Submitted:
20 December 2023
Posted:
21 December 2023
You are already at the latest version
Abstract
Shrews (Soricidae) are common small wild mammals with a significant overlap in their habitats with humans and domestic animals. Currently, viruses from 24 families have been identified in shrews, including Adenoviridae, Arenaviridae, Arteriviridae, Astroviridae, Bornaviridae, Caliciviridae, Circoviridae, Coronaviridae, Filoviridae, Flaviviridae, Hantaviridae, Hepadnaviridae, Hepeviridae, Nairoviridae, Orthoherpesviridae, Paramyxoviridae, Parvoviridae, Phenuiviridae, Picornaviridae, Polyomaviridae, Poxviridae, Rhabdoviridae, Sedoreoviridae, and Spinareoviridae, which pose potential threats to humans and domestic animals. This review compiled fundamental information about shrews and provided a comprehensive summary of the viruses that have been detected in shrews, with the aim to facilitating a deeper understanding of shrews and the diversity and risks of their viruses for researchers and the general public.
Keywords:
shrew
; virus
; diversity
; domestic animal
; human
1. Introduction of Shrews (Soricidae)
Shrews are usually termed long-nosed mice, but they are not rodents. The largest species of shrews, Suncus murinus (Asian house shrews) (Figure 1), attains a length of approximately 15 cm and a weight of around 100 g, while Suncus etruscus (Etruscan shrews) could represent the smallest extant terrestrial mammals, measuring a mere 3.5 cm in length and weighing approximately 1.8 g [1].
In taxonomy, shrews are closer in phylogenetics to hedgehogs and moles than to rodents. They are taxonomically classified within the family Soricidae in the order Eulipotyphla. Currently, Soricidae encompasses three subfamilies, 26 genera, and 376 recognized species (Table 1, Figure 2). The three subfamilies are Crocidurinae (white-toothed shrews), Soricinae (red-toothed shrews), and Myosoricinae (African shrews). These three subfamilies comprise nine genera with 210 species, 14 genera with 148 species, and three genera with 18 species, respectively. The genus Crocidura in the subfamily Crocidurinae is the largest genus in Soricidae and covers 172 species. The genus Sorex is the largest genus in Soricidae and covers 77 species. The common names of some shrew species are listed in Table 2.
It is worth emphasizing that tree shrews, otter shrews, elephant shrews, West Indies shrews, and marsupial shrews are not shrews, because they do not fall within the Soricidae family [2].
Shrews are distributed globally across major tropical and temperate landmasses (Table 1), with notable exceptions in New Guinea, Australia, and New Zealand, where indigenous shrew populations are absent. In South America, shrews are confined to the northern Andes region and were introduced during the Great American Interchange.
In behaviors, shrews have a lifespan of 12 to 30 months, and they exhibit a relatively high metabolic rate, higher than some mammals of similar body sizes. Consequently, shrews have a relatively large food intake. Shrews do not hibernate but can enter a torpid state. During the winter, many shrews undergo significant changes in morphology, with body weight decreasing by 30% to 50%, and both skeletal and organ sizes noticeably shrinking [3]. Shrews are mostly found in cool and humid environments, with many being terrestrial, while some are semi-aquatic or burrowing. They may be active during both day and night or primarily nocturnal. Shrews are typically solitary creatures, coming together only for mating purposes.
In reproduction, female shrews can give birth to as many as 10 litters each year. In tropical regions, they are capable of mating year-round, while in temperate regions, they cease reproduction during the winter. The gestation period for shrews ranges from 17 to 32 days. Female shrews can become pregnant again within one to two days after giving birth and can lactate and nurse their offspring during pregnancy [4].
Shrews possess two characteristics distinct from most mammals. Firstly, certain shrew species secrete venom [5], which contains various compounds. For example, the venom of American short-tailed shrews can kill 200 mice when administered intravenously. Some shrews in China also produce venom. Secondly, similar to bats and toothed whales, some shrews are capable of echolocation [6]. Currently, two genera, namely Sorex (long-tailed shrews) and Blarina (blarina shrews) possess echolocation abilities. Species within these two genera include Eurasian water shrews (Neomys fodiens), northern short-tailed shrews (Blarina brevicauda), and American water shrews (Sorex palustris), among others [7].
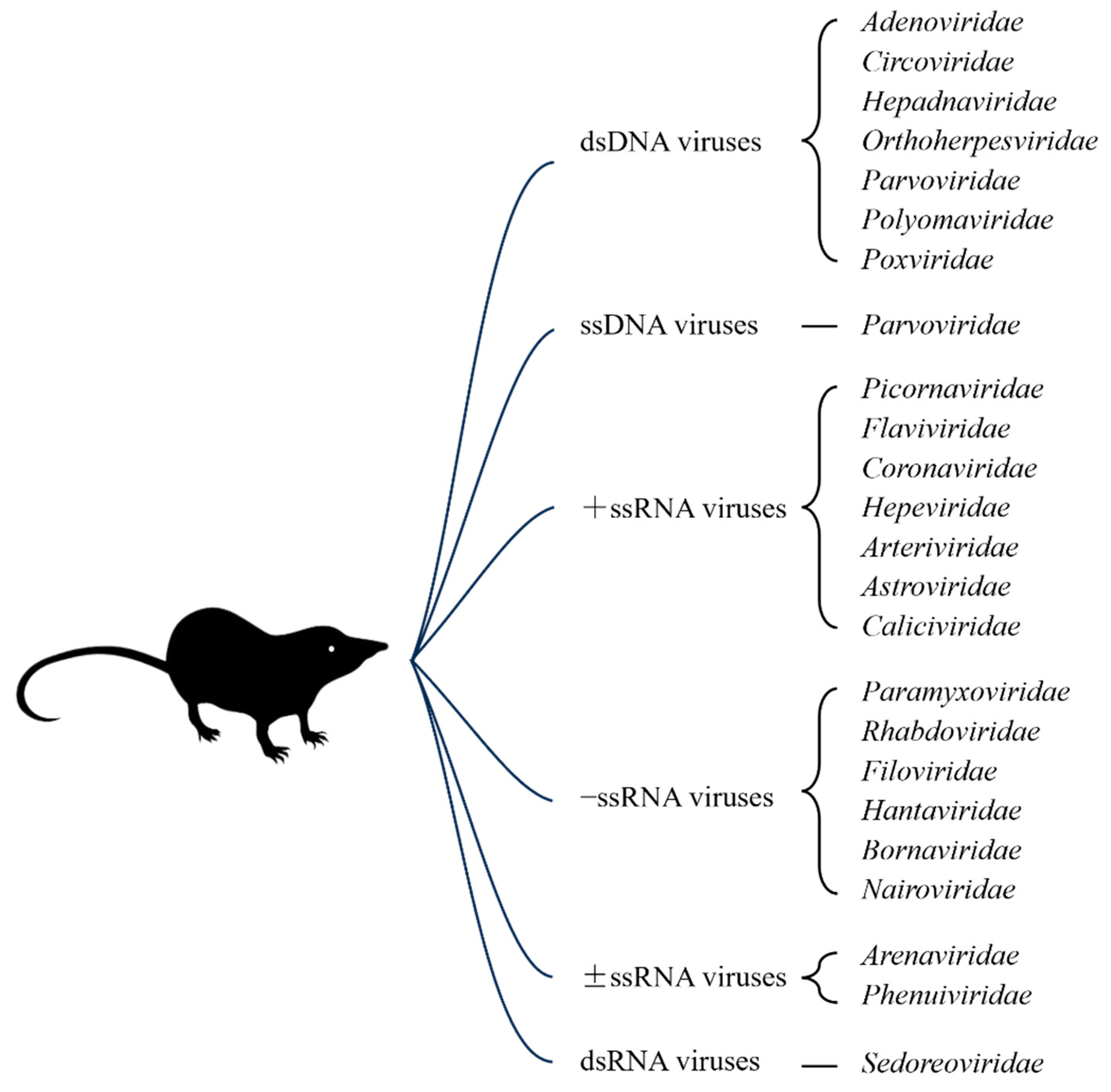
The viruses in shrews have exhibited a high diversity, and many viruses in shrews have not been identified [8]. Currently, as detailed below and summarized in Figure 2 and Table 3, viruses from 24 families have been identified in shrews, including Adenoviridae, Arenaviridae, Arteriviridae, Astroviridae, Bornaviridae, Caliciviridae, Circoviridae, Coronaviridae, Filoviridae, Flaviviridae, Hantaviridae, Hepadnaviridae, Hepeviridae, Nairoviridae, Orthoherpesviridae, Paramyxoviridae, Parvoviridae, Phenuiviridae, Picornaviridae, Polyomaviridae, Poxviridae, Rhabdoviridae, Sedoreoviridae, and Spinareoviridae.
Figure 3.
The virus families that contain viruses having been identified in shrews.
2. Paramyxoviruses in Shrews
Paramyxoviridae family contains several important viruses that are pathogenic to both humans and livestock. Viruses within this family are enveloped, approximately spherical or pleomorphic in shape, with a diameter of around 150 nm or larger. The genomes of Paramyxoviridae viruses consist of single-stranded, negative-sense RNA that is non-segmented, with a length of approximately 15 to 21 kb. These genomes encode six to ten different proteins, including the nucleocapsid protein (NP), phosphoprotein (P), large polymerase protein (L), matrix protein (M), fusion protein (F), and hemagglutinin-neuraminidase protein (HN) [9].
The International Committee on Taxonomy of Viruses (ICTV) currently classifies the Paramyxoviridae into 17 genera. The Paraavulavirus genus contains two species of avian viruses. The Henipavirus genus comprises 10 mammalian viruses, including the pathogenic Hendra virus, Nipah virus, and Langya virus. The Jeilongvirus genus includes 15 mammalian viruses, such as J virus and Beilong virus. Currently, seven species of paramyxoviruses have been identified in shrews, with six belonging to the Henipavirus genus and one belonging to the Jeilongvirus genus.
Langya virus, a member of the Henipavirus, was first reported in China in 2022, with shrews potentially serving as its natural reservoir host, detected in two shrew species, Crocidura lasiura (Ussuri white-toothed shrews) and Crocidura shantungensis (Asian lesser white-toothed shrews). The study identified 35 acute Langya virus cases among patients in Shandong and Henan provinces, manifesting symptoms such as fever (100%), fatigue (54%), cough (50%), loss of appetite (50%), myalgia (46%), nausea (38%), headache (35%), vomiting (35%), thrombocytopenia (35%), leukopenia (54%), and liver (35%) and kidney dysfunction (8%). Research encompassed 25 wild animal species and indicated a high virus infection rate among shrews (71/262 [27%]). Nevertheless, the virus's distribution in shrews and other animals across diverse Chinese regions remains poorly elucidated [10].
