Submitted:
13 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
Cinnamate 4-hydroxylase (C4H) is a cytochrome P450-dependent monooxygenase enzyme that catalyzes a key step in phenylpropanoid metabolism in plants. However, there is very limited information on the function of C4H in medicinal plants. In the present study, the C4H gene from the Ferula pseudalliacea genome (named FpC4H) was cloned and characterized. In addition, the expression levels of FpC4H in the roots, stems, leaves, flowers and seeds were investigated via real-time PCR. FpC4H showed the most similarity with C4H proteins from Daucus carota. The results predicted that FpC4H, as well as its orthologs, is an unstable and hydrophilic protein. A high genetic distance was observed between FpC4H and C4H from monocots based on evolutionary analysis. Moreover, a variable region related to the variable function of C4H was detected in conserved domain region of C4H proteins. Protein structure analysis revealed that isoleucine and leucine residues are frequently present in the binding region of FpC4H, and two ion binding regions, cysteine258 and asparagine217, were identified in the 3D structure of FpC4H. Expression analysis revealed that the FpC4H gene is expressed in all organs of F. pseudallicea. A low expression level of FpC4H was recorded in leaves, and a high expression level was observed in flower tissues. Based on the expression profile, FpC4H seems to be linked to a high content of phenolic compounds in flowers.
Keywords:
Phenylpropanoid pathway
; Cinnamate-4-hydroxylase
; Apiaceae
; Secondary metabolism
1. Introduction
Medicinal plants have been used for thousands of years as traditional remedies worldwide. The bioactive compounds of these plants have proven beneficial for treating human diseases and promoting health. Endemic medicinal plants found only in certain geographic regions often have unique properties that have evolved in their isolated habitats. Iran, located at the intersection of Central Asia, West Asia, and the Middle East, harbors more than 8,000 plant species, approximately 1,900 of which are endemic [1]. Many of these endemic species have a long history of use in traditional Iranian medicine for treating conditions such as infections, inflammation, pain, and digestive issues [2]. As rich sources of specialized metabolites and bioactive compounds, Iran's endemic medicinal flora represent an immense repository of plant chemical diversity waiting to be explored for new drug leads and treatment possibilities. Understanding these endemic plants through genomic, phytochemical and pharmacological studies can provide insights for developing improved therapies and advancing human health [3].
Apiaceae, also known as Umbelliferae, is an economically important family of flowering plants consisting of more than 3,700 species, including crops, herbs, spices, and medicinal plants. Major Apiaceae crops include carrots, celery, parsley, parsnip, and fennel, which are cultivated worldwide for their edible taproots, stems, leaves, or seeds that add flavor, nutrients and medicinal value to human diets [4]. Many Apiaceae species contain bioactive compounds and essential oils that give them antimicrobial, insecticidal, and pharmaceutical properties. Popular medicinal Apiaceae plants include anise, caraway, cumin, coriander, dill, and asafetida and are used extensively in traditional medicine systems and herbal remedies globally. The Apiaceae family has a worldwide distribution but is most abundant in temperate climates. Several genera, such as Ferula, Prangos and Kelussia, predominantly occur around the Mediterranean and across West and Central Asia [5,6]. Understanding the phytochemical diversity of Apiaceae species through genomic and metabolomic studies can aid in selective breeding efforts and expand the utilization of medicinal members within this vital plant family for improved food security and enhanced medication use [6,7].
The genus Ferula belongs to the economically significant Apiaceae family and comprises more than 170 species of flowering plants. Most members of this genus are perennial herbaceous plants that grow in arid grasslands and mountains ranging from Central Asia to northern Africa [8]. The characteristic features of Ferula species include thick roots, sturdy hollow stems, and feathery leaves, and they produce resins and gums that contain bioactive compounds such as terpenoids, coumarins, and alkaloids [9]. These specialized metabolites confer antispasmodic, antimicrobial, and anti-inflammatory effects to Ferula plants, underlying their extensive use in traditional medicine systems in Iran and neighboring countries [9,10]. Ferula pseudalliacea is an indigenous medicinal plant species endemic to the Zagros mountainous regions of Iran's Kurdistan Province and western border areas. This species has been traditionally utilized in folk medicine for various purposes, including as an antiparasitic and antiulcer remedy [11]. Several new bioactive sesquiterpene coumarins, including kamonolol acetate, farnesiferol B, sanandajin, ethyl galbanate, methyl galbanate, and fekrynol acetate, have been isolated and identified from F. pseudalliacea [12]. These compounds have exhibited potent anticancer, antiplasmodial, antibacterial and strong antioxidant effects, revealing the medicinal potential of these endemic species [13,14]. However, despite harboring such valuable specialized metabolites, F. pseudalliacea has a narrow geographical range with scattered fragmented populations, leading to its classification as a threatened plant [15]. With rich phytochemical diversity and proven pharmacological properties, further conservation efforts and molecular research on F. pseudalliacea can facilitate sustainable utilization of this medicinal species as well as the discovery of novel bioactive lead compounds.
Cinnamate 4-hydroxylase (C4H) is a cytochrome P450-dependent monooxygenase that catalyzes a key step in phenylpropanoid metabolism across plant species. It controls the conversion of trans-cinnamic acid to p-coumaric acid, funneling metabolites toward myriad phenolic end products such as lignins, coumarins, flavonoids, anthocyanins, and stilbenoids [16,17]. These phenolic compounds act as structural barriers or chemical defenses in plants while also exerting beneficial antioxidant, anti-inflammatory, and antimicrobial effects. C4H belongs to the CYP73A enzyme family and is encoded by a multigene family whose members are induced differentially by biotic and abiotic stresses [18]. Complete genomic sequences and individual C4H gene functions have already been well characterized in model crop species using molecular genetics tools [19]. Structural modeling revealed conserved heme-binding and substrate recognition sites across C4H orthologs from diverse taxa [20]. However, C4H has been cloned and sequenced in several medicinal plant species, such as Salvia miltiorrhiza [21], Scrophularia striata [22], Euphorbia maculata [23], and Carthamus tinctorius [24]; its sequence has not been explored among key medicinal species, such as Ferula, which are rich reservoirs of phenylpropanoid-derived chemicals. Elucidating C4H genes in such plants can reveal their evolutionary relationships and regulatory patterns governing species-specific phenolic secondary metabolism. Overall, advances in C4H research have allowed improved metabolic engineering strategies for enhancing the biosynthesis of high-value phenolics in medicinal and crop plants. Currently, little information exists regarding the sequence and expression patterns of C4H in industrially and conservationally important Ferula species, particularly F. pseudalliacea. Hence, the current study aimed to i) isolate and sequence the C4H gene from F. pseudalliacea using 3’-RACE methods; ii) conduct bioinformatic analysis of the obtained C4H sequences, including conserved domains, evolutionary relationships, and predicted structural features; and iii) quantify the expression profiles of individual C4H genes across roots, stems, leaves, flowers and seeds of F. pseudalliacea plants via real-time PCR.
2. Materials and Methods
2.1. Plant material
Multiple tissues, including roots, stems, leaves, flowers and seeds, were harvested from healthy, uniform Ferula pseudalliacea individuals (Figure 1) growing in their native habitat site located in Gazne village (10 km west of Sanandaj, coordinates 35°08'38.1"N 46°57'45.4"E), Iran. The collected samples were promptly frozen in liquid nitrogen upon site extraction and transported to the biotechnology laboratory at the University of Kurdistan (UOK) under cold chain maintenance at -80°C for subsequent experiments. Botanical authentication of the F. pseudalliacea specimens utilized in the current investigation was performed by Eng. Hossein Maroofi (Agriculture and Natural Resources Research Center). Key morphological details have been deposited in the herbarium collection housed in the Faculty of Agriculture, University of Kurdistan, Iran, for future verification.
