
Submitted:
12 December 2023
Posted:
12 December 2023
You are already at the latest version
Abstract
Stroke remains one of the leading causes of death and disability worldwide. Current reperfusion treatments for ischaemic stroke are limited due to their narrow therapeutic window in rescuing ischaemic penumbra. Stem cell therapy offers a promising alternative. As a regenerative medicine, stem cells offer a wider range of treatment strategies, including long-term intervention for chronic patients, through the reparation and replacement of injured cells via mechanisms of differentiation and proliferation. The purpose of this review is to evaluate the therapeutic role of stem cells for ischaemic stroke. The paper discusses the pathology during acute, subacute, and chronic phases of cerebral ischaemic injury, highlights the mechanisms involved in mesenchymal, endothelial, haematopoietic, and neural stem cell-mediated cerebrovascular regeneration and evaluates the pre-clinical and clinical data concerning the safety and efficacy of stem cell-based treatments. Treatment of stroke patients with different type of stem cells appears to be safe and efficacious even at relatively higher concentrations irrespective of the route and timing of administration. Priming or preconditioning of cells prior to administration appears to help augment their therapeutic impact. However, larger patient cohorts and later phase trials are required to consolidate these findings.
Keywords:
stem cells
; cell-based therapy
; endothelial progenitor cells
; ischaemic stroke
1. Introduction
Stroke continues to be one of the leading causes of mortality and morbidity in the world, with around thirty-two thousand stroke-related deaths in England alone each year [1]. As the population ages, the prevalence of stroke-related death and disability will continue to rise, presenting a substantial public health burden. Stroke occurs when cerebral blood supply is disrupted as a result of an occlusion (ischaemic strokes) or rupture (haemorrhagic strokes) of an artery leading to, within or on the surface of the brain. The brain is particularly susceptible to damage when it is starved of oxygen and glucose even for shorter periods of time, leading to sudden appearance of contralateral hemiparesis, speech difficulties, confusion, visual disturbances, balance problems and a severe headache. In chronic phases, the patients manifest persistent aphasia, amnesia and problems with emotional functioning. This marked reduction in the quality of life is a catalyst for stroke research community to discover new agents or interventions for stroke.
While a range of effective prophylactic medicines exist, including aspirin, clopidogrel, warfarin and other anticoagulants, the current curative therapeutic options are restricted to thrombolysis, thrombectomy or bridging treatment. Thrombolysis is realised by intravenous (IV) administration of recombinant tissue-plasminogen activator (rt-PA) to eligible patients to restore cerebral blood flow. Though proven to be safe and effective in improving clinical outcomes at three months [2], the therapeutic window for thrombolysis is limited. To minimise the damage to ischaemic penumbra, rt-PA must be administered within the first 4.5 h of an ischaemic stroke [3]. Beyond this therapeutic window, intravenous thrombolysis (IVT) may further compromise the integrity of the blood-brain barrier (BBB), consequently giving rise to symptomatic intracerebral haemorrhage [4]. Endovascular thrombectomy (EVT) is an invasive procedure which involves the insertion of a catheter into an artery to surgically remove thrombus for recanalization. Beneficial effect of EVT were determined in patients with acute ischaemic stroke who received treatment 6 to 24 hours after they had last been known to be well [5]. In addition to the narrow time window, patients with most types active haemorrhage are not eligible to both IVT, EVT or other anticoagulatory treatments [6].
In this regard, stem cells, with their self-renewing capabilities and capacity to differentiate and repair damaged tissue, present an exciting alternative. Instead of targeting the cause of stroke, stem cells act to reverse or remedy the pathological damage caused by ischaemic damage. By presenting a therapeutic option beyond the acute phase of stroke, stem cells may be of significant value to extend the interventional strategies to all patient profiles. This review examines a selection of different stem cell types and their therapeutic relevance in ischaemic stroke. The niches discussed in this paper include mesenchymal stem cells (MSCs), endothelial progenitor cells (EPCs), haematopoietic stem cells (HSCs) and neural stem cells (NSCs).
2. Pathology of Ischaemic Stroke
It is essential to comprehend different mechanisms and networks involved in the pathogenesis and outcome of stroke to appreciate the therapeutic use and value of stem cells. The mechanisms discussed below occur in many different cell types ranging from nervous tissue, involved in sensory and motor communications, to cells of the BBB, responsible for protecting the CNS.
