Submitted:
17 December 2023
Posted:
18 December 2023
You are already at the latest version
Abstract
Nitric oxide (NO) is synthesized and is involved in various physiological and developmental process regulation in all kingdoms of life. Fungi have been less explored in terms of endogenous NO biology than mammals, plants, and bacteria. In this review, we summarize and discuss the studies to date on intracellular NO biosynthesis and function in fungi. Two mechanisms for NO biosynthesis, NO synthase (NOS)-mediated arginine oxidation, and nitrate and nitrite reductase-mediated nitrite reduction, are most frequently reported. Furthermore, we summarize the multifaceted functions of NO in fungi, as well as its role as a signaling molecule in fungal growth regulation, development, abiotic stress, virulence regulation, and metabolism. Finally, we present potential directions for future research on fungal NO biology.
Keywords:
Nitric oxide
; Fungi
; Endogenous production
; Nitric oxide synthase
; Nitrite reductase
; Nitrate reductase
; Biological function
; Signaling molecule
1. Introduction
Nitric oxide (NO) is a diatomic gas synthesized by bacteria, fungi, plants, and mammals. Although NO biosynthesis mechanisms vary among species, increasing evidence has demonstrated the conserved role of endogenous NO as a signaling molecule that regulates many physiological and differential processes [1,2,3]. In mammals, NO is produced by NO synthase (NOS), which plays a crucial role in vasodilation, neurotransmission, and the immune response [4,5]. NO produced by endothelial cells located within blood vessels induces vasodilation, increases blood flow, and regulates blood pressure [6]. In the nervous system, neuron-produced NO acts as a neurotransmitter, participates in synaptic transmission and plasticity, and ultimately affects learning and memory processes [7]. During immune responses, immune cell-produced NO enhances the anti-microbial activity of macrophages and regulates inflammatory factor and chemokine expression [8,9].
In plants, NO is an important signaling molecule that regulates plant growth, maturation, and stress, as well as seed germination, root formation, stomatal aperture, flowering, and senescence [3,10,11,12]. During embryonic development, NO participates in seed dormancy and germination by regulating protein tyrosine nitration and cysteine S-nitrosylation [13]. The root cells at the root tip also generate NO, which is implicated in root hair development and lateral root formation [14,15]. The opening and closing of stomata, gas exchange, and water loss can be controlled by regulating the NO levels in guard cells [16]. NO interacts with plant hormones (auxins, abscisic acid, and gibberellins) to regulate plant growth and development [13,15,16]. However, NO synthesis in plant cells does not appear to follow the same pathway as that in mammalian cells. NOS-like enzyme activities (oxidation of arginine to produce citrulline and NO) are also found in plants, but the sequence homology between plant and mammalian NOS is low [17]. Other biosynthetic pathways have also been identified in plants. NO is synthesized via nitrate reduction to NO by nitrate and nitrite reductases, representing the dominant plant mechanism of NO synthesis [18].
NO production has also been observed in prokaryotic bacterial cells [19,20]. Bacterial NO is generated by nitrite (NO2-) reduction by nitrite reductase during denitrification and ammonia (NH3) oxidation by hydroxylamine (NH2OH) oxidoreductase [21,22]. In addition, NOS, which contains a region homologous to the oxygenase domain of mammalian NOS, has been found in the genome of many bacteria, and NO is produced by NOS-mediated arginine oxidation, as demonstrated in mammalian cells [23,24,25,26]. Endogenous NO in bacterial cells regulates pathogenicity, toxin biosynthesis, and morphological differentiation [25,27].
Compared with other organisms, fungi have received less attention with respect to endogenous NO production and function [2]. In recent years, accumulating experimental data have demonstrated that fungi can produce endogenous NO and may be involved in fungal physiology, cell differentiation, and pathogenicity regulation [2,28,29]. NO appears to be a universal signaling molecule conserved in organisms of all kingdoms. However, the biosynthetic pathways and functions of endogenous NO in fungal cells are not fully understood [2,28]. Fungi exhibit species diversity and functional complexity, which may lead to various aspects of NO production and function [30,31]. In this review, we summarized the findings of studies on the functions and mechanisms of NO production in various fungi. Endogenous NO is likely to be a universal signaling molecule that is well conserved in all organisms. To understand the universal and conserved roles and fate of NO in prokaryotic and eukaryotic cells, it is important to review the current literature.
2. Fungal Endogenous NO Production
NO biosynthesis within the fungal cells has not yet been clearly elucidated. This may be attributable to the lack of NOS, which shows high homology with mammalian NOS in the fungal genome. NOS-like activity has often been observed in various fungi, through measuring enzyme activity or using mammalian NOS enzyme inhibitors [2,29]. Like plants, fungi are likely to have NOS-independent mechanisms for NO biosynthesis such as nitrite reduction by nitrite reductase during denitrification. However, the different molecular structures of the putative NOS proteins and other NOS-independent mechanisms indicate that further studies should be performed to better understand NO biosynthesis in fungi.
2.1. Arginine-Dependent NO Formation
L-arginine can be oxidized to L-citrulline and NO via NOS [32]. NOS-mediated NO synthesis is well established in mammalian cells [5]. Enzymes homologous to mammalian NOS have been found in plant, bacterial, and fungal genomes; however, their sequence homology to mammalian NOS is low [17,19,29]. The involvement of NOS in NO synthesis in fungi has been examined by measuring biochemical enzyme activity and inhibiting enzyme activity (Table 1). Enzyme activity was assessed by determining the L-arginine to L-citrulline rate of conversion [33,34,35,36,37,38,39,40,41,42,43]. NO synthase activity in the fruiting bodies of Flammulina velutipes can reach 500 pmol/mg/min [34], whereas NO synthase activity in the mycelia of Phycomyces blakesleeanu and Neurospora crassa is only 3 and 18 pmol/mg/min, respectively [44]. Mammalian NOS inhibitors such as L-NAME (NG-nitro-L-arginine methyl ester), L-NMMA (NG-methyl-L-arginine acetate salt), L-NNA (Nw-nitro-L-arginine), and AG (aminoguanidine) reduce intracellular NO levels indicating the NOS involvement in NO synthesis [33,35,39,44,45,46,47,48,49,50,51]. In several fungi, NOS-dependent NO production only occurs under specific environmental conditions. Intracellular NO levels in Pleurotus eryngii var. tuoliensis increased with NOS activity under heat stress [49].
Fungal genome analysis revealed that NOS-like genes with high sequence homology to mammalian NOS were rarely found in the fungal genome. However, in recent studies, NOS proteins were purified from F. velutipes and S. cerevisiae using affinity chromatography [33,34], and NOS genes were identified in the genomes of Shiraia sp. Slf14, M. phaseolina and I. obliquus [39,50,55]. Fungal NOSs have a degree of homology or functional similarity to mammalian NOS but may have significant differences in structure, regulation, and substrate specificity [29,56]. NOS-like enzymes in fungi are highly regulated and influenced by various environmental factors, including changes in oxygen and cofactor levels [29,57].
