jawless fish. Palaeontology. 61, 1-13. [CrossRef]
- Clarkson, E.N.K., Harper, D.T., 2016. Silurian of the Midland Valley of Scotland and Ireland. Geology Today 32(5), 195-200. [CrossRef]
- Cochran, P. A., 2008. Observations on giant American Brook Lamprey (Lampetra appendix). Journal of Freshwater Ecology 23, 161–164. [CrossRef]
- Denison, R. N., 1961. Feeding mechanisms of agnatha and early gnathostomes. American Zoologist 1, 177–182. [CrossRef]
- Dewey, J.F., Strachan R.A., 2003. Changing Silurian–Devonian relative plate motion in the Caledonides: sinistral transpression to sinistral transtension. Journal of the Geological Society, London 160, 219–229. [CrossRef]
- Dineley, D., 1999. Silurian fossils fish sites of Scotland. In: Dineley, D. & Metcalf, S (eds). Fossil Fishes of Great Britain. Chapter 2. Geological Conservation Review Series 16, 33-62. [CrossRef]
- Donoghue, P. C. J., Keating, J.N., 2014. Early vertebrate evolution. Palaeontology 57 (5), 879-893. [CrossRef]
- Donoghue, P. C. J., Purnell, M. A., 2009. Distinguishing heat from light in debate over controversial fossils. BioEssays 31(2), 178–189. [CrossRef]
- Donoghue, P. C. J., Smith, M. P., 2001. The anatomy of Turinia pagei (Powrie), and the phylogenetic status of the Thelodonti. Transactions of the Royal Society of Edinburgh: Earth Sciences 92, 15–37.
- Donoghue, P. C. J., Forey, P. L, Aldridge, R. J., 2000. Conodont affinity and chordate phylogeny. Biological Reviews 75, 191–251. [CrossRef]
- Donoghue, P.C.J., Smith, M. P., Sansom, I. J., 2003. The origin and early evolution of chordates: molecular clocks and the fossil record, in Donoghue, P. C. J., Smith, M. P. (Eds), Telling the evolutionary time: molecular clocks and the fossil record. CRC Press, London, pp.190–223.
- Ferrón, H.G., Botella, H., 2017. Squamation and ecology of thelodonts. PLoS One12(2), e0172781. [CrossRef]
- Fletcher, T., Altringham, J., Peakall, J., Wignall, P., Dorrell, R., 2014. Hydrodynamics of fossil fishes. Proceedings of the Royal Society B281,20140703. [CrossRef]
- Forey, P. L., 1984. Yet more reflections on agnatha-gnathostome relationships. Journal of Vertebrate Paleontology 4, 330–343. [CrossRef]
- Forey, P. L., 1995. Agnathans recent and fossil, and the origin of jawed vertebrates. Reviews in Fish Biology and Fisheries 5, 267–303. [CrossRef]
- Forey, P. L., Janvier, P. 1993. Agnathans and the origin of jawed vertebrates. Nature 361, 129–134. [CrossRef]
- Fuller, P., Neilson, M., 2015. Lethenteron appendix. USGS Nonindigenous Aquatic Species Database, Gainesville, FL. http://nas.er.usgs.gov/queries/FactSheet.aspx?speciesID=835.
- Gess, R. W., Coates, M. I., Rubidge, B. S., 2006. A lamprey from the Devonian period of South Africa. Nature 443, 981–984. [CrossRef]
- Gilmore, B., 1992 Scroll coprolites from the Silurian of Ireland and the feeding of early invertebrates. Palaeontology 35 (2), 319-333.