2.2. DNA and RNA extraction and cDNA synthesis
Genomic DNA was extracted from the leaf samples using the cetyltrimethylammonium bromide (CTAB) protocol [25], with certain modifications. Purified DNA was subjected to digestion with Ribonuclease A (34390, SERVA, Heidelberg, Germany) at 37°C for 1 hour to eliminate contaminating RNA. Total RNA was isolated from the assorted frozen Ferula pseudalliacea tissues, including roots, stems, leaves, flowers and seeds, utilizing the methodology delineated by Mazzara and James [26]. The RNA samples were further treated with DNase I (Pishgam Biotech, Tehran, Iran) to eliminate DNA contamination. Quantification and integrity analyses of the extracted DNA and RNA samples were carried out with a NanoDrop OneC device (Thermo Scientific TM) coupled with agarose gel electrophoresis visualization. Subsequently, first-strand complementary DNA (cDNA) was synthesized from 1 μg of pure RNA template from each sample using a commercially available reverse transcriptase kit (Yekta Tajhiz Azma Co., Tehran, Iran) per the manufacturer's protocols.
2.3. Primer design and gene cloning and sequencing
Due to the lack of genomic resources for F. pseudalliacea, primer design targeted highly conserved regions within cinnamate 4-hydroxylase (C4H) coding sequences and actin reference gene sequences from other Apiaceae taxa, including Angelica gigas, Angelica dahurica, Daucus carota, and Petroselinum crispum, with available C4H protein data. Multiple sequence alignments were constructed using Clustal Omega [27] to identify consensus protein sequences, which were further utilized to develop degenerate primers via Primer3 [28]. In silico analyses enabled screening for self-dimers, secondary structures and mispriming sites using the Oligo Analyzer v3.1 online platform (http://eu.idtdna.com). Initial PCR was used to amplify a core FpC4H fragment, which was subsequently amplified via 3’-RACE to obtain full-length transcripts via the use of an oligo-dT anchor primer paired with a specific forward primer (Table 1). Amplicons representing putative FpC4H genes were gel-purified using the N0041101 Gene MARK Kit (BioElegen Tech, Taichung, Taiwan) and TA-cloned and inserted into the pTG19-T vector. The constructs were subsequently transformed into E. coli DH5ɑ cells via heat shock and subsequently plated for blue–white screening and colony PCR validation using M13 universal and gene-specific primers [29]. Bidirectional Sanger sequencing of 10 positive clone plasmids per transformation was performed (Applied Biosystems, Foster City, CA, USA), yielding common FpC4H sequences that were edited with Geneious software [30] and deposited into the NCBI under accession number MH987776.1. A BLASTx homology search was used to confirm sequence identity against nonredundant databases.
2.4. Phylogeny analysis
To investigate the evolutionary relationships of the FpC4H proteins, multiple sequence alignments of the deduced FpC4H proteins and known orthologs from Arabidopsis thaliana, Oryza sativa, Brachypodium distachyon, Daucus carota, Brassica napus, Helianthus annuus and Camelina sativa were constructed using Clustal Omega. The alignment was analyzed using the IQ-TREE web server [31] for maximum likelihood phylogenetic reconstruction with 1000 bootstrap replicates. The resulting phylogenetic tree was visualized using iTOL online software [32]. Conserved protein domain prediction of FpC4H sequences was also carried out using the InterProScan tool [33].
2.5. Sequence structure analysis
The prediction of putative phosphorylation sites in FpC4H relied on the NetPhos 3.1 server [34], for which a threshold score ≥ 0.90 was used. A protein interaction network analysis was performed for FpC4H based on the STRING database [35] and carrot ortholog information. Significant biological processes and molecular function terms linked to the network nodes were identified using an FDR ≤ 0.05. Additionally, various physicochemical properties of the FpC4H protein and their orthologous sequences were computed using the ProtParam online tool [36]. In addition, the 3D structure and binding sites of the FpC4H protein were investigated using the online Phyre2 tool [37].
2.6. Real-time PCR analysis
Quantitative real-time PCR (qPCR) analysis was performed to examine and compare the expression profiles of FpC4H across different F. pseudalliacea organs, including seeds, flowers, stems, leaves, and roots. Gene-specific primers targeting 3’-UTR regions were utilized for amplification from synthesized cDNA templates. The FpACTIN housekeeping gene was coamplified in parallel reactions as an internal normalization control. qPCR was performed in three biological replicates with two technical replicates per sample in 15 μL volumes by applying SYBR Green chemistry (Yekta Tajhiz Azma, Tehran, Iran) on an ABI StepOne Plus thermal cycler (Life Technologies, CA, USA). The cycling conditions were set to initial denaturation at 95°C for 3 min, followed by 40 cycles of 95°C for 20 sec and 60°C for 30 min. Melt curve analysis validated the single peak and specific amplification. Relative quantification relied on the ΔΔCt method [38] using FpACTIN for normalization, with the lowest expressing tissue set used as a calibrator. Statistical analyses included one-way ANOVA accompanied by Tukey's post hoc test performed with SPSS software (ver. 27.0.1.0).
3. Results
3.1. Screening and confirmation of clones
In this study, one isoform of the C4H gene in the F. pseudaliacea genome (named FpC4H) was identified and registered in the NCBI database with accession number MH987776. The sequence of FpC4H was most similar to that of C4H proteins from Daucus carota. In the present study, the physicochemical properties of FpC4H and its orthologs were compared (Table 2). The results revealed that all the studied C4H proteins are unstable and have a negative GRAVY, indicating that the C4Hs are more hydrophilic proteins. According to the isoelectric point (pI) value, the FpC4H protein was predicted to be active in near-neutral environments, while other orthologs were more active under alkaline conditions.
3.2. Phylogenetic analysis
Evolutionary analysis of C4H genes revealed that this gene has undergone a different evolutionary process in monocotyledonous and dicotyledonous plants (Figure 2); thus, FpC4H orthologs in monocotyledonous plants showed greater genetic distance than orthologs in dicotyledonous plants. Based on the genetic distance, the C4H genes were divided into four groups. FpC4H and its orthologs from carrot and sugar beet were placed in group 3, which also showed less genetic distance from the sunflower C4H genes in group 2. C4Hs from monocotyledonous plants were placed in group 1, and Arabidopsis C4H, along with its orthologs in rapeseed and camelina, were placed in group four. A separate multiple sequence alignment analysis was performed for the members of groups 2 and 3 along with Arabidopsis C4H to better understand the sequence structure of C4Hs. The results revealed that there is a conserved region associated with the corresponding domain (cytochrome P450) in these proteins (Figure 3). Interestingly, a region of 11 amino acids was identified in the sequences, which was presented as a variable region (VR) that might be related to the variable function of C4Hs in different plants, although further research is needed.
3.3. Three-dimensional structure and binding sites of FpC4H
The three-dimensional structure of FpC4H was predicted based on a homolog model with high identity and 100% confidence in the model (Figure 4). In addition, the binding regions that affect the activity and possible interactions were identified (Figure 4a). The results revealed that isoleucine (I) and leucine (L) residues were more abundant in the binding region of the FpC4H protein. In addition, two regions, C258 (cysteine 258) and N217 (asparagine 217), were predicted to be ion binding regions (Figure 4b). In addition, the mutation sensitivity of the FpC4H structure was analyzed, the results of which showed that the FpC4H structure has low sensitivity to mutations (Figure 4b).
3.4. FpC4H protein phosphorylation events
Posttranslational modifications affect the activity and stability of target proteins. In the present study, the possible phosphorylation sites of FpC4H within its close orthologs were identified (Figure 5). Four serine residues were identified as phosphorylation sites in FpC4H as well as in its close ortholog in carrot (DCAR_018641). The phosphorylation sites ranged between two sites (HanXRQr2_Chr06g0265981 and Os05g0320700) and nine sites in Arabidopsis C4H (Figure 5a). The distribution pattern of the phosphorylation regions of the C4H proteins was somewhat variable, although some regions were almost common to most proteins, such as serine 266, 197, and 477 (Figure 5b). These results suggest that some posttranslational events in these proteins have remained conserved during the evolutionary process.
