Submitted:
11 December 2023
Posted:
18 December 2023
You are already at the latest version
Abstract
The mechanism of brain information processing unfolds within spatial and temporal domains inherently linked to the concept of symmetry. Biological evolution, beginning with the prevalent molecular chirality, results in the handedness of cognitive and psychological functions (the phenomena known as biochirality). The key element in the chain of chirality transfer from the downstream to upstream processes is the pyramidal neuron (PyrN) morphology-function paradigm (archetype). The most apparent landmark of PyrN is the geometry of the cell soma. However, “why/how PyrN soma gains the shape of quasi-tetrahedral symmetry” has never been explicitly articulated. Resolving the above inquiry is only possible based on the broad-view assumption that encoding 3D-space requires specific 3D-geometry of the neuronal detector and corresponding network. Accordingly, our hypothesis states that if the primary function of PyrNs, at the organism level, is sensory space perception, then the pyramidal shape of soma is the best evolutionary-selected geometry to support sensory-motor coupling.
Keywords:
Artificial Intelligence
; Bilaterality
; Biochirality
; Biological Intelligence
; Prevalent Molecular Chirality
; Pyramidal Neuron
; Evolution
; Biopsychology
; Cognitive Psychology
1. Introduction
Advances in molecular biology promise a new opportunity in developing AI devices based on bio-molecular building blocks (DNA and proteins) functioning in micro- and nano-scale. To achieve this goal, the broad-view revision of new results in diverse branches of biology is in demand.
Generalizing Darwinism to the Evolution of the Universe is a challenging and attractive idea, elevating the analytical description of bio-molecular, cellular, and organism levels of events at a new qualitative level. As an integrative approach, it is necessary and beneficial to understand the common principles of biological and non-biological existence, the hierarchical organization of biological systems, and the relations between biological and artificial intelligence. From the holistic view, space-time symmetry and relativity (STSR) are indispensable forms of existence, which can neither be created nor destroyed but only transformed, suggesting that physics and biology are imminently associated branches of science. The ground of the above-mentioned association is that the most common fundamental determinants of organic and non-organic objects are space, time, symmetry, and relativity. Below, we will focus on the spatial symmetry determinant in biology, known as biological chirality or biochirality (Figure 1). Mirror or reflection symmetry (also referring to chirality or handedness) is widely recognized as a fundamental mathematical tool and universal degree of freedom in physical and the principal concept in biology [1,2]. Accordingly, symmetry’s physical roots appearing in biological patterns are frequently addressed, and chirality centric view on physical and biological processes steadily gain recognition. [3,4]. It is a common agreement that animal morphogenesis is regulated by the gene regulatory networks and functional requirement [5]. Homochirality of biomolecules (DNA, RNA, and proteins) is linked to the origin of life, i.e., to the beginning of biological evolution (Figure 2). Consequently, the entire hierarchy of biological processes (including bilaterality) can be associated with the interplay of molecular, cellular, and higher orders of biological organization, influencing by initial molecular chirality, and factors related to the movement of organism [6,7].
2. Hierarchical Chain of Chirality Transfer
The stereospecific nature of living matter, beginning at the molecular level, from discriminating between the alternative enantiomers and culminating in the laterality of human cognitive functions, has long been viewed as the most striking feature challenging the curiosity of biologists, psychiatrists, and physicists [8]. The contemporary view on biochirality is based on the explicit integrative assessment of the spatial symmetry determinants persistently working in evolution and development. The association of molecular and cellular level biological processes with mental health and illness points to the reciprocal relations between two opposite poles of the biological hierarchy, molecular biology (I) and the phenomenon of the conscious mind (V). Biological psychology (biopsychology, psychobiology, psychophysiology, neuropsychobiology) and biological psychiatry represent the branches of science dealing with this fundamental link. The molecular and cell physiology of pyramidal neurons (PyrNs) provides a natural bridge between molecular biology and higher cognitive and psychological function. At the cellular level, biological processes within CNS are grounded in the physiology and function of PyrNs [9]. In biology, many legitimate questions still need appropriate answers. But some questions have not been ever expressed. One such inquiry is why/how PyrN, implicated in the spatial navigation of bilaterians, gains quasi-tetrahedral symmetry of soma. The objective reason for such silence is the combination of several factors. First. The long-time limited availability of molecular chiral discriminative tools. Second. An enormous volume of information describing the link between molecular chirality and brain laterality is dispersed in many scientific journals covering various sciences, from quantum physics and group theory to molecular biology and neurosciences. Third. The lack of a broader (generalized or philosophical) view on the subject matter (biochirality). This review is an attempt to improve the situation. In our view, the shape of soma is presumably associated with the tetrahedral geometry of homochiral biomolecular condensates. To my knowledge, the first attempt to approach this inquiry (examination) was made just recently [10]. In our view, the question should be resolved based on the understanding of the evolution of CNS [11]. Among the principal down-stream factors contributing to pathways of biomolecular condensates is the geometry of chiral carbon atoms in proteinogenic AAs. Tetrahedral (sp3-hybridized) carbons with four different substituents are the principal components of biological molecules exhibiting homochirality. Chiral of carbon atom promote the tetrahedral molecular assembly, observed also in DNA structures and structure of “biological” water (Figure 2) [12,13,14,15,16]. Unfortunately, the geometrical definition of soma shape was not explicitly clarified. Consequently, whether the pyramidal shape of soma was evolutionarily selected, or the current state is “an appendix of the CNS” has never been debated. Evolutionary selected biochemical homochirality is based on the chiral stereoselectivity of biosynthetic and metabolic reactions [17,18].
Chirality is the right keyword in the search for the integrity of PyrNs origin, structure, and functions. Such an assumption agrees that chiral asymmetry appears in nature at all levels, from elementary particles and AAs to mammals’ morphology and even galaxies’ levels [7,19,20]. The chirality of multi-dimensional elementary particles is the principal concern of string and gauge theories projecting beyond the Standard Model physics [21]. The well-known Feynman diagram reflect space-time-dependent symmetry transformations in the elementary particle events. The origin of chirality at the cellular level is attributed to biomolecular homochirality [21]. Molecular homochirality and cellular chirality are the internal “forces” driving an organism’s left–right asymmetric development [22,23] in bilaterians. The hierarchical chain of chirality transfer from the atomic to the organism level is one of the challenging targets of contemporary biological sciences [24,25,26]. The attention to the possible all-embracing role of the symmetry determinant in biology (from the biomolecular chirality to the bilaterality of human cognitive function) has become a significant trend in cognitive neuroscience [20,27,28,29]. However, surprisingly, in this chirality-oriented stream of biological research, the question “why/how does PyrNs gain tetrahedral geometry?” remains (as mentioned before) unarticulated. In this situation, the value of questioning exceeds the value of the particular hypothesis because the first question triggers the string of the next one. The essential secondary-wave question is “What is the mutual orientation of the corresponding PyrNs somas in the left and right hemisphere”.
Below, we introduce some relevant hypotheses based on the generalized view of biological symmetry. A broad view of biological symmetry assumes that chirality transfer from the molecular to macromolecular and cellular levels plays a vital role in physiology underlying perceptual and cognitive functions linked with the bilateral organisms’ emotional and behavioral expression [7,20,30,31]. The cellular and molecular mechanisms of psychological states are fast-developing branches of life science. From the time of Freud, Jung, and Assagioli, perceptual, cognitive, and behavioral functions have been considered the essential determinants of psychological processes [32,33,34]. Regardless of the view on the relationship between psychology’s cognitive and behavioral domains, it is a commonly agreed that both exhibit a hierarchy, with the bottom referencing the basic sensory and perceptual processes and the top referencing the higher levels of cognitive and executive functioning control. From the top view, the hierarchical structure is the chain of downstream processes, including motor functions, perceptual abilities, neuronal circuits, and underlying molecular biology. Interpreting sensory perception and motor functions within cellular and molecular biology relies significantly on the laws of space-time symmetry [7,35,36,37,38]. Following this logic, the shape and functions of PyrNs deserve specific attention. Before considering the downstream molecular determinants, let us review fundamental up-stream factors.
3. Space, Time, Symmetry, and Relativity
Throughout history, our notion of space and time has undergone dramatic transformations. Following the intuitive view of Euclid, Newton concluded that the external to the observer space is infinite, isotropic, uniform, perfectly penetrable, and immovable (i.e., absolute quantity) where he located the relative motion and studied its variation [39,40]. The Galilean principle of relativity formulated the first deviation from the intuitive interpretation of space and time. Later, the principle of relativity was complemented by the Noether theorem framing the fundamental role of symmetry. All advanced physical theories directly or indirectly employ an inevitable link between four determinants of nature: space, time, symmetry, and relativity (STSR), bringing a deeper interpretation to the concepts of thermodynamics and entropy. As a consequence, the concept of STSR has gained attention in biology.
Biological systems are structurally organized according to patterns repeated at each hierarchical level [41,42]. Notably and prominently, the most fundamental features, common for all hierarchical levels, are STSR and notion of chirality, as a specific form of geometric symmetry, plays a critical role [7,43,44]. In this article, we assume that the unifying concept of STSR reflects the fundamental properties of nature [26,45,46,47,48] and primary determinants of life [7,44,49,50]. Now we are prepared for an overview of the downstream molecular determinants.
4. Biological Evolution
4.1. Molecular Chirality
Despite the long history of attention, the origin of the single chirality of the Earth’s biosystem is still referred to as a great puzzle. Molecular homochirality (more precisely, prevalent molecular chirality), feeding the roots of the evolutionary tree, pre-determines its branching structure and behavior of each individual sub-division (Fig 2). Numerous theories of origin molecular (biological and non-biological) homochirality fall into two categories. One is searching for the possible coincidence of natural circumstances [51,52], and the second is for fundamental determinants of biological symmetry [7]. We will examine accumulating evidence in favor of indispensable causal force. Narrowing attention to the spatial symmetry at the molecular level, we can state that majority of biological molecules (such as AAs and sugars) and macromolecules (including DNA, RNA, proteins, and lipids) exhibit the prevalent form of spatial symmetry in three domains of geometry: chirality, fractality, and topology [53,54,55,56,57]. Enormous complexity of biochirality illustrated by the fact that entire complex of chiral molecules modulates diverse forms of cell chirality. An additional example of such complexity is the fact that molecular compounds of chiral and achiral structures frequently demonstrate strong chiro-optical and chiro-magnetic effects [58]. The most studied biomolecules implicated in the effects of chirality are proteinogenic AAs, enzyme-substrate protein complexes, and non-proteinogenic AAs in the CNS [59,60,61,62]. The chiral non-proteinogenic AAs play an essential role in the physiology of plants, insects, and animals. An example of non-proteinogenic chiral AAs is DOPA, which is part of the normal biology of some plants and all animals. Homochiral synthesis of L-DOPA isoform in humans occurs via biosynthesis from L-tyrosine (L-Tyr) by the tyrosine hydroxylase enzyme [63]. Protein stereochemistry is represented by two classes of stereo-isomers: chiral enantiomers (prevalent L-isoform) and achiral diastereomers (prevalent cis-stereo-form) (Figure 3A). The range of chirality-specific phase-transitions at the molecular level includes chiral inversion and spontaneous chiral symmetry breaking in diverse states of protein folding and aggregation. Evolution branched single-cell ancestors into two main domains of life: Procaryotes and Eukarya. Structurally and functionally, Eukaryotes are organized into more hierarchical levels than Prokaryotes (bacteria and archaea) [64]. Protein chirality is strongly associated with the chirality of membrane phospholipids. The main component of biological membranes glycerophospholipids are glycerol-based phospholipids that have differential chirality: D-glycerol for bacteria and eukaryotes and L-L-glycerol for archaea [65]. At the cellular level we restrict our attention to eukaryotes’ evolution, leading to the emergence of all neuronal cell types, including PyrNs. At the molecular level we primarily focused of the contribution of protein handedness to the shape of PyrNs. The prevalent molecular chirality of organisms at the protein level occurs through the fine-balanced interaction of L-(major) and D-(minor) isoforms. Up-regulation of D-AAs is implicated in organism aging [60,62,66], psychiatric disorders [67], and cancers [68,69]. The most known adverse impacts are attributed to the up-regulation of three AAs: D-aspartic acid, (D-Asp), D-serine (D-Ser), and D-alanine (D-Ala). All of them (and, potentially, by D-glutamate) are agonists for NMDA (N-methyl-D-aspartate) receptors of PyrNs [70,71,72,73,74,75]. Continuous spontaneous and induced assembly of homochiral proteins from simple to complex arrangements includes the tetrahedral complexes [76] (well-known in the hemoglobin conformational dynamics) [77,78]. Notably, the tetrahedral geometry of molecular compounds is not accidental but is the consequence of the symmetry of atomic electron orbitals. But how tetrahedral molecular complexes can result in the pyramidal shape of soma remains unknown. We will return to this question later in the text.
4.2. Link of Physiological and Psychological Functions
Before closely considering PyrNs morphology let’s analyze their contribution to upstream biological processes. The cellular ground of association between brain physiology and psychology is the function of PyrNs. Meaning that PyrNs provide an opportunity to basis of sensory processes (including visual, auditory, gustatory, olfactory, and cutaneous systems) to participate in the higher cognitive functions (such as emotion, learning, and memory) [79,80]. Indeed, populations of PyrNs observed in low-level sensory cortices including, primary somatosensory, visual, and auditory cortex, with PyrNs located in high-level areas such as the prefrontal cortex (PFC). Such a conceptual view allows us to trace a link between chirality-dependent molecular events in PyrNs and their role in perceptual, cognitive, and behavioral aspects of psychological functions (PFs) object perception or object recognition is the process of meaningful interpretation of the sensory input underlying our ability to act in the world. Psychophysical and physiological evidence suggests that object recognition relay on perceptual constancy, referring to the fact conservation of perceived geometrical characteristics of objects during relative motion of subject and object. Perceptual constancy is the invariance under spatial transformations and is closely associated with the mathematical definition of symmetries. Therefore, it is not accidental that CNS of bilateral animals possess the specific neuronal mechanism of symmetry / dissymmetry perception [81,82,83]. Prevalent molecular chirality plays pivotal role in human physiology. It can be illustrated by the regulation of intra-body dynamics of D-AAs. It is well known that D-Ser is involved in bio-chemical interaction of main organs of the body, including kidneys, liver, gut, urine, stomal, lung, and brain [11,18,84]. The dependence of CNS on D/L-AAs metabolism allows us to trace the interplay between physiology and complex of perceptual, cognitive, emotions, and psychological functioning [30,33,83,84,85,86,87,88]. Analyzing the types of personalities, Jung differentiated them based on the complex cognitive functions, including thinking, filing, sensation, intuition, and aesthetical judgment [27]. The expressions psychological functions (PFs), faculties, abilities, agents, states, conditions, and types are widely used in psychological sciences and to a lesser degree, in clinical practice.
