
Submitted:
11 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
The South of France is a biodiversity hotspot within Europe. Here, we present a comprehensive review of surveys conducted in the Cent Fonts aquifer, an overlooked subterranean biodiversity hotspot embedded in a region rich in stygobiotic species and threatened by climate change and water abstraction projects. Key studies, spanning from 1950 to 2006, show a progression in survey methods and results, although troglobiotic species remain poorly documented. With 43 stygobiotic species recorded, the Cent Fonts is the second richest stygobiont hotspot in Europe. Most species are regional endemics, a quarter of which are considered vulnerable by the IUCN. The Cent Fonts also hosts several relict species and is the type locality of four species. Such a high biological value clearly deserves to be preserved. Our analysis warns of a possible decline in biodiversity, as 8 of the species recorded in the 20th century were absent from the 2006 survey, suggesting potential threats of unknown origin. The capture of the Cent Font springs for water abstraction is discussed as a potential threat to this ecosystem and its unique biodiversity. Three new species of stygobiotic molluscs are described, one of which was collected in the Cent Fonts.
Keywords:
stygobiont
; troglobiont
; conservation
; karst
; subterranean diversity
; water abstraction
1. Introduction
1.1. Karst and caves of the north-Montpellier region
The Cent Fonts aquifer is located in the southern region of France, in the Mediterranean basin, and is part of a larger karstic system comprising the Hérault (2,600 km2), Vidourle (800 km2) and Lez (200 km2) river basins. These regions are already acknowledged for their abundant and remarkable subterranean biodiversity [1,2].
1.2. Description of the Cent Fonts system
The karst system supplying the Cent Fonts is located in the western part of the northern Montpellier garrigues, formed by the limestone and dolomitic massifs located between Montpellier and the Cévennes. This karstic system develops within massive dolomites and oolitic limestones of Bathonian age (Middle Jurassic). The Cent Fonts aquifer is a binary karstic system, receiving its water supply from both the rainfall on the Causse-de-la-Selle plateau and a sinkhole from the Buèges River, a tributary of the Hérault, situated more than 8 km upstream (Figure 1). The average altitude of the plateau that forms the Cent Fonts catchment area is about 300 m; the Cent Fonts springs are located at an altitude of 81 m on the right bank of the Hérault River. These springs emerge in the Bathonian dolomite, close to a fault. The system consists of about ten resurgences spread over a 300m front, two observation points located a few meters higher, and the Cent Fonts cave, the entrance of which is situated a few meters above the observation points.
The spring has been explored by cave divers, one of whom died in 1984. The divers have reached a depth of -95m, about 150m from the cave entrance, and were blocked by a narrow passage.
The Cent Fonts system is the most important emergence of the Causse-de-la-Selle plateau. The land use in its catchment area consists mainly of evergreen oak forests (Quercus ilex L., 1753) and extensive pastures. Human settlement in this area is very limited, and the presumed anthropogenic impacts are low. The Cent Fonts site falls within several protected areas (Natura 2000 site FR9101388 – Gorges de l’Hérault; classified site; Grand Site de France; ZNIEFF [Natural Areas of Floristic and Faunistic Interest]). This site stands out for its particular vegetation associations (Salzman Pine forest), rare bird species such as the Bonelli eagle Aquila fasciata Vieillot, 1822 and the Cinereous vulture Aegypius monachus (Linnaeus, 1766), and some rare insect species, including an endemic beetle, Cryptocephalus mayeti Marseul, 1878. Rarely mentioned however is its exceptional richness in stygobiotic invertebrates.
1.3. History of biological studies
Following the description of the subterranean crustacean Gallocaris inermis (Fage, 1837) in the Gard department and its subsequent discovery in other aquifers bordering the Hérault, the aquatic fauna of the Cent Fonts massif has undergone more extensive exploration. Initially, it was the subject of sporadic investigations utilizing rudimentary tools such as nets and baited traps [3,4,5], which revealed the presence of four large-sized crustacean species: Gallocaris inermis, Faucheria faucheri (Dollfus & Viré, 1900), Sphaeromides raymondi Dollfus, 1897, Niphargus virei Chevreux, 1896), and an ostracod species, Sphaeromicola cebennica Rémy, 1948, a parasite of Sphaeromidesi (Table 1).
A second inventory dedicated to the Cent Fonts aquifer dates from 1967-1968 [6]. The system was studied under natural conditions, including all the springs and the cave. This study aimed primarily to characterize the stygobiotic fauna within the submerged zone of the karst. More methods were used and the inventory of the stygobiotic fauna was more comprehensive. Thirty-nine crustacean species were collected, including 20 stygobiotic species belonging to the orders Decapoda, Amphipoda, Isopoda and Copepoda (Table 1). The authors concluded that the Cent Fonts aquifer was "exceptionally rich”. The molluscs were not mentioned in the paper [6]. Their diversity in the northern Montpellier region was only studied later by Prié [7,8,9] and Girardi [10,11], but without focusing on the Cent Fonts aquifer.
In 2004, a water resource exploitation project prompted additional studies. A more thorough inventory of the stygobiotic fauna was conducted between 2005 and 2006 [12], employing extended filtrations of effluents during low-flow and flood periods, along with experimental pumping.
1.4. Threats
As mentioned above, the landscape surrounding the Cent Fonts system is relatively unaffected by human activities. However, this system is seen by the authorities as a major water resource for the entire department [13]. This human pressure on the water supply is believed to increase in the future, especially as the local climate is already dry and drought is expected to increase in a context of global warming.
1.5. Objectives
The aims of this paper are (i) to summarise the biospeleological studies carried out at Cent Fonts and in the surrounding caves, in particular the work of Olivier et al. [12], which has never been scientifically published, (ii) to update the taxonomy of the species present, with the description of new gastropod species, (iii) to highlight the interest of the site as a biodiversity hotspot and (iv) to discuss the impact that aquifer exploitation project could have on this hotspot.
2. Materials and Methods
We define the "Cent Fonts system" as the area drained by the Cent Fonts springs, extending from the Buèges River in the north to the Hérault River in the east and south, and an inactive valley to the east that, together with the rivers, outlines the boundaries of the "Causse de la Selle." The surveys were all carried out in the Cent Fonts springs, which are closed to the Hérault River south-east of the Causse de la Selle (Figure 1).
The first surveys (1950 – 1951) used very simple methods such as dip nets and baited traps. Rouch et al. [6] in 1968 used a more comprehensive range of methods, including dip nets and baited traps, but also sight-hunting in each siphon (method only valid for large crustaceans); fine-netting, carried out in all siphons using a Bluter silk net; pumping with the Bou-Rouch pump [14], which allows large quantities of water to be pumped from the interstitial zone, at a depth of about 60 cm in the sandy clay alluvium at the bottom of the siphon and 1 to 2 m from the edge in the gravel at the front of the spring formed by the exsurgences and in the alluvia of the Hérault river, a little downstream of the Cent Fonts springs; Karaman-Chappuis boreholes drilled in the siphon banks; filtering of all the exsurgences with Bluter silk nets of various mesh sizes, left in place and lifted every week. Some water outlets were filtered almost continuously from 15 November 1967 to 23 February 1968.
The same methods were used in the years 2005 and 2006: Bou-Rouch pumping (Figure 2), surbers, spring water filtration; baited traps, sight hunting, plus sediment sampling for mollusc shells in the springs and in the subterranean environment (Cent Fonts cave). The latter method consists of sampling sediment and leaving it in a bucket of spring water for a few days, in a cool and dark environment (eg. a house cellar). As oxygen becomes scarce, the animals will try to return to the surface and can be caught on the sides of the bucket with flexible forceps. After a few days, when no live snails are found, the sediment is dried out and poured into water again. The grains of sand will sink, and the empty shells will float to the surface and can be collected with a sieve.
All the data presented here have been deposited in the Inventaire National du Patrimoine Naturel (https://inpn.mnhn.fr) database. The site number of the Cent Fonts is 2047774. The sequences produced for the description of the new species (Appendix A) are deposited in GenBank, accession numbers XXX to XXX.
3. Results
Following this study, the Cent Fonts aquifer appears as one of the richest systems for stygobiotic taxa, with 43 cave-restricted species (Table 1). The terrestrial taxa, which are presumably not as rich as the aquatic ones, have not been studied in the Cent Fonts system itself. We present here the results of surveys carried out in neighbouring caves located on the right bank of the Hérault valley, in the same geological context.
Where available, the IUCN Red List category is given for each species at global and national levels. Mollusc species were assessed at global level in 2010 [18] and at regional level in 2021 (French Red List [19]). Although most species are regional endemics, the 2010 (global) and 2021 (French) assessments sometimes differ. This is mainly due to an increased awareness of the threats to aquatic ecosystems, as human and climate change threats are increasingly documented. Most crustacean species have not been assessed at the global level, but a regional level assessment is available.
3.1. Stygobionts
3.1.1. Clitellata Michaelsen, 1919; Arhynchobdellida Blanchard, 1894
- Trocheta taunensis Grosser, 2015 (= T. bykowskii)
Several populations of leeches named T. bykowskii have been discovered in Central and Western Europe [15]. Sket [16] was the first to suggest that T. bykowskii actually represents a species complex. Following Grosser [17], Lecaplain [15] considers the French populations to belong to T. taunensis. However, the records of T. taunensis in France are only from eastern France. The taxonomic status of the Cent Fonts population remains to be confirmed. The species was found in the Cent Fonts cave by F. Malard in 2002 (unpublished data).
3.1.2. Gastropoda Cuvier, 1795; Littorinimorpha Golikov & Starobogatov, 1975
3.1.2.1. Amnicolidae Tryon, 1863
- Bythinella sp.
A species of Bythinella was found in abundance in the springs of Cent Fonts (Figure 3a). It was considered as a new species by Olivier et al. [12], based on the fact that it lives in a different aquifer from the regional stygobiotic Bythinella species described so far, i.e. Bythinella navacellensis Prié & Bichain 2009 endemic to the Larzac plateau (north-west of Cent Fonts) and B. eutrepha (Paladilhe, 1867) endemic to the Lez karst (south-west). Its identity remains unclear as no genetic data have been collected.
3.1.2.2 Hydrobiidae Stimpson, 1865
- Heraultiella exilis (Paladilhe, 1867)
Heraultiella exilis lives in the hyporheic zone [7]. It sometimes occurs in springs but has never been found deep inside the caves. Here, it has been found in the hyporheic zone of the Hérault River, and marginally in the springs. This species is protected in France and considered Vulnerable on both the international [18]and French Red List [19].
- Islamia cf. moquiniana (Dupuy, 1851)
The genus Islamia is awaiting a molecular revision. Islamia moquiniana is described from the department of Lozère (type locality “… alluvions du Lot à Mende”), far from the Cent Fonts, and the specimens collected in the Hérault basin are morphologically different from those from the department of Lozère. It is therefore likely that the population found in the Cent Fonts is part of an undescribed species.
3.1.2.3 Moitessieriidae Bourguignat, 1864
- Paladilhia pleurotoma Bourguignat, 1865
P. pleurotoma is restricted to a few karst areas east of the Hérault River and west of the Rhône River. It is a cave specialist and has never been collected alive in the hyporheic zone. It is not certain that the hyporheic zone can be used by this species as a corridor, as is the case for Bythiospeum species. Only one shell was found in the Cent Fonts cave (Figure 3b). This shell could be allogenic (transported there by flood). This species is protected in France. It was listed as Least Concerned in the IUCN international red list in 2010 [20] but re-evaluated as Vulnerable on the French Red List in 2021[19].
- Bythiospeum bourguignati (Paladilhe, 1866)
This species lives mainly in the karst on the left bank of the Hérault [8]. It is found in the hyporheic zone of the Hérault and has been marginally collected in the sediments of the springs of the Cent Fonts. It is thought to reach its westernmost distribution limit in the Cent Font, which is also the westernmost limit of the genus. This species is protected in France. It was listed as Least Concerned in the IUCN international red list in 2010 ([21]) but re-evaluated as Near threatened on the French Red List in 2021 [19].
