Submitted:
21 December 2023
Posted:
22 December 2023
You are already at the latest version
Abstract
Telomeres are specialized structures at the ends of linear chromosomes that protect them from degradation and fusion. Telomere replication is a complex process that involves both DNA polymerases and a specialized enzyme called telomerase. Telomerase is a ribonucleoprotein complex that synthesizes telomeric DNA by using an internal RNA template. However, telomerase alone cannot fully replicate the telomeric DNA, and requires the cooperation of other factors, such as shelterin, CST, and DNA repair proteins. Moreover, telomere replication is tightly regulated by various mechanisms, such as cell cycle checkpoints, telomere length homeostasis, and telomere position effect. Dysregulation of telomere replication can lead to genomic instability, cellular senescence, and cancer. Therefore, understanding the molecular details of telomere replication is crucial for elucidating the role of telomeres in aging and disease.
Keywords:
telomere
; DNA replication
; chromosome
; shelterin
; molecular biology
Challenges and Dynamics of Telomere Replication
Eukaryotic cells have linear chromosomes that enable the shuffling of alleles between homologous chromosomes during meiosis, which increases genetic diversity [1]. However, linear chromosomes also have telomeres, which are vulnerable regions at the ends of chromosomes that consist of thousands of repeats of the sequence 5′-TTAGGG-3′, with a single-stranded 3′ overhang that can form a loop structure by invading the double-stranded repeats [3]. Telomeres are bound by the shelterin complex, which includes TRF1, TRF2, POT1, TIN2, RAP1, and TPP1, and protects telomeres from DNA damage responses and end joining, which can cause genomic instability, cell cycle arrest, senescence, or cell death [3]. Telomeres are shortened by the end replication problem, which occurs during the leading-strand synthesis and removes some of the telomeric repeats [2,4]. To prevent telomere erosion, shelterin recruits telomerase, a reverse transcriptase that adds repeats to the overhang using its RNA component (TERC) [5]. Telomerase is regulated by the CST complex, which also promotes the lagging-strand synthesis [6]. Cancer cells can bypass telomere shortening by activating telomerase or using the ALT mechanism [7]. Telomeres are challenging regions for DNA replication, as they present multiple obstacles for the replication machinery. Replication forks often slow down and stall near the telomeric chromatin, and may collapse if not resolved, leading to double-strand breaks and homologous recombination [2,8]. This can result in telomere loss or aberrations, which can be detected by FISH on metaphase chromosomes [9]. These abnormal structures may also reflect telomere entanglement or incomplete replication. Telomere replication is therefore a source of stress that threatens telomere integrity and stability. The timing of telomere replication is also crucial for telomere homeostasis and telomerase regulation [2]. In mammalian cells, telomeres replicate throughout the S phase, whereas in yeasts, they replicate at the end of the S phase [10]. However, short telomeres or global replication perturbations can advance the replication of telomeres, altering the telomere length equilibrium [11].
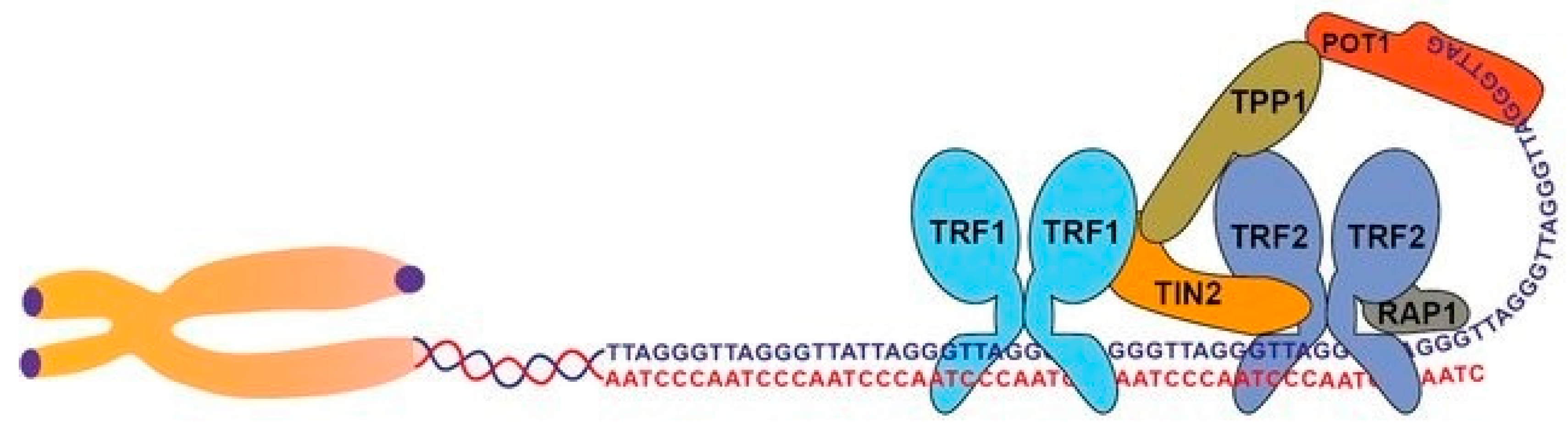
Figure 1.