3.5. Interaction network of FpC4H
An interaction network for FpC4H was constructed based on available data from carrot plants (Figure 6). The FpC4H homolog, a key point in the network, strongly interacts with proteins involved in diverse pathways. C4H-1 interacted with other proteins from the phenylpropanoid pathway, including C4H-2, PAL1, PAL3, and PAL4 (Figure 6a). In addition, several interactions were observed between CHS (chalcone synthase) and 4CL3 (4-coumarate:coenzyme A ligase). In addition, Gene Ontology (GO) enrichment analysis was performed to determine the molecular functions and biological processes associated with the C4H network elements (Figure 6b and 6c). The molecular function terms chalcone synthase activity, oxidoreductase activity, cinnamoyl-CoA reductase activity, acyltransferase activity, 4-coumarate-CoA ligase activity and phenylalanine ammonia-lyase activity were strongly linked with the C4H network (Figure 6b). In addition, several biological process terms, such as secondary metabolic process, phenylpropanoid metabolic/biosynthetic process, polyketide biosynthetic process, flavonoid biosynthetic process, aromatic amino acid family catabolic process, and cinnamic acid biosynthetic process, were significantly enriched (Figure 6c). These findings revealed that C4H is involved in several key metabolite pathways.
3.6. Analysis of FpC4H gene expression
In the present study, the expression levels of the FpC4H gene were evaluated in different organs of F. pseudallicea (Figure 7). Compared with those in other tissues, FpC4H in leaves was expressed at lower levels. The FpC4H gene was more highly expressed in flower tissues, and the FpC4H transcript levels were equal in stem and root tissues. In addition, the FpC4H gene was expressed at lower levels in seed tissues than in flowers, stems, and roots. The variable expression of the FpC4H gene indicated that this gene is more strongly induced in reproductive organs and is involved in the increase in linked metabolites in flower tissues.
4. Discussion
C4H is involved in the biosynthesis of phenylpropanoids, which are important active plant compounds that participate in the production of valuable secondary metabolites. There is not enough information on the C4H gene as a key gene in the biosynthesis of several secondary metabolites in the Ferula genus. In this study, the isolation and identification of this gene and its expression level in different organs of Ferula pseudalliacea plants were carried out. The results revealed that F. pseudaliacea has one isoform of the C4H gene (FpC4H). Only one member of this gene has been identified in Arabidopsis [39], parsley [39], cowpea [40] and black raspberry [41]. However, in sycamore [42], subabel tree [43] and rapeseed [44], C4H is encoded by a small gene family. For instance, four C4H genes were found in the soybean genome [20] and Populus kitakamiensis genome [45]. The number of copies of a gene can be related to polyploidy events and segmental and tandem duplications [46,47]. Increasing the number of isoforms of a gene can be effective at increasing functional diversity and related interactions [48]. Phylogenetic analysis of C4H proteins revealed that there is a large genetic distance between the C4H genes of monocot and dicot plants, suggesting that they have undergone a different evolutionary process. Most likely, after the derivation of dicotyledons and monocotyledons, structural diversity occurred in the C4H genes. Despite this genetic difference, C4Hs have similar physicochemical characteristics, revealing that the conditions and environment of activity are the same. A region (consisting of eleven amino acids) was identified in the conserved region of C4H proteins, which showed high variation between orthologs. This region could be involved in the functional diversity of orthologs, although further molecular studies on this region are needed.
An investigation of the FpC4H protein structure revealed that there is an extensive binding region in this protein. The presence of many binding regions in a protein can affect its extent of activity and interaction. Additionally, the amino acids isoleucine and leucine likely play more prominent roles in the binding and interaction of FpC4H. In addition, two ion binding regions, cysteine 258 and asparagine 217, were recognized in the FpC4H structure. The common feature of CYP450 family members is the presence of an iron core in their protein structure, which is connected to a cysteine amino acid [49]. Additionally, four serine residues were identified as potential phosphorylation sites in the FpC4H protein. Phosphorylation is a posttranslational modification that affects the activity and life of a protein [50]. For proteins such as C4H, which is an upstream element in metabolite synthesis pathways, phosphorylation plays a critical role in controlling downstream components by influencing these proteins. The results of the interaction network confirmed that FpC4H interacts with proteins involved in the biosynthesis of secondary metabolites. For example, biological process terms, including phenylpropanoid metabolic/biosynthetic process, polyketide biosynthetic process, flavonoid biosynthetic process, aromatic amino acid family catabolic process, and cinnamic acid biosynthetic process, were significantly enriched in FpC4H (Figure 6c). Phenylpropanoids and flavonoids play key roles in the response to adverse conditions, including abiotic and biotic stresses [51], as well as in plant growth processes [52]. Previous studies have shown that the expression of the C4H gene in the leaves of Lepidium apetalum after methyl jasmonate treatment is correlated with the flavonoid biosynthesis pathway [53]. Additionally, the flavonoid content at different stages in the tea plant Camellia sinensis was correlated with the expression pattern of C4Hb [54]. In addition, downregulation of the C4H gene in Nicotiana tabacum via the use of CRISPRi could increase flavonoid biosynthesis [55]. According to these results, it is hypothesized that the C4H gene is an important element in the biosynthesis of secondary metabolites and that this gene affects the improvement of plant resistance to adverse environmental conditions.
The FpC4H gene exhibited diverse expression levels in different organs, including the roots, stems, leaves, flowers, and seeds of F. pseudallicea, and the expression of this gene was detected in all organs (Figure 7). The results revealed that FpC4H is expressed in a tissue-specific manner. For example, the FpC4H gene was more strongly induced in floral tissues than in other tissues. In contrast, the C4H-2 gene was more highly expressed in rapeseed seeds than in roots and flowers [44]. Moreover, diverse expression levels were observed between four C4H genes in the soybean genome [20]. Metabolomics analysis of different organs of F. pseudallicea revealed that phenolic compounds accumulated more in flowers than in other organs [11]. The high expression level of the FpC4H gene in reproductive organs such as flowers suggested that this gene is positively correlated with the production of phenolic compounds, which can be considered in metabolite engineering studies.
5. Conclusion
In the present study, cinnamate 4-hydroxylase (FpC4H) was cloned and characterized in Ferula pseudalliacea. The physicochemical properties revealed that FpC4H is an unstable and hydrophilic protein. Phylogenetic analysis revealed that FpC4H and C4H proteins from Daucus carota underwent almost the same evolutionary process. In addition, the isoleucine and leucine residues were identified as key elements affecting the interaction and function of FpC4H. Expression analysis revealed that the FpC4H gene can be induced in all organs of F. pseudallicea, and a high expression level was observed in the flower tissues. Overall, these findings suggested that FpC4H is associated with the concentration of phenolic compounds in flowers.