Gamak virus and Daeryong virus, both belonging to the Henipavirus, which were discovered in South Korea in 2021 from Ussuri white-toothed shrews and Asian lesser white-toothed shrews, respectively. Gamak virus has the ability to infect and replicate in human lung epithelial cell line A549 cells and rapidly induce the secretion of type I/III interferons, ISGs, and pro-inflammatory cytokines from these cells [11].
The viruses Melian virus and Denwin virus, belonging to the Henipavirus, were respectively discovered in 2022 in large-headed shrews (Crocidura grandiceps) in Guinea and greater white-toothed shrews (Crocidura russula) in Belgium. Further studies on their pathogenicity have not been conducted yet [12].
Ninorex virus, belonging to the Henipavirus, were respectively discovered in 2023 in Sorex minutus in Belgian. Further studies on their pathogenicity have not been conducted yet [13].
Beilong virus, belonging to the Jeilongvirus, has also been detected in shrews, although it was initially found in rodents. A study conducted between September 2013 and May 2019, covering eleven provinces in China, examined small mammals. The PCR testing showed a positivity rate of 28.57% (2/7) in the Asian lesser white-toothed shrews and 17.57% (13/74) in the Suncus murinus. Currently, there is no evidence of human infection with Beilong virus, but its potential risk to humans and livestock should not be underestimated [14].
3. Hantaviruses in Shrews
Hantaviridae family includes many pathogenic viruses for both humans and animals. These viruses are enveloped and are typically spherical or tubular, with a diameter ranging from 120 to 160 nm. The viral genome is a single-stranded negative-sense RNA virus (−ssRNA) with a size ranging from approximately 10,000 to 15,000 nucleotides. It consists of three segments: large (L), medium (M), and small (S), which collectively encode at least four structural proteins [15]. The largest segment (L) of the viral genome encodes an RNA-dependent RNA polymerase (RdRp). The medium segment (M) encodes a glycoprotein precursor (GPC), which is co-translationally cleaved by host cell signal peptidases in the endoplasmic reticulum to produce two envelope glycoproteins (Gn and Gc, or G1 and G2). The small segment (S) encodes the nucleocapsid protein (N), responsible for assembling the viral genomic RNA [16].
Hantaviridae is currently classified into seven genera. The Orthohantavirus genus has 38 mammalian viruses, including 15 that are zoonotic. The Mobatvirus genus contains 5 animal viruses. The Thottimvirus genus contains 2 mammalian viruses.
Lena mobatvirus, belonging to the Mobatvirus, was discovered in 2021 in the Sorex caecutiens and Sorex roboratus in the Siberian and Far Eastern regions of Russia. Its zoonotic potential has not been analyzed [17].
Asikkala virus, belonging to the Orthohantavirus, was discovered in 2010 in Eurasian pygmy shrews (Sorex minutus) in Finland. Subsequently, partial L segment sequences of three new hantaviruses were obtained in the Czech Republic and Germany. Genetic evolutionary analysis indicated that all of these hantaviruses belong to the Asikkala virus group, originally identified in Finland [18].
Bowé virus, belonging to the Orthohantavirus, was discovered in 2012 in the muscle tissue of the Doucet's musk shrews (Crocidura douceti) in southwestern Guinea. It shares a common ancestor with the unclassified Tanganya virus, and its zoonotic potential is currently unknown [19].
Cao Bang virus, a member of the Orthohantavirus, was discovered in 2006 in the lung tissue of the Chinese mole shrews (Anourosorex squamipes) in China. Sequencing of the M and partial S segments revealed that it is genetically distantly related to hantaviruses carried by rodents and other mole shrews hantaviruses. Its zoonotic potential is currently unknown [20].
Imjin virus, a member of the Orthohantavirus, was discovered in 2009 in Ussuri white-toothed shrews in South Korea. Its zoonotic potential is currently unknown [21].
Jeju virus, a member of the Orthohantavirus, was discovered in 2012 in Ussuri white-toothed shrews on Jeju Island, South Korea. Genetic evolution analysis revealed that it is distantly related to the previously discovered Imjin virus in South Korea, making it a new member of the Orthohantavirus. Its zoonotic potential is currently unknown [22].
Kenkeme virus, a member of the Orthohantavirus, was discovered in 2010 on Sorex roboratus in the northeastern part of the Sakha Republic, Siberia. Sequence analysis of the full-length S segment and partial M and L segments indicated that Kenkeme virus represents a novel Orthohantavirus in terms of its genetic makeup and phylogenetic relationship. Kenkeme virus exhibits distinct differences in terms of genetics and phylogeny when compared to other hantaviruses found in Eurasian common shrews, as well as in other rodents, shrews, and moles. Its zoonotic potential is currently unknown [23].
Seewis virus, a member of the Orthohantavirus, was discovered in 2006 in Eurasian common shrews (Sorex araneus) in Switzerland. Phylogenetic analysis of the complete S segment and partial M and L segments of Seewis virus revealed that it is genetically distant from hantaviruses found in rodents but more closely related to hantaviruses found in shrews. The zoonotic potential of Seewis virus is currently unknown [24].
Yakeshi virus, a member of the Orthohantavirus, was discovered in 2013 on long-clawed shrews (Sorex unguiculatus) in China. Lianghe virus, an unclassified virus within the Orthohantavirus, was found in Chinese mole shrews (Anourosorex squamipes). The zoonotic potential of these viruses is currently unknown [25].
Seoul virus, a member of the Orthohantavirus, was first reported in 2014 in Ussuri white-toothed shrews in China. This virus is known to be infectious to humans [26].
Altai virus, an unclassified member of the Orthohantavirus, was amplified from the tissue of an Eurasian common shrew in the Altai Republic in 2007. Initially, this virus was classified as Seewis virus, but subsequent findings indicated that it did not align with the host shrew species and geographical location. Through nucleotide pairing comparisons and analysis of a 300-nucleotide region of the L segment, it was confirmed to be a unique Hantavirus species [27].
Artybash virus, an unclassified member of the Orthohantavirus, was detected in a very limited manner in the Sorex caecutiens captured near Lake Teletskoye in the Altai Republic of Western Siberia in 2006. Initially referred to as the Amga virus, genetic and phylogenetic analysis of both partial and complete genome sequences identified it as a genetic variant of the Artybash virus. Its zoonotic potential remains unknown [28].
Xinyi virus, an unclassified member of the Orthohantavirus, was reported in 2016 and discovered in the Anourosorex yamashinai. Phylogenetic analysis based on the S segment, M segment, and L segment sequences suggests that Xinyi virus shares a common ancestor with the Cao Bang virus. Their pathogenicity to humans and animals is currently unknown [29].
The unclassified Ash River virus and Jemez Springs virus, both members of the Orthohantavirus, were discovered in 2008 on masked shrews (Sorex cinereus) and dusky shrews (Sorex monticolus) in the United States. Their pathogenicity to humans and animals is currently unknown [30].
The unclassified Camp Ripley virus, a member of the Orthohantavirus, was discovered in 2007 in the northern short-tailed shrews in the United States. Its pathogenicity to humans and animals is currently unknown [31].
The unclassified Boginia virus, a member of the Orthohantavirus, was reported in 2013 in Eurasian water shrews in Poland. Its pathogenicity to humans and animals is currently unknown [32].
The unclassified Azagny virus, a member of the Orthohantavirus, was discovered in 2011 in West African pygmy shrews (Crocidura obscurior) in Azagny Park, Côte d'Ivoire. Its pathogenicity to humans and animals is currently unknown [33].
The unclassified Puumala virus, a member of the Orthohantavirus genus, was discovered in 2014 in Eurasian common shrews in Finland. Its pathogenicity to humans and animals is currently unknown [34].
The unclassified Kilimanjaro virus and Uluguru virus, members of the Orthohantavirus, were discovered in 2014 in the Myosorex geata and Myosorex zinki in West Africa. Their pathogenicity to humans and animals is currently unknown [35].
The unclassified Qian hu shan virus, a member of the Orthohantavirus, was detected in the lung tissue of stripe-backed shrews (Sorex cylindricauda) in China in 2014. Its pathogenicity to humans and animals is currently unknown [36].
Thottapalayam virus (TPMV), a member of the Thottimvirus, was first isolated in 1965 from Asian house shrews in India. This virus represents the earliest isolation of a hantavirus from shrews [37].
4. Sedoreoviruses in Shrews
The viruses in Sedoreoviridae family include numerous pathogens for humans and animals. These viruses lack an envelope, have an icosahedral shape, and measure approximately 60 to 100 nm in diameter [38]. The genome of Sedoreoviridae consists of 10 to 12 linear double-stranded RNA segments, with a total length ranging from 18 to 26 kilobase pairs (kp). Individual segments vary in length from 0.6 to 5.8 kp. Each double-stranded positive-sense RNA carries a type 1 cap structure at the 5' end but lacks a 3' polyadenylated tail. Virus RNAs are typically monogenic and feature relatively short 5' and 3' non-coding regions, although some segments may contain a second or third functional open reading frame [39].
Sedoreoviridae is currently classified into six genera. The Orbivirus genus contains a total of 22 animal viruses, including 16 mammalian viruses such as Bluetongue virus; the Rotavirus genus contains 9 rotaviruses, including 3 that are pathogenic to humans, 3 to birds, and 3 to other mammals.
Rotavirus A, belonging to the Rotavirus, was detected in Asian house shrews in 2016. This virus can infect humans, causing severe diarrhea in infants and young children. It can also infect domestic and wild mammals as well as birds [40].
Bluetongue virus, a member of the Orbivirus, was discovered in an unknown shrew species in Africa in 1994. This virus primarily infects sheep and various ruminant animals, with a very high fatality rate in sheep [41].
5. Parvoviruses in Shrews
The Parvoviridae family contains several significant viruses that can cause diseases in humans and animals. Viruses in this family are non-enveloped and typically have small, symmetrical icosahedral shapes with a diameter of approximately 23 to 28 nm [42]. The genome of parvoviruses consists of a linear, non-circular, single-stranded DNA molecule with a length ranging from 4 to 6 kb. The capsid is composed of sixty VP proteins that assemble into a nested arrangement, usually encoded by a single structural gene. This gene includes the entire coding sequence for VP1, which typically ranges from 75 to 100 kDa. Additionally, there are one or more smaller forms (VP2-5) that share a common C-terminal sequence but have varying lengths of N-terminal truncation [43].
Parvoviridae is currently classified into 14 genera. The Bocaparvovirus genus encompasses 31 mammalian viruses. The Dependoparvovirus genus comprises 13 animal viruses, including 3 avian viruses and 8 mammalian viruses. The Protoparvovirus genus includes 18 mammalian viruses, such as canine parvovirus, porcine parvovirus, and 3 human parvoviruses.
Bufavirus, a member of the Protoparvovirus, was discovered in the lesser red musk shrews (Crocidura hirta) in Zambia in 2015. This virus is capable of infecting humans, primarily causing diarrhea in children [44].