The role of shelterin in protecting and regulating telomeres. This is a diagram of how the shelterin complex attaches to the telomeric DNA. The shelterin components TRF1 and TRF2 form dimers that bind to specific regions of the telomeric DNA. Image source: Doksani Y. (2019). The Response to DNA Damage at Telomeric Repeats and Its Consequences for Telomere Function. Genes, 10(4), 318. https://doi.org/10.3390/genes10040318
Figure 1.
The role of shelterin in protecting and regulating telomeres. This is a diagram of how the shelterin complex attaches to the telomeric DNA. The shelterin components TRF1 and TRF2 form dimers that bind to specific regions of the telomeric DNA. Image source: Doksani Y. (2019). The Response to DNA Damage at Telomeric Repeats and Its Consequences for Telomere Function. Genes, 10(4), 318. https://doi.org/10.3390/genes10040318
Unwinding of G4 Structures for Telomere Replication
The obstacles that slows down the replication fork include heterochromatin, T-loop, TERRA, RNA:DNA hybrids, and nuclear envelope attachment. One of the most challenging obstacles is the G-quadruplex (G4) structure, which is formed by four guanines stacking together in a planar arrangement. G4 can occur in the single-stranded G-rich lagging strand template during replication or transcription, and can block the fork or cause it to break [2]. This can lead to chromosome instability and telomere loss [12]. To prevent this, cells have several strategies to overcome the telomere replication problem, such as helicases, nucleases, and fork protection complex (FPC). The FPC is part of the replisome and ensures proper fork pausing and passage [13]. The shelterin complex also helps to promote efficient telomere replication and prevent fork stalling and collapse [2,14]. Thus, replisome and shelterin cooperate to maintain telomere stability. To prevent the interference of G4 structures with telomere replication, several helicases and single-strand DNA binding proteins (SSB) are recruited to unwind G4. For example, WRN and BLM, which are 3′–5′-directed helicases from the RecQ family that are mutated in Werner’s and Bloom’s syndromes, respectively [2,15]. WRN may be involved in G4 resolution at telomeres by interacting with replication factor A complex (RPA), PCNA, Pol δ, and TRF2 [16]. RPA can also bind and unfold G4 structures by itself, or recruit other helicases through physical interactions [17]. Telomeric proteins, such as POT1 and the shelterin components TRF1 and TRF2, may also prevent G4 formation by binding to telomeric tails or acting as scaffolds for replication factors [18,19]. The proliferating cell nuclear antigen (PCNA) may coordinate this network by recruiting different factors to the replisome [19].
BLM may collaborate with TRF1, which has the FxLxP motif for BLM binding [20]. TRF1 may also recruit BLM to remove G4 and avoid telomere fragility [20]. Another helicase that can resolve G4 with a 5′–3′ polarity is RTEL1, which is essential for DNA replication and recombination [21]. RTEL1 may be associated with the replisome by its PIP box domain that binds PCNA [21]. BLM and RTEL1 have different roles, as their deficiency causes additive telomere fragility [2,21]. Therefore, helicases that are linked to the replisome or shelterin can unwind G4 and ensure telomere replication.The Pif1 helicase family is widespread in eukaryotes and has various roles in DNA metabolism, including G4 unwinding. In yeast, there are two Pif1 family members: ScPif1 and Rrm3. ScPif1 is a potent G4 unwinder that inhibits telomerase by displacing its RNA component from telomeric ends [22]. Rrm3 travels with the replication fork and helps replicate telomeric repeats [23]. In humans and mice, PIF1 also unwinds G4 and interacts with TERT [24]. In fission yeast, Pfh1 is essential for replicating difficult regions and resolving G4 at telomeres [2,25]. Another protein that may process G4 at telomeres is DNA2, a 5′–3′ helicase/nuclease that cleaves G4 in vitro and co-immunoprecipitates with TRF1–TRF2 [26].
