Submitted:
24 December 2023
Posted:
26 December 2023
You are already at the latest version
Abstract
Non-coding RNAs, specifically microRNAs (miRNAs), exhibit altered expression and contribute significantly to the pathological processes observed in numerous diseases, encompassing various types of human cancers. Recent discoveries regarding miRNAs underscore their crucial roles in tumor pathogenesis and their responses to diverse therapeutic interventions. miRNAs are a subset of non-coding RNAs that regulate the expression of a multitude of genes post-transcriptionally and thus are potential diagnostic, prognostic, and predictive biomarkers and have also emerged as potential therapeutics. Because miRNAs are involved in the post-transcriptional regulation of their target mRNAs via repressing gene expression, defects in the miRNA biogenesis pathway and miRNA expression perturb the expression of a multitude of oncogenic or tumor-suppressive target genes that are involved in the pathogenesis of various cancers. As such, numerous miRNAs have been identified to be downregulated or upregulated in many types of cancers functioning as either oncomiRs or oncosuppressor miRs. Moreover, dysregulation of miRNA biogenesis pathways can also change miRNA expression and function in cancer.
Profiling deregulated miRNAs in many cancer types has been shown to correlate with disease diagnosis, indicate optimal treatment options, and predict response to a specific therapy. Specific miRNA signatures can track all stages of disease including many cancer types and hold potential as biomarkers and therapeutic targets as well as therapeutics as miRNA mimics and inhibitors (antagomirs). As such, identifying specific miRNAs and mRNAs they regulate in many types of cancer along with downstream pathways can be used as potential therapeutic targets.
Because a single miRNA can regulate a pool of targets involved in similar cellular processes and pathways, thereby amplifying the cellular response, this potentially makes miRNAs powerful therapeutics to restore cell functions that are altered as part of a disease phenotype. However, because a single miRNA regulates multiple gene expressions their cellular effects are numerous leading to potential off-target effects. Besides their intended targeting issues, there is a need to reduce the immunogenic reactions and determine the minimal dosing to achieve the desired effect while minimizing side effects. As such, a careful risk evaluation of miRNA therapeutics is needed to minimize off-target effects and to avoid overdosing of miRNA drugs. Furthermore, a limited understanding and validation of the specific roles of miRNAs limits their clinical application. In addition, there are many challenges concerning the delivery, sensitivity, specificity, toxicity, and applicability of the potential utility of miRNAs as therapeutic targets or therapeutics. Thus, future work will warrant if miRNAs can be used as cancer biomarkers as well as therapeutics for clinical application.
Keywords:
microRNAs
; miRNAs
; post-transcriptional gene regulation
; miRNA therapeutics
; miRNA mimics
; antagomirs
; antimirs
1. Introduction
Although the majority of oncology research is still focused on the dynamic variations of proteins and protein coding RNAs (their corresponding coding sequences only account for ∼2% of the genome), the role of non-coding RNAs (ncRNAs) transcribed from the remaining 98% of the genome including microRNAs (miRNAs) plays key roles in a multitude of biological processes in normal physiological states but also the development of various types of diseases including cancer underscoring the importance of miRNAs and other ncRNAs in tumor initiation and progression. While most of the research in oncology predominantly centers around the ever-changing aspects of proteins and the RNA molecules responsible for coding those proteins, it’s important to note that these coding sequences account for only about 2% of the genome (https://www.genomicseducation.hee.nhs.uk/genotes/knowledge-hub/non-coding-dna/) [1,2,3]. However, the remaining 98% of the genome, which includes non-coding RNAs (ncRNAs) such as miRNAs, plays pivotal roles in numerous biological processes during typical physiological conditions, as well as in the onset and advancement of different diseases, including cancer [4]. This emphasizes the significance of miRNAs and other non-coding RNAs in the initiation and progression of tumors.
In addition, miRNAs play key roles in the regulation of gene expression at the transcriptional [5,6,7] and post-transcriptional [8,9,10,11] levels, and exhibit tissue-specific [12,13] and developmental expression patterns [14,15,16] and play key functional roles in a broad range of biological processes within cells and organisms. Altered expression of miRNAs has emerged as an additional molecular mechanism implicated in the pathogenesis of numerous diseases [17,18,19], spanning innate immunity [20], autoimmunity and autoimmune diseases [21], viral infections [22,23,24,25], acute hepatitis [26], depression [27], anxiety [28], Alzheimer’s disease [29], Huntington’s disease [30], metabolic and cardiovascular diseases [31,32,33,34], diabetes [8,33,34,35,36,37,38] and a many types of cancers [12,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69]. Consequently, these miRNAs can serve as indicators for the presence of a pathological condition, as well as provide insights into its stage, progression, or genetic associations.
More recently, there is an emerging evidence suggesting that diet-derived exogenous miRNAs (or “xenomiRs”) can enter the circulatory system and tissues, potentially influencing gene expression and biological functions [70,71,72,73,74,75]. Uptake of miRNAs by gastric and intestinal cells as well as their potential effects on the gut microbiota by orally delivered miRNAs and their potential immunomodulatory properties indicate the possibility of cross-species or cross-kingdom communication through miRNAs [75]. Because of these observations, one potential method of administering miRNAs is orally. MiRNAs are often associated with extracellular vesicles (EVs), RNA-binding proteins, lipoproteins, or lipid derivatives, along with nanoparticles. These protective elements shield miRNAs from the adverse gastrointestinal environment, including salivary and pancreatic RNases, the stomach’s low pH, digestive enzymes, peristaltic activity, and microbial enzymes. Such protection likely facilitates the absorption of miRNAs from the digestive tract [75]. However, there is ongoing debate surrounding the absorption, stability, and physiological impact of these food-derived miRNAs and there are contrary findings regarding the bioavailability and the in-human functionality of miRNAs contained in plant food [76,77].
Ongoing research continues to unravel new insights into the molecular mechanisms underpinning the dysregulation of miRNA biogenesis and expression in cancer. For example, it is widely acknowledged that genetic deletions or amplifications, epigenetic methylation of miRNA genomic loci, and modifications influencing the regulation of primary miRNAs (pri-miRNA) by transcription factors, as well as components involved in the miRNA biogenesis pathway frequently alter miRNA expression and function in many cancers [56,78,79]. Furthermore, additional factors, such as oncogenic drivers like mutations in the KRAS gene, can also affect global miRNA biogenesis and effector function, contributing to the broader dysregulation of miRNAs [80]. Consequently, miRNAs and their dysregulation have garnered significant interest from both academia and industry as a focal area of research for both understanding of disease biology and explore their applications as potential diagnostic, prognostic, and predictive biomarkers [68] as well as well as their roles as drug targets or therapeutic agents [81].
It is well-established that miRNAs are powerful genetic regulators of diverse biological and developmental processes, in addition to playing a pivotal role in the pathogenesis of various diseases. This is due to the ability of a single miRNA to regulate entire cellular pathways by interacting with numerous target genes [77]. Because of this, miRNAs have emerged as a novel class of therapeutic agents with the potential to restore disrupted cellular functions, particularly in various malignancies, including cancer. However, the very potency of miRNAs can be a double-edged sword. Their far-reaching effects, while beneficial, can also lead to off-target effects in non-targeted tissues, a concern documented in recent clinical trials [82,83,84]. Managing these off-target effects represents a significant challenge to be addressed. Take, for instance, MRX34, a miR-34a mimic encapsulated within a liposome-formulated nanoparticle (NOV40) was evaluated in a first-in-human, Phase 1 study in patients with advanced solid tumors, including melanoma NSCLC, hepatocellular carcinoma, renal carcinoma.
Although MRX34 exhibited significant efficacy, with three patients achieving prolonged confirmed partial responses and 14 patients maintaining stable disease (median duration, 136 days) [85]; however, the clinical trial was terminated due to the occurrence of serious immune-mediated adverse events which resulted in the deaths of four patients (NCT01829971) [82,83,84]. Despite the setback, dose-dependent modulation of pertinent target genes provides proof-of-concept for miRNA-based cancer therapy.
This review discusses the dysregulation of miRNA expression in cancer and their potential as therapeutics and further discusses the main challenges and strategies to address the problems that must be overcome to fully harness the therapeutic potential of miRNAs.
2. miRNAs
Following the discovery of lin-4, the first miRNA in 1993 in Caenorhabditis elegans [86,87] [79,80], it became evident that miRNAs are widespread in the animal and plant kingdoms, some of which exhibit high levels of conservation across species [88,89,90].
MiRNAs, which are short non-coding RNA molecules typically composed of approximately 22 nucleotides, are naturally encoded in the genomes of various species [88,89,90,91] and play vital roles in regulating gene expression at both transcriptional [5,6,7] and post-transcriptional [8,9,10,11,92] levels of their target mRNAs [8,10] by and via the modulation of the stability and translation of mRNA [93] in a broad range of biological processes [94], impacting activities such as cell differentiation, proliferation, angiogenesis, and apoptosis. Furthermore, miRNAs display specific patterns of expression in different tissues [12,13] and during various stages of development [14,15,16].
There are currently estimated to be more than 2588 mature human miRNAs present in human cells [95], each with a unique temporal and tissue-dependent expression pattern. These miRNAs are estimated to control over 60% of human gene expression, showcasing their significant regulatory roles in diverse physiological processes. Because a single microRNA can regulate multiple genes, many miRNAs can contribute to the development of many human diseases when they become dysfunctional [2,8,18,20,21,22,23,24,25,26,28,30,31,32,33,34,35,37,66,96,97,98] including many types of cancer [39,41,42,43,44,47,50,51,53,55,56,57,58,59,60,61,62,63,64,65,66,67,69,99,100,101,102,103,104].
However, determining the precise relevance of individual miRNAs has been challenging, despite their evident significance as regulatory molecules [105]. Studies investigating miRNA functions by either overexpressing or silencing specific miRNAs have generated data that sometimes conflict with findings from loss-of-function models [105]. For example, studies in Caenorhabditis elegans involving systematic miRNA deletions suggest that fewer than 10% of the miRNAs are individually essential for normal development or viability [106] and this trend appears consistent in mice as well [97].
As illustrated in Figure 1 and discussed in detail in the literature, miRNAs are primarily transcribed from DNA sequences into primary miRNAs (pri-miRNAs), which undergo initial processing by DROSHA in the nucleus to generate precursor miRNAs (pre-miRNAs) [8,68,107]. It’s worth noting that as many as 40% of miRNA genes may be located within the introns or exons of other genes [108]. Pre-miRNAs are subsequently transported from the nucleus to the cytoplasm by exportin 5 (XPO5), where they are further processed by DICER, resulting in small RNA duplexes with specific 3’ overhangs of 2 nucleotides. These double-stranded RNA duplexes are loaded onto the Argonaute (AGO) protein, which retains one mature miRNA strand while discarding the other [10]. The AGO-miRNA complex, along with co-factors like GW182 (TNRC6A), forms the RNA-induced silencing complex (RISC) [92], responsible for mRNA transcript degradation and translational inhibition through interaction with complementary mRNA target sequences, typically located within the 3’-untranslated region (3’-UTR) of mRNAs (Figure 1) [109,110,111,112]. The interaction between miRNA and target mRNA typically takes place at the 5′ end of the miRNA, known as the ‘seed’ region. Yet, recent evidence points to a unique group of target mRNAs that bind the miRNA not just through the seed but also via a complementary region at the 3′ end of miRNAs. This extended complementarity displaces the miRNA from Ago2, rendering it vulnerable to enzymatic degradation. This process is referred to as the target-directed miRNA degradation mechanism (TDMD) [113,114].
miRNAs are regarded as master regulators of the genome due to their ability to bind to and alter the expression of many protein-coding RNAs [115]. Because of this, a single miRNA can potentially regulate distinct mRNAs (anywhere from 10 to 100 protein-coding RNAs) due to their ability to bind to target mRNAs even when the pairing is not perfect [55,116]. As a result, a single miRNA can regulate a pool of targets involved in similar cellular processes and pathways, thereby amplifying the cellular response and potentially making miRNAs powerful therapeutics to restore cell functions that are altered as part of a disease phenotype. Conversely, a specific messenger RNA can become the target of many miRNAs, whether concurrently or in a context-dependent manner [117], leading to a collaborative repression effect [118,119]. Bioinformatic analyses indicate that a single miRNA can potentially bind to as many as 200 distinct gene targets with various functions, such as transcription factors, receptors, and more (https://bitesizebio.com/24926/mysterious-mirna-identifying-mirnas-and-their-targets/).
3. miRNAs role in cancer
Cancer is a complex and heterogeneous disease characterized by a series of genetic and genomic aberrations that promote tumorigenesis [120]. These changes within the genome, which affect gene function, often arise from genomic aberrations like chromosomal translocations, insertions, deletions, amplifications, single-nucleotide mutations, or in the epigenome. These genetic and epigenetic aberrations frequently lead to the activation of oncogenes and the suppression of tumor suppressor genes [121]. In addition, miRNAs have been identified as additional genomic regulators that also play a crucial role in various aspects of organismal development, normal physiological processes, and the development of disease, including many types of cancers [68]. It has been demonstrated that miRNAs play a pivotal role in all the known processes involved in cancer including proliferation, survival, metastasis, and apoptosis [115]. Data suggest that dysregulation of miRNA function, either through its loss or gain, contributes to cancer development by either upregulating or silencing specific target genes. As a consequence, utilizing miRNAs either as miRNA mimics or antagomirs could present a potent therapeutic strategy to interfere with key molecular pathways associated with cancer as such miRNAs have the capacity to regulate all the recognized hallmarks of cancer, either acting as tumor suppressors or promoting oncogenic processes. Several of these cancer hallmarks influenced by miRNAs are discussed in detail in literature [65,66].
It is generally acknowledged that various changes in miRNA genes and their expression, such as genetic deletions or amplifications, epigenetic methylation of miRNA gene locations, and modifications that influence the regulation of primary miRNA (pri-miRNA) through transcription factors, as well as factors involved in miRNA biogenesis process frequently alter miRNA expression and function in a wide range of cancer types [66].
In addition, changes in the miRNA biogenesis process can also impact the availability of target mRNAs, including those associated with the development of cancer (Figure 3) [122]. When miRNAs or the machinery involved in miRNA processing are altered or dysregulated this often leads to the loss of normal homeostatic state, leading to malignant transformation, including various types of cancer [51,52,56,65,66,67,123].
Because of miRNAs key role in controlling the expression of numerous genes that are involved in cellular responses to environmental stressors such as DNA damage, hypoxia, oxidative stress, and nutrient deprivation, they can function either as oncogenes (oncomiRs), or as tumor suppressors (onco-suppressor miRs). This is supported by recent findings that have identified miRNAs with oncogenic and tumor-suppressing roles in a range of neoplastic malignancies, and the dysregulation of miRNA expression is closely linked to the initiation, progression, and metastasis of cancer [43,45,104].
Furthermore, dysregulated circulating miRNAs have been shown to be associated with the origin, progression, treatment response, and the survival of patients with the disease [124,125]. For instance, the unique tissue-specificity of miRNAs [13], which is essential for maintaining normal cells and tissues [40], renders them promising candidates as potential biomarkers for diagnosing cancers of unknown primary [126,127]. Furthermore, given the frequent genetic and epigenetic alterations observed in specific miRNAs and components of the miRNA biogenesis process in different cancer types, the oncogenic and tumor suppressor miRNAs have now emerged as promising candidates for miRNA-based therapeutics and diagnostic applications.
4. RNA therapeutics
As discussed in detail in literature, in the recent years, more than 50 siRNA-based drugs have entered clinical trials (phase I, II, and III) [128,129]. Among those, approximately 15 phase I-, II-, and III-programs based on siRNA therapeutics are being explored for the treatment of various cancer types [129]. Two siRNA-based drugs Patisiran and Givosiran (Alnylam Pharmaceuticals) were approved by the Food and Drug Administration (FDA) in 2018 and 2019 for hereditary transthyretin-mediated amyloidosis and acute hepatic porphyria, respectively [130,131]. The first example of an FDA approved RNA-based drug, a siRNA-based therapy developed by Alnylam Pharmaceuticals is Patisiran, sold under the brand name Onpattro™ for the treatment of polyneuropathy of hereditary transthyretin-mediated amyloidosis in adults. Based on the completion of a successful Phase III APOLLO trial, Onpattro™ was approved by the US FDA in August 2018. Onpattro™ contains patisaran, which comprises a siRNA targeting transthyretin (TTR) mRNA conjugated with a lipid complex which leads to decrease in TTR protein levels in the liver thus resulting in a reduction in amyloid deposits. Patisiran specifically targets and binds to a genetically conserved sequence in the 3’ untranslated region (3’UTR) of mutant and wild-type TTR mRNA [132]. Findings from the APOLLO trial, a placebo (77 patients) controlled Phase III trial which enrolled 225 patients showed that 51% of patients receiving Onpattro™ (148 patients, once every three weeks (0.3 mg/ kg body weight)) exhibited an improved quality of life (measured using the Norfolk Quality of Life Diabetic Neuropathy (QoL-DN)), as compared to only 10% in the control arm of the patients that received a placebo drug [132,133].
With the onset of the COVID-19 pandemic, mRNA technology emerged as a pivotal force, serving as the cornerstone for remarkably effective mRNA-based vaccines that have played a crucial role in mitigating the spread of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). The groundbreaking science behind mRNA vaccines earned Katalin Karikó and Drew Weissman the prestigious 2023 Nobel Prize in Physiology or Medicine for their pioneering work on nucleoside base modifications, enabling the development of these impactful COVID-19 vaccines.
The evolution of cap analogs has vastly improved mRNA translation, while advancements in purification, packaging, and delivery methods have revolutionized medicine. Visionaries like Katalin Karikó, Drew Weissman, Edward Darzynkiewicz, Robert Rhodes, Ugur Sahin, and Ozlem Tureci made pivotal early contributions to mRNA research, deserving recognition for their pioneering efforts. This mRNA narrative charts a remarkable journey of breakthroughs in a field holding immense promise for the future of medicine.
The success of mRNA vaccines has paved the way for mRNA-based technology in personalized neoantigen vaccines, seamlessly integrating them into standard oncological workflow [134,135]. These mRNA-based vaccines can be tailored and manufactured as individualized vaccines with multiple neoantigens [136], and can effectively stimulate antigen-presenting cells [137,138,139,140] and be delivered using clinical-stage delivery formulations [141]. The studies and insights from the mRNA-based COVID-19 vaccines highlight the promise of RNA therapeutics as an innovative class of treatments.
However, the effectiveness of miRNA and other nucleic acid-based therapies hinges on a reliable delivery method with minimal adverse events and drug or treatment related toxicity. Delivering miRNA treatments into cells presents challenges due to the need for precise targeting of diseased cells while avoiding healthy ones. Unlike mRNA COVID-19 vaccines, which are taken up by scavenging immune cells, miRNA therapeutics must evade immune recognition to reach their target cells without triggering an immune response.
5. miRNA therapeutics
The discovery of the link between miRNAs and human diseases in 2002 sparked a strong interest in their potential as a new class of therapies. Consequently, interdisciplinary fields encompassing biology, chemistry, and medical science have made significant investments in the development of miRNA-based therapies.
As illustrated in Figure 2 and Table 3, and discussed in literature in detail [77,122,128], there are only a few miRNA therapeutics that have entered clinical trials with none of them entering Phase III or being approved by the FDA and several of them were terminated due to toxicity. Despite the significant progress made in preclinical research, the progress in the field of miRNA-based diagnostic [68] and therapeutic applications remain at an early stage and only a few of these miRNA-based therapies have advanced to clinical development. Given this situation, several efforts in the biotechnology and pharmaceutical industry have integrated miRNAs into their development pipelines focusing on the development of two categories of miRNA drugs, miRNA mimics and inhibitors (antagomirs or antimirs) (Figure 2, Table 2 and Table 3) [77,142]. As a consequence, several of miRNA-based therapeutics that are being tested in clinical trials are continuously growing for the treatments of a variety of genetic, metabolic, and oncological conditions [143,144,145].
MiRNA therapeutics are a type of RNAi-based therapeutic that targets and modulates the activity of specific endogenous miRNAs in the body. Because miRNAs play a crucial role in regulating gene expression in normal health and disease states, by targeting and manipulating specific miRNAs, miRNA therapeutics aim to treat various diseases by restoring (miRNA mimics) or correcting miRNA expression patterns (antagomir). MiRNA-based therapeutic programs for cancer are predominantly conducted by a few biopharmaceutical companies, including Santaris Pharma, Roche Pharmaceuticals, Regulus therapeutics (San Diego, CA, USA), Mirna Therapeutics Inc. (Carlsbad, CA, USA), miRagen Therapeutics (Boulder, CO, USA), and EnGeneIC (Sydney, Australia).
Immune evasion and chemotherapy resistance is a challenge in cancer therapy and this resistance can be mediated by various factors including miRNAs induced by tumor microenvironment stimuli, like hypoxia or cell–cell communication [146].
Hypoxia has been shown to influence microRNA expression in cancer and stromal cells in the tumor microenvironment (TME) via downregulation of factors involved in miRNA biogenesis machinery or regulation of transcription factors that control miRNA expression. Accordingly, many hypoxia-regulated miRNAs and their role in tumor progression have been reported. These hypoxia-regulated miRNAs including miR-26a, miR-181b, miR-210, miR301-a, miR-424, and miR-519c have also been associated with chemo- or radiotherapy response in different cancers [147,148,149,150]. Therefore, therapeutic targeting of these miRNAs could be a strategy to re-sensitize hypoxic tumors to chemo and other therapies. For example, in the hypoxic pancreatic cancer microenvironment, HIF-1α induces gemcitabine resistance. A recent study showed that delivery of miR-519c, which is downregulated in pancreatic cancer, could inhibit HIF1-α in gemcitabine-resistant pancreatic cancer cells under hypoxia [151]. Furthermore, a redox-sensitive nanoplatform used to co-deliver miR-159c and gemcitabine was demonstrated to inhibit the expression of HIF-1α and genes responsible for glucose uptake and cancer cell metabolism, thus inhibiting orthotopic desmoplastic pancreatic cancer growth in NSG mice and reversing hypoxia-induced chemotherapy resistance [151]. Similarly, tumor suppressor miR-34a has been shown to downregulate the expression of more than 30 oncogenes across multiple oncogenic pathways, as well as genes implicated in tumor immune evasion, but is lost or under-expressed in many malignancies [82]. However, if miRNAs are to be used for the treatment of a cancer, miRNAs must be delivered to the target tissue, not trigger an immune response and be economically feasible so that wide-spread adoption of these nano therapies can be realized. Although there has been some success in clinical development, several clinical trials have been terminated mostly due to various serious adverse events, indicating that there are still several challenges to overcome before the clinical application of RNAi-based therapies becomes widespread.
5.1. Examples of miRNA therapeutics in clinical trials
As illustrated in Figure 2, and Table 2 and Table 3, and discussed in detail in the literature [85,145], there are several miRNA-based therapeutics being tested both in preclinical studies (Table 2) or in human clinical trials (Table 3).
Miravirsen: The first miRNA-based therapeutic entering clinical trials was Miravirsen (SPC3649), a miR-122 15-mer LNA-PS-modified modified ASO antagomir of miR-122, as a therapy against Hepatitis C Virus (HCV) infections developed by Santaris Pharma, Roche Pharmaceuticals. miR-122 was shown to play a role in HCV replication [152]. Phase II clinical trials were conducted to evaluate the safety and antiviral efficacy of Miravirsen in patients with chronic HCV infection. Miravirsen showed strong efficacy by reducing viremia in patients with HCV [155,156,157] and underwent multiple phase II clinical trials (NCT01200420, NCT01872936, NCT02031133, NCT02508090). However, due to severe side effects the trial was halted [145,153].