Author Contributions
Conceptualization, P.S., B.B., Y.V. and D.D.; methodology, P.S., B.B. and Y.V.; validation, B.B., Y.V. and D.D.; formal analysis, P.S., B.B., Y.V. and P.H.; investigation, P.S., B.B. and Y.V.; writing—original draft preparation, Y.V. and P.H.; writing—review and editing, Y.V. and P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data generated or analyzed in this study are included in this article. Other materials that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank the University of Kurdistan for financially supporting and providing the needed facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Buso, P.; Manfredini, S.; Reza Ahmadi-Ashtiani, H.; Sciabica, S.; Buzzi, R.; Vertuani, S.; Baldisserotto, A. Iranian medicinal plants: From ethnomedicine to actual studies. Medicina 2020, 56, 97. [Google Scholar] [CrossRef]
- Amiri, M.S.; Yazdi, M.E.T.; Rahnama, M. Medicinal plants and phytotherapy in Iran: Glorious history, current status and future prospects. Plant Sci. Today 2021, 8, 95–111. [Google Scholar] [CrossRef]
- Hassanpouraghdam, M.B.; Ghorbani, H.; Esmaeilpour, M.; Alford, M.H.; Strzemski, M.; Dresler, S. Diversity and distribution patterns of endemic medicinal and aromatic plants of Iran: Implications for conservation and habitat management. Int. J. Environ. Res. Public Health 2022, 19, 1552. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.S.; Joharchi, M.R. Ethnobotanical knowledge of Apiaceae family in Iran: A review. Avicenna J. Phytomedicine 2016, 6, 621. [Google Scholar]
- Sayed-Ahmad, B.; Talou, T.; Saad, Z.; Hijazi, A.; Merah, O. The Apiaceae: Ethnomedicinal family as source for industrial uses. Ind. Crops Prod. 2017, 109, 661–671. [Google Scholar] [CrossRef]
- Wang, X.-J.; Luo, Q.; Li, T.; Meng, P.-H.; Pu, Y.-T.; Liu, J.-X.; Zhang, J.; Liu, H.; Tan, G.-F.; Xiong, A.-S. Origin, evolution, breeding, and omics of Apiaceae: A family of vegetables and medicinal plants. Hortic. Res. 2022, 9, uhac076. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, F.; Vannozzi, A.; Barcaccia, G. Impact of genomic and transcriptomic resources on apiaceae crop breeding strategies. Int. J. Mol. Sci. 2021, 22, 9713. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.J.; Chou, S.-C. The structural diversity of phthalides from the Apiaceae. J. Nat. Prod. 2007, 70, 891–900. [Google Scholar] [CrossRef]
- Salehi, M.; Naghavi, M.R.; Bahmankar, M. A review of Ferula species: Biochemical characteristics, pharmaceutical and industrial applications, and suggestions for biotechnologists. Ind. Crops Prod. 2019, 139, 111511. [Google Scholar] [CrossRef]
- Mohammadhosseini, M.; Venditti, A.; Sarker, S.D.; Nahar, L.; Akbarzadeh, A. The genus Ferula: Ethnobotany, phytochemistry and bioactivities—A review. Ind. Crops Prod. 2019, 129, 350–394. [Google Scholar] [CrossRef]
- Dastan, D.; Hamah-Ameen, B.A.; Salehi, P.; Ghaderi, H.; Miran, M. Chemical composition and bioactivities of essential oils from different plant parts of Ferula pseudalliacea Rech. f. as an endemic plant from Iran. Nat. Prod. Res. 2022, 36, 1311–1316. [Google Scholar] [CrossRef]
- Dastan, D.; Salehi, P.; Gohari, A.R.; Zimmermann, S.; Kaiser, M.; Hamburger, M.; Khavasi, H.R.; Ebrahimi, S.N. Disesquiterpene and sesquiterpene coumarins from Ferula pseudalliacea, and determination of their absolute configurations. Phytochemistry 2012, 78, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Dastan, D.; Salehi, P.; Gohari, A.R.; Ebrahimi, S.N.; Aliahmadi, A.; Hamburger, M. Bioactive sesquiterpene coumarins from Ferula pseudalliacea. Planta Med. 2014, 80, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Mahaki, H.; Tanzadehpanah, H.; Abou-Zied, O.K.; Moghadam, N.H.; Bahmani, A.; Salehzadeh, S.; Dastan, D.; Saidijam, M. Cytotoxicity and antioxidant activity of Kamolonol acetate from Ferula pseudalliacea, and studying its interactions with calf thymus DNA (ct-DNA) and human serum albumin (HSA) by spectroscopic and molecular docking techniques. Process Biochem. 2019, 79, 203–213. [Google Scholar] [CrossRef]
- JafariA, S.H.; SepehryB, A.; SoltanlooC, H.; KarimianD, A.A. Effect of topography and soil properties on distribution of Ferula pseudalliacea (Bitter Asafetida) in Yazd Province, Iran. J. Rangel. Sci. 2019, 9, 184. [Google Scholar]
- Ehlting, J.; Hamberger, B.; Million-Rousseau, R.; Werck-Reichhart, D. Cytochromes P450 in phenolic metabolism. Phytochem. Rev. 2006, 5, 239–270. [Google Scholar] [CrossRef]
- Shuab, R.; Lone, R.; Koul, K.K. Cinnamate and cinnamate derivatives in plants. Acta Physiol. Plant. 2016, 38, 1–9. [Google Scholar] [CrossRef]
- Kumar, K.; Debnath, P.; Singh, S.; Kumar, N. An overview of plant phenolics and their involvement in abiotic stress tolerance. Stresses 2023, 3, 570–585. [Google Scholar] [CrossRef]
- Hansen, C.C.; Nelson, D.R.; Møller, B.L.; Werck-Reichhart, D. Plant cytochrome P450 plasticity and evolution. Mol. Plant 2021, 14, 1244–1265. [Google Scholar] [CrossRef]
- Khatri, P.; Chen, L.; Rajcan, I.; Dhaubhadel, S. Functional characterization of Cinnamate 4-hydroxylase gene family in soybean (Glycine max). PLoS ONE 2023, 18, e0285698. [Google Scholar] [CrossRef]
- Huang, B.; Duan, Y.; Yi, B.; Sun, L.; Lu, B.; Yu, X.; Sun, H.; Zhang, H.; Chen, W. Characterization and expression profiling of cinnamate 4-hydroxylase gene from Salvia miltiorrhiza in rosmarinic acid biosynthesis pathway. Russ. J. Plant Physiol. 2008, 55, 390–399. [Google Scholar] [CrossRef]