Overcoming Replication Challenges at Telomeres
The T-loop is a structure formed by the invasion of the telomeric 3′ overhang into the double-stranded part of the telomere, creating a D-loop. This protects the telomere from degradation, but also poses a challenge for DNA replication. To avoid replication fork collision and allow telomerase access, the T-loop needs to be disassembled in a timely manner. RTEL1 is a helicase that participates in this process by interacting with the shelterin protein TRF2, that binds to the T-loop base [27]. RTEL1 also associates with the replisome through PCNA to promote replication [28]. How RTEL1 coordinates its interactions with PCNA and TRF2 throughout the cell cycle is unclear, as well as how it distinguishes between different replication barriers such as G4, T-loops, or others. Helicases, such as WRN, BLM, and RECQL4, may also be involved in T-loop resolution [2,29]. If RTEL1 fails, the SLX1–SLX4 nucleases resolve the T-loop inappropriately, causing telomere instability [30]. TRF2 also recruits Apollo, a 5′-exonuclease that prevents topological stress at the T-loop base [31]. The regulation of T-loop resolution likely depends on a complex network of post-translational modifications, involving the shelterin proteins. TERRA is a type of non-coding RNA that is transcribed from the subtelomeric regions to the TTAGGG repeats at the ends of eukaryotic chromosomes [32]. TERRA can form RNA:DNA hybrids with the telomeric DNA, displacing the G-rich strand and creating R-loops [33]. This R-loop can interfere with the replication of telomeric repeats and cause telomere fragility and genomic instability [34]. To prevent this, TERRA levels are regulated during the cell cycle, peaking at G1–S and declining from S to G2 [2,35]. Moreover, several factors are involved in resolving TERRA R-loops, such as RNase H, which degrades the RNA strand [36], ATRX, which is a chromatin remodeler that may recognize or modify G4 structures [36], and UPF1, which is a helicase that participates in telomere replication [37]. These mechanisms ensure that TERRA does not impair the completion of leading-strand telomere replication and maintain telomere integrity.
TERRA also has many positive roles in telomere biology, such as regulating telomere length, replication, protection, chromatin structure, and mobility [38]. Therefore, TERRA levels and R-loop formation must be tightly controlled to avoid replication–transcription conflicts [2]. Several proteins can degrade or displace TERRA, such as Pif1 and FEN1 helicases, but the coordination and regulation of these mechanisms are not fully understood [39]. Telomeres also form a compact chromatin structure that protects them from DNA damage response, but also poses a barrier to the replication fork. TRF2 binds to telomeric DNA, modulates the topological state of telomeres and cooperates with Apollo and topoisomerase 2α to remove superhelical constraints [40]. Telomere anchoring is another source of topological stress that needs to be resolved during replication. The nuclear envelope (NE) and the nuclear matrix (NM) are two structures that constrain the localization and movement of telomeres, the ends of chromosomes. Telomeres are attached to the NE on one side of the nucleus and centromeres on the other in yeast cells [41]. This attachment is mediated by different proteins, such as Esc1–Sir4–Rap1 and yKu–Mps3 in budding yeast, and Bqt4 and Rap1 in fission yeast [2]. Fft3, a chromatin remodeler, also contributes to telomere anchoring [42]. Human telomeres, however, are distributed throughout the nucleus and interact with the NM via shelterin and lamins [43]. Only some telomeres are found at the NE [10]. To replicate telomeres, these topological constraints have to be overcome by detaching telomeres from the NE or NM. This is a potential research topic for the future.