- Moitessieria vidourlensis n. sp. (= Moitessieria rolandiana Bourguignat, 1864)
Most authors consider M. rolandiana as a widespread species in southern France, west of the Rhône River. However, Prié [9] showed that there is a strong genetic structure within the area of occurrence of M. rolandiana, which reflects the structure of the hydrographic network. A description based on morphometric and molecular data is provided hereafter (Appendix A). M. vidourlensis n. sp. (Figure 3c) is morphologically close to M. rolandiana, but can be distinguished by morphometric analysis. This species is protected in France under the name Moitessieria rolandiana.
- Moitessieria guilhemensis Girardi & Boeters, 2017
This species was first recognized by Prié [22] based on morphological data (shells larger and smaller than that of M. rolandiana, Figure 3d), but not described as a new species because no genetic data was available, and morphology has proven to be misleading for stygobiont species. However, Girardi and Boeters [23] couldn’t wait and described the species as M. guilhemensis. This species is protected in France under the name Moitessieria rolandiana.
- Moitessieria n. sp. ?
A spectacular shell was collected at the Cent Fonts (Figure 3e), perhaps a monstrous form, perhaps something new. As this is a single shell, we prefer not to consider it as a new species, pending further data, but we do report this remarkable form.
3.1.3. Malacostracea Latreille, 1802
3.1.3.1 Decapoda Latreille, 1802
- Gallocaris (Troglocaris) inermis (Fage, 1937)
A spectacular species (Figure 4), measuring up to 2 cm long; one of only wo species of stygobiont decapod in France, endemic to a few aquifers in the Gard and Hérault valleys where it is known from less than 10 localities. Interestingly, Rouch et al. [6] noted that this species only occurs in streamless waters inside the cave. Its supposed rheophobia may explain why it has never been collected in the springs, even during floods. It is listed as Near Threatened on the IUCN global Red List [24]; Vulnerable on the French Red List [25].
3.1.3.2. Isopoda Latreille, 1816
- Stenasellus buili Rémy, 1949
Described from the department of Aude, with isolated populations in the Corbières mountains and here in the Hérault valley. It is not evaluated at the international level but listed as Near Threatened on the French Red List [25].
- Proasellus cavaticus (Leydig, 1871)
Widespread in Western Europe, occurring in France along the Rhône-Rhine axis and in the Haut-Languedoc (marginally present in the Atlantic basin). According to Henry [26], the population of Cent Fonts belongs to P. cavaticus cavaticus and is remarkable because it is the most western and the only place where Stenasellus and Proasellus cavaticus occur in syntopy. The species is considered Least Concern on the French Red List [25].
- Microcharon doueti Coineau, 1968
Discovered by Rouch by filtering the exsurgences of the Cents-Fonts (type locality) and then collected in the water table of the Orb River, west of the Hérault River. Listed as Vulnerable on the French Red List [25].
- Faucheria faucheri (Dollfus & Viré, 1900)
Originally described by Adrien Dollfus and Armand Viré in 1900 as Cæcosphæroma faucheri (family Sphaeromatidae), it was reclassified by the authors in 1905 in the family Cirolanidae. Bertrand [27] lists a total of 21 stations, 10 in the upper Vidourle valley and the Hérault gorges and 11 in the eastern Corbières (Agly basin and its tributary the Verdouble). We (C.A.) add here another locality, the outlet of the Avencas cave, near Issensac, which extends the distribution of the species south to the coast. Least concern on the French Red List [25].
- Sphaeromides raymondi Dollfus, 1897
S. raymondi (Figure 5) is a large species, up to 3 cm, known from a few caves in the Hérault department and the right bank drainage of the Rhône River, up to the Ardèche River. This species is mentioned in the literature from the 1950s and by Rouch et al. [6] but was not found during the 2006 sampling. Listed as Near Threatened on the French Red List [25].
3.1.3.3 Amphipoda Latreille, 1816
- Niphargus laisi Schellenberg, 1936
The species is widespread in France and Germany. In France, its distribution is sporadic, from the Alsace in the north to the Rhône River aquifer near Lyon, and in the south in the Hérault basin. Its habitat is mainly represented by the hyporheic and phreatic zones. Considered Data Deficient on the French Red List [25].
- Niphargus gallicus Schellenberg, 1935
This species is scarce in the southern half of France, where it lives in the porous aquifers of large alluvial floodplains (Rhône) and small streams (e.g. Triouzoune, St Angel), both in the phreatic and hyporheic zones. It has also been collected in karst areas (e.g. Prades-Le-Lez). Least Concern on the French Red List [25].
- Niphargus kochianus Bate, 1859
Niphargus kochianus had several subspecies, of which N. k. dimorphopus (present in Belgium and the Netherlands) and N. k. irlandicus (restricted to Ireland), are now considered as separate species. Niphargus. k. kochianus is well established in Great Britain. Its presence in France, although frequently reported, is considered doubtful as it would not have a transcontinental distribution [28]. McInerney et al. [29] defined four distinct clades (A, B, C, D) based on molecular analysis. N. kochianus “D” would be the lineage present in France. The French form is sparsely distributed from the extreme north to the south (Pyrenees region), but is more common in the Rhône basin, Jura and Ardèche regions. Given the large number of sites and specimens reported from France, it is difficult to provide a clear taxonomic status for the N. kochianus collected in the Cent Fonts system. Moreover for this French “lineage D” the number of sites and specimens in the 2014 study [29] appears to be very low.
From an ecological point of view, it is a small species, typically interstitial, living in cool waters and stable flow conditions. In French aquifers N. kochianus is often found in the upwelling zones of rivers (e.g., Rhône), or in deep alluvial and phreatic zones (e.g. wells in the Albarine valley, Jura). It is also reported in karst, where it may find conditions for an interstitial lifestyle. Least Concern on the French Red List [25].
- Niphargus pachypus Schellenberg, 1933
Previously described as a subspecies of N. kochianus, it has been raised to species level and is now recognised as a highly divergent lineage [30]. It was collected from only few sites in the Netherlands and is also reported from Belgium and Luxembourg. In France, it is widespread, with more than a hundred localities. As a small-sized species, it is typically interstitial and particularly prefers cool and hydrologically stable areas, which explains its abundance in the deep alluvia of streams and in the phreatic zone. However, it has also been collected in the karst, where it is probably associated with alluvial deposits. Least Concern on the French Red List [25].
- Niphargus cf. virei Chevreux, 1896 Clade A
Found mainly in France, but also in a few places in the Netherlands, Belgium and Switzerland. In France it is typically a karstic species. Niphargus virei has never been found in the porous aquifer, except for one specimen collected in the alluvia of the Rhône-Ardèche confluence, probably drifted from the surrounding karst. It is particularly common and abundant in the Jura and Ardèche massifs. Genetic studies have revealed the presence of three cryptic species in the French virei group [31]. The population of the Cent Fonts karst system belong to the cryptic species “A”, located at the extreme south of the group's geographical distribution. N. virei has been described from specimens collected in caves of the Jura mountains (grottes d’Arbois, Baumes-les-Messieurs and Baumes-les-Dames). The nominal species should then belong to clade “B” of Lefebure et al. [31]. The most widespread of these cryptic species- (Clade A), which is found from the Hérault to the Rhône and Moselle Rivers, is still awaiting formal description. Species considered Least Concern on the French Red List [25].
- Salentinella angelieri Delamare-Deboutteville & Ruffo, 1952
With a wide geographical distribution in Greece, Italy and Spain, it is less common in France, where it is mainly recorded from the Rhône basin and the Hérault region. A stand is also reported from Corsica. Two subspecies have been described from Croatia and Spain. In the Rhône floodplain it is always collected in upwelling, i.e. in cool and stable interstitial water. Listed as Near Threatened on the French Red List [25].
- Salentinella delamarei Coineau, 1962
Described from the phreatic waters of the Tech River in the department of Pyrénées-Orientales. Reported only from France along the Rhône River, the Ardèche and Hérault areas. Two subspecies: S. delamarei delamarei and S. delamarei macrocheles. Species considered Least Concern on the French Red List [25].
3.1.6 Ingolfiellida Hansen, 1903
- Ingolfiella thibaudi Coineau, 1968
This species has been reported from less than fifteen sites, from Ruoms in the Ardèche to Tarbes and the Saint Girons area in the Pyrenees region. It has been collected in both karst and porous aquifers (hyporheic and phreatic zones). In the Cent Fonts system, several specimens have been found in the spring sediments and in the hyporheic zone of the Hérault river (Figure 7). Species considered Least Concern on the French Red List [25].
3.1.7 Ostracoda Latreille, 1802; Podocopida Sars, 1866
- Fabaeformiscandona cf. breuili (Paris, 1920)
This species is widespread in Europe, from Poland to Spain, and certainly represents a number of subspecies or cryptic species. The taxonomic status of the Cent Fonts population needs to be clarified. On a European scale, F. breuili has been sampled in different habitats: wells, springs, hyporheic zone of rivers and, more rarely, in caves. In the Cent Fonts, F. breuili has only been sampled with exsurgence filtering, but not in the hyporheic zone. Species Least Concerned for the UICN (French Red List [25]).
- Marmocandona (Pseudocandona) cf. zschokkei (Wolf, 1920)
Originally described in the genus Candona, then included in the genus Pseudocandona, Danielpol et al. [32] proposed the genus Marmocandona (whose type species is Candona zschokkei Wolf, 1920) for four stygobiotic species. This species is widespread in Western Europe: in Switzerland, Germany, Belgium, and France. The taxonomic status of this southern population needs to be clarified. M. zschokkei was often sampled in the hyporheic zone of large rivers, but also occurred in springs and wells. In the Cent Fonts, M. zschokkei was sampled with exsurgence filtering, but not in the hyporheic zone of the river. Species Least Concern on the UICN French Red List [25].
- Schellencandona cf. simililampadis (Danielpol, 1978)
Species previously restricted to an artificial cave associated with the Vidourle spring at Sauve (Gard department). The taxonomic status of the population sampled in the Cent Fonts needs to be clarified. Species sampled with exsurgence filtering. Listed as Vulnerable on the French Red List [25].
- Sphaeromicola cebennica juberthiei Danielpol, 1977
Currently known from only two sites in the Hérault valley: the Cent Fonts and another cave a few kilometres upstream, also on the right bank of the Hérault River. This species is mentioned in the literature from the 1950s and by Rouch et al. [6] but was not found during the 2006 sampling. Listed as Vulnerable on the French Red List [25].
- Candoninae sp. 1, 2, 3
Three other species of the subfamily Candoninae were sampled during the 2006 Cent Fonts study, but only with juveniles: a “long” form, related to the genus Cryptocandona; a “bean-shaped” form, related to Pseudocandona; and a triangular form, related to the Pseudocandona group eremita. Their taxonomic status still needs to be established by examination of adult specimens. However, although they could not be formally identified to species level, they represent other species than those listed above.
3.1.8. Bathynellacea Chappuis, 1915
- Gallobathynella (Clamousella) delayi Serban, Coineau & Delamare Deboutteville 1971
This species was previously considered strictly endemic from the Clamouse Cave, a few kilometers downstream the Hérault Valley, also on the right bank. Listed as Vulnerable on the French Red List [25].
3.1.9. Copepoda Milne Edwards, 1840
3.1.9.1. Cyclopoida Burmeister, 1834
- Acanthocyclops rhenanus Kiefer, 1936
This obligate groundwater cyclopoid shows a wide distribution in many groundwater habitat types of Europe. Its distribution covers several countries in central-eastern Europe, from France to Poland. The species shows no apparent habitat specialisation being recorded from almost all the groundwater habitat types. This species is mentioned by Rouch et al. [6] but was not found during the 2006 sampling. Least Concern in France [25].