RG-012: RG012 is an anti-miR-21 therapy developed by Regulus Therapeutics for the management of Alport syndrome. miR-21 has been shown to be up-regulated in fibrotic kidney disease. Preclinical studies have shown that treatment with an anti-miR-21 significantly reduced kidney failure by reducing the rate of progression of renal fibrosis. RG-012 has been granted orphan drug status in the US and Europe. However, some sequence-independent side effects have been reported in context with phosphorothioate-modified oligonucleotides [[154].
RG-101: RG-101, an antagomir of miR-122 developed by Regulus Therapeutics is an N-acetyl-d-galactosamine(GalNAc)-conjugated synthetic RNA oligonucleotide that targets and inhibits miR-122, which is involved in HCV replication for patients with HCV. in addition to its essential role for HCV replication, miR-122, a liver-specific miRNA, has relevant functions in liver metabolism [155] [20]. This miRNA is also an essential host factor for HCV.
Clinical trials were conducted to evaluate RG-101’s safety and efficacy as a potential treatment for chronic HCV infection. Although RG-101 showed considerable efficacy and showed a significant reduction in viral loads in chronic HCV subjects [156]; however, the trial was terminated due to some serious adverse events of severe hyperbilirubinemia [156].
MRG-201: MRG-201 is a synthetic RNA oligonucleotide that targets and activates miR-29, which has been shown to inhibit fibrosis. Clinical trials were conducted to assess MRG-201’s safety and efficacy in treating fibrotic disorders such as hypertrophic scars and idiopathic pulmonary fibrosis.
MRX34: MRX34, developed by miRNA Therapeutics Inc. is a synthetic miRNA mimic designed to mimic the activity of a tumor suppressor miR-34a encapsulated into a liposome-formulated nanoparticle (NOV40) for the treatment of advanced solid tumors including melanoma, NSCLC, hepatocellular carcinoma, and renal carcinoma. miR-34a is a naturally occurring tumor-suppressor miRNA expressed at reduced levels in many tumor types. MRX34, is considered to be a first-in-class miRNA mimic for the treatment of many cancers, such as non-small cell lung cancer, hepatocellular carcinoma, colon cancer, ovarian cancer, cervical cancer, and others. The formulation was tested in a phase I clinical trial [82].
Although MRX34 displayed strong activity, at the end of the trial, only three patients achieved prolonged confirmed partial responses, and 14 patients presented with stable disease (median duration- 136 days) [85]. However, the trial was terminated due to serious immune-mediated adverse events that resulted in four patient deaths (NCT01829971) [82,83,84]. After this, MiRNA Therapeutics ceased operations in 2017 and agreed to merge with Synlogic Inc.
Cobomarsen (MRG-106): MRG-106 (Cobomarsen), an LNA-based antagomir of miR-155 was developed by Miragen Therapeutics (Viridian Therapeutics Inc) that aimed to inhibit the activity of miR-155 [157,158] in several lymphoma subtypes, as well as in diffuse large B-cell lymphoma [102] [83] where miR-155 is up-regulated. Phase II clinical trials are being conducted to assess its effectiveness in treating certain cancers and immune disorders including cutaneous T-cell lymphoma (CTCL), chronic lymphocytic leukemia, diffuse large B-cell lymphoma, and mycosis fungoides (NCT03837457), and adult T-cell leukemia/lymphoma (NCT02580552, NCT03713320). While Phase I trial was completed, two of the Phase II studies were terminated. The study was terminated early for business reasons, and not due to concerns regarding safety or lack of efficacy. (https://classic.clinicaltrials.gov/ct2/show/NCT03713320).
MRG-107: MRG-107 an antagomir of miR-155 was developed by Miragen Therapeutics (Viridian Therapeutics Inc) that aimed to inhibit the activity of miR-155. miR-155 plays relevant functions in the immune mechanisms and inflammation processes in amyotrophic lateral sclerosis (ALS) and miR-155 is elevated in the spinal cords of ALS patients. In preclinical models of ALS, inhibition of miR-155 has reduced the ALS symptoms and extended survival [145].
MRG-110: MRG-110 is a synthetic antagomir of miRNA-92a developed by MiRagen Therapeutics in collaboration with Servier to treat ischemic conditions such as heart failure [159]. MRG-110 is developed to promote the growth of new blood vessels by inhibiting miR-92a. MRG-110 is being studied to determine if it can accelerate healing of wounds by improving blood flow into the wound area. A phase I clinical trial that tested safety, tolerability, pharmacokinetics, and pharmacodynamics of MRG-110 following intradermal Injection in healthy Volunteers was recently completed (NCT03603431).
Remlarsen (MRG-201): Remlarsen (MRG-201) is an LNA RNA mimic of miR-29 developed by MiRagen Therapeutics for keloid disorder and decreases the expression of collagen and other proteins that are involved in scar formation and has been shown to inhibit fibrosis [160]. miR-29 family members are typically downregulated in fibrotic diseases [161]. Clinical trials is being conducted to assess Remlarsen’s safety and efficacy in treating fibrotic disorders such as hypertrophic scars and idiopathic pulmonary fibrosis when administered by intradermal injection at the site of an excisional wound [160] 90]. The phase II clinical trial is currently underway to determine if it can limit the formation of fibrous scar tissue in certain diseases (NCT03601052).
TargomiRs: The MesomiR 1 trial (NCT02369198) tested the safety and efficacy of miR-15/16 encapsulated in bacterial minicells (TangomiRs) in patients with recurrent malignant pleural mesothelioma (MPM. TangomiRs developed by EnGeneIC to deliver miR16 mimics encapsulated in TargomiRs composed of nonliving bacterial nanocells with anti-EGFR bispecific antibody to target EGFR-expressing cancer cells were tested as 2nd or 3rd Line Treatment for patients with recurrent malignant pleural mesothelioma (MPM) and non-small cell lung cancer (NSCLC) (NCT02369198) [162]. More specifically, mir-16 mimic was encapsulated in EnGeneIC’s bacterially-derived EDV™ (EDV)TM nanocells (a 400 nm particle of bacterial origin able to carry a drug cargo) and targeted with EGFR antibodies (TargomiRs). miR-15/16 are implicated as tumor suppressors in MPM. Although variable response rates were observed with 5% of the patients showing partial response, 68% showing stable disease and 27% showing progressive disease after low dose systemic administration of TargomiRs; however, dose-dependent toxicities were observed (i.e., anaphylaxis, inflammation as well as cardiac events) [128,162].
MGN-1374: MGN-1374, an 8-mer LNA ASO developed by miRAgen Therapeutics, is designed to specifically target the seed region of the miR-15 family and is currently in the preclinical phase for the control of postmyocardial infarction remodeling.
RGLS4326: RGLS4326 is a single-stranded, chemically modified, 9-mer ASO with full complementarity to the seed sequence of miR-17. RGLS4326 is designed to inhibit the pathologic functions of the miR-17 family in Polycystic kidney disease (ADPKD) [163] [87], an autosomal dominant disease, one of the most frequent monogenic disorders, caused by mutations in the PKD1 or PKD2 gene and therapeutic options for the treatment of ADPKD are limited. A phase I clinical trial of RGLS4326 was recently completed (NCT04536688).
Additionally, Table 2 highlights several miRNA-based drugs currently under preclinical investigation, targeting various diseases such as Peripheral Arterial Disease, Chronic Heart Failure, Amyotrophic Lateral Sclerosis (ALS), among others.
Moreover, miRNA therapeutics in combination with chemotherapeutic agents have also been explored to overcome cancer therapy resistance [142]. Studies indicate that combining therapeutic miRNAs with chemotherapy can decrease the required drug doses for cancer treatment [164,165]. For example, miR-3622b-5p, when paired with cisplatin, not only enhances apoptosis but also sensitizes ovarian tumor organoids to cisplatin [166], suggesting the potential of miRNAs in combination with chemotherapy to address cancer treatment and counteract drug resistance.
5.2. Small molecule modulators of miRNA expression
Because altered levels of miRNA expression is associated with many cancers, restoring the function of tumor suppressor miRNAs by overexpressing or introducing of miRNA mimics to restore to their relatively normal physiological levels or function or by inhibiting overexpressed oncogenic miRNAs by miRNA inhibitors (antagomirs), or miRNA sponges in cancer represents two major strategies for miRNA therapeutics in cancer [122,167] (Figure 3).
The function of repressed miRNAs can also be restored to their relatively normal physiological levels and function by using some small molecules that can transcriptionally activate the expression of miRNA genes leading to expression of endogenous miRNAs and restoring the expression of tumor suppressive miRNAs. Conversely, overexpressed oncogenic miRNAs can also be suppressed by small-molecule inhibitors.
As illustrated in Figure 3 and Table 1 and discussed in literature [122], because nucleic acid-based therapeutics have poor cell-permeability for drug delivery, in recent years small-molecule drugs in the regulation of miRNA expression have been explored since they can cross the cell membrane by free diffusion and can modulate the expression of miRNAs and also traditional drug development can be applied for the development of novel miRNA inhibitors (or activators) [168] (Wu, 2020b).
Various small molecule inhibitors of various miRNAs with various chemical structures and different mechanisms of action (Table 1) have been described with different target sites of inhibition and interference across the whole process of miRNA biogenesis such as processing, maturation, and function (Figure X) [122].
For example, as discussed in literature [122], reduced expression of tumor suppressor miRNAs can be reactivated to their normal physiological levels by some small molecule compounds, such as hypomethylating agents [169].
Decitabine or 5-azacytidine are two drugs for the treatment of myelodysplastic syndrome and were shown to upregulate the expression of several miRNAs [106].
Similarly, enoxacin was shown to activate the expression of several miRNAs in vitro [170] and to suppress tumor growth by upregulating the expression of 24 miRNAs in vivo in mice xenograft models [170] suggesting that small molecule compounds can potentially restore miRNA expression and function to a more physiological setting.
For example, miR-21 is one of the tumor-associated miRNAs (oncomiR) has been shown to be upregulated in a variety of tumor cells, including breast cancer, ovarian cancer, colon cancer, pancreatic cancer, thyroid cancer, and others and its high expression in cancer which is closely associated with tumorigenesis [171,172,173,174].
Trypaflavine (TPF), a small molecule inhibitor of miR-21 was shown to suppress the expression of miR-21 [175].
Trypaflavine was shown to inhibit the formation of RISC by blocking the assembly of miR-21 and AGO2 protein, leading to the suppression of the expression level of miR-21.
Similarly, Kanamycin A was shown to inhibit the expression of let-7 by binding to pre-let-7 and interfering with Dicer [176].
Furthermore, small-molecule inhibitors of miR-122, NSC 158959 and NSC 5476 [177] which may be involved in the modulation of transcription of miR-122 gene to pri-miR-122.
Importantly, mir-122 is a liver specific miRNA, accounting for about 72% of the total miRNA in the adult liver and is one of the earliest miRNAs with tissue-specific and high abundance expression [178].
miR-122 plays a key role in the regulation of cholesterol and fatty-acid metabolism in the adult liver suggesting that miR-122 may be an attractive therapeutic target for metabolic disease [179].
Moreover, it was also shown that miR-122 also plays a key role in the development of various types of liver diseases including acute and chronic liver injury,, liver tumor and hepatitis C virus (HCV) infection, liver cirrhosis, and alcoholic hepatitis [180].
miR-1 which is abundantly expressed in skeletal muscle cells plays a role in the regulation of the formation of skeletal muscle cells and the development of muscle and is closely associated with the development of the heart [181,182].
Various small molecular Inhibitors of miR-1 have been identified from photocycloadducts of acetylenes with 2-methoxy-1,4-naphthalenequinone as the basic skeleton by photocyclization reaction [183]. For example, the small molecule 2-methoxy-1,4-naphthalenequinone was demonstrated to exert specific inhibitory effects on miR-1 and was shown to significantly inhibit the expression of mature miR-1 in cells. However, the specific mechanism of action of 2-methoxy-1,4-naphthalenequin for its inhibitory function on miR-21 remains to be elucidated.
6. Advances in delivery of miRNA therapeutics
Although a few phase 1 and 2 clinical trials involving miRNA-targeting or miRNA-based therapeutics, there are no miRNA-based therapeutics in phase III trials so far. This is partly due to the challenge of targeted delivery of miRNAs to specific cell types, tissues, and organs. Although targeting of miRNAs to a cell of interest by using various methods (e.g. antibodies, nanoparticles, or ligands) have been reported to improve the efficacy of miRNAs and reduce off-target effects (e.g., immunotoxicity [184], there are still limitations and challenges for miRNA therapeutics warranting more research in this field.
As illustrated in Figure 4 and discussed in detail in the literature in detail [77,84,185,186], there are various strategies being explored as a mechanism to deliver miRNA therapeutics (mimics and antagomirs) to the indented tissue and to improve the pharmacokinetic mechanisms, and avoid off-target effects. These include vector-based and non-vector-based methods including lipid-based nanoparticles, polymeric vectors/dendrimer-based vectors, cell-derived membrane vesicles, 3D scaffold-based delivery systems, and other nanoparticles derived from polymers and metals that are biodegradable and biocompatible.
As such, there is a need for an efficient delivery system for developing miRNA-based therapeutics.
As discussed in detail in literature [187], several different strategies have been explored to efficiently deliver RNA-based therapeutics (Figure 4) including vector-based and non-vector-based delivery systems including adeno-associated virus, lentivirus, bacterial nanocells [188], bacteriophages, cationic lipid-based liposomes, including monovalent and multivalent lipids, natural polymer-based nanoparticles, polymer based nanoparticles conjugated with polyethylene glycol (PEG), extracellular vesicles (EVs) or exosomes, nanocomplex-forming functionalized metals such as gold nanoparticles, and carbon nanotubes and many others that are engineered to contain biomolecule conjugates for improved stability and pharmacokinetics and target delivery to the intended cell or tissue type [84,186,187,189,190]. A recent report demonstrated enhanced antitumor potency of STING agonists after covalent conjugation of cyclic dinucleotides (CDN) to polymer nanoparticles (poly(β-amino ester) formulation for the intravenous delivery [191].
Non pathogenic recombinant viral vectors including retroviruses and lentiviruses (which bears a risk for genomic integration), adenovirus, and adeno-associated viruses (only remains transiently stable in a episomal form in the host cell’s nucleus [192,193] encoding the intended RNA are also being explored for intracellular delivery of miRNA-based therapeutics and hence become a major focus of attention [194]. A Phase II trial is currently testing an adeno-associated viral vector for the delivery of the miRNA drug AMT-130 for the treatment of Huntington’s disease (ClinicalTrials.gov identifier NCT04120493) [195,196,197]. Despite their potential for the delivery and expression of miRNAs, there are various side effects with the use of viral vectors such as immunogenicity and transgene-related immune responses [198].
Packaging of the negatively charged nucleic acids in liposome nanoparticles mask their negative charge and also protects against serum nuclease degradation [190,199]. Delivery of miRNAs using liposome nanoparticles has already been applied in several clinical studies, such as MRX34 (NCT01829971, NCT02862145) [82,83].
Similarly, miRNA-loaded bacterial minicells were used for the delivery of miR-16 mimics in a phase 1 trial in patients with recurrent malignant pleural mesothelioma (MesomiR 1, NCT02369198) [162,200]. However, the study also reported several side effects including dose-limiting toxicities, decreased lymphocyte counts, or cardiac events [162].
Extracellular vesicles (and exosomes) that can be loaded with a desired cargo to permit in-body cellular transfer are also being explored as drug delivery systems [201]. EVs derived from mesenchymal stromal cells from human adipose tissue were engineered to package miR-125b which consequently was shown to inhibit the proliferation of human hepatocarcinoma cells [202].
In addition, different modalities of drug delivery systems have been explored for the delivery of miRNA-based drugs such as core-shell magnetic nanoparticles, quantum dot nanocrystals, polymeric micelles, and mesoporous silica nanoparticles are among the other examples of nanocarriers as drug-delivery systems to improve the therapeutic effectiveness and specificity, and tissue targeting of miRNA and other nucleic acid therapeutics [145].
For example, covalent conjugation of miRNAs and other nucleic acid-based drugs and biomolecules to lipids, peptides, or sugars that work via receptor-mediated endocytosis mechanisms is another promising strategy [190].
Similarly, lipophilic cholesterol conjugate was used for a cell type–independent delivery of miR-29–based mimic (remlarsen/MRG-201) to human skin fibroblasts by skin injection in a Phase II trial on keloid disorder to repress extracellular matrix expression and fibroplasia in the skin ( NCT02603224, NCT03601052) [160].
Aptamer conjugates coupled to corresponding miRNA therapeutics by simple sticky-end annealing [203] is yet another strategy for the delivery of miRNAs to intended cell type. Aptamers are single-stranded nucleic acids that are developed as high-affinity ligands specific to a cell surface receptors to facilitate the delivery of therapeutic cargo including miRNAs through receptor-mediated transport [190,203]. Aptamer-conjugated miRNAs, including the Aptamer-miR-34c conjugate (GL21.T-miR-34c) are currently being explored in preclinical studies in non-small-cell lung cancer cells [204].
Other biomolecule conjugates including N-acetylgalactosamine (GalNAc) which enables a targeted delivery of nucleic acid therapeutics via endocytosis by the stimulation of the liver cell–specific asialoglycoprotein receptors have also been explored in clinical trials [205,206]. GalNAc conjugated to a miR-122 inhibitor (RG-101) and to an miR-103/107 inhibitor (RG-125/AZD4076) were explored in clinical trials for chronic HCV [EU Clinical Trials Register (clinicaltrialsregister.eu) EudraCT numbers 2015-001535-21, 2015-004702-42, 2016-002069-77] and steatohepatitis (NCT02612662, NCT02826525), respectively [206,207,208]. However, because of some side effects (i.e., jaundice cases), the RG-101 clinical trial was halted and the cause of which is still under investigation [156,209,210].
Other examples of the use of GalNAc-Conjugated LNA, anti-miR-122 antisense oligonucleotides, or nano-carrier vehicles in combination with cell type-specific biomolecule conjugates or miR-155 inhibitors by gold nanoparticles functionalized with antagomir and AS1411 aptamer have been explored in recent preclinical studies [211,212].
In addition, the 3D matrices for delivering nucleic acid–based therapeutics and conventional drugs are currently undergoing optimization with diverse design features. This encompasses various application routes, such as edible or injectable carriers [77,213,214,215]. One potential method of administering miRNAs is orally [75]. MiRNAs are often associated with extracellular vesicles (EVs), RNA-binding proteins, lipoproteins, or lipid derivatives, along with nanoparticles which shield miRNAs from the adverse gastrointestinal environment, including salivary and pancreatic RNases, the stomach’s low pH, digestive enzymes, peristaltic activity, and microbial enzymes. Such protection likely facilitates the absorption of miRNAs from the digestive tract [75]. However, there is ongoing debate surrounding the absorption, stability, and physiological impact of these food-derived miRNAs.
7. Progress in chemical modifications of miRNAs for improved stability and cellular uptake
As illustrated in Figure 4 and discussed in the literature [77], a combination of chemical modifications, biomolecule conjugation, or the use of carriers improves site-directed and efficient cell targeting of miRNA and other nucleic acid therapeutics. In addition, various types of chemical modifications of nucleobases, ribose sugar, or the phosphate backbone can mask the negative charge of the miRNAs and other nucleic acids and increase their adhesion to the cell surface, thereby facilitating cellular uptake [190,216] and also improving their stability.
Among the commonly used nucleic acid modifications, locked nucleic acid (LNA) bases [216] are characterized by the introduction of methylene bridges which reduce the flexibility of the ribose ring, resulting in a locked conformation of the modified nucleotides [217,218].
LNA-modified RNA-based therapeutics are more resistant to ribonucleases and exhibit improved cellular uptake, primarily through an endocytosis mechanism that remains not fully understood [219]. The locked conformation enhances the capability of RNA-based therapeutics to establish stable duplexes by binding to and inhibiting the function of the target miRNA.
Consequently, LNA-modified RNAs are frequently employed in single-stranded antagomirs, such as antisense oligonucleotides (ASOs) and this locked conformation enhances the capability of ASOs to establish stable duplexes by binding to and inhibiting the function of the target miRNA [216,219]. Due to these benefits, LNA-modified oligonucleotides have emerged as one of the primary approaches for inhibitory therapeutics targeting both miRNA and mRNA.
Phosphorothioate modifications, involving the introduction of a sulfur atom into the oligonucleotide’s phosphodiester backbone, have emerged as a promising method for improving oligonucleotide stability and promoting endosomal uptake, particularly through stabilin receptors found on cell surfaces (e.g., in kidney cells) [220,221]. The latter approach has been utilized for the targeted renal delivery of synthetic miR-21–anti-miR (RG-012/lademirsen/SAR339375) in a clinical investigation involving Alport syndrome (ClinicalTrials.gov identifiers NCT03373786, NCT02855268) [222]. However, certain sequence-independent effects have been documented in association with phosphorothioate-modified oligonucleotides [154]. A recent investigation has described a fully modified form of miR-34a (FM-miR-34a) that effectively addresses issues related to miR-34a stability, non-specific delivery, and delivery-related toxicity [69]. FM-miR-34a demonstrated a potent inhibition of proliferation and invasion, leading to sustained suppression of its target genes for over 120 hours following the in vivo delivery of FM-miR-34a conjugated to folate (FM-FolamiR-34a). This treatment resulted in the inhibition of tumor growth, leading to complete cures in some mice [69]. No significant changes observed in the body weight of mice throughout the study indicating the safety of FM-FolamiR-34. These results have the ability to revitalize miR-34a as an anti-cancer agent, providing a strong rationale for clinical testing.
8. Progress in predicting and validating miRNA targets
As discussed in the literature [85] the mechanistic functions of candidate miRNAs can be evaluated through bioinformatic analysis and/or in vitro experiments before progressing to testing in preclinical animal models. In addition. multiple databases and algorithms have been developed and are available for predicting their targets associated with each miRNA [109,223]. To enhance predictive accuracy of miRNA target prediction, it is a common practice to employ multiple distinct algorithms simultaneously to predict miRNA binding sites in protein-coding genes and relevant biological pathways and networks. An example algorithms is TargetScan [109], which predicts miRNA targets based on seed regions that are essential for mRNA binding. TargetScan encompasses nearly all miRNA sequences documented in miRBase to date.
Likewise, advances in high-throughput screens and bioinformatic tools for target predictions have significantly facilitated the study of miRNAs and prediction of their putative targets and biological pathways. For example, bioinformatics tools such as KEGG and Ingenuity Pathway Analysis not only predict potential biological pathways but, in some instances, also identify disease states that may be influenced by miRNAs.
Furthermore, various computational tools can be used for calculating the free energy between the RNA sequences of interest [224]. For example, a lower free energy, typically around -20 or less, is indicative of a more robust binding [224]. Consequently, the integration of clinical research databases with miRNA bioinformatics platforms could further improve the identification and assessing potential therapeutic candidates.
As for the preclinical models, various human cell lines and induced-pluripotent stem (IPS) cells have been used to investigate the mechanisms, toxicity, and potential therapeutic efficacy of miRNA candidates as well as epigenetic manipulation of target transcripts [85]. For example, use of IPS cells enable modulation of biological pathways along distinct stem cell lineages from readily available skin tissue source [225,226]. Furthermore, the availability of various animal models through academic laboratories or commercially facilitated efficient validation of findings from in vitro miRNA studies. Other animal models including nonhuman primate models have also been successfully used for the preclinical safety and toxicology testing of miRNA therapeutics supporting the initiation of several human miRNA therapeutic clinical trials.