Conclusion
The replication of telomeres, the ends of chromosomes, is a challenging process that requires overcoming several obstacles. These include secondary structures (G4 and T-loops), transcription, and topological constraints due to compaction and anchoring of telomeric chromatin. These factors can cause replication stress and fork stalling at telomeres. Shelterin, a complex of telomere-binding proteins, protects telomeres from replication stress by modulating two distinct pathways. TRF1 prevents fork stalling and ATR activation in S phase [9], while TRF2 resolves supercoiling generated by fork progression [31]. The coordination and regulation of these pathways, as well as the molecular interactions between shelterin and the replisome, are not fully understood and require further investigation. Post-translational modifications of TRF1 and TRF2 [44] may play a key role in this process. Telomere replication also influences telomere length maintenance by telomerase, an enzyme that adds DNA repeats to telomeres. Two models have been proposed to explain how telomerase elongates short telomeres preferentially [45]. Both models involve the association of telomerase with the replication fork and the dissociation of telomerase due to natural barriers at telomeres. Thus telomere replication and elongation are tightly linked processes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- de Lange, T. T-loops and the origin of telomeres. Nature reviews. Molecular cell biology 2004, 5, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Maestroni, L., Matmati, S., & Coulon, S. Solving the Telomere Replication Problem. Genes 2017, 8, 55. [CrossRef] [PubMed]
- de Lange, T. Shelterin: the protein complex that shapes and safeguards human telomeres. Genes & development 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Soudet, J., Jolivet, P., & Teixeira, M. T. Elucidation of the DNA end-replication problem in Saccharomyces cerevisiae. Molecular cell 2014, 53, 954–964. [CrossRef] [PubMed]
- Nandakumar, J. , Bell, C. F., Weidenfeld, I., Zaug, A. J., Leinwand, L. A., & Cech, T. R. The TEL patch of telomere protein TPP1 mediates telomerase recruitment and processivity. Nature 2012, 492, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Chen, L. Y. , Redon, S., & Lingner, J. The human CST complex is a terminator of telomerase activity. Nature 2012, 488, 540–544. [Google Scholar] [CrossRef]
- Herbig, U. , Jobling, W. A., Chen, B. P., Chen, D. J., & Sedivy, J. M. Telomere shortening triggers senescence of human cells through a pathway involving ATM, p53, and p21(CIP1), but not p16(INK4a). Molecular cell 2004, 14, 501–513. [Google Scholar] [CrossRef]
- Miller, K. M. , Rog, O., & Cooper, J. P. Semi-conservative DNA replication through telomeres requires Taz1. Nature 2006, 440, 824–828. [Google Scholar] [CrossRef]
- Sfeir, A. , Kosiyatrakul, S. T., Hockemeyer, D., MacRae, S. L., Karlseder, J., Schildkraut, C. L., & de Lange, T. Mammalian telomeres resemble fragile sites and require TRF1 for efficient replication. Cell 2009, 138, 90–103. [Google Scholar] [CrossRef]
- Arnoult, N. , Schluth-Bolard, C., Letessier, A., Drascovic, I., Bouarich-Bourimi, R., Campisi, J., Kim, S. H., Boussouar, A., Ottaviani, A., Magdinier, F., Gilson, E., & Londoño-Vallejo, A. Replication timing of human telomeres is chromosome arm-specific, influenced by subtelomeric structures and connected to nuclear localization. PLoS genetics 2010, 6, e1000920. [Google Scholar] [CrossRef]
- Chang, Y. T. , Moser, B. A., & Nakamura, T. M. Fission yeast shelterin regulates DNA polymerases and Rad3(ATR) kinase to limit telomere extension. PLoS genetics 2013, 9, e1003936. [Google Scholar] [CrossRef] [PubMed]
- Tarsounas, M. , & Tijsterman, M. Genomes and G-quadruplexes: for better or for worse. Journal of molecular biology 2013, 425, 4782–4789. [Google Scholar] [CrossRef] [PubMed]
- Leman, A. R. , & Noguchi, E. Local and global functions of Timeless and Tipin in replication fork protection. Cell cycle (Georgetown Tex.), 2012, 11, 3945–3955. [Google Scholar] [CrossRef] [PubMed]