- Rostami, Z.; Fazeli, A.; Hojati, Z. The isolation and expression analysis of cinnamate 4-hydroxylase and chalcone synthase genes of Scrophularia striata under different abiotic elicitors. Sci. Rep. 2022, 12, 8128. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.B.; Huang, S.H.; Zou, J.X.; Liu, H.S.; Quan, W.J.; Yang, H. Cloning and analysis of cinnamate acid 4-hydroxylase gene and its promoter from Euphorbia maculata L. Mol Plant Breed 2023, 21, 819–825. [Google Scholar]
- Hou, Y.; Wang, Y.; Liu, X.; Ahmad, N.; Wang, N.; Jin, L.; Yao, N.; Liu, X. A Cinnamate 4-HYDROXYLASE1 from Safflower Promotes Flavonoids Accumulation and Stimulates Antioxidant Defense System in Arabidopsis. Int. J. Mol. Sci. 2023, 24, 5393. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual; Springer: Dordrecht, The Netherlands, 1989; pp. 73–83. [Google Scholar]
- Mazzara, M.; James, D.J. The influence of photoperiodic growth condition on isolation of RNA from strawberry (Fragaria× ananassa Duch.) tissue. Mol. Biotechnol. 2000, 15, 237–241. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal omega. Curr. Protoc. Bioinforma. 2014, 48, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115–e115. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Mizutani, M.; Ohta, D.; Sato, R. Isolation of a cDNA and a genomic clone encoding cinnamate 4-hydroxylase from Arabidopsis and its expression manner in planta. Plant Physiol. 1997, 113, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Park, N.I.; Li, X.; Kim, Y.K.; Lee, S.Y.; Park, S.U. Molecular cloning and characterization of phenylalanine ammonia-lyase, cinnamate 4-hydroxylase and genes involved in flavone biosynthesis in Scutellaria baicalensis. Bioresour. Technol. 2010, 101, 9715–9722. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.-H.; Chung, B.-Y.; Kim, J.-H.; Kim, J.-S.; Lee, S.-S.; An, B.-C.; Lee, I.-J.; Kim, T.-H. cDNA cloning and expression pattern of cinnamate-4-hydroxylase in the Korean black raspberry. BMB Rep. 2008, 41, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, L.; Jiang, L.; Zhang, G.; Luo, Y. Molecular cloning and functional characterization of a cinnamate 4-hydroxylase-encoding gene from Camptotheca acuminata. Acta Physiol. Plant. 2016, 38, 1–9. [Google Scholar] [CrossRef]
- Kumar, S.; Omer, S.; Patel, K.; Khan, B.M. Cinnamate 4-Hydroxylase (C4H) genes from Leucaena leucocephala: A pulp yielding leguminous tree. Mol. Biol. Rep. 2013, 40, 1265–1274. [Google Scholar] [CrossRef]
- Chen, A.-H.; Chai, Y.-R.; Li, J.-N.; Chen, L. Molecular cloning of two genes encoding cinnamate 4-hydroxylase (C4H) from oilseed rape (Brassica napus). J. Biochem. Mol. Biol. 2007, 40, 247–260. [Google Scholar] [CrossRef]
- Lu, S.; Zhou, Y.; Li, L.; Chiang, V.L. Distinct roles of cinnamate 4-hydroxylase genes in Populus. Plant Cell Physiol. 2006, 47, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Yaghobi, M.; Heidari, P. Genome-Wide Analysis of Aquaporin Gene Family in Triticum turgidum and Its Expression Profile in Response to Salt Stress. Genes 2023, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Puresmaeli, F.; Heidari, P.; Lawson, S. Insights into the Sulfate Transporter Gene Family and Its Expression Patterns in Durum Wheat Seedlings under Salinity. Genes 2023, 14, 333. [Google Scholar] [CrossRef] [PubMed]
- Heidari, P.; Puresmaeli, F.; Vafaee, Y.; Ahmadizadeh, M.; Ensani, M.; Ahmadinia, H. Comparative Analysis of Phospholipase D (PLD) Gene Family in Camelina sativa and Brassica napus and Its Responses in Camelina Seedlings under Salt Stress. Agronomy 2023, 13, 2616. [Google Scholar] [CrossRef]
- Poulos, T.L. Heme enzyme structure and function. Chem. Rev. 2014, 114, 3919–3962. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, S.; Ahmadizadeh, M.; Heidari, P. Genome-wide characterization, expression profiling, and post- transcriptional study of GASA gene family. Gene Rep. 2020, 20, 100795. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Levy, H.L.; Sarkissian, C.N.; Scriver, C.R. Phenylalanine ammonia lyase (PAL): From discovery to enzyme substitution therapy for phenylketonuria. Mol. Genet. Metab. 2018, 124, 223–229. [Google Scholar] [CrossRef]
- Zhao, L.; Mali, G.; Yang, Z.A.; Feng, W.S.; Zheng, X.K. expression analysis of C4H gene from Lepidium apetalum. Acta Pharmacol. Sin 2017, 5, 821–831. [Google Scholar]
- Xia, J.; Liu, Y.; Yao, S.; Li, M.; Zhu, M.; Huang, K.; Gao, L.; Xia, T. Characterization and expression profiling of Camellia sinensis cinnamate 4-hydroxylase genes in phenylpropanoid pathways. Genes 2017, 8, 193. [Google Scholar] [CrossRef]
- Karlson, C.K.S.; Mohd Noor, S.N.; Khalid, N.; Tan, B.C. CRISPRi-mediated down-regulation of the cinnamate-4-hydroxylase (C4H) gene enhances the flavonoid biosynthesis in Nicotiana tabacum. Biology 2022, 11, 1127. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphology of Ferula pseudalliaceae. A. Young leaf development; B. Fully developed inflorescence covered for self-pollination and protection.
Figure 1.
Morphology of Ferula pseudalliaceae. A. Young leaf development; B. Fully developed inflorescence covered for self-pollination and protection.

Figure 2.
Phylogenetic tree of C4H in different plants. C4H genes from O. sativa (starting with Os), Brachypodium distachyon (starting with BRADI), Helianthus annuus (starting with HanXRQr), Beta vulgaris (starting with BVRB), Daucus carota (starting with DCAR), B. napus (starting with Bna), Arabidopsis (starting with AT), and C. sativa (starting with Csa) were used.
Figure 2.
Phylogenetic tree of C4H in different plants. C4H genes from O. sativa (starting with Os), Brachypodium distachyon (starting with BRADI), Helianthus annuus (starting with HanXRQr), Beta vulgaris (starting with BVRB), Daucus carota (starting with DCAR), B. napus (starting with Bna), Arabidopsis (starting with AT), and C. sativa (starting with Csa) were used.
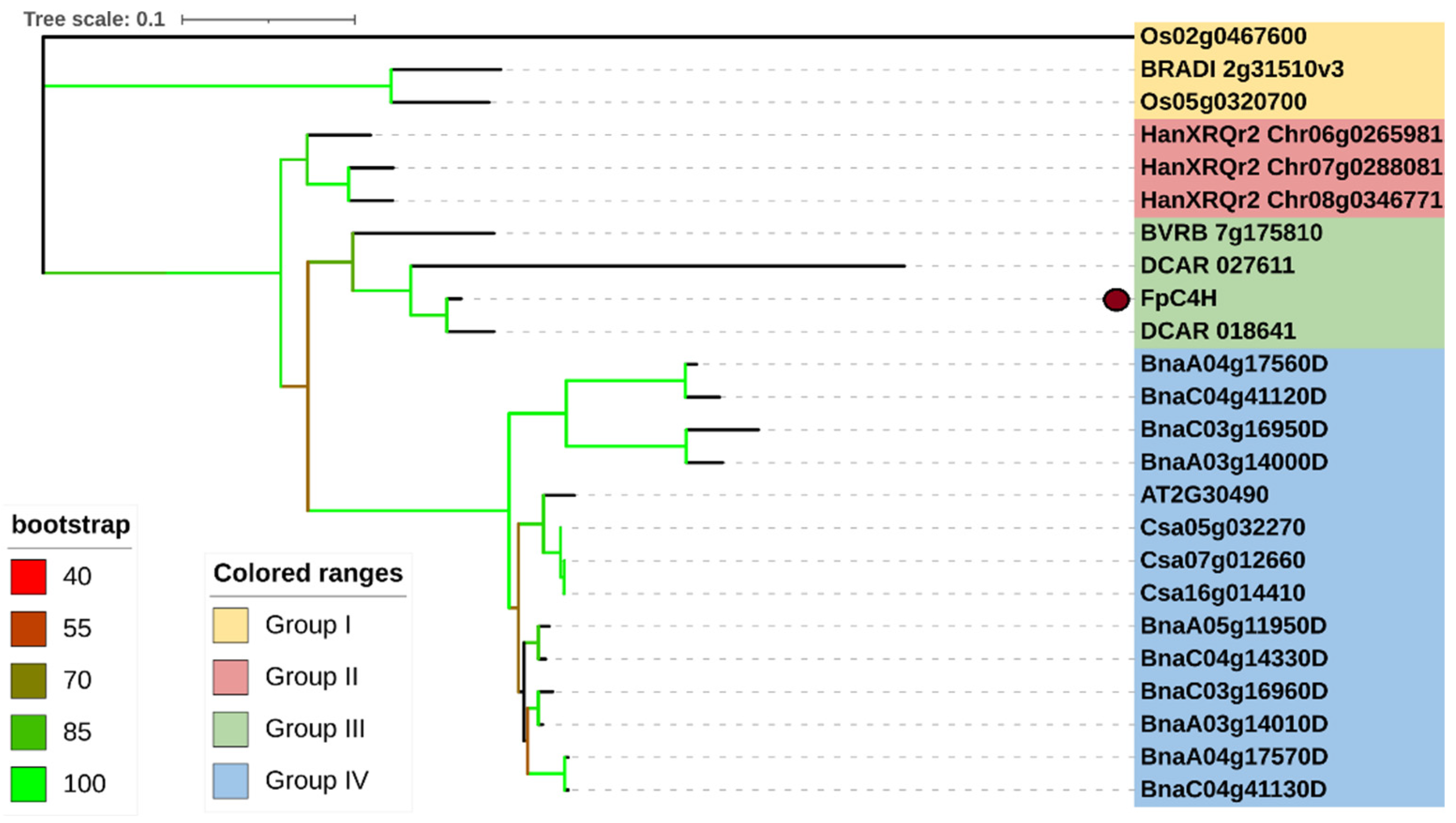
Figure 3.