4.3. Bilaterians: Symmetry-Function Interplay
The first bilateral organisms originate in invertebrates. In vertebrates, characterized by more diverse molecular homochirality, differentiation of all 3-D axes is promoted to more advanced forms, signifying progress in space perception, voluntary movement, and navigation. In humans the conjunction of cognitive and psychological functions allows projecting all aspect of molecular biology and space-perception of on the area of psychology and psychopathology [89,90]. Such a view brings us to the understanding that the bilateral CNS of humans unavoidably exhibits bilateral patterns of physiological, cognitive, emotional, and behavioral functions underlying the psychological state of individuals (in health and disease conditions). It means that every individual has a window for the spontaneous and intended inputs influencing the state of bilateral CNS. This is practically exploited in bilateral/unilateral activation of the sensory system and corresponding cognitive functions in the healing of post-traumatic stress disorders [90] and mediating fear conditions [91]. The symmetry in biology is extended to the balanced arrangement of body parts around the 3-D axis. The Bilateria or bilaterians are animals revealing bilateral symmetry (chirality) from the embryonic state. Neuroimaging studies reveal hemispheric asymmetry of association and limbic fiber tracts in utero [92,93]. The body plan of Bilateria has right and left sides that are mirror images of each other. The evolutionary origin of biochirality points to several major contributing factors. Three of them, relevant to the topic of our discussion, are prevalent molecular chirality, the necessity of space-time orientation of moving body with the possibility to distinguish left and right (we point to the left/right because it is related to a left-right mirror symmetry at the molecular level), a sensory system capable of space-time localization of objects representing food or social partner. From a biochirality view, it is essential to note that plants and animals, representing immobile and mobile organisms, exhibit different evolution pathways [94,95], which, in our view, are deferentially driven by the determinants of spatial symmetry. The focus of our consideration is specifically the evolution of mobile animal creatures. According to fossil evidence, bilateral body symmetry in animals took place about 500 million years ago. In the course of evolution, bilateral body inevitably leads to bilateral CNS. This fact convinces many scientists that the laterality of brain functions cannot be accidental [96]. Symmetry in CNS is the balanced and co-related spatial arrangement of molecular, cellular, and anatomical components, providing the opportunity for the laterality of cognitive functions. The laterality of cognitive functions experiences the “symmetry pressure” from two sides: from the internal determinants and behavioral-environmental determinants. One is strongly associated with the genetic mechanism, and another evidently with epigenetic factors [97]. Epigenetics represents the network of molecular interfaces mediating gene-environmental interactions [98]. In the language of symmetry, any epigenetic mechanism is the factor (symmetry changing factor) breaking genetically imposed homochirality of ribosomal protein synthesis [99]. Exploring human brain lateralization with attention to molecular chirality [29], cell chirality [100], genetics and non-genetic factors in association with the underlying physical laws concerning space-time symmetry confirms that the origin of relative chiral homogeneity of biological molecules is somehow connected to the origin and evolution of life [19,28,91]. For moving bilateral organisms, persistent asymmetry in the activation of the sensory system (as the vision in the birds) becomes the environmental factor of the development of perception, cognition, and action, and, consequently, the object of intuitive attraction (attention). In humans, this attraction takes various shapes, from pragmatic skills of spatial orientation to aesthetic feeling. At the cellular level molecular chirality is known as the factor promoting neuronal proliferation [101]. Space-time perception is the most influential force driving the direction of individual organism development and the directionality of biological evolution [19,102]. Both ontogeny and phylogeny can be traced at the molecular, cellular and organism levels. In the case of humans, the interplay of prevalent molecular chirality and the sensory-motor functions is what drives evolution. Hence, the sensory-motor system’s mechanism supports higher cognitive abilities, consciousness, and psychological state. The human brain is the integrative bilaterally asymmetric machinery of space-time perception of moving organisms accompanied by executive function. In the CNS, the link of molecular chirality to the organism’s function at the cell level is mediated by multiple subtypes of neurons with unique morphologies, electrical properties, and molecular identities [22,103].
4.4. Cell Chirality
4.4.1. Cell Evolution
Prokaryotic and eukaryotic cells are descended from a single primordial ancestor but have a specificity at morphological and molecular levels. At the morphological level, eukaryotes contain a nucleus separating genetic material from the cytoplasm and cytoplasmic organelles. At the molecular level, prokaryotic and eukaryotes share two major classes of informational macromolecules: nucleic acids and proteins. After splitting animal-plant-fungi-protists pathways, animal cells of bilaterians undergo several steps of additional modifications. Bilateral organisms acquire signaling proteins necessary for neuronal proliferation, formation synapse, and CNS, allowing efficient environmental navigation and movement. In this sense, the human CNS is the final product of evolution. The tuning of spatial navigation occurs through the development of internal factors (including molecular diversity) and the appearance of sophisticated sensory systems that allow communication with the environment These two constantly interacting factors are the primary determinants of specific neuronal morphology, differing from the shape-functions link of other cell types, such as red blood cells.
4.4.2. Pyramidal Neurons
PyrNs within bilateral organisms are studied in reptiles, fish, birds, and mammals (Figure 2) [104]. PyrNs gain their name due to the easily recognizable geometrical shape of the soma. PyrNs soma shape is sharply distinct from other neuronal (Purkinje and granular cells) and non-neuronal (microglia, astrocytes, epithelial, or red blood cell) cell types. Based on this fact, it is reasonable to assume that the specificity of PyrNs functions accounts for this difference. The term PyrNs refers to all major classes of excitatory multipolar glutamatergic cell types sharing common (pyramidal-like) soma shape despite the different degree of deviation from the perfect geometrical form [105]. Even in the publications explicitly devoted to cell-kind-dependent chirality and cell morphology [100,105,106], the shape of PyrNs soma is out of discussion. In the most detailed description of the PyrNs’ soma shape, it is usually characterized as a “teardrop or rounded pyramid” [107], extremely elongated rod-shaped” [108], and most frequently pyramid-shaped [109]. The base geometry of the pyramid (triangular or square) is never experimentally expored or theoretically predicted. One of the best experimental images of PyrNs shows the pyramidal shape of soma (Figure 3B.) [110]. However, confirmation of the tetrahedral geometry requires additional goal-oriented efforts. In such a situation, it is relevant to say that PyrNs soma has relative similarity with the pyramid’s geometry in general or tetrahedron, in particular. There are many arguments that the genetic chirality-centric architecture of the structural left-right asymmetry of the bilateral human brain is evolutionarily preserved for the optimum function of sensory perception of the spatial environment [111]. However, the tetrahedral shape of PyrNs may be induced by evolutionary tendency secondary to the sensory perception of the spatial environment. At the cellular level, space-time perception occurs through the activity of PyrNs in close collaboration with other cell types and first with the astrocytes. Astrocytes are gatekeepers for maintaining PyrN excitability by glutamate biosynthesis, uptake, and release [112,113], synthesis of lactate (taken by neurons for energy production [114]), and as the primary source of D-Ser [115]. The disruption of astrocyte functions triggers cross-talk of many neurodegenerative pathways sharing a common feature – prevalent molecular chirality, which is vulnerable to spontaneous racemization. The primary functions of PyrNs includes the evaluation of the distance, direction, and left-right discrimination of movement. In agreement with this view, the spatial arrangement of the PyrNs firing was found to be affected by environmental geometry [116]. Notable, that the aging-related cognitive deficits are attributable to reduced activity of PyrNs [117]. Left–Right discrimination of brain functions is well represented in the behavioral study [117,118]. This knowledge is supported by new results, showing brain asymmetry at the molecular and cellular levels, but additional studies are required. PyrNs are the primary excitatory multipolar cell type abundant in the brain cortex [117], hippocampus [119,120], and amygdala [121]. The heterogeneity of the PyrN family is defined by their distinct axonal projections, dendritic arborization, and types of receptors. But how the neuronal soma develops its pyramidal morphology, which directs the proper neurite orientation, was poorly understood for a long time. Examination of all traditional graphical representations of the PyrNs community suggests that most (if not all) studies assume coherent orientation of soma in the plane orthogonal to apical dendrites axis [122]. However, such an assumption requires experimental verification.
The cortical circuit network predominantly comprises pyramidal-to-pyramidal neuron connections, yet their assembly during embryonic development has yet to be entirely understood. PyrNs of cortex are distributed in the sensory, motor, association, and executive areas and found in all cortical layers except layer I [9,123,124]. Each PyrN receives input from thousands of excitatory synapses segregated onto dendritic branches. It has been previously proposed that sophisticated neuronal circuits associated with non-linear properties of dendrites enable cortical neurons to recognize multiple insequence patterns and robust sequence memory [125]. Dysfunctional PyrN circuitry has been associated with perception, cognition, and psychological conditions abnormalities. Glutamatergic signaling of PyrN occurs through neurotransmitters AAs L-glutamate (L-Glu) [126] and D-Ser [127] in close interaction with the complex of catecholaminergic systems {with corresponding neurotransmitters: L-dopamine, L-norepinephrine (noradrenaline), and L-epinephrine (adrenaline)} [128]. Despite the involved of neuronal circuits of different natures, the most significant contribution to the morphological asymmetry of brain hemispheres and left-right differentiation of neural pathways is attributed to PyrN. The fact that PyrN are the most populated neuronal type in the human cerebral cortex and hippocampus suggests their primary role in processing space-time information utilized in sensory-motor functions. PyrN signaling is necessary for normal development and essential functions of mature organisms [129]. While the distortion of neuronal geometry and formation of aberrant synapses are associated with pathological conditions [130], including impairment of visual perception [131] and mental retardation [132]. The bilateral cortex and hippocampus, containing the majority of PyrN, are studied in brain regions involved in a wide range of hemisphere-specific functions, including spatial coding, navigation, spatial memory, decision-making [133,134,135], and intelligence [136]. The evolutionary selected system for space-time information processing, including the morphology and spatial orientation of PyrN, is the fundamental feature underlying the function of CNS. Experimentally observed hemispheric asymmetry of synaptic morphology of PyrNs explains well-known functional laterality of human perceptual and cognitive functions [137,138,139,140,141]. The morphology of PyrNs concerning the function was the focus of long-term attention in neuroscience. The major studied structural features were dendritic arborization, synaptic connectivity, and axonal network [103,139,140,142]. Apical and basal segments of the dendritic tree, complemented by the relative orientation of presynaptic and postsynaptic neurons, were carefully studied [15,23,118,142,143,144,145,146,147,148]. Two dendritic arbors have distinct morphology, orientation and are involved in different synaptic circuits [118]. The dendritic orientation of PyrN is sublayer specific and exhibits dorsal-ven and front-back differentiation [149]. The experimental parameters characterizing the cell body include soma size, spatial distributions, the density of soma, and pyramidal somatic integrative zones. The shape and spatial orientation of pyramidal soma has little attention, partly due to the void of reliable experimental control. In bilateral organisms, beginning from C elegans, CNS contains PyrNs. During neurogenesis from the ventricular zone (VZ), before adopting pyramidal morphology, neurons pass through several intermediate stages (multipolar, bipolar) [150,151].
Presumably, many molecular correlates contribute to the pyramidal shape of soma, but all cell morphology alterations are supported/assisted by the molecular dynamics of homochiral enzyme-substrate complexes. A commonly accepted axiom is that molecular chirality drives cell chirality [152]. Experimental evidence suggests that such a chirality transfer occurs with the participation of diverse cytoskeleton-based long-lived structures. Indeed, it was shown that the interaction of the diverse family of cytoskeleton filaments (septin [153], actin [154,155], microtubules [156,157], and that neurofilaments (NF) [158] provides fundamental cell morphogenetic mechanisms, including the shape and spatial orientation of PyrNs soma [155]. It was shown that NF (and another intermediate filament proteins) contain in their in their N-terminal domains the motifs that bind unassembled tubulin. Peptides containing such motifs inhibit microtubules’ in vitro polymerization leading to altered cell shapes [159]. This fact suggests that NF-microtubules interaction can contribute to the shape of PyrN soma. The prominent candidate for attention is actin cytoskeleton. Actin filament network is involved in vital neuronal processes associated with various memory functions in different organisms, from invertebrates to mammals. The cellular processes, mediated by its activity, include cellular motility, division, intracellular transport, synaptic plasticity, and morphogenesis [160]. Actin and actin-binding proteins (α-actinin and synaptopodin) are present in the typical cisternal organelle of an axon initial segment (AIS) in subpopulation pyramidal neurons [161]. In agreement with our generalized function-based hypothesis, the recent publication shows that developing a pyramidally shaped soma is linked to septin functions [162]. So, molecular mechanisms involved in the neurogenesis and the development of pyramidal neurons soma are on the way to being clarified in detail.
The homochirality of actin-myosin cytoskeletons allows the cells to develop polarity and left-right asymmetry [152]. However, the bidirectional impact of prevalent molecular chirality (internal determinant) and bilaterality of CNS (window to the external epigenetic factors) on the pyramidality of PyrNs soma has never been considered. Notable that dendritic arborization exhibits cortical layer-dependent orientation preference towards the anterior orientation [146]. However, the spatial orientation of pyramidal soma in two brain hemispheres and their relation to space-time information processing have yet to be experimentally studied or theoretically discussed. Based on the lateralization of perceptual and cognitive functions, we can expect differential bilateral asymmetry in the morphology and orientation of PyrN. Indeed, currently, the hemispheric difference is experimentally observed in the number/volume [163] and synaptic organization [164]. Asymmetric hemispheric allocation of NMDA receptor subunits in hippocampal PyrN complements the whole picture [118,120]. The fact that PyrNs of the healthy human brain have a significantly greater density, larger size, and are more spherical in shape on the left- than on the right-side point to the meaningful link between two kinds of biological events [165]. Notable that bilateral asymmetry of brain activity indicates a state of the CNS system concerning mood and anxiety. For example, studies of brain EEG associated with PyrNs firing suggest that high levels of beta in the right hemisphere are associated with anxiety symptoms. In contrast, high levels of alpha in the left hemisphere indicates depressive features [166,167]. Studying PyrNs functions in biological information processing is necessary for designing A) strategy [104,168].