Porcine bocavirus G4, a member of the Bocaparvovirus, was discovered in Asian house shrews in 2019 in China. This virus can infect pigs, leading to respiratory and gastrointestinal diseases [45].
Adeno-associated virus (AAV), a member of the Dependoparvovirus, was discovered in Asian house shrews in 2020 in China. The Infection status in humans and other animals is unknown [46].
6. Nairoviruses in Shrews
The Nairoviridae family includes various significant viruses that are harmful to both humans and animals. Viruses in this family are enveloped, spherical in shape, with a diameter of 80-120 nm. The envelope is covered with glycoproteins Gn and Gc. The isolated ribonucleoprotein (RNP) complex consists of segmented genomic RNA, each wrapped in nucleoprotein (N) [47]. The Nairovirus genome comprises three single-stranded negative-sense RNA molecules, with these three genomic segments labeled as L (large), M (medium), and S (small). These RNAs code for proteins on the viral complementary strands, including N, glycoprotein precursor (GPC), and L (containing RdRp, helicase, and nuclease domains) [48].
Nairoviridae is currently classified into three genera. The Orthonairovirus genus comprises 55 animal viruses.
The Orthonairovirus Lamusara virus and Lamgora virus were discovered in Crocidura sp. in Gabon, Central Africa, in 2022. Their pathogenicity to humans and animals is currently unknown [49].
Erve virus, belonging to the Orthonairovirus, was discovered in greater white-toothed shrews in France in 1989. It can infect humans and various domestic animals, leading to neurological symptoms in humans [50].
Thiafora virus, belonging to the Orthonairovirus, was discovered in Crocidura sp. in Senegal in 2015. Its pathogenicity to humans and animals is currently unknown [51].
Cencurut virus, belonging to the Orthonairovirus, was discovered in Asian house shrews in Singapore in 2023. Its pathogenicity to humans and animals is currently unknown [52].
8. Flaviviruses in Shrews
The Flaviviridae family comprises several significant viruses that are harmful to humans and animals. Viruses in this family are enveloped and spherical, with a viral particle diameter ranging from 40 to 60 nm [58]. The viral genome is a positive-sense, single-stranded RNA, with a length of 8.9 to 13.0 kb. It consists of a single large open reading frame (ORF) flanked by 3' and 5' untranslated regions (UTRs), and it possesses a 5' cap structure. This single ORF encodes a polyprotein that undergoes processing and cleavage by viral protease NS2B/NS3 and cellular proteases. This cleavage results in the formation of three structural proteins, including the C protein of the viral capsid, the M (membrane) protein, and the E (membrane) protein. Additionally, seven non-structural proteins play essential roles in viral replication. These proteins are NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5 [59].
Flaviviridae is currently classified into four genera. The Orthoflavivirus genus includes 52 animal viruses, including several significant human pathogens such as Yellow Fever virus, Dengue virus, Zika virus, Japanese Encephalitis virus, West Nile virus, and Tick-Borne Encephalitis virus. It also has pathogenicity for livestock and poultry (turkeys, pigs, horses, and sheep), causing economically important diseases such as encephalitis viruses in various livestock. The Hepacivirus genus contains 14 mammalian viruses, with human hepatitis C virus being among those capable of infecting humans. The Pegivirus genus comprises 11 mammalian viruses.
Usutu virus, belonging to the Orthoflavivirus, was first reported in Crocidura sp. in Senegal in 2019. At the time of collection, no pathogenic infection or symptoms were observed in shrews. This virus was initially discovered in 1959 and is associated with Culex mosquitoes as its vectors. Currently, the virus has been isolated from birds, arthropods, and humans in Europe and Africa, and subsequently detected in bats and horses [60].
Powassan virus type 2, classified within the Orthoflavivirus, was first discovered in 2021 in Blarina brevicauda in the United States. Previously, it had been found primarily in ticks and humans. This virus can lead to encephalitis in humans and is primarily transmitted by arthropod vectors [61].
Tick-borne encephalitis virus, classified within the Orthoflavivirus, was first discovered in 1967 in Eurasian common shrews in Slovakia. This virus can infect humans, and when it reaches the central nervous system, neurological symptoms can occur. Infection can manifest as meningitis, encephalitis, or meningoencephalitis. In nature, tick-borne encephalitis virus circulates between ticks and small mammals [62].
Japanese encephalitis virus, classified within the Orthoflavivirus, was first discovered to infect Asian house shrews in 1984. This virus can infect humans and various livestock. Infected individuals may exhibit mild symptoms such as fever and headache, or they may be asymptomatic. However, approximately 1 in every 250 infected individuals may develop severe illness. Initial symptoms in children can include abdominal pain and vomiting. Severe illness is characterized by a rapid rise in body temperature, headache, neck stiffness, loss of orientation, coma, seizures, spasmodic paralysis, and may lead to death. The fatality rate for individuals displaying disease symptoms can be as high as 30% [63].
Suncus murinus hepacivirus, belonging to the Hepacivirus, was discovered in Asian house shrews in 2016. Currently, its zoonotic potential is unknown [64].
Rat pegivirus, belonging to the Pegivirus, was discovered in Chinese Asian house shrews in 2020. This virus primarily targets lymphocytes and causes asymptomatic infections in humans and other animals, presenting as benign [65].
9. Arteriviruses in Shrews
The Arteriviridae family comprises several significant veterinary pathogens. The viruses in this family are enveloped and generally spherical, with an estimated diameter ranging from 50 to 74 nm [66]. The viral genome is a linear positive-sense RNA, approximately 12 to 16 kb in length. The infectious genome RNA contains a 5' type I cap structure and a 3' polyadenylate sequence, surrounding multiple and mostly overlapping open reading frames (ORFs). The two largest ORFs closest to the 5' end, known as ORF1a and ORF1b, encode non-structural proteins, while the ORFs at the 3' end encode structural viral proteins.
Arteriviridae is currently classified into 13 genera. The Muarterivirus genus contains only one shrew arterivirus, Olivier's shrew virus 1, which was discovered in 2018 in Olivier's shrews (Crocidura olivieri guineensis) in Guinea. Both phylogenetic analysis and pairwise sequence comparison indicate that this virus is distinctly different from the established arterivirus genera and belongs to a new genus [67].
10. Other Viruses in Shrews
Shrews serve as the hosts for multiple other viruses, such as adenovirus, hepatitis B virus, herpesvirus, Ebola virus, pegivirus, and astrovirus, as listed in Table 2.
11. Discussion
Shrews share overlapping habitats with humans and livestock, highlighting the importance of shrew virus research for public health and animal welfare. This review shows that shrew populations host at least 24 families of viruses, and 23 species of hantaviruses, seven species of paramyxoviruses, six species of flaviviruses, and five species of nairoviruses in shrews have been identified. 17 species of viruses in 14 families in Asian house shrews (Suncus murinus) have been identified. Eight species of viruses in six families in Eurasian common shrews (Sorex araneus) have been identified. Six species of viruses in two zoonotic families in Ussuri white-toothed shrews (Crocidura lasiura) have been identified. These three species of shrews could host more species of zoonotic viruses than other shrew species.
Future research should assess the transmission risks of shrew viruses from shrews to humans and domestic animals. Future research should conduct broader surveys to delve into the diversity, evolution, potential pathogenicity, and effective measures of these viruses. This will enhance our capacity to respond swiftly to the potential outbreaks of shrew viruses in humans or domestic animals.
Shrews' small size and ease of husbandry make them ideal for studying viruses of shrews and other animals including humans. Their biological traits, like reproductive cycles, lifespan, and immune systems, are valuable for investigating the transmission, pathogenicity, and potential therapies of viral diseases.
Author Contributions
H.-Y.G., R.-X.C., S.-M.T., and X.W. wrote the first version of this manuscript under the direction of J.-M.C., Y.-L.Z., M.L. and H.-Y.G. drew the tables and figures. J.-M.C., Y.-L.Z., and M.L. conceived, designed, and financially supported this study and revised this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by by the High-Level Talent Fund of Foshan University [No. 20210036] and the Open Competition Program of Top Ten Critical Priorities of Agricultural Science and Technology Innovation for the 14th Five-Year Plan of Guangdong Province [2022SDZG02]. The APC was funded by the second funder.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author Ji-Ming Chen upon reasonable request.