2.2. Nitrite (NO2-)-Dependent NO Formation
Plant and bacteria-based studies have demonstrated that endogenous NO can be synthesized by nitrite (NO2-) reduction during denitrification. Similar observations were made for fungi (Table 1). Fungi produce NO by reducing inorganic nitrite via denitrification [58,59,60]. The key enzymes involved in this process are nitrate reductase (NR) and nitrite reductase (NiR) [61,62,63]. NR catalyzes the nitrate (NO3-) reduction to nitrite (NO2-) (2NO3- + 2H+ + 2e- → 2NO2- + H2O), and NiR mediates nitrite (NO2-) reduction to NO (2NO2- + 2H+ + 2e- → 2NO + H2O) [60,64,65,66]. Endogenous NO production is related to NR and NiR (NIRK/NIRS) gene expression in the endophytic fungus, Preussia sp. BSL-10 [53]. Furthermore, the NR gene, niaD, is essential for NO production in A. nidulans from the vegetative to early developmental stages [51]. G. lucidum can also produce NO via NR with methyl jasmonate induction [52]. In the endophytic fungus, Shiraia sp. Slf14 NR activity and expression were enhanced by an increase in L-arginine levels, promoting NO production [42]. Many fungi contain genes that encode NR and NiR. However, these genes may not contribute to NO production in certain fungi. For example, NO formation in the rice blast fungus Magnaporthe oryzae is not associated with a nitrite-dependent pathway [67]. NO2--dependent NO production occurs only under specific environmental conditions, as shown in S. cerevisiae, where NO2--dependent NO production was observed only under hypoxic conditions [54]. This may be because the NR and NiR expression and activity are upregulated during denitrification under hypoxic conditions, as observed in Fusarium oxysporum [68].
Interestingly, an acidic environment promotes nitrite (NO2-) conversion to NO via non-enzymatic processes (2 HNO2 ↔ NO + NO2 + H2O ↔ 2NO + 2O2 + H2O) [69]. Currently, there is no evidence of non-enzymatic NO production by fungi. However, non-enzymatic NO production has often been observed in the human stomach, oral cavity, skin surface, urine, and plant cytoplasmic apoplasm [69,70]. This may be because the pKa of nitrite is approximately 3.2 and the pH values in these areas are < 4.5, which is a suitable condition for non-enzymatic NO formation [71].
3. Function of Endogenous NO and NO Signaling in Fungi
3.1. Growth and Development Regulation
Cellularly produced NO is implicated in the regulation of various aspects of fungal growth and development, such as hyphal extension, sporulation, and differentiation (Table 2) [28,29]. Furthermore, it can act as a signaling molecule in developmental processes [72,73]. In Pleurotus ostreatus (edible mushrooms), NO negatively regulates the rate of primordium formation by inhibiting the expression and enzymatic activity of mitochondrial aconitase, thereby reducing ATP production [74]. In A. nidulans, NO is produced via NR that is upregulated upon light-regulated conidiation and catabolized by flavohaemoglobins [51]. Balance between biosynthesis and catabolism of NO results in NO homeostasis in fungal cells, and deviation from NO homeostasis can be a cue for developmental processes [51]. Increase in NO level reduces conidiation and increases sexual development [51]. The balance between light-induced conidiation (asexual reproduction) and sexual reproduction is influenced by intracellular NO level through regulating asexual and sexual development regulator expression [51,75]. Similarly, intracellular NO inhibits the light-induced conidiation and NOS activity is not changed during photocarotenogenesis and photoconidiation regulation in N. crassa [41,44]. However, it is also reported that intracellular NO is highly detected in conidiophores and transcription level of highly expressed genes during conidiation is reduced upon intracellular NO scavenging in N. crassa [76]. Endogenous NO in N. crassa seems to promote the hyphae growth, which may be related to the elevated mss-4 and gel-3 expression, as demonstrated in recent studies [76,77]. Other evidence of NO regulation during light-induced development has been demonstrated in P. blakesleeanus [35]. In this fungus, light induces macrosporangiophore formation and citrulline production from arginine, which are inhibited by NOS inhibitors. In C. coccodes, NO was detected in germinating conidia and might regulate conidial germination [45].
Studies have also demonstrated an association between cGMP, a downstream molecule generated by NO in mammalian cells, and endogenous NO in fungi. In the aquatic fungus B. emersonii, the intracellular level of NO increases during sporulation, and the level of NO is reduced by the addition of an NOS inhibitor. Furthermore, cyclic guanosine monophosphate (cGMP) inhibition prevents zoospore generation [37]. In addition, calcium ions are required for NOS activity [37]. This suggests that the Ca2+-NO-cGMP signaling pathway, in which NO is synthesized by the mediation of NOS and calcium ions, induces cGMP production, eventually regulating zoospore biogenesis. A close association between NOS activity and cGMP levels has also been demonstrated in C. minitans, a sclerotial parasite of the plant pathogenic fungus Sclerotinia sclerotiorum, where cGMP functions as a secondary messenger in NO-mediated conidiation through the NO-sGC(guanylate cyclase)-cGMP signaling pathway [38].In C. minitans, L-arginine drives the formation of endogenous NO through NOS, and NO mediates conidia formation in C. minitans, [46]. Extracellular vesicles (EVs) secreted by the pathogenic fungus C.albicans can promote its own growth through the L-arginine/NO pathway, enhancing pathogenesis [78]. L-arginine was also found to be a key factor in EVs promoting of C.albicans conidia formation [78]. During the 5-day starvation period needed to induce sporulation competence, NOS expression was strongly up-regulated in macroplasmodia of Physarum polycephalum, and sporulation competence was inhibited by NOS inhibitors(l-N6–(1-iminoethyl)-lysine (NIL), indicating the involvement of endogenous NO in sporulation competence [79]. Furthermore, endogenous NO can also regulate fungal growth and development by regulating ROS levels. During pre-infection development of Puccinia striiformis Westend f.sp. tritici (Pst) (the wheat stripe rust pathogen), NO and ROS serve as key signaling molecules to regulate the polar growth of germ tubes [80]. In C.albicans, EVs reduce the intracellular ROS and cell apoptosis by upregulating the expression of NO dioxygenase gene YHB1 [78].
3.2. Response to Stressors
NO acts as a signaling molecule in the fungal response to stress by regulating stress-related gene expression and contributing to cellular defense mechanisms against stress-induced damage (Table 2). Under heat stress, endogenous NO can resist oxidative damage by regulating trehalose accumulation, as has been observed in P. eryngii var. tuoliensis [49,84]. In G. lucidum, the polyamine putrescine alleviates heat shock stress by adjusting intracellular NO accumulation, which influences cellular glutamine levels [82]. In addition, researchers found that the expression of a new gene encoding an inducible NOS-like protein (iNOSL) in Shiraia sp. Slf14(w) was significantly increased by heat stress treatment, thereby producing more endogenous NO, and NO can promote the biosynthesis and release of perylenequinones (PQs) [55]. Similarly, under heat shock, high hydrostatic pressure, and hypoxia, the level of endogenous NO, a response signaling molecule, significantly increased and protected S. cerevisiae cells during stress [85,86]. The pH value also has an impact on NO concentration. When the pH was 3.0, the NO content in the culture liquid of L. edodes and G. frondosa decreased [83]. When the pH is 10.0 (alkaline medium), the NO content increase significantly [83]. However, the NO concentration did not change under temperature stress, carbon stress, and nitrogen stress. [83]. This seems to imply that NO changes differently under the influence of different stress factors. In S. cerevisiae, NO2--dependent NO synthesis is induced by the catalysis of cytochrome c oxidase in mitochondria, regulating the expression of hypoxia-related genes when cells are exposed to hypoxic conditions [54]. H2O2 (oxidative stress) induced apoptotic S. cerevisiae cells synthesize NO through nitric oxide synthase (NOS)-like activity, and NO mediates GAPDH S-nitrosation leading to cell death during chronological lifespan [36]. After stimulation with 420 nm intense pulsed light IPL, the levels of nitric oxide synthase (NOS) and NO increased, while asymmetric dimethylarginine (ADMA) level, keratinase activity, and fungal growth decreased in T. rubrum [40]. Upon exposure to antifungal agents, A. fumigatus responds by increasing NO production in the exposed hyphae [81]. Interestingly, the arbuscular mycorrhizal fungus R. irregularis can enhance rice NR and NOS activity, increase intracellular NO accumulation in symbionts, and improve the tolerance of rice plants to low-temperature stress by regulating proline metabolism [43]. In conclusion, the different responses triggered by NO in fungi may be related to the different nature of the stress.