- Leman, A. R. , Dheekollu, J., Deng, Z., Lee, S. W., Das, M. M., Lieberman, P. M., & Noguchi, E. Timeless preserves telomere length by promoting efficient DNA replication through human telomeres. Cell cycle (Georgetown Tex.), 2012, 11, 2337–2347. [Google Scholar] [CrossRef]
- Huber, M. D. , Lee, D. C., & Maizels, N. G4 DNA unwinding by BLM and Sgs1p: substrate specificity and substrate-specific inhibition. Nucleic acids research 2002, 30, 3954–3961. [Google Scholar] [CrossRef] [PubMed]
- [16] Arnoult, N. , Saintome, C., Ourliac-Garnier, I., Riou, J. F., & Londoño-Vallejo, A. Human POT1 is required for efficient telomere C-rich strand replication in the absence of WRN. Genes & development 2009, 23, 2915–2924. [Google Scholar] [CrossRef] [PubMed]
- Safa, L. , Gueddouda, N. M., Thiébaut, F., Delagoutte, E., Petruseva, I., Lavrik, O., Mendoza, O., Bourdoncle, A., Alberti, P., Riou, J. F., & Saintomé, C. 5' to 3' Unfolding Directionality of DNA Secondary Structures by Replication Protein A: G-QUADRUPLEXES AND DUPLEXES. The Journal of biological chemistry 2016, 291, 21246–21256. [Google Scholar] [CrossRef]
- Wang, H. , Nora, G. J., Ghodke, H., & Opresko, P. L. Single molecule studies of physiologically relevant telomeric tails reveal POT1 mechanism for promoting G-quadruplex unfolding. The Journal of biological chemistry 2011, 286, 7479–7489. [Google Scholar] [CrossRef] [PubMed]
- Déjardin, J. , & Kingston, R. E. Purification of proteins associated with specific genomic Loci. Cell 2009, 136, 175–186. [Google Scholar] [CrossRef]
- Zimmermann, M. , Kibe, T., Kabir, S., & de Lange, T. TRF1 negotiates TTAGGG repeat-associated replication problems by recruiting the BLM helicase and the TPP1/POT1 repressor of ATR signaling. Genes & development 2014, 28, 2477–2491. [Google Scholar] [CrossRef]
- Vannier, J. B. , Sarek, G., & Boulton, S. J. RTEL1: functions of a disease-associated helicase. Trends in cell biology 2014, 24, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R. , Zhang, J., Bochman, M. L., Zakian, V. A., & Ha, T. Periodic DNA patrolling underlies diverse functions of Pif1 on R-loops and G-rich DNA. eLife 2014, 3, e02190. [Google Scholar] [CrossRef]
- Paeschke, K. , Bochman, M. L., Garcia, P. D., Cejka, P., Friedman, K. L., Kowalczykowski, S. C., & Zakian, V. A. Pif1 family helicases suppress genome instability at G-quadruplex motifs. Nature 2013, 497, 458–462. [Google Scholar] [CrossRef]
- Rodriguez, R. , Miller, K. M., Forment, J. V., Bradshaw, C. R., Nikan, M., Britton, S., Oelschlaegel, T., Xhemalce, B., Balasubramanian, S., & Jackson, S. P. Small-molecule-induced DNA damage identifies alternative DNA structures in human genes. Nature chemical biology 2012, 8, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Wallgren, M. , Mohammad, J. B., Yan, K. P., Pourbozorgi-Langroudi, P., Ebrahimi, M., & Sabouri, N. G-rich telomeric and ribosomal DNA sequences from the fission yeast genome form stable G-quadruplex DNA structures in vitro and are unwound by the Pfh1 DNA helicase. Nucleic acids research 2016, 44, 6213–6231. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C. , Pourmal, S., & Pavletich, N. P. Dna2 nuclease-helicase structure, mechanism and regulation by Rpa. eLife 2015, 4, e09832. [Google Scholar] [CrossRef] [PubMed]
- Sarek, G. , Vannier, J. B., Panier, S., Petrini, J. H. J., & Boulton, S. J. TRF2 recruits RTEL1 to telomeres in S phase to promote t-loop unwinding. Molecular cell 2015, 57, 622–635. [Google Scholar] [CrossRef]
- Vannier, J. B., Sandhu, S., Petalcorin, M. I., Wu, X., Nabi, Z., Ding, H., & Boulton, S. J. RTEL1 is a replisome-associated helicase that promotes telomere and genome-wide replication. Science 2013, 342, 239–242. [CrossRef]
- Ghosh, A. K. , Rossi, M. L., Singh, D. K., Dunn, C., Ramamoorthy, M., Croteau, D. L., Liu, Y., & Bohr, V. A. RECQL4, the protein mutated in Rothmund-Thomson syndrome, functions in telomere maintenance. The Journal of biological chemistry 2012, 287, 196–209. [Google Scholar] [CrossRef]
- Vannier, J. B. , Pavicic-Kaltenbrunner, V., Petalcorin, M. I., Ding, H., & Boulton, S. J. RTEL1 dismantles T loops and counteracts telomeric G4-DNA to maintain telomere integrity. Cell 2012, 149, 795–806. [Google Scholar] [CrossRef]