4.4.3. From Molecular to Cell Chirality
We consider the evolution of life in terms of the sequence of transformation of molecular chirality into cell chirality and bilaterality of organisms [22,25,169,170,171]. Our hypothesis highlights formal consequences of three co-existent events: molecular chirality, PyNs’ shape, and bilateral body-brain morphology [172]. Indeed, episodic memory, allowing mental navigation in space and time, is based on the hemisphere-asymmetrical activity of hippocampal PyrNs [134]. At the same time, from a geometrical standpoint, PyN soma (with four non-equivalent vertexes) represents a highly asymmetric (chiral) tetragonal structure. The anatomical symmetry and asymmetry of the PyrN structure imposes fundamental information processing capabilities. The essential point is that two mirror images of PyrNs (two seemingly identical PyrNs), located in the cortex or hippocampus, are not superimposable (i.e., not identical) (see Figure 3C). Furthermore, the chirality of PyrN provides a formal opportunity for hemisphere-specific information processing contributed by correlative protein and lipid chirality [7,173,174]. Currently, no data confirms or negates any hemispheric asymmetry (amount, content, or chirality) of lipids in animal brains. Examination of all traditional graphical representations of the PyrNs community suggests that most (if not all) studies assume coherent orientation of soma (withing brain hemisphere) in the plane orthogonal to apical dendrites axis (Figure 3D) [122]. However, such an assumption requires experimental verification.
5. Self-Assembly of Biomolecules
The idea of molecular chirality contribution to the soma-shape of PyrNs is based on the fact that organic and non-organic molecular nano-scale complexes can spontaneously adopt various shapes, including square-pyramidal [175], triangular-prism [176,177], octahedral, icosahedral., and tetrahedral [178,179,180,181] geometry. Indeed, after Le Bel and Van’t Hoff showed that the substituents of a tetravalent carbon center occupy the vertices of a tetrahedron, numerous experimental results illustrated that chiral covalent stereogenic units of molecular structures are responsible for the axial, planar, helical, and tetrahedral motifs in biological polymers [182,183]. Accumulating evidence shows the ability of biomolecules to self-assemble into the range of structures (from nanoscale to microscale) of distinct geometrical symmetry [184,185]. The mechanisms of self-assembly were studied for (DNA [186], nucleic acids [185], proteins [187], phospholipids [188], and their combinations. The most common DNA nanostructures are the tetrahedral DNA nanostructure (TDN). In the biological environment (cytosol), TDN exhibits maximum stability. The unique spatial structure of TDN allows it to penetrate cell membranes in abundance and regulate essential cellular processes, including proliferation, migration, and differentiation. Modifying TDN vertices, tetrahedral arms, or DNA tetrahedral cages by bioactive ligands enables TDN to be used as a nanocarrier for targeted therapies, molecular diagnosis, biosensing, antibacterial treatment, antitumor strategies, and tissue regeneration [186,189]. Peptides and proteins can also self-organize into highly ordered supramolecular architectures, such as nanofibrils, nanobelts, nanotubes, nanowires, and vesicles [190]. All facts mentioned above suggest the possibility of the natural occurrence of tetrahedral molecular aggregate in living cells as the molecular determinants of cell morphology and functions. A known example is the ferritin protein cages found in nearly all life forms. The most general structures show octahedral and tetrahedral symmetry [188]. Human ferritin cage has an outer diameter of ~12 nm and an inner cavity of 7–8 nm.
Molecular-Cellular Co-Evolution
Many physical and biological objects, including elementary particles (such as electrons and photons), molecular structures, and cell aggregates, have an intrinsic degree of freedom – chirality [184]. Evolutionary selected molecular chirality provides all available resources for the efficient mechanism of chirality transfers across all levels of biological organization [182]. Notably, the common symmetry principles guide the external and internal determinants of cell morphology, assembly, and functions of intracellular molecules. Understanding this fundamental commonality is necessary for neuroscience-inspired AI [191,192,193]. Protein assemblies adopting tetrahedral symmetry create shell-like architectures [194]. Such structures serve as enclosures for viral genomes and potentially can serve as internal determinants of cell shape. In addition to ferritins [187,195], well-known examples of tetrahedral protein assembly are small heat shock proteins (sHSPs) [196] and globular hemoglobin (Hb) [197,198,199,200,201]. Hb is a highly conserved globular protein in all life forms and functionally tied to aerobic organisms utilizing oxygen from the atmosphere and delivering it to cells. The expression of mitochondrial Hb (Hba-a2 and Hbb) in neurons (including nigral dopaminergic neurons, striatal γ-aminobutyric acid GABA-ergic neurons, and cortical PyrNs) was experimentally observed [199,200,201,202]. Along with natural (organic molecules, many artificial metal-organic tetrahedral (filled and porous) structures synthesized recently exhibit a chiral degree of freedom [203]. Porous molecular cages are successfully used for intracellular drug delivery [204]. Observation of amyloid beta peptides, which are prone to tetrahedral coordination of metal ions (including Cu, Zn, and Fe), suggest an active role of tetrahedral protein structures in cell physiology [204].
It is reasonable to assume that the shape of the soma can adapt to the demands of the cytoskeleton’s tetrahedral mesh-like gel, undergoing a chain of structural phase transitions [178,201,202]. In neuronal cells, the assembly of cytoskeleton proteins can be considered an internal determinant of soma’s shape. Cytoskeleton-based molecular (CMC) cages, as a form of membrane-bound and membrane-less organelles, are a well-known phenomenon in the cell physiology of intracellular segregation. CMC surrounds and protects various intracellular compartments, including the nucleus [205] and mitochondria [206], intracytoplasmic bacteria [Mostowy et al. 2010], and lipid droplets [207]. The diverse combination of cytoskeletal-based cages participates in intracellular physiology, including actin-based [205,206,208], vimentin based [209], septins-based [209,210] and microtubules based [211] complexes. However, no experimental evidence confirms the cytoskeleton’s direct involvement in the PyrN soma morphology.
In many cells type (including PyrN), the liquid-liquid phase separation, (LLPS) of biomolecular condensates (proteins and lipids assemblies) are involved in diverse functions, including an asymmetric cell division, the establishment of neuronal stem cell (NSC) polarity, neuronal proliferation, synaptic transmission and, likely, soma morphology of PyrNs. It became evident that sophisticated biophysical and biochemical processes govern cell-environment (interface) chirality sensation in a living system. Human embryonic stem cells (hESCs), differentiating into heart, gut, and brain tissues, show intrinsic [212] and extrinsic [213,214] cell chirality induced by the molecular (peptide-protein) chirality of intracellular and extracellular constituents. Accumulated experimental evidence accumulated for different cell types suggests that the pyramidal shape of neuronal soma is a direct result of such bidirectional impact. The size scales range for protein-lipid tetrahedral complexes can be from several nanometers to several microns [199,215,216,217,218,219,220,221,222]. Despite such a size is smaller than size of PyrNs soma, these experimental facts point to principal opportunity for the molecular structure to form the corners of pyramidal soma. From a formal geometrical view, it is possible to compose a 3-D tetrahedral structure of unlimited size (Figure 1] from the small tetrahedral units [221]. Notably, that chirality of elementary trihedron can be transferred to the lager-scale structure. Such tetrahedral units are known for biological and nonbiological molecules. Presently, three types of biological tetrahedral structures containing proteins, DNA, and a mixture of both are widely explored [222] (tetrahedral structures for phospholipids are the next target for exploration). Protein-based tetrahedral structures comprise helical, beta sheets, and other components [178]. Therefore, theoretically, all studied protein-based tetrahedral structures can be assembled in 3-dimensional cell-size complexes, supporting the shape of pyramidal neurons. When the base geometry of the pyramid (triangular or square) is never experimentally characterized or theoretically predicted, it is relevant to say that PyrNs soma is similar to the pyramid’s geometry in general or tetrahedron, in particular. However, additional experimental verification of such pathways is required. The co-appearance of all the above-mentioned spatial arrangements of PyrNs in the visual cortex suggests that the corresponding primary function is the involvement in the space-time orientation of moving organisms. In neuronal cells, the assembly of cytoskeleton proteins can be considered an internal determinant of soma’s shape.
6. Conclusions
The significance of spatial (i.e., geometrical) determinants in biology is supported by the fact that group theory is routinely used to describe the symmetries and conformational behavior of various systems ranging from the quantum mechanics of atoms, protein folding, neuronal signaling and brain functions levels to organism-environment interaction [223]. The waves of new scientific evidence bring the research community to the conclusion that there is no cell type, tissue, organ, or system of organism indifferent to the physiological balance of molecular chirality [25,224,225]. The mystery of PyrNs, in a broad sense, is on the way to be solved. The broad-view PyrN’s hypothesis will help to guide future studies. What remains is the need for complex, goal-oriented experimental advances, focusing on the complex of stereospecific effects illuminating the link between molecular chirality (such as of L-/D-actin cytoskeleton dynamics in dendritic spines or chirality of mGluR7 receptors/ligands complex), the shape of PyrNs, and laterality of sensory perception. We have introduced readers to the spectrum of facts and thoughts, the fundamental link of which is frequently hidden from the broad research audience. Now, it is time for your own judgment regarding the hypothesis on the consideration and for a new experimental design. Let’s point attention to two areas of science where PyrN’s hypothesis is in high demand. Until recently, one of the notable differences between biological and artificial systems were that biomaterials consist primarily of chiral molecules governing neuronal functions. Indeed, in the animal kingdom, the cognitive decision-making process is based on the same basic cellular elements of CNS – neurons as a unit integrating all advantages of prevalent molecular chirality to advance intelligence. Decades of molecular assembly-based nanotechnology linked with the chiral light-matter interaction dramatically changed the situation [226,227]. Evolutionary transitions between humans and AI fuel hope of symbiotic entities in the future and become a matter of fact [228]. In the prolonged period, the left-right asymmetry of the brain has been studied chiefly through psychological examination and behavioral study leaving its molecular and synaptic aspects of PyrNs largely unaddressed. In 2008, Shinohara and colleagues showed that “hippocampal CA1 pyramidal cell synapses differ in size, shape, and glutamate receptor expression depending on the laterality of presynaptic origin. CA1 synapses receiving neuronal input from the right CA3 PyrNs are larger and have more perforated PSD and a GluR1 expression level twice as high as those receiving input from the left CA3 ‘’ [229]. After the first convincing evidence, many great studies were done to link synaptic hemispheric asymmetry to function of PyrNs For all of them, the knowledge of hemisphere-specific soma orientation could be beneficial [121,230,231,232,233,234,235,236,237,238,239].
Author Contributions
Main contribution is done by Dyakin V.V. and Dyakina-Fagnano N.V.
Acknowledgments
The author gratefully thanks Pamela Butler, Stephen Ginsberg, and Csaba Vadasz for the valuable suggestions and corrections.
Conflicts of Interest
The authors declare no conflict of interest. Main contribution is done by Dyakin V.V. and Dyakina-Fagnano.
Abbreviations
Amino acids (AAs). Artificial Intelligence (AI). Biological Intelligence (BI). Central nervous system (CNS). Electroencephalogram (EEG). Human embryonic stem cells (hESCs). Liquid-liquid phase separation (LLPS). Neurofilaments (NFs). Neuronal stem cell (NSC). Pyramidal neurons (PyNs). Psychological functions (PFs). Space-time symmetry and relativity (STSR). D-aspartic acid, (D-Asp). D-serine (D-Ser). D-alanine (D-Ala). L-glutamate (L-Glu) D-proline (D-Pro). D-tyrosine (D-Tyr).
References
- Michael, G.; John, S. and Witten, Edward (2012). Superstring theory. Vol. 2: Loop amplitudes, anomalies and phenomenology. Cambridge University Press. 2019.
- Ma, Y.; Shi, L.; Yue, H. and Gao, G. Recognition at chiral interfaces: From molecules to cells. Colloids and Surfaces B: Biointerfaces. 2020, 195, 111268. [CrossRef]
- Li, R. and Bowerman, B. Symmetry Breaking in Biology Cold Spring Harb Perspect Biol. 2010, 2(3). a003475. [CrossRef]
- Reasoning for Symmetry in Biological Systems. SCE Scholary Community Encyclopedia, 2023. [CrossRef]
- Hollo, G. A new paradigm for animal symmetry. Interface Focus 2015, 5, 20150032. [CrossRef]
- Xin, Z.; Cai, Y.; Dang, L.T.’ Burke, H.M.S.; Revote, J.; Charitakis, N.; Bienroth, D.; Nim, H.T.; Li, Y-F. & Ramialison, M. Mona, GO: a novel gene ontology enrichment analysis visualisation system. BMC Bioinformatics 2022, 23(1), 69. [CrossRef]
- Dyakin V.V. Fundamental Cause of Bio-Chirality: Space-Time Symmetry—Concept Review. Symmetry 2023, 15(1), 79. [CrossRef]
- Palladino, P. Stereochemistry and the Nature of Life: Mechanist, Vitalist, and Evolutionary Perspectives. JSTOR, 1990, 81(1), 44-67, https://www.jstor.org/stable/234082.