In contrast, exogenous NO addition increased the stress tolerance capacity of the fungus. The addition of an NO-producing chemical (sodium nitroprusside, SNP) can improve the resistance of P. eryngii var. tuoliensis and Ganoderma oregonense under high-temperature stress [49,94]. Under metal stress (Cu2+ or Cd2+), the exogenous addition of NO exerted a protective effect against S. cerevisiae and P. eryngii [95,96].
3.3. Metabolism Regulation
NO regulates multiple metabolic pathways in fungi, including energy, nitrogen, and secondary metabolite production (Table 2). Many fungal secondary metabolites have been used in medicine, agriculture, and industry, including penicillin (antibiotics from Penicillium), cephalosporins (antibiotics from Acremonium and Cephalosporium), taxanes (anticancer compounds from endophytic fungi), and industrially useful enzymes, such as cellulase, amylase, and flavor/aroma compounds [97]. In the endophytic fungus, Shiraia sp. Slf14(w), an endogenous NO derived from arginine, serves as a signaling molecule and can regulate the biosynthesis of secondary metabolite perylenequinones (anti-microbial, anticancer, and antiviral photodynamic therapy agents) via the NO - cGMP – protein kinage G (PKG) signaling pathway [42,55]. In G. lucidum, NR-dependent endogenous NO production increases methyl jasmonate-induced biosynthesis of ganoderic acid (an important secondary metabolite) [52]. In extractive Shiraia fermentation, elevated levels of endogenous NO significantly increase and regulate hypocrellin A expression (a new photosensitizer for anticancer photodynamic therapy) [89]. NO is involved in expression of biosynthetic genes, Mono, PKS, and Omef for hypocrellin A production and upregulates expression the transporter genes, MFS and ABC, for hypocrellin A exudation [89]. In addition, NO donor addition (sodium nitroprusside) increased hypocrellin A content in the mycelium by increasing intracellular NO levels [90]. Similar results were found in A. nidulans, where the exogenous addition of NO increased mycotoxin production [87]. Endogenous NO also mediates the biosynthesis of antioxidant polyphenols, including inoscavins, phelligridins, davallialactone, and methyldavallialactone [48]. These active substances can be used to treat human diseases caused by oxidative stress, such as cancer, hypertension, neurodegenerative diseases, and autoimmune diseases [48,98]. In N. crassa, intracellular NO is actively involved in cellulase production, and cAMP participates in this regulatory effect [88]. N. crassa transcriptome analysis demonstrates that endogenous NO regulates carbohydrate and amino acid metabolism, including pentose and glucuronate interconversion, as well as fructose, mannose, galactose, amino and nucleotide sugar, arginine, proline, and tyrosine metabolisms [77].Preussia sp. BSL-10, an endophytic fungal strain that produces endogenous NO, indole-3-acetic acid (IAA), and gibberellins (GA4, GA7, GA15, and GA53), promotes edge crop growth and yield [53]. Through RT-PCR, NO biosynthesis is validated by expression of ent-desaturase oxidase (P450-4), GA14 synthase (P450-1) and nitrite reductase (NIRK/NIRS), cytochrome P450 (P450nor), nitrate reductase (NR), NOS-like (NOL), and nitric oxide reductase (QNOR/CNOR) [53]. Synthesis of phytohormones is also verified by expression of enzymes mediating the synthesis process such as tryptophan synthase (TRP), indole-3-acetamide hydrolase (IAAH), tryptophan-2-monooxygenase (IAAM), aldehyde dehydrogenase (ALD), GA4 desaturase (DES), and geranylgeranyl-diphosphate synthase (GGS2) [53]. However, it is unclear whether the production of plant hormones is related to the production of NO [53]. In co-culture of I. obliquus and P. morii, biosynthesis of phenylpropanoids that have antioxidant, anti-inflammatory, anti-diabetic, anti-tumor, and anti-viral properties is enhanced, and endogenous NO participates in fungal interspecies interactions [39]. Co-culture of the two fungi triggered the expression of a gene encoding inducible NOS-like protein (iNOSL) on the genome of I. obliquus. iNOSL is more responsible for NO production in I. obliquus and may serve as important regulators controlling phenylpropanoids production during fungal interspecies interactions [39]. NO biosynthesis is enhanced in two co-cultured fungi with subsequent expression of phenylalanine ammonia lyase (PAL) and 4-coumaric acid-CoA ligase (4CL), and upregulation of styrylpyrone polyphenol biosynthesis in I. obliquus [39].
3.4. Virulence and Pathogenicity
Pathogenicity is defined as the capacity of a microbe to cause damage in a host, while virulence is viewed to be a relative term and defined as the relative capacity of a microbe to cause damage in a host. NO in pathogenic microorganisms seems to play a role in both pathogenicity and virulence. In bacteria, endogenous NO is known to regulate toxin biosynthesis and host infection [19]. For fungi, some pathogenic fungi can cause direct damage to tissues by extending their hyphae into host cells or secreting toxins, and NO plays a role in regulating virulence and interactions with host organisms [67,87]. In the hemibiotrophic fungal pathogen M. oryzae, endogenous NO regulates spore germination and appressorium formation during the initial stages of infection, and NO scavenging significantly reduces the formation of barley (Hordeum vulgare) lesions [67]. In addition, a study demonstrates that genes encoding enzymes involved in arginine biosynthetic pathway are essential for pathogenicity, and NO is generated during conidial germination and appressorium formation (infectious morphogenesis) in M. oryzae [93]. However, researchers state that this NO is not generated through arginine-dependent pathway [93]. In the interaction between plant host and fungal pathogen, NO appears to be an important mediator causing both plant defense and pathogen escape. Because plants produce NO in response to pathogen attacks, pathogens should protect themselves against damages induced by plant generated NO. Metabolizing NO may be a way for pathogens to escape NO generated damages. In M. oryzae, S-(hydroxymethyl)-glutathione dehydrogenase is involved in metabolizing NO by catalyzing the reduction of S-nitrosoglutathione (GSNO) in plant [99]. The fungal deletion mutant of this enzyme shows the increase in sensitivity to exogenous NO in formaldehyde-containing medium and the decrease in turgor pressure of spores and appressoria and toxicity to rice plant, indicating that S-(hydroxymethyl)-glutathione dehydrogenase-mediated NO metabolism is critical for virulence of M. oryzae [99]. In a phytopathogenic fungus B. graminis f.sp. hordei, intracellular NO is a determinant of powdery mildew disease in barley as it controls fungal appressorium structure formation, thereby affecting host infections [47]. Fungal pathogen-produced NO can penetrate plant cells, causing host cell death owing to allergic reactions, and this may facilitate the fungal colonization in plant tissue. In the necrotrophic pathogen B. cinerea, NO is produced inside the germinating spores and mycelium and in the surrounding medium in vitro [91]. Intracellular NO can diffuse outside the fungal cells, stimulating fungal colonization of plant tissues [91]. The fungal pathogen, B. ellipsoidum, induces programmed cell death in lilies, and intracellular NO accumulation is observed in both fungal pathogens and plant cells during infection [92].