Like other drug classes, the development of miRNA-based drugs must go through a sequence of developmental stages, spanning from discovery to preclinical studies, toxicology assessment, pre-IND, and multiple phases of human clinical trials prior to approval by regulatory agencies before market entry (Figure 5) [142].
In the traditional drug development process, the timeline from identifying a drug target to drug discovery and lead development through preclinical and phase 1-3 human clinical trials, FDA approval, and subsequent Phase 4 studies often spans several years. However, the escalating costs and time required in this process have become unsustainable, urging the imperative to hasten drug discovery and development while curbing associated expenses and timeframes.
Conversely, leveraging RNA-based methodologies, particularly miRNA-based approaches, holds promise in expediting both the discovery and development of drugs, potentially mitigating attrition rates, reducing time constraints, and cutting costs.
9. Progress in preclinical validation of miRNA therapeutics
As discussed in the literature [77,85], single miRNAs not only can regulate an entire pathway and multitude of mRNA targets but also each mRNA may also be targeted by multiple miRNAs [227]. It has been suggested that miRNAs can regulate a diverse range of RNAs stems from their ability to bind to target mRNAs even when the pairing is not perfect. As a result, a single miRNA can regulate a pool of targets involved in similar cellular processes, thereby amplifying the cellular response Although a single miRNA can inhibit hundreds of genes, the effect on each gene is generally mild [116] and multiple miRNAs can regulate the same gene [117,227] thereby amplifying the cellular response.
Because a single miRNA has the potential to bind to as many as 200 target mRNAs, each with diverse functions, including transcription factors, receptors and many others, consequently, entire signaling pathways can be regulated by individual miRNAs [117] or miRNA clusters [228].
The pivotal role of miRNAs in the regulation of multiple genes and their far-reaching effects within regulatory networks presents significant challenges for miRNA therapeutics [85,229]. Therefore, the successful development of miRNA-based therapeutics requires a comprehensive functional characterization and validation of the molecular effects of each miRNA prior to their application as therapeutics [77,85]. Toward the functional characterization of each miRNA, many steps must be undertaken, ranging from the confirmation of the authenticity of an miRNA as a true miRNA, the enhancement of miRNA target prediction algorithms, and the experimental validation of their functional effect on their intended targets in relevant preclinical models [77].
10. Off-target effects of miRNA therapeutics
Among the challenges inherent in miRNA-based therapeutics, off-target effects and associated toxicities emerge due to the capacity of each miRNA to regulate the expression of multiple genes and are one of the main challenges associated with miRNA therapeutics. Thus, further research is essential for the development of miRNAs as effective cancer therapeutics. Additionally, numerous miRNAs are dysregulated not only in cancer cells but also in other cells within the tumor microenvironment (TME), where they might have opposing functions.
Furthermore, depending on the route of administration and the way to enable an intracellular delivery, the miRNA therapeutics are not necessarily restricted to the intended tissue or cells but can also lead to systemic side effects [77]. For example, MRX34, a synthetic miR-34a mimic as a tumor suppressor [230] systemically administered by a liposomal amphoteric (i.e., pH-dependent) delivery, which takes advantage of the low-pH environment of tumorous tissues [231] used for the treatment of various solid tumors and hematologic malignancies (NCT01829971) was terminated prematurely because of severe immune-related side effects leading to the death of four patients [82,83]. Previous animal studies, however, demonstrated that a miR-34a mimic was not only taken up by tumor tissue but also by bone marrow and spleen [232,233] which are involved in the generation and preservation of immune cells. Supporting these preclinical observations, the clinical testing of miR-34a mimic demonstrated a dose-dependent change in several target genes in white blood cells [83]. Consequently, miR-34a mimic not only functions as a tumor suppressor but also impacts the immune cells by modulating calcium or chemokine signaling such as CXCL10/CXCL11/CXCR3-axis in CD4+, CD8+ T cells, and M1 macrophages [234,235].
Despite the lack of the direct causative link between the patient death, the miR-34a function in immune cells has not been yet established. However, the serious and deadly adverse effects of miR-34a mimic MRX34 underscores the need for a priori risk assessment of miRNA therapeutics, specifically their potential off-target effects in other unintended tissues highlighting the need for the development of more precise tissue target delivery systems.
11. Challenges and future perspectives
In addition to their potential utility as biomarkers [68], miRNA mimics, and inhibitors provide a significant potential as therapeutics as many miRNAs function as oncomirs or tumor suppressors, and restoring deregulated miRNA levels to that of healthy tissue levels could potentially help in maintaining the endogenous anti-tumor regulatory mechanisms. While many new immunotherapeutic such as antibodies, recombinant proteins, cell therapies, and small molecules have shown their success in the treatment of various types of cancers [236], because of the global success of mRNA vaccines as modulators of immune stimulation for tackling COVID-19 pandemic, there has been a resurgence of RNA-based cancer immunotherapies [187]. Moreover, further improvements in RNA chemistry and delivery are opening new opportunities for RNA-based immunotherapy. Although, drug development, including RNA-based drugs, often requires many years and substantial costs before approval by the FDA or other regulatory agencies (Figure 5), the global threat of the COVID-19 pandemic has catalyzed the extremely rapid development of a new class of mRNA-based vaccines. These mRNA vaccines whether developed by BioNtech in partnership with Pfizer or Moderna have shown to deliver on their promise i.e., they can be developed extremely fast, can be manufactured under GMP-compliant manufacturing processes, and can be scaled for rapid availability of large numbers of doses, were safe and active at a relatively low dose range [141,187,237,238]. Because of this, many companies are now leveraging the experience gained from the COVID-19 vaccine development to develop RNA–based cancer therapies and potentially for other diseases. This of course have reignited the potential of miRNA as both diagnostics and therapeutics. The approval of several antisense, small interfering RNA (siRNA), and mRNA-based drugs and vaccines have validated their potential and opened the door for expansion into new indications. Research surrounding RNA-based therapies comprises a broad ecosystem, ranging from RNA engineering, and RNA chemistry including various modifications to improve pharmacokinetics and reduce non-specific undesirable side effects to delivery technologies. Furthermore, novel RNA constructs including those self-amplifying RNA, circular RNA, siRNA, as well as gene editing (Cas9 mRNA, single guide RNA [sgRNA]) all hold promise for next-generation cancer immunotherapy.
However, despite this potential over many years, there are several challenges including sensitivity, specificity, toxicity immunogenicity, delivery among many others which are a significant barrier to exploiting the full potential of miRNAs as therapeutics.
In addition, the development of novel targeted delivery systems would be vital for the delivery of miRNAs. The delivery method must be target-specific, and be able to deliver the miRNA drugs to the targeted cells or tissues [142]. As illustrated in Figure 3, the future lies in the targeted delivery vehicles, including lipid and polymer nanoparticles, cell or extracellular vesicle-based packaging, and hybrid systems, as well as viral vectors that will likely increase the therapeutic potency of various RNA-based therapies while decreasing side effects.
In addition, there are other challenges that need to be addressed with regard to the sensitivity, specificity, toxicity, and applicability of potential utility of miRNAs as therapeutics and therapeutic targets. Because each miRNA regulates more than one gene, sometimes a single miRNA can regulate entire cellular pathways via interacting with multiple target genes. Likewise, each mRNA is regulated by more than one miRNA [77]. This phenomenon is referred to as “too many targets for miRNA effect” (TMTME) [129]. Although this characteristics of miRNAs make them a powerful new class of therapeutic, it also represents a major challenge in terms of controlling adverse effects that have been observed in clinical trials [77]. Because of this, Zhang et al. [129] proposed that adverse events observed in terminated clinical trials involving miRNA therapeutics could be attributed to the broad-ranging effects of miRNAs.
In addition, the type of miRNAs might change during the course and stage of cancer which further complicates the target prediction, however, can also be beneficial for assigning a specific miRNA or several miRNAs to a specific stage of cancer. Therefore, more innovative approaches for predicting miRNA targets might be required to validate the predicted targets.
Additional issues that need to be resolved include immunogenic reactions. Although viral delivery systems undoubtedly enhance cellular uptake and expression of miRNAs, they are associated with various side effects, including immunogenicity [198]. To address this issue, there needs to be a better understanding on the prevalence of immunogenic reactions resulting from viral transfer systems.
In addition, further research is needed to investigate whether severe immunogenic reactions are also possible due to modifications in miRNAs, such as LNA miRNAs and artificial miRNAs (amiRNAs), or through miRNA interfering molecules like small cell-permeable molecules, application systems including biodegradable 3D matrices, carriers like functionalized metals, viral transfer systems, or biomolecule conjugates such as aptamers? Additionally, is it feasible to diminish immunogenic reactions by concealing or masking reactive components?
Additionally, as detailed, as summarized in Table 4 and discussed in literature, [77] there are many other key outstanding issues that must be overcome before miRNA therapeutics can become widely adopted as novel therapeutics in the clinic.
These findings emphasize the need for further investigation in developing miRNAs as both novel therapeutics and therapeutic targets for cancer.
12. Conclusions
Because many miRNAs is abnormally expressed or mutated in many cancers acting as oncogenes or a tumor suppressors [99,239,240,241], they have emerged as potential biomarkers [68], therapeutic targets, and therapeutics [81]. However, there are many outstanding issues and challenges with respect to specificity and associated off-targeting effects of miRNA-based therapeutics since each miRNA appears to regulate more than target and each target is regulated more than one miRNA leading to undesired toxicity hence limiting their use as therapeutics.
Currently, most of the miRNA therapeutics are still in early phases of human clinical trials; as such, it is awaited to see how other miRNA therapeutics perform in human clinical trials in terms of toxicity or side effects. Notably, a few recent clinical trials using miRNA therapeutics have reported some serious adverse events. For example, MRX34, a microRNA liposomal injection developed by Mirna Therapeutics, Inc. evaluated in a Phase 1 clinical trial for its efficacy against melanoma was withdrawn (NCT02862145) or terminated (NCT01829971) [82,83,84] due to serious adverse events. As a result, numerous challenges must be addressed to bring therapeutic miRNAs into clinical practice. These include establishing miRNA specificity to their intended targets, reducing immunogenic reactions and adverse events, determining optimal dosing for the desired therapeutic effect while minimizing side effects [77], and developing improved methods for targeted delivery.
Despite these significant challenges, the potential of miRNAs as a therapeutic approach for various diseases is clear. Further research will be necessary to establish whether miRNAs can effectively serve as therapeutics or therapeutic targets for clinical applications.
Abbreviations and acronyms
| AGO | Argonaute |
| AGO2 | Argonaute RISC Catalytic Component 2 |
| ALS | Amyotrophic lateral sclerosis |
| ASO | Antisense oligonucleotides |
| CLL | Chronic Lymphocytic Leukemia |
| CTCL | Cutaneous T-cell Lymphoma |
| ATLL | Adult T-Cell Leukemia/Lymphoma |
| COVID-19 | coronavirus disease 2019 |
| DICER1 | Dicer 1, Ribonuclease III |
| DLBCL | Diffuse Large B-Cell Lymphoma |
| EGFR | Epidermal growth factor receptor |
| DGCR8 | DiGeorge Syndrome Critical Region 8 |
| DROSHA | Drosha Ribonuclease III |
| EVs | Extracellular vesicles |
| GalNAc | N-acetylgalactosamine |
| HCV | Hepatitis C virus |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LNA | Locked nucleic acid (LNA) |
| MF | Mycosis fungoides |
| mRNA | messenger RNA |
| miRNA | microRNA |
| ncRNAs | non-coding RNAs |
| P-bodies | Processing bodies |
| pre-miRNA | precursor microRNA |
| pri-miRNA | primary microRNA |
| RNA Pol II | RNA polymerase II |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| sgRNA | Single guide RNA |
| siRNA | Small interfering RNA |
| TDMD | Target-directed miRNA degradation mechanism |
| TME | Tumor microenvironment |
| TRBP | The TAR RNA-binding protein |
| TTR | Transthyretin |
| UTR | Untranslated region |
| XenomiRs | Exogenous miRNAs |
| XPO5 | Exportin 5 |
Funding
This research received no external funding.
Institutional Review Board Statement
This article does not contain any studies with human participants or animals performed by any of the authors. Informed consent was not required for the preparation of this review article as it used secondary sources only.
Informed Consent Statement
The authors grant the publisher the sole and exclusive license of the full copyright in the Contribution, which license the Publisher hereby accepts.
Data Availability Statement
The data presented in this study are openly available in https://seer.cancer.gov, https://pubmed.ncbi.nlm.nih.gov/, https://clinicaltrials.gov/, and https://www.genomicseducation.hee.nhs.uk/genotes/knowledge-hub/non-coding-dna, all accessed on 26 July 2023.
Acknowledgments
The author thanks Legorreta Cancer Center at Brown University for providing the intellectual and collaborative environment, as well as the necessary resources, to prepare this review article.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Review criteria
Publicly available information such as PubMed and Internet were used for the literature review. We focused on identifying articles published on the use of miRNAs or miRNA-targeting as therapeutics. The research was restricted to the most recent studies in this field and all research was limited to human studies published in English.
References
- Rands, C. M.; Meader, S.; Ponting, C. P.; Lunter, G. 8.2% of the Human genome is constrained: variation in rates of turnover across functional element classes in the human lineage. PLoS Genet 2014, 10, e1004525. [Google Scholar] [CrossRef]
- Dunham, I.; Kundaje, A.; Aldred, S. F.; Collins, P. J.; Davis, C. A.; Doyle, F.; Epstein, C. B.; Frietze, S.; Harrow, J.; Kaul, R.; Khatun, J.; Lajoie, B. R.; Landt, S. G.; Lee, B.-K.; Pauli, F.; Rosenbloom, K. R.; Sabo, P.; Safi, A.; Sanyal, A.; Shoresh, N.; Simon, J. M.; Song, L.; Trinklein, N. D.; Altshuler, R. C.; Birney, E.; Brown, J. B.; Cheng, C.; Djebali, S.; Dong, X.; Dunham, I.; Ernst, J.; Furey, T. S.; Gerstein, M.; Giardine, B.; Greven, M.; Hardison, R. C.; Harris, R. S.; Herrero, J.; Hoffman, M. M.; Iyer, S.; Kellis, M.; Khatun, J.; Kheradpour, P.; Kundaje, A.; Lassmann, T.; Li, Q.; Lin, X.; Marinov, G. K.; Merkel, A.; Mortazavi, A.; Parker, S. C. J.; Reddy, T. E.; Rozowsky, J.; Schlesinger, F.; Thurman, R. E.; Wang, J.; Ward, L. D.; Whitfield, T. W.; Wilder, S. P.; Wu, W.; Xi, H. S.; Yip, K. Y.; Zhuang, J.; Bernstein, B. E.; Birney, E.; Dunham, I.; Green, E. D.; Gunter, C.; Snyder, M.; Pazin, M. J.; Lowdon, R. F.; Dillon, L. A. L.; Adams, L. B.; Kelly, C. J.; Zhang, J.; Wexler, J. R.; Green, E. D.; Good, P. J.; Feingold, E. A.; Bernstein, B. E.; Birney, E.; Crawford, G. E.; Dekker, J.; Elnitski, L.; Farnham, P. J.; Gerstein, M.; Giddings, M. C.; Gingeras, T. R.; Green, E. D.; Guigó, R.; Hardison, R. C.; Hubbard, T. J.; Kellis, M.; Kent, W. J.; Lieb, J. D.; Margulies, E. H.; Myers, R. M.; Snyder, M.; Stamatoyannopoulos, J. A.; Tenenbaum, S. A.; Weng, Z.; White, K. P.; Wold, B.; Khatun, J.; Yu, Y.; Wrobel, J.; Risk, B. A.; Gunawardena, H. P.; Kuiper, H. C.; Maier, C. W.; Xie, L.; Chen, X.; Giddings, M. C.; Bernstein, B. E.; Epstein, C. B.; Shoresh, N.; Ernst, J.; Kheradpour, P.; Mikkelsen, T. S.; Gillespie, S.; Goren, A.; Ram, O.; Zhang, X.; Wang, L.; Issner, R.; Coyne, M. J.; Durham, T.; Ku, M.; Truong, T.; Ward, L. D.; Altshuler, R. C.; Eaton, M. L.; Kellis, M.; Djebali, S.; Davis, C. A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; Xue, C.; Marinov, G. K.; Khatun, J.; Williams, B. A.; Zaleski, C.; Rozowsky, J.; Röder, M.; Kokocinski, F.; Abdelhamid, R. F.; Alioto, T.; Antoshechkin, I.; Baer, M. T.; Batut, P.; Bell, I.; Bell, K.; Chakrabortty, S.; Chen, X.; Chrast, J.; Curado, J.; Derrien, T.; Drenkow, J.; Dumais, E.; Dumais, J.; Duttagupta, R.; Fastuca, M.; Fejes-Toth, K.; Ferreira, P.; Foissac, S.; Fullwood, M. J.; Gao, H.; Gonzalez, D.; Gordon, A.; Gunawardena, H. P.; Howald, C.; Jha, S.; Johnson, R.; Kapranov, P.; King, B.; Kingswood, C.; Li, G.; Luo, O. J.; Park, E.; Preall, J. B.; Presaud, K.; Ribeca, P.; Risk, B. A.; Robyr, D.; Ruan, X.; Sammeth, M.; Sandhu, K. S.; Schaeffer, L.; See, L.-H.; Shahab, A.; Skancke, J.; Suzuki, A. M.; Takahashi, H.; Tilgner, H.; Trout, D.; Walters, N.; Wang, H.; Wrobel, J.; Yu, Y.; Hayashizaki, Y.; Harrow, J.; Gerstein, M.; Hubbard, T. J.; Reymond, A.; Antonarakis, S. E.; Hannon, G. J.; Giddings, M. C.; Ruan, Y.; Wold, B.; Carninci, P.; Guigó, R.; Gingeras, T. R.; Rosenbloom, K. R.; Sloan, C. A.; Learned, K.; Malladi, V. S.; Wong, M. C.; Barber, G. P.; Cline, M. S.; Dreszer, T. R.; Heitner, S. G.; Karolchik, D.; Kent, W. J.; Kirkup, V. M.; Meyer, L. R.; Long, J. C.; Maddren, M.; Raney, B. J.; Furey, T. S.; Song, L.; Grasfeder, L. L.; Giresi, P. G.; Lee, B.-K.; Battenhouse, A.; Sheffield, N. C.; Simon, J. M.; Showers, K. A.; Safi, A.; London, D.; Bhinge, A. A.; Shestak, C.; Schaner, M. R.; Ki Kim, S.; Zhang, Z. Z.; Mieczkowski, P. A.; Mieczkowska, J. O.; Liu, Z.; McDaniell, R. M.; Ni, Y.; Rashid, N. U.; Kim, M. J.; Adar, S.; Zhang, Z.; Wang, T.; Winter, D.; Keefe, D.; Birney, E.; Iyer, V. R.; Lieb, J. D.; Crawford, G. E.; Li, G.; Sandhu, K. S.; Zheng, M.; Wang, P.; Luo, O. J.; Shahab, A.; Fullwood, M. J.; Ruan, X.; Ruan, Y.; Myers, R. M.; Pauli, F.; Williams, B. A.; Gertz, J.; Marinov, G. K.; Reddy, T. E.; Vielmetter, J.; Partridge, E.; Trout, D.; Varley, K. E.; Gasper, C.; The, E. P. C.; Overall, c.; Data production, l.; Lead, a.; Writing, g.; management, N. p.; Principal, i.; Boise State, U. ; University of North Carolina at Chapel Hill Proteomics, g.; Broad Institute, G.; Cold Spring Harbor, U. o. G. C. f. G. R. B. R. S. I. U. o. L. G. I. o. S. g.; Data coordination center at, U. C. S. C.; Duke University, E. B. I. U. o. T. A. U. o. N. C.-C. H. g.; Genome Institute of Singapore, g.; HudsonAlpha Institute, C. U. C. I. S. g. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar]