- Acanthocyclops venustus (stammeri) cf. westfalicus (Kiefer, 1931)
This species has an alternate representation in the current literature, and at present knowledge the accepted name for the subspecies westfalicus is A. venustus venustus [33]. The venustus group of the genus Acanthocyclops is in need of revision, and pending a clearer taxonomic assessment, the subspecies name westfalicus is here provisionally maintained. This subspecies has been recorded from Germany, Belgium, and France, and collected from phreatic habitats, the hyporheic zone of rivers, and aquifers in unconsolidated sediments. A. venustus is considered as Vulnerable on the French Red List [25].
- Graeteriella boui Lescher-Moutoué, 1974
This species is known only from France, with 11 records from both alluvial and karst aquifers, with a higher incidence in the saturated karst. It was originally described on the basis of specimens collected in the Gard department, but in the description the author mentions the Ardèche and Hérault populations as belonging to the same species (“[The description of Graeteriella boui is based on individuals caught in the Gard department. Other forms collected in neighbouring departments reproduce the same characteristics; some differences, not sufficient to introduce new systematic subdivisions, are noted below]”). The population studied in the Hérault basin is that of the Cent Fonts. And Lescher-Moutoué [34] concludes: "The presence of G. boui in the Cent-Fonts karstic system is all the more remarkable because two species of this genus have also been recorded in the same system: G. unisetigera and G. (Paragraeteriella) vandeli Lescher-Moutoué, 1969". The species has also been collected from the karst aquifer of the Lez River. It is considered as Vulnerable on the French Red List [25].
- Graeteriella unisetigera (Graeter, 1908)
This species is considered by Fiers & Ghenne [35] to be a member of the cryptozoic fauna, as it has also been found in leaf litter and in other surface habitats (e.g. mosses) in Belgium, usually with some connection to groundwater. In spite of this situation, the species has several morphological characteristics that make it a good candidate for a widespread stygobiotic species in Europe, able to live in true groundwater habitats as well as in surface ecosystems dependent on groundwater. It is mentioned by Rouch et al. [6] but was not collected again during the 2006 sampling. Least Concern in the French Red List [25].
- Graeteriella (Paragraeteriella) vandeli Lescher-Moutoué, 1969
Rouch et al. [6] mention “Paragraeteriella n. sp.”, without any further details. It was later described as Paragraeteriella vandeli by Lescher-Moutoué [36]. The type locality is the Cent Fonts. It is known only from a single record from the Cent Fonts karstic system, which makes it spot endemic to this restricted area and rare in terms of abundance. At present it has only been collected from the saturated karst. It was not found during the 2006 survey. Listed as Vulnerable on the French Red List [25].
- Kieferiella delamarei (Lescher-Moutoué, 1971)
This cyclopid species has exceptional stygomorphic features, such as a slender body, completely depigmented, long antennules and long setae on the swimming legs, which make it a typical planktonic species swimming in underground karst lakes. This species is known from the Lez karst system and has also been collected from the Cent Fonts karst springs. The genus Kieferiella is monotypic and the only known species is from this restricted area in the south of France, making it a priority for conservation. Listed as Vulnerable on the French Red List [25].
- Speocyclops racovitzai (Chappuis, 1923)
This species is mentioned by Rouch et al. 1968 as “Speocyclops sp. (en cours de determination)”. It was not found during the 2006 sampling. S. racovitzai is present throughout southern France [37]. It shows a high degree of diversification in morphological micro-characteristics and is therefore divided into several subspecies with subtle morphological differences. No less important, some subspecies show overlapping distributions, raising doubts about their subspecific identity. The currently recognized subspecies need a taxonomic redefinition, but all are considered stygobionts. The nominotypical species also shows a wide distribution in the Pyrenees. It is listed as Least Concern on the French Red List [25].
3.1.9.2. Harpacticoida Sars G.O., 1903
- Ceuthonectes gallicus Chappuis, 1928
This species is widespread in France and always associated with groundwater habitats, both in alluvial and karst aquifers, with some preference for the latter [38]. French endemic, of Least Concern on the French Red List [25].
- Elaphoidella leruthi meridionalis Chappuis, 1953
The genus Elaphoidella is one of the most diverse harpacticoid genera in groundwater environments. In the study area, E. leruthi meridionalis is the only species recorded. It is known from several sites, mainly in southern France, with a clear preference for karstic groundwater, both in the saturated and unsaturated zones. E. leruthi is considered Data Deficient on the French Red List [25].
- Nitocrella omega Hertzog, 1936
The ameirid genus Nitocrella is considered to be of ancient direct marine origin and almost all species of this genus are known only from groundwater habitats [39]. This species collected from the Cent Fonts is rare in terms of occurrence and abundance, being known from only a few localities in France, Germany and Hungary. Listed as Vulnerable on the French Red List [25].
- Nitocrella hirta Chappuis, 1924
This species is widespread throughout Europe, with more than forty localities and collected from many groundwater habitat types. Four subspecies have been described. Not evaluated on the French IUCN Red list.
- Pseudectinosoma vandeli (Rouch, 1969)
This minute harpacticoid was the first Pseudectinosoma species discovered in groundwater worldwide. The species was first mentioned by Rouch et al. in 1968 as “Ectinosomidae sp. ». A year later, Rouch described it and placed it in the marine genus Sigmatidium. It was only later that Galassi et al. [40] re-analysed the type material of the type species of the marine genus Sigmatidium on the occasion of the discovery of the second representative of the genus Pseudectinosoma in France, and definitively placed this species in the genus Pseudectinosoma. The genus Pseudectinosoma is considered to be an ancient Tethyan relic found in the groundwater of Europe and Australia, probably the only remnant of an ancient fauna of direct marine origin. P. vandeli is known only from this area and has been collected in large numbers from the Cent Fonts karst system. The Cent Fonts is the type locality of the species. Listed as Vulnerable on the French Red List [25].
3.1.10. Arachnida, Acari
- Soldanellonyx chappuisi Walter, 1917
This species is mentioned by Rouch et al. [6] but was not found (but not sought for) during the 2006 sampling.
3.2. Troglobionts
Unlike stygobionts, troglobionts have not been inventoried in the Cent Fonts system. The only troglobiont species collected in the Cent Fonts cave is the carabidae beetle Laemostenus (Actenipus) oblongus balmae. There is currently no report of other troglobitic taxa in the Cent Fonts system itself. On the assumption that troglobionts are less drainage-dependent than stygobionts, we list below species which most likely occur in the Cent Fonts, since they occupy many caves in the surroundings.
3.2.1. Araneae
- Palliduphantes sanctivincenti (Simon, 1873)
Endemic from southern France, widespread between the Pyrenees and the Alpes.
3.2.2. Opiliones
- Peltonychia clavigera (Simon, 1872)
The genus Peltonychia contains the first described travunioid species. This polyphyletic genus is only known from Central European caves in the Pyrenees and the Alps. Peltonychia clavigera is distributed on both slopes of the Pyrenees and in the Cevennes where it is sporadic (Figure 8A).
3.2.3. Pseudoscorpiones
- Neobisium tuzetae Vachon, 1947
Described from the Signal de la Montete cave towards Quissac in the Gard depart-ment, this species is found in a large number of caves from the Hérault valley to the Larzac plateau (Figure 8B).
3.2.4. Isopoda
- Trichoniscoides bonneti Vandel, 1946
This endemic species is quite common in the caves of the limestone edge of the Cevennes, between the Hérault and Vidourle rivers (Figure 8C).
3.2.5. Diplura
- Plusiocampa balsani Conde, 1947
An endemic species found in many caves in the Massif Central. It is very common in all the caves of the Hérault valley (Figure 8D).
3.2.6. Collembola
- Pseudosinella denisi Gisin, 1954
Endemic from the sub-region (Gard, Ardèche and Hérault Departments). Widespread in caves around the Cent-Fonts. Clearly troglomorphic: eyeless, unpigmented, with elongated appendages and elongated claw.
- Onychiurus ortus Denis, 1935
Endemic from the sub-region, departments of Hérault, Gard and Aveyron. Widespread in caves around the Cent-Fonts. Clearly troglomorphic: eyeless, unpigmented and elongated claw.
3.2.6. Coleoptera
- Laemostenus (Actenipus) oblongus balmae (Delarouzée, 1860)
A widely distributed species, known from the Pyrenees to the southern and eastern edge of the Massif Central. The subspecies balmae is known from a few caves in the Gard and Hérault Departments, with one location in the Ardèche Department (Païolive).
3.3. Stygophilic taxa
Five stygophilic species of Cyclopids have been collected in the Cent Fonts according to Lescher-Moutoué [37]:
- Eucyclops serrulatus (Fisher, 1851), stygophile.
- Paracyclops fimbriatus (Fisher, 1853), stygophile.
- Acanthocyclops vernalis (Fisher, 1853), stygophile.
- Megacyclops viridis (Jurine, 1820), stygophile, in the hyporheic zone of the Hérault River nearby the Cent Fonts exurgences.
- Diacyclops languidoides Lilljeborg, 1901, stygophile.
3.4. Troglophilic taxa and parasites
Several other troglophilic taxa are expected to be found in the Cent Fonts system, of which the most important are listed below.
Ixodida
- Eschatocephalus vespertilionis (Koch, 1844), bat parasite, common.
Araneae
- Lessertia dentichelis (Simon, 1884), troglophile, very common in the caves throughout the Hérault valley.
- Meta bourneti Simon, 1922, troglophile, very common in all caves in the area.
- Meta menardi (Latreille, 1804), troglophile, very common in all caves in the area.
Opiliones
- Sabacon paradoxus Simon, 1879, troglophile, found in cave entrances in France and Spain. Very common in most caves in the Cévennes and in the Hérault karsts.
Julida
- Blaniulus guttulatus (Fabricius, 1798), troglophile, common in all caves of the region.
Isopoda
- Oritoniscus delmasi delmasi Vandel, 1933, endogeous and troglophilic species. Endemic to the southern Cévennes between the Vidourle and Lergue rivers.
- Phymatoniscus propinquus (Brian, 1908), troglophile. The ocular area of this species is generally provided with a large single eyespot but in specimens of the Cents Fonts cave, eyes are completely invisible to external examination [41]. The species is common throughout the Cévennes in the Ardèche, Gard and Hérault departments.
Coleoptera
- Leptinus testaceus P.W. Müller, 1817, is a troglophile, ectoparasite and commensal of many species of micromammals, lives mainly in subterranean mammal nests as well as in caves on bat guano. The species is sporadic but known from many caves around the Cent Fonts system.
4. Discussion
4.1. A biodiversity hotspot embedded in a stygobiont species-rich area
Only five stygobiont species were recorded from the Cent Fonts in the 1950s. Then Rouch et al. [6] carried out a more extensive survey and found 20 stygobiont species. About 50 years later, another survey was triggered by an impact study of an important water extraction project, resulted in a total of 43 stygobionts [12], making the Cent Fonts system the second richest hotspot of subterranean biodiversity in Europe (Table 1). Its stygobiont richness is higher than that of the better known Lez system (40 stygobionts), considered one of the most important biodiversity hotspots in the world [1,2]. The third rich area of the southern Massif Central in France is the Sauve karstic system (29 stygobionts), close to the Cent Fonts (Figure 1B). For the terrestrial fauna, it is expected that additional species will be found in the Cent Fonts, especially among the troglobionts known to occur in the surrounding caves (see 3.2.5), as the Cent Fonts cave has been quickly sampled for troglobionts.
As pointed out by Rouch et al. [6], the rich fauna observed in the Cent Fonts includes groundwater genera of undoubtedly freshwater origin, such as Elaphoidella, Ceuthonectes, Speocyclops, Graeteriella, for copepods and Gallocaris for the decapods, and genera of no less certain marine origin such as Microcharon and Sphaeromides for isopods, Ingolfiella for ingolfiellids and Salentinella for amphipods. Once again, the cave environment proves to be "the place of arrival of lineages of very different origins" [42]. In the stygobiotic molluscs, the origin of the family Moitessieriidae is still unclear, as all the published phylogenies have failed to anchor it in the global phylogenies of freshwater molluscs: the node linking it to the other taxa was not supported (e.g. [43]). This raises the question of the origin of this family, which could also be of marine origin.