2.1. Excitotoxic Cell Death
Hypoxia that occurs immediate aftermath of an ischaemic attack triggers excitotoxic cell death. Hypoxic conditions downregulate ATP production by inhibiting plasma membrane Na+/K+/ATPase and Ca2+/ATPase pumps [7,8]. Receptor malfunction increases intracellular Na+ and Ca2+, causing cellular depolarisation and the propagation of action potentials. Na+ influx results in K+ efflux, further stimulating peri-infarct depolarisation. High levels of intracellular Ca2+ triggers glutamate exocytosis into the synaptic cleft, this accretion stimulates postsynaptic glutamate receptors, further increasing intracellular Ca2+ in the postsynaptic neurone [9,10]. Excessive Ca2+ load results in mitochondrial dysfunction, stimulating proteolysis and NADPH oxidase enzyme induction, triggering oxidative stress accompanied by excessive release of reactive oxygen species (ROS). Once generated, ROS promote inflammatory mechanisms by attracting cytokines and leukocytes infiltrate the brain as the BBB degrades [11,12,13,14]. Microglial cells, which are activated under oxidative stress, along with cytokines, also recruit matrix metalloproteinases (MMPs), a family of protease enzymes, further aiding local inflammation of ischaemic tissue [15]. Both activated microglia and reactive astrocytes are major components of the immune system in the brain and the crosstalk between them reinforces the release of several proinflammatory factors, including IL-1β, IL-6, TNF-α, IL-15 and MMPs [16,17]. This homeostatic upset, inflammation and uncontrolled enzymatic degradation inevitably damages the cellular structure and function and adversely affects the surrounding microenvironment.
2.2. Apoptosis, Necrosis and Necroptosis Pathways
Lack of Ca2+ homeostasis also stimulates numerous cellular death pathways. Ischaemia induces apoptosis via the release of cytochrome C from dysfunctional mitochondria followed by the activation of caspase-3 and the downstream hydrolases [18]. The cell enters the execution phase of apoptosis, cytoplasm begins to shrink and display cytomorphological changes, including nuclear condensation [19]. Alternatively, the cell may undergo necrosis. This is often described as premature cell death and occurs due to Na+ influx accompanying Na+/K+ pump and Ca2+/ATP-pump failure. Intracellular Na+ and Ca2+ aggregation leads to cellular oedema, swelling and loss of lysosomal membrane integrity and cell rupture. Exposed cellular components attract digestive molecules for cell lysis, further contributing to local inflammation. It is noteworthy that unlike apoptosis, necrosis is independent of caspase activity [20,21]. Another recognised route of cell death in stroke is necroptosis, otherwise known as ‘programmed necrosis’. Though currently unidentified mechanisms, necroptosis appears to be regulated by receptor-interacting protein kinase 1 (RIPK1) and involves swelling of the cell and lysosomal rupture (mimicking necrosis) and similar to apoptosis requires caspase and cytochrome C activity [22].
Autophagy or auto-phagocytosis is another degradation pathway often witnessed in ischaemic stroke. Autophagy is the breakdown and phagocytosis of cellular organelles to maintain homeostasis [15,23]. Its catabolic effects allow for the emergency production of energy and nutrients during hypoxia and other stressful conditions. Though autophagy is generally recognised as a protective pathway, evidence linking it to neurodegeneration also exists [24].