Acknowledgments
This study was supported by the High-Level Talent Fund of Foshan University (No. 20210036) and the Open Competition Program of Top Ten Critical Priorities of Agricultural Science and Technology Innovation for the 14th Five-Year Plan of Guangdong Province (2023SDZG02).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jürgens, K. D. , Etruscan shrew muscle: The consequences of being small. J Exp Biol 2002, 205 Pt 15, 2161–2166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z. , Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness (Addenda 2013). Zootaxa 2013, 3703, 1–82. [Google Scholar] [CrossRef] [PubMed]
- Chung, D. J.; Madison, G. P.; Aponte, A. M.; Singh, K.; Li, Y.; Pirooznia, M.; Bleck, C. K. E.; Darmani, N. A.; Balaban, R. S. , Metabolic design in a mammalian model of extreme metabolism, the North American least shrew (Cryptotis parva). J Physiol 2022, 600, 547–567. [Google Scholar] [CrossRef] [PubMed]
- Bedford, J. M.; Mock, O. B.; Goodman, S. M. , Novelties of conception in insectivorous mammals (Lipotyphla), particularly shrews. Biol Rev Camb Philos Soc 2004, 79, 891–909. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, K.; Rychlik, L. , Venom Use in Eulipotyphlans: An Evolutionary and Ecological Approach. Toxins (Basel) 2021, 13, (3). [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Tian, R.; Rong, X.; Li, G.; Chen, B.; Ren, W.; Xu, S.; Yang, G. , Evidence of Echolocation in the Common Shrew from Molecular Convergence with Other Echolocating Mammals. Zool Stud 2020, 59, e4. [Google Scholar] [PubMed]
- Siemers, B. M.; Schauermann, G.; Turni, H.; von Merten, S. , Why do shrews twitter? Communication or simple echo-based orientation. Biol Lett 2009, 5, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y. M.; Hu, S. J.; Lin, X. D.; Tian, J. H.; Lv, J. X.; Wang, M. R.; Luo, X. Q.; Pei, Y. Y.; Hu, R. X.; Song, Z. G.; Holmes, E. C.; Zhang, Y. Z. , Host traits shape virome composition and virus transmission in wild small mammals. Cell 2023, 186, 4662–4675. [Google Scholar] [CrossRef] [PubMed]
- Thibault, P. A.; Watkinson, R. E.; Moreira-Soto, A.; Drexler, J. F.; Lee, B. , Zoonotic Potential of Emerging Paramyxoviruses: Knowns and Unknowns. Adv Virus Res 2017, 98, 1–55. [Google Scholar] [PubMed]
- Zhang, X. A.; Li, H.; Jiang, F. C.; Zhu, F.; Zhang, Y. F.; Chen, J. J.; Tan, C. W.; Anderson, D. E.; Fan, H.; Dong, L. Y.; Li, C.; Zhang, P. H.; Li, Y.; Ding, H.; Fang, L. Q.; Wang, L. F.; Liu, W. , A Zoonotic Henipavirus in Febrile Patients in China. N Engl J Med 2022, 387, 470–472. [Google Scholar] [CrossRef]
- Lee, S. H.; Kim, K.; Kim, J.; No, J. S.; Park, K.; Budhathoki, S.; Lee, S. H.; Lee, J.; Cho, S. H.; Cho, S.; Lee, G. Y.; Hwang, J.; Kim, H. C.; Klein, T. A.; Uhm, C. S.; Kim, W. K.; Song, J. W. , Discovery and Genetic Characterization of Novel Paramyxoviruses Related to the Genus Henipavirus in Crocidura Species in the Republic of Korea. Viruses 2021, 13, (10). [Google Scholar] [CrossRef] [PubMed]
- Vanmechelen, B.; Meurs, S.; Horemans, M.; Loosen, A.; Joly Maes, T.; Laenen, L.; Vergote, V.; Koundouno, F. R.; Magassouba, N.; Konde, M. K.; Condé, I. S.; Carroll, M. W.; Maes, P. , The characterization of multiple novel paramyxoviruses highlights the diverse nature of the subfamily Orthoparamyxovirinae. Virus Evol 2022, 8, veac061. [Google Scholar] [CrossRef] [PubMed]
- Horemans, M.; Van Bets, J.; Joly Maes, T.; Maes, P.; Vanmechelen, B. , Discovery and genome characterization of six new orthoparamyxoviruses in small Belgian mammals. Virus Evol 2023, 9, vead065. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. J.; Zhang, X. A.; Fan, H.; Jiang, F. C.; Jin, M. Z.; Dai, K.; Wang, N.; Zhang, P. H.; Li, X. K.; Li, H.; Shi, W.; Yang, Z. C.; Fang, L. Q.; Zhou, H. S.; Wei, Y. H.; Liu, W. , Distribution and characteristics of Beilong virus among wild rodents and shrews in China. Infect Genet Evol 2020, 85, 104454. [Google Scholar] [CrossRef] [PubMed]
- Parvate, A.; Williams, E. P.; Taylor, M. K.; Chu, Y. K.; Lanman, J.; Saphire, E. O.; Jonsson, C. B. , Diverse Morphology and Structural Features of Old and New World Hantaviruses. Viruses 2019, 11, (9). [Google Scholar] [CrossRef] [PubMed]
- Muyangwa, M.; Martynova, E. V.; Khaiboullina, S. F.; Morzunov, S. P.; Rizvanov, A. A. , Hantaviral Proteins: Structure, Functions, and Role in Hantavirus Infection. Front Microbiol 2015, 6, 1326. [Google Scholar] [CrossRef] [PubMed]
- Yashina, L. N.; Abramov, S. A.; Zhigalin, A. V.; Smetannikova, N. A.; Dupal, T. A.; Krivopalov, A. V.; Kikuchi, F.; Senoo, K.; Arai, S.; Mizutani, T.; Suzuki, M.; Cook, J. A.; Yanagihara, R. , Geographic Distribution and Phylogeny of Soricine Shrew-Borne Seewis Virus and Altai Virus in Russia. Viruses 2021, 13, (7). [Google Scholar] [CrossRef] [PubMed]
- Radosa, L.; Schlegel, M.; Gebauer, P.; Ansorge, H.; Heroldová, M.; Jánová, E.; Stanko, M.; Mošanský, L.; Fričová, J.; Pejčoch, M.; Suchomel, J.; Purchart, L.; Groschup, M. H.; Krüger, D. H.; Ulrich, R. G.; Klempa, B. , Detection of shrew-borne hantavirus in Eurasian pygmy shrew (Sorex minutus) in Central Europe. Infect Genet Evol 2013, 19, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Gu, S. H.; Nicolas, V.; Lalis, A.; Sathirapongsasuti, N.; Yanagihara, R. , Complete genome sequence and molecular phylogeny of a newfound hantavirus harbored by the Doucet's musk shrew (Crocidura douceti) in Guinea. Infect Genet Evol 2013, 20, 118–123. [Google Scholar] [CrossRef]
- Song, J. W.; Kang, H. J.; Song, K. J.; Truong, T. T.; Bennett, S. N.; Arai, S.; Truong, N. U.; Yanagihara, R. , Newfound hantavirus in Chinese mole shrew, Vietnam. Emerg Infect Dis 2007, 13, 1784–1787. [Google Scholar] [CrossRef]
- Song, J. W.; Kang, H. J.; Gu, S. H.; Moon, S. S.; Bennett, S. N.; Song, K. J.; Baek, L. J.; Kim, H. C.; O'Guinn, M. L.; Chong, S. T.; Klein, T. A.; Yanagihara, R. , Characterization of Imjin virus, a newly isolated hantavirus from the Ussuri white-toothed shrew (Crocidura lasiura). J Virol 2009, 83, 6184–6191. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Gu, S. H.; Baek, L. J.; Tabara, K.; Bennett, S. N.; Oh, H. S.; Takada, N.; Kang, H. J.; Tanaka-Taya, K.; Morikawa, S.; Okabe, N.; Yanagihara, R.; Song, J. W. , Divergent ancestral lineages of newfound hantaviruses harbored by phylogenetically related crocidurine shrew species in Korea. Virology 2012, 424, 99–105. [Google Scholar] [CrossRef]
- Kang, H. J.; Arai, S.; Hope, A. G.; Cook, J. A.; Yanagihara, R. , Novel hantavirus in the flat-skulled shrew (Sorex roboratus). Vector Borne Zoonotic Dis 2010, 10, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Song, J. W.; Gu, S. H.; Bennett, S. N.; Arai, S.; Puorger, M.; Hilbe, M.; Yanagihara, R. , Seewis virus, a genetically distinct hantavirus in the Eurasian common shrew (Sorex araneus). Virol J 2007, 4, 114. [Google Scholar] [CrossRef]
- Guo, W. P.; Lin, X. D.; Wang, W.; Tian, J. H.; Cong, M. L.; Zhang, H. L.; Wang, M. R.; Zhou, R. H.; Wang, J. B.; Li, M. H.; Xu, J.; Holmes, E. C.; Zhang, Y. Z. , Phylogeny and origins of hantaviruses harbored by bats, insectivores, and rodents. PLoS Pathog 2013, 9, e1003159. [Google Scholar] [CrossRef]
- Sun, X. F.; Zhao, L.; Zhang, Z. T.; Liu, M. M.; Xue, Z. F.; Wen, H. L.; Ma, D. Q.; Huang, Y. T.; Sun, Y.; Zhou, C. M.; Luo, L. M.; Liu, J. W.; Li, W. Q.; Yu, H.; Yu, X. J. , Detection of Imjin Virus and Seoul Virus in Crocidurine Shrews in Shandong Province, China. Vector Borne Zoonotic Dis 2017, 17, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Kang, H. J.; Gu, S. H.; Yashina, L. N.; Cook, J. A.; Yanagihara, R. , Highly Divergent Genetic Variants of Soricid-Borne Altai Virus (Hantaviridae) in Eurasia Suggest Ancient Host-Switching Events. Viruses 2019, 11, (9). [Google Scholar] [CrossRef]
- Arai, S.; Kang, H. J.; Gu, S. H.; Ohdachi, S. D.; Cook, J. A.; Yashina, L. N.; Tanaka-Taya, K.; Abramov, S. A.; Morikawa, S.; Okabe, N.; Oishi, K.; Yanagihara, R. , Genetic Diversity of Artybash Virus in the Laxmann's Shrew (Sorex caecutiens). Vector Borne Zoonotic Dis 2016, 16, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Gu, S. H.; Arai, S.; Yu, H. T.; Lim, B. K.; Kang, H. J.; Yanagihara, R. , Genetic variants of Cao Bang hantavirus in the Chinese mole shrew (Anourosorex squamipes) and Taiwanese mole shrew (Anourosorex yamashinai). Infect Genet Evol 2016, 40, 113–118. [Google Scholar] [CrossRef]
- Arai, S.; Bennett, S. N.; Sumibcay, L.; Cook, J. A.; Song, J. W.; Hope, A.; Parmenter, C.; Nerurkar, V. R.; Yates, T. L.; Yanagihara, R. , Phylogenetically distinct hantaviruses in the masked shrew (Sorex cinereus) and dusky shrew (Sorex monticolus) in the United States. Am J Trop Med Hyg 2008, 78, 348–351. [Google Scholar] [CrossRef]