4.2. Conservation issues and threats
The Cent Fons system is the second richest biodiversity hotspots in Europe for stygobiotic species and deserves conservation measures for this reason alone. A quarter of these 43 species are considered Vulnerable by the IUCN Red List. The Cent Fonts also hosts several relict species. Furthermore, it is the type locality of four taxa: Sphaeromicola cebennica juberthiei, Graeteriella vandeli, Microcharon doueti and Pseudectinosoma vandeli. Type localities should be preserved for future taxonomic work. Such a high biological value clearly deserves special attention.
Interestingly, eight species collected by Rouch et al. [6] (Sphaeromides raymondi, Sphaeromicola cebennica juberthiei, Acanthocyclops rhenanus, Graeteriella vandeli, Graeteriella unisetigera, Speocyclops racovitzai, Diacyclops languidoides and Soldanellonyx chappuisi were not collected in the 2006 inventory. This could be due to a lower probability of detection in 2006, but the sampling was more intensive, with water filtered for two years, a large team of experienced people, both in the field and for the identification of the taxa, the use of improved collection methods… So if not the probability of detection, the absence of these species in 2006 could be due to local extirpation. It cannot be ruled out that these species are indeed in decline, but the reason for this is unknown. Although relatively well preserved in terms of land use, the Cent Fonts hydrosystem may be under unknown threats.
Water pollution from the karst surface is likely to be low, as this karst has a very low human density. Climate change, which began in the early 1900s, could be a significant threat, but its effects on subterranean ecosystems are still poorly documented. However, severe droughts combined with increasing human pressure on the water resource especially in summer, with extreme fluctuations in water levels, are likely to affect subterranean ecosystems. Indeed, a short-term threat is the prospect of using this aquifer for drinking water. The Cent Fonts massif is recognised for the importance of its water supply and the quality of its water. An assessment of the volume of this resource and the possibilities for its exploitation was carried out in 2005 [13]. This study concluded that the drinking water reserve of the aquifer could not be mobilised for exploitation. However, this study can be disputed [44] and future needs may require greater resources.
4.2. Future prospects
Troglobionts have been under-sampled, and it is likely that many more species will be found in future surveys, as described above. Intensive surveys by Rouch et al. [6] and Olivier et al. [12] have allowed the collection of many stygobiotic species, and only a few are expected to be added. However, some of the species collected in 1968 were not collected again in 2006. This may be due to local extinction and/or bias in the probability of detection. Intensive and regular surveys would give us a clearer picture of the biodiversity of the Cent Fonts and allow us to document its evolution and threats. However, these surveys require significant investment and are unlikely to be undertaken in the near future to monitor the stygobitic fauna.
Environmental DNA is the topical, cost-effective answer to unsatisfactory detection probabilities and lack of taxonomic expertise. Several studies [45,46] have demonstrated its ability to detect up to 95% of aquatic organisms in surface streams, provided that optimised methods are implemented. Preliminary tests carried out in this karst with optimised methods (up to 250 liters filtered, 12 PCR replicates, coverage of 300 000…) were promising, allowing the detection of most, but not all, of the gastropod and crustacean species known to occur in the area. Extensive work on sampling methods is needed to improve the detection probability. This approach deserves to be explored further and is probably the future for surveying and monitoring the fascinating stygobiotic ecosystems.
Author Contributions
Conceptualization, V.P. and M.-J.D.O.; methodology, C.B. , V.P. and M.-J.D.O.; investigation, C.B. , V.P. and M.-J.D.O.; data curation, C.B. , V.P., C.A., P.M., D.M.P.G. and M.-J.D.O.; writing—original draft preparation, V.P.; writing—review and editing, V.P.; visualization, V.P. All authors have read and agreed to the published version of the manuscript.
Funding
The 2006 research was funded by Bureau de Recherches Geologiques et Minières (BRGM), service EAU, unite RMD.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data are available in the online open access database of the Inventaire National du Patrimoine Naturel (https://inpn.mnhn.fr/accueil/index); INPN locality # 2047774.
Acknowledgments
We would like to thank Danielle Defaye for checking the Copepods and Ostracods. Thanks to Louis Deharveng and Anne Bedos for sharing the bibliography and for constructive discussions.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
We present here a brief description of three new species of the genus Moitessieria. These new species are described in more detail in Prié (2013) in a Ph.D. thesis that is not considered an official publication according to the International Code of Zoological Nomenclature (ICZN). The names used in this earlier work are therefore nomina nuda. The descriptions are reproduced here with proposed new names, in line with the current trend to avoid eponyms when describing new species.
Moitessieria species are very rarely collected alive and when they are, they are difficult to preserve, because ethanol doesn’t penetrate into the shells - hence the paucity of available sequences on Genbank. A Moitessieria shell can have 7 to 8 whorls, but the animal will retract to the first three whorls when stressed. As the opening at the shell mouth is less than 1/4 mm wide, it is unlikely that the ethanol ever comes into contact with the flesh. This probably explains why, in most cases, DNA amplification fails from Moitessieria specimens, or only one or two genes amplify. The genetic data presented here are therefore incomplete.
The gastropod family Moitessieriidae is the only family composed entirely of stygobionts. Moitessieria rolandiana was considered to be a widespread species in southern France [47]. This wide distribution contrasts with that of other species in the family, which are often restricted to a small karstic area, due to the fragmentation of subterranean habitat. Prié [9] showed that M. rolandiana is actually composed of three cryptic species, each occupying a distinct karstic area, which supports their reproductive isolation by geographic barriers. They can be distinguished morphometrically, and molecular data corroborate their reproductive isolation. The species delimitation is based on morphometry, molecular and distribution data.
Material and methods
Material and methods are described in detail in Prié [9]. Biogeographic analyses are based on the SANDRE database [48], which describes the subsurface hydrogeological units. Morphometric analyses are based on the measurements of the shells, as shown in Figure 9. The three genes studied, COI, 16S and 28S were amplified using universal primers and conventional methods.
Biogeography
The region north of Montpellier in southern France is composed of distinct karst units, that have given rise to distinct faunal assemblages [8], with species delimitation based on obvious morphological characters). Not surprisingly, these distinct hydrosystems also suport distinct species. The Moitessieria populations studied belong to four adjacent basins. One, the Tarn basin in the west, flows into the Atlantic. The Atlantic and Mediterranean drainages are the most isolated, especially because no stygobiont gastropods are found in the upstream hydrosystems. On the Mediterranean side the Hérault (west), the Lez (centre) and the Vidourle (east) flow. Within these main basins, different hydrogeological units can be distinguished (Figure 10). While the surface relief creates ridge lines that distinguish these catchment areas, there may or may not be underground connections between the hydrogeological units. For example, the Larzac plateau flows north to the Vis River and south to the Lergue river, but the fauna is the same on both sides, reflecting the known subterranean connections between the two drainages. The same seems to happen between the upper Hérault and Vidourle drainages.
Morphometry
Both PCA and LDA multivariate analyses allowed the populations from the Lez source (type locality of Moitessieria rolandiana), from the Larzac plateau, and from the upper Hérault / Vidourle rivers to be distinguished. The populations from the upper Hérault and the upper Vidourle had the same morphology and could not be distinguished by morphometric analyses (Figure 11).
Genetics
Amplification was unsuccessful for several specimens or some of the genes studied, probably because Moitessieria species retract very deep into their shells, preventing contact between the tissues and the ethanol. The successful sequences of COI, 16S and 28S obtained are given in table 2. They support the biogeographic and morphometric analysis.

Table 2.
Results of the tentative amplification of the three genes for the live specimens of Moitessieria collected. Location according to Fig. 9.
Table 2.
Results of the tentative amplification of the three genes for the live specimens of Moitessieria collected. Location according to Fig. 9.
| Specimen | Location | COI | 16S | 28S |
|---|---|---|---|---|
| Moitessieria rolandiana | 4 1 | X | ||
| Moitessieria rolandiana | 4 1 | X | X | |
| Moitessieria larzacensis n. sp. | 1 | X | X | |
| Moitessieria larzacensis n. sp. | 6 | X | X | |
| Moitessieria atlantica n. sp. | 5 | X | ||
| Moitessieria atlantica n. sp. | 5 | X | ||
| Moitessieria atlantica n. sp. | 5 | X | X | |
| Moitessieria atlantica n. sp. | 5 | X | ||
| Moitessieria vidourlensis n. sp. | 3 | X | ||
| Moitessieria vidourlensis n. sp. | 2 | X | ||
| Moitessieria vidourlensis n. sp. | 2 | X |
1 Type locality of Moitessieria rolandiana.
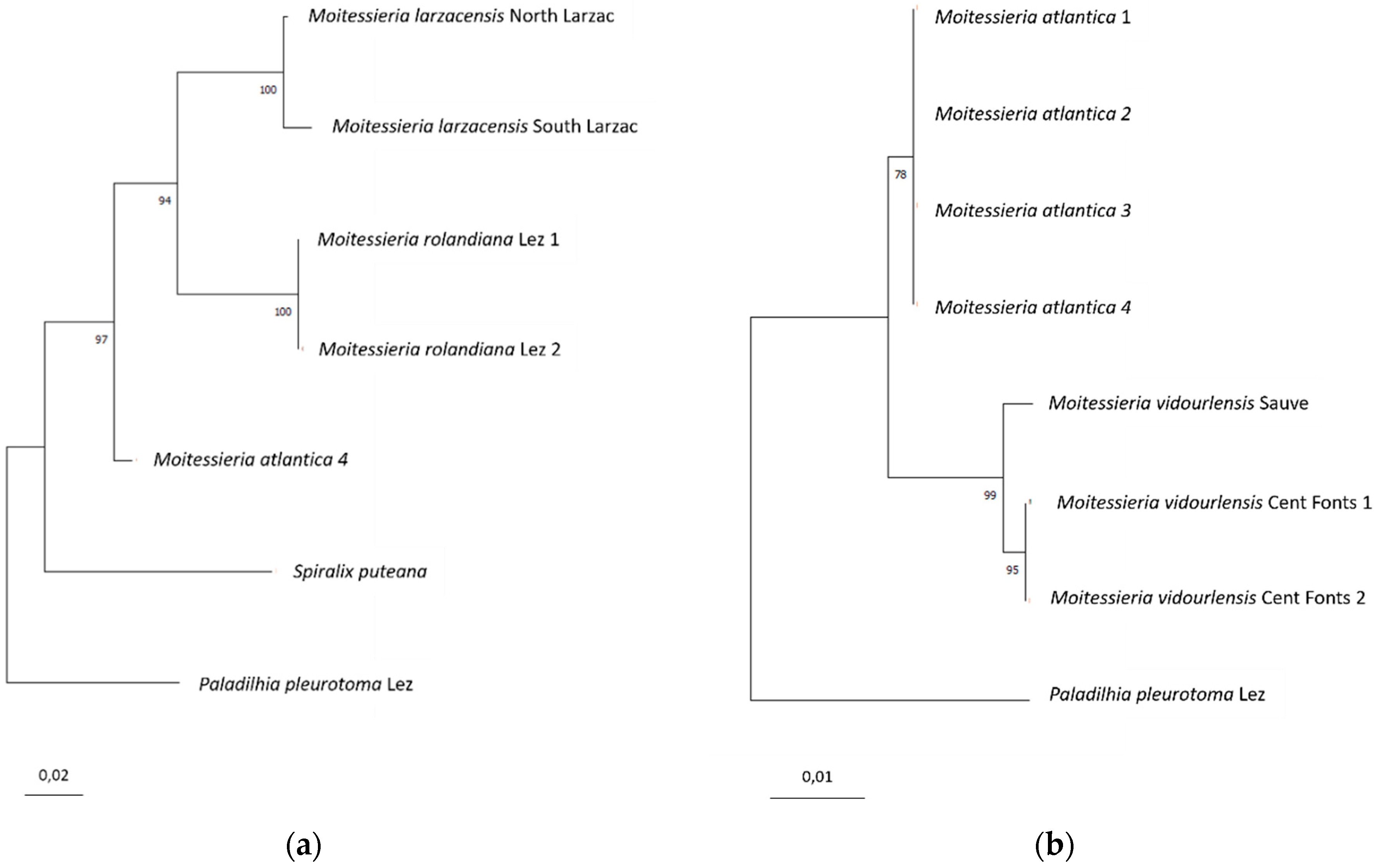
The mitochondrial genes (COI and 16S concatenated, Figure 12a) suggest that the population from the Larzac plateau is a separate species from Moitessieria rolandiana from the Lez drainage (type locality), as they diverge of over 10% in COI, a threshold largely over interspecific divergences in related taxa [49,50,51]. This corresponds to a divergence of 3.8 to 4.7% in the 16S gene. No COI was available to compare the Atlantic population to M. rolandiana, but the 16S results can be transposed, as all mitochondrial genes share the same history. The Atlantic population’s divergence to M. rolandiana type population is even higher, 6%. These results are congruent with the organisation of the hydrogeological networks.