- Ye, J. , Lenain, C., Bauwens, S., Rizzo, A., Saint-Léger, A., Poulet, A., Benarroch, D., Magdinier, F., Morere, J., Amiard, S., Verhoeyen, E., Britton, S., Calsou, P., Salles, B., Bizard, A., Nadal, M., Salvati, E., Sabatier, L., Wu, Y., Biroccio, A., … Gilson, E. TRF2 and apollo cooperate with topoisomerase 2alpha to protect human telomeres from replicative damage. Cell 2010, 142, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C. M. , Reichenbach, P., Khoriauli, L., Giulotto, E., & Lingner, J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Costantino, L. , & Koshland, D. The Yin and Yang of R-loop biology. Current opinion in cell biology 2015, 34, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Rippe, K. , & Luke, B. TERRA and the state of the telomere. Nature structural & molecular biology 2015, 22, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R. L. , Cox, K. E., Jeitany, M., Wakimoto, H., Bryll, A. R., Ganem, N. J., Bersani, F., Pineda, J. R., Suvà, M. L., Benes, C. H., Haber, D. A., Boussin, F. D., & Zou, L. Alternative lengthening of telomeres renders cancer cells hypersensitive to ATR inhibitors. Science 2015, 347, 273–277. [Google Scholar] [CrossRef]
- Arora, R. , Lee, Y., Wischnewski, H., Brun, C. M., Schwarz, T., & Azzalin, C. M. RNaseH1 regulates TERRA-telomeric DNA hybrids and telomere maintenance in ALT tumour cells. Nature communications 2014, 5, 5220. [Google Scholar] [CrossRef]
- Clynes, D. , Jelinska, C., Xella, B., Ayyub, H., Scott, C., Mitson, M., Taylor, S., Higgs, D. R., & Gibbons, R. J. Suppression of the alternative lengthening of telomere pathway by the chromatin remodelling factor ATRX. Nature communications 2015, 6, 7538. [Google Scholar] [CrossRef]
- Chawla, R. , Redon, S., Raftopoulou, C., Wischnewski, H., Gagos, S., & Azzalin, C. M. Human UPF1 interacts with TPP1 and telomerase and sustains telomere leading-strand replication. The EMBO journal 2011, 30, 4047–4058. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C. M. , & Lingner, J. Telomere functions grounding on TERRA firma. Trends in cell biology 2015, 25, 29–36. [Google Scholar] [CrossRef]
- Teasley, D. C. , Parajuli, S., Nguyen, M., Moore, H. R., Alspach, E., Lock, Y. J., Honaker, Y., Saharia, A., Piwnica-Worms, H., & Stewart, S. A. Flap Endonuclease 1 Limits Telomere Fragility on the Leading Strand. The Journal of biological chemistry 2015, 290, 15133–15145. [Google Scholar] [CrossRef]
- Benarroch-Popivker, D. , Pisano, S., Mendez-Bermudez, A., Lototska, L., Kaur, P., Bauwens, S., Djerbi, N., Latrick, C. M., Fraisier, V., Pei, B., Gay, A., Jaune, E., Foucher, K., Cherfils-Vicini, J., Aeby, E., Miron, S., Londoño-Vallejo, A., Ye, J., Le Du, M. H., Wang, H., … Giraud-Panis, M. J. TRF2-Mediated Control of Telomere DNA Topology as a Mechanism for Chromosome-End Protection. Molecular cell 2016, 61, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Nagai, S. , Heun, P., & Gasser, S. M. Roles for nuclear organization in the maintenance of genome stability. Epigenomics 2010, 2, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Steglich, B. , Strålfors, A., Khorosjutina, O., Persson, J., Smialowska, A., Javerzat, J. P., & Ekwall, K. The Fun30 chromatin remodeler Fft3 controls nuclear organization and chromatin structure of insulators and subtelomeres in fission yeast. PLoS genetics 2015, 11, e1005101. [Google Scholar] [CrossRef] [PubMed]
- Kaminker, P. G., Kim, S. H., Desprez, P. Y., & Campisi, J. A novel form of the telomere-associated protein TIN2 localizes to the nuclear matrix. Cell cycle (Georgetown Tex.) 2009, 8, 931–939. [CrossRef] [PubMed]
- Walker, J. R. , & Zhu, X. D. Post-translational modifications of TRF1 and TRF2 and their roles in telomere maintenance. Mechanisms of ageing and development 2012, 133, 421–434. [Google Scholar] [CrossRef]
- Greider C., W. Regulating telomere length from the inside out: the replication fork model. Genes & development 2016, 30, 1483–1491. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