- Halldorsson, B. V.; Eggertsson, H. P.; Moore, K. H. S.; Hauswedell, H.; Eiriksson, O.; Ulfarsson, M. O.; Palsson, G.; Hardarson, M. T.; Oddsson, A.; Jensson, B. O.; Kristmundsdottir, S.; Sigurpalsdottir, B. D.; Stefansson, O. A.; Beyter, D.; Holley, G.; Tragante, V.; Gylfason, A.; Olason, P. I.; Zink, F.; Asgeirsdottir, M.; Sverrisson, S. T.; Sigurdsson, B.; Gudjonsson, S. A.; Sigurdsson, G. T.; Halldorsson, G. H.; Sveinbjornsson, G.; Norland, K.; Styrkarsdottir, U.; Magnusdottir, D. N.; Snorradottir, S.; Kristinsson, K.; Sobech, E.; Jonsson, H.; Geirsson, A. J.; Olafsson, I.; Jonsson, P.; Pedersen, O. B.; Erikstrup, C.; Brunak, S.; Ostrowski, S. R.; Andersen, S.; Banasik, K.; Burgdorf, K.; Didriksen, M.; Dinh, K. M.; Erikstrup, C.; Gudbjartsson, D.; Hansen, T. F.; Hjalgrim, H.; Jemec, G.; Jennum, P.; Johansson, P. I.; Larsen, M. A. H.; Mikkelsen, S.; Nielsen, K. R.; Nyegaard, M.; Ostrowski, S. R.; Sækmose, S.; Sørensen, E.; Thorsteinsdottir, U.; Brun, M. T.; Ullum, H.; Werge, T.; Thorleifsson, G.; Jonsson, F.; Melsted, P.; Jonsdottir, I.; Rafnar, T.; Holm, H.; Stefansson, H.; Saemundsdottir, J.; Gudbjartsson, D. F.; Magnusson, O. T.; Masson, G.; Thorsteinsdottir, U.; Helgason, A.; Jonsson, H.; Sulem, P.; Stefansson, K.; Consortium, D. G. The sequences of 150,119 genomes in the UK Biobank. Nature 2022, 607, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nature Reviews Molecular Cell Biology 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Arif, M. A.; Seumel, G. I.; Ossowski, S.; Weigel, D.; Reski, R.; Frank, W. Transcriptional control of gene expression by microRNAs. Cell 2010, 140, 111–22. [Google Scholar] [CrossRef] [PubMed]
- Samad, A. F. A.; Sajad, M.; Nazaruddin, N.; Fauzi, I. A.; Murad, A. M. A.; Zainal, Z.; Ismail, I. MicroRNA and Transcription Factor: Key Players in Plant Regulatory Network. Front Plant Sci 2017, 8, 565. [Google Scholar] [CrossRef]
- Tong, Z.; Cui, Q.; Wang, J.; Zhou, Y. TransmiR v2.0: an updated transcription factor-microRNA regulation database. Nucleic Acids Research 2018, 47, D253–D258. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, A. A. microRNAs with different functions and roles in disease development and as potential biomarkers of diabetes: progress and challenges. Mol Biosyst 2015, 11, 1217–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, P. Y.; Meister, G. microRNA-guided posttranscriptional gene regulation. 2005, 386, 1205–1218. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int J Mol Sci 2016, 17, 1712. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front Endocrinol (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, Q.; Qian, J.; Li, M.; Gu, C.; Yang, Y. Review: RNA-based diagnostic markers discovery and therapeutic targets development in cancer. Pharmacol Ther 2022, 234, 108123. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Maki, M.; Ding, R.; Yang, Y.; zhang, B.; Xiong, L. Genome-wide survey of tissue-specific microRNA and transcription factor regulatory networks in 12 tissues. Scientific Reports 2014, 4, 5150. [Google Scholar] [CrossRef] [PubMed]
- Wienholds, E.; Kloosterman, W. P.; Miska, E.; Alvarez-Saavedra, E.; Berezikov, E.; de Bruijn, E.; Horvitz, H. R.; Kauppinen, S.; Plasterk, R. H. MicroRNA expression in zebrafish embryonic development. Science 2005, 309, 310–1. [Google Scholar] [CrossRef]
- Aboobaker, A. A.; Tomancak, P.; Patel, N.; Rubin, G. M.; Lai, E. C. Drosophila microRNAs exhibit diverse spatial expression patterns during embryonic development. Proc Natl Acad Sci U S A 2005, 102, 18017–22. [Google Scholar] [CrossRef] [PubMed]
- Walker, J. C.; Harland, R. M. Expression of microRNAs during embryonic development of Xenopus tropicalis. Gene Expr Patterns 2008, 8, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kowdley, K. V. MicroRNAs in Common Human Diseases. Genomics, Proteomics & Bioinformatics 2012, 10, 246–253. [Google Scholar]
- De Guire, V.; Robitaille, R.; Tetreault, N.; Guerin, R.; Menard, C.; Bambace, N.; Sapieha, P. Circulating miRNAs as sensitive and specific biomarkers for the diagnosis and monitoring of human diseases: promises and challenges. Clin Biochem 2013, 46, 846–60. [Google Scholar] [CrossRef]
- Condrat, C. E.; Thompson, D. C.; Barbu, M. G.; Bugnar, O. L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S. M.; Voinea, S. C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Gareev, I.; de Jesus Encarnacion Ramirez, M.; Goncharov, E.; Ivliev, D.; Shumadalova, A.; Ilyasova, T.; Wang, C. MiRNAs and lncRNAs in the regulation of innate immune signaling. Noncoding RNA Res 2023, 8, 534–541. [Google Scholar] [CrossRef]
- Pauley, K. M.; Cha, S.; Chan, E. K. MicroRNA in autoimmunity and autoimmune diseases. J Autoimmun 2009, 32, 189–94. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C. S.; Ganem, D. MicroRNAs and viral infection. Mol Cell 2005, 20, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Skalsky, R. L.; Cullen, B. R. Viruses, microRNAs, and host interactions. Annu Rev Microbiol 2010, 64, 123–41. [Google Scholar] [CrossRef] [PubMed]
- Barbu, M. G.; Condrat, C. E.; Thompson, D. C.; Bugnar, O. L.; Cretoiu, D.; Toader, O. D.; Suciu, N.; Voinea, S. C. MicroRNA Involvement in Signaling Pathways During Viral Infection. Front Cell Dev Biol 2020, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Abu-Izneid, T.; AlHajri, N.; Ibrahim, A. M.; Javed, M. N.; Salem, K. M.; Pottoo, F. H.; Kamal, M. A. Micro-RNAs in the regulation of immune response against SARS CoV-2 and other viral infections. Journal of Advanced Research 2021, 30, 133–145. [Google Scholar] [CrossRef]
- Elfimova, N.; Schlattjan, M.; Sowa, J. P.; Dienes, H. P.; Canbay, A.; Odenthal, M. Circulating microRNAs: promising candidates serving as novel biomarkers of acute hepatitis. Front Physiol 2012, 3, 476. [Google Scholar] [CrossRef]
- Li, Y. J.; Xu, M.; Gao, Z. H.; Wang, Y. Q.; Yue, Z.; Zhang, Y. X.; Li, X. X.; Zhang, C.; Xie, S. Y.; Wang, P. Y. Alterations of serum levels of BDNF-related miRNAs in patients with depression. PLoS One 2013, 8, e63648. [Google Scholar] [CrossRef]
- Scott, K. A.; Hoban, A. E.; Clarke, G.; Moloney, G. M.; Dinan, T. G.; Cryan, J. F. Thinking small: towards microRNA-based therapeutics for anxiety disorders. Expert Opin Investig Drugs 2015, 24, 529–42. [Google Scholar] [CrossRef]
- Liu, W.; Liu, C.; Zhu, J.; Shu, P.; Yin, B.; Gong, Y.; Qiang, B.; Yuan, J.; Peng, X. MicroRNA-16 targets amyloid precursor protein to potentially modulate Alzheimer’s-associated pathogenesis in SAMP8 mice. Neurobiol Aging 2012, 33, 522–34. [Google Scholar] [CrossRef] [PubMed]
- Weir, D. W.; Sturrock, A.; Leavitt, B. R. Development of biomarkers for Huntington’s disease. Lancet Neurol 2011, 10, 573–90. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, N.; Zhang, Y.; Ran, Y.; Pu, J. Circulating microRNAs are promising novel biomarkers of acute myocardial infarction. Intern Med 2011, 50, 1789–95. [Google Scholar] [CrossRef]
- Recchioni, R.; Marcheselli, F.; Olivieri, F.; Ricci, S.; Procopio, A. D.; Antonicelli, R. Conventional and novel diagnostic biomarkers of acute myocardial infarction: a promising role for circulating microRNAs. Biomarkers 2013, 18, 547–58. [Google Scholar] [CrossRef] [PubMed]
- Nunez Lopez, Y. O.; Coen, P. M.; Goodpaster, B. H.; Seyhan, A. A. Gastric bypass surgery with exercise alters plasma microRNAs that predict improvements in cardiometabolic risk. Int J Obes (Lond) 2017, 41, 1121–1130. [Google Scholar] [CrossRef]
- Nunez Lopez, Y. O.; Garufi, G.; Pasarica, M.; Seyhan, A. A. Elevated and Correlated Expressions of miR-24, miR-30d, miR-146a, and SFRP-4 in Human Abdominal Adipose Tissue Play a Role in Adiposity and Insulin Resistance. Int J Endocrinol 2018, 2018, 7351902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Y.; Minto, A. W.; Wang, J.; Shi, Q.; Li, X.; Quigg, R. J. MicroRNA-377 is up-regulated and can lead to increased fibronectin production in diabetic nephropathy. Faseb j 2008, 22, 4126–35. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, A. A.; Nunez Lopez, Y. O.; Xie, H.; Yi, F.; Mathews, C.; Pasarica, M.; Pratley, R. E. Pancreas-enriched miRNAs are altered in the circulation of subjects with diabetes: a pilot cross-sectional study. Sci Rep 2016, 6, 31479. [Google Scholar] [CrossRef] [PubMed]
- Nunez Lopez, Y. O.; Garufi, G.; Seyhan, A. A. Altered levels of circulating cytokines and microRNAs in lean and obese individuals with prediabetes and type 2 diabetes. Mol Biosyst 2016, 13, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Nunez Lopez, Y. O.; Pittas, A. G.; Pratley, R. E.; Seyhan, A. A. Circulating levels of miR-7, miR-152 and miR-192 respond to vitamin D supplementation in adults with prediabetes and correlate with improvements in glycemic control. J Nutr Biochem 2017, 49, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Kluiver, J.; Poppema, S.; de Jong, D.; Blokzijl, T.; Harms, G.; Jacobs, S.; Kroesen, B. J.; van den Berg, A. BIC and miR-155 are highly expressed in Hodgkin, primary mediastinal and diffuse large B cell lymphomas. J Pathol 2005, 207, 243–9. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E. A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B. L.; Mak, R. H.; Ferrando, A. A.; Downing, J. R.; Jacks, T.; Horvitz, H. R.; Golub, T. R. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–8. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F. J. Oncomirs — microRNAs with a role in cancer. Nature Reviews Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Lawler, S.; Chiocca, E. A. Emerging functions of microRNAs in glioblastoma. J Neurooncol 2009, 92, 297–306. [Google Scholar] [CrossRef]
- Ventura, A.; Jacks, T. MicroRNAs and cancer: short RNAs go a long way. Cell 2009, 136, 586–91. [Google Scholar] [CrossRef]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. Epigenetic alteration and microRNA dysregulation in cancer. Front Genet 2013, 4, 258. [Google Scholar] [CrossRef]
- Di Leva, G.; Garofalo, M.; Croce, C. M. MicroRNAs in cancer. Annu Rev Pathol 2014, 9, 287–314. [Google Scholar] [CrossRef]
- Hayes, J.; Peruzzi, P. P.; Lawler, S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med 2014, 20, 460–9. [Google Scholar] [CrossRef]
- Graveel, C. R.; Calderone, H. M.; Westerhuis, J. J.; Winn, M. E.; Sempere, L. F. Critical analysis of the potential for microRNA biomarkers in breast cancer management. Breast Cancer (Dove Med Press) 2015, 7, 59–79. [Google Scholar]
- Hata, A.; Lieberman, J. Dysregulation of microRNA biogenesis and gene silencing in cancer. Sci Signal 2015, 8, re3. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R. I. MicroRNA biogenesis pathways in cancer. Nat Rev Cancer 2015, 15, 321–33. [Google Scholar] [CrossRef] [PubMed]
- Hata, A.; Kashima, R. Dysregulation of microRNA biogenesis machinery in cancer. Crit Rev Biochem Mol Biol 2016, 51, 121–34. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C. M. The role of MicroRNAs in human cancer. Signal Transduction and Targeted Therapy 2016, 1, 15004. [Google Scholar] [CrossRef]
- Lulla, A. R.; Slifker, M. J.; Zhou, Y.; Lev, A.; Einarson, M. B.; Dicker, D. T.; El-Deiry, W. S. miR-6883 Family miRNAs Target CDK4/6 to Induce G(1) Phase Cell-Cycle Arrest in Colon Cancer Cells. Cancer Res 2017, 77, 6902–6913. [Google Scholar] [CrossRef]
- Biswas, S. MicroRNAs as Therapeutic Agents: The Future of the Battle Against Cancer. Curr Top Med Chem 2018, 18, 2544–2554. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Tan, C.; He, Y.; Zhang, G.; Xu, Y.; Tang, J. Functional miRNAs in breast cancer drug resistance. Onco Targets Ther 2018, 11, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- Plotnikova, O.; Baranova, A.; Skoblov, M. Comprehensive Analysis of Human microRNA-mRNA Interactome. Front Genet 2019, 10, 933. [Google Scholar] [CrossRef]
- Ali Syeda, Z.; Langden, S. S. S.; Munkhzul, C.; Lee, M.; Song, S. J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int J Mol Sci 2020, 21, 1723. [Google Scholar] [CrossRef]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. microRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front Oncol 2020, 10, 581007. [Google Scholar] [CrossRef]
- Cao, D.; Di, M.; Liang, J.; Shi, S.; Tan, Q.; Wang, Z. MicroRNA-183 in Cancer Progression. J Cancer 2020, 11, 1315–1324. [Google Scholar] [CrossRef]
- Fathi, M.; Ghafouri-Fard, S.; Abak, A.; Taheri, M. Emerging roles of miRNAs in the development of pancreatic cancer. Biomedicine & Pharmacotherapy 2021, 141, 111914. [Google Scholar]
- Galka-Marciniak, P.; Urbanek-Trzeciak, M. O.; Nawrocka, P. M.; Kozlowski, P. A pan-cancer atlas of somatic mutations in miRNA biogenesis genes. Nucleic Acids Res 2021, 49, 601–620. [Google Scholar] [CrossRef]
- Inoue, J.; Inazawa, J. Cancer-associated miRNAs and their therapeutic potential. Journal of Human Genetics 2021, 66, 937–945. [Google Scholar] [CrossRef]
- Pajares, M. J.; Alemany-Cosme, E.; Goni, S.; Bandres, E.; Palanca-Ballester, C.; Sandoval, J. Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside. Int J Mol Sci 2021, 22, 7350. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Hogendorf, P. The Role of microRNA in Pancreatic Cancer. Biomedicines 2021, 9, 1322. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. miRNAs in Cancer (Review of Literature). Int J Mol Sci 2022, 23, 2805. [Google Scholar] [CrossRef]
- Menon, A.; Abd-Aziz, N.; Khalid, K.; Poh, C. L.; Naidu, R. miRNA: A Promising Therapeutic Target in Cancer. Int J Mol Sci 2022, 23, 11502. [Google Scholar] [CrossRef] [PubMed]
- Raufi, A. G.; May, M. S.; Hadfield, M. J.; Seyhan, A. A.; El-Deiry, W. S. Advances in Liquid Biopsy Technology and Implications for Pancreatic Cancer. Int J Mol Sci 2023, 24, 4238. [Google Scholar] [CrossRef] [PubMed]
- Ricarte-Filho, J. C.; Casado-Medrano, V.; Reichenberger, E.; Spangler, Z.; Scheerer, M.; Isaza, A.; Baran, J.; Patel, T.; MacFarland, S. P.; Brodeur, G. M.; Stewart, D. R.; Baloch, Z.; Bauer, A. J.; Wasserman, J. D.; Franco, A. T. DICER1 RNase IIIb domain mutations trigger widespread miRNA dysregulation and MAPK activation in pediatric thyroid cancer. Front Endocrinol (Lausanne) 2023, 14, 1083382. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, A. A. Circulating microRNAs as Potential Biomarkers in Pancreatic Cancer-Advances and Challenges. Int J Mol Sci 2023, 24, 13340. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, A. M.; Sohal, I. S.; Iyer, S.; Sudarshan, K.; Kothandaraman, H.; Lanman, N. A.; Low, P. S.; Kasinski, A. L. A first-in-class fully modified version of miR-34a with outstanding stability, activity, and anti-tumor efficacy. Oncogene 2023, 42, 2985–2999. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; Yin, Y.; Wang, C.; Zhang, T.; Zhu, D.; Zhang, D.; Xu, J.; Chen, Q.; Ba, Y.; Liu, J.; Wang, Q.; Chen, J.; Wang, J.; Wang, M.; Zhang, Q.; Zhang, J.; Zen, K.; Zhang, C. Y. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Res 2012, 22, 107–26. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K. W. XenomiRs and miRNA homeostasis in health and disease: evidence that diet and dietary miRNAs directly and indirectly influence circulating miRNA profiles. RNA Biol 2012, 9, 1147–54. [Google Scholar] [CrossRef]
- Wagner, A. E.; Piegholdt, S.; Ferraro, M.; Pallauf, K.; Rimbach, G. Food derived microRNAs. Food Funct 2015, 6, 714–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, T.; Yin, Y.; Zhang, C. Y.; Zhang, Y. L. Dietary microRNA-A Novel Functional Component of Food. Adv Nutr 2019, 10, 711–721. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J. I.; Martínez, J. A.; Milagro, F. I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front Nutr 2021, 8, 586564. [Google Scholar] [CrossRef] [PubMed]
- Cieślik, M.; Bryniarski, K.; Nazimek, K. Dietary and orally-delivered miRNAs: are they functional and ready to modulate immunity? AIMS Allergy and Immunology 2023, 7, 104–131. [Google Scholar] [CrossRef]
- Dickinson, B.; Zhang, Y.; Petrick, J. S.; Heck, G.; Ivashuta, S.; Marshall, W. S. Lack of detectable oral bioavailability of plant microRNAs after feeding in mice. Nat Biotechnol 2013, 31, 965–7. [Google Scholar] [CrossRef] [PubMed]
- Diener, C.; Keller, A.; Meese, E. Emerging concepts of miRNA therapeutics: from cells to clinic. Trends Genet 2022, 38, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Arif, K. M. T.; Elliott, E. K.; Haupt, L. M.; Griffiths, L. R. Regulatory Mechanisms of Epigenetic miRNA Relationships in Human Cancer and Potential as Therapeutic Targets. Cancers (Basel) 2020, 12, 2922. [Google Scholar] [CrossRef] [PubMed]
- Machowska, M.; Galka-Marciniak, P.; Kozlowski, P. Consequences of genetic variants in miRNA genes. Computational and Structural Biotechnology Journal 2022, 20, 6443–6457. [Google Scholar] [CrossRef]
- Bortoletto, A. S.; Parchem, R. J. KRAS Hijacks the miRNA Regulatory Pathway in Cancer. Cancer Research 2023, 83, 1563–1572. [Google Scholar] [CrossRef]
- Li, L. J.; Leng, R. X.; Fan, Y. G.; Pan, H. F.; Ye, D. Q. Translation of noncoding RNAs: Focus on lncRNAs, pri-miRNAs, and circRNAs. Exp Cell Res 2017, 361, 1–8. [Google Scholar] [CrossRef]
- Beg, M. S.; Brenner, A. J.; Sachdev, J.; Borad, M.; Kang, Y. K.; Stoudemire, J.; Smith, S.; Bader, A. G.; Kim, S.; Hong, D. S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Invest New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef]
- Hong, D. S.; Kang, Y. K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H. Y.; Brenner, A. J.; Park, K.; Lee, J. L.; Kim, T. Y.; Shin, S.; Becerra, C. R.; Falchook, G.; Stoudemire, J.; Martin, D.; Kelnar, K.; Peltier, H.; Bonato, V.; Bader, A. G.; Smith, S.; Kim, S.; O’Neill, V.; Beg, M. S. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Desantis, V.; Saltarella, I.; Lamanuzzi, A.; Melaccio, A.; Solimando, A. G.; Mariggiò, M. A.; Racanelli, V.; Paradiso, A.; Vacca, A.; Frassanito, M. A. MicroRNAs-Based Nano-Strategies as New Therapeutic Approach in Multiple Myeloma to Overcome Disease Progression and Drug Resistance. International Journal of Molecular Sciences 2020, 21, 3084. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G. S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front Genet 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Lee, R. C.; Feinbaum, R. L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–54. [Google Scholar] [CrossRef] [PubMed]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–62. [Google Scholar] [CrossRef]