The nuclear gene (28S, Figure 12b), although it should be less variable, distinguishes the Atlantic population from the Mediterranean ones (only the upper Hérault and Vidourle basins analysed here), but also, with a smaller divergence, the upper Hérault and upper Vidourle populations. This incongruence between the nuclear and mitochondrial genes analyses for the upper Hérault and upper Vidourle populations is not explained. Molecular data from more specimens are needed to state whereas the upper Vidourle and the upper Hérault populations constitute different species. I here adopt a conservative attitude and consider the upper Vidourle and the cent Fons populations as belonging to the same species.
In a nutshell, genetic data shows that:
- (i)
- M. larzacensis n. sp. differs from M. rolandiana based on both COI and 16S.
- (ii)
- M. atlantica n. sp. differs from M. rolandiana and M. larzacensis based on 16S.
- (iii)
- M. vidourlensis differs from M. atlantica based on 28S, but cannot be compared to M. rolandiana nor M. larzacensis from the available molecular data. Only morphological differences, that is supported by eographical isolation, allow to separate this species from M. rolandiana and M. larzacensis.
Figure 12.
Phylogenetic tree of the genes studied. (a) COI+16S concatenated; (b) 28S.
Species delimitation
Moitessieria rolandiana was considered to be a widespread species, distributed from the western tributaries of the Rhône River to the Garonne drainage [47]. Our results show that the name Moitessieria rolandiana should be restricted to the populations from the Lez drainage system. To the east, the adjacent Vidourle system hosts a distinct species, M. vidourlensis n. sp. The population sampled from the karst systems on the right bank of the Hérault River was morphologically and genetically similar to the Vidourle population and was therefore considered to belong to the same species. This distribution pattern involving two coastal river basins was unexpected. However, it is reminiscent of the distribution of some stygobiotic shrimps Gallocaris inermis, Proasellus cavaticus or Faucheria faucheri… a total of 12 crustacean species also known from both the Cent Fonts and Sauve (Vidourle) springs. It is therefore likely that hydrological connections exist, at least sporadically, in the complex karstic network of the upstream Hérault and Vidourle drainages. M. larzacensis n. sp. is likely to live in the subterranean basin of whole Vis River, probably on both sides, as Moitessieria species are known to live in the hyporheic zone and can therefore easily colonise the hydrosystems of both sides of the river. The hydrosystems of the Vis drainage are isolated from the Atlantic drainages to the west. It is therefore not surprising that the population of the Atlantic drainage belongs to a distinct species, here described as Moitessieria atlantica n. sp., due to geographic barriers.
Species turbo-taxonomy
Moitessieria vidourlensis n. sp. Prié 2023
Type material: holotype IM-2000-30145; paratypes: 28 shells (IM-2000-30146), deposited at the Museum national d’Histoire naturelle in Paris (Figure 13).
Type locality: the Sauve cave, in the Sauve (Gard department) municipality; 43° 56’ 27.2394’ N; 3° 56’ 58.1568” E. The live specimen was collected in scuba diving, on the ground, with forceps, a few tens of meters from the entrance of the cave.
Etymology: This species was initially dedicated to Frank Vasseur, an outstandingly skilled subterranean scuba diver, who collected material from inside caves for us (including M. larzacensis), and escorted me scuba diving into the Sauve cave, ensuring my safety while I was collecting live specimens of Moitessieria. I here prefer to avoid eponyms and give a name reflecting its distribution.
Distribution: pending further studies, the name Moitessieria vidourlensis should apply to the populations of Moitessieria from the Vidourle drainage, and the populations from the Causse-de-la-Selle. The species’ distribution probably includes part of the Hortus karstic plateau between the Hérault and Vidourle drainages. Its eastward distribution limit is unknown.
Distinctive characters: site 3 - Vidourle: shell height: 1.67 (1.47 – 1.98) mm; shell width: 0.75 (0.65 – 0.98) mm; last whorl width: 0.65 (0.60 – 0.73) mm; N=31; site 2 – Cent Fonts: shell height: 1.74 (1.57 – 2.08) mm; shell width: 0.73 (0.65 – 0.82) mm; last whorl width: 0.62 (0.57 – 0.69) mm; N=19.
Sequences GenBank accession numbers: XXX, XXX
Moitessieria larzacensis n. sp. Prié 2023
Type material: holotype IM-2000-30143; paratypes: 19 shells (IM-2000-30144), deposited at the Museum national d’Histoire naturelle in Paris (Figure 14).
Type locality: The live specimen used for DNA analyses was collected scuba-diving in the nearby Gourneyras cave, 43° 51’ 42.624 N; 3° 31’ 34.100 E.
Etymology: This species was initially dedicated to Annie Tillier, who successfully amplified the specimen collected from Gourneyras, at a time when Moitessieria specimen amplifications were systematically failing. We here prefer to avoid eponyms and give a name reflecting its distribution.
Distribution: known only from type locality but believed to be widespread in the hydrologic network of the Larzac plateau that drains into the Vis River. As Moitessieria species are known to occur in the hyporheic zone, the species may also occur in the Blandas plateau, left bank of the Vis River.
Distinctive characters: shell height: 1.87 (1.65 – 2.33) mm; width: 0.63 (0.55 – 0.74) mm; last whorl width: 0.56 (0.50 – 0.67) mm; N=20. Measurements come from another population, located a few hundred of meters upstream, as only a single specimen, alive, has been collected in the type locality.
Sequences GenBank accession numbers: XXX, XXX
Moitessieria atlantica n. sp. Prié 2023
Type material: holotype IM-2000-30147; paratypes: 11 shells (IM-2000-30148), deposited at the Museum national d’Histoire naturelle in Paris (Figure 15).
Type locality: The Gloriette spring, in the Sorgue drainage, municipality of Cornus (Aveyron department), 43° 54’ 28.5114” N; 3° 10’ 38.0634 E.
Etymology: this species was initially dedicated to Henri Girardi, a famous French malacologist, author of many subterranean snails’ descriptions. I here prefer to avoid eponyms and give a name reflecting its distribution.
Distribution: known only from the type locality, but presumably present elsewhere in the Sorgue River karstic drainages. Shells from a population sampled downstream (Saint-Paul-des-Fonts) fall within the morphological range of M. atlantica and could belong to the same species.
Interestingly, no subterranean snail has been collected despite important sampling in the vicinity of the limit between the Atlantic and Mediterranean watershed. M. atlantica is therefore geographically isolated from the other Moitessieria species described here.
Distinctive characters: shell height: 1.72 (1.58 – 1.93) mm; shell width: 0.68 (0.66 – 0.73) mm; last whorl width: 0.62 (0.60 – 0.67) mm; N=9. There is no biometric data on this population as the only specimens collected were used for DNA analyses (i.e. shells had to be crushed).
Sequences GenBank accession numbers: XXX, XXX
References
- Malard, F.; Gibert, J.; Laurent, R. L’aquifère de la source du Lez : un réservoir d’eau… et de biodiversité. Karstologia 1997, 30, 49–54. [Google Scholar] [CrossRef]
- Culver, D.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Cave Karst Stud. 2000, 62, 11–17. [Google Scholar]
- Tuzet, O.; Bonnet, A.; Bournier, E.; Du Cailar, J. Troisième contribution à la faune du Languedoc méditerranéen. Notes biospéologiques 1950, 5, 85–95. [Google Scholar]
- Balazuc, J.; Bonnet, A.; Bournier, E.; Du Cailar, J. Crustacés des eaux souterraines du Languedoc. Remarques sur leur répartition ? Bull. Soc. Hist. Nat. Toulouse 1951, 86, 80–87. [Google Scholar]
- Bonnet, A.; Bournier, E.; Du Cailar, J.; Quezel, P. Sur quatre crustacés aquatiques et troglobies d’une résurgence des gorges de l’Héralut. Soc. Mér. Spéléologie et Préhistoire 1951, 86, 341–346. [Google Scholar]
- Rouch, R.; Juberthie-Jupeau, L.; Juberthie, C. Recherche sur les eaux souterraines – 3 – Essai d’étude du peuplement de la zone noyée d’un karst. Ann. Spéléol. 1968, 23/4, 717–733. [Google Scholar]
- Prié, V. Répartition de Heraultiella exilis (Paladilhe, 1867) (Gastropoda, Caenogastropoda, Rissooidea). MalaCo 2005, 2005 1, 8–9. [Google Scholar]
- Prié, V. Les mollusques souterrains comme traceurs des hydrosystèmes : application aux hydrosystèmes karstiques des massifs nord-montpelliérains. Karstologia 2009, 52, 7–16. [Google Scholar] [CrossRef]
- Prié, V. 2013.- Taxonomie et biogéographie des mollusques patrimoniaux: quelles échelles pour la délimitation des taxons et des unités de gestion? Thèse de doctorat Muséum national d’Histoire naturelle à Paris.
- Girardi, H. Moitessieria wienini n. sp. des eaux de l’Aquifère de la Montagne de la Sellette, sur la rivière Hérault, (F. 34), (Mollusca : Gastéropoda : Moitessieriidae). Documents Malacologiques 2001, 2, 3–10. [Google Scholar]
- Girardi, H. Contribution à l’étude des gastéropodes stygobies de France. 3 – Paladilhia conica (PALADILHE, 1867) (Gastropoda : Moitessieriidae). Documents Malacologiques 2003, 4, 89–90. [Google Scholar]
- Olivier, M.-J.; Martin, D.; Bou, C.; Prié, V. Interprétation du suivi hydrobiologique de la faune stygobie, réalisé sur le système karstique des Cent Fonts lors du pompage d’essai. BRGM/RP+54865-FR. 2006, BRGM/RP+54865-FR, 42. [Google Scholar]
- Ladouche, B.; Maréchal, J.C.; Dörfliger, N.; Lachassagne, P.; Lanini, S.; Le Strat, P. Pompages d’essai sur le système karstique des Cent Fonts (Cne de Causse de la Selle, Hérault), Présentation et interprétation des données recueillies, BRGM/RP 54426-FR, 2005, 245 pp.
- Bou, C.; Rouch, R. Un nouveau champ de recherches sur la faune aquatique souterraine. C. R. Acad. Sci. Paris 1967, 265m, 369–370. [Google Scholar]
- Lecaplain, B. Sur la présence en France de Trocheta taunensis Grosser, 2015 (Hirudinida, Erpobdellidae). Naturae 2021, 25, 345–349. [Google Scholar] [CrossRef]
- Sket, B. K Poznavanju Favne Pijavk (Hirudinea) v Jugoslaviji, Zur Kenntnis der Egel-Fauna (Hirudinea) Jugoslawiens. Academia Scientiarum et Artium Slovenica Classis IV: Historia Naturalis et Medicina 1968, 9, 127–197. [Google Scholar]
- Grosser, C. Differentiation of some similar species of the subfamily Trochetinae (Hirudinida: Erpobdellidae). Ecol. Montenegrina 2015, 2(1), 29–41. [Google Scholar] [CrossRef]
- Prié, V. Prié, V. 2011. Heraultiella exilis. The IUCN Red List of Threatened Species 2011: e.T2092A9236035. DOI: 10.2305/IUCN.UK.2011-1.RLTS.T2092A9236035.en (accessed on 24 November 2023).