- González, C., Dupont, L.M., Behling, H., Wefer, G., 2008. Neotropical vegetation response to rapid climate changes during the last glacial period: Palynological evidence from the Cariaco Basin. Quaternary Research 69 (2), 217-230. [CrossRef]
- Harms, J.C., Southard, J.B., Spearing, D.R., Walker, R.G., 1975. Depositional Environments as Interpreted from Primary Sedimentary Structures and Stratification Sequences. Society for Economic Petrology and Mineralogy, Tulsa, Oklahoma. 161p. [CrossRef]
- Haug, C., Briggs, D.E.G., Mikulic, D.G., Kluessendorf, J., Huag, J.T., 2014. The implications of a Silurian and other thylacocephalan crustaceans for the functional morphology and systematic affinities of the group. BMC Evol Biol 14, 159. [CrossRef]
- Hughen, K.A., Overpeck, J.T., Peterson, L.C., Anderson, R.F., 1996a. The nature of varved sedimentation in the Cariaco Basin, Venezuela, and its paleoclimatic significance. Geological Society Special Publication 116, 171–183. [CrossRef]
- Hunter, J.R.S., 1884. Notes on a new fossil scorpion (Palaeophonus caledonicus) from the Upper Silurian Shales, Logan Water, Lesmahagow. Transactions of the Edinburgh Geological Society 5, 187 – 191. [CrossRef]
- James, K., 1990. The Venezuelan Hydrocarbon Habitat. Geological Society of London Special Publication 50. 9-35. [CrossRef]
- Janvier, P., 1981. The phylogeny of the Craniata, with reference to the significance of fossil ‘agnathans’. Journal of Vertebrate Paleontology 1, 121–159. [CrossRef]
- Janvier, P. 1996. Early Vertebrates. Oxford Monographs on Geology and Geophysics 33. Clarendon Press, Oxford. 393pp. [CrossRef]
- Janvier, P., 2008. Early Jawless Vertebrates and Cyclostome Origins. Zoological Science 25(10), 1045-1056. [CrossRef]
- Janvier, P., Arsenault, M., 2007. The anatomy of Euphanerops longaevus Woodward, 1900, an anaspid-like jawless vertebrate from the Upper Devonian of Miguasha, Quebec, Canada. Godiversitas 29, 143–216.
- Jennings, J.S. 1961. The geology of the eastern part of the Lesmahagow inlier. Univ. Edinburgh. Unpub. Ph.D. thesis, 306p.
- Keating, J.N., Donaghue, P.C.J., 2016. Histology and affinity of anaspids, and the early evolution of the vertebrate dermal skeleton. Proceedimngs of the royal society B283, 20152917. [CrossRef]
- Kermack, K.A. Kermack, K.A., 1943. The functional dignificance of the hypocercal Tail in Pteraspis rostrata. Journal of Experimental Biology 20 (1), 23-27. [CrossRef]
- Kuraku, S., Kuratani, S., 2006. Time Scale for Cyclostome Evolution Inferred with a Phylogenetic Diagnosis of Hagfish and Lamprey cDNA Sequences. Zoological Science 23 (12), 1053-1064. [CrossRef]
- Lammons, M.L., 2009. Mud and Mucus: Feeding selectivity in a suspension-feeding detritivorous fish. M.Sc., The College of William and Mary. 101p.
- Linsley, R. M., 1977. Some "laws" of gastropod shell form. Paleobiology 3, 196-206. [CrossRef]
- Lovelock, C.E., 1998. Sedimentary environments and biofacies of the Silurian inlier at Lesmahagow Midland Valley of Scotland. Unpub PhD thesis, University of Edinburgh, 448p.
- Mallat, J., 1984. Feeding ecology of the earliest vertebrates. Zoological Journal of the Linnean Society 82, 261–272. [CrossRef]
- Mallatt, J. (1985). Reconstructing the Life Cycle and the Feeding of Ancestral Vertebrates. In: Foreman, R.E., Gorbman, A., Dodd, J.M., Olsson, R. (eds) Evolutionary Biology of Primitive Fishes. NATO ASI Series, vol 103, 59-68. Springer, Boston, MA. [CrossRef]
- Mallat, J., 2023. Vertebrate origins are informed by larval lampreys (ammocoetes): a response to Miyashita et al., 2021. Zoological Journal of the Linnean Society 197, 287–321. [CrossRef]
- Miyashita, T., Coates, M., Farrar, R., Larson, P., Manning, P., Wogelius, R., Edwards, N., Anné, J., Bergmann, U., Palmer, A., Currie, P., 2019. Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological-molecular conflict in early vertebrate phylogeny. Proceedings of the National Academy of Sciences. 116, e2146-2151. [CrossRef]
- Miyashita, T., Gess, R.W., Tietjen, K., Coates. M.I., 2021 Non-ammocoete larvae of Palaeozoic stem lampreys. Nature 591, 408–412. [CrossRef]
- Muller-Karger, F. E., Varela, R., Thunell, R., Scranton, M., Bohrer, R., Taylor, G., Capelo, J., Astor, Y., Tappa, E., Ho, T.Y., Walsh, J.J., 2001. Annual cycle of primary production in the Cariaco Basin: response to upwelling and implications for vertical export. Journal of Geophysical Research 106C3, 4527-4542. [CrossRef]
- Oleh, M., 2018. Laboratory Manual on General and Special Ichthyology. World News of Natural Sciences 18 (1), 1-51.