- Pasquinelli, A. E.; Reinhart, B. J.; Slack, F.; Martindale, M. Q.; Kuroda, M. I.; Maller, B.; Hayward, D. C.; Ball, E. E.; Degnan, B.; Müller, P.; Spring, J.; Srinivasan, A.; Fishman, M.; Finnerty, J.; Corbo, J.; Levine, M.; Leahy, P.; Davidson, E.; Ruvkun, G. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Li, S. C.; Chan, W. C.; Hu, L. Y.; Lai, C. H.; Hsu, C. N.; Lin, W. C. Identification of homologous microRNAs in 56 animal genomes. Genomics 2010, 96, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M. R.; Lizano, E.; Houben, A. J. S.; Bezdan, D.; Báñez-Coronel, M.; Kudla, G.; Mateu-Huertas, E.; Kagerbauer, B.; González, J.; Chen, K. C.; LeProust, E. M.; Martí, E.; Estivill, X. Evidence for the biogenesis of more than 1,000 novel human microRNAs. Genome Biology 2014, 15, R57. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S.; Grocock, R. J.; van Dongen, S.; Bateman, A.; Enright, A. J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res 2006, 34, D140–4. [Google Scholar] [CrossRef]
- Pu, M.; Chen, J.; Tao, Z.; Miao, L.; Qi, X.; Wang, Y.; Ren, J. Regulatory network of miRNA on its target: coordination between transcriptional and post-transcriptional regulation of gene expression. Cellular and Molecular Life Sciences 2019, 76, 441–451. [Google Scholar] [CrossRef]
- van Rooij, E. The art of microRNA research. Circ Res 2011, 108, 219–34. [Google Scholar] [CrossRef]
- Friedman, R. C.; Farh, K. K.; Burge, C. B.; Bartel, D. P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 2009, 19, 92–105. [Google Scholar] [CrossRef]
- de Rie, D.; Abugessaisa, I.; Alam, T.; Arner, E.; Arner, P.; Ashoor, H.; Åström, G.; Babina, M.; Bertin, N.; Burroughs, A. M.; Carlisle, A. J.; Daub, C. O.; Detmar, M.; Deviatiiarov, R.; Fort, A.; Gebhard, C.; Goldowitz, D.; Guhl, S.; Ha, T. J.; Harshbarger, J.; Hasegawa, A.; Hashimoto, K.; Herlyn, M.; Heutink, P.; Hitchens, K. J.; Hon, C. C.; Huang, E.; Ishizu, Y.; Kai, C.; Kasukawa, T.; Klinken, P.; Lassmann, T.; Lecellier, C. H.; Lee, W.; Lizio, M.; Makeev, V.; Mathelier, A.; Medvedeva, Y. A.; Mejhert, N.; Mungall, C. J.; Noma, S.; Ohshima, M.; Okada-Hatakeyama, M.; Persson, H.; Rizzu, P.; Roudnicky, F.; Sætrom, P.; Sato, H.; Severin, J.; Shin, J. W.; Swoboda, R. K.; Tarui, H.; Toyoda, H.; Vitting-Seerup, K.; Winteringham, L.; Yamaguchi, Y.; Yasuzawa, K.; Yoneda, M.; Yumoto, N.; Zabierowski, S.; Zhang, P. G.; Wells, C. A.; Summers, K. M.; Kawaji, H.; Sandelin, A.; Rehli, M.; Hayashizaki, Y.; Carninci, P.; Forrest, A. R. R.; de Hoon, M. J. L. An integrated expression atlas of miRNAs and their promoters in human and mouse. Nat Biotechnol 2017, 35, 872–878. [Google Scholar] [CrossRef]
- Gambardella, S.; Rinaldi, F.; Lepore, S. M.; Viola, A.; Loro, E.; Angelini, C.; Vergani, L.; Novelli, G.; Botta, A. Overexpression of microRNA-206 in the skeletal muscle from myotonic dystrophy type 1 patients. J Transl Med 2010, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Baer, C.; Claus, R.; Frenzel, L. P.; Zucknick, M.; Park, Y. J.; Gu, L.; Weichenhan, D.; Fischer, M.; Pallasch, C. P.; Herpel, E.; Rehli, M.; Byrd, J. C.; Wendtner, C. M.; Plass, C. Extensive promoter DNA hypermethylation and hypomethylation is associated with aberrant microRNA expression in chronic lymphocytic leukemia. Cancer Res 2012, 72, 3775–85. [Google Scholar] [CrossRef] [PubMed]
- Achey, R. L.; Khanna, V.; Ostrom, Q. T.; Kruchko, C.; Barnholtz-Sloan, J. S. Incidence and survival trends in oligodendrogliomas and anaplastic oligodendrogliomas in the United States from 2000 to 2013: a CBTRUS Report. J Neurooncol 2017, 133, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Eder, M.; Scherr, M. MicroRNA and lung cancer. N Engl J Med 2005, 352, 2446–8. [Google Scholar] [CrossRef]
- Brennan, C. W.; Verhaak, R. G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S. R.; Zheng, S.; Chakravarty, D.; Sanborn, J. Z.; Berman, S. H.; Beroukhim, R.; Bernard, B.; Wu, C. J.; Genovese, G.; Shmulevich, I.; Barnholtz-Sloan, J.; Zou, L.; Vegesna, R.; Shukla, S. A.; Ciriello, G.; Yung, W. K.; Zhang, W.; Sougnez, C.; Mikkelsen, T.; Aldape, K.; Bigner, D. D.; Van Meir, E. G.; Prados, M.; Sloan, A.; Black, K. L.; Eschbacher, J.; Finocchiaro, G.; Friedman, W.; Andrews, D. W.; Guha, A.; Iacocca, M.; O’Neill, B. P.; Foltz, G.; Myers, J.; Weisenberger, D. J.; Penny, R.; Kucherlapati, R.; Perou, C. M.; Hayes, D. N.; Gibbs, R.; Marra, M.; Mills, G. B.; Lander, E.; Spellman, P.; Wilson, R.; Sander, C.; Weinstein, J.; Meyerson, M.; Gabriel, S.; Laird, P. W.; Haussler, D.; Getz, G.; Chin, L.; Network, T. R. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–77. [Google Scholar] [CrossRef]
- Allen, J. E.; Crowder, R. N.; El-Deiry, W. S. First-In-Class Small Molecule ONC201 Induces DR5 and Cell Death in Tumor but Not Normal Cells to Provide a Wide Therapeutic Index as an Anti-Cancer Agent. PLoS One 2015, 10, e0143082. [Google Scholar] [CrossRef] [PubMed]
- Bedewy, A. M. L.; Elmaghraby, S. M.; Shehata, A. A.; Kandil, N. S. Prognostic Value of miRNA-155 Expression in B-Cell Non-Hodgkin Lymphoma. Turk J Haematol 2017, 34, 207–212. [Google Scholar] [PubMed]
- Kline, C. L. B.; Ralff, M. D.; Lulla, A. R.; Wagner, J. M.; Abbosh, P. H.; Dicker, D. T.; Allen, J. E.; El-Deiry, W. S. Role of Dopamine Receptors in the Anticancer Activity of ONC201. Neoplasia 2018, 20, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Zhao, X.; Zhang, Z.; Xiong, M.; Wang, Y.; Sun, Y.; He, B.; Zhu, J. The diagnostic and prognostic value of the miR-17-92 cluster in hepatocellular carcinoma: A meta-analysis. Front Genet 2022, 13, 927079. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, J. A.; Ventura, A. The biological functions of miRNAs: lessons from in vivo studies. Trends Cell Biol 2015, 25, 137–47. [Google Scholar] [CrossRef] [PubMed]
- Lujambio, A.; Ropero, S.; Ballestar, E.; Fraga, M. F.; Cerrato, C.; Setien, F.; Casado, S.; Suarez-Gauthier, A.; Sanchez-Cespedes, M.; Git, A.; Spiteri, I.; Das, P. P.; Caldas, C.; Miska, E.; Esteller, M. Genetic unmasking of an epigenetically silenced microRNA in human cancer cells. Cancer Res 2007, 67, 1424–9. [Google Scholar] [CrossRef]
- O’Carroll, D.; Schaefer, A. General principals of miRNA biogenesis and regulation in the brain. Neuropsychopharmacology 2013, 38, 39–54. [Google Scholar] [CrossRef]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J. L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res 2004, 14, 1902–10. [Google Scholar] [CrossRef]
- Bartel, D. P. MicroRNAs: target recognition and regulatory functions. Cell 2009, 136, 215–33. [Google Scholar] [CrossRef]
- Iwasaki, S.; Kobayashi, M.; Yoda, M.; Sakaguchi, Y.; Katsuma, S.; Suzuki, T.; Tomari, Y. Hsc70/Hsp90 chaperone machinery mediates ATP-dependent RISC loading of small RNA duplexes. Mol Cell 2010, 39, 292–9. [Google Scholar] [CrossRef]
- Shukla, G. C.; Singh, J.; Barik, S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol Cell Pharmacol 2011, 3, 83–92. [Google Scholar] [PubMed]
- Akgul, B.; Erdogan, I. Intracytoplasmic Re-localization of miRISC Complexes. Front Genet 2018, 9, 403. [Google Scholar] [CrossRef] [PubMed]
- Ghini, F.; Rubolino, C.; Climent, M.; Simeone, I.; Marzi, M. J.; Nicassio, F. Endogenous transcripts control miRNA levels and activity in mammalian cells by target-directed miRNA degradation. Nature Communications 2018, 9, 3119. [Google Scholar] [CrossRef] [PubMed]
- Sheu-Gruttadauria, J.; Pawlica, P.; Klum, S. M.; Wang, S.; Yario, T. A.; Schirle Oakdale, N. T.; Steitz, J. A.; MacRae, I. J. Structural Basis for Target-Directed MicroRNA Degradation. Mol Cell 2019, 75, 1243–1255.e7. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, M.; Croce, C. M. microRNAs: Master regulators as potential therapeutics in cancer. Annu Rev Pharmacol Toxicol 2011, 51, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, S.; Mannsperger, H.; Zhang, J. D.; Horvat, E.; Schmidt, C.; Küblbeck, M.; Henjes, F.; Ward, A.; Tschulena, U.; Zweig, K.; Korf, U.; Wiemann, S.; Sahin, O. Global microRNA level regulation of EGFR-driven cell-cycle protein network in breast cancer. Mol Syst Biol 2012, 8, 570. [Google Scholar] [CrossRef]
- Grimson, A.; Farh, K. K.; Johnston, W. K.; Garrett-Engele, P.; Lim, L. P.; Bartel, D. P. MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell 2007, 27, 91–105. [Google Scholar] [CrossRef]
- Saetrom, P.; Heale, B. S.; Snøve, O. Jr.; Aagaard, L.; Alluin, J.; Rossi, J. J. Distance constraints between microRNA target sites dictate efficacy and cooperativity. Nucleic Acids Res 2007, 35, 2333–42. [Google Scholar] [CrossRef]
- Desterro, J.; Bak-Gordon, P.; Carmo-Fonseca, M. Targeting mRNA processing as an anticancer strategy. Nature Reviews Drug Discovery 2020, 19, 112–129. [Google Scholar] [CrossRef]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V. E.; Zhou, S.; Diaz, L. A., Jr.; Kinzler, K. W. Cancer genome landscapes. Science 2013, 339, 1546–58. [Google Scholar] [CrossRef]
- Fu, Z.; Wang, L.; Li, S.; Chen, F.; Au-Yeung, K. K.; Shi, C. MicroRNA as an Important Target for Anticancer Drug Development. Front Pharmacol 2021, 12, 736323. [Google Scholar] [CrossRef]
- Ding, J.; Cao, Y.; Qi, C.; Zong, Z. Dysregulated microRNAs participate in the crosstalk between colorectal cancer and atrial fibrillation. Hum Cell 2023. [Google Scholar]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: the advantage and disadvantage. Clinical Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y.; Zhao, Y.; Wang, Y.; Ding, H.; Xue, S.; Li, P. Circulating miRNAs as biomarkers for early diagnosis of coronary artery disease. Expert Opin Ther Pat 2018, 28, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A. O.; Landthaler, M.; Lin, C.; Socci, N. D.; Hermida, L.; Fulci, V.; Chiaretti, S.; Foa, R.; Schliwka, J.; Fuchs, U.; Novosel, A.; Muller, R. U.; Schermer, B.; Bissels, U.; Inman, J.; Phan, Q.; Chien, M.; Weir, D. B.; Choksi, R.; De Vita, G.; Frezzetti, D.; Trompeter, H. I.; Hornung, V.; Teng, G.; Hartmann, G.; Palkovits, M.; Di Lauro, R.; Wernet, P.; Macino, G.; Rogler, C. E.; Nagle, J. W.; Ju, J.; Papavasiliou, F. N.; Benzing, T.; Lichter, P.; Tam, W.; Brownstein, M. J.; Bosio, A.; Borkhardt, A.; Russo, J. J.; Sander, C.; Zavolan, M.; Tuschl, T. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–14. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stahler, C.; Meese, E.; Keller, A. Distribution of miRNA expression across human tissues. Nucleic Acids Res 2016, 44, 3865–77. [Google Scholar] [CrossRef]
- Reda El Sayed, S.; Cristante, J.; Guyon, L.; Denis, J.; Chabre, O.; Cherradi, N. MicroRNA Therapeutics in Cancer: Current Advances and Challenges. Cancers (Basel) 2021, 13, 2680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cheng, Z.; Wang, Y.; Han, T. The Risks of miRNA Therapeutics: In a Drug Target Perspective. Drug Des Devel Ther 2021, 15, 721–733. [Google Scholar] [CrossRef]
- Mullard, A. 2018 FDA drug approvals. Nat Rev Drug Discov 2019, 18, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. 2019 FDA drug approvals. Nat Rev Drug Discov 2020, 19, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Urits, I.; Swanson, D.; Swett, M. C.; Patel, A.; Berardino, K.; Amgalan, A.; Berger, A. A.; Kassem, H.; Kaye, A. D.; Viswanath, O. A Review of Patisiran (ONPATTRO®) for the Treatment of Polyneuropathy in People with Hereditary Transthyretin Amyloidosis. Neurol Ther 2020, 9, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Conceição, I.; González-Duarte, A.; Obici, L.; Schmidt, H. H.; Simoneau, D.; Ong, M. L.; Amass, L. “Red-flag” symptom clusters in transthyretin familial amyloid polyneuropathy. J Peripher Nerv Syst 2016, 21, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Vormehr, M.; Türeci, Ö.; Sahin, U. Harnessing Tumor Mutations for Truly Individualized Cancer Vaccines. Annu Rev Med 2019, 70, 395–407. [Google Scholar] [CrossRef]
- Rojas, L. A.; Sethna, Z.; Soares, K. C.; Olcese, C.; Pang, N.; Patterson, E.; Lihm, J.; Ceglia, N.; Guasp, P.; Chu, A.; Yu, R.; Chandra, A. K.; Waters, T.; Ruan, J.; Amisaki, M.; Zebboudj, A.; Odgerel, Z.; Payne, G.; Derhovanessian, E.; Müller, F.; Rhee, I.; Yadav, M.; Dobrin, A.; Sadelain, M.; Łuksza, M.; Cohen, N.; Tang, L.; Basturk, O.; Gönen, M.; Katz, S.; Do, R. K.; Epstein, A. S.; Momtaz, P.; Park, W.; Sugarman, R.; Varghese, A. M.; Won, E.; Desai, A.; Wei, A. C.; D’Angelica, M. I.; Kingham, T. P.; Mellman, I.; Merghoub, T.; Wolchok, J. D.; Sahin, U.; Türeci, Ö.; Greenbaum, B. D.; Jarnagin, W. R.; Drebin, J.; O’Reilly, E. M.; Balachandran, V. P. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature 2023, 618, 144–150. [Google Scholar] [CrossRef]
- Sahin, U.; Derhovanessian, E.; Miller, M.; Kloke, B. P.; Simon, P.; Löwer, M.; Bukur, V.; Tadmor, A. D.; Luxemburger, U.; Schrörs, B.; Omokoko, T.; Vormehr, M.; Albrecht, C.; Paruzynski, A.; Kuhn, A. N.; Buck, J.; Heesch, S.; Schreeb, K. H.; Müller, F.; Ortseifer, I.; Vogler, I.; Godehardt, E.; Attig, S.; Rae, R.; Breitkreuz, A.; Tolliver, C.; Suchan, M.; Martic, G.; Hohberger, A.; Sorn, P.; Diekmann, J.; Ciesla, J.; Waksmann, O.; Brück, A. K.; Witt, M.; Zillgen, M.; Rothermel, A.; Kasemann, B.; Langer, D.; Bolte, S.; Diken, M.; Kreiter, S.; Nemecek, R.; Gebhardt, C.; Grabbe, S.; Höller, C.; Utikal, J.; Huber, C.; Loquai, C.; Türeci, Ö. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature 2017, 547, 222–226. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–9. [Google Scholar] [CrossRef]
- Diebold, S. S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–31. [Google Scholar] [CrossRef]
- Lund, O.; Nielsen, M.; Kesmir, C.; Petersen, A. G.; Lundegaard, C.; Worning, P.; Sylvester-Hvid, C.; Lamberth, K.; Røder, G.; Justesen, S.; Buus, S.; Brunak, S. Definition of supertypes for HLA molecules using clustering of specificity matrices. Immunogenetics 2004, 55, 797–810. [Google Scholar] [CrossRef]
- Kranz, L. M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K. C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; Grunwitz, C.; Vormehr, M.; Hüsemann, Y.; Selmi, A.; Kuhn, A. N.; Buck, J.; Derhovanessian, E.; Rae, R.; Attig, S.; Diekmann, J.; Jabulowsky, R. A.; Heesch, S.; Hassel, J.; Langguth, P.; Grabbe, S.; Huber, C.; Türeci, Ö.; Sahin, U. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016, 534, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Polack, F. P.; Thomas, S. J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J. L.; Pérez Marc, G.; Moreira, E. D.; Zerbini, C.; Bailey, R.; Swanson, K. A.; Roychoudhury, S.; Koury, K.; Li, P.; Kalina, W. V.; Cooper, D.; Frenck, R. W.; Hammitt, L. L.; Türeci, Ö.; Nell, H.; Schaefer, A.; Ünal, S.; Tresnan, D. B.; Mather, S.; Dormitzer, P. R.; Şahin, U.; Jansen, K. U.; Gruber, W. C. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New England Journal of Medicine 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A. R.; Sharma, G.; Lee, S.-S. Therapeutic advances of miRNAs: A preclinical and clinical update. Journal of Advanced Research 2021, 28, 127–138. [Google Scholar] [CrossRef]
- Dhuri, K.; Bechtold, C.; Quijano, E.; Pham, H.; Gupta, A.; Vikram, A.; Bahal, R. Antisense Oligonucleotides: An Emerging Area in Drug Discovery and Development. J Clin Med 2020, 9, 2004. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L. R.; Frampton, A. E.; Jacob, J.; Pellegrino, L.; Krell, J.; Giamas, G.; Tsim, N.; Vlavianos, P.; Cohen, P.; Ahmad, R.; Keller, A.; Habib, N. A.; Stebbing, J.; Castellano, L. MicroRNAs targeting oncogenes are down-regulated in pancreatic malignant transformation from benign tumors. PLoS One 2012, 7, e32068. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, G. miRNAs: The Road from Bench to Bedside. Genes (Basel) 2023, 14, 314. [Google Scholar] [CrossRef] [PubMed]
- Raue, R.; Frank, A. C.; Syed, S. N.; Brune, B. Therapeutic Targeting of MicroRNAs in the Tumor Microenvironment. Int J Mol Sci 2021, 22, 2210. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Pan, M. H.; Wang, L.; Li, W.; Jiang, C.; He, J.; Abouzid, K.; Liu, L. Z.; Shi, Z.; Jiang, B. H. Hypoxia-mediated mitochondria apoptosis inhibition induces temozolomide treatment resistance through miR-26a/Bad/Bax axis. Cell Death Dis 2018, 9, 1128. [Google Scholar] [CrossRef]
- Gee, H. E.; Ivan, C.; Calin, G. A.; Ivan, M. HypoxamiRs and cancer: from biology to targeted therapy. Antioxid Redox Signal 2014, 21, 1220–38. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Xia, X.; Wang, X.; Zhang, K.; Cao, J.; Jiang, T.; Zhao, Q.; Qiu, Z. miR-301a plays a pivotal role in hypoxia-induced gemcitabine resistance in pancreatic cancer. Exp Cell Res 2018, 369, 120–128. [Google Scholar] [CrossRef]
- Zhang, J.; Jin, H.; Liu, H.; Lv, S.; Wang, B.; Wang, R.; Liu, H.; Ding, M.; Yang, Y.; Li, L.; Zhang, J.; Fu, S.; Xie, D.; Wu, M.; Zhou, W.; Qian, Q. MiRNA-99a directly regulates AGO2 through translational repression in hepatocellular carcinoma. Oncogenesis 2014, 3, e97. [Google Scholar] [CrossRef]
- Xin, X.; Kumar, V.; Lin, F.; Kumar, V.; Bhattarai, R.; Bhatt, V. R.; Tan, C.; Mahato, R. I. Redox-responsive nanoplatform for codelivery of miR-519c and gemcitabine for pancreatic cancer therapy. Sci Adv 2020, 6, (46). [Google Scholar] [CrossRef]
- Gebert, L. F.; Rebhan, M. A.; Crivelli, S. E.; Denzler, R.; Stoffel, M.; Hall, J. Miravirsen (SPC3649) can inhibit the biogenesis of miR-122. Nucleic Acids Res 2014, 42, 609–21. [Google Scholar] [CrossRef] [PubMed]
- Huang, P. S.; Liao, C. J.; Huang, Y. H.; Yeh, C. T.; Chen, C. Y.; Tang, H. C.; Chang, C. C.; Lin, K. H. Functional and Clinical Significance of Dysregulated microRNAs in Liver Cancer. Cancers (Basel), 2021; 13, (21). [Google Scholar]