- Kushchayev, S.V.; Moskalenko, V.F.; Wiener, P.C.; Tsymbaliuk, V.I.; Cherkasov, V.G.; Dzyavulska, I.V.; Kovalchuk, O.I.; Sonntag, V.K.H.; Spetzler, R.F. and Preul, M.C. The discovery of the pyramidal neurons: Vladimir Betz and a new era of neuroscience. Braim. A Journal of Neurology 2012, 135; 285–300. [CrossRef]
- Radler, M.R.; Liu, X.; Peng, M.; Doyle, B.; Toyo-Oka, K. and Spiliotis, E.T. Pyramidal neuron morphogenesis requires a septin network that stabilizes filopodia and suppresses lamellipodia during neurite initiation. Current Biology 2023, 33(3), 434-448.e8. [CrossRef]
- Sousa, A.M.M.; Meyer, K.A.; Santpere, G.; Gulden, F.O. and Sestan, N. Evolution of the Human Nervous System Function, Structure, and Development. Cell. 2017 Jul 13; 170(2): 226–247. [CrossRef]
- Dahanayake, J.N. and Mitchell-Koch, K.R. Entropy connects water structure and dynamics in protein hydration layer. Phys.Chem.Chem.Phys., 2018, 20, 14765. [CrossRef]
- Marvan, T.; Polák, M.; Bachmann, T. and Phillips, W.A. Apical amplification—a cellular mechanism of conscious perception? Neuroscience of Consciousness, 2021, 2021(2), 2021. [CrossRef]
- Kumar, P.; Buldyrev, S.V. and H. Eugene, H. A tetrahedral entropy for water. PNAS. 2009, 106 (52) 22130-22134. [CrossRef]
- Wang, Y.; Ye, M.; Kuang, X.; Li, Y. and Hu, S. A simplified morphological classification scheme for pyramidal cells in six layers of primary somatosensory cortex of juvenile rats. IBRO Rep. 2018, 74–90. [CrossRef]
- Luine, V. and Frankfurt, M. Interactions between estradiol, BDNF and dendritic spines in promoting memory. Neuroscience. 2013, 239, 34-45. [CrossRef]
- Mason, S.F. The development of concepts of chiral discrimination. Chirality 1989, 1(3), 183-191. [CrossRef]
- Bastings, J.J.A.J.; van Eijk, H.M.; Damink. S.W.O and Sander S. Rensen, S.S. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11(9), 2205. [CrossRef]
- Kondepudi, D. Chiral Asymmetry in Nature, Ch. 1 in Chiral Analysis (Second Edition). Advances in Spectroscopy, Chromatography and Emerging Methods 2018, pg. 3-28. [CrossRef]
- Cristadoro, G., Degli Esposti, M. & Altmann, E.G. The common origin of symmetry and structure in genetic sequences. Sci Reports 2018, 8, 15817. [CrossRef]
- Book by Ibanez, L.E and Uranga, A.M. String theory and particle physics: An introduction to string phenomenology. 688 pages. Cambridge University Press 2012.
- Inaki, M.; Liu, J. and Matsuno, K. Cell chirality: its origin and roles in left–right asymmetric development. Philos Trans R Soc Lond B Biol Sci. 2016, 371(1710). [CrossRef]
- Lin, YM. Creating chirality. Nat Chem Biol. 2008, 4, 330. [CrossRef]
- Takaoka, K.; Yamamoto, M.; and Hamada, H. Origin of body axes in the mouse embryo. Current Opinion in Genetics & Development 2007, 17(4}44-350. [CrossRef]
- Dyakin, V.V.; Lucas, J.; Dyakina-Fagnano, N.V.; Posner, E.P.; Vadasz, C. The Chain of Chirality Transfer as Determinant of Brain Functional Laterality. Breaking the Chirality Silence: Search for New Generation of Biomarkers; Relevance to Neurodegenerative Diseases, Cognitive Psychology, and Nutrition Science. Neurology and Neuroscience Research. 2017, 1(1),2. [CrossRef]
- Duan, P.; Cao, H.; Zhang, L. and Liu, M. Gelation induced supramolecular chirality: chirality transfer, amplification and application. Soft Matter. 2014, 10, 5428. [CrossRef]
- Vallortigara G. The evolutionary psychology of left and right: costs and benefits of lateralization Dev Psychobiol. 2006 Sep;48(6):418-27. [CrossRef]
- Francks, C. Exploring human brain lateralization with molecular genetics and genomics Annals of The New York Academy of Sciences Issue: The Year in Cognitive Neuroscience. 2015, 1359, 1-13. [CrossRef]
- Stacho, M. and Manahan-Vaughan, D. Mechanistic flexibility of the retrosplenial cortex enables its contribution to spatial cognition. Trends in Neuroscience. 2022, 45(4), P284-296. [CrossRef]
- Lee, T-W; Dolan, R.J.; and Critchley, H.D. Controlling Emotional Expression: Behavioral and Neural Correlates of Nonimitative Emotional Responses. Cerebral Cortex, 2008, 18(1), 104–113. [CrossRef]
- Yuan, J.; Lu, X.; Zhang, S.; Zheng, F.; Deng, Q.; Han, L. and Lu, Q. Molecular Chirality and Morphological Structural Chirality of Exogenous Chirality-Induced Liquid Crystalline Block Copolymers. Macromolecules 2022, 55, 5, 1566–1575. [CrossRef]
- Book by C.G. Jung. Psychological Types (The Collected Works of C. G. Jung, Vol. 6) (Bollingen Series XX). (Part of the Jung’s Collected Works (#6) Series and Dzieła (#2) Series). Publisher: Princeton University Press. 1976. 1977.
- Assagioli, A. Dynamic Psychology and Psychosynthesis. Publisher: Psychosynthesis Research Foundation, inc 1958. Roberto Assagioli.
- Myers, S. The five functions of psychological type. Analytical Psychology 2016, 61(2), 183-202. [CrossRef]
- Pillai, A.S. and Jirsa, V.K. Perspective. Symmetry Breaking in Space-Time Hierarchies Shapes Brain Dynamics and Behavior. Neuron 2017, 94(5), 1010-1026. [CrossRef]
- Jirsa, V. and Sheheitli, H. Entropy, free energy, symmetry and dynamics in the brain. Journal of Physics: Complexity 2022 3(1), 015007. [CrossRef]
- Dyakin, V.V. and Uversky, V.N. Arrow of Time, Entropy, and Protein Folding: Holistic View on Biochirality. Int J Mol Sci. 2022, 23(7):3687. [CrossRef]
- Iohnston, I.G.; Dingle, K.; Greenbury, S.F.; Camargo, C.Q.; Doye, J.P.K.; Ahnert, S.E. and Louis, A.A. Symmetry and simplicity spontaneously emerge from the algorithmic nature of evolution. Proc Natl Acad Sci U S A. 2022, 119(11), e2113883119. [CrossRef]
- Jammer, M. Concepts of Space: The history of Theories of Space in Physics, 3rd ed.; Dover: New York, NY, USA, 1993.
- Capecchi, D. Development of the Concept of Space up to Newton Encyclopedia 2022, 2(3), 1528-1544. [CrossRef]
- Nederbr, H. Hierarchical Organization of Biological Systems and the Structure of Adaptation in Evolution and Tumorigenesis. Journal of Theoretical Biology 1997, 184(2), 149-156. [CrossRef]
- Eronen, M.I. Levels of Organization in Biology. Sranf. Encycl. of Philos. 2023. {https://plato.stanford.edu/entries/levels-org-biology/}.
- Blackmond, D.G. The Origin of Biological Homochirality Cold Spring Harb Perspect Biol. 2010, 2(5): a002147. [CrossRef]
- Ocklenburg & Mundorf. Ocklenburg S. and Mundorf, A. Symmetry and asymmetry in biological structures Proc. Natl. Acad. Sci. U.S.A. PNAS. 2022, 119 (28), e2204881119. [CrossRef]
- Tasson. J.D. What Do We Know About Lorentz Invariance? Rep. Prog. Phys. 2014, 77, 062901. [CrossRef]
- Comte, C. Symmetry, relativity and quantum mechanics. Nuov Cim B 1996, 111, 937–956. [CrossRef]
- Field, J.N. Space-Time Exchange Invariance: Special Relativity as a Symmetry Principle. American Journal of Physics 2001, 69, 569. [CrossRef]
- Ajaltouni Z.J. Symmetry and relativity: From classical mechanics to modern particle physics. Natural Science. 2014, 6(4), Article ID:43343,7 pages. [CrossRef]
- Auffray, C. and Nottale, L. Review. Scale relativity theory and integrative systems biology: 1: Founding principles and scale laws. Progress in Biophysics and Molecular Biology 2008, 97(1), 79-114. [CrossRef]
- Noble, R.; Tasaki, K.; Noble, P.J. and Noble, D. Biological Relativity Requires Circular Causality but Not Symmetry of Causation: So, Where, What and When Are the Boundaries? Front. Physiol. Sec. Integrative Physiology 2019, 10. [CrossRef]
- Devínsky, F. Chirality and the Origin of Life. Symmetry 2021, 13(12), 2277. [CrossRef]
- Piñeros, W.D. and Tlusty, T. Spontaneous chiral symmetry breaking in a random driven chemical system. Nat Commun 2022, 13, 2244. [CrossRef]
- Cornelissen, J.J.L.M.; Rowan, A.E.; Nolte, R.J.M. and Sommerdijk, N.A.J.M. Chiral Architectures from Macromolecular Building Blocks Chem. Rev. 2001, 101, 12, 4039–4070. [CrossRef]
- Todoroff, N.; Kunze, J.; Schreuder, H.; Hessler, G.; Baringhaus, K-H. and Schneider, G. (2014). Fractal Dimensions of Macromolecular Structures. Mol Inform. 2014, 33(9): 588–596. [CrossRef]
- Wang, J., Panagiotou, E. The protein folding rate and the geometry and topology of the native state. Sci Rep 12, 6384. [CrossRef]
- Pandey, S., Mandal, S., Danielsen, M.B. et al. Chirality transmission in macromolecular domains. Nat Common 2022, 13, 76. [CrossRef]
- Kim, N.H., Choi, H., Shahzad, Z.M. et al. Supramolecular assembly of protein building blocks: from folding to function. Nano Convergence 2022, 9(4). [CrossRef]
- MacKenzie, L.E. and Stachelek, P. The twists and turns of chiral chemistry. Nat. Chem. 2021, 13, 521–522. [CrossRef]
- Dhanavade, M.J. and Sonawane K.D. Amyloid beta peptide-degrading microbial enzymes and its implication in drug design. 3 Biotech. 2020, 10(6), 247. [CrossRef]
- Reetz, M.T. and Garcia-Borràs, M. The Unexplored Importance of Fleeting Chiral Intermediates in Enzyme-Catalyzed Reactions. J. Am. Chem. Soc. 2021, 143, 37, 14939–14950. [CrossRef]
- Dyakin, V.V.; Dyakina-Fagnano, N.V.; Mcintire, L.B. and Uversky, V.N. Fundamental Clock of Biological Aging: Convergence of Molecular, Neurodegenerative, Cognitive and Psychiatric Pathways: Non-Equilibrium Thermodynamics Meet Psychology. Int J Mol Sci. 2022, 23(1), 285. [CrossRef]
- Dyakin, V.V.; Wisniewski T.M. and Lajtha, A. Chiral Interface of Amyloid Beta (Aβ): Relevance to Protein Aging, Aggregation and Neurodegeneration. Symmetry 2020, 12(4), 585. [CrossRef]
- Stansley, B. L and Yamamoto, B.K. L-dopa-induced dopamine synthesis and oxidative stress in serotonergic cells. Neuropharmacology. 2013, 67:243-51. [CrossRef]
- van Hooff, J.J.E. Towards unraveling the origins of eukaryotic nuclear genome organization. Trends in Cell Biology 2023, 33(10), 820-823. [CrossRef]
- Caforio, A. and Driessen, A.J.M. Archaeal phospholipids: Structural properties and biosynthesis. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 2017, 1862 (11), 1325–1339. [CrossRef]
- Fujii, N.; Takata, T.; Fujii,N.; Aki K. & Sakaue, H. D-Amino Acid Residues in Proteins Related to Aging and Age-Related Diseases and a New Analysis of the Isomers in Proteins Chapter in the book (pg 241-245) by Yoshimura, T., Nishikawa, T., Homma, H. (eds) D-Amino Acids Springer, Tokyo. [CrossRef]
- Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Kitagishi,Y. and Matsuda, S. Omar Cauli, Academic Editor and Soraya L. Valles, Academic Editor D-Amino Acids as a Biomarker in Schizophrenia. Diseases. 2022, 10(1), 9. [CrossRef]
- Jacco J.A.J. Bastings, Hans M. van Eijk, Steven W. Olde Damink,and Sander S. Rensen. D-amino Acids in Health and Disease: A Focus on Cancer. Nutrients. 2019, 11(9): 2205. [CrossRef]
- Murtas, G. and Pollegioni, L. D-Amino Acids and Cancer: Friends or Foes? Int. J. Mol. Sci. 2023, 24(4), 3274. [CrossRef]
- Wolosker, H., Balu, D.T. D-Serine as the gatekeeper of NMDA receptor activity: implications for the pharmacologic management of anxiety disorders. Transl Psychiatry 2020 10, 184. [CrossRef]
- Li, Y.; Han, H.; Yin, J.; Li, T.and Yina, Y. Role of D-aspartate on biosynthesis, racemization, and potential functions: A mini-review. Anim Nutr. 2018, 4(3): 311–315. [CrossRef]
- Saitoh, Y.; Katane, M.; Miyamoto, T.; Sekine, M.; Sakai-Kato, K. and Homma, H. D-Serine and D-Alanine Regulate Adaptive Foraging Behavior in Caenorhabditis elegans via the NMDA Recepto,r Journal of Neuroscience 2020, 40 (39) 7531-7544. [CrossRef]
- Kera, Y.; Aoyama, H.; Matsumura, H.; Hasegawa, Hisae Nagasaki, H. and Yamada, R. Presence of free D-glutamate and D-aspartate in rat tissues. Biochimica et Biophysica Acta (BBA) 1995, 1243(2), 282-286. [CrossRef]
- Mangas, A.; Coveñas,R.; Bodet, D.; Geffard, M.; Aguilar, L.A. and Yajeya, J. Immunocytochemical visualization of d-glutamate in the rat brain. Neuroscience 2007, 144(2), 654-664. [CrossRef]
- Lin CH, Yang HT, Lane HY (2019) d-glutamate, D-serine, and d-alanine differ in their roles in cognitive decline in patients with Alzheimer’s disease or mild cognitive impairment. Pharmacol Biochem Behav 2019,185: 172760. [CrossRef]
- Luo, Q.; Hou, C.; Bai, Y.; Wang, R. and Liu, J. Protein Assembly: Versatile Approaches to Construct Highly Ordered Nanostructures. Chem. Rev. (American Chemical Society) 2016, 116, 22, 13571–13632. [CrossRef]
- Riccio, A.; Vitagliano, L.; di Prisco, G.; Zagari, A. and Mazzarella, L. The crystal structure of a tetrameric hemoglobin in a partial hemichrome state. Proc Natl Acad Sci U S A. 2002, 99(15), 9801–9806. [CrossRef]
- Ha, C-E. and Bhagavan, N.V. Hemoglobin Chapter in Essentials of Medical Biochemistry. eBook ISBN: 9780124166974. Sec Ed. Elsevier 2015.