- Arai, S.; Song, J. W.; Sumibcay, L.; Bennett, S. N.; Nerurkar, V. R.; Parmenter, C.; Cook, J. A.; Yates, T. L.; Yanagihara, R. , Hantavirus in northern short-tailed shrew, United States. Emerg Infect Dis 2007, 13, 1420–1423. [Google Scholar] [CrossRef] [PubMed]
- Gu, S. H.; Markowski, J.; Kang, H. J.; Hejduk, J.; Sikorska, B.; Liberski, P. P.; Yanagihara, R. , Boginia virus, a newfound hantavirus harbored by the Eurasian water shrew (Neomys fodiens) in Poland. Virol J 2013, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Kang, H. J.; Kadjo, B.; Dubey, S.; Jacquet, F.; Yanagihara, R. , Molecular evolution of Azagny virus, a newfound hantavirus harbored by the West African pygmy shrew (Crocidura obscurior) in Côte d'Ivoire. Virol J 2011, 8, 373. [Google Scholar] [CrossRef]
- Ling, J.; Sironen, T.; Voutilainen, L.; Hepojoki, S.; Niemimaa, J.; Isoviita, V. M.; Vaheri, A.; Henttonen, H.; Vapalahti, O. , Hantaviruses in Finnish soricomorphs: Evidence for two distinct hantaviruses carried by Sorex araneus suggesting ancient host-switch. Infect Genet Evol 2014, 27, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Kang, H. J.; Stanley, W. T.; Esselstyn, J. A.; Gu, S. H.; Yanagihara, R. , Expanded host diversity and geographic distribution of hantaviruses in sub-Saharan Africa. J Virol 2014, 88, 7663–7667. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S. Q.; Gong, Z. D.; Fang, L. Q.; Jiang, J. F.; Zhang, J. S.; Zhao, Q. M.; Cao, W. C. , A new hantavirus from the stripe-backed shrew (Sorex cylindricauda) in the People's Republic of China. Virus Res 2014, 184, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Carey, D. E.; Reuben, R.; Panicker, K. N.; Shope, R. E.; Myers, R. M. , Thottapalayam virus: A presumptive arbovirus isolated from a shrew in India. Indian J Med Res 1971, 59, 1758–1760. [Google Scholar] [PubMed]
- Jaafar, F. M.; Attoui, H.; Mertens, P. P. C.; de Micco, P.; de Lamballerie, X. , Structural organization of an encephalitic human isolate of Banna virus (genus Seadornavirus, family Reoviridae). J Gen Virol 2005, 86 Pt 4, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A. E.; Grimes, J. M.; Mertens, P. P.; Attoui, H. , Detection of a fourth orbivirus non-structural protein. PLoS ONE 2011, 6, e25697. [Google Scholar] [CrossRef]
- Li, K.; Lin, X. D.; Huang, K. Y.; Zhang, B.; Shi, M.; Guo, W. P.; Wang, M. R.; Wang, W.; Xing, J. G.; Li, M. H.; Hong, W. S.; Holmes, E. C.; Zhang, Y. Z. , Identification of novel and diverse rotaviruses in rodents and insectivores, and evidence of cross-species transmission into humans. Virology 2016, 494, 168–177. [Google Scholar] [CrossRef]
- Alexander, K. A.; MacLachlan, N. J.; Kat, P. W.; House, C.; O'Brien, S. J.; Lerche, N. W.; Sawyer, M.; Frank, L. G.; Holekamp, K.; Smale, L.; et al. , Evidence of natural bluetongue virus infection among African carnivores. Am J Trop Med Hyg 1994, 51, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S. F.; Tattersall, P. , Parvoviruses: Small Does Not Mean Simple. Annu Rev Virol 2014, 1, 517–537. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S. F.; Tattersall, P. , Genome packaging sense is controlled by the efficiency of the nick site in the right-end replication origin of parvoviruses minute virus of mice and LuIII. J Virol 2005, 79, 2287–2300. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Orba, Y.; Anindita, P. D.; Ishii, A.; Ueno, K.; Hang'ombe, B. M.; Mweene, A. S.; Ito, K.; Sawa, H. , Distinct Lineages of Bufavirus in Wild Shrews and Nonhuman Primates. Emerg Infect Dis 2015, 21, 1230–1233. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y. Q.; You, F. F.; Chen, X. J.; Chen, Y. X.; Wen, Y. Q.; Chen, Q. , Detection and phylogenetic analysis of porcine bocaviruses carried by murine rodents and house shrews in China. Transbound Emerg Dis 2019, 66, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y. Q.; Zhang, M. Y.; Zhou, J. H.; Li, Y. Z.; You, F. F.; Wen, Y. Q.; He, W. Q.; Chen, Q. , A Molecular Epidemiological Investigation of Carriage of the Adeno-Associated Virus in Murine Rodents and House Shrews in China. Intervirology 2018, 61, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, X.; Wang, X.; Dong, H.; Ma, C.; Wang, J.; Liu, B.; Mao, Y.; Wang, Y.; Li, T.; Yang, C.; Guo, Y. , Structural and Functional Diversity of Nairovirus-Encoded Nucleoproteins. J Virol 2015, 89, 11740–11749. [Google Scholar] [CrossRef]
- Garrison, A. R.; Alkhovsky Альхoвский Сергей Владимирoвич, S. V.; Avšič-Županc, T.; Bente, D. A.; Bergeron, É.; Burt, F.; Di Paola, N.; Ergünay, K.; Hewson, R.; Kuhn, J. H.; Mirazimi, A.; Papa, A.; Sall, A. A.; Spengler, J. R.; Palacios, G.; Consortium, I. R., ICTV Virus Taxonomy Profile: Nairoviridae. J Gen Virol 2020, 101, 798–799. [Google Scholar] [CrossRef]
- Ozeki, T.; Abe, H.; Ushijima, Y.; Nze-Nkogue, C.; Akomo-Okoue, E. F.; Ella, G. W. E.; Koumba, L. B. M.; Nso, B.; Mintsa-Nguema, R.; Makouloutou-Nzassi, P.; Makanga, B. K.; Nguelet, F. L. M.; Ondo, G. N.; Mbadinga, M.; Igasaki, Y.; Okada, S.; Hirano, M.; Yoshii, K.; Lell, B.; Bonney, L. C.; Hewson, R.; Kurosaki, Y.; Yasuda, J. , Identification of novel orthonairoviruses from rodents and shrews in Gabon, Central Africa. J Gen Virol 2022, 103, (10). [Google Scholar] [CrossRef]
- Chastel, C.; Main, A. J.; Richard, P.; Le Lay, G.; Legrand-Quillien, M. C.; Beaucournu, J. C. , Erve virus, a probable member of Bunyaviridae family isolated from shrews (Crocidura russula) in France. Acta Virol 1989, 33, 270–280. [Google Scholar]
- Walker, P. J.; Widen, S. G.; Firth, C.; Blasdell, K. R.; Wood, T. G.; Travassos da Rosa, A. P.; Guzman, H.; Tesh, R. B.; Vasilakis, N. , Genomic Characterization of Yogue, Kasokero, Issyk-Kul, Keterah, Gossas, and Thiafora Viruses: Nairoviruses Naturally Infecting Bats, Shrews, and Ticks. Am J Trop Med Hyg 2015, 93, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Low, D. H. W.; Ch'ng, L.; Su, Y. C. F.; Linster, M.; Zhang, R.; Zhuang, Y.; Kwak, M. L.; Borthwick, S. A.; Hitch, A. T.; Smith, G. J. D.; Mendenhall, I. H. , Cencurut virus: A novel Orthonairovirus from Asian house shrews (Suncus murinus) in Singapore. One Health 2023, 16, 100529. [Google Scholar] [CrossRef] [PubMed]
- Bárcena, M.; Oostergetel, G. T.; Bartelink, W.; Faas, F. G.; Verkleij, A.; Rottier, P. J.; Koster, A. J.; Bosch, B. J. , Cryo-electron tomography of mouse hepatitis virus: Insights into the structure of the coronavirion. Proc Natl Acad Sci U S A 2009, 106, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, K.; Mulley, G.; Gulyaeva, A. A.; Zhao, L.; Shu, G.; Jiang, J.; Neuman, B. W. , Description and initial characterization of metatranscriptomic nidovirus-like genomes from the proposed new family Abyssoviridae, and from a sister group to the Coronavirinae, the proposed genus Alphaletovirus. Virology 2018, 524, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Ntumvi, N. F.; Mbala Kingebeni, P.; Tamoufe, U.; Kumakamba, C.; Ndze, V.; Ngay Lukusa, I.; LeBreton, M.; Atibu Losoma, J.; Le Doux Diffo, J.; N'Kawa, F.; Takuo, J. M.; Mulembakani, P.; Nwobegahay, J.; Makuwa, M.; Muyembe Tamfum, J. J.; Gillis, A.; Harris, S.; Rimoin, A. W.; Hoff, N. A.; Fair, J. M.; Monagin, C.; Ayukekbong, J.; Rubin, E. M.; Wolfe, N. D.; Lange, C. E. , High Herpesvirus Diversity in Wild Rodent and Shrew Species in Central Africa. Intervirology 2018, 61, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Tsoleridis, T.; Onianwa, O.; Horncastle, E.; Dayman, E.; Zhu, M.; Danjittrong, T.; Wachtl, M.; Behnke, J. M.; Chapman, S.; Strong, V.; Dobbs, P.; Ball, J. K.; Tarlinton, R. E.; McClure, C. P. , Discovery of Novel Alphacoronaviruses in European Rodents and Shrews. Viruses 2016, 8, 84. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Lu, L.; Du, J.; Yang, L.; Ren, X.; Liu, B.; Jiang, J.; Yang, J.; Dong, J.; Sun, L.; Zhu, Y.; Li, Y.; Zheng, D.; Zhang, C.; Su, H.; Zheng, Y.; Zhou, H.; Zhu, G.; Li, H.; Chmura, A.; Yang, F.; Daszak, P.; Wang, J.; Liu, Q.; Jin, Q. , Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome 2018, 6, 178. [Google Scholar] [CrossRef] [PubMed]
- Barrows, N. J.; Campos, R. K.; Liao, K. C.; Prasanth, K. R.; Soto-Acosta, R.; Yeh, S. C.; Schott-Lerner, G.; Pompon, J.; Sessions, O. M.; Bradrick, S. S.; Garcia-Blanco, M. A. , Biochemistry and Molecular Biology of Flaviviruses. Chem Rev 2018, 118, 4448–4482. [Google Scholar] [CrossRef] [PubMed]
- Bartholomeusz, A. I.; Wright, P. J. , Synthesis of dengue virus RNA in vitro: Initiation and the involvement of proteins NS3 and NS5. Arch Virol 1993, 128, (1–2), 111. [Google Scholar] [CrossRef]
- Diagne, M. M.; Ndione, M. H. D.; Di Paola, N.; Fall, G.; Bedekelabou, A. P.; Sembène, P. M.; Faye, O.; Zanotto, P. M. A.; Sall, A. A. , Usutu Virus Isolated from Rodents in Senegal. Viruses 2019, 11, (2). [Google Scholar] [CrossRef]