- Winkler, J.; Stessl, M.; Amartey, J.; Noe, C. R. Off-target effects related to the phosphorothioate modification of nucleic acids. ChemMedChem 2010, 5, 1344–52. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, G.; Siani, A. Role of microRNAs in obesity and obesity-related diseases. Genes Nutr 2017, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- van der Ree, M. H.; de Vree, J. M.; Stelma, F.; Willemse, S.; van der Valk, M.; Rietdijk, S.; Molenkamp, R.; Schinkel, J.; van Nuenen, A. C.; Beuers, U.; Hadi, S.; Harbers, M.; van der Veer, E.; Liu, K.; Grundy, J.; Patick, A. K.; Pavlicek, A.; Blem, J.; Huang, M.; Grint, P.; Neben, S.; Gibson, N. W.; Kootstra, N. A.; Reesink, H. W. Safety, tolerability, and antiviral effect of RG-101 in patients with chronic hepatitis C: a phase 1B, double-blind, randomised controlled trial. Lancet 2017, 389, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Seto, A. G.; Beatty, X.; Lynch, J. M.; Hermreck, M.; Tetzlaff, M.; Duvic, M.; Jackson, A. L. Cobomarsen, an oligonucleotide inhibitor of miR-155, co-ordinately regulates multiple survival pathways to reduce cellular proliferation and survival in cutaneous T-cell lymphoma. Br J Haematol 2018, 183, 428–444. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadou, E.; Seto, A. G.; Beatty, X.; Hermreck, M.; Gilles, M. E.; Stroopinsky, D.; Pinter-Brown, L. C.; Pestano, L.; Marchese, C.; Avigan, D.; Trivedi, P.; Escolar, D. M.; Jackson, A. L.; Slack, F. J. Cobomarsen, an Oligonucleotide Inhibitor of miR-155, Slows DLBCL Tumor Cell Growth In Vitro and In Vivo. Clin Cancer Res 2021, 27, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Gallant-Behm, C. L.; Piper, J.; Dickinson, B. A.; Dalby, C. M.; Pestano, L. A.; Jackson, A. L. A synthetic microRNA-92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen 2018, 26, 311–323. [Google Scholar] [CrossRef]
- Gallant-Behm, C. L.; Piper, J.; Lynch, J. M.; Seto, A. G.; Hong, S. J.; Mustoe, T. A.; Maari, C.; Pestano, L. A.; Dalby, C. M.; Jackson, A. L.; Rubin, P.; Marshall, W. S. A MicroRNA-29 Mimic (Remlarsen) Represses Extracellular Matrix Expression and Fibroplasia in the Skin. J Invest Dermatol 2019, 139, 1073–1081. [Google Scholar] [CrossRef]
- Kriegel, A. J.; Liu, Y.; Cohen, B.; Usa, K.; Liu, Y.; Liang, M. MiR-382 targeting of kallikrein 5 contributes to renal inner medullary interstitial fibrosis. Physiol Genomics 2012, 44, 259–67. [Google Scholar] [CrossRef]
- van Zandwijk, N.; Pavlakis, N.; Kao, S. C.; Linton, A.; Boyer, M. J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M. J.; Bailey, D. L.; Cooper, W. A.; Kritharides, L.; Ridley, L.; Pattison, S. T.; MacDiarmid, J.; Brahmbhatt, H.; Reid, G. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: a first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Bais, T.; Gansevoort, R. T.; Meijer, E. Drugs in Clinical Development to Treat Autosomal Dominant Polycystic Kidney Disease. Drugs 2022, 82, 1095–1115. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A. R.; Sharma, G.; Sarkar, B. K.; Lee, S. S. The novel strategies for next-generation cancer treatment: miRNA combined with chemotherapeutic agents for the treatment of cancer. Oncotarget 2018, 9, 10164–10174. [Google Scholar] [CrossRef] [PubMed]
- Yu, H. W.; Cho, W. C. The emerging role of miRNAs in combined cancer therapy. Expert Opin Biol Ther 2015, 15, 923–5. [Google Scholar] [CrossRef] [PubMed]
- Vernon, M.; Lambert, B.; Meryet-Figuière, M.; Brotin, E.; Weiswald, L. B.; Paysant, H.; Vigneron, N.; Wambecke, A.; Abeilard, E.; Giffard, F.; Louis, M. H.; Blanc-Fournier, C.; Gauduchon, P.; Poulain, L.; Denoyelle, C. Functional miRNA Screening Identifies Wide-ranging Antitumor Properties of miR-3622b-5p and Reveals a New Therapeutic Combination Strategy in Ovarian Tumor Organoids. Mol Cancer Ther 2020, 19, 1506–1519. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; Wang, X. miRNA-based biomarkers, therapies, and resistance in Cancer. Int J Biol Sci 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Wu, P. Inhibition of RNA-binding proteins with small molecules. Nat Rev Chem 2020, 4, 441–458. [Google Scholar] [CrossRef]
- Galm, O.; Herman, J. G.; Baylin, S. B. The fundamental role of epigenetics in hematopoietic malignancies. Blood Rev 2006, 20, 1–13. [Google Scholar] [CrossRef]
- Melo, S.; Villanueva, A.; Moutinho, C.; Davalos, V.; Spizzo, R.; Ivan, C.; Rossi, S.; Setien, F.; Casanovas, O.; Simo-Riudalbas, L.; Carmona, J.; Carrere, J.; Vidal, A.; Aytes, A.; Puertas, S.; Ropero, S.; Kalluri, R.; Croce, C. M.; Calin, G. A.; Esteller, M. Small molecule enoxacin is a cancer-specific growth inhibitor that acts by enhancing TAR RNA-binding protein 2-mediated microRNA processing. Proc Natl Acad Sci U S A 2011, 108, 4394–9. [Google Scholar] [CrossRef]
- Roldo, C.; Missiaglia, E.; Hagan, J. P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G. A.; Volinia, S.; Liu, C. G.; Scarpa, A.; Croce, C. M. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J Clin Oncol 2006, 24, 4677–84. [Google Scholar] [CrossRef]
- Tetzlaff, M. T.; Liu, A.; Xu, X.; Master, S. R.; Baldwin, D. A.; Tobias, J. W.; Livolsi, V. A.; Baloch, Z. W. Differential expression of miRNAs in papillary thyroid carcinoma compared to multinodular goiter using formalin fixed paraffin embedded tissues. Endocr Pathol 2007, 18, 163–73. [Google Scholar] [CrossRef]
- Asangani, I. A.; Rasheed, S. A.; Nikolova, D. A.; Leupold, J. H.; Colburn, N. H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008, 27, 2128–36. [Google Scholar] [CrossRef]
- Pan, X.; Wang, Z. X.; Wang, R. MicroRNA-21: a novel therapeutic target in human cancer. Cancer Biol Ther 2010, 10, 1224–32. [Google Scholar] [CrossRef]
- Watashi, K.; Yeung, M. L.; Starost, M. F.; Hosmane, R. S.; Jeang, K. T. Identification of small molecules that suppress microRNA function and reverse tumorigenesis. J Biol Chem 2010, 285, 24707–16. [Google Scholar] [CrossRef]
- Davies, B. P.; Arenz, C. A homogenous assay for micro RNA maturation. Angew Chem Int Ed Engl 2006, 45, 5550–2. [Google Scholar] [CrossRef] [PubMed]
- Young, D. D.; Connelly, C. M.; Grohmann, C.; Deiters, A. Small molecule modifiers of microRNA miR-122 function for the treatment of hepatitis C virus infection and hepatocellular carcinoma. J Am Chem Soc 2010, 132, 7976–81. [Google Scholar] [CrossRef]
- Girard, M.; Jacquemin, E.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. miR-122, a paradigm for the role of microRNAs in the liver. J Hepatol 2008, 48, 648–56. [Google Scholar] [CrossRef]
- Esau, C.; Davis, S.; Murray, S. F.; Yu, X. X.; Pandey, S. K.; Pear, M.; Watts, L.; Booten, S. L.; Graham, M.; McKay, R.; Subramaniam, A.; Propp, S.; Lollo, B. A.; Freier, S.; Bennett, C. F.; Bhanot, S.; Monia, B. P. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab 2006, 3, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C. L.; Yi, M.; Lancaster, A. M.; Lemon, S. M.; Sarnow, P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science 2005, 309, 1577–81. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Catalucci, D.; Bauersachs, J. MicroRNAs: novel regulators in cardiac development and disease. Cardiovasc Res 2008, 79, 562–70. [Google Scholar] [CrossRef]
- Wystub, K.; Besser, J.; Bachmann, A.; Boettger, T.; Braun, T. miR-1/133a clusters cooperatively specify the cardiomyogenic lineage by adjustment of myocardin levels during embryonic heart development. PLoS Genet 2013, 9, e1003793. [Google Scholar] [CrossRef] [PubMed]
- Tan, S. B.; Huang, C.; Chen, X.; Wu, Y.; Zhou, M.; Zhang, C.; Zhang, Y. Small molecular inhibitors of miR-1 identified from photocycloadducts of acetylenes with 2-methoxy-1,4-naphthalenequinone. Bioorg Med Chem 2013, 21, 6124–31. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, D. Y.; Huang, L. In vivo delivery of miRNAs for cancer therapy: challenges and strategies. Adv Drug Deliv Rev 2015, 81, 128–41. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, E. S.; Lu, C.; Wang, H.; Luber, B.; Nakazawa, M.; Roeser, J. C.; Chen, Y.; Mohammad, T. A.; Chen, Y.; Fedor, H. L.; Lotan, T. L.; Zheng, Q.; De Marzo, A. M.; Isaacs, J. T.; Isaacs, W. B.; Nadal, R.; Paller, C. J.; Denmeade, S. R.; Carducci, M. A.; Eisenberger, M. A.; Luo, J. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med 2014, 371, 1028–38. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Chen, J.; Huang, Z. Recent progress in microRNA-based delivery systems for the treatment of human disease. ExRNA 2019, 1, 24. [Google Scholar] [CrossRef]
- Beck, J. D.; Reidenbach, D.; Salomon, N.; Sahin, U.; Türeci, Ö.; Vormehr, M.; Kranz, L. M. mRNA therapeutics in cancer immunotherapy. Mol Cancer 2021, 20, 69. [Google Scholar] [CrossRef] [PubMed]
- Jivrajani, M.; Nivsarkar, M. Ligand-targeted bacterial minicells: Futuristic nano-sized drug delivery system for the efficient and cost effective delivery of shRNA to cancer cells. Nanomedicine 2016, 12, 2485–2498. [Google Scholar] [CrossRef]
- Dasgupta, I.; Chatterjee, A. Recent Advances in miRNA Delivery Systems. Methods Protoc 2021, 4, 10. [Google Scholar] [CrossRef]
- Roberts, T. C.; Langer, R.; Wood, M. J. A. Advances in oligonucleotide drug delivery. Nat Rev Drug Discov 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Dosta, P.; Cryer, A. M.; Dion, M. Z.; Shiraishi, T.; Langston, S. P.; Lok, D.; Wang, J.; Harrison, S.; Hatten, T.; Ganno, M. L.; Appleman, V. A.; Taboada, G. M.; Puigmal, N.; Ferber, S.; Kalash, S.; Prado, M.; Rodríguez, A. L.; Kamoun, W. S.; Abu-Yousif, A. O.; Artzi, N. Investigation of the enhanced antitumour potency of STING agonist after conjugation to polymer nanoparticles. Nature Nanotechnology 2023, 18, 1351–1363. [Google Scholar] [CrossRef]
- Schultz, B. R.; Chamberlain, J. S. Recombinant adeno-associated virus transduction and integration. Mol Ther 2008, 16, 1189–99. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Carrillo, E.; Liu, Y. P.; Berkhout, B. Improving miRNA Delivery by Optimizing miRNA Expression Cassettes in Diverse Virus Vectors. Hum Gene Ther Methods 2017, 28, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Mascellino, M. T.; Di Timoteo, F.; De Angelis, M.; Oliva, A. Overview of the Main Anti-SARS-CoV-2 Vaccines: Mechanism of Action, Efficacy and Safety. Infect Drug Resist 2021, 14, 3459–3476. [Google Scholar] [CrossRef] [PubMed]
- Keskin, S.; Brouwers, C. C.; Sogorb-Gonzalez, M.; Martier, R.; Depla, J. A.; Vallès, A.; van Deventer, S. J.; Konstantinova, P.; Evers, M. M. AAV5-miHTT Lowers Huntingtin mRNA and Protein without Off-Target Effects in Patient-Derived Neuronal Cultures and Astrocytes. Mol Ther Methods Clin Dev 2019, 15, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Miniarikova, J.; Zanella, I.; Huseinovic, A.; van der Zon, T.; Hanemaaijer, E.; Martier, R.; Koornneef, A.; Southwell, A. L.; Hayden, M. R.; van Deventer, S. J.; Petry, H.; Konstantinova, P. Design, Characterization, and Lead Selection of Therapeutic miRNAs Targeting Huntingtin for Development of Gene Therapy for Huntington’s Disease. Mol Ther Nucleic Acids 2016, 5, e297. [Google Scholar] [CrossRef]
- Samaranch, L.; Blits, B.; San Sebastian, W.; Hadaczek, P.; Bringas, J.; Sudhakar, V.; Macayan, M.; Pivirotto, P. J.; Petry, H.; Bankiewicz, K. S. MR-guided parenchymal delivery of adeno-associated viral vector serotype 5 in non-human primate brain. Gene Ther 2017, 24, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Monahan, P. E.; Négrier, C.; Tarantino, M.; Valentino, L. A.; Mingozzi, F. Emerging Immunogenicity and Genotoxicity Considerations of Adeno-Associated Virus Vector Gene Therapy for Hemophilia. J Clin Med 2021, 10, 2471. [Google Scholar] [CrossRef]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat Biotechnol 2015, 33, 941–51. [Google Scholar] [CrossRef]
- Reid, G.; Kao, S. C.; Pavlakis, N.; Brahmbhatt, H.; MacDiarmid, J.; Clarke, S.; Boyer, M.; van Zandwijk, N. Clinical development of TargomiRs, a miRNA mimic-based treatment for patients with recurrent thoracic cancer. Epigenomics 2016, 8, 1079–85. [Google Scholar] [CrossRef]
- Elsharkasy, O. M.; Nordin, J. Z.; Hagey, D. W.; de Jong, O. G.; Schiffelers, R. M.; Andaloussi, S. E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how? Adv Drug Deliv Rev 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Baldari, S.; Di Rocco, G.; Magenta, A.; Picozza, M.; Toietta, G. Extracellular Vesicles-Encapsulated MicroRNA-125b Produced in Genetically Modified Mesenchymal Stromal Cells Inhibits Hepatocellular Carcinoma Cell Proliferation. Cells 2019, 8, 1560. [Google Scholar] [CrossRef]
- Zhou, J.; Rossi, J. Aptamers as targeted therapeutics: current potential and challenges. Nat Rev Drug Discov 2017, 16, 181–202. [Google Scholar] [CrossRef]
- Russo, V.; Paciocco, A.; Affinito, A.; Roscigno, G.; Fiore, D.; Palma, F.; Galasso, M.; Volinia, S.; Fiorelli, A.; Esposito, C. L.; Nuzzo, S.; Inghirami, G.; de Franciscis, V.; Condorelli, G. Aptamer-miR-34c Conjugate Affects Cell Proliferation of Non-Small-Cell Lung Cancer Cells. Mol Ther Nucleic Acids 2018, 13, 334–346. [Google Scholar] [CrossRef]
- Prakash, T. P.; Graham, M. J.; Yu, J.; Carty, R.; Low, A.; Chappell, A.; Schmidt, K.; Zhao, C.; Aghajan, M.; Murray, H. F.; Riney, S.; Booten, S. L.; Murray, S. F.; Gaus, H.; Crosby, J.; Lima, W. F.; Guo, S.; Monia, B. P.; Swayze, E. E.; Seth, P. P. Targeted delivery of antisense oligonucleotides to hepatocytes using triantennary N-acetyl galactosamine improves potency 10-fold in mice. Nucleic Acids Res 2014, 42, 8796–807. [Google Scholar] [CrossRef]
- Biessen, E. A.; Vietsch, H.; Rump, E. T.; Fluiter, K.; Kuiper, J.; Bijsterbosch, M. K.; van Berkel, T. J. Targeted delivery of oligodeoxynucleotides to parenchymal liver cells in vivo. Biochem J 1999, 340 (Pt 3) Pt 3, 783–92. [Google Scholar] [CrossRef]
- Drenth, J. P. H.; Schattenberg, J. M. The nonalcoholic steatohepatitis (NASH) drug development graveyard: established hurdles and planning for future success. Expert Opin Investig Drugs 2020, 29, 1365–1375. [Google Scholar] [CrossRef]
- Huang, Y. Preclinical and Clinical Advances of GalNAc-Decorated Nucleic Acid Therapeutics. Mol Ther Nucleic Acids 2017, 6, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Kelnar, K.; Peltier, H. J.; Leatherbury, N.; Stoudemire, J.; Bader, A. G. Quantification of therapeutic miRNA mimics in whole blood from nonhuman primates. Anal Chem 2014, 86, 1534–42. [Google Scholar] [CrossRef] [PubMed]
- Stelma, F.; van der Ree, M. H.; Sinnige, M. J.; Brown, A.; Swadling, L.; de Vree, J. M. L.; Willemse, S. B.; van der Valk, M.; Grint, P.; Neben, S.; Klenerman, P.; Barnes, E.; Kootstra, N. A.; Reesink, H. W. Immune phenotype and function of natural killer and T cells in chronic hepatitis C patients who received a single dose of anti-MicroRNA-122, RG-101. Hepatology 2017, 66, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Mukai, Y.; Wada, F.; Terada, C.; Kayaba, Y.; Oh, K.; Yamayoshi, A.; Obika, S.; Harada-Shiba, M. Highly Potent GalNAc-Conjugated Tiny LNA Anti-miRNA-122 Antisense Oligonucleotides. Pharmaceutics 2021, 13, 817. [Google Scholar] [CrossRef] [PubMed]
- Kardani, A.; Yaghoobi, H.; Alibakhshi, A.; Khatami, M. Inhibition of miR-155 in MCF-7 breast cancer cell line by gold nanoparticles functionalized with antagomir and AS1411 aptamer. J Cell Physiol 2020, 235, 6887–6895. [Google Scholar] [CrossRef]
- Nikolova, M. P.; Chavali, M. S. Recent advances in biomaterials for 3D scaffolds: A review. Bioact Mater 2019, 4, 271–292. [Google Scholar] [CrossRef]
- Aguilar-de-Leyva, Á.; Linares, V.; Casas, M.; Caraballo, I. 3D Printed Drug Delivery Systems Based on Natural Products. Pharmaceutics 2020, 12, 620. [Google Scholar] [CrossRef]
- Shende, P.; Trivedi, R. 3D Printed Bioconstructs: Regenerative Modulation for Genetic Expression. Stem Cell Rev Rep 2021, 17, 1239–1250. [Google Scholar] [CrossRef]
- Lennox, K. A.; Behlke, M. A. Chemical modification and design of anti-miRNA oligonucleotides. Gene Ther 2011, 18, 1111–20. [Google Scholar] [CrossRef]
- Grünweller, A.; Hartmann, R. K. Locked nucleic acid oligonucleotides: the next generation of antisense agents? BioDrugs 2007, 21, 235–43. [Google Scholar] [CrossRef] [PubMed]
- Koshkin, A. A.; Singh, S. K.; Nielsen, P.; Rajwanshi, V. K.; Kumar, R.; Meldgaard, M.; Olsen, C. E.; Wengel, J. LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition. Tetrahedron 1998, 54, 3607–3630. [Google Scholar] [CrossRef]
- Deprey, K.; Batistatou, N.; Kritzer, J. A. A critical analysis of methods used to investigate the cellular uptake and subcellular localization of RNA therapeutics. Nucleic Acids Res 2020, 48, 7623–7639. [Google Scholar] [CrossRef] [PubMed]
- Miller, C. M.; Tanowitz, M.; Donner, A. J.; Prakash, T. P.; Swayze, E. E.; Harris, E. N.; Seth, P. P. Receptor-Mediated Uptake of Phosphorothioate Antisense Oligonucleotides in Different Cell Types of the Liver. Nucleic Acid Ther 2018, 28, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, J.; Hanss, B.; Kopp, J. B.; Copeland, T. D.; Bruggeman, L. A.; Coffman, T. M.; Klotman, P. E. Transport of phosphorothioate oligonucleotides in kidney: implications for molecular therapy. Kidney Int 1995, 47, 1462–9. [Google Scholar] [CrossRef] [PubMed]
- Gomez, I. G.; MacKenna, D. A.; Johnson, B. G.; Kaimal, V.; Roach, A. M.; Ren, S.; Nakagawa, N.; Xin, C.; Newitt, R.; Pandya, S.; Xia, T. H.; Liu, X.; Borza, D. B.; Grafals, M.; Shankland, S. J.; Himmelfarb, J.; Portilla, D.; Liu, S.; Chau, B. N.; Duffield, J. S. Anti-microRNA-21 oligonucleotides prevent Alport nephropathy progression by stimulating metabolic pathways. J Clin Invest 2015, 125, 141–56. [Google Scholar] [CrossRef] [PubMed]
- Washietl, S.; Will, S.; Hendrix, D. A.; Goff, L. A.; Rinn, J. L.; Berger, B.; Kellis, M. Computational analysis of noncoding RNAs. Wiley Interdiscip Rev RNA 2012, 3, 759–78. [Google Scholar] [CrossRef]
- Lekprasert, P.; Mayhew, M.; Ohler, U. Assessing the utility of thermodynamic features for microRNA target prediction under relaxed seed and no conservation requirements. PLoS One 2011, 6, e20622. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, P.; Takahashi, K.; Saito, M.; Yoshida, Y.; Okita, K.; Watanabe, A.; Inoue, H.; Yamashita, J. K.; Todani, M.; Nakagawa, M.; Osawa, M.; Yashiro, Y.; Yamanaka, S.; Osafune, K. Induced Pluripotent Stem Cells and Their Use in Human Models of Disease and Development. Physiol Rev 2019, 99, 79–114. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, O.; Sugai, K.; Yamaguchi, R.; Tashiro, S.; Nagoshi, N.; Kohyama, J.; Iida, T.; Ohkubo, T.; Itakura, G.; Isoda, M.; Shinozaki, M.; Fujiyoshi, K.; Kanemura, Y.; Yamanaka, S.; Nakamura, M.; Okano, H. Concise Review: Laying the Groundwork for a First-In-Human Study of an Induced Pluripotent Stem Cell-Based Intervention for Spinal Cord Injury. Stem Cells 2019, 37, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L. F. R.; MacRae, I. J. Regulation of microRNA function in animals. Nat Rev Mol Cell Biol 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, P.; Boström, A. K.; Impens, F.; Fredlund, E.; Van Peer, G.; De Antonellis, P.; von Stedingk, K.; Ghesquière, B.; Schulte, S.; Dews, M.; Thomas-Tikhonenko, A.; Schulte, J. H.; Zollo, M.; Schramm, A.; Gevaert, K.; Axelson, H.; Speleman, F.; Vandesompele, J. The miR-17-92 microRNA cluster regulates multiple components of the TGF-β pathway in neuroblastoma. Mol Cell 2010, 40, 762–73. [Google Scholar] [CrossRef]
- Borsuk, R.; Zhou, L.; Chang, W. I.; Zhang, Y.; Sharma, A.; Prabhu, V. V.; Tapinos, N.; Lulla, R. R.; El-Deiry, W. S. Potent preclinical sensitivity to imipridone-based combination therapies in oncohistone H3K27M-mutant diffuse intrinsic pontine glioma is associated with induction of the integrated stress response, TRAIL death receptor DR5, reduced ClpX and apoptosis. Am J Cancer Res 2021, 11, 4607–4623. [Google Scholar]