- UICN Comite français, OFB & MNHN (2021). La Liste rouge des espèces menacées en France – Chapitre Mollusques continentaux de France métropolitaine. Paris, France.
- Prié, V. Paladilhia pleurotoma. The IUCN Red List of Threatened Species 2010: e.T15876A5275513. DOI: 10.2305/IUCN.UK.2010-4.RLTS.T15876A5275513.en (accessed on 24 November 2023).
- Prié, V. Bythiospeum bourguignati. The IUCN Red List of Threatened Species 2010: e.T61315A12461687. DOI: 10.2305/IUCN.UK.2010-4.RLTS.T61315A12461687.en (Accessed on 24 November 2023). 2010. [Google Scholar]
- Prié, V. Systématique et micro-répartition des mollusques stygobies des karsts du Nord-Montpelliérain. Master de l’École Pratique des Hautes Études, École Pratique des hautes Études, 2006 Paris Sorbonne.
- Callot-Girardi, H.; Boeters, H. D. Moitessieria guilhemensis, nouvelle espèce de la résurgence du Cabrier à Saint-Guilhem-le Désert, Hérault, France. (Mollusca: Caenogastropoda: Moitessieriidae). Avenionia 2017, 2, 42–63. [Google Scholar]
- De Grave, S. Gallocaris inermis. The IUCN Red List of Threatened Species 2013: e.T198319A2520643 . 2013. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T198319A2520643.en (accessed on 01 December 2023).
- UICN France & MNHN (2012). La Liste rouge des espèces menacées en France - Chapitre Crustacés d’eau douce de France métropolitaine. Paris, France.
- Henry, J.-P. Contribution à l’étude du genre Proasellus (crustacea isopoda asellidae) : le groupe cavaticus. Vie et Milieu 1971, XXII, 33–77. [Google Scholar]
- Bertrand, J.-Y. Recherches sur l’écologie de Recherches sur l'écologie de Faucheria faucheri (Crustacés, Cirolanides). Thèse de 3° cycle, Université de Paris VI 1974, 123. [Google Scholar]
- Fiser, C.; Zagmajster, M.; Dethier, M. Overview of Niphargidae (Crustacea: Amphipoda) in Belgium: distribution, taxonomic notes and conservation issues. Zootaxa 2018, 4387, 26. [Google Scholar] [CrossRef] [PubMed]
- McInerney, C.E.; Maurice, L.; Robertson, A.L., Knight; Arnscheidt, J.; Venditti, C.; Dooley, J.; Mathers, T.C.; Matthijs, S.; Rrikkson, K.; Proulove, G.; Hänfling, B. The ancient Britons: groundwater fauna survived extreme climate change over tens of millions of years across NW Europe. Mol. Ecol. 2014, 23, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Stock, J. H.; Gledhill, T. The Niphargus kochianus-group in North-Western Europe. Crustaceana supplement 1977, 4, 212–243. [Google Scholar]
- Lefébure, T.; Douady, C.J.; Gouy, M.; Trontelj, P.; Briolay, J.; Gibert, J. Phylogeography of a subterranean amphipod reveals cryptic diversity and dynamic evolution in extreme environments. Mol. Ecol. 2006, 15, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Danielpol, D. L.; Namiotko, T.; Meisch, C. Marmocandona nov. gen. (Ostracoda, Candoninae), with comments on the contribution of stygobitic organisms to micropalaeontological studies. Kölner Forum Geol. Paläont 2012, 21, 13–16. [Google Scholar]
- Walter, T.C.; Boxshall, G.; World of Copepods Database. Acanthocyclops venustus venustus (Norman & Scott T., 1906). 2023, Accessed through: World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=729873 (accessed on 11 novembre 2023).
- Lescher-Moutoué, F. Recherches sur les eaux souterraines – 21 – Un Cyclopide nouveau du genre Graeteriella. Ann. Spéléol 1974, 71–76. [Google Scholar]
- Fiers, F.; Ghenne, V. Cryptozoic copepods from Belgium: diversity and biogeographic implications. Belg. J. of Zool. 2000, 130, 11–19. [Google Scholar]
- Lescher-Moutoué, F. Recherches sur les eaux souterraines – 7 - Les cyclopides de la zone noyée d'un karst. I Graeteriella (Paragraeteriella) vandeli n. sp. Ann. Spéléol 1969, 24, 429–438. [Google Scholar]
- Lescher-Moutoué, F. Sur la biologie et l'écologie des Copépodes Cyclopides hypogés. Ann. Spéléol 1973, 28, 429–502, 581–674. [Google Scholar]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Spatial distribution of stygobitic crustacean harpacticoids at the boundaries of groundwater habitat types in Europe. Sci Rep. 2020, 10, 19043. [Google Scholar] [CrossRef] [PubMed]
- Galassi, D.M.P.; Huys, R.; Reid, J.W. Diversity, ecology and evolution of groundwater copepods. Freshw. Biol 2009, 54, 691–708. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; De Laurentiis, P.; Dole-Olivier, M.-J. Phylogeny and biogeography of the genus Pseudectinosoma, and description of P. janineae sp. n. (Crustacea, Copepoda, Ectinosomatidae). Zool. Scr. 1999, 28, 289–303. [Google Scholar] [CrossRef]
- Vandel, A. Isopodes terrestres (Première partie). Office central de faunistique, Fédération française des Sociétés de Sciences naturelles. Paris, Lechevallier, Faune de France 64, 1960 pp. 417.
- Delamare Debouteville, C. Biologie des eaux souterraines littorales et continentales. Hermann 1960, 740. [Google Scholar]
- Wilke, T.; Davis, G.; Falniowski, A.; Giusti, F.; Bodon, M.; Szarowska, M. Molecular systematics of Hydrobiidae (Mollusca: Gastropoda: Rissooidea): Testing monophyly and phylogenetic relationships. Proc. Acad. Nat. Sci. USA 2009, 151, 1–21. [Google Scholar] [CrossRef]
- Machetel, P.; Yuen D., A. Revisiting Cent-Fonts Fluviokarst Hydrological Properties with Conservative Temperature Approximation. Hydrology 2017, 4, 6. [Google Scholar] [CrossRef]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; Gaboriaud, C.; Jean, P.; Poulet, N.; Roset, N.; Copp, G.H.; Geniez, P.; Pont, D.; Argillier, C.; Baudoin, J. M.; Peroux, T.; Crivelli, A.J.; Olivier, A.; Acqueberge, M.; Le Brun, M.; Møller, P.M.; Willerslev, E.; Dejean, T. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25(4), 929–942. [Google Scholar] [CrossRef] [PubMed]
- Prié, V.; Danet, A.; Valentini, A.; Lopes-Lima, M.; Taberlet, P.; Besnard, A.; Roset, N.; Gargominy, O.; Dejean, T. Conservation assessment based on large-scale monitoring of eDNA: application to freshwater mussels. Biol. Cons. 2023, 283, 110089. [Google Scholar] [CrossRef]
- Bertrand, A. Atlas préliminaire de répartition géographique des mollusques stygobies de la faune de France (MOLLUSCA : RISSOIDEA : CAENOGASTROPODA). Documents malacologiques 2015. [Google Scholar] [CrossRef]
- SANDRE. Base de données sur l’hydrographie. 2007. Available online: http://sandre.eaufrance.fr/ (accessed on 14 novembre 2023).
- Hershler, R.; Liu H., P.; Thompson, F., G. Phylogenetic relationships of North American nymphophiline gastropods based on mitochondrial DNA sequences. Zool. Scr. 2003, 32, 357–366. [Google Scholar] [CrossRef]
- Hurt, C. R. Genetic divergence, population structure and historical demography of rare spring-snails (Pyrgulopsis) in the lower Colorado River basin. Mol. Ecol. 2004, 13, 1173–1187. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.P.; Hershler, R.; Clift, K. Mitochondrial DNA sequences reveal extensive cryptic diversity within a western American springsnail. Mol. Ecol. 2003, 12, 2771–2782. [Google Scholar] [CrossRef]
Figure 1.
Location of the Cent Fonts springs. A: in Western Europe. B: biodiversity hotspots of the Montpellier region, numbers refer to the number of stygobiont species, 43 in the Cent Fonts, 40 in the Lez aquifer and 29 in the Sauve spring. C: the Causse de la Selle, aquifer of the Cent Fonts. Blue dots: springs; black line: cave topography. In bluish, the Causse-de-la-Selle plateau, which is the impluvium of the Cent Fonts system. To the north is the Buèges River, part of whose waters flow into the underground water system of the Cent Fonts (dotted blue line).
Figure 1.
Location of the Cent Fonts springs. A: in Western Europe. B: biodiversity hotspots of the Montpellier region, numbers refer to the number of stygobiont species, 43 in the Cent Fonts, 40 in the Lez aquifer and 29 in the Sauve spring. C: the Causse de la Selle, aquifer of the Cent Fonts. Blue dots: springs; black line: cave topography. In bluish, the Causse-de-la-Selle plateau, which is the impluvium of the Cent Fonts system. To the north is the Buèges River, part of whose waters flow into the underground water system of the Cent Fonts (dotted blue line).
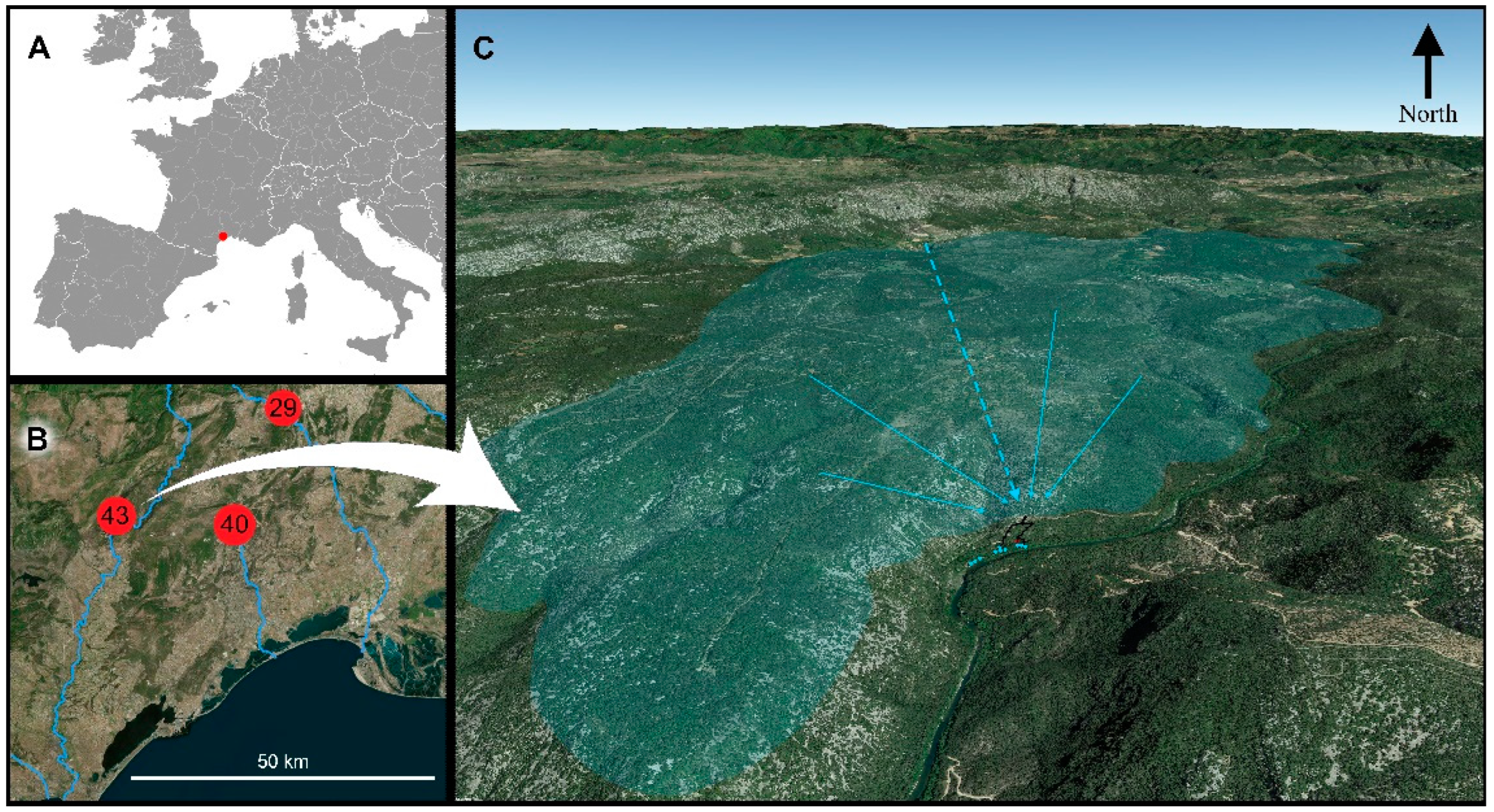
Figure 2.