- Parrington, F. R. 1958. On the nature of the Anaspida. In T. S. Westoll, (ed.), Studies on fossil vertebrates. Athlone Press, London. p. 108-128.
- Peach, B.N., Horne, J. 1899. The Silurian rocks of Britain, Vol.1 Scotland. (with petrological chapters and notes by J.J.H. Teall). Memoirs of the Geological Survey of the United Kingdom. Glasgow, HMSO. 749 pp.
- Plotnick, R.E., 1999. Habitat of Llandoverian-Lochkovian eurypterids. In: A. J. Boucot, J. D. Lawson (eds.), Paleocommunities - a case study from the Silurian and Lower Devonian. p. 106-136.
- Potter, I. C., Gill, H. S., Renaud, C. B., Haoucher, D., 2015, The Taxonomy, Phylogeny, and Distribution of Lampreys. In: Docker, M. F. (ed.), Lampreys: Biology, Conservation and Control, Springer Netherlands, pp. 35–73. [CrossRef]
- Reeves, J.C., Sansom, R.S., 2023. Multivariate mapping of ontogeny, taphonomy and phylogeny to reconstruct problematic fossil taxa. Proceedings of the Royal Society B.290, 20230333. [CrossRef]
- Reeves, J.C., Wogelius, R., Keating, J., Sansom, R. S., 2023. Lasanius , an exceptionally preserved Silurian jawless fish from Scotland. Palaeontology 66, e12643. [CrossRef]
- Richardson, M.K., Admiraal, J., Wright, G.M., 2010. Developmental anatomy of lampreys. Biological Reviews 85 (1), 1-206. [CrossRef]
- Ritchie, A., 1960. A new interpretation of Jamoytius kerwoodi White. Nature 188, 647–649. [CrossRef]
- Ritchie, A., 1963. Palaeontological studies on Scottish Silurian fish beds. Upublished Ph.D. thesis, University of Edinburgh.170pp.
- Ritchie, A., 1968a. New evidence on Jamoytius kerwoodi White, an important ostracoderm from the Silurian of Lanarkshire, Scotland. Palaeontology 11, 21–39.
- Ritchie, A., 1968b. Lanarkopterus dolichoschelus (Störmer) gen. nov., a mixopterid eurypterid from the Upper Silurian of the Lesmahagow and Hagshaw Hills inliers, Scotland. Scottish Journal of Geology 4, 317 – 338. [CrossRef]
- Ritchie, A., 1984. Conflicting interpretations of the Silurian agnathan, Jamoytius. Scottish Journal of Geology 20, 249–256. [CrossRef]
- Rolfe, W.D.I., Beckett, E.C.M., 1984. Autecology of Silurian Xiphosurida, Scorpionida, Cirripedia, and Phyllocarida. Special Papers in Paleontology 32: 27–37.
- Sansom, R.S., Freedman, K., Gabbott, S.E., Aldridge, R.J., Purnell, M.A., 2010. Taphonomy and affinity of an enigmatic Silurian vertebrate, Jamoytius kerwoodi White. Palaeontology 53, 1393-1409. [CrossRef]
- Schmidt, M., Melzer, R.R., Plotnick, R.E., Bicknell, R.D.C., 2022. Spines and baskets in apex predatory sea scorpions uncover unique feeding strategies using 3D-kinematics. iScience 25 (1), 103662. [CrossRef]
- Schweitzer, M.H., Zheng, W., Moyer, A.E., Sjövall, P., Lindgren, J., 2018. Preservation potential of keratin in deep time. PloS one, 13:e0206569. [CrossRef]
- Scourfield, D.J., 1937. An anomalous fossil organism, possibly a new type of chordate, from the Upper Silurian of Lesmahagow, Lanarkshire – Ainiktozoon loganense, gen. et sp. nov. Proceedings of the Royal Society B. 121 (825): 533–547. [CrossRef]
- Shu, D., Luo, H-L., Conway-Morris, S., Zhang, X., Hu, S., Han, J., Zhu, M., Li, Y., L-Z., 1999. A Lower Cambrian vertebrate from South China. Nature. 402. 42-46. [CrossRef]
- Stanley, S.M., 1972. Functional morphology and evolution of byssally attached bivalves. Journal of Paleontology 46 (2), 165-212.