- Saito, Y.; Nakaoka, T.; Saito, H. microRNA-34a as a Therapeutic Agent against Human Cancer. J Clin Med 2015, 4, 1951. [Google Scholar] [CrossRef]
- Bouchie, A. First microRNA mimic enters clinic. Nat Biotechnol 2013, 31, 577. [Google Scholar] [CrossRef]
- Daige, C. L.; Wiggins, J. F.; Priddy, L.; Nelligan-Davis, T.; Zhao, J.; Brown, D. Systemic delivery of a miR34a mimic as a potential therapeutic for liver cancer. Mol Cancer Ther 2014, 13, 2352–60. [Google Scholar] [CrossRef]
- Kelnar, K.; Bader, A. G. A qRT-PCR Method for Determining the Biodistribution Profile of a miR-34a Mimic. Methods Mol Biol 2015, 1317, 125–33. [Google Scholar] [PubMed]
- Diener, C.; Hart, M.; Alansary, D.; Poth, V.; Walch-Rückheim, B.; Menegatti, J.; Grässer, F.; Fehlmann, T.; Rheinheimer, S.; Niemeyer, B. A.; Lenhof, H. P.; Keller, A.; Meese, E. Modulation of intracellular calcium signaling by microRNA-34a-5p. Cell Death Dis 2018, 9, 1008. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.; Nickl, L.; Walch-Rueckheim, B.; Krammes, L.; Rheinheimer, S.; Diener, C.; Taenzer, T.; Kehl, T.; Sester, M.; Lenhof, H. P.; Keller, A.; Meese, E. Wrinkle in the plan: miR-34a-5p impacts chemokine signaling by modulating CXCL10/CXCL11/CXCR3-axis in CD4(+), CD8(+) T cells, and M1 macrophages. J Immunother Cancer 2020, 8, (2). [Google Scholar] [CrossRef] [PubMed]
- Raimondo, T. M.; Reed, K.; Shi, D.; Langer, R.; Anderson, D. G. Delivering the next generation of cancer immunotherapies with RNA. Cell 2023, 186, 1535–1540. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E. E.; Frenck, R. W.; Falsey, A. R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M. J.; Bailey, R.; Swanson, K. A.; Li, P.; Koury, K.; Kalina, W.; Cooper, D.; Fontes-Garfias, C.; Shi, P.-Y.; Türeci, Ö.; Tompkins, K. R.; Lyke, K. E.; Raabe, V.; Dormitzer, P. R.; Jansen, K. U.; Şahin, U.; Gruber, W. C. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. New England Journal of Medicine 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E. J.; Rouphael, N. G.; Widge, A. T.; Jackson, L. A.; Roberts, P. C.; Makhene, M.; Chappell, J. D.; Denison, M. R.; Stevens, L. J.; Pruijssers, A. J.; McDermott, A. B.; Flach, B.; Lin, B. C.; Doria-Rose, N. A.; O’Dell, S.; Schmidt, S. D.; Corbett, K. S.; Swanson, P. A.; Padilla, M.; Neuzil, K. M.; Bennett, H.; Leav, B.; Makowski, M.; Albert, J.; Cross, K.; Edara, V. V.; Floyd, K.; Suthar, M. S.; Martinez, D. R.; Baric, R.; Buchanan, W.; Luke, C. J.; Phadke, V. K.; Rostad, C. A.; Ledgerwood, J. E.; Graham, B. S.; Beigel, J. H. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. New England Journal of Medicine 2020, 383, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Aloy, E.; Connerty, P.; Salik, B.; Liu, B.; Woo, A. J.; Haber, M.; Norris, M. D.; Wang, J.; Wang, J. Y. miR-101 suppresses the development of MLL-rearranged acute myeloid leukemia. Haematologica 2019, 104, e296–e299. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Cheng, R.; Yang, F.; Yu, M.; Wang, C.; Cui, S.; Hong, Y.; Liang, H.; Liu, M.; Zhao, C.; Ding, M.; Sun, W.; Liu, Z.; Sun, F.; Zhang, C.; Zhou, Z.; Jiang, X.; Chen, X. The Jun/miR-22/HuR regulatory axis contributes to tumourigenesis in colorectal cancer. Mol Cancer 2018, 17, 11. [Google Scholar] [CrossRef]
- Zhu, S.; Cheng, X.; Wang, R.; Tan, Y.; Ge, M.; Li, D.; Xu, Q.; Sun, Y.; Zhao, C.; Chen, S.; Liu, H. Restoration of microRNA function impairs MYC-dependent maintenance of MLL leukemia. Leukemia 2020, 34, 2484–2488. [Google Scholar] [CrossRef]
- Bose, D.; Jayaraj, G.; Suryawanshi, H.; Agarwala, P.; Pore, S. K.; Banerjee, R.; Maiti, S. The tuberculosis drug streptomycin as a potential cancer therapeutic: inhibition of miR-21 function by directly targeting its precursor. Angew Chem Int Ed Engl 2012, 51, 1019–23. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, J.; Qian, X.; Han, L.; Zhang, K.; Chen, L.; Liu, J.; Ren, Y.; Yang, M.; Zhang, A.; Pu, P.; Kang, C. AC1MMYR2, an inhibitor of dicer-mediated biogenesis of Oncomir miR-21, reverses epithelial-mesenchymal transition and suppresses tumor growth and progression. Cancer Res 2013, 73, 5519–31. [Google Scholar] [CrossRef]
- Gumireddy, K.; Young, D. D.; Xiong, X.; Hogenesch, J. B.; Huang, Q.; Deiters, A. Small-molecule inhibitors of microrna miR-21 function. Angew Chem Int Ed Engl 2008, 47, 7482–4. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, N. S.; Manavalan, T. T.; Dougherty, S. M.; Riggs, K. A.; Li, Y.; Klinge, C. M. Estradiol downregulates miR-21 expression and increases miR-21 target gene expression in MCF-7 breast cancer cells. Nucleic Acids Res 2009, 37, 2584–95. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C. S.; Wang, X. M.; Zhang, S. Q.; Meng, L. S.; Zhu, W. H.; Xu, J.; Lu, S. M. Discovery of 4-benzoylamino-N-(prop-2-yn-1-yl)benzamides as novel microRNA-21 inhibitors. Bioorg Med Chem 2015, 23, 6510–9. [Google Scholar] [CrossRef] [PubMed]
- Naro, Y.; Thomas, M.; Stephens, M. D.; Connelly, C. M.; Deiters, A. Aryl amide small-molecule inhibitors of microRNA miR-21 function. Bioorg Med Chem Lett 2015, 25, 4793–4796. [Google Scholar] [CrossRef] [PubMed]
- Velagapudi, S. P.; Gallo, S. M.; Disney, M. D. Sequence-based design of bioactive small molecules that target precursor microRNAs. Nat Chem Biol 2014, 10, 291–7. [Google Scholar] [CrossRef]
- Zhang, S.; Ma, C.; Pang, H.; Zeng, F.; Cheng, L.; Fang, B.; Ma, J.; Shi, Y.; Hong, H.; Chen, J.; Wang, Z.; Xia, J. Arsenic trioxide suppresses cell growth and migration via inhibition of miR-27a in breast cancer cells. Biochem Biophys Res Commun 2016, 469, 55–61. [Google Scholar] [CrossRef]
- Bose, D.; Jayaraj, G. G.; Kumar, S.; Maiti, S. A molecular-beacon-based screen for small molecule inhibitors of miRNA maturation. ACS Chem Biol 2013, 8, 930–8. [Google Scholar] [CrossRef]
- Childs-Disney, J. L.; Disney, M. D. Small Molecule Targeting of a MicroRNA Associated with Hepatocellular Carcinoma. ACS Chem Biol 2016, 11, 375–80. [Google Scholar] [CrossRef]
- Diaz, J. P.; Chirayil, R.; Chirayil, S.; Tom, M.; Head, K. J.; Luebke, K. J. Association of a peptoid ligand with the apical loop of pri-miR-21 inhibits cleavage by Drosha. Rna 2014, 20, 528–39. [Google Scholar] [CrossRef]
- Kashtan, C. E.; Gross, O. Clinical practice recommendations for the diagnosis and management of Alport syndrome in children, adolescents, and young adults-an update for 2020. Pediatr Nephrol 2021, 36, 711–719. [Google Scholar] [CrossRef]
- Kashtan, C. E.; Gross, O. Correction to: Clinical practice recommendations for the diagnosis and management of Alport syndrome in children, adolescents, and young adults-an update for 2020. Pediatr Nephrol 2021, 36, 731. [Google Scholar] [CrossRef]
- Abplanalp, W. T.; Fischer, A.; John, D.; Zeiher, A. M.; Gosgnach, W.; Darville, H.; Montgomery, R.; Pestano, L.; Allée, G.; Paty, I.; Fougerousse, F.; Dimmeler, S. Efficiency and Target Derepression of Anti-miR-92a: Results of a First in Human Study. Nucleic Acid Ther 2020, 30, 335–345. [Google Scholar] [CrossRef]
- Reid, G.; Pel, M. E.; Kirschner, M. B.; Cheng, Y. Y.; Mugridge, N.; Weiss, J.; Williams, M.; Wright, C.; Edelman, J. J.; Vallely, M. P.; McCaughan, B. C.; Klebe, S.; Brahmbhatt, H.; MacDiarmid, J. A.; van Zandwijk, N. Restoring expression of miR-16: a novel approach to therapy for malignant pleural mesothelioma. Ann Oncol 2013, 24, 3128–35. [Google Scholar] [CrossRef] [PubMed]
- Täubel, J.; Hauke, W.; Rump, S.; Viereck, J.; Batkai, S.; Poetzsch, J.; Rode, L.; Weigt, H.; Genschel, C.; Lorch, U.; Theek, C.; Levin, A. A.; Bauersachs, J.; Solomon, S. D.; Thum, T. Novel antisense therapy targeting microRNA-132 in patients with heart failure: results of a first-in-human Phase 1b randomized, double-blind, placebo-controlled study. Eur Heart J 2021, 42, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Batkai, S.; Genschel, C.; Viereck, J.; Rump, S.; Bär, C.; Borchert, T.; Traxler, D.; Riesenhuber, M.; Spannbauer, A.; Lukovic, D.; Zlabinger, K.; Hašimbegović, E.; Winkler, J.; Garamvölgyi, R.; Neitzel, S.; Gyöngyösi, M.; Thum, T. CDR132L improves systolic and diastolic function in a large animal model of chronic heart failure. Eur Heart J 2021, 42, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Olena, A. F.; Patton, J. G. Genomic organization of microRNAs. J Cell Physiol 2010, 222, 540–5. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V. N. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol 2014, 15, 509–24. [Google Scholar] [CrossRef] [PubMed]
- Ottosen, S.; Parsley, T. B.; Yang, L.; Zeh, K.; van Doorn, L. J.; van der Veer, E.; Raney, A. K.; Hodges, M. R.; Patick, A. K. In vitro antiviral activity and preclinical and clinical resistance profile of miravirsen, a novel anti-hepatitis C virus therapeutic targeting the human factor miR-122. Antimicrob Agents Chemother 2015, 59, 599–608. [Google Scholar] [CrossRef]
- Elmén, J.; Lindow, M.; Schütz, S.; Lawrence, M.; Petri, A.; Obad, S.; Lindholm, M.; Hedtjärn, M.; Hansen, H. F.; Berger, U.; Gullans, S.; Kearney, P.; Sarnow, P.; Straarup, E. M.; Kauppinen, S. LNA-mediated microRNA silencing in non-human primates. Nature 2008, 452, 896–9. [Google Scholar] [CrossRef] [PubMed]
- Lanford, R. E.; Hildebrandt-Eriksen, E. S.; Petri, A.; Persson, R.; Lindow, M.; Munk, M. E.; Kauppinen, S.; Ørum, H. Therapeutic silencing of microRNA-122 in primates with chronic hepatitis C virus infection. Science 2010, 327, 198–201. [Google Scholar] [CrossRef]
- Janssen, H. L.; Reesink, H. W.; Lawitz, E. J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A. J.; Patick, A. K.; Chen, A.; Zhou, Y.; Persson, R.; King, B. D.; Kauppinen, S.; Levin, A. A.; Hodges, M. R. Treatment of HCV infection by targeting microRNA. N Engl J Med 2013, 368, 1685–94. [Google Scholar] [CrossRef]
- James, A. M.; Ruckman, J.; Pestano, L. A.; Hopkins, R. D.; Rodgers, R. C.; Marshall, W. S.; Rubin, P.; Escolar, D. SOLAR: A PHASE 2, GLOBAL, RANDOMIZED, ACTIVE COMPARATOR STUDY TO INVESTIGATE THE EFFICACY AND SAFETY OF COBOMARSEN IN SUBJECTS WITH MYCOSIS FUNGOIDES (MF). Hematological Oncology 2019, 37, 562–563. [Google Scholar] [CrossRef]
- Querfeld, C.; Pacheco, T.; Foss, F. M.; Halwani, A. S.; Porcu, P.; Seto, A. G.; Ruckman, J.; Landry, M. L.; Jackson, A. L.; Pestano, L. A.; Dickinson, B. A.; Sanseverino, M.; Rodman, D. M.; Gordon, G.; Marshall, W. Preliminary Results of a Phase 1 Trial Evaluating MRG-106, a Synthetic microRNA Antagonist (LNA antimiR) of microRNA-155, in Patients with CTCL. Blood 2016, 128, 1829–1829. [Google Scholar] [CrossRef]
- Adams, D.; Suhr, O. B.; Dyck, P. J.; Litchy, W. J.; Leahy, R. G.; Chen, J.; Gollob, J.; Coelho, T. Trial design and rationale for APOLLO, a Phase 3, placebo-controlled study of patisiran in patients with hereditary ATTR amyloidosis with polyneuropathy. BMC Neurol 2017, 17, 181. [Google Scholar] [CrossRef]
- Sousa, D. P.; Conde, J. Gold Nanoconjugates for miRNA Modulation in Cancer Therapy: From miRNA Silencing to miRNA Mimics. ACS Materials Au 2022, 2, 626–640. [Google Scholar] [CrossRef]
- Durso, M.; Gaglione, M.; Piras, L.; Mercurio, M. E.; Terreri, S.; Olivieri, M.; Marinelli, L.; Novellino, E.; Incoronato, M.; Grieco, P.; Orsini, G.; Tonon, G.; Messere, A.; Cimmino, A. Chemical modifications in the seed region of miRNAs 221/222 increase the silencing performances in gastrointestinal stromal tumor cells. European Journal of Medicinal Chemistry 2016, 111, 15–25. [Google Scholar] [CrossRef]
Figure 1.
Illustration of miRNA biogenesis, processing, and target RNA translational suppression or degradation via various mechanisms. Created with BioRender.com. miRNAs are a class of small, single–stranded non-coding RNAs that function as a guide molecule in RNA silencing and hence modulate the expression of most mRNAs. The miRNA:target mRNA interaction usually occurs at the 5′ end of the miRNA (i.e., ‘seed’ region). However, recent evidence suggests that there is a special class of target mRNAs which bind the miRNA not only through the ‘seed’ region, but also through a second region of complementarity at the 3′ end of the miRNA. The extended complementarity forces the miRNA out of Ago2, where it becomes accessible to enzymatic degradation. This phenomenon is referred to as target-directed miRNA degradation mechanism (TDMD) [113,114].
Figure 1.
Illustration of miRNA biogenesis, processing, and target RNA translational suppression or degradation via various mechanisms. Created with BioRender.com. miRNAs are a class of small, single–stranded non-coding RNAs that function as a guide molecule in RNA silencing and hence modulate the expression of most mRNAs. The miRNA:target mRNA interaction usually occurs at the 5′ end of the miRNA (i.e., ‘seed’ region). However, recent evidence suggests that there is a special class of target mRNAs which bind the miRNA not only through the ‘seed’ region, but also through a second region of complementarity at the 3′ end of the miRNA. The extended complementarity forces the miRNA out of Ago2, where it becomes accessible to enzymatic degradation. This phenomenon is referred to as target-directed miRNA degradation mechanism (TDMD) [113,114].
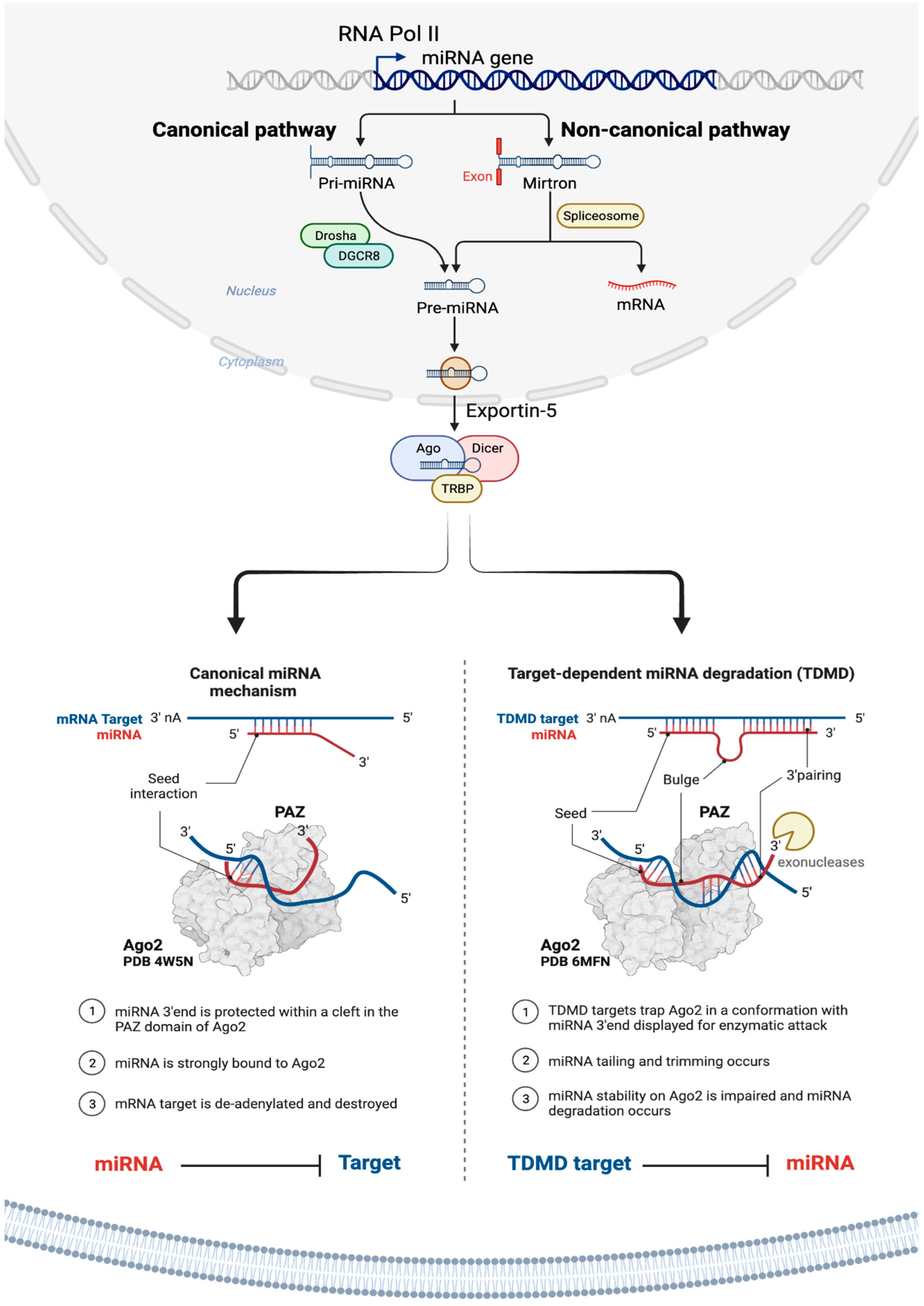
Figure 3.
Schematic illustration of different inhibition mechanisms of miRNA specific small molecule inhibitors. Created with BioRender.com.
Figure 3.
Schematic illustration of different inhibition mechanisms of miRNA specific small molecule inhibitors. Created with BioRender.com.
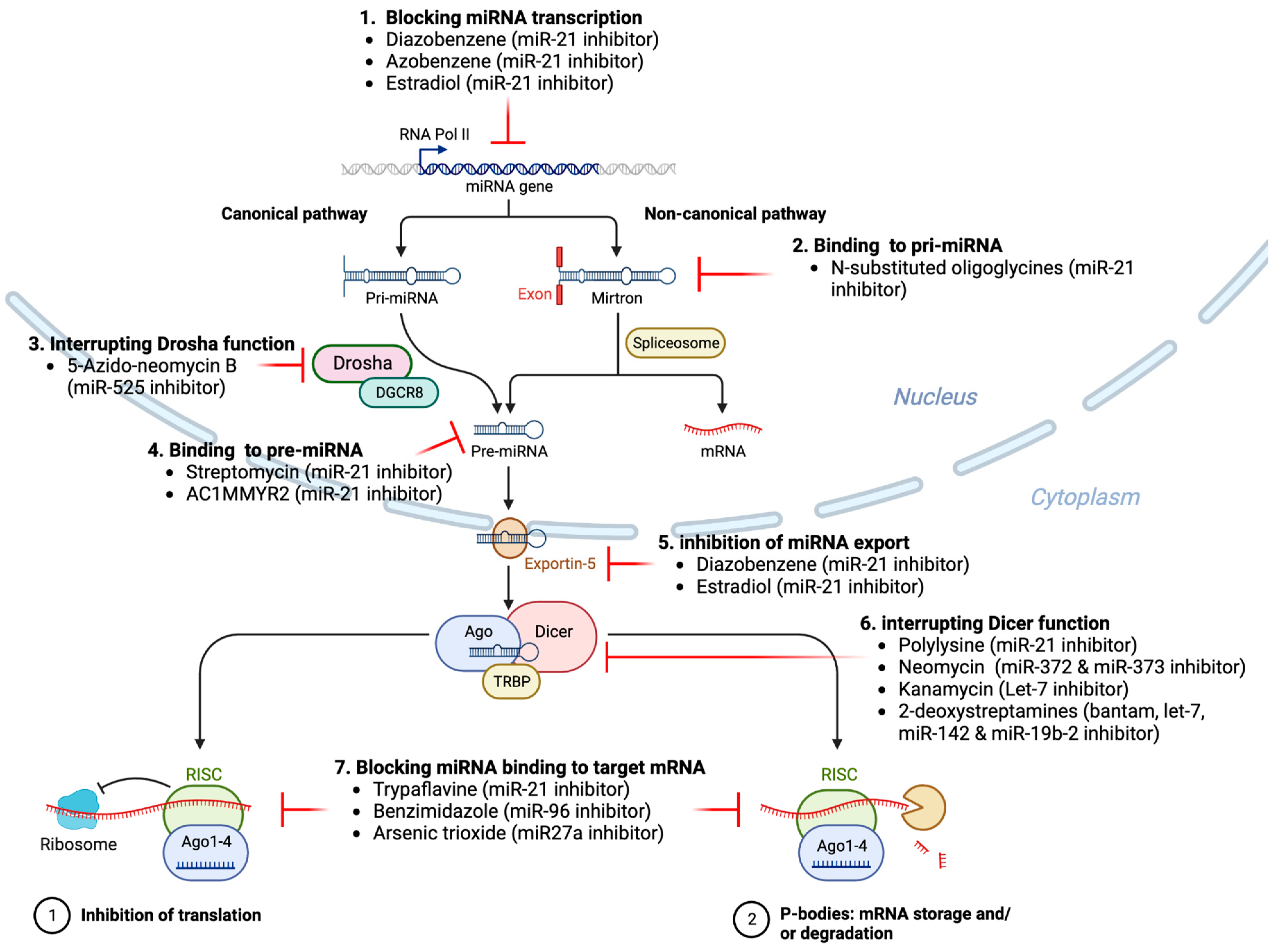
Figure 2.
Schematic overview of miRNA therapeutic strategies to regulate the function of oncogenic and tumor suppressor miRNAs involved in cancer. Top panel: The strategy of miRNA therapeutics is based on the strategy of restoring the balance of oncogenic miRNAs and tumor suppressor miRNAs by downregulation of oncomiRNAs or the restoring of tumor suppressor miRNAs. Bottom panel: Therapeutic miRNA manipulations can target the expression or function of pathologically relevant miRNAs via miRNA inhibitors (anti-miRs) mediating degradation or functional blocking of endogenous miRNAs, synthetic miRNA mimics imitating endogenous miRNA double-strands, viral vector expressed miRNAs, small molecules inhibitors interfering with miRNA biogenesis, or miRNA sponges causing functional inhibition by diverting endogenous miRNAs from their mRNA targets. In addition, combining miRNAs with chemotherapies, immunotherapies and other conventional drugs or siRNAs is another strategy to overcome drug resistance. Created with BioRender.com.
Figure 2.
Schematic overview of miRNA therapeutic strategies to regulate the function of oncogenic and tumor suppressor miRNAs involved in cancer. Top panel: The strategy of miRNA therapeutics is based on the strategy of restoring the balance of oncogenic miRNAs and tumor suppressor miRNAs by downregulation of oncomiRNAs or the restoring of tumor suppressor miRNAs. Bottom panel: Therapeutic miRNA manipulations can target the expression or function of pathologically relevant miRNAs via miRNA inhibitors (anti-miRs) mediating degradation or functional blocking of endogenous miRNAs, synthetic miRNA mimics imitating endogenous miRNA double-strands, viral vector expressed miRNAs, small molecules inhibitors interfering with miRNA biogenesis, or miRNA sponges causing functional inhibition by diverting endogenous miRNAs from their mRNA targets. In addition, combining miRNAs with chemotherapies, immunotherapies and other conventional drugs or siRNAs is another strategy to overcome drug resistance. Created with BioRender.com.

Figure 4.
Examples of miRNA delivery systems. MiRNA therapeutics can be administered orally or intranasally or through venous (intravenously) or muscle (intramuscularly) or skin (subcutaneously) injections, or via cell-/tissue-directed approaches, or adoptive cell transfer, or the implantation of 3D matrices that release miRNA therapeutics, or other extracorporeal miRNA delivery strategies [77]. Other modes of delivery of miRNA therapeutics include vector based and non-vector-based delivery systems including adeno-associated virus. (A), lentivirus (B), bacterial nanocells (C), bacteriophages (D) liposomes, including monovalent and multivalent lipids such as cationic lipid-based ligand-targeted nanoparticles (E), natural polymer-based nanoparticles (F), polymer based nanoparticles (natural, green and synthetic, blue) conjugated with polyethylene glycol (PEG) (G), extracellular vesicles or exosomes (H), gold nanoparticles [268] (I) carbon nanotubes (J), quantum dot nanoparticles (K), core-shell magnetic nanoparticles (L), and others such as polymeric micelles, and mesoporous silica nanoparticles are the examples of nanocarriers as drug-delivery systems [145]. Moreover, there have been efforts to improve the serum stability, pharmacokinetics, and tissue specificity by targeted delivery of miRNA mimics, miRNA inhibitors and other nucleic acid therapeutics by incorporation of various chemical modifications and/or conjugation of these RNA and nucleic acid therapeutics to biomolecules to facilitate receptor-mediated uptake such as N-acetylgalactosamine (GalNAc), 2′-O-methyl nucleotide, phosphorothioate, cholesterol, locked nucleic acid (LNA), and aptamer moieties are also shown as examples [77,84,186,208,269]. Created with BioRender.com.
Figure 4.
Examples of miRNA delivery systems. MiRNA therapeutics can be administered orally or intranasally or through venous (intravenously) or muscle (intramuscularly) or skin (subcutaneously) injections, or via cell-/tissue-directed approaches, or adoptive cell transfer, or the implantation of 3D matrices that release miRNA therapeutics, or other extracorporeal miRNA delivery strategies [77]. Other modes of delivery of miRNA therapeutics include vector based and non-vector-based delivery systems including adeno-associated virus. (A), lentivirus (B), bacterial nanocells (C), bacteriophages (D) liposomes, including monovalent and multivalent lipids such as cationic lipid-based ligand-targeted nanoparticles (E), natural polymer-based nanoparticles (F), polymer based nanoparticles (natural, green and synthetic, blue) conjugated with polyethylene glycol (PEG) (G), extracellular vesicles or exosomes (H), gold nanoparticles [268] (I) carbon nanotubes (J), quantum dot nanoparticles (K), core-shell magnetic nanoparticles (L), and others such as polymeric micelles, and mesoporous silica nanoparticles are the examples of nanocarriers as drug-delivery systems [145]. Moreover, there have been efforts to improve the serum stability, pharmacokinetics, and tissue specificity by targeted delivery of miRNA mimics, miRNA inhibitors and other nucleic acid therapeutics by incorporation of various chemical modifications and/or conjugation of these RNA and nucleic acid therapeutics to biomolecules to facilitate receptor-mediated uptake such as N-acetylgalactosamine (GalNAc), 2′-O-methyl nucleotide, phosphorothioate, cholesterol, locked nucleic acid (LNA), and aptamer moieties are also shown as examples [77,84,186,208,269]. Created with BioRender.com.