Endogenous NO production can also influence A. nidulans virulence via mycotoxin biosynthesis regulation [87]. Mycotoxins seriously threaten human health, and ingesting food contaminated with mycotoxins can cause acute or chronic toxicity to humans and animals. NO increases the ability of Aspergillus to produce mycotoxins, which means that NO increases the virulence of the fungus [87].
4. Conclusions and Future Perspectives
Limited data are available on NO production and its function in fungal cells. Regardless, an increasing number of studies have demonstrated that NO is synthesized in fungal cells and acts as a highly reactive signaling molecule that plays crucial roles in fungal growth and development, metabolic control, virulence enhancement, and environmental adaptation (Figure 1). NO is a universal intracellular regulator of biological functions in all kingdoms of life. However, their biosynthetic pathways do not appear to be well-conserved among kingdoms. Compared to the functional analysis of endogenous fungal NO, the biosynthetic mechanisms for fungal NO are more controversial because NOS with high sequence homology to those of mammals, plants, and bacteria has rarely been found in fungal genomes, and nitrite reduction is another mechanism for NO synthesis. NOS-independent synthesis has also been observed in both plants and bacteria. There may be some general mechanisms for NO synthesis that are well conserved among species, but the lifestyle of the species and environmental conditions can affect the generation of different mechanisms. NO can be generated as a byproduct of cellular metabolic pathways, such as mitochondrial respiration and denitrification processes, as well as other non-enzymatic reactions. The NO of some plant species is produced via these pathways. This can also be a future research subject for elucidating fungal NO biosynthesis pathways. The production and function of endogenous NO remain poorly understood in fungi, and future studies are required to establish conserved NO biology in all kingdoms of life.
Author Contributions
Conceptualization, N.-N.Y. and G.P.; investigation, N.-N.Y. and G.P.; writing—original draft preparation, N.-N.Y.; writing—review and editing, G.P.; supervision, G.P.; project administration, G.P.; and funding acquisition, G.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Foundation of Korea (NRF) 2020R1F1A1070942 and 2021R1A6A1A03038785.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Editage (www.editage.co.kr) for English language editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lamattina, L.; Garcia-Mata, C.; Graziano, M.; Pagnussat, G. Nitric oxide: the versatility of an extensive signal molecule. Annu Rev Plant Biol 2003, 54, 109–136. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, D.; Marcos, J.F.; Marcos, A.T.; Strauss, J. Nitric oxide in fungi: is there NO light at the end of the tunnel? Curr Genet 2016, 62, 513–518. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Yun, B.W. Nitric oxide acts as a key signaling molecule in plant development under stressful conditions. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Bredt, D.S. Endogenous nitric oxide synthesis: biological functions and pathophysiology. Free Radic Res 1999, 31, 577–596. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: regulation and function. Eur Heart J 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Ahmad, A.; Dempsey, S.K.; Daneva, Z.; Azam, M.; Li, N.; Li, P.L.; Ritter, J.K. Role of nitric oxide in the cardiovascular and renal systems. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Esplugues, J.V. NO as a signalling molecule in the nervous system. Br J Pharmacol 2002, 135, 1079–1095. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The metabolic signature of macrophage responses. Front immunol 2019, 10, 1462. [Google Scholar] [CrossRef]
- Kobayashi, Y. The regulatory role of nitric oxide in proinflammatory cytokine expression during the induction and resolution of inflammation. J Leukoc Biol 2010, 88, 1157–1162. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Hancock, J.T. Nitric oxide signalling in plants. New Phytol 2003, 159, 11–35. [Google Scholar] [CrossRef]
- Palavan-Unsal, N.; Arisan, D. Nitric oxide signalling in plants. Bot Rev 2009, 75, 203–229. [Google Scholar] [CrossRef]
- Hancock, J.T. Nitric oxide signaling in plants. Plants (Basel) 2020, 9, 1550. [Google Scholar] [CrossRef]
- Arc, E.; Galland, M.; Godin, B.; Cueff, G.; Rajjou, L. Nitric oxide implication in the control of seed dormancy and germination. Front Plant Sci 2013, 4, 346. [Google Scholar] [CrossRef]
- Correa-Aragunde, N.; Graziano, M.; Lamattina, L. Nitric oxide plays a central role in determining lateral root development in tomato. Planta 2004, 218, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.C.; Graziano, M.; Polacco, J.C.; Lamattina, L. Nitric oxide functions as a positive regulator of root hair development. Plant Signal Behav 2006, 1, 28–33. [Google Scholar] [CrossRef]
- Gayatri, G.; Agurla, S.; Raghavendra, A.S. Nitric oxide in guard cells as an important secondary messenger during stomatal closure. Front Plant Sci 2013, 4, 425. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: an update. J Exp Bot 2018, 69, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Crane, B.R.; Sudhamsu, J.; Patel, B.A. Bacterial nitric oxide synthases. Annu Rev Biochem 2010, 79, 445–470. [Google Scholar] [CrossRef]
- Chen, J.H.; Liu, L.L.; Wang, W.W.; Gao, H.C. Nitric oxide, nitric oxide formers and their physiological impacts in bacteria. Int J Mol Sci 2022, 23, 21. [Google Scholar] [CrossRef]
- Coleman, R.E.; Lancaster, K.M. Heme P460: A (cross) link to nitric oxide. Acc Chem Res 2020, 53, 2925–2935. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.K. Flavohaemoglobin: the pre-eminent nitric oxide-detoxifying machine of microorganisms. F1000Res 2020, 9, F1000 Faculty Rev-7–7. [Google Scholar] [CrossRef]