- Goldstein, E. B. Crosstalk between psychophysics and physiology in the study of perception. In E. B. Goldstein (Ed.). Blackwell handbook of perception (pp. 1–23). Blackwell Publishing. 2001.
- Xu, X.; Hanganu-Opatz,I.L. and Bieler, M. Cross-Talk of Low-Level Sensory and High-Level Cognitive Processing: Development, Mechanisms, and Relevance for Cross-Modal Abilities of the Brain. Front. Neurorobot., 2020, 14. [CrossRef]
- Logothetis, N.K. and Pauls, J. Psychophysical and Physiological Evidence for Viewer-centered Object Representations in the Primate. Cerebral Cortex 1995, 3, 270-288, 1047-3211/95/S4.OO. [CrossRef]
- Mascalzoni, E.; Osorio, D.; Regolinm L. and Giorgio Vallortigara, G. Symmetry perception by poultry chicks and its implications for three-dimensional object recognition. Proc Biol Sci. 2012. 7;279(1730), 841-6. [CrossRef]
- Pizlo, Z. and de Barros, J. A. The Concept of Symmetry and the Theory of Perception Front. Comput. Neurosci., 2021, 15. [CrossRef]
- Nakade, Y.; Iwata,Y.; Furuichi, K.; Mita, M.; Hamase, K.; Konno, R. et al. Gut microbiota-derived D-serine protects against acute kidney injury. The Journal of Clinical Investigation Insight 2018 18, 3(20), e97957. [CrossRef]
- Freud S. An outline of psychoanalysis. Std. Edn. Vol. 23. London: Vintage. 1940.
- Wada, K.; Yamamoto, M. and Nakashima, K. Psychological function in aging. Nihon Rinsho (Janan) 2013, 71(10).1713-9.
- Bottaccioli, A.G.; Bologna, M. and Bottaccioli, F. Psychic Life-Biological Molecule Bidirectional Relationship: Pathways, Mechanisms, and Consequences for Medical and Psychological Sciences—A Narrative Review. Int. J. Mol. Sci. 2022, 23, 3932. [CrossRef]
- Corballis, M. C., & Beale, I. L. (1970). Bilateral symmetry and behavior. Psychological Review, 77(5), 451–464. [CrossRef]
- Delvenne, J-F.; Castronovo, J.; Demeyere, N. and umphreys, G.W. Bilateral Field Advantage in Visual Enumeration PLOS 2011. [CrossRef]
- Tamaki Amano and Motomi Toichi. Zoi Kapoula, Editor. The Role of Alternating Bilateral Stimulation in Establishing Positive Cognition in EMDR. Therapy: A Multi-Channel Near-Infrared Spectroscopy Study. PLoS One. 2016; 11(10): e0162735. [CrossRef]
- Boukezzi, S.; Silva, C.; Nazarian, B.; Rousseau, P-F.; Guedj, E. and Valenzuela-Moguillansky, C. Bilateral Alternating Auditory Stimulations Facilitate Fear Extinction and Retrieval. Front. Psychol. Sec. Psychology for Clinical Settings 2017, 8. [CrossRef]
- Kasprian G, Langs G, Brugger PC, Bittner M, Weber M, Arantes M, et al. The prenatal origin of hemispheric asymmetry: an in-utero neuroimaging study. Cereb Cortex. 2011, 21(5), 1076–83. pmid:20851852. [CrossRef]
- Mitter, C.; Prayer, D.; Brugger, P.C.; Weber, M. and Kaspria, G. In Vivo Tractography of Fetal Association Fibers. PLOS. ONE 2015, 10(3), e011953. [CrossRef]
- Nakamura, M. and Hashimoto, T. Mechanistic Insights into Plant Chiral Growth. Symmetry 2020, 12(12), 2056. [CrossRef]
- Zagórska-Marek, B.; Sokołowska, K. and Turzańska, M. Chiral events in developing gametophores of Physcomitrella patens and other moss species are driven by an unknown, universal direction-sensing mechanism. Am. J. of Botany 2018. 105(12), 1986-1994. [CrossRef]
- Than, K. 2005. Symmetry in Nature: Fundamental Fact or Human Bias. Live Science. 2005.{https://www.livescience.com/4002-symmetry-nature-fundamental-fact-human-bias.html}.
- Gunturkun, O and Ocklenburg, S. Ontogenesis of Lateralization. Neuron, 2017, 94(2), 249–263. [CrossRef]
- Mehler, M.F. Epigenetic Principles and Mechanisms Underlying Nervous System Functions in Health and Disease. Prog Neurobiol. 2008, 86(4): 305–341. [CrossRef]
- Zion, E. and Chen, X. Breaking Symmetry: The Asymmetries in Epigenetic Inheritance. Biochem (Lond). 2021. 43(1): 14–19. [CrossRef]
- Wan, L.Q.; Chin, A.S.; Worley, K.E. and Ray, P. Cell chirality: emergence of asymmetry from cell culture. Philos Trans R Soc Lond B Biol Sci. 2016, 371(1710), 20150413. [CrossRef]
- Sun. Na.; Dou, X.; Tang, Z.; Zhang, D.; Ni, N.; Wang, J.; Gao, H.; Ju, Y.; Dai, X.; Zhao, C.; Gu, P.; Ji, J. and Feng, C. Bio-inspired chiral self-assemblies promoted neuronal differentiation of retinal progenitor cells through activation of metabolic pathway. Bioactive Materials 2021, 6(4), 990-997. [CrossRef]
- Dong, C., Madar, A.D. & Sheffield, M.E.J. Distinct place cell dynamics in CA1 and CA3 encode experience in new environments. Nat Commun 2021, 12, 2977. [CrossRef]
- Graham, H.K. and Duan, X. Molecular mechanisms regulating synaptic specificity and retinal circuit formation. Wiley Interdiscip Rev Dev Biol. 2021, 10(1), e379. [CrossRef]
- Striedter, G.F. Review. Special Issue: Cortical Evolution. Evolution of the hippocampus in reptiles and birds. JCN (Res. in System Neuroscience) 2016, 524(3), 496-517. [CrossRef]
- Matho,K.S; Huilgol, D.; Galbavy, W.; He, M.; Kim, G.; Xu An, Xu.; Lu, J. et al. Genetic dissection of the glutamatergic neuron system in cerebral cortex. Nature. 2021, 598, 7879):182-187. [CrossRef]
- Zhang, H. and Wan, L.Q. Cell Chirality as A Novel Measure for Cytotoxicity 2022, 6(1): e2101088. [CrossRef]
- Bekkers. J.M. Pyramidal neurons. Current biology Curr Biol. 2011, 21(24), R975. [CrossRef]
- Banovac, I.; Sedmak, D.; Džaja, D.; Jalšovec, D.; Milošević, N.J.; Rašin, [M.R. and Petanjek, Z. Somato-dendritic morphology and axon origin site specify von Economo neurons as a subclass of modified pyramidal neurons in the human anterior cingulate cortex. Journal of Anatomy 2019, 235, 3, 651-666. [CrossRef]
- Johns, P. Neurons and glial cells. Chapter 5 in Clinical Neuroscience, 2014, 61-69. [CrossRef]
- Barger, N.; Sheley, MF. and Schumann, C.M. Stereological study of pyramidal neurons in the human superior temporal gyrus from childhood to adulthood. J Comp Neurol. 2015, 523(7), 054–1072. [CrossRef]
- Sha, Z.; Schijven, D.; Carrion-Castillo, A.; Joliot, M.; Mazoyer, B.; Fisher, S.E.; Crivello, F. and Francks, C. The genetic architecture of structural left-right asymmetry of the human brain. Nature Human Behaviour 2021, 5, 1226–1239. [CrossRef]
- Ottersen, O.P.; Zhang, N. and Walberg, F. Metabolic compartmentation of glutamate and glutamine: morphological evidence obtained by quantitative immunocytochemistry in rat cerebellum. Neuroscience. 1992, 46(3),519-34. [CrossRef]
- Sanghai, N. and Tranmer, G.K. Biochemical and Molecular Pathways in Neurodegenerative Diseases: An Integrated View. Cell 2023, 12(18), 2318. [CrossRef]
- Rabinowitz, J.D. and Enerbäck, S. Lactate: the ugly duckling of energy metabolism Nat Metab. 2020, 2(7): 566–571. [CrossRef]
- [Sultan et al. 2015] Sultan, S.; Li, L.; Moss, J.; Petrelli, F.; Cassé, F.; Gebara, E.; Lopatar, J.; Pfrieger, F.W.; Bezzi, P.; Bischofberger, J.; et al. Synaptic Integration of Adult-Born Hippocampal Neurons Is Locally Controlled by Astrocytes. Neuron 2015, 88, 957–972. [CrossRef]
- Krupic, J.; Bauza, M.; Burton, S.; Barry,C. and O’Keefe, J. Grid cell symmetry is shaped by environmental geometry. Nature. 2015, 518(7538): 232–235. [CrossRef]
- Oh, M.M.; Simkin, D. and Disterhoft, J.F. Intrinsic Hippocampal Excitability Changes of Opposite Signs and Different Origins in CA1 and CA3 Pyramidal Neurons Underlie Aging-Related Cognitive Deficits. Front. Syst. Neurosci. 2016, 10. [CrossRef]
- Constant M. and Mellet, E. The Impact of Handedness, Sex, and Cognitive Abilities on Left–Right Discrimination: A Behavioral Study. Front. Psychol. 2018, 9, 405. [CrossRef]
- Merrick, C.M.; Dixon, T.C.; Breska, A.; Lin, J.; Chang, E.F.; King-Stephens, D.; Laxer, K.D.; Weber, P.B.; Carmena, J.; Knight, R.T. and Ivry, R.B. Left hemisphere dominance for bilateral kinematic encoding in the human brain. eLife 2022, 11, e69977. [CrossRef]
- Jiang, S., Guan, Y., Chen, S. et al. Anatomically revealed morphological patterns of pyramidal neurons in layer 5 of the motor cortex. Sci Rep 2020, 10, 7916. [CrossRef]
- Kawakami, R.; Shinohara, Y.; Kato, Y.; Sugiyama, H.; Shigemoto, R, and Itoauthors, I. Asymmetrical allocation of NMDA receptor ε2 subunits in hippocampal sircuitry. Science 2003, 300(5621), 990-994. [CrossRef]
- Galakhova, A.A; Hunt, A.S.; Wilbers, R.; de Kock, C.P.J.; Mansvelder, H.D.; Goriounova, N.A. et al. Evolution of cortical neurons supporting human cognition. Trands in Cognit. Science 2022. [CrossRef]
- Shinohara, Y.; Hirase, H.; Watanabe, M, Makoto Itakura, M.; Mmi Takahashi, M.and Shigemoto, R. Edited by Huganir, R.L. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. PNAS. 2008,105 (49) 19498-19503. [CrossRef]
- Muller, J.F.; Mascagni, F. and McDonald. A.J. Pyramidal Cells of the Rat Basolateral Amygdala. Synaptology and Innervation by Parvalbumin-immunoreactive Interneurons. J Comp Neurol. 2006, 494(4): 635–650. [CrossRef]
- Lorente de Nó R, L. Cerebral cortex: architecture, intracortical connections and motor projections. In: Physiology of the nervous system, 3rd edn (Fulton JF, ed.), pp. 288–330. Oxford: Oxford University Press. 1949.
- Chan, C.H.; Godinho, L.N.; Thomaidou, D.; Tan, S.S.; Gulisano, M and Parnavelas J.G. Emx1 is a Marker for Pyramidal Neurons of the Cerebral Cortex. Cerebral Cortex 2001, 11(12), 1191–1198. [CrossRef]
- Hawkins, J. and Ahmad, S. Hypothesis and theory article. Why Neurons Have Thousands of Synapses, a Theory of Sequence Memory in Neocortex. Front. Neural Circuits, 2016, 10. [CrossRef]
- Jia, W.; Hu, C.; Wang, Y.; Liu, Y.; Wang, L.; Zhang, S.; Zhu, Q.; Gu, Y.; Zhang, P.; Ma, J.; Chen, et al. Identification of Single-Molecule Catecholamine Enantiomers Using a Programmable Nanopore. ACS Nano 2022, 16, 4, 6615–6624. [CrossRef]
- Parnavelas JG.; Dinopoulos A. and Davies SW. The central visual pathways. In: Handbook of chemical neuroanatomy, vol. 7. Integrated systems of the CNS, Part II (Björklund A, Hökfelt T, Swanson LW, eds), pp. 1–164. Amsterdam: Elsevier. 1989.