Sampling the hyporheic zone of the Hérault River with the Bou-Rouch pump.

Figure 3.
(a) Bythinella sp.; (b) Paladilhia pleurotoma; (c) Moitessieria vidourlensis n. sp.; (d) Moitessieria guilhemensis ; (e) Moitessieria sp. nov.? or a monstrous form of Moitessieria sp. All specimens collected in the Cent Fonts sources.
Figure 3.
(a) Bythinella sp.; (b) Paladilhia pleurotoma; (c) Moitessieria vidourlensis n. sp.; (d) Moitessieria guilhemensis ; (e) Moitessieria sp. nov.? or a monstrous form of Moitessieria sp. All specimens collected in the Cent Fonts sources.

Figure 4.
Gallocaris inermis Source of Sauve (Vidourle), ≈ 12 mm. © C. Alonso.

Figure 5.
Sphaeromides raymondi, Grotte exsurgence de l'Avencas, Brissac, ≈ 18 mm. © J.C Queneau.

Figure 6.
Niphargus cf. virei, Grotte exsurgence de l'Avencas, Brissac, ≈ 15 mm. © J.C Queneau.

Figure 7.
Ingolfiella thibaudi, Cent Fonts, ≈ 2 mm. © M.-J. Dole-Olivier.

Figure 8.
Some troglobite taxa of the Cent Fonts kast system. (A) Opiliones Peltonychia clavigera; (B) Pseudoscorpiones Neobisium tuzetae; (C) Isopoda Trichoniscoides bonneti; and (D) Diplura Plusiocampa balsani. © C. Alonso.
Figure 8.
Some troglobite taxa of the Cent Fonts kast system. (A) Opiliones Peltonychia clavigera; (B) Pseudoscorpiones Neobisium tuzetae; (C) Isopoda Trichoniscoides bonneti; and (D) Diplura Plusiocampa balsani. © C. Alonso.

Figure 9.
Measurements of a Moitessieria shell. H: height of the shell; HdT: height of the last whorl; Ho: height of the aperture; Lo: width of the aperture; LdS: width of the suture of the last whorl; LdT: width of the last whorl; L width of the shell.
Figure 9.
Measurements of a Moitessieria shell. H: height of the shell; HdT: height of the last whorl; Ho: height of the aperture; Lo: width of the aperture; LdS: width of the suture of the last whorl; LdT: width of the last whorl; L width of the shell.
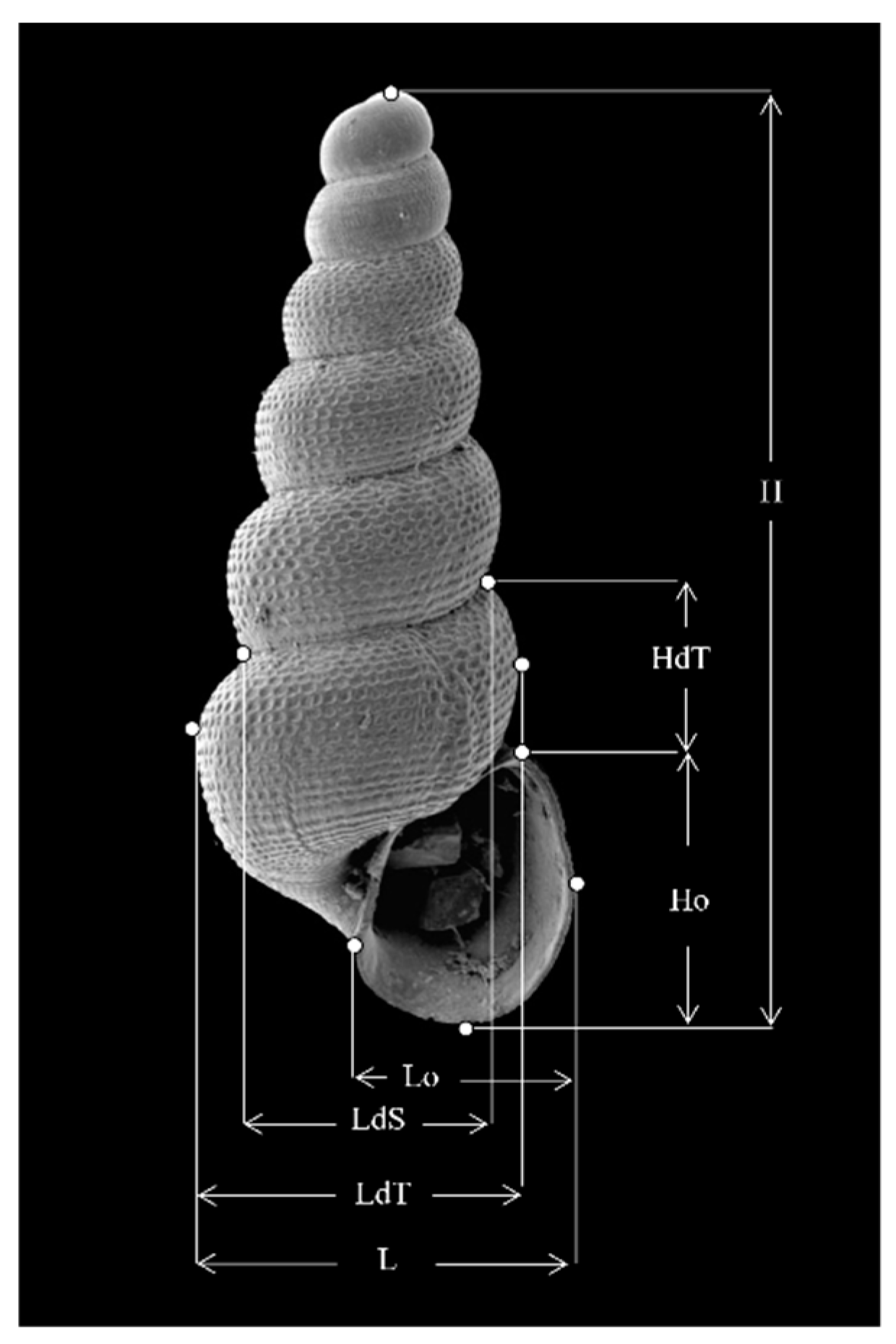
Figure 10.
Biogeography of the subregion. Bold black line: separation between the Atlantic and Mediterranean drainages; thin dotted lines: separation between the major river drainages; blue lines: rivers; red dots: sampled populations, with numbers referring to the populations for which morphometric analysis was performed. The Mediterranean rivers basins are highlighted in grey. The hydrogeological units are based on the SANDRE database. The numbers refer to the locations of the populations for which morphometric and/or molecular analyses were carried out (Figure 11 and Figure 12).
Figure 10.
Biogeography of the subregion. Bold black line: separation between the Atlantic and Mediterranean drainages; thin dotted lines: separation between the major river drainages; blue lines: rivers; red dots: sampled populations, with numbers referring to the populations for which morphometric analysis was performed. The Mediterranean rivers basins are highlighted in grey. The hydrogeological units are based on the SANDRE database. The numbers refer to the locations of the populations for which morphometric and/or molecular analyses were carried out (Figure 11 and Figure 12).
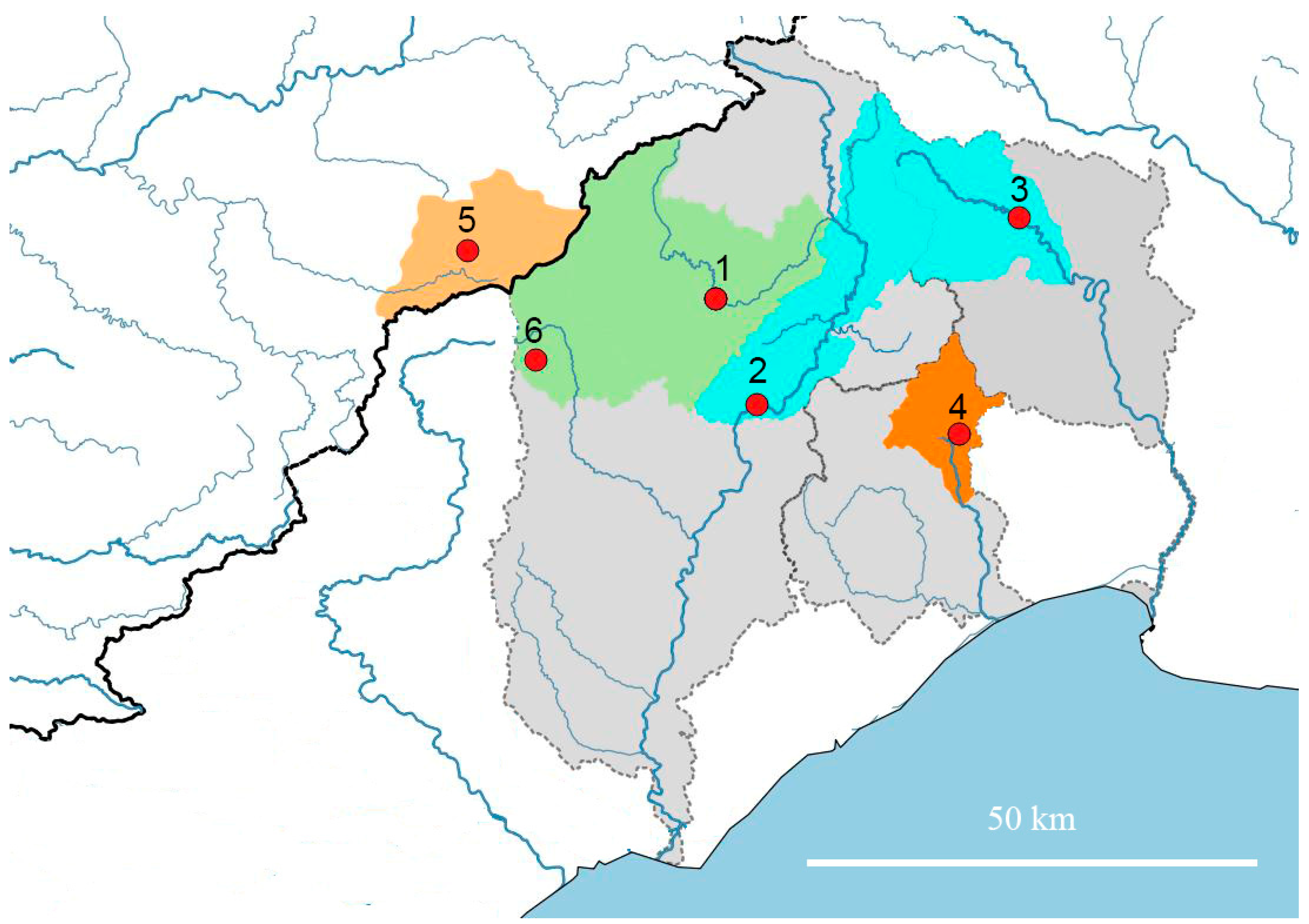
Figure 11.