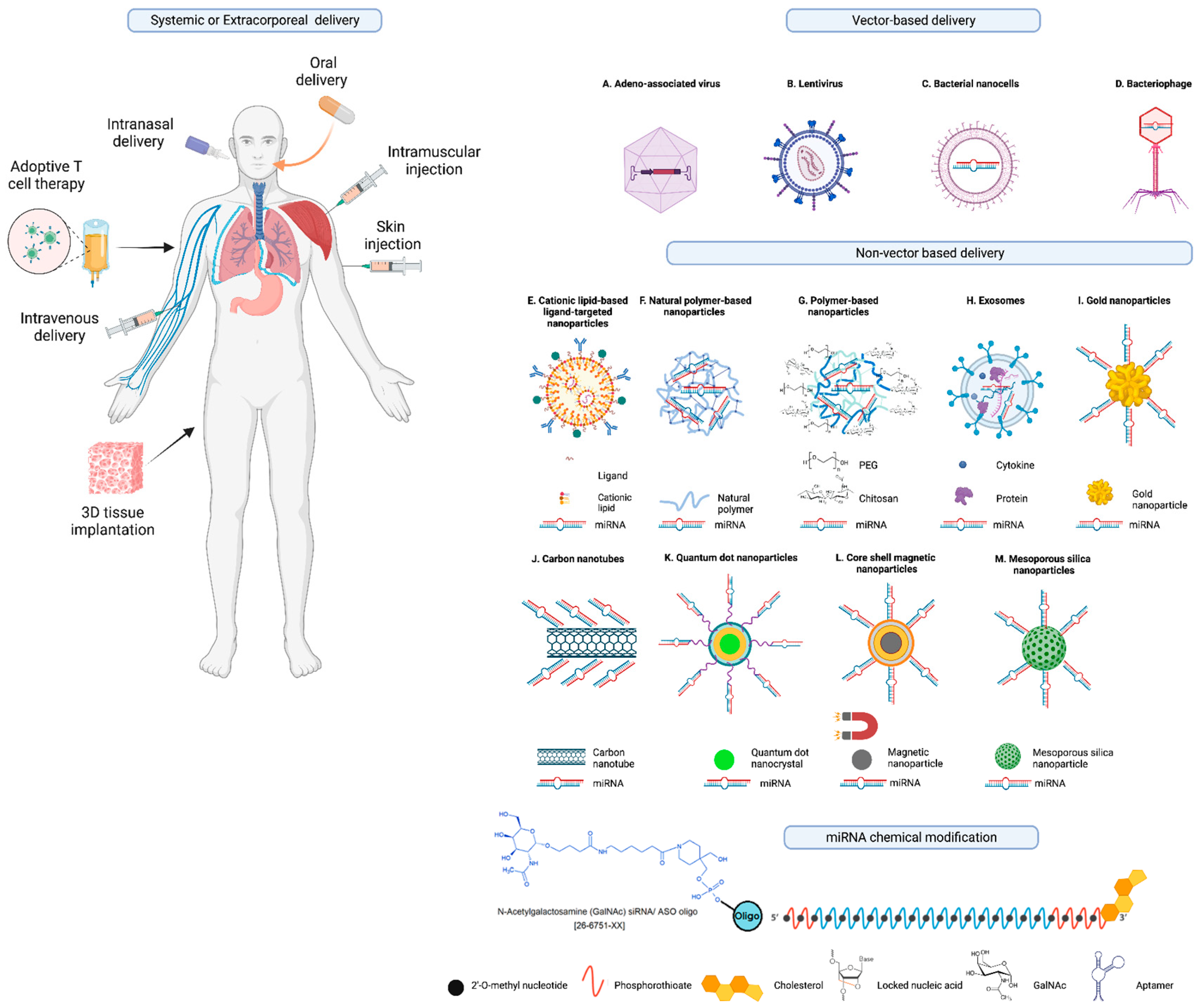
Figure 5.
Illustration of the miRNA drugs discovery and development process beginning from the target identification and miRNA discovery to FDA approved miRNA therapeutics in the market. In the traditional drug development process, the timeline from target, drug discovery to phase 1-3 human clinical trials and, ultimately, FDA approval, followed by Phase 4 studies can go on for several years. Conversely, RNA-based and more specifically, miRNA-based drug development can accelerate miRNA-based drug discovery and development, potentially mitigating attrition rates, time constraints, and costs. Created with BioRender.com.
Figure 5.
Illustration of the miRNA drugs discovery and development process beginning from the target identification and miRNA discovery to FDA approved miRNA therapeutics in the market. In the traditional drug development process, the timeline from target, drug discovery to phase 1-3 human clinical trials and, ultimately, FDA approval, followed by Phase 4 studies can go on for several years. Conversely, RNA-based and more specifically, miRNA-based drug development can accelerate miRNA-based drug discovery and development, potentially mitigating attrition rates, time constraints, and costs. Created with BioRender.com.
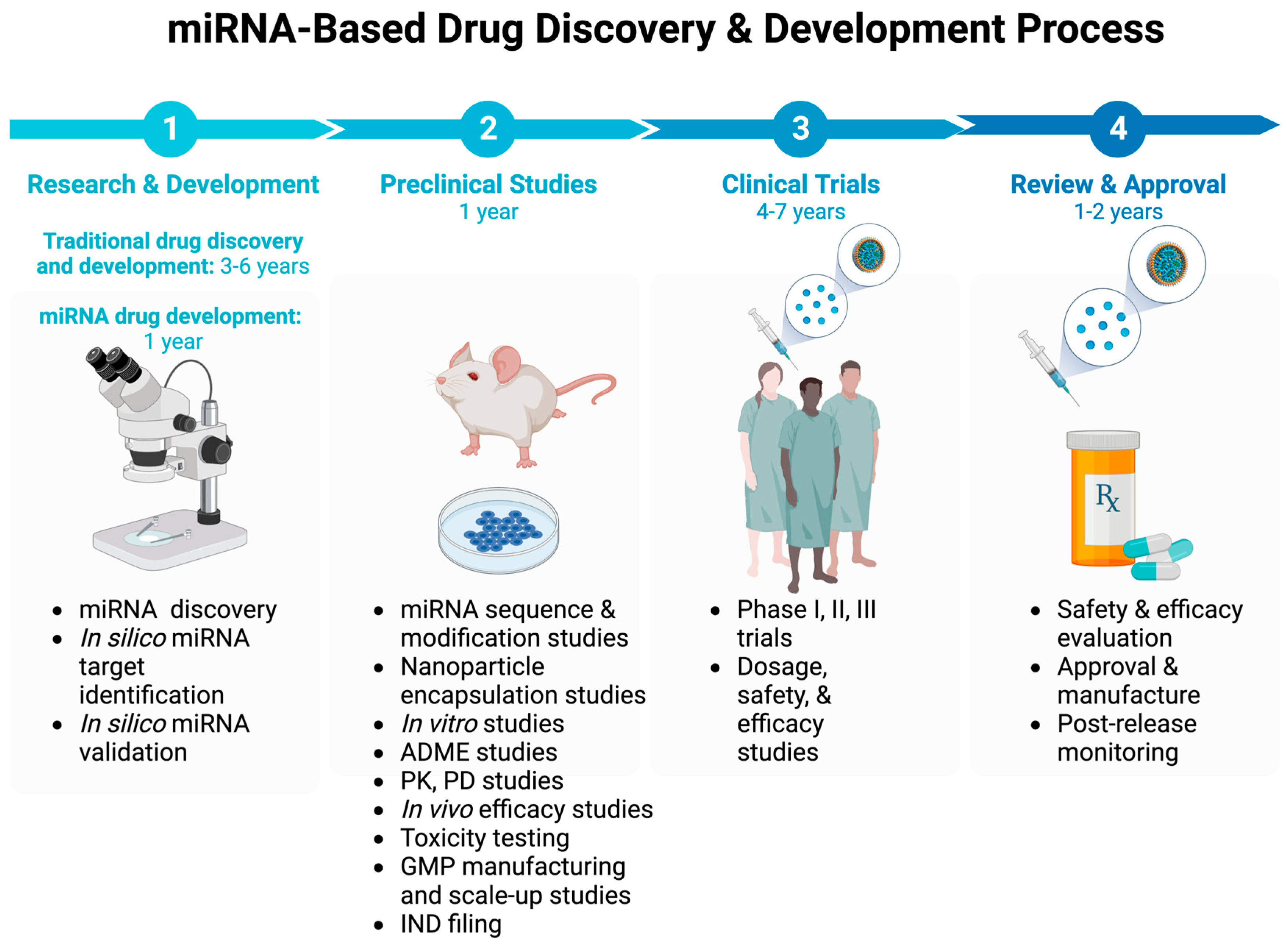

Table 2.
Major miRNA based therapeutics in various stages of the preclinical development phase for human malignancies.
Table 2.
Major miRNA based therapeutics in various stages of the preclinical development phase for human malignancies.
| Therapeutic molecule | Target miRNA | Disease | Biopharmaceutical company | Stage of development |
|---|---|---|---|---|
| RG-012 | miRNA-21 | Alport nephropathy | Regulus therapeutics (with the strategic alliance with Genzyme) | Preclinical stage |
| MGN-1374 | miRNA-15 and miR-195 | Post-myocardial infarction | miRagen therapeutics | Preclinical stage |
| MGN-2677 | miR-143/145 | Vascular disease | miRagen therapeutics | Preclinical stage |
| MGN-4220 | miR-29 | Cardiac fibrosis | miRagen therapeutics | Preclinical stage |
| MGN-4893 | miR-451 | For the treatment of disorders like abnormal red blood cell production | miRagen therapeutics. | Preclinical stage |
| MGN-5804 | miR-378 | Cardiometabolic disease | miRagen therapeutics | Preclinical stage |
| MGN-6114 | miR-92 | Peripheral arterial disease | miRagen therapeutics | Preclinical stage |
| MGN-9103 | miR-208 | Chronic heart failure | miRagen therapeutics | Preclinical stage |
| MRG-107 | miR-155 | Amyotrophic lateral sclerosis (ALS) | miRagen therapeutics | Completed preclinical stage |
Table 3.
Clinical trials with miRNA therapeutics in various diseases. NCT numbered trials are registered at ClinicalTrials.gov; EudraCT numbered trials are registered at EU Clinical Trials Register (clinicaltrialsregister.eu).
Table 3.
Clinical trials with miRNA therapeutics in various diseases. NCT numbered trials are registered at ClinicalTrials.gov; EudraCT numbered trials are registered at EU Clinical Trials Register (clinicaltrialsregister.eu).
| miRNA drug name | Targeted miRNA | Study Title | Mode of action | Disease/condition | Mode of delivery | Phase | Status | Clinical trial number(s) | References |
|---|---|---|---|---|---|---|---|---|---|
| miR-10b | miR-10b | Evaluating the Expression Levels of MicroRNA-10b in Patients With Gliomas | miR-10b as diagnostic and in vitro testing of anti-mir-10b as therapeutic | AstrocytomaOligodendrogliomaOligoastrocytomaAnaplastic AstrocytomaAnaplastic OligodendrogliomaAnaplastic OligoastrocytomaGlioblastomaBrain TumorsBrain Cancer | Observational | Recruiting | NCT01849952 | ||
| INT-1B3 | miR-193a-3p mimic | First-in-Human Study of INT-1B3 in Patients With Advanced Solid Tumors | miRNA mimic | Advanced solid tumors | Phase I | Recruiting | NCT04675996 | NA | |
| AMT-130 | Artificial miRNA | Safety and Proof-of-Concept (POC) Study With AMT-130 in Adults With Early Manifest Huntington’s Disease | A miRNA expression | Huntington disease | Stereotaxic infusion/viral transfer (adeno-associated vector) | Phase I | Ongoing | NCT04120493 | [23,24,25,195,196,197] |
| RG-012/lademirsen/SAR339375 | miR-21 | A Study of RG-012 in Subjects With Alport Syndrome | Anti-miR-21 Lademirsen—also known as RG-012, RG456070 or (SAR339375) | Alport syndrome | Subcutaneous injection/chemical modification (phosphorothioate) | Phase II | Completed | NCT03373786 | [209,222,253,254] |
| RG-012/lademirsen/SAR339375 | miR-21 | Study of Lademirsen (SAR339375) in Patients With Alport Syndrome | Anti-miR-21 Lademirsen—also known as RG-012, RG456070 or (SAR339375) | Alport syndrome | Subcutaneous injection/chemical modification (phosphorothioate) | Phase II | Terminated | NCT02855268 | [209,222,253,254] |
| RGLS4326 | miR-17 | A Study of RGLS4326 in Patients With Autosomal Dominant Polycystic Kidney Disease | Anti-miR-17 | Autosomal dominant polycystic kidney disease | Administered via subcutaneous injection | Phase I | Completed | NCT04536688 | |
| RG-125/AZD4076 | miR-103/107 | A Study to Assess the Safety and Tolerability of Single Doses of AZD4076 in Healthy Male Subjects | Anti-miR | Non-alcoholic Steatohepatitis (NASH) | Subcutaneous injection/biomolecule conjugation (GalNAc) | Phase I | Active, not recruiting | NCT02612662, NCT02826525 | [207,208,209] |
| RG-125/AZD4076 | miR-103/107 | AZD4076 in Type 2 Diabetic Subjects With Non-Alcoholic Fatty Liver Disease | Anti-miR | T2DM With NAFLD | Subcutaneous injection/biomolecule conjugation (GalNAc) | Phase I | Completed | NCT02826525 | [207,208,209] |
| MRG-110 | miR-92a | Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of MRG-110 Following Intradermal Injection in Healthy Volunteers | Anti-miR | Healthy Volunteer | Skin injection/chemical modification (LNA) | Phase I | Completed | NCT03603431 | [159,255] |
| MesomiR 1 | miR-16 | MesomiR 1: A Phase I Study of TargomiRs as 2nd or 3rd Line Treatment for Patients With Recurrent MPM and NSCLC | miRNA mimic | Malignant pleural mesothelioma, non–small cell lung cancer | Intravenously/vehicle transfer (nonliving bacterial nanocells (EDVs or TargomiRs) | Phase I | Completed | NCT02369198 | [162,200,256] |
| CDR132L | miR-132 | Clinical Study to Assess Safety, PK and PD Parameters of CDR132L | Anti-miR | Heart failure | Intravenously/chemical modification (LNA) | Phase I | Completed | NCT04045405 | [257,258] |
| Remlarsen/MRG-201 | miR-29 | Efficacy, Safety, and Tolerability of Remlarsen (MRG-201) Following Intradermal Injection in Subjects With a History of Keloids | miRNA mimic | Keloid disorder | Skin injection/biomolecule conjugation (cholesterol) | Phase II | Completed | NCT03601052 | [160,259,260] |
| Miravirsen/SPC3649 | miR-122 | Long-Term Extension Study of Miravirsen Among Participants With Genotype 1 Chronic Hepatitis C (CHC) Who Have Not Responded to Pegylated-Interferon Alpha Plus Ribavirin | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/chemical modification (LNA) | Phase IIPhase IIPhase IIPhase IIPhase IIPhase I | CompletedCompletedCompletedUnknownUnknownCompleted | NCT02508090NCT02508090, NCT02452814, NCT01200420, NCT01872936, NCT01727934, NCT01646489 | [152,261,262,263,264] |
| Miravirsen/SPC3649 | miR-122 | Long Term Extension Study is Designed to Monitor Long-Term Efficacy and Safety of Miravirsen Sodium in Combination With Telaprevir and Ribavirin in Subjects With Chronic Hepatitis C Virus Genotype 1 Infection | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/chemical modification (LNA) | Phase II | Completed | NCT02452814 | [152,261,262,263,264] |
| Miravirsen/SPC3649 | miR-122 | Multiple Ascending Dose Study of Miravirsen in Treatment-Naïve Chronic Hepatitis C Subjects | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/chemical modification (LNA) | Phase IIPhase IIPhase IIPhase I | CompletedUnknownUnknownCompleted | NCT01200420 | [152,261,262,263,264] |
| Miravirsen/SPC3649 | miR-122 | Miravirsen in Combination With Telaprevir and Ribavirin in Null Responder to Pegylated-Interferon Alpha Plus Ribavirin Subjects With Chronic Hepatitis C Virus Infection | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/chemical modification (LNA) | Phase IIPhase IIPhase I | Unknown | NCT01872936 | [152,261,262,263,264] |
| Miravirsen/SPC3649 | miR-122 | Miravirsen Study in Null Responder to Pegylated Interferon Alpha Plus Ribavirin Subjects With Chronic Hepatitis C | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/chemical modification (LNA) | Phase IIPhase IIPhase I | Unknown | NCT01727934 | [152,261,262,263,264] |
| Miravirsen/SPC3649 | miR-122 | Drug Interaction Study to Assess the Effect of Co-Administered Miravirsen and Telaprevir in Healthy Subjects | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/chemical modification (LNA) | Phase IIPhase IIPhase I | Completed | NCT01646489 | [152,261,262,263,264] |
| RG-101 | miR-122 | A Randomized, Multi-Center, Phase 2 Study to Evaluate Safety and Efficacy of Subcutaneous Injections of RG-101 in Combination with Oral Agents in Treatment Naïve, Genotype 1 and 4, Chronic Hepatitis. | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/biomolecule conjugation (GalNAc) | Phase II | EudraCT numbers 2015-001535-21, 2015-004702-42, 2016-002069-77 | [156,209,210] | |
| RG-101 | miR-122 | A Multi-Center, Parallel Group, Open-Label, Phase 2 Study to Evaluate the Efficacy and Safety of a Single Subcutaneous Injection of RG-101 Combined with Oral GSK2878175 | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/biomolecule conjugation (GalNAc) | Phase II | EudraCT numbers 2015-004702-42 | [156,209,210] | |
| RG-101 | miR-122 | An Observational Long-Term Safety and Efficacy Follow-Up Study of Subjects Who Have Previously Received RG-101 | Anti-miR | Chronic hepatitis C virus | Subcutaneous injection/biomolecule conjugation (GalNAc) | Observational | Unknown | EudraCT numbers 2016-002069-77 | [156,209,210] |
| MRX34 | miR-34a | A Multicenter Phase I Study of MRX34, MicroRNA miR-RX34 Liposomal Injection | miRNA mimic | Primary Liver CancerSCLCLymphomaMelanomaMultiple MyelomaRenal Cell CarcinomaNSCLC | Intravenously/vehicle transfer (liposomal) | Phase I | Terminated (5 immune related serious adverse events) | NCT01829971 | [29,32,133][65,82,83] |
| MRX34 | miR-34a | Pharmacodynamics Study of MRX34, MicroRNA Liposomal Injection in Melanoma Patients With Biopsy Accessible Lesions | miRNA mimic | Solid tumors (e.g., hepatocellular carcinoma, melanoma, SCLC, NSCLC, lymphoma, multiple myeloma, renal cell carcinoma | Intravenously/vehicle transfer (liposomal) | Phase IPhase II | Withdrawn (5 immune related serious adverse events in Phase I) | NCT02862145 | [29,32,133][65,82,83] |
| Cobomarsen/MRG-106 | miR-155 | Anti-miR | Mycosis fungoides (MF)Cutaneous T-cell Lymphoma (CTCL)Chronic Lymphocytic Leukemia (CLL)Diffuse Large B-Cell Lymphoma (DLBCL), ABC SubtypeAdult T-Cell Leukemia/Lymphoma (ATLL) | Intravenously/chemical modification (LNA) | Phase IPhase IIPhase II | CompletedTerminatedTerminated | NCT02580552, NCT03713320, NCT03837457 | [157,265,266] | |
| Serum MicroRNA-25 | miR-25 | Serum miR-25 as diagnostic | Pancreatic cancer | Serum miR-25 | Observational | Not yet recruiting | NCT03432624 | ||
| Patisiran (ALN-TTR02), | RNAi therapeutic | Transthyretin (TTR)-Mediated Amyloidosis | ALN-TTR02 administered by intravenous infusion | Phase III | Completed | NCT01960348 | [267] | ||
| miR-10 | miR-10 | Evaluating the Expression Levels of MicroRNA-10b in Patients With Gliomas | anti-miR-10 | Glioma | Evaluating the expression levels of microRNA-10b in patients with gliomas | Observational | Recruiting | NCT01849952 |
Table 1.
Representative examples of small-molecule miRNA inhibitors.
| Small molecule inhibitors of miRNAs | Target miRNAs | Mechanism of action | References |
|---|---|---|---|
| Trypaflavine | miR-21 | Blocking the assembly of miR-21 with Ago2 | [175] |
| Streptomycin | miR-21/ miR-27a | Blocking the cleavage of pre-miR-21 by Dicer | [242] |
| AC1MMYR2 | miR-21 | Blocking the cleavage of pre-miR-21 to produce mature miR-21 | [243] |
| Diazobenzene | miR-21 | Inhibition the transcription of miR-21 gene | [244] |
| Azobenzene | miR-21 | Inhibition the transcription of miR-21 gene | [244] |
| Estradiol | miR-21 | Inhibition the transcription of miR-21 gene | [245] |
| Polylysine | miR-21 | Blocking the formation of mature of pre-miR-21 by the inhibition of Dicer | [175] |
| 4-benzoylamino-N-(prop-2-yn- 1-yl)benzamides | miR-21 | Up-regulation of PDCD4, the function target of miR-21 | [246] |
| Arylamide derivatives | miR-21 | Blocking the mature of pre-miR-21 | [247] |
| Kanamycin A | Let-7/ miR-27a | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 1 | Let-7 | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 2 | Let-7 | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 3 | Let-7 | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 4 | Let-7 | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 5 | Let-7 | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 6 | Let-7 | Binding to pre-let-7 and blocking the function of Dicer | [176] |
| 2-DOS Compound 7 | Bantam | Binding to pre-bantam and blocking the function of Dicer | [176] |
| 2-DOS Compound 8 | miR-142 | Binding to pre-miR-142 and blocking the function of Dicer | [176] |
| 2-DOS Compound 9 | miR-19b-2 | Binding to pre-miR-19b-2 and blocking the function of Dicer | [176] |
| NSC 158959 | miR-122 | Inhibition of the transcription of miR-122 | [177] |
| NSC 5476 | miR-122 | Inhibition of the transcription of miR-122 | [177]) |
| Benzimidazole | miR-96 | Up-regulation of FOXO1, the function target of miR-21 | [248] |
| 2-methoxy-1,4-naphthalenequin | miR-1 | Down-regulation the expression level of miR-1 | [183] |
| Arsenic trioxide | miR-27a | Down-regulation the expression level of miR-27a | [249] |
| Neomycin | miR-27a | Blocking the mature of miR-27a by the inhibition of Dicer | [250] |
| Amikacin | miR-27a | Blocking the mature of miR-27a by the inhibition of Dicer | [250] |
| Tobramycin | miR-27a | Blocking the mature of miR-27a by the inhibition of Dicer | [250] |
| 5″-azido-neomycin B | miR-525 | Binding to the processing site of Drosha to block the generation of pre- miR-525 | [251] |
| N-substituted oligoglycines | miR-21 | A specific ligand binding with pri-miR-21. | [252] |
Table 4.
Key questions to address before miRNA therapeutics become clinically impactful.
| 1 | What methods can be used to effectively guide therapeutic miRNAs/miRNA inhibitors to their intended target tissue and cells in vivo? |
| 2 | How can the design of miRNA/miRNA-based drugs and delivery vehicles be optimized to reduce or, ideally, eliminate unintended impacts on non-targeted cells? |
| 3 | What other strategies can be used to improve more accurate targeting for miRNA/miRNA inhibitor therapeutics? |
| 4 | Is there a risk of incompatibilities when using diverse carrier materials for advanced miRNA/miRNA inhibitor-based drug delivery, which may lead to undesired interactions between the materials and miRNA therapeutics? |
| 5 | Is there a risk of incompatibilities when using miRNA/miRNA inhibitor therapeutics in combination with traditional drugs pose the risk of incompatibilities? |
| 6 | Do modifications of miRNA/miRNA inhibitors such as LNA miRNA/miRNA inhibitors and other synthetic miRNAs, as well as agents such cell-permeable molecules, delivery methods such as biodegradable 3D matrices, carriers like functionalized metals, viral transfer systems, or biomolecule combinations such as aptamers invoke immunogenic responses? If so, can the activation of immunogenic responses be ameliorated through the masking of reactive components or moieties? |
| 7 | What is the level of risk associated with genomic integrations of viral transduction constructs that carry miRNA or miRNA inhibitors? |
| 8 | What is the impact of the expression of endogenous miRNAs and mRNAs on exogenously delivered therapeutic miRNAs and miRNA inhibitors which may be also affected by factors like cell type, cell cycle, and the cellular environment? |
| 9 | What is the necessary dosage for particular administration techniques for miRNA/miRNA inhibitors, such as skin injection, infusion, or inhalation, and for carrier-based methods like biodegradable 3D matrices? |
| 10 | How can the administration of miRNA/miRNA inhibitor therapeutic doses be regulated along intricate in vivo delivery pathways? |
| 11 | Is it possible to achieve consistent and sustainable rates of cellular uptake of miRNA/miRNA inhibitor therapeutics under varying in vivo conditions? |
| 12 | In what ways can dosing of miRNA mimics and inhibitors support the desired gene targeting outcome? |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