- Dallérac, G.; Li, X.; Lecouflet, P.; Morisot, N.; Sacchi, S.; Asselot, R.; Thu Ha Pham, T.H. et al. Dopaminergic neuromodulation of prefrontal cortex activity requires the NMDA receptor coagonist D-serine. Proc Natl Acad Sci U S A. 2021, 118(23): e2023750118. [CrossRef]
- O’Keefe J. and Krupic, J. Do hippocampal pyramidal cells respond to nonspatial stimuli? Physiological review 2021, 101(3), 1427-145. [CrossRef]
- Purpura, D.P. and Suzuki, K. Distortion of neuronal geometry and formation of aberrant synapses in neuronal storage disease. Brain Research 1976, 116(1), 1-2. [CrossRef]
- Adámek, P.; Langová, V. & Horáček, J. Early-stage visual perception impairment in schizophrenia, bottom-up and back again. Schizophrenia 2022 8, 27. [CrossRef]
- Granato, A. and De Giorgio, A. Alterations of neocortical pyramidal neurons: turning points in the genesis of mental retardation. Front. Pediatr. 2014. [CrossRef]
- Rennó-Costa, C. and Adriano Tort, A.B.L. Place and Grid Cells in a Loop: Implications for Memory Function and Spatial Coding. J Neurosci. 2017, 37(34), 8062-8076. [CrossRef]
- Kitanishi, T., Ito, H.T., Hayashi, Y. et al. Network mechanisms of hippocampal laterality, place coding, and goal-directed navigation. J Physiol Sci 2017, 67, 247–258. [CrossRef]
- Zhong, S.; He, Y.; Shu, H. and Gaolang Gong, G. Developmental Changes in Topological Asymmetry Between Hemispheric Brain White Matter Networks from Adolescence to Young Adulthood. Cerebral Cortex, 2017, 7(4), 2560–2570. [CrossRef]
- Goriounova, N.A.; Heyer, D.B.; Wilbers, R.; Verhoog, M.B.; Giugliano, M.; Christophe Verbist, C. et al. Large and fast human pyramidal neurons associated with intelligence. Elife. 2018, 18(7), e41714. [CrossRef]
- Shipton, O. A., El-Gaby, M., Apergis-Schoute, J., Deisseroth, K., Bannerman, D. M., Paulsen, O., & Kohl, M. M. Left–right dissociation of hippocampal memory processes in mice. PNAS 2014, 111(42), 15238–15243. [CrossRef]
- Sá, M.J.; Ruela, C. and Madeira, M.D. Dendritic right/left asymmetries in the neurons of the human hippocampal formation: a quantitative Golgi study. Arq Neuropsiquiatr 2007, 65(4B), 1105-13. [CrossRef]
- Shinohara, Y.; Hirase, H.; Watanabe, M.; +2, and Ryuichi Shigemoto, R. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. PNAS 2008, 105 (49) 19498-19503. [CrossRef]
- Goto, K.; Kurashima R.; Gokan, H.; Inoue, N.; Ito, I. and Watanabe, S. Left−Right Asymmetry Defect in the Hippocampal Circuitry Impairs Spatial Learning and Working Memory in iv Mice. PLOS ONE 2010, 5(11), e15468. [CrossRef]
- Ukai, H.; Kawahara, A.; Hirayama, K.; Show all 17 Ito Isao, I. ItPirB regulates asymmetries in hippocampal circuitry. PLOS ONE 2017, 12(6). [CrossRef]
- Luengo-Sanchez, S.; Bielza, C.; Benavides-Piccione, R.; Fernaud-Espinosa, I.;DeFelipe, J. and Larrañaga, P. A univocal definition of the neuronal soma morphology using Gaussian mixture models. Front. Neuroanat., 2015, 9. [CrossRef]
- Rockland, K.S. Pyramidal Neurons: Looking for the origins of axons. Evolutionary Biology. Neuroscience 2022. [CrossRef]
- Abrous, D.N., Koehl, M. & Lemoine, M. A. Baldwin interpretation of adult hippocampal neurogenesis: from functional relevance to physiopathology. Mol Psychiatry 2022. 27, 383–402. [CrossRef]
- Leguey, I.; Benavides-Piccione, R.; Rojo, C.; Larrañaga, P.; Bielza, C. and DeFelipe, J. Patterns of Dendritic Basal Field Orientation of Pyramidal Neurons in the Rat Somatosensory Cortex. eNeuro 2018, 5 (6). [CrossRef]
- Weiler, S.; Nilo, D.G.; Bonhoeffer, T.; Hübener, M.; Rose, T. and Scheuss, V. Orientation and direction tuning align with dendritic morphology and spatial connectivity in mouse visual cortex. Current Biology 2020, 32(8), 1743-1753.e7. [CrossRef]
- Wahle, P.; Sobierajski, E.; Gasterstädt, I.; Lehmann,N.; Weber, S.; Lübke, H.R.; Engelhardt, M.; Distler, C. and Meyer, G. Neocortical pyramidal neurons with axons emerging from dendrites are frequent in non-primates, but rare in monkey and human. eLife. 2022, 11, e76101. [CrossRef]
- Musall, S., Sun, X.R., Mohan, H. et al. Pyramidal cell types drive functionally distinct cortical activity patterns during decision-making. Nat Neurosci 2023. [CrossRef]
- Park, J.; Papoutsi, A.; Ash, R.T. et al. Contribution of apical and basal dendrites to orientation encoding in mouse V1 L2/3 pyramidal neurons. Nat Commun 10, 5372 (2019). [CrossRef]
- Elaine M. Pinheiro, E.M.; Xie, Z.; Norovich, A.L.; Vidaki, M.; Tsai, L-H. and Gertler, F.B. Lpd depletion reveals that SRF specifies radial versus tangential migration of pyramidal neurons. Nat Cell Biol. 2011, 13(8), 989–995. [CrossRef]
- Hobert, O. Development of left/right asymmetry in the Caenorhabditis elegans nervous system: From zygote to postmitotic neuron. Genesis 2014, 52(6), 528-43. [CrossRef]
- Tee, Y., Shemesh, T., Thiagarajan, V. et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nature Cell Biolohy 2015, 17, 445 – 457. [CrossRef]
- Falk,J.; Boubakar, L. and Valérie Castellani, V. Septin functions during neuro-development, a yeast perspective Current Opinion in Neurobiology 2019. 57, 102-109. [CrossRef]
- Inaki, M.; Sasamura. T. and Kenji Matsuno, K. Review. Cell Chirality Drives Left-Right Asymmetric Morphogenesis. Front. Cell Dev. Biol. 2018, 6. [CrossRef]
- Konietzny,A.; Bär, J. and Mikhaylova, M. Review. Dendritic Actin Cytoskeleton: Structure, Functions, and Regulations. Front. Cell. Neurosci. Sec. Cellular Neurophysiology 2017, 11. [CrossRef]
- Satir, P. Chirality of the cytoskeleton in the origins of cellular asymmetry Philos Trans R Soc Lond B Biol Sci. 2016, 371(1710), 20150408. [CrossRef]
- Parato, J. and Bartolini, F.The microtubule cytoskeleton at the synapse Neurosci Lett. 2021, 14;753,135850. [CrossRef]
- Lamprecht, R. The actin cytoskeleton in memory formation. Progress in Neurobiology 2014, 117, 1-19. [CrossRef]
- Sánchez-Ponce, D.; Blázquez-Llorca, L.; DeFelipe, J.; Garrido, J.J. and Muñoz, A. Colocalization of α-actinin and Synaptopodin in the Pyramidal Cell Axon Initial Segment. Cerebral Cortex 2012, 22(7), 1648–1661. [CrossRef]
- Radler, M.R.; Liu, X.; Peng, M.; Doyle, B. and Toyo-Oka, K. Pyramidal neuron morphogenesis requires a septin network that stabilizes filopodia and suppresses lamellipodia during neurite initiation. Curr Biol. 2023 Feb 6;33(3):434-448.e8. [CrossRef]
- Bocquet, A.; Berges, R.; Frank, R.; Robert, P.; Peterson, A.C. and Eyer, J. Neurofilaments Bind Tubulin and Modulate Its Polymerization. J Neurosci. 2009, 29(35), 11043–11054. [CrossRef]
- Uylings, H.B.; Jacobsen, A.M.; Zilles, K. and Amunts, K. Left-right asymmetry in volume and number of neurons in adult Broca’s area. Cortex. 2006, 42(4), 652-8. [CrossRef]
- El-Gaby, M.; Reeve, H.M.; Lopes-dos-Santos,V.; Campo-Urriza, N.; Perestenko, P.V. Morley, A.; Strickland, L.A.M.; Lukács, I.P.; Paulsen, O. and Dupret, D. An Emergent Neural Coactivity Code for Dynamic Memory. Nat Neurosci. 2021, 24(5): 694–704. [CrossRef]
- Cullen, T.J.; Walker, M.A.; Eastwood, S.L.; Esiri, M.M.; Harrison, P.J. and Crow, T.J. Anomalies of asymmetry of pyramidal cell density and structure in dorsolateral prefrontal cortex in schizophrenia Br J Psychiatry 2006, 188:26-31. [CrossRef]
- Souta, R. An Introductory Perspective on the Emerging Application of qEEG. In the book Introduction to Quantitative EEG and Neurofeedback. Advsanced Theory and Applications. By Budzynski, T.H.; Washington, P.; Budzynski, H.K.; Washington, P.; Evans, J.R. and Abarbanel, A. Sec Ed. Acad. Press. 2009.
- Beniaguev, D.; Segev, I. and London, M. Single cortical neurons as deep artificial neural networks. Neuron 2021. 109(17), 2727-2739.e3. [CrossRef]
- Toxvaerd, S. Scilit. The emergence of the bilateral symmetry in animals: A review and a new hypothesis. Symmetry, 2021, 13(2), 261. [CrossRef]
- Matsuo, K.; Tamura, R.; Hotta, K.; Okada,M.; Takeuchi, A.; Yanlin Wu, Y. et al. Bilaterally Asymmetric Helical Myofibrils in Ascidian Tadpole Larvae. Front. Cell Dev. Biol. Sec. Morphogenesis and Patterning 2021, 9 – 2021. [CrossRef]
- Hoffmann, L.A, and Giomi, L. Theory of cellular homochirality and trait evolution in flocking systems. 2923, arXiv:2307.14360 [q-bio.CB]. [CrossRef]
- Zhu, DY., Cao, TT., Fan, HW. et al. The increased in vivo firing of pyramidal cells but not interneurons in the anterior cingulate cortex after neuropathic pain. Mol Brain. 2022, 15, 12. [CrossRef]
- Koga, K. Early evolution of membrane lipids: how did the lipid divide occur? Journal of Molecular Evolution, 2011, 72(3), 274-82. [CrossRef]
- Qi, H-X.; Jain, N.; Preuss, T.M. and Kaas, J.K. Inverted pyramidal neurons in chimpanzee sensorimotor cortex are revealed by immunostaining with monoclonal antibody SMI-32. Somatosensory & Motor Research 1999; 16(1), 49-56. [CrossRef]
- Steger, R.; Ramos, R.L.; Cao, R.; Yang, Q,; Chen, Ch-Ch.; Dominici, J. and Brumberg, J.C. Physiology and morphology of inverted pyramidal neurons in the rodent neocortex. Neuroscience. 2013, 17; 0: 165–179. [CrossRef]
- Martin, H.S.; Podolsky, K.A. and Devaraj, N.K. Probing the role of chirality in phospholipid membranes. ChemBioChem 2021, 22(22). 3148-315. [CrossRef]
- Kong, X-Z.; Postema, M.C.; Guadalupe, T.; de Kovel, C.; Boedhoe, P.C.W.; Hoogman, M.; Mathias, S.R.; van Rooij, D.; Schijven, D.; Glahn, D.C.; Medland, S.E. et al., Mapping brain asymmety in health and disease through the ENIGMA consortium. Hum Brain Mapp. 2022, 43(1), 167–181. [CrossRef]
- Kay, J.W. and Phillips, W.A. Contextual Modulation in Mammalian Neocortex is Asymmetric. Symmetry 2020, 12(5), 815. [CrossRef]
- Tripathi, S.; Dey,A.; Shanmugam, M.; Narayanan, R.S.and Chandrsekhar, V. Cobalt (II) Complexes as Single-Ion Magnets. Topics in Organometallic Chem 2019, 64. [CrossRef]
- Kagamiyama, H. and Hayashi, H. Crystallographic Structures. Branched-Chain Amino Acids, Part B. In the book Methods in Enzymoligy. Elsevier 2000.
- Housecroft, C. E.; Sharpe, A. G. (2004). Inorganic Chemistry (2nd ed.). Prentice Hall. ISBN 978-0-13-039913-7.
- Gradišar, H.; Božič, S.; Doles, T.; Vengust, D.; Hafner-Bratkovič, I.; Mertelj, A.; Webb, B.; Šali, A.; Klavžar, S. and Roman Jerala, R. Design of a single-chain polypeptide tetrahedron assembled from coiled-coil segments. Nat Chem Biol. 2013, 9(6), 362–366. [CrossRef]
- Le Bel, J.A. Bull. Soc. Chim. Fr., 1874, 22, 337.
- Mun, J.; Kim, M.; Yang, Y.; Badloe, T.; Ni, J.; Chen, Y.; Qiu C., and Rho, J, Electromagnetic chirality: From fundamentals to nontraditional chiroptical phenomena. Light. Sci. Appl. 2020, 9,139. [CrossRef]
- Wheeler RJ. Use of chiral cell shape to ensure highly directional swimming in trypanosomes. PLoS Comput Biol 2017, 13(1), e1005353. [CrossRef]
- Jacobus Henricus Van‘t Hoff, J.H. Arch. Neérl. Sci. Exactes. Nat., 1874, 9(1), 445-454, https://www.chemteam.info/Chem-History/Van’t-Hoff-1874.html.