- Crane, B.R.; Arvai, A.S.; Ghosh, D.K.; Wu, C.; Getzoff, E.D.; Stuehr, D.J.; Tainer, J.A. Structure of nitric oxide synthase oxygenase dimer with pterin and substrate. Science 1998, 279, 2121–2126. [Google Scholar] [CrossRef]
- Adak, S.; Aulak, K.S.; Stuehr, D.J. Direct evidence for nitric oxide production by a nitric-oxide synthase-like protein from Bacillus subtilis. J Biol Chem 2002, 277, 16167–16171. [Google Scholar] [CrossRef] [PubMed]
- Kers, J.A.; Wach, M.J.; Krasnoff, S.B.; Widom, J.; Cameron, K.D.; Bukhalid, R.A.; Gibson, D.M.; Crane, B.R.; Loria, R. Nitration of a peptide phytotoxin by bacterial nitric oxide synthase. Nature 2004, 429, 79–82. [Google Scholar] [CrossRef]
- Li, H.; Poulos, T.L. Structure-function studies on nitric oxide synthases. J Inorg Biochem 2005, 99, 293–305. [Google Scholar] [CrossRef]
- Honma, S.; Ito, S.; Yajima, S.; Sasaki, Y. Nitric oxide signaling for aerial mycelium formation in Streptomyces coelicolor A3 (2). Appl Environ Microbiol 2022, 88, e0122222. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.X.; Lim, J.; Xu, J.Y.; Yu, J.H.; Zheng, W.F. Nitric oxide as a developmental and metabolic signal in filamentous fungi. Mol Microbiol 2020, 113, 872–882. [Google Scholar] [CrossRef]
- Filippovich, S.Y.; Bachurina, G.P. Nitric oxide in fungal metabolism (Review). Appl Biochem Microbiol 2021, 57, 694–705. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; van der Heijden, M.G.A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nature Comm 2019, 10, 4841. [Google Scholar] [CrossRef]
- Zak, J.C.; Willig, M.R. 5 - Fungal biodiversity patterns. In Biodiversity of Fungi; Mueller, G.M., Bills, G.F., Foster, M.S., Eds.; Academic Press: Cambridge, 2004; pp. 59–75. [Google Scholar]
- Boucher, J.L.; Moali, C.; Tenu, J.P. Nitric oxide biosynthesis, nitric oxide synthase inhibitors and arginase competition for L-arginine utilization. Cell Mol Life Sci 1999, 55, 1015–1028. [Google Scholar] [CrossRef]
- Kanadia, R.N.; Kuo, W.N.; Mcnabb, M.; Botchway, A. Constitutive nitric oxide synthase in Saccharomyces cerevisiae. Biochem Mol Biol Int 1998, 45, 1081–1087. [Google Scholar] [CrossRef]
- Song, N.K.; Jeong, C.S.; Choi, H.S. Identification of nitric oxide synthase in Flammulina velutipes. Mycologia 2000, 92, 1027–1032. [Google Scholar] [CrossRef]
- Maier, J.; Hecker, R.; Rockel, P.; Ninnemann, H. Role of nitric oxide synthase in the light-induced development of sporangiophores in Phycomyces blakesleeanus. Plant Physiol 2001, 126, 1323–1330. [Google Scholar] [CrossRef]
- Almeida, B.; Buttner, S.; Ohlmeier, S.; Silva, A.; Mesquita, A.; Sampaio-Marques, B.; Osório, N.S.; Kollau, A.; Mayer, B.; Leão, C.; et al. NO-mediated apoptosis in yeast. J Cell Sci 2007, 120, 3279–3288. [Google Scholar] [CrossRef]
- Vieira, A.L.; Linares, E.; Augusto, O.; Gomes, S.L. Evidence of a Ca2+-NO-cGMP signaling pathway controlling zoospore biogenesis in the aquatic fungus Blastocladiella emersonii. Fungal Genet Biol 2009, 46, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fu, Y.P.; Jiang, D.H.; Xie, J.T.; Cheng, J.S.; Li, G.Q.; Hamid, M.I.; Yi, X.H. Cyclic GMP as a second messenger in the nitric oxide-mediated conidiation of the mycoparasite Coniothyrium minitans. Appl Environ Microbiol 2010, 76, 2830–2836. [Google Scholar] [CrossRef]
- Zhao, Y.; Xi, Q.; Xu, Q.; He, M.; Ding, J.; Dai, Y.; Keller, N.P.; Zheng, W. Correlation of nitric oxide produced by an inducible nitric oxide synthase-like protein with enhanced expression of the phenylpropanoid pathway in Inonotus obliquus cocultured with Phellinus morii. Appl Microbiol Biotechnol 2015, 99, 4361–4372. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Huang, M.; Lv, W.; Hu, Y.; Wang, R.; Zheng, X.; Ma, Y.; Chen, C.; Tang, H. Inhibition of Trichophyton rubrum by 420-nm intense pulsed light: In vitro activity and the role of nitric oxide in fungal death. Front Pharmacol 2019, 10, 1143. [Google Scholar] [CrossRef]
- Filippovich, S.Y.; Onufriev, M.V.; Peregud, D.I.; Bachurina, G.P.; Kritsky, M.S. Nitric-oxide synthase activity in the photomorphogenesis of Neurospora crassa. Appl Biochem Microbiol 2020, 56, 446–452. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, C.; Yang, H.; Liu, Z.; Zhang, Z.; Yan, R.; Zhu, D. L-Arginine enhanced perylenequinone production in the endophytic fungus Shiraia sp. Slf14(w) via NO signaling pathway. Appl Microbiol Biotechnol 2022, 106, 2619–2636. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Bi, S.; Meng, J.; Liu, T.; Li, P.; Yu, C.; Peng, X. Arbuscular mycorrhizal fungi enhanced rice proline metabolism under low temperature with nitric oxide involvement. Front Plant Sci 2022, 13, 962460. [Google Scholar] [CrossRef] [PubMed]
- Ninnemann, H.; Maier, J. Indications for the occurrence of nitric oxide synthases in fungi and plants and the involvement in photoconidiation of Neurospora crassa. Photochem Photobiol 1996, 64, 393–398. [Google Scholar] [CrossRef]
- Wang, J.; Higgins, V.J. Nitric oxide has a regulatory effect in the germination of conidia of Colletotrichum coccodes. Fungal Genet Biol 2005, 42, 284–292. [Google Scholar] [CrossRef]
- Gong, X.Y.; Fu, Y.P.; Jiang, D.H.; Li, G.Q.; Yi, X.H.; Peng, Y.L. L-arginine is essential for conidiation in the filamentous fungus Coniothyrium minitans. Fungal Genet Biol 2007, 44, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Prats, E.; Carver, T.L.; Mur, L.A. Pathogen-derived nitric oxide influences formation of the appressorium infection structure in the phytopathogenic fungus Blumeria graminis. Res Microbiol 2008, 159, 476–480. [Google Scholar] [CrossRef]
- Zheng, W.; Miao, K.; Zhang, Y.; Pan, S.; Zhang, M.; Jiang, H. Nitric oxide mediates the fungal-elicitor-enhanced biosynthesis of antioxidant polyphenols in submerged cultures of Inonotus obliquus. Microbiology (Reading) 2009, 155, 3440–3448. [Google Scholar] [CrossRef]
- Kong, W.; Huang, C.; Chen, Q.; Zou, Y.; Zhang, J. Nitric oxide alleviates heat stress-induced oxidative damage in Pleurotus eryngii var. tuoliensis. Fungal Genet Biol 2012, 49, 15–20. [Google Scholar] [CrossRef]