- Goethert, H. K.; Mather, T. N.; Johnson, R. W.; Telford, S. R. , 3rd, Incrimination of shrews as a reservoir for Powassan virus. Commun Biol 2021, 4, 1319. [Google Scholar] [CrossRef] [PubMed]
- Nosek, J.; Grulich, I. , The relationship between the tick-borne encephalitis virus and the ticks and mammals of the Tribec mountain range. Bull World Health Organ 1967, (Suppl 1), 31–47. [Google Scholar]
- Banerjee, K.; Ilkal, M. A.; Deshmukh, P. K. , Susceptibility of Cynopterus sphinx (frugivorus bat) & Suncus murinus (house shrew) to Japanese encephalitis virus. Indian J Med Res 1984, 79, 8–12. [Google Scholar] [PubMed]
- Guo, H.; Cai, C.; Wang, B.; Zhuo, F.; Jiang, R.; Wang, N.; Li, B.; Zhang, W.; Zhu, Y.; Fan, Y.; Chen, W.; Chen, W.; Yang, X.; Shi, Z. , Novel hepacivirus in Asian house shrew, China. Sci China Life Sci 2019, 62, 701–704. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y. W.; Wan, Z. W.; Wu, Y.; Li, X. F.; Tang, S. X. , PCR-based screening and phylogenetic analysis of rat pegivirus (RPgV) carried by rodents in China. J Vet Med Sci 2020, 82, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Spilman, M. S.; Welbon, C.; Nelson, E.; Dokland, T. , Cryo-electron tomography of porcine reproductive and respiratory syndrome virus: Organization of the nucleocapsid. J Gen Virol 2009, 90 Pt 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Vanmechelen, B.; Vergote, V.; Laenen, L.; Koundouno, F. R.; Bore, J. A.; Wada, J.; Kuhn, J. H.; Carroll, M. W.; Maes, P. , Expanding the Arterivirus Host Spectrum: Olivier's Shrew Virus 1, A Novel Arterivirus Discovered in African Giant Shrews. Sci Rep 2018, 8, 11171. [Google Scholar] [CrossRef] [PubMed]
- Calvete, C.; Mendoza, M.; Sarto, M. P.; Bagüés, M. P. J.; Luján, L.; Molín, J.; Calvo, A. J.; Monroy, F.; Calvo, J. H. , Detection of Rabbit Hemorrhagic Disease Virus GI.2/RHDV2/b in the Mediterranean Pine Vole ( Microtus duodecimcostatus) and White-Toothed Shrew ( Crocidura russula). J Wildl Dis 2019, 55, 467–472. [Google Scholar] [PubMed]
- Guo, L.; Liu, S.; Song, J.; Han, L.; Zhang, H.; Wu, C.; Wang, C.; Zhou, H.; Wang, J. , Seroprevalence of Wenzhou virus in China. Biosaf Health 2020, 2, 152–156. [Google Scholar] [CrossRef]
- Li, K.; Lin, X. D.; Wang, W.; Shi, M.; Guo, W. P.; Zhang, X. H.; Xing, J. G.; He, J. R.; Wang, K.; Li, M. H.; Cao, J. H.; Jiang, M. L.; Holmes, E. C.; Zhang, Y. Z. , Isolation and characterization of a novel arenavirus harbored by Rodents and Shrews in Zhejiang province, China. Virology 2015, 476, 37–42. [Google Scholar] [CrossRef]
- Ushijima, Y.; Abe, H.; Ozeki, T.; Ondo, G. N.; Mbadinga, M.; Bikangui, R.; Nze-Nkogue, C.; Akomo-Okoue, E. F.; Ella, G. W. E.; Koumba, L. B. M.; Nso, B.; Mintsa-Nguema, R.; Makouloutou-Nzassi, P.; Makanga, B. K.; Nguelet, F. L. M.; Zadeh, V. R.; Urata, S.; Mbouna, A. V. N.; Massinga-Loembe, M.; Agnandji, S. T.; Lell, B.; Yasuda, J. , Identification of potential novel hosts and the risk of infection with lymphocytic choriomeningitis virus in humans in Gabon, Central Africa. Int J Infect Dis 2021, 105, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Hilbe, M.; Herrsche, R.; Kolodziejek, J.; Nowotny, N.; Zlinszky, K.; Ehrensperger, F. , Shrews as reservoir hosts of borna disease virus. Emerg Infect Dis 2006, 12, 675–677. [Google Scholar] [CrossRef]
- Xiong, Y. Q.; Mo, Y.; Chen, M. J.; Cai, W.; He, W. Q.; Chen, Q. , Detection and phylogenetic analysis of torque teno virus (TTV) carried by murine rodents and house shrews in China. Virology 2018, 516, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Fevola, C.; Rossi, C.; Rosso, F.; Girardi, M.; Rosà, R.; Manica, M.; Delucchi, L.; Rocchini, D.; Garzon-Lopez, C. X.; Arnoldi, D.; Bianchi, A.; Buzan, E.; Charbonnel, N.; Collini, M.; Ďureje, L.; Ecke, F.; Ferrari, N.; Fischer, S.; Gillingham, E. L.; Hörnfeldt, B.; Kazimírová, M.; Konečný, A.; Maas, M.; Magnusson, M.; Miller, A.; Niemimaa, J.; Nordström, Å.; Obiegala, A.; Olsson, G.; Pedrini, P.; Piálek, J.; Reusken, C. B.; Rizzolli, F.; Romeo, C.; Silaghi, C.; Sironen, T.; Stanko, M.; Tagliapietra, V.; Ulrich, R. G.; Vapalahti, O.; Voutilainen, L.; Wauters, L.; Rizzoli, A.; Vaheri, A.; Jääskeläinen, A. J.; Henttonen, H.; Hauffe, H. C. , Geographical Distribution of Ljungan Virus in Small Mammals in Europe. Vector Borne Zoonotic Dis 2020, 20, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J. F.; Corman, V. M.; Lukashev, A. N.; van den Brand, J. M.; Gmyl, A. P.; Brünink, S.; Rasche, A.; Seggewiβ, N.; Feng, H.; Leijten, L. M.; Vallo, P.; Kuiken, T.; Dotzauer, A.; Ulrich, R. G.; Lemon, S. M.; Drosten, C. , Evolutionary origins of hepatitis A virus in small mammals. Proc Natl Acad Sci U S A 2015, 112, 15190–15195. [Google Scholar] [CrossRef] [PubMed]
- Gedvilaite, A.; Tryland, M.; Ulrich, R. G.; Schneider, J.; Kurmauskaite, V.; Moens, U.; Preugschas, H.; Calvignac-Spencer, S.; Ehlers, B. , Novel polyomaviruses in shrews (Soricidae) with close similarity to human polyomavirus 12. J Gen Virol 2017, 98, 3060–3067. [Google Scholar] [CrossRef] [PubMed]
- Orba, Y.; Sasaki, M.; Yamaguchi, H.; Ishii, A.; Thomas, Y.; Ogawa, H.; Hang'ombe, B. M.; Mweene, A. S.; Morikawa, S.; Saijo, M.; Sawa, H. , Orthopoxvirus infection among wildlife in Zambia. J Gen Virol 2015, 96 Pt 2, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, T.; Tryland, M.; Hansen, H.; Mehl, R.; Moens, U.; Olsvik, O.; Traavik, T. , Naturally occurring orthopoxviruses: Potential for recombination with vaccine vectors. J Clin Microbiol 1998, 36, 2542–2547. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Chmura, A. A.; Li, J.; Zhu, G.; Desmond, J. S.; Zhang, Y.; Zhang, W.; Epstein, J. H.; Daszak, P.; Shi, Z. , Detection of diverse novel astroviruses from small mammals in China. J Gen Virol 2014, 95 Pt 11, 2442–2449. [Google Scholar] [CrossRef] [PubMed]
- Morvan, J. M.; Deubel, V.; Gounon, P.; Nakouné, E.; Barrière, P.; Murri, S.; Perpète, O.; Selekon, B.; Coudrier, D.; Gautier-Hion, A.; Colyn, M.; Volehkov, V. , Identification of Ebola virus sequences present as RNA or DNA in organs of terrestrial small mammals of the Central African Republic. Microbes Infect 1999, 1, 1193–1201. [Google Scholar] [CrossRef]
- Liu, J. W.; Wen, H. L.; Fang, L. Z.; Zhang, Z. T.; He, S. T.; Xue, Z. F.; Ma, D. Q.; Zhang, X. S.; Wang, T.; Yu, H.; Zhang, Y.; Zhao, L.; Yu, X. J. , Prevalence of SFTSV among Asian house shrews and rodents, China, January-August 2013. Emerg Infect Dis 2014, 20, 2126–2128. [Google Scholar] [CrossRef] [PubMed]
- Kemp, G. E.; Causey, O. R.; Moore, D. L.; Odelola, A.; Fabiyi, A. , Mokola virus. Further studies on IbAn 27377, a new rabies-related etiologic agent of zoonosis in nigeria. Am J Trop Med Hyg 1972, 21, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; Li, W.; Su, J.; Fang, L.; Takeda, N.; Wakita, T.; Li, T. C.; Ke, C. , Asian musk shrew as a reservoir of rat hepatitis E virus, China. Emerg Infect Dis 2013, 19, 1341–1343. [Google Scholar] [CrossRef] [PubMed]
- Nie, F. Y.; Tian, J. H.; Lin, X. D.; Yu, B.; Xing, J. G.; Cao, J. H.; Holmes, E. C.; Ma, R. Z.; Zhang, Y. Z. , Discovery of a highly divergent hepadnavirus in shrews from China. Virology 2019, 531, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X. Y.; Qiu, M.; Ke, X. M.; Guan, W. J.; Li, J. M.; Huo, S. T.; Chen, S. W.; Zhong, X. S.; Zhou, W.; Xiong, Y. Q.; Ge, J.; Chen, Q. , Detection of novel adenoviruses in fecal specimens from rodents and shrews in southern China. Virus Genes 2016, 52, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Diffo, J.; Ndze, V. N.; Ntumvi, N. F.; Takuo, J. M.; Mouiche, M. M. M.; Tamoufe, U.; Nwobegahay, J.; LeBreton, M.; Gillis, A.; Schneider, B. S.; Fair, J. M.; Monagin, C.; McIver, D. J.; Joly, D. O.; Wolfe, N. D.; Rubin, E. M.; Lange, C. E. , DNA of diverse adenoviruses detected in Cameroonian rodent and shrew species. Arch Virol 2019, 164, 2359–2366. [Google Scholar] [CrossRef]
- Harima, H.; Sasaki, M.; Kajihara, M.; Mori-Kajihara, A.; Hang'ombe, B. M.; Changula, K.; Orba, Y.; Ogawa, H.; Simuunza, M.; Yoshida, R.; Mweene, A.; Takada, A.; Sawa, H. , Detection of novel orthoreovirus genomes in shrew (Crocidura hirta) and fruit bat (Rousettus aegyptiacus). J Vet Med Sci 2020, 82, 162–167. [Google Scholar] [CrossRef]
Figure 1.
Three Asian house shrews we raised in a barrel with soil.

Figure 2.
Numbers of known host species and known virus species in each shrew genus. The genera shown in red, blue, and green colors belongs to the subfamilies of Crocidurinae (white-toothed shrews), Soricinae (red-toothed shrews), and Myosoricinae (African shrews), respectively.
Figure 2.
Numbers of known host species and known virus species in each shrew genus. The genera shown in red, blue, and green colors belongs to the subfamilies of Crocidurinae (white-toothed shrews), Soricinae (red-toothed shrews), and Myosoricinae (African shrews), respectively.