Multiple sequence alignment analysis of FpC4H and its close orthologs.
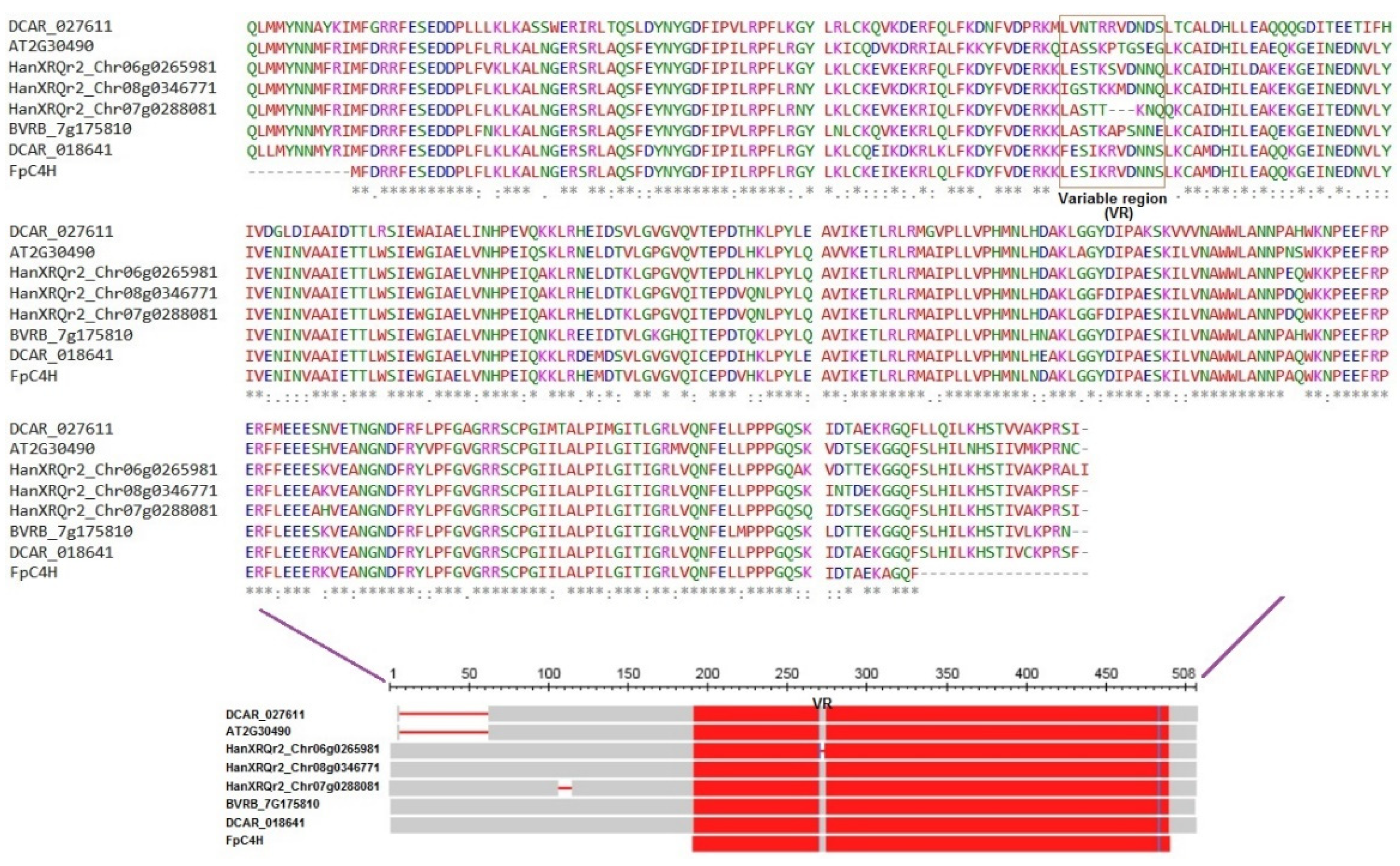
Figure 4.
3D structure of the FpC4H protein based on the homology method. Binding site of the FpC4H protein based on pocket analysis (a) and rate of mutation sensitivity of the structure of the FpC4H protein (b). Abbreviations: Alanine (A); Cysteine (C); Aspartic acid (D); Glutamic acid (E); Phenylalanine (F); Glycine (G); Histidine (H); Isoleucine (I); Lysine (K); Leucine (L); Methionine (M); Asparagine (N); Proline (P); Glutamine (Q); Arginine (R); Serine (S); Threonine (T); Valine (V); Tryptophan (W); Tyrosine (Y).
Figure 4.
3D structure of the FpC4H protein based on the homology method. Binding site of the FpC4H protein based on pocket analysis (a) and rate of mutation sensitivity of the structure of the FpC4H protein (b). Abbreviations: Alanine (A); Cysteine (C); Aspartic acid (D); Glutamic acid (E); Phenylalanine (F); Glycine (G); Histidine (H); Isoleucine (I); Lysine (K); Leucine (L); Methionine (M); Asparagine (N); Proline (P); Glutamine (Q); Arginine (R); Serine (S); Threonine (T); Valine (V); Tryptophan (W); Tyrosine (Y).
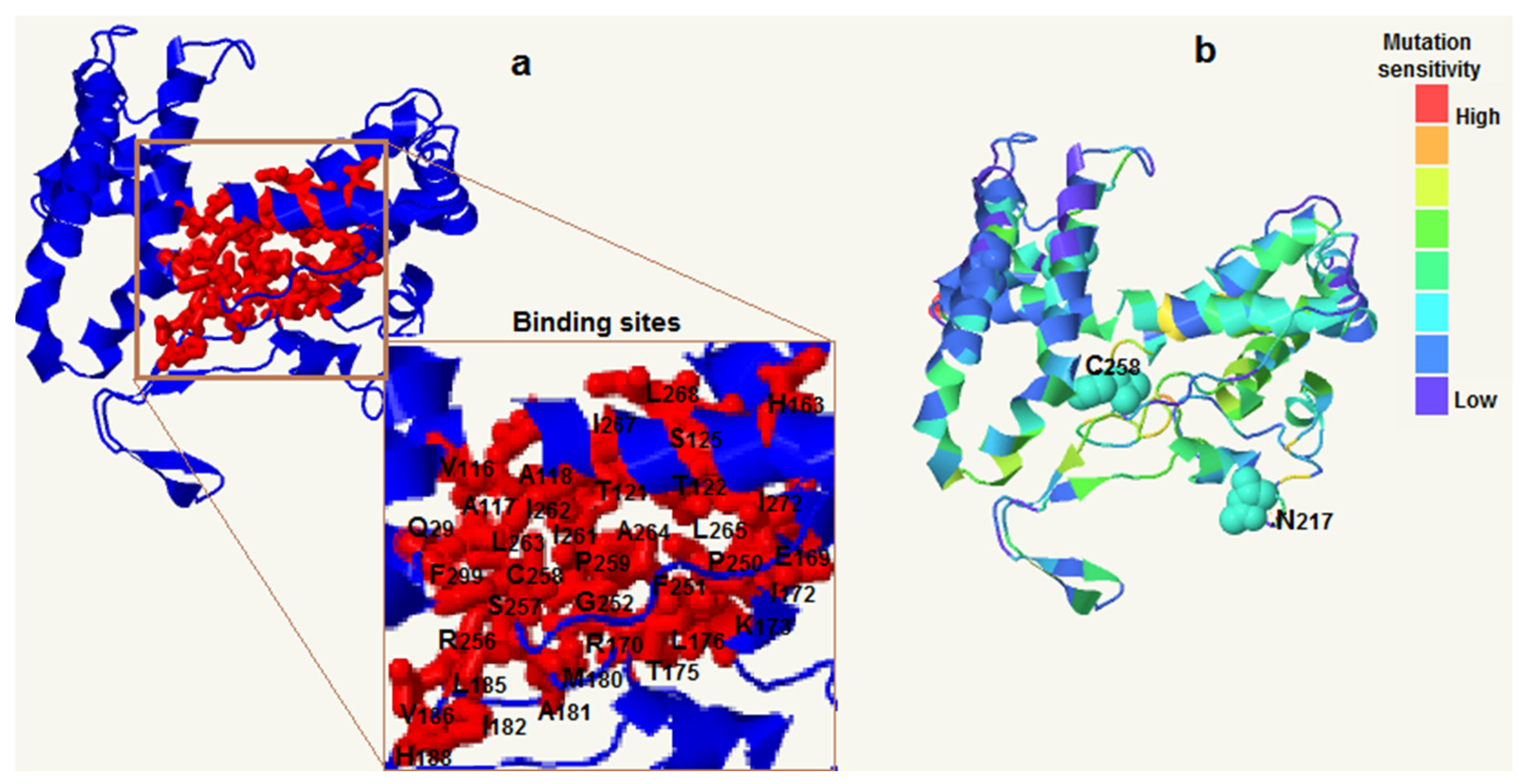
Figure 5.