- Sarkar, T.S.; Biswas, P.; Ghosh, S.K.; Ghosh, S. Nitric oxide production by necrotrophic pathogen Macrophomina phaseolina and the host plant in charcoal rot disease of jute: complexity of the interplay between necrotroph-host plant interactions. PLOS ONE 2014, 9, e107348. [Google Scholar] [CrossRef]
- Marcos, A.T.; Ramos, M.S.; Marcos, J.F.; Carmona, L.; Strauss, J.; Cánovas, D. Nitric oxide synthesis by nitrate reductase is regulated during development in Aspergillus. Mol Microbiol 2016, 99, 15–33. [Google Scholar] [CrossRef]
- Shi, L.; Yue, S.; Gao, T.; Zhu, J.; Ren, A.; Yu, H.; Wang, H.; Zhao, M. Nitrate reductase-dependent nitric oxide plays a key role on MeJA-induced ganoderic acid biosynthesis in Ganoderma lucidum. Appl Microbiol Biotechnol 2020, 104, 10737–10753. [Google Scholar] [CrossRef]
- Al-Hosni, K.; Shahzad, R.; Khan, A.L.; Imran, Q.M.; Al Harrasi, A.; Al Rawahi, A.; Asaf, S.; Kang, S.M.; Yun, B.W.; Lee, I.J. Preussia sp. BSL-10 producing nitric oxide, gibberellins, and indole acetic acid and improving rice plant growth. J Plant Interact 2018, 13, 112–118. [Google Scholar] [CrossRef]
- Castello, P.R.; Woo, D.K.; Ball, K.; Wojcik, J.; Liu, L.; Poyton, R.O. Oxygen-regulated isoforms of cytochrome c oxidase have differential effects on its nitric oxide production and on hypoxic signaling. PNAS 2008, 105, 8203–8208. [Google Scholar] [CrossRef]
- Xu, C.; Lin, W.; Chen, Y.; Gao, B.; Zhang, Z.; Zhu, D. Heat stress enhanced perylenequinones biosynthesis of Shiraia sp. Slf14(w) through nitric oxide formation. Appl Microbiol Biotechnol 2023, 107, 3745–3761. [Google Scholar] [CrossRef]
- Astuti, R.I.; Nasuno, R.; Takagi, H. Nitric oxide signaling in yeast. Appl Microbiol Biotechnol 2016, 100, 9483–9497. [Google Scholar] [CrossRef]
- Franco-Cano, A.; Marcos, A.T.; Strauss, J.; Canovas, D. Evidence for an arginine-dependent route for the synthesis of NO in the model filamentous fungus Aspergillus nidulans. Environ Microbiol 2021, 23, 6924–6939. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Fushinobu, S.; Zhou, S.M.; Wakagi, T.; Shoun, H. The possible involvement of copper-containing nitrite reductase (NirK) and flavohemoglobin in denitrification by the fungus Cylindrocarpon tonkinense. Biosci Biotechnol Biochem 2010, 74, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Shoun, H.; Fushinobu, S.; Jiang, L.; Kim, S.-W.; Wakagi, T. Fungal denitrification and nitric oxide reductase cytochrome P450nor. Phil Trans R Soc B 2012, 367, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Aldossari, N.; Ishii, S. Fungal denitrification revisited - Recent advancements and future opportunities. Soil Biol Biochem 2021, 157, 108250. [Google Scholar] [CrossRef]
- Kobayashi, M.; Shoun, H. The copper-containing dissimilatory nitrite reductase involved in the denitrifying system of the fungus Fusarium oxysporum. J Biol Chem 1995, 270, 4146–4151. [Google Scholar] [CrossRef] [PubMed]
- Morozkina, E.V.; Kurakov, A.V. Dissimilatory nitrate reduction in fungi under conditions of hypoxia and anoxia: a review. Appl Biochem Microbiol 2007, 43, 607–613. [Google Scholar] [CrossRef]
- Schinko, T.; Berger, H.; Lee, W.; Gallmetzer, A.; Pirker, K.; Pachlinger, R.; Buchner, I.; Reichenauer, T.; Guldener, U.; Strauss, J. Transcriptome analysis of nitrate assimilation in Aspergillus nidulans reveals connections to nitric oxide metabolism. Mol Microbiol 2010, 78, 720–738. [Google Scholar] [CrossRef]
- Horchani, F.; Prévot, M.; Boscari, A.; Evangelisti, E.; Meilhoc, E.; Bruand, C.; Raymond, P.; Boncompagni, E.; Aschi-Smiti, S.; Puppo, A.; et al. Both plant and bacterial nitrate reductases contribute to nitric oxide production in Medicago truncatula nitrogen-fixing nodules. Plant Physiol 2011, 155, 1023–1036. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Jimenez, M.; Llamas, A.; Galvan, A.; Fernandez, E. Role of nitrate reductase in NO production in photosynthetic eukaryotes. Plants (Basel) 2019, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Kulbir, *!!! REPLACE !!!*; Das, S.; Devi, T.; Goswami, M.; Yenuganti, M.; Bhardwaj, P.; Ghosh, S.; Sahoo, S.C.; Kumar, P. Oxygen atom transfer promoted nitrate to nitric oxide transformation: a step-wise reduction of nitrate → nitrite → nitric oxide. Chem Sci 2021, 12, 10605–10612. [Google Scholar] [CrossRef] [PubMed]
- Samalova, M.; Johnson, J.; Illes, M.; Kelly, S.; Fricker, M.; Gurr, S. Nitric oxide generated by the rice blast fungus Magnaporthe oryzae drives plant infection. New Phytol 2013, 197, 207–222. [Google Scholar] [CrossRef]
- Fujii, T.; Takaya, N. Denitrification by the fungus Fusarium oxysporum involves NADH-nitrate reductase. Biosci Biotechnol Biochem 2008, 72, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Planchet, E.; Kaiser, W.M. Nitric oxide production in plants: facts and fictions. Plant Signal Behav 2006, 1, 46–51. [Google Scholar] [CrossRef]
- Bethke, P.C.; Badger, M.R.; Jones, R.L. Apoplastic synthesis of nitric oxide by plant tissues. Plant Cell 2004, 16, 332–341. [Google Scholar] [CrossRef]
- Weitzberg, E.; Lundberg, J.O. Nonenzymatic nitric oxide production in humans. Nitric Oxide 1998, 2, 1–7. [Google Scholar] [CrossRef]
- Koch, B.; Barugahare, A.A.; Lo, T.L.; Huang, C.; Schittenhelm, R.B.; Powell, D.R.; Beilharz, T.H.; Traven, A. A metabolic checkpoint for the yeast-to-hyphae developmental switch regulated by endogenous nitric oxide signaling. Cell Rep 2018, 25, 2244–2258.e7. [Google Scholar] [CrossRef]
- Jedelska, T.; Luhova, L.; Petrivalsky, M. Nitric oxide signalling in plant interactions with pathogenic fungi and oomycetes. J Exp Bot 2021, 72, 848–863. [Google Scholar] [CrossRef]
- Hou, L.; Huang, C.; Wu, X.; Zhang, J.; Zhao, M. Nitric oxide negatively regulates the rapid formation of Pleurotus ostreatus primordia by inhibiting the mitochondrial aco gene. J Fungi 2022, 8, 1055. [Google Scholar] [CrossRef]
- Marcos, A.T.; Ramos, M.S.; Schinko, T.; Strauss, J.; Cánovas, D. Nitric oxide homeostasis is required for light-dependent regulation of conidiation in Aspergillus. Fungal Genet Biol 2020, 137, 103337. [Google Scholar] [CrossRef] [PubMed]