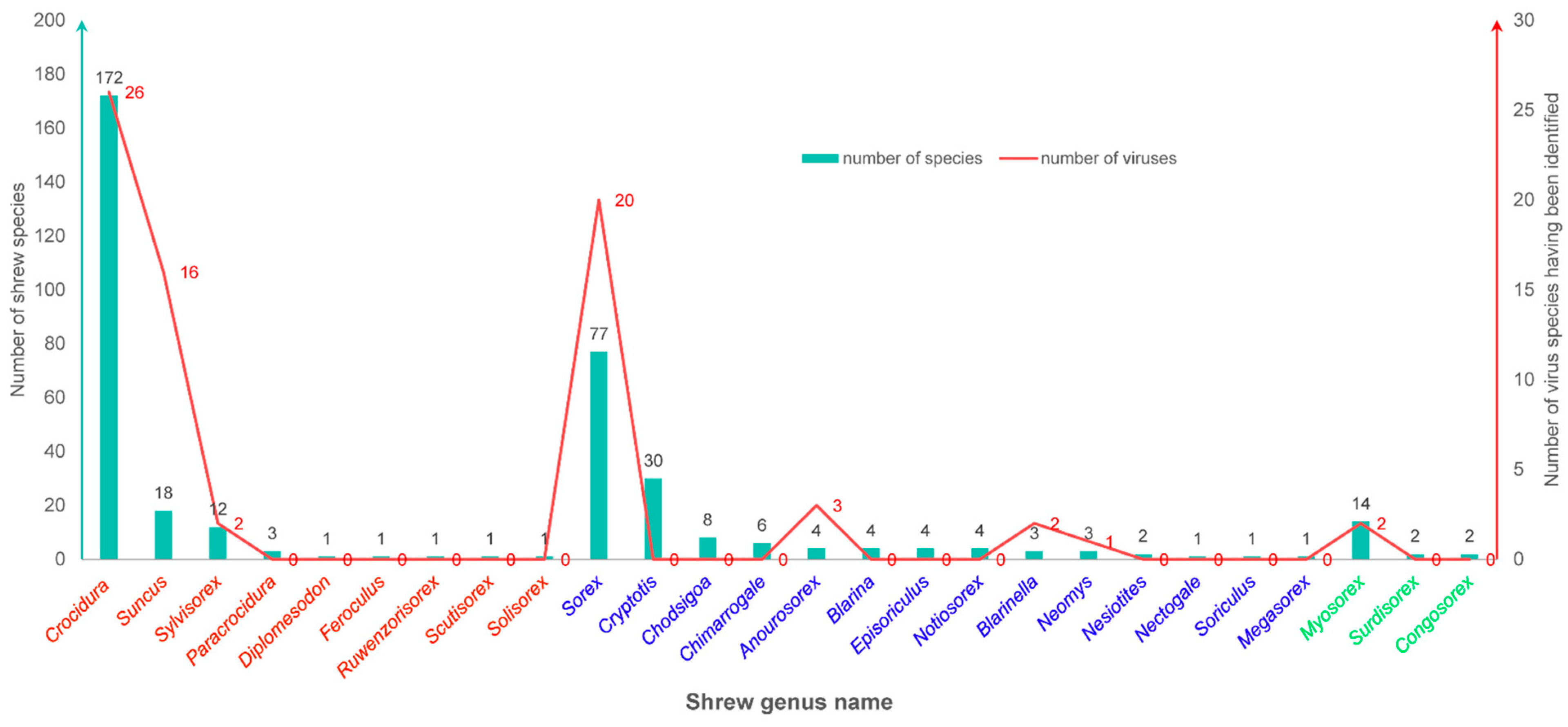

Table 1.
Geographic and viral distribution of 26 shrew genera.
| Shrew Genus | Geographical Distribution |
Viral Distribution (Shrew Host Species Were Given in Parentheses) |
| Crocidura in the subfamily Crocidurinae covers 172 species | Distributed across all tropical and temperate regions of Africa, Europe, and Asia, from northern South Africa to Europe, and across the entire eastern part of Asia, extending to the Malay Archipelago | Langya virus (Crocidura shantungensis, Crocidura lasiura); Gamak virus and Daeryong virus (Crocidura shantungensis, Crocidura lasiura); Melian virus (Crocidura grandiceps); Denwin virus (Crocidura russula); Beilong virus (Crocidura shantungensis); Bowé virus (Crocidura douceti); Imjin virus (Crocidura lasiura); Jeju virus (Crocidura lasiura); Seoul virus (Crocidura lasiura); Azagny virus (Crocidura obscurior); Bufavirus (Crocidura hirta); Lamusara virus (Crocidura sp.); Lamgora virus (Crocidura sp.); Erve virus (Crocidura russula); Thiafora virus (Crocidura spp.); Usutu virus (Crocidura sp.); Olivier's shrew virus 1 (Crocidura olivieri guineensis); lymphocytic choriomeningitis virus (Crocidura goliath); Borna disease virus (Crocidura leucodon); Ljungan virus (Crocidura leucodon); Monkeypox virus (Crocidura spp.); Shrew herpesviruses (Crocidura spp.); Mokola virus (Crocidura spp.); rabbit hemorrhagic disease virus 2b (Crocidura russula); Shrew orthoreovirus (Crocidura hirta) |
| Suncus in the subfamily Crocidurinae covers 18 species | Distributed in all continents except Oceania | Thottapalayam virus (Suncus murinus); Rotavirus A (Suncus murinus); Porcine Bocavirus G4 (Suncus murinus); adeno-associated virus (Suncus murinus); Japanese encephalitis virus (Suncus murinus); Suncus murinus hepacivirus (Suncus murinus); rat pegivirus (Suncus murinus); Wenzhou virus (Suncus murinus); torque teno virus (Suncus murinus); astroviruses (Suncus murinus); severe fever with thrombocytopenia syndrome virus (Suncus murinus); hepatitis E virus (Suncus murinus); shrew hepatitis B virus (Suncus murinus); Asian house shrew adenovirus (Suncus murinus); Cencurut virus (Suncus murinus); Wénchéng shrew virus (Suncus murinus) |
| Sylvisorex in the subfamily Crocidurinae covers 12 species | Distributed in Africa | Ebola virus (Sylvisorex ollula); Unnamed shrew adenovirus (Sylvisorex sp.) |
|
Diplomesodon in the subfamily Crocidurinae covers 1 species |
Distributed in Caspian region | No viruses have been identified in this shrew genus |
|
Feroculus in the subfamily Crocidurinae covers 1 species |
Distributed in southern Sri Lanka and southern India | No viruses have been identified in this shrew genus |
| Paracrocidura in the subfamily Crocidurinae covers 3 species | Distributed in Burundi, the Democratic Republic of the Congo, Rwanda, Uganda, Gabon, the Central African Republic, and Equatorial Guinea in Africa | No viruses have been identified in this shrew genus |
|
Ruwenzorisorex in the subfamily Crocidurinae covers 1 species |
Distributed in Found in Burundi, the Democratic Republic of the Congo, Rwanda, and Uganda | No viruses have been identified in this shrew genus |
|
Scutisorex in the subfamily Crocidurinae covers 1 species |
Distributed in the Republic of the Congo | No viruses have been identified in this shrew genus |
|
Solisorex in the subfamily Crocidurinae covers 1 species |
Distributed in Sri Lanka and southern India | No viruses have been identified in this shrew genus |
|
Sorex in the subfamily Soricinae covers 77 species |
Distributed Eurasia and North America | Ninorex virus (Sorex minutus); Lena mobatvirus (Sorex caecutiens, Sorex roboratus); Asikkala virus (Sorex minutus); Asikkala virus (Sorex minutus); Kenkeme virus (Sorex roboratus); Seewis virus (Sorex araneus); Yakeshi virus (Sorex unguiculatus); Altai virus (Sorex araneus); Artybash virus (Sorex caecutiens); Artybash virus (Sorex caecutiens); Ash River virus (Sorex cinereus) Jemez Springs virus (Sorex monticolus); Puumala virus (Sorex araneus); Qian hu shan virus (Sorex cylindricauda); common shrew coronavirus Tibet-2014 (Sorex araneus); Ljungan Virus (Sorex antinorii); Hepatitis A virus (sorex araneus); human polyomavirus 12 (sorex araneus, sorex coronatus, sorex minutus); cowpox virus (Sorex araneus); tick-borne encephalitis virus (Sorex araneus) |
|
Anourosorex in the subfamily Soricinae covers 4 species |
Distributed in China, Taiwan, India, and Indochina | Cao Bang virus (Anourosorex squamipes); Lianghe virus (Anourosorex squamipes); Xinyi virus (Anourosorex yamashinai) |
|
Blarinella in the subfamily Soricinae covers 3 species |
Distributed in India, China, and Myanmar | Camp Ripley virus (Blarina brevicauda); Powassan virus type 2 (Blarina brevicauda) |
|
Neomys in the subfamily Soricinae covers 3 species |
Distributed in Europe and northern Asia | Boginia virus (Neomys fodiens) |
|
Blarina in the subfamily Soricinae covers 4 species |
Distributed in North America | No viruses have been identified in this shrew genus |
|
Cryptotis in the subfamily Soricinae covers 30 species |
Distributed in North America and Central America | No viruses have been identified in this shrew genus |
|
Chimarrogale in the subfamily Soricinae covers 6 species |
Distributed in Asia | No viruses have been identified in this shrew genus |
|
Chodsigoa in the subfamily Soricinae covers 8 species |
Distributed in Asia | No viruses have been identified in this shrew genus |
|
Episoriculus in the subfamily Soricinae covers 4 species |
Distributed in Asia | No viruses have been identified in this shrew genus |
|
Nectogale in the subfamily Soricinae covers 1 species |
Distributed in India and China | No viruses have been identified in this shrew genus |
|
Nesiotites in the subfamily Soricinae covers 2 species |
Distributed in Europe and North Africa | No viruses have been identified in this shrew genus |
|
Soriculus in the subfamily Soricinae covers 1 species |
Distributed in Asia | No viruses have been identified in this shrew genus |
|
Megasorex in the subfamily Soricinae covers 1 species |
Distributed in the United States and Mexico | No viruses have been identified in this shrew genus |
|
Notiosorex in the subfamily Soricinae covers 4 species |
Distributed in the United States and Mexico | No viruses have been identified in this shrew genus |
|
Myosorex in the subfamily Myosoricinae covers 14 species |
Distributed in Africa | Kilimanjaro virus (Myosorex geata); Uluguru virus (Myosorex zinki) |
|
Surdisorex in the subfamily Myosoricinae covers 2 species |
Distributed in Kenya | No viruses have been identified in this shrew genus |
|
Congosorex in the subfamily Myosoricinae covers 2 species |
Distributed in Gabon, Central African Republic, Democratic Congo, Cameroon, Tanzania | No viruses have been identified in this shrew genus |
Table 2.
The common names of some shrew species.
| Common Name of Shrews | Species Name of Shrews |
| American water shrew | Sorex palustris |
| Asian house shrews | Suncus murinus |
| Asian lesser white-toothed shrews | Crocidura shantungensis |
| bicolored shrews | Crocidura leucodon |
| Chinese mole shrews | Anourosorex squamipes |
| Doucet's musk shrews | Crocidura douceti |
| dusky shrews | Sorex monticolus |
| Eurasian pygmy shrews | Sorex minutus |
| Eurasian common shrews | Sorex Araneus |
| Eurasian water shrews | Neomys fodiens |
| flat-skulled shrews | Sorex roboratus |
| Geata mouse shrews | Myosorex geata |
| Goliath shrews | Crocidura goliath |
| greater forest shrews | Sylvisorex ollula |
| greater white-toothed shrews | Crocidura russula |
| Kilimanjaro mouse shrews | Myosorex zinki |
| large-headed shrews | Crocidura grandiceps |
| Laxmann's shrews | Sorex caecutiens |
| lesser red musk shrews | Crocidura hirta |
| long-clawed shrews | Sorex unguiculatus |
| masked shrews | Sorex cinereus |
| Millet's shrews | Sorex coronatus |
| northern short-tailed shrews | Blarina brevicauda |
| Olivier's shrews | Crocidura olivieri guineensis |
| stripe-backed shrews | Sorex cylindricauda |
| Taiwanese mole shrews | Anourosorex yamashinai |
| Ussuri white-toothed shrews | Crocidura lasiura |
| Valais shrews | Sorex antinorii |
| West African pygmy shrews | Crocidura obscurior |
Table 2.