Multivariate analyses of the morphometrics of Moitessieria populations: (a) Principal Component Analysis; (b) Linear Discriminant Analysis. 1: Vis River, Larzac plateau, Hérault basin; 2: Cabrier source, same system as the Cent Fonts, Hérault basin; 3: Sauve source, upper Vidourle; 4: Lez source, type locality of Moitessieria rolandiana (see Figure 9).
Figure 11.
Multivariate analyses of the morphometrics of Moitessieria populations: (a) Principal Component Analysis; (b) Linear Discriminant Analysis. 1: Vis River, Larzac plateau, Hérault basin; 2: Cabrier source, same system as the Cent Fonts, Hérault basin; 3: Sauve source, upper Vidourle; 4: Lez source, type locality of Moitessieria rolandiana (see Figure 9).
Figure 13.
Moitessieria vidourlensis: (a) General view of the holotype (left) and of a paratype (right), scale = 500µm; (b) details of the protoconch of the holotype (above) and of a paratype (below), scale = 50 µm.
Figure 13.
Moitessieria vidourlensis: (a) General view of the holotype (left) and of a paratype (right), scale = 500µm; (b) details of the protoconch of the holotype (above) and of a paratype (below), scale = 50 µm.

Figure 14.
Moitessieria larzacensis: (a) General view of the holotype (left) and of a paratype (right), scale = 500µm; (b) details of the protoconch of the holotype (above) and of a paratype (below), scale = 50 µm.
Figure 14.
Moitessieria larzacensis: (a) General view of the holotype (left) and of a paratype (right), scale = 500µm; (b) details of the protoconch of the holotype (above) and of a paratype (below), scale = 50 µm.

Figure 15.
Moitessieria atlantica: (a) General view of the holotype (left) and of a paratype (right), scale = 500µm; (b) details of the protoconch of the holotype (above) and of a paratype (below), scale = 50 µm.
Figure 15.
Moitessieria atlantica: (a) General view of the holotype (left) and of a paratype (right), scale = 500µm; (b) details of the protoconch of the holotype (above) and of a paratype (below), scale = 50 µm.

Table 1.
List of the species recorded in the Cent Fonts system from 1950 to 2006 (NB: all stygobiont were recorded in the Cent Fonts system itself; most troglobionts were recorded only in surrounding caves).
Table 1.
List of the species recorded in the Cent Fonts system from 1950 to 2006 (NB: all stygobiont were recorded in the Cent Fonts system itself; most troglobionts were recorded only in surrounding caves).
| Classe | Sous-classe | Ordre | Balazuc, Bonnet etc. 1950 | Rouch et al. 1968 | Olivier et al. 2006 | This paper | IUCN France | IUCN Global | |
|---|---|---|---|---|---|---|---|---|---|
| Clitellata | Hirudinea | Arhynchobdellida | Stygobiotic | ? | ? | - | Trocheta taunensis Grosser, 2015 | NE | NE |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Bythinella n. sp. | Bythinella sp. | NA | NA |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Heraultiella exilis | Heraultiella exilis(Paladilhe, 1867) | VU | VU |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | lslamia moquiniana | lslamia cf. moquiniana | NA | NA |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Paladilhia pleurotoma | Paladilhia pleurotoma Bourguignat, 1865 | VU | LC |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Bythiospeum bourguignati | Bythiospeum bourguignati (Paladilhe, 1866) | NT | LC |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Moitessieria rolandiana | Moitessieria vidourlensis | NE | NE |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Moitessieria n.sp.1 | Moitessieria guilhemensis Girardi & Boeters, 2017 | NE | NE |
| Gastropoda | Caenogastropoda | Littorinimorpha | Stygobiotic | - | - | Moitessieria n.sp.2 ? | Moitessieria sp. | NA | NA |
| Malacostraca | Eumalacostraca | Decapoda | Stygobiotic | Troglocaris inermis | Troglocaris inermis | Troglocaris inermis | Gallocaris (Troglocaris) inermis (Fage, 1937) | VU | NT |
| Malacostraca | Eumalacostraca | Isopoda | Stygobiotic | - | Stenasellus buili | Stenasellus buili | Stenasellus buili Rémy, 1949 | NT | NE |
| Malacostraca | Eumalacostraca | Isopoda | Stygobiotic | - | Proasellus cavaticus cavaticus | Proasellus cavaticus | Proasellus cavaticus (Leydig, 1871) | LC | NE |
| Malacostraca | Eumalacostraca | Isopoda | Stygobiotic | - | Microcharon douetin. sp. | Microcharon doueti | Microcharon doueti Coineau, 1968 | VU | NE |
| Malacostraca | Eumalacostraca | Isopoda | Stygobiotic | Faucheria faucheri | Faucheria faucheri | Faucheria faucheri | Faucheria faucheri (Dollfus & Viré, 1900) | LC | NE |
| Malacostraca | Eumalacostraca | Isopoda | Stygobiotic | Sphaeromides raymondi | Sphaeromides raymondi | - | Sphaeromides raymondi Dollfus, 1897 | NT | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | - | - | Niphargus laisi | Niphargus laisi Schellenberg, 1936 | DD | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | - | - | Niphargus gallicus | Niphargus gallicus Schellenberg, 1935 | LC | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | - | - | Niphargus kochianus | Niphargus kochianus Bate, 1859 | LC | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | - | - | Niphargus pachypus | Niphargus pachypus Schellenberg, 1933 | LC | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | Niphargus orcinus virei | Niphargus orcinus virei | Niphargus virei | Niphargus cf. virei Chevreux, 1896 clade A | LC | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | - | Salentinella sp. | Salentinella angelieri | Salentinella angelieri Delamare-Deboutteville & Ruffo, 1952 | NT | NE |
| Malacostraca | Eumalacostraca | Amphipoda | Stygobiotic | - | Salentinella delamarei | Salentinella delamarei Coineau, 1962 | LC | NE | |
| Malacostraca | Eumalacostraca | Ingolfiellida | Stygobiotic | - | Ingofiella sp. | Ingolfiella thibaudi | Ingolfiella thibaudi Coineau, 1968 | LC | NE |
| Ostracoda | Podocopa | Podocopida | Stygobiotic | - | - | Fabaeformiscandona cf. breuili | Fabaeformiscandona cf. breuili (Paris, 1920) | LC | NE |
| Ostracoda | Podocopa | Podocopida | Stygobiotic | - | - | Pseudocandona zschokkei | Marmocandona (Pseudocandona) cf. zschokkei (Wolf, 1920) | LC | NE |
| Ostracoda | Podocopa | Podocopida | Stygobiotic | - | - | Schellencandona cf. simililampadis | Schellencandona cf. simililampadis (Danielpol, 1978) | VU | NE |
| Ostracoda | Podocopa | Podocopida | Stygobiotic | Sphaeromicola cebennica Remy | Sphaeromicola cebennica Remy | - | Sphaeromicola cebennica juberthiei Danielpol, 1977 | VU | NE |
| Ostracoda | Podocopa | Podocopida | Stygobiotic | Candoninae sp. 1, long, related to Cryptocandona | Candoninae sp. 1, cf. Cryptocandona | NA | NA | ||
| Ostracoda | Podocopa | Podocopida | Stygobiotic | Candoninae sp. 2, bean-shaped, related to Pseudocandona | Candoninae sp. 2, cf. Pseudocandona | NA | NA | ||
| Ostracoda | Podocopa | Podocopida | Stygobiotic | Candoninae sp. 3, triangular, related to Pseudocandona group eremita | Candoninae sp. 3, cf. Pseudocandona group eremita | NA | NA | ||
| Malacostraca | Eumalacostraca | Bathynellacea | Stygobiotic | - | - | Clamousella cf. delayi | Gallobathynella delayi Serban, Coineau & Delamare Deboutteville | VU | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | Acanthocyclops rhenanus | - | Acanthocyclops rhenanus Kiefer, 1936 | LC | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | Acanthocyclops stammeri westfalicus | Acanthocyclops venustus westfalicus | Acanthocyclops venustus westfalicus (Kiefer, 1931) | VU | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | - | Graeteriella (Graeteriella) cf. boui | Graeteriella (Graeteriella) boui Lescher-Moutoué, 1969 | VU | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | Graeteriella unisetiger | - | Graeteriella (Graeteriella) unisetigera Graeter, 1910 | LC | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | Paragraeteriella n. sp. | - | Graeteriella (Paragraeteriella) vandeli Lescher-Moutoué, 1969 | VU | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | - | Kieferiella delamarei | Kieferiella delamarei (Lescher-Moutoué, 1971) | VU | NE |
| Copepoda | Neocopepoda | Cyclopoida | Stygobiotic | - | Speleocyclops sp. | - | Speocyclops racovitzai Chappuis, 1923 | LC | NE |
| Copepoda | Neocopepoda | Harpacticoida | Stygobiotic | - | Ceuthonectes gallicus | Ceuthonectes gallicus | Ceuthonectes gallicus Chappuis, 1928 | LC | NE |
| Copepoda | Neocopepoda | Harpacticoida | Stygobiotic | - | Elaphoidella leruthi meridionalis | Elaphoidella leruthi meridionalis | Elaphoidella leruthi meridionalis Chappuis, 1953 | DD | NE |
| Copepoda | Neocopepoda | Harpacticoida | Stygobiotic | - | - | Nitocrella omega | Nitocrella omega Hertzog, 1936 | VU | NE |
| Copepoda | Neocopepoda | Harpacticoida | Stygobiotic | - | Nitocrella sp. groupe hirta | Nitocrella hirta hirta | Nitocrella hirta Chappuis, 1924 | NE | NE |
| Copepoda | Neocopepoda | Harpacticoida | Stygobiotic | - | Ectinosomidae sp. | Pseudectinosoma vandeli | Pseudectinosoma vandeli (Rouch, 1969) | VU | NE |
| Arachnida | Acari | Trombidiformes | Stygobiotic | - | Soldanellonyx chappuisi | - | Soldanellonyx chappuisi Walter, 1917 | NE | NE |
| Arachnida | Micrura | Araneae | Troglobiotic | Palliduphantes sanctivincenti (Simon, 1873) | NE | NE | |||
| Arachnida | Dromopoda | Opiliones | Troglobiotic | Peltonychia clavigera (Simon, 1872) | NE | NE | |||
| Arachnida | Dromopoda | Pseudoscorpiones | Troglobiotic | Neobisium tuzetae Vachon, 1947 | NE | NE | |||
| Malacostraca | Eumalacostraca | Isopoda | Troglobiotic | Trichoniscoides bonneti Vandel, 1946 | NE | NE | |||
| Entognatha | - | Diplura | Troglobiotic | Plusiocampa balsani Conde, 1947 | NE | NE | |||
| Entognatha | Elliplura | Colembola | Troglobiotic | Pseudosinella denisi Gisin, 1954 | NE | NE | |||
| Entognatha | Elliplura | Colembola | Troglobiotic | Onychiurus ortus Denis, 1935 | NE | NE | |||
| Insecta | Pterygota | Coleoptera | Troglobiotic | Laemostenus (Actenipus) oblongus balmae (Delarouzée, 1860) | NE | NE | |||
| Total Stygobiotic | 43 | ||||||||
| Total Troglobiotic | 5 | 21 | 36 | 8 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