- Pengkit, A.; Jeon, S.S.; Son, S.J.; Shin, J.H.; Baik, K.Y.; Choi, E.H.; Park, G. Identification and functional analysis of endogenous nitric oxide in a filamentous fungus. Sci Rep 2016, 6, 30037. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.N.; Veerana, M.; Ketya, W.; Sun, H.N.; Park, G. RNA-seq-based transcriptome analysis of nitric oxide scavenging response in Neurospora crassa. J Fungi 2023, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wang, Z.; Liu, Y.Q.; Liao, B.Y.; Zong, Y.W.; Shi, Y.Y.; Liao, M.; Wang, J.N.; Zhou, X.D.; Cheng, L.; et al. Extracellular vesicles of Candida albicans regulate its own growth through the l-arginine/nitric oxide pathway. Appl Microbiol Biotechnol 2023, 107, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Golderer, G.; Werner, E.R.; Leitner, S.; Gröbner, P.; Werner-Felmayer, G. Nitric oxide synthase is induced in sporulation of Physarum polycephalum. Genes Dev 2001, 15, 1299–1309. [Google Scholar] [CrossRef]
- Yin, S.; Gao, Z.; Wang, C.; Huang, L.; Kang, Z.; Zhang, H. Nitric oxide and reactive oxygen species coordinately regulate the germination of Puccinia striiformis f. sp. tritici urediniospores. Front Microbiol 2016, 7, 178. [Google Scholar] [CrossRef]
- Oiki, S.; Nasuno, R.; Urayama, S.-I.; Takagi, H.; Hagiwara, D. Intracellular production of reactive oxygen species and a DAF-FM-related compound in Aspergillus fumigatus in response to antifungal agent exposure. Sci Rep 2022, 12, 13516. [Google Scholar] [CrossRef]
- Xia, J.L.; Wu, C.G.; Ren, A.; Hu, Y.R.; Wang, S.L.; Han, X.F.; Shi, L.; Zhu, J.; Zhao, M.W. Putrescine regulates nitric oxide accumulation in Ganoderma lucidum partly by influencing cellular glutamine levels under heat stress. Microbiol Res 2020, 239, 11. [Google Scholar] [CrossRef]
- Loshchinina, E.A.; Nikitina, V.E. Role of the NO synthase system in response to abiotic stress factors for Basidiomycetes Lentinula edodes and Grifola frondosa. Mikrobiologiia 2016, 85, 154–161. [Google Scholar] [CrossRef]
- Kong, W.W.; Huang, C.Y.; Chen, Q.; Zou, Y.J.; Zhao, M.R.; Zhang, J.X. Nitric oxide is involved in the regulation of trehalose accumulation under heat stress in Pleurotus eryngii var. tuoliensis. Biotechnol Lett 2012, 34, 1915–1919. [Google Scholar] [CrossRef]
- Domitrovic, T.; Palhano, F.L.; Barja-Fidalgo, C.; DeFreitas, M.; Orlando, M.T.; Fernandes, P.M. Role of nitric oxide in the response of Saccharomyces cerevisiae cells to heat shock and high hydrostatic pressure. FEMS Yeast Res 2003, 3, 341–346. [Google Scholar] [CrossRef]
- Castello, P.R.; David, P.S.; McClure, T.; Crook, Z.; Poyton, R.O. Mitochondrial cytochrome oxidase produces nitric oxide under hypoxic conditions: Implications for oxygen sensing and hypoxic signaling in eukaryotes. Cell Metab 2006, 3, 277–287. [Google Scholar] [CrossRef]
- Baidya, S.; Cary, J.W.; Grayburn, W.S.; Calvo, A.M. Role of nitric oxide and flavohemoglobin homolog genes in Aspergillus nidulans sexual development and mycotoxin production. Appl Environ Microbiol 2011, 77, 5524–5528. [Google Scholar] [CrossRef]
- Yu, N.N.; Ketya, W.; Park, G. Intracellular nitric oxide and cAMP are involved in cellulolytic enzyme production in Neurospora crassa. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Wang, Y.; Ma, Y.J.; Wang, J.W.; Zheng, L.P. Nitric oxide and hydrogen peroxide signaling in extractive Shiraia fermentation by Triton X-100 for hypocrellin A production. Int J Mol Sci 2020, 21, 882. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Li, X.P.; Wang, Y.; Wang, J.W. Nitric oxide donor sodium nitroprusside-induced transcriptional changes and hypocrellin biosynthesis of Shiraia sp. S9. Microb Cell Factories 2021, 20, 92. [Google Scholar] [CrossRef] [PubMed]
- Turrion-Gomez, J.L.; Benito, E.P. Flux of nitric oxide between the necrotrophic pathogen Botrytis cinerea and the host plant. Mol Plant Pathol 2011, 12, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Van Baarlen, P.; Staats, M.; Van Kan, J.A.L. Induction of programmed cell death in lily by the fungal pathogen Botrytis elliptica. Mol Plant Pathol 2004, 5, 559–574. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, H.; Liang, S.; Ning, G.; Xu, N.; Lu, J.; Liu, X.; Lin, F. MoARG1, MoARG5,6 and MoARG7 involved in arginine biosynthesis are essential for growth, conidiogenesis, sexual reproduction, and pathogenicity in Magnaporthe oryzae. Microbiol Res 2015, 180, 11–22. [Google Scholar] [CrossRef]
- Chen, C.; Li, Q.; Wang, Q.F.; Lu, D.H.; Zhang, H.; Wang, J.; Fu, R.T. Transcriptional profiling provides new insights into the role of nitric oxide in enhancing Ganoderma oregonense resistance to heat stress. Sci Rep 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Huang, W.; Xiong, C.; Zhao, J. Transcriptome analysis reveals the role of nitric oxide in Pleurotus eryngii responses to Cd2+ stress. Chemosphere 2018, 201, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Chiang, K.T.; Shinyashiki, M.; Switzer, C.H.; Valentine, J.S.; Gralla, E.B.; Thiele, D.J.; Fukuto, J.M. Effects of nitric oxide on the copper-responsive transcription factor Ace1 in Saccharomyces cerevisiae: cytotoxic and cytoprotective actions of nitric oxide. Arch Biochem Biophys 2000, 377, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.; Demain, A.L. Bioactive products from fungi. Food Bioactives 2017, 2017 Jan 11, 59–87. [Google Scholar] [CrossRef]
- Zheng, W.F.; Zhang, M.M.; Zhao, Y.X.; Wang, Y.; Miao, K.J.; Wei, Z.W. Accumulation of antioxidant phenolic constituents in submerged cultures of Inonotus obliquus. Bioresour Technol 2009, 100, 1327–1335. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.Y.; Chai, R.Y.; Qiu, H.P.; Jiang, H.; Mao, X.Q.; Wang, Y.L.; Liu, F.Q.; Sun, G.C. An S-(hydroxymethyl) glutathione dehydrogenase is involved in conidiation and full virulence in the rice blast fungus. Plos One 2015, 10. [Google Scholar] [CrossRef]
Figure 1.
Summary diagram of NO production and function in fungi.
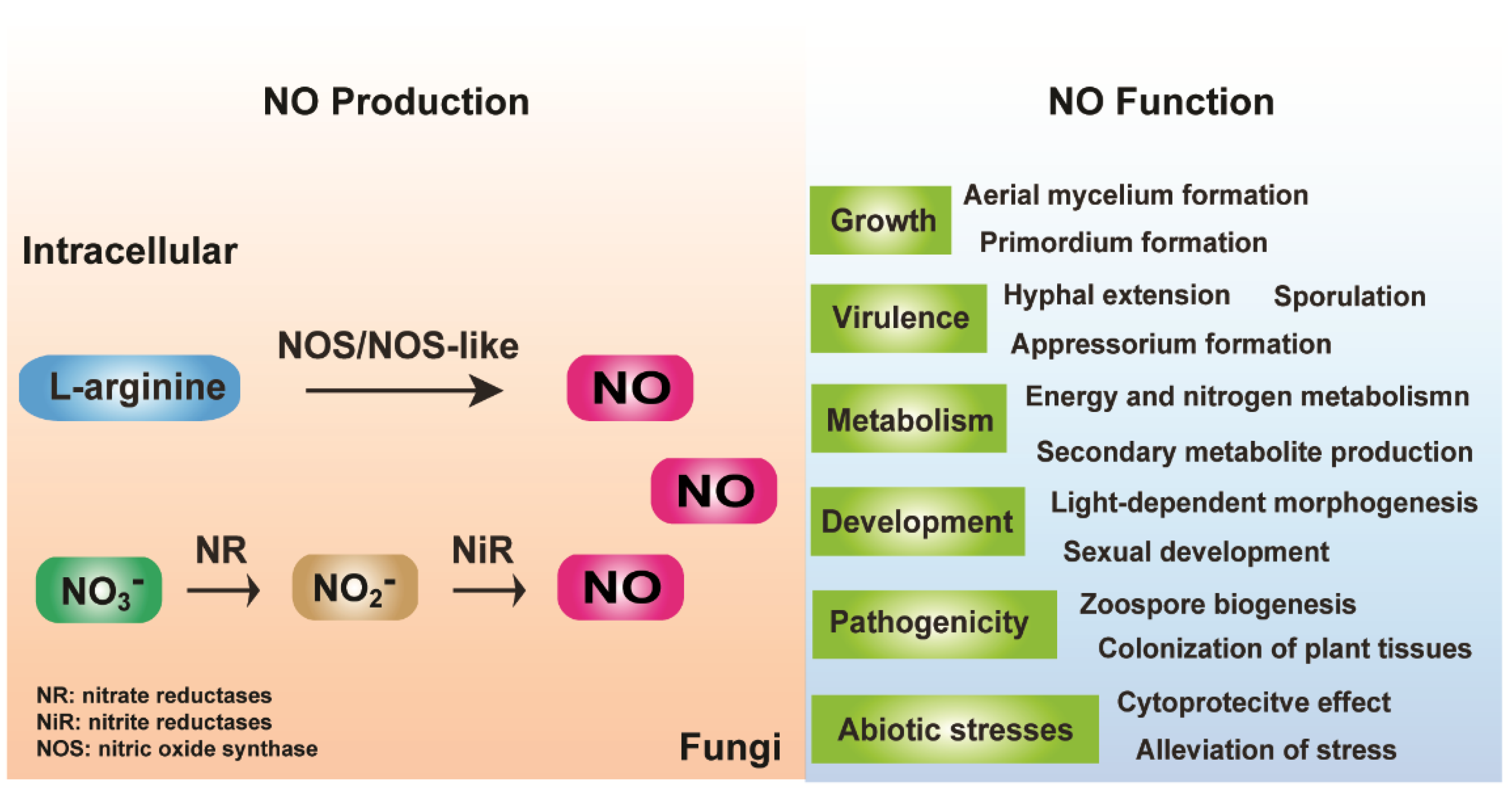

Table 1.