Frequency (a) and distribution (b) of the FpC4H phosphorylation site. Phosphorylation regions were estimated with a confidence level above 90%.
Figure 5.
Frequency (a) and distribution (b) of the FpC4H phosphorylation site. Phosphorylation regions were estimated with a confidence level above 90%.
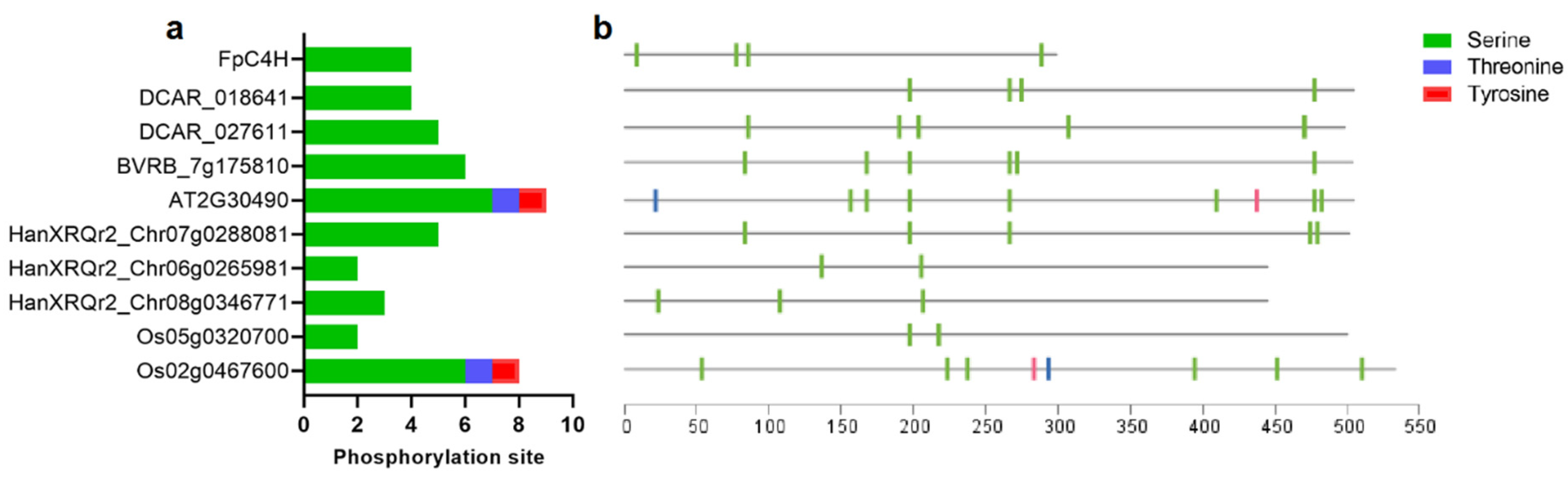
Figure 6.
Interaction network of the FpC4H protein based on available data from Daucus carota (a). GO enrichment analysis of network nodes in terms of molecular function (b) and biological process terms (c).
Figure 6.
Interaction network of the FpC4H protein based on available data from Daucus carota (a). GO enrichment analysis of network nodes in terms of molecular function (b) and biological process terms (c).
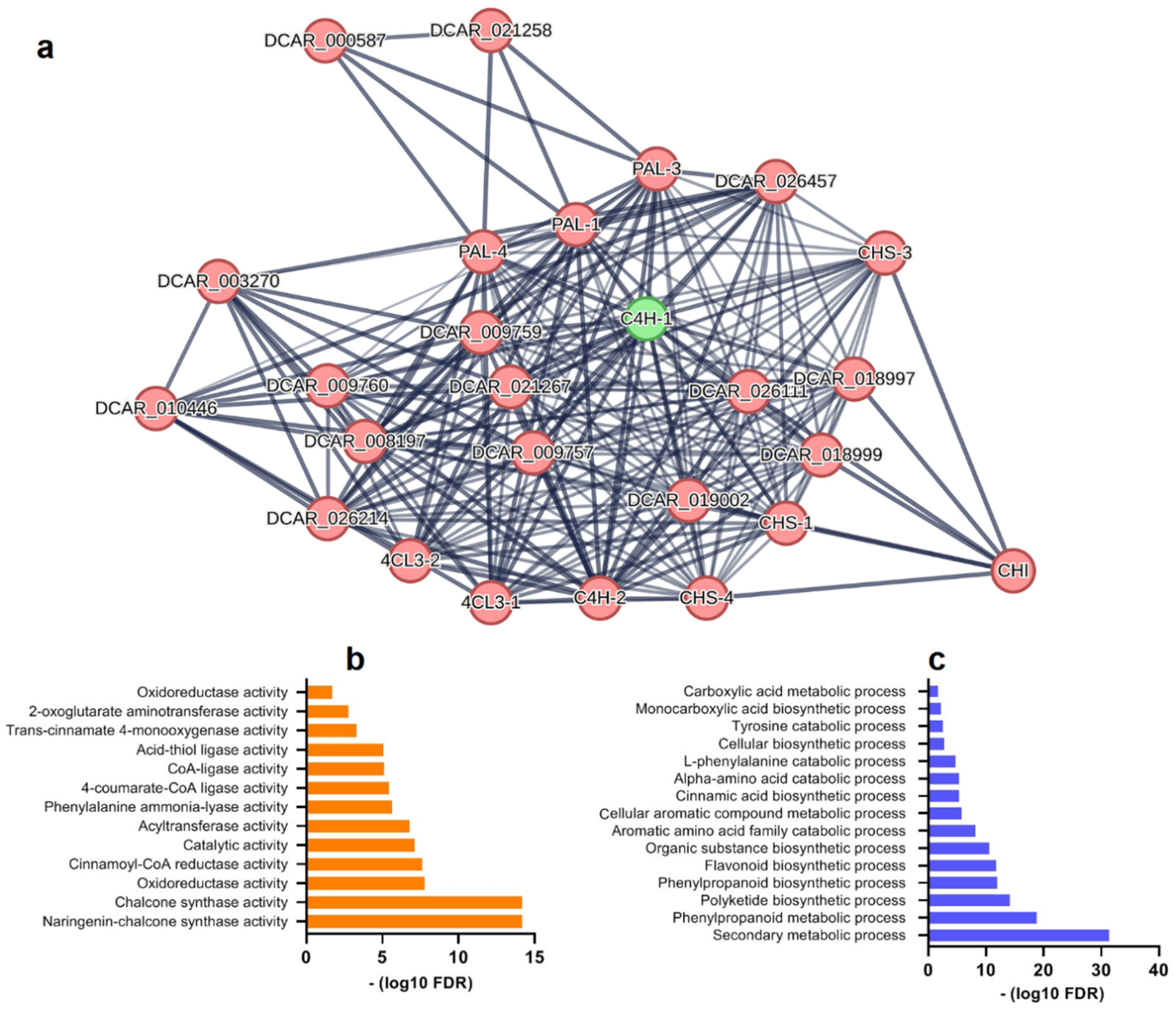
Figure 7.