The taxonomy and zoonotic potential of the viruses having been identified in shrews.
| Family | Genus | Virus | Zoonotic Potential | Year / Country / Host Species |
| Paramyxoviridae | Henipavirus | Langya virus | Infects humans and some other mammals | 2022 / China / Crocidura lasiura, Crocidura shantungensis [10] |
| Paramyxoviridae | Henipavirus | Gamak virus | Unknown | 2021 / China / Crocidura lasiura, Crocidura shantungensis [11] |
| Paramyxoviridae | Henipavirus | Daeryong virus | Unknown | 2021 / China / Crocidura lasiura, Crocidura shantungensis [11] |
| Paramyxoviridae | Henipavirus | Melian virus | Unknown | 2022 / Guinea / Crocidura grandiceps [12] |
| Paramyxoviridae | Henipavirus | Denwin virus | Unknown | 2022 / Belgium / Crocidura russula [12] |
| Paramyxoviridae | Henipavirus | Ninorex virus | Unknown | 2023 / Belgian / Sorex minutus [13] |
| Paramyxoviridae | Jeilongvirus | Beilong virus | Unknown | 2019 / China / Crocidura shantungensis, Suncus murinus [14] |
| Hantaviridae | Mobatvirus | Lena mobatvirus | Unknown | 2021 / Siberian and Russia / Sorex caecutiens, Sorex roboratus [17] |
| Hantaviridae | Orthohantavirus | Asikkala virus | Unknown | 2010 / Finland / Sorex minutus [18] |
| Hantaviridae | Orthohantavirus | Bowé virus | Unknown | 2012 / Guinea / Crocidura douceti [19] |
| Hantaviridae | Orthohantavirus | Cao Bang virus | Unknown | 2006 / China / Anourosorex squamipes [20] |
| Hantaviridae | Orthohantavirus | Imjin virus | Unknown | 2009 / South Korea / Crocidura lasiura [21] |
| Hantaviridae | Orthohantavirus | Jeju virus | Unknown | 2012 / South Korea / Crocidura lasiura [22] |
| Hantaviridae | Orthohantavirus | Kenkeme virus | Unknown | 2010 / Sakha Republic, Siberia / Sorex roboratus [23] |
| Hantaviridae | Orthohantavirus | Seewis virus | Unknown | 2006 / Switzerland / Sorex araneus [24] |
| Hantaviridae | Orthohantavirus | Yakeshi virus | Unknown | 2013 / China / Sorex unguiculatus [25] |
| Hantaviridae | Orthohantavirus | Lianghe virus | Unknown | 2013 / China / Anourosorex squamipes [25] |
| Hantaviridae | Orthohantavirus | Seoul virus | Infects humans | 2014 / China / Crocidura lasiura [26] |
| Hantaviridae | Orthohantavirus | Altai virus | Unknown | 2007 / Altai Republic / Sorex araneus [27] |
| Hantaviridae | Orthohantavirus | Artybash virus | Unknown | 2006 / Altai Republic / Sorex caecutiens [28] |
| Hantaviridae | Orthohantavirus | Xinyi virus | Unknown | 2016 / Chian / Anourosorex yamashinai [29] |
| Hantaviridae | Orthohantavirus | Ash River virus | Unknown | 2008 / United States / Sorex cinereus, Sorex monticolus [30] |
| Hantaviridae | Orthohantavirus | Camp Ripley virus | Unknown | 2007 / United States / Blarina brevicauda [31] |
| Hantaviridae | Orthohantavirus | Boginia virus | Unknown | 2013 / Poland / Neomys fodiens [32] |
| Hantaviridae | Orthohantavirus | Azagny virus | Unknown | 2011 / Côte d'Ivoire / Crocidura obscurior [33] |
| Hantaviridae | Orthohantavirus | Puumala virus | Unknown | 2014 / Finland / Sorex araneus [34] |
| Hantaviridae | Orthohantavirus | Kilimanjaro virus | Unknown | 2014 / West Africa / Myosorex geata [35] |
| Hantaviridae | Orthohantavirus | Uluguru virus | Unknown | 2014 / West Africa / Myosorex zinki [35] |
| Hantaviridae | Orthohantavirus | Qian hu shan virus | Unknown | 2014 / China / Sorex cylindricauda [36] |
| Hantaviridae | Thottimvirus | Thottapalayam virus | Unknown | 1965 / India / Suncus murinus [37] |
| Sedoreoviridae | Rotavirus | Rotavirus A | Infects humans | 2016 / China / Suncus murinus [40] |
| Sedoreoviridae | Orbivirus | Bluetongue virus | Unknown | 1994 / Africa / unknown shrew species [41] |
| Parvoviridae | Protoparvovirus | Bufavirus | Infects humans | 2015 / Zambia / Crocidura hirta [44] |
| Parvoviridae | Bocaparvovirus | porcine bocavirus G4 | Infects pigs | 2019 / China / Suncus murinus [45] |
| Parvoviridae | Dependoparvovirus | adeno-associated virus | Unknown | 2020 / China / Suncus murinus [46] |
| Nairoviridae | Orthonairovirus | Lamusara virus | Unknown | 2022 / Gabon / Crocidura sp. [49] |
| Nairoviridae | Orthonairovirus | Lamgora virus | Unknown | 2022 / Gabon / Crocidura sp. [49] |
| Nairoviridae | Orthonairovirus | Erve virus | Unknown | 1989 / France / Crocidura russula [50] |
| Nairoviridae | Orthonairovirus | Thiafora virus | Unknown | 2015 / Senegal / Crocidura sp. [51] |
| Nairoviridae | Orthonairovirus | Cencurut virus | Unknown | 2023 / Singapore / Suncus murinus [52] |
| Coronaviridae | Alphacoronavirus | Wencheng shrew virus | Unknown | 2017 / China / Suncus murinus [55] |
| Coronaviridae | Alphacoronavirus | unnamed shrew coronavirus | Unknown | 2016 / United Kingdom / Sorex araneus [56] |
| Coronaviridae | Alphacoronavirus | common shrew coronavirus Tibet-2014 | Unknown | 2014 / China / Sorex araneus [57] |
| Flaviviridae | Orthoflavivirus | Usutu virus | Infects birds, arthropods, humans, bats, and horses | 2019 / Senegal / Crocidura sp. [60] |
| Flaviviridae | Orthoflavivirus | Powassan virus type 2 | Infects humans and causes encephalitis | 2021 / United States / Blarina brevicauda [61] |
| Flaviviridae | Orthoflavivirus | tick-borne encephalitis virus | Infects humans and causes encephalitis | 1967 / Slovakia / Sorex araneus [62] |
| Flaviviridae | Orthoflavivirus | Japanese encephalitis virus | Infects humans and various domestic animals and causes human encephalitis | 1984 / Japan / Suncus murinus [63] |
| Flaviviridae | Hepacivirus | Suncus murinus hepacivirus | Unknown | 2016 / China / Suncus murinus [64] |
| Flaviviridae | Pegivirus | rat pegivirus | Infects humans and other animals without symptoms | 2020 / China / Suncus murinus [65] |
| Arteriviridae | Muarterivirus | Olivier's shrew virus 1 | Unknown | 2018 / Guinea / Crocidura olivieri guineensis [67] |
| Caliciviridae | Lagovirus | Rabbit hemorrhagic disease virus | Infects rabbits | 2019 / Spain / Crocidura russula [68] |
| Arenaviridae | Mammarenavirus | Wenzhou virus | Infects humans | 2015 / China / Suncus murinus [69,70] |
| Arenaviridae | Mammarenavirus | lymphocytic choriomeningitis virus | Infects humans and various animals and dangerous to pregnant women and infants | 2021 / Gabon / Crocidura goliath [71] |
| Bornaviridae | Orthobornavirus | Borna disease virus | Infects humans, horses, sheep, and various other mammals and causes encephalitis | 2006 / Switzerland / Crocidura leucodon [72] |
| Circoviridae | Anellovirus | torque teno virus | Infects humans and causes various symptoms | 2018 / China / Suncus murinus [73] |
| Picornaviridae | Parechovirus | Ljungan Virus | Infects humans and causes various symptoms | 2020 / Italy / Crocidura leucodon, Sorex antinorii [74] |
| Picornaviridae | Hepatovirus | Hepatitis A virus | Unknown | 2015 / Germany / sorex araneus [75] |
| Polyomaviridae | Betapolyomavirus | human polyomavirus 12 | Infects humans | 2017 / Germany, Norway / sorex Araneus, sorex coronatus, sorex minutus [76] |
| Poxviridae | Orthopoxvirus | monkeypox virus | Infects humans and other primates | 2015 / Zambia, Congo / Crocidura spp. [77] |
| Poxviridae | Orthopoxvirus | shrew cowpox virus | Unknown | 1998 / Norway / Sorex araneus [78] |
| Astroviridae | Mamastrovirus | shrew astrovirus | Unknown | 2014 / China / Suncus murinus [79] |
| Filoviridae | Ebolavirus | Ebola virus | Infects humans with a high case fatality rate | 1999 / Central Africa / Sylvisorex ollula [80] |
| Phenuiviridae | Phlebovirus | severe fever with thrombocytopenia syndrome virus | Infects humans with a high case fatality rate | 2014 / China / Suncus murinus [81] |
| Herpesviridae | Unknown | shrew herpesvirus | Unknown | 2018 / Central Africa / Crocidura spp. [55] |
| Rhabdoviridae | Lyssavirus | Mokola virus | Unknown | 1968 / Nigeria / Crocidura spp. [82] |
| Hepeviridae | Paslahepevirus | shrew hepatitis E virus | Unknown | 2013 / China / Suncus murinus [83] |
| Hepadnaviridae | Orthohepadnavirus | shrew hepatitis B virus | Unknown | 2019 / China / Suncus murinus [84] |
| Adenoviridae | Atadenovirus | Asian house shrew adenovirus | Unknown | 2016 / China / Suncus murinus [85] |
| Adenoviridae | Atadenovirus | shrew adenovirus | Unknown | 2019 / Cameroon / Sylvisorex sp. [86] |
| Spinareoviridae | Orthoreovirus | shrew orthoreovirus | Unknown | 2019 / Zambia / Crocidura hirta [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