- Stensiö, E.A., 1958. Les cyclostomes fossiles ou ostracodermes. In: Grasse, P.-P. (editor) Traite de zoologie 13. Paris: Masson. p.173-125.
- Störmer, L., 1935. Dictyocaris salter, a large crustacean from the Upper Silunan and. Downtownian. Norsk Geologisk Tidsskrift 15, 267-298.
- Strahan, A., 1963. The behaviour of Mixinoids. Acta zoologica Stockholm 44, 73–102.
- Tarlo, L. B. H. 1967. Agnatha.. In W. B. Harland (ed.). The fossil record. London Geological Society, London, 629–636.
- Traquair R. H., 1899. Report on fossil fishes collected by the Geological Survey of Scotland in the Silurian rocks of the south of Scotland. Transactions of the Royal society of Edinburgh 39, 827-864. [CrossRef]
- Turner, S., 1982. A new articulated thelodont (Agnatha) from the Early Devonian of Britain. Palaeontology 25, 879-889.
- Turner, S., 1999. Early Silurian to Early Devonian thelodont assemblages and their possible ecological significance. In Boucot, A.J. and Lawson, J. (eds.). Palaeocommunities: a case study from the Silurian and Lower Devonian -International Geological Correlation Programme 53, Project Ecostratigraphy, Final Report. Cambridge University Press, Cambridge. pp 42-78.
- Turner, S., 2004. Early vertebrates: analysis from microfossil evidence. Recent Advances in the Origin and Early Radiation of Vertebrates G. Arratia, M. V. H. Wilson & R. Cloutier (eds.): pp. 67-94, 7 figs., 2 tabs. Verlag Dr. Friedrich Pfeil, München, Germany.
- van der Brugghen, G.,1995. Dictyocaris, een enigmatisch fossiel uit het Silur. Grondboor en Harner 1, 18-22.
- van der Brugghen, G., 2015. Ciderius cooperi gen. nov., sp. nov., the earliest known euphaneropid from the Lower Silurian of Scotland. Netherlands Journal of Geosciences 94, 1-10. [CrossRef]
- van der Brugghen, G., Schram, F.R., Marthill, D.M., 1997. The Fossil Ainiktozoon is an arthropod. Nature 385, 589–91.
- Walker, I., 2009. Omnivory and resource-sharing in nutrient-deficient Rio Negro waters: Stabilization of biodiversity? Acta Amazonica 39, 617-626. [CrossRef]
- Walton E. K.& Oliver, G. J. H. 1991. — Lower Palaeozoic stratigraphy, in Craig G. Y. (ed.), Geology of Scotland. 3rd ed. Scottish Academic Press, Edinburgh: 161-193.
- White, E. I., 1946. Jamoytius kerwoodi, a new chordate from the Silurian of Lanarkshire. Geological Magazine 83, 89– 97. [CrossRef]
- Wu, F., Janvier, P., Zhang, C., 2023. The rise of predation in Jurassic lampreys. Nature Communications 14 (1), 6652. [CrossRef]
- Zolitschka, B., Francus, P., Antti, E.K.O., Schimmelmann, A., 2015. Varves in lake sediments a review. Quaternary Science Reviews 117, 1-41. [CrossRef]

Table 1.
Taxa, ecology and abundance of Jamoytius association biota.
| Taxa | Feeding strategy | Frequency |
| Arthropods | ||
| Ceratiocaris papilio | nektonic omnivore | very common |
| Slimonia acuminata | nektonic scavenger | rare |
| Erretopterus bilobus | nektonic carnivore | very rare |
| Ainiktozoon loganense | unknown | common |
| Beyrichias sp. (1 specimen) detritivore | very rare | |
| Chordata | ||
| Loganellia scotica | nektonic detritus/herbivore? | common |
| Jamoytius kerwoodi | nektonic detritus/herbivore? | rare |
| Loganellia grossi | nektonic detritus/herbivore? | very rare |
| Cephalopoda | ||
| ?Orthocone indeterminate | nektonic carnivore | very rare |
| Small (2 specimens) | ||
| Gastropoda | ||
| Platyschisma helicites | mobile herbivore, grazer | rare |
| (7 specimens) | ||
| Bivalvia | ||
| Pteritonella sp. | Bysally attached suspension | rare |
| feeder | ||
| Unknown | ||
| Dictyocaris slimoni | plant primary producer? | very common |
| Plant | ||
| Tatia catena | primary producer | very rare |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