Expression levels of the FpC4H gene in different organs of F. pseudaliacea. Columns with common letters have no significant difference at 5% probability level.
Figure 7.
Expression levels of the FpC4H gene in different organs of F. pseudaliacea. Columns with common letters have no significant difference at 5% probability level.
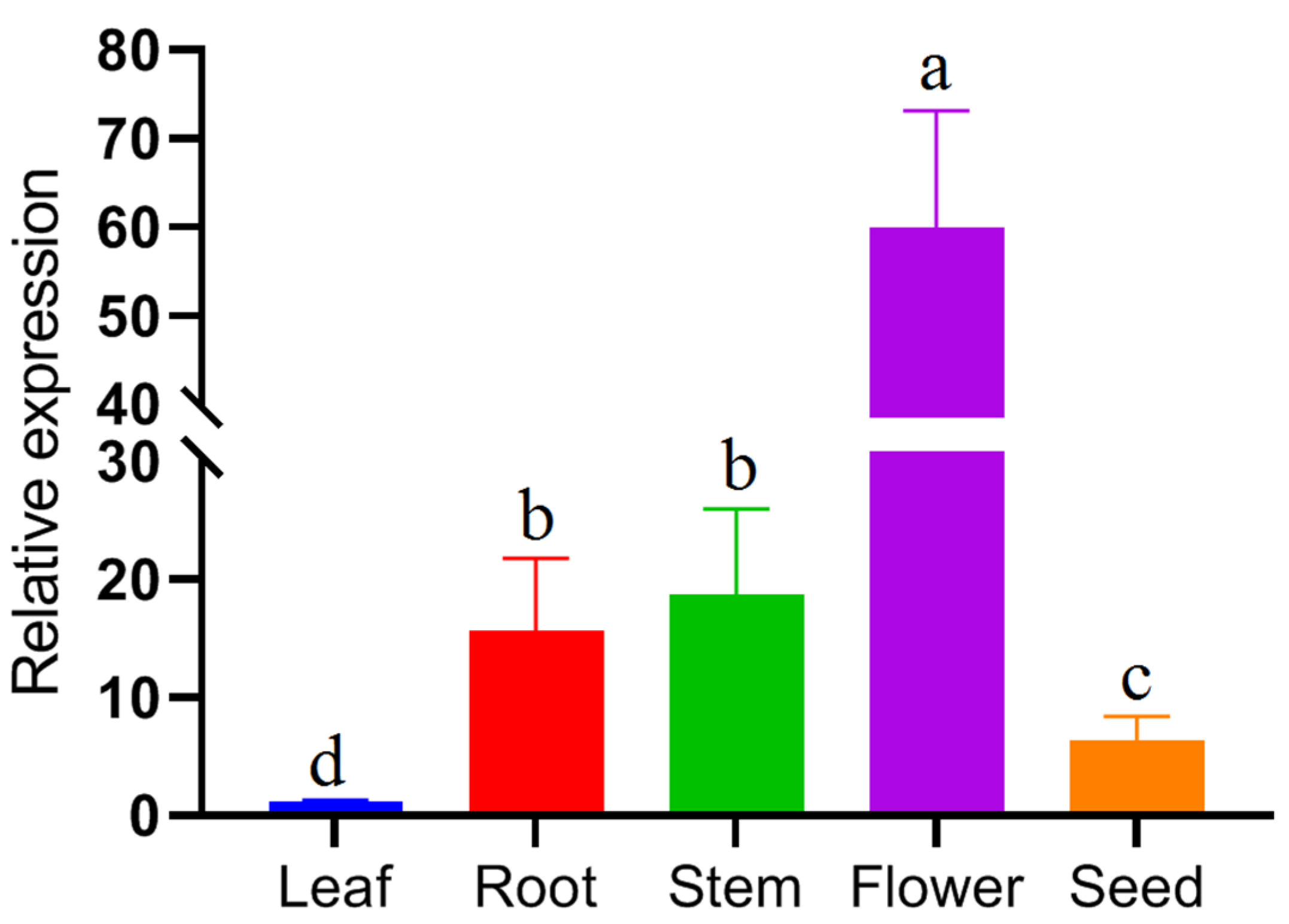

Table 1.
List of primers used in the present study to clone the FpC4H gene and to study the expression patterns of the genes in F. pseudalliacea organs. F: forward primer, R: reverse primer, RT: real-time PCR.
Table 1.
List of primers used in the present study to clone the FpC4H gene and to study the expression patterns of the genes in F. pseudalliacea organs. F: forward primer, R: reverse primer, RT: real-time PCR.
| Primer name | Sequence (5’-3’) | tm °C | Product size |
|---|---|---|---|
| FpC4H-F | TCATGTTTGATAGAAGGTTTGAGAG | 61 | 890 |
| FpC4H-R | GAACTGTCCTSCTTTCTCTGCTG | ||
| Oligo(dT) primer | GACCACGCGTATCGATGTCGACTTTTTTTTTTTTTTTTV | ||
| PCR anchor primer | GACCACGCGTATCGATGTCGAC | ||
| F-FpC4H.RT | ACAGCTTGAAGTGTGCCATG | 56 | 185 |
| R-FpC4H.RT | GCACAGTGTCCATCTCATGC | ||
| F-FpACTIN.RT | GCCATCTATGATTGGGAATGG | 56 | 190 |
| R-FpACTIN.RT | GCCACCACCTTGATCTTCATG |
Table 2.
Physicochemical properties of FpC4H and its orthologs.
| Gene ID | Plant | Length (aa) | MW (kDa) | Instability index | GRAVY | pI |
|---|---|---|---|---|---|---|
| FpC4H (MH987776) | F. pseudaliacea | 299 | 34.54 | Unstable | -0.342 | 5.88 |
| DCAR_018641 | Daucus carota | 505 | 58.10 | Unstable | -0.215 | 9.00 |
| DCAR_027611 | Daucus carota | 498 | 57.00 | Unstable | -0.216 | 9.21 |
| BVRB_7g175810 | Beta vulgaris | 504 | 58.09 | Unstable | -0.298 | 9.14 |
| HanXRQr2_Chr08g0346771 | Helianthus annuus | 505 | 57.93 | Unstable | -0.216 | 9.16 |
| HanXRQr2_Chr07g0288081 | Helianthus annuus | 502 | 57.55 | Unstable | -0.217 | 8.92 |
| HanXRQr2_Chr06g0265981 | Helianthus annuus | 505 | 57.93 | Unstable | -0.204 | 9.07 |
| AT2G30490 | A. thaliana | 505 | 57.79 | Unstable | -0.185 | 8.89 |
| Os02g0467600 | O. sativa | 533 | 59.48 | Unstable | 0.007 | 8.77 |
| Os05g0320700 | O. sativa | 500 | 56.88 | Unstable | -0.102 | 9.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