- Xie, N.; Liu, S.; Yang, X.; Xiaoxiao He.; Huang, J. and Kemin Wang, K. DNA tetrahedron nanostructures for biological applications: biosensors and drug delivery. Analyst 2017, 18(142), 3322-3332. [CrossRef]
- Mathur, D.; Rogers, K.E.; Díaz, S.A.; Muroski, M.E.; Klein, W.P.; Nag, O.K.; Lee, K.; Field, L.D.; Delehanty, J.B. and Medin, I.L. Determining the Cytosolic Stability of Small DNA Nanostructures in Cellula. Nano Lett. 2022 22(12): 5037–5045. [CrossRef]
- Aumiller, Jr., W.A.; Masaki Uchida, and Trevor DouglasProtein cage assembly across multiple length scales. Chem Soc Rev. 2018 May 21; 47(10): 3433–3469. [CrossRef]
- Tenchov, R.; Bird, R.; Curtze, A.E. and Zhou, Q. Lipid Nanoparticles─From Liposomes to mRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 11, 16982–17015. [CrossRef]
- Hong, S.; Jiang, W.; Ding, Q.; Lin, K.; Zhao, C. and Wang, X. The Current Progress of Tetrahedral DNA Nanostructure for Antibacterial Application and Bone Tissue Regeneration. Int J Nanomedicine. 2023; 18: 3761–3780. [CrossRef]
- Zheng, Y.; Mao, K.; Chen,S. and Zhu, H. Chirality Effects in Peptide Assembly Structures. Front in Bioengineering and Biothechnology. 2021, 9, 703004. [CrossRef]
- Hassabis, D.; Kumaran, D.; Summerfield, C. and Botvinick, M. Neuroscience-Inspired Artificial Intelligence. Neuron 2017, 95(2), P245-258. [CrossRef]
- Zhao, J.; Wu, M.; Zhou, L.; Wang, X. and Jia, J. REVIEW. The Application of Artificial Intelligence in Brain-Computer Interface and Neural System Rehabilitation, Front. Neurosci., Sec. Neuroprosthetics 2022, 16. Research-Topic Cognitive psychology-based artificial intelligence review. [CrossRef]
- Surianarayanan, C.; Lawrence, J.J.; Chelliah, P.R.; Prakash, E. and Hewage, C. Gwanggil Jeon, Academic Editor. Convergence of Artificial Intelligence and Neuroscience towards the Diagnosis of Neurological Disorders—A Scoping Review. Sensors (Basel). 2023, 23(6): 3062. [CrossRef]
- Strychalski, W. 3D Computational Modeling of Bleb Initiation Dynamics. Front. Phys., Sec. Biophysics 2021, 9. [CrossRef]
- Johnson, E.; Cascio, D.; Sawaya, M.R.; Gingery, M. and Schröder, I. Crystal Structures of a Tetrahedral Open Pore Ferritin from the Hyperthermophilic Archaeon Archaeoglobus fulgidus. Structure. 2005 IF = 13(4), 637-648. [CrossRef]
- Zhang, K., Ezemaduka, A., Wang, Z. et al. A Novel Mechanism for Small Heat Shock Proteins to Function as Molecular Chaperones. Sci Rep 2015, 5, 8811 (2015). [CrossRef]
- Yoo, S.H. and Lee, H-S. Foldectures: 3D Molecular Architectures from Self-Assembly of Peptide Foldamers. Acc. Chem. Res. 2017, 50, 4, 832–841. [CrossRef]
- Sasaki, E., Böhringer, D., van de Waterbeemd, M. et al. Structure and assembly of scalable porous protein cages. Nat Commun 2017, 8, 14663. [CrossRef]
- Quaye, I.K. Extracellular hemoglobin: the case of a friend turned foe. Front Physiol. 2015; 6, 96. [CrossRef]
- Eaton, A.W. Retrospective on statistical mechanical models for hemoglobin allostery editors-pick The Journal of Chemical Physics 2022, 157, 184104. [CrossRef]
- Richter, F.; Meurers, B.H.; Zhu, C.; Medvedeva, V.P. and Chesselet, M-F. Neurons Express Hemoglobin α- and β-Chains in Rat and Human Brains. J Comp Neurol. 2009, 15(5): 538–547. [CrossRef]
- Zhang, D.; Ronson, T.K.; Mosquera, J.; Martinez, A.; Guy, L. and Nitschke, J.R. Anion Binding in Water Drives Structural Adaptation in an Azaphosphatrane-Functionalized FeII4L4 Tetrahedron. J. Am. Chem. Soc. 2017, 139, 19, 6574–6577. [CrossRef]
- 205. Shao X, Li Q, Mogilner A, Bershadsky AD, and Shivashankar GV. Mechanical stimulation induces formin-dependent assembly of a perinuclear actin rim. Proc. Natl. Acad. Sci. U.S.A. 2015; 112(20):E2595-601. [CrossRef]
- Kruppa, A.J.; Kishi-Itakura, C.; Masters, T.A.; Rorbach, J.E.; Grice, G.L.; Kendrick-Jones, J.; James A. Nathan, J.A. et al. Myosin VI-Dependent Actin Cages Encapsulate Parkin-Positive Damaged Mitochondria. Dev Cell. 2018. 26; 44(4): 484–499.e6. [CrossRef]
- Heid H, Rickelt S, Zimbelmann R, Winter S, Schumacher H, Dörflinger Y, Kuhn C, and Franke WW. On the formation of lipid droplets in human adipocytes: the organization of the perilipin-vimentin cortex. PLoS ONE 2014; 9(2), e90386. [PMID: 24587346]. [CrossRef]
- Tessarz, P., Schwarz, M., Mogk, A. & Bukau, B. The yeast AAA+ chaperone Hsp104 is part of a network that links the actin cytoskeleton with the inheritance of damaged proteins. Mol. Cell Biol. 2009, 29, 3738–3745. [CrossRef]
- Franke WW, Hergt M, and Grund C. Rearrangement of the vimentin cytoskeleton during adipose conversion: formation of an intermediate filament cage around lipid globules. Cell 1987; 49(1):131-41. [PMID: 3548999]. [CrossRef]
- Mostowy S, Bonazzi M, Hamon MA, Tham TN, Mallet A, Lelek M, Gouin E, Demangel C, Brosch R, Zimmer C, Sartori A, Kinoshita M, Lecuit M, and Cossart P. Entrapment of intracytosolic bacteria by septin cage-like structures. Cell Host Microbe 2010; 8(5):433-44. [CrossRef]
- Biliński, S.M. and Jaglarz, M.K. Organization and possible functions of microtubule cytoskeleton in hymenopteran nurse cells. Cell Motil Cytoskeleton. 1999, 43(3):213-20. [CrossRef]
- Xia, Z.; Wang, P.; Liu, X.; Liu, T.; Yan, Y.; Juan Yan, J. et al. Tumor-Penetrating Peptide-Modified DNA Tetrahedron for Targeting Drug Delivery. Biochemistry 2016, 55, 9, 1326–1331. [CrossRef]
- Chen, Gf., Xu, Th., Yan, Y. et al. Amyloid beta: structure, biology and structure-based therapeutic development. Acta Pharmacol Sin 2017, 38, 1205–1235. [CrossRef]
- Bao, Y.; Wu, S.; Chu, L.T.; Kwong, H.K.; Hartanto, H.; Huang, Y. et al. Early Committed Clockwise Cell Chirality Upregulates Adipogenic Differentiation of Mesenchymal Stem Cells. Adv Biosyst. 2020, 10, e2000161. [CrossRef]
- Yao, X.; Hu, Y.; Cao, B.; Peng, R. and Ding, J. Effects of surface molecular chirality on adhesion and differentiation of stem cells. Biomaterials 2013, 34 9001-9009. [CrossRef]
- Zheng, H.; Yoshitomi, T. and Yoshimoto, K. Analysis of Chirality Effects on Stem Cell Fate Using Three-dimensional Fibrous Peptide Hydrogels ACS Appl. Bio Mater. 2018, 1, 3, 538–543. [CrossRef]
- Walser, M.; Svensson, J.; Karlsson, Motalleb, R.; Åberg, M.; Kuhn, H.G.; Isgaard, J. and Åberg, N.D. Growth Hormone and Neuronal Hemoglobin in the Brain—Roles in Neuroprotection and Neurodegenerative Diseases. Front. Endocrinol. Sec. Neuroendocrine Science 2021, 11. [CrossRef]
- Zheng, R.; Yan, Y.; Pu, J. and Zhang, B. Physiological and Pathological Functions of Neuronal Hemoglobin: A Key Underappreciated Protein in Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23(16), 9088. [CrossRef]
- Flynn, K.S. The cytoskeleton and neurite initiation. Bioarchitecture. 2013, 3(4): 86–109. [CrossRef]
- Giuliana Indelicato, Newton Wahome, Philippe Ringler. Shirley A. Müller, Mu-Ping Nieh, Peter Burkhard, Reidun Twarock. Principles Governing the Self-Assembly of Coiled-Coil Protein Nanoparticles. Biophysical Journal 2016, 110(3), P646-660. [CrossRef]
- Heberle, F.A. and Feigenson, G.W. Phase Separation in Lipid Membranes. Cold Spring Harb Perspect Biol. 2011, 3(4), a004630. [CrossRef]
- Wu, X.; Cai, Q.;Feng, Z. and Mingjie Zhang, M. Review. Liquid-Liquid Phase Separation in Neuronal Development and Synaptic Signaling. Developmental Cell 2020, 55(1), 18-29. [CrossRef]
- Wonka, P. and Gervautz. M. Ray Tracing of Nonlinear Fractals. Journal of WSCG. 1998, 6(1-3), 424-431, http://wscg.zcu.cz/wscg1998/wscg98.htm.
- Taylor, L.LK; Riddell, I.A. and Smulders, M.M.J. Review. Self-Assembly of Functional Discrete Three-Dimensional Architectures in Water. Angewanter Chemie (Int Edition) 2019, 58(5), 1280-1307. [CrossRef]
- Chirikjian, G.S. Group theory and biomolecular conformation: I. Mathematical and computational models. J Phys Condens Matter. 2010, 22(32): 323103. [CrossRef]
- Dyakin V.V. and Posner, E.V. Molecular Chirality as an Integral Biomarker of Lactation, Nutrition and Cognitive Development Express-Review. Biomedical Science & Research. 2020, 8(5). [CrossRef]
- Liu, Y.; Wu, Z.; Armstrong, D.W.; Wolosker, H. and Zheng, Y. Detection and analysis of chiral molecules as disease biomarkers. Nat Rev Chem. 2023, 7(5): 355–373. [CrossRef]
- Ariga, K.; Richards, G.J.; Ishihara, S.; Izawa, H. and Jonathan P. Hill, J.P. Intelligent Chiral Sensing Based on Supramolecular and Interfacial Concepts Sensors (Basel). 2010; 10(7): 6796–6820. [CrossRef]
- Li, J., Li, J., Liu, R. et al. Autonomous discovery of optically active chiral inorganic perovskite nanocrystals through an intelligent cloud lab. Nat Commun 2020. 11, 2046. [CrossRef]
- Rainey, P.B. Major evolutionary transitions in individuality between humans and AI. Phil. Trans.of Royal Soc. B. 2023, 378 (1872). [CrossRef]
- Shinohara, Y.; Hirase, H.; Watanabe, M.; Itakura, M.; Takahashi, M. and Shigemoto, R. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. Proc Natl Acad Sci U S A. 2008. 105(49):19498-503. [CrossRef]
- Goto K, Kurashima R, Gokan H, Inoue N, Ito I, Watanabe S. Left−Right Asymmetry Defect in the Hippocampal Circuitry Impairs Spatial Learning and Working Memory in iv Mice. PLoS ONE 2010, 5(11): e15468. [CrossRef]
- El-Gaby, M.; Shipton, O.A. Paulsen, O. Synaptic Plasticity and Memory: New Insights from Hippocampal Left–Right Asymmetries, The Sage Journal. The Neuroscientist 2014, 21(5).
- Shimbo, A.; Kosaki, Y.; Ito, I. and Watanabe, S. Mice lacking hippocampal left-right asymmetry show non-spatial learning deficits. Behavioural Brain Research 2018, 336(15) 156-165. [CrossRef]
- Shipton, O. A., El-Gaby, M., Apergis-Schoute, J., Deisseroth, K., Bannerman, D. M., Paulsen, O., & Kohl, M. M. (2014). Left–right dissociation of hippocampal memory processes in mice. PNAS 2014, 111(42), 15238–15243. [CrossRef]
- Sha, Z., Schijven, D., Carrion-Castillo, A. et al. The genetic architecture of structural left–right asymmetry of the human brain. Nat Hum Behav 2021, 5, 1226–1239. [CrossRef]
- Jianguo Wang, Sidi Ma, Peijie Yu, Xionglei He Author NotesEvolution of Human Brain Left–Right Asymmetry: Old Genes with New Functions. Molecular Biology and Evolution, 2023, 40(9), msad181. [CrossRef]
- Cathryn A. Cutia, Leanna K. Leverton and Catherine A. Christian-HinmanSex and Estrous Cycle Stage Shape Left-Right Asymmetry in Chronic Hippocampal Seizures in Mice. eNeuro 29 May 2023, 10 (6). [CrossRef]
- Shiptona, O.A.; Tanga, C.S.; Paulsena, O. and Vargas-Caballero, M. Differential vulnerability of hippocampal CA3-CA1 synapses to Ab. Acta Neuropathologica Communications 2022, 10, 45. [CrossRef]
Figure 1.
(Graphical Abstract). Allegorical/metaphorical representation of four fundamental determinants (driving forces) of biological evolution and pyramidal neuron morphology: space, time, symmetry, and relativity. Space, time, symmetry, and relativity are considered as the elements of tetrahedral meta-structure.
Figure 1.
(Graphical Abstract). Allegorical/metaphorical representation of four fundamental determinants (driving forces) of biological evolution and pyramidal neuron morphology: space, time, symmetry, and relativity. Space, time, symmetry, and relativity are considered as the elements of tetrahedral meta-structure.

Figure 2.