Mechanisms for NO synthesis in fungi.
| Fungus | Mechanism for NO Synthesis | Reference |
|---|---|---|
| Aspergillus nidulans | NO2- dependent: nitrate reductase activity | [51] |
| Blastocladiella emersonii | NOS dependent: enzyme activity | [37] |
| Blumeria graminis | NOS dependent: enzyme inhibition | [47] |
| Colletotrichum coccodes | NOS dependent: enzyme inhibition | [45] |
| Coniothyrium minitans | NOS dependent: enzyme activity / inhibition | [38,46] |
| Cryphonectria parasitica | NOS dependent: enzyme inhibition | [46] |
| Flammulina velutipes | NOS dependent: protein isolation and enzyme activity | [34] |
| Ganoderma lucidum | NO2- dependent: nitrate reductase activity | [52] |
| Inonotus obliquus | NOS dependent: enzyme inhibition | [48] |
| Inonotus obliquus cocultured with Phellinus morii | NOS dependent: gene identification and enzyme activity / inhibition | [39] |
| Macrophomina phaseolina | NOS dependent: gene identification and enzyme inhibition | [50] |
| Neurospora crassa | NOS dependent: enzyme inhibition | [44] |
| NOS dependent: enzyme activity | [41] | |
| Phycomyces blakesleeanus | NOS dependent: enzyme activity/inhibition | [35] |
| Pleurotus eryngii var. tuoliensis | NOS dependent: enzyme inhibition | [49] |
| Preussia sp. BSL-10 | Expression of NOS, nitrate reductase, and nitrite reductase | [53] |
| Saccharomyces cerevisiae | NOS dependent: enzyme activity | [36] |
| NOS dependent: protein isolation and enzyme activity/inhibition | [33] | |
| NO2- dependent: mitochondrial cytochrome c oxidase and nitrite reductase under hypoxia condition |
[54] | |
| Shiraia sp. Slf14 | NOS dependent: gene identification | [55] |
| NOS dependent: enzyme activity and gene expression NO2- dependent: nitrate reductase activity |
[42] | |
| Trichophyton rubrum | NOS dependent: enzyme activity | [40] |
Table 2.
Endogenous NO function in fungi.
| Function | Fungus | Reference | |
|---|---|---|---|
| Growth and development | Aspergillus nidulans | Reduced conidiation and induced the formation of cleistothecia | [51] |
| The light regulation of conidiation | [75] | ||
| Blastocladiella emersonii | Controlling zoospore biogenesis | [37] | |
| Candida albicans | Growth promotion and pathogenesis by extracellular vesicles | [78] | |
| Colletotrichum coccodes | Regulation of spore germination | [45] | |
| Coniothyrium minitans | The nitric oxide-mediated conidiation | [38,46] | |
| Neurospora crassa | The light induced conidiation and carotenogenesis | [41,44] | |
| Regulate mycelial development and conidia formation. | [76] | ||
| Impacting the growth and development of hyphae (vegetative growth) | [77] | ||
| Phycomyces blakesleeanus | The light-induced development of sporangiophores | [35] | |
| Physarum polycephalum | Sporulation | [79] | |
| Pleurotus ostreatus | Primordia formation | [74] | |
| Puccinia striiformis f.sp. tritici | Induce spore germination | [80] | |
| Response to stresses | Aspergillus fumigatus | Effects of antifungal agent (farnesol) on germination | [81] |
| Ganoderma lucidum | Heat stress -induced ganoderic acids levels | [82] | |
| Lentinula edodes and Grifola frondosa | Tolerance to superoptimal pH and in nitrogen-limitation | [83] | |
| Pleurotus eryngii var. tuoliensis | Heat stress-induced oxidative damage | [49] | |
| Heat stress-induced trehalose accumulation | [84] | ||
| Rhizophagus irregularis | Enhanced host plant tolerance to low temperature stress by regulating proline accumulation in plant | [43] | |
| Saccharomyces cerevisiae | Cytoprotective effect from heat-shock or high hydrostatic pressure | [85] | |
| Hypoxia signaling | [54,86] | ||
| H2O2-induced apoptosis | [36] | ||
| Shiraia sp. Slf14(w) | Heat stress enhanced perylenequinones biosynthesis | [55] | |
| Trichophyton rubrum | Reduction in fungal viability by 420-nm intense pulsed light | [40] | |
| Metabolism | Aspergillus nidulans | Mycotoxin production | [87] |
| Ganoderma lucidum | Methyl jasmonate -induced ganoderic acid biosynthesis | [52] | |
| Inonotus obliquus | Biosynthesis of antioxidant polyphenols / Accumulation of antioxidant phenolic constituents | [48] | |
|
Inonotus obliquus and Phellinus morii |
Increase in level of styrylpyrone polyphenols in fungal interspecific interaction | [39] | |
| Neurospora crassa | Cellulolytic enzyme production | [88] | |
| The pentose and glucuronate interconversion, fructose and mannose metabolism, galactose metabolism, amino and nucleotide sugar metabolism, arginine and proline metabolism and tyrosine metabolism | [77] | ||
| Preussia sp. BSL-10 | Improve rice plant growth and related gene expression | [53] | |
| Shiraia sp. S9 | Hypocrellin A production | [89,90] | |
| Shiraia sp. Slf14(w) | Production of secondary metabolite perylenequinone | [42,55] | |
| Virulence and pathogenicity | Aspergillus nidulans | Mycotoxin production | [87] |
| Blumeria graminis | Influences formation of the appressorium infection structure | [47] | |
| Botrytis cinerea | Saprophytic growth and plant infection | [91] | |
| Botrytis elliptica | Induction of programmed cell death in lily | [92] | |
| Magnaporthe oryzae | Drives plant infection (delays germling development and reduces disease lesion numbers) | [67] | |
| Conidial germination and appressorium formation (infectious morphogenesis) | [93] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