Biological evolution from the bio-chirality perspective. A broad view of biological symmetry assumes that chirality transfer from the molecular to macromolecular and cellular levels plays a vital role in human physiology underlying perceptual and cognitive function linked with bilateral organisms’ emotional and behavioral expression. Biologists have made long-term efforts to explain the common origin of the homochiral world of DNA code and protein sequence resulting in the emergence of the human mind. Quite unexpectedly, for many scientists, it was realized that the Ariadne thread is the concept of symmetry [7,20]. On the other hand, all theories ignoring the symmetry principle occurred to be irrelevant. Indeed, the evolutionary history of life exhibits the chain of successive events (chain of chirality transfer) linked by the apparent common association with the notion of symmetry. The four most significant are indicated in Figure 2. Events I, II, and IV are the long-term concern of the scientists. The appearance and evolution of PyrN (event III) attracts increasing attention but does not focus on the specific geometric shape of soma. Top. Transfer of symmetry constraints from molecular to cellular level. a) Tetrahedron - geometrical shape which can be transformed from chiral to achiral version. b) Tetrahedral (sp3-hybridized) carbons with four different substituents are the principal components of biological molecules exhibiting homochirality. c) Tetrahedral structure of water forming about 70 % of cytosol. d) L/D amino acids. e) Tetrahedral structure of hemoglobin (Hb) expressed in neuronal and red blood cells. f) Tetrahedral assembly of DNA. g) Pyramidal neuron. Adopted with changes from [15,16]. PNs were discovered by the Ukrainian anatomist and histologist Vladimir A. Betz (1834–1894) [9]. Middle. Tree of life. Successive biologically essential evolutionary selections: I – prevalent molecular chirality, II - bilaterality, III - pyramidal neurons, IV - human mind. Adopted from Encyclopedia Britannica Inc. 2012 with alterations. The evolution of bilaterian CNS occurs through the differentiation of cell types and network organization. A line of phylogenomic evidence suggests that some organisms (such as sponges and placozoans) may benefit from the loss and/or modification of their neural cell types. But the mainstream of evolution demonstrates the benefit of the CNS for managing voluntary movement (events in the space-time domain). At the cellular level, the origin of CNS is characterized by the appearance of pyramidal neurons (PyrNs). However, currently, there is no systematic information concerning the differentiation of PyrNs between mammal groups (placentals, marsupials, and monotremes). Bottom. Time-Arrow of biological evolution. Learning the congruence of the molecular and cellular events helps to understand the link between voluntary movement, space perception CNS, and PyrNs morphology.
Figure 2.
Biological evolution from the bio-chirality perspective. A broad view of biological symmetry assumes that chirality transfer from the molecular to macromolecular and cellular levels plays a vital role in human physiology underlying perceptual and cognitive function linked with bilateral organisms’ emotional and behavioral expression. Biologists have made long-term efforts to explain the common origin of the homochiral world of DNA code and protein sequence resulting in the emergence of the human mind. Quite unexpectedly, for many scientists, it was realized that the Ariadne thread is the concept of symmetry [7,20]. On the other hand, all theories ignoring the symmetry principle occurred to be irrelevant. Indeed, the evolutionary history of life exhibits the chain of successive events (chain of chirality transfer) linked by the apparent common association with the notion of symmetry. The four most significant are indicated in Figure 2. Events I, II, and IV are the long-term concern of the scientists. The appearance and evolution of PyrN (event III) attracts increasing attention but does not focus on the specific geometric shape of soma. Top. Transfer of symmetry constraints from molecular to cellular level. a) Tetrahedron - geometrical shape which can be transformed from chiral to achiral version. b) Tetrahedral (sp3-hybridized) carbons with four different substituents are the principal components of biological molecules exhibiting homochirality. c) Tetrahedral structure of water forming about 70 % of cytosol. d) L/D amino acids. e) Tetrahedral structure of hemoglobin (Hb) expressed in neuronal and red blood cells. f) Tetrahedral assembly of DNA. g) Pyramidal neuron. Adopted with changes from [15,16]. PNs were discovered by the Ukrainian anatomist and histologist Vladimir A. Betz (1834–1894) [9]. Middle. Tree of life. Successive biologically essential evolutionary selections: I – prevalent molecular chirality, II - bilaterality, III - pyramidal neurons, IV - human mind. Adopted from Encyclopedia Britannica Inc. 2012 with alterations. The evolution of bilaterian CNS occurs through the differentiation of cell types and network organization. A line of phylogenomic evidence suggests that some organisms (such as sponges and placozoans) may benefit from the loss and/or modification of their neural cell types. But the mainstream of evolution demonstrates the benefit of the CNS for managing voluntary movement (events in the space-time domain). At the cellular level, the origin of CNS is characterized by the appearance of pyramidal neurons (PyrNs). However, currently, there is no systematic information concerning the differentiation of PyrNs between mammal groups (placentals, marsupials, and monotremes). Bottom. Time-Arrow of biological evolution. Learning the congruence of the molecular and cellular events helps to understand the link between voluntary movement, space perception CNS, and PyrNs morphology.
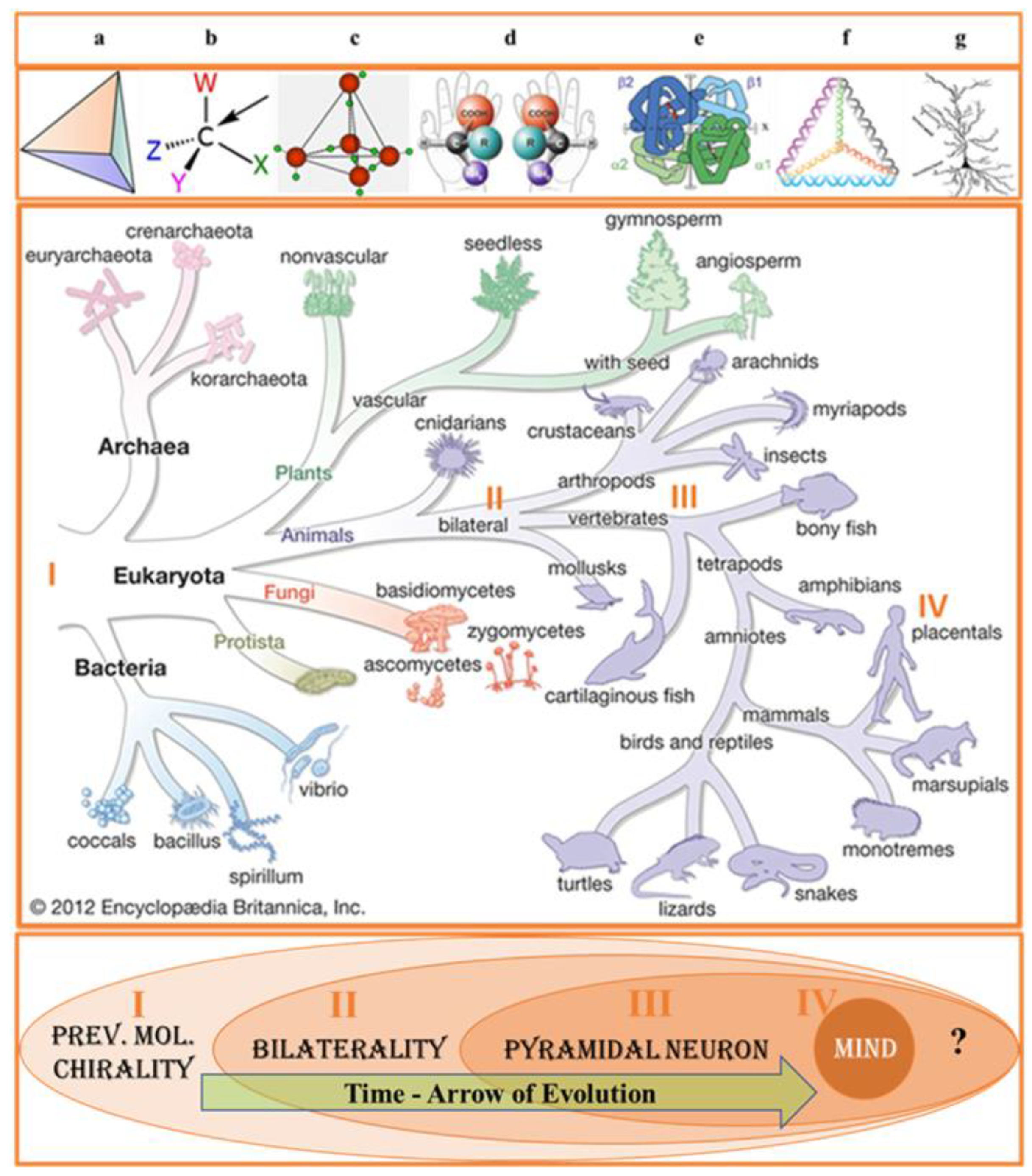
Figure 3.
(A, B, C) From molecular to cell chirality. A. Biological (specific) isomerism exhibits many forms of not equivalency in the functions of two molecular isomers. Biological symmetry preference among stereoisomers exists in two forms: the enantiomers for L-enantiomers (chiral) and cis diastereomers (achiral). While enantiomers can only come in pairs, many diastereomers exist for a given molecule. The most significant difference in biochemistry is attributed to enantiomers characterized by prevalent handedness in biological objects. B. Pyramidal neurons (arrow) were identified by their large size, triangular shape, and the presence of a pially oriented apical dendrite opposite one or more basilar dendrites. The scale bar represents 10 μm. Adopted from [110]. C. Arbitrary variants of PyrN orientation in the bilateral brain may provide different spatial navigation information processing strategies. Most sensorimotor cortical neurons in the chimpanzees had typical vertically oriented apical dendrites that extended towards the pial surface [174,175]. However, the question of hemispheric symmetry in PyrN orientation has yet to be analyzed. We are unfamiliar with studying the PyrN’s soma orientation in the horizontal plane. A few papers describe the orientation of basal dendritic arbors of pyramidal cells concerning the anterior, dorsal, posterior, and ventral directions [148].
Figure 3.
(A, B, C) From molecular to cell chirality. A. Biological (specific) isomerism exhibits many forms of not equivalency in the functions of two molecular isomers. Biological symmetry preference among stereoisomers exists in two forms: the enantiomers for L-enantiomers (chiral) and cis diastereomers (achiral). While enantiomers can only come in pairs, many diastereomers exist for a given molecule. The most significant difference in biochemistry is attributed to enantiomers characterized by prevalent handedness in biological objects. B. Pyramidal neurons (arrow) were identified by their large size, triangular shape, and the presence of a pially oriented apical dendrite opposite one or more basilar dendrites. The scale bar represents 10 μm. Adopted from [110]. C. Arbitrary variants of PyrN orientation in the bilateral brain may provide different spatial navigation information processing strategies. Most sensorimotor cortical neurons in the chimpanzees had typical vertically oriented apical dendrites that extended towards the pial surface [174,175]. However, the question of hemispheric symmetry in PyrN orientation has yet to be analyzed. We are unfamiliar with studying the PyrN’s soma orientation in the horizontal plane. A few papers describe the orientation of basal dendritic arbors of pyramidal cells concerning the anterior, dorsal, posterior, and ventral directions [148].
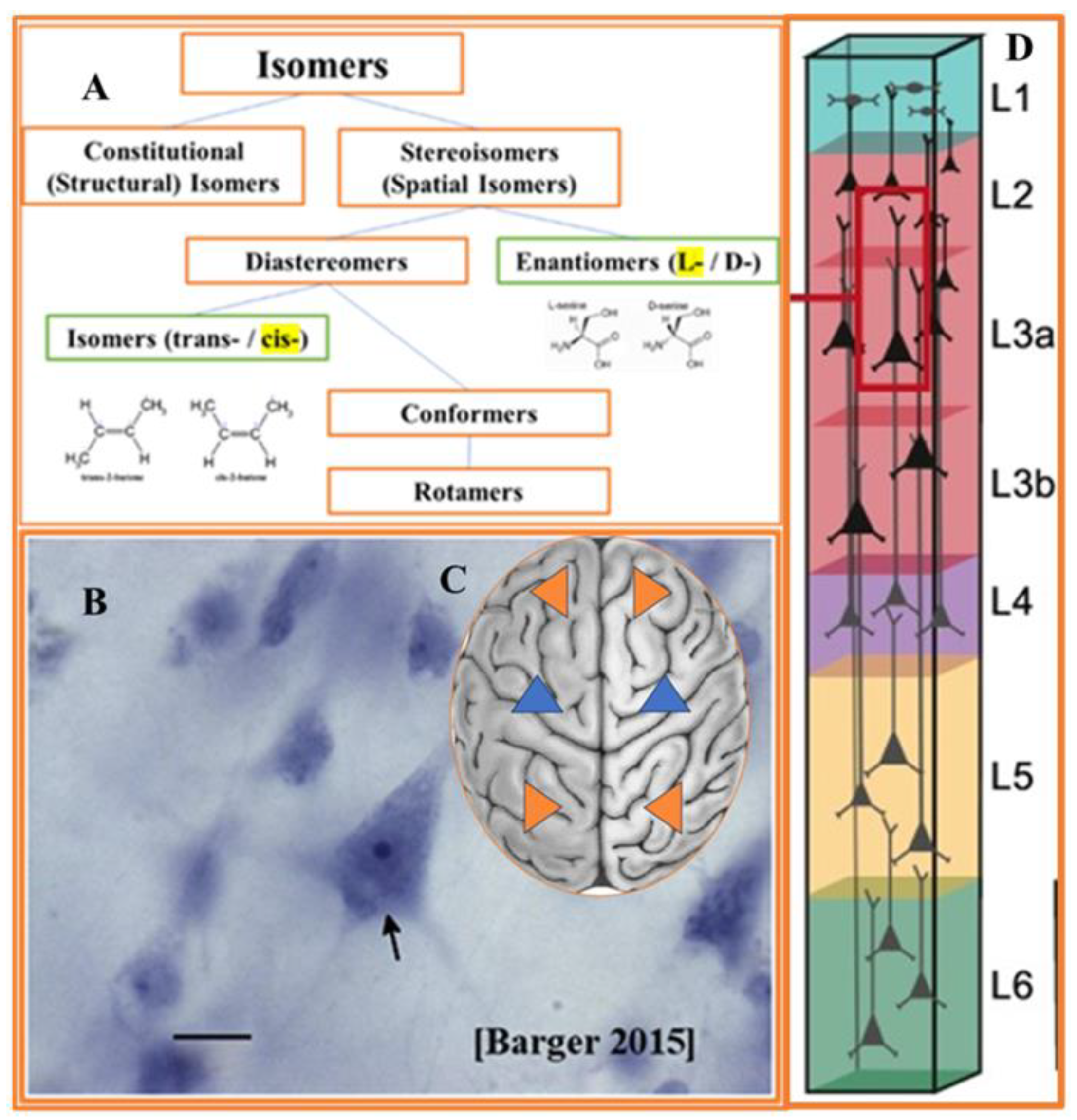
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