3. Blood-Brain Barrier
The BBB, an integral component of the neurovascular unit, regulates the selective passage of compounds between the blood and the brain parenchyma [15,25]. The BBB consists of pericytes, astrocytes and endothelial cells (ECs) and is paracellularly sealed by tight junctions (TJs). These protein complexes are primarily composed of the transmembrane proteins claudins, occludins, junction adhesion molecules (JAMs) and zone occludens (ZO), an accessory protein responsible for manoeuvring cytoskeletal interactions [26]. The claudin family demonstrate a variety of transmembrane domains, of which the claudin-5 isoform is most greatly expressed, showing direct responsibility in tightening the BBB against small molecules (<800Da) [26,27]. Occludins form dimers and oligomers, aiding paracellular permeability and stabilising barrier function to which JAMs provide further support. The degradation of these tight junction constituents, catalysed by activated MMPs, compromises the BBB. Hypoxia and exaggerated local cytokine availability augment MMP expression, with elevated MMP-2 and MMP-9 levels identified in stroke patients [15,28,29,30,31,32]. Restoration of BBB integrity during post-ischaemic period by MMP inhibition highlights this relationship [33,34] Furthermore, the autophagy of claudin-5 is associated with significant increases in solute permeability of the BBB [15,30]. Decreased expression of ZO-1, occludin and claudin-5 in senescent ECs leads to the impairment of BBB integrity [35]. The disruption of other TJ factors such as the JAM VE-cadherin, resulted in BBB compromise [27]. Furthermore, the pathology of pericytes which mature and maintain the BBB and astrocytes, involved in the maintenance of osmotic BBB conditions, further precipitate BBB breakdown [36].
Destruction of the basement membrane is another pathology to consider in ischaemic stroke. Basement membrane is a non-cellular complex consisting of a sheath of extracellular matrix and a series of proteins, namely collagen IV, nidogen, perlecan, agrin and laminin. Although how basement membrane gets damaged during ischaemic stroke remains vague, it is presumed that the membrane undergoes dissolution thereby exacerbating loss of BBB and vascular integrity [37,38,39].
4. Stem Cell as Therapeutics
A literature search using the key MeSH terms “stem cells”, “ischaemic stroke”, “stroke pathology”, “mesenchymal stem cells” (MSCs), “endothelial progenitor cells” (EPCs), “haematopoietic stem cells” (HSCs), and “neural stem cells” (NSCs) on the PubMed database identified relevant studies. Nottingham University search and Google Scholar were also used to collect pertinent studies. This paper has critically evaluated a variety of pre-clinical and clinical studies to ascertain the role of stem cells as therapeutics for stroke.
Figure 1.
Mechanisms of stem cell therapy for ischaemic stroke. Excitotoxic cell damages, activation of immune cells, inflammatory reaction, breakdown of blood-brain barrier, mitochondrial dysfunction and oxidative stress are involved in the pathophysiology of stroke. Stem cell therapy have the potential to ameliorate these processes via differentiation into various cells to replace the damaged cells, secrete cytokines and growth factors to promote angiogenesis, neurogenesis and immunomodulation. HGF, hepatocyte growth factor; BDNF, brain-derived neurotrophic factor; VEGF, vascular endothelial growth factor; PDGF, Platelet-derived growth factor; GDNF, glial cell-derived neurotrophic factor; IGF-1, insulin-like growth factor 1; TGF-β1, transforming growth factor beta-1; GM-CFS, granulocyte-macrophage colony-stimulating factor; G-CSF, granulocyte colony stimulating factor; PBEF, pre-B cell-enhancing factor; ECs, endothelial cells; SMCs, smooth muscle cells; NGF, nerve growth factor; IL, interleukin; TNF-α, tumor necrosis factor alpha; MMPs, matrix metalloproteinases; ROS, reactive oxygen species.
Figure 1.
Mechanisms of stem cell therapy for ischaemic stroke. Excitotoxic cell damages, activation of immune cells, inflammatory reaction, breakdown of blood-brain barrier, mitochondrial dysfunction and oxidative stress are involved in the pathophysiology of stroke. Stem cell therapy have the potential to ameliorate these processes via differentiation into various cells to replace the damaged cells, secrete cytokines and growth factors to promote angiogenesis, neurogenesis and immunomodulation. HGF, hepatocyte growth factor; BDNF, brain-derived neurotrophic factor; VEGF, vascular endothelial growth factor; PDGF, Platelet-derived growth factor; GDNF, glial cell-derived neurotrophic factor; IGF-1, insulin-like growth factor 1; TGF-β1, transforming growth factor beta-1; GM-CFS, granulocyte-macrophage colony-stimulating factor; G-CSF, granulocyte colony stimulating factor; PBEF, pre-B cell-enhancing factor; ECs, endothelial cells; SMCs, smooth muscle cells; NGF, nerve growth factor; IL, interleukin; TNF-α, tumor necrosis factor alpha; MMPs, matrix metalloproteinases; ROS, reactive oxygen species.
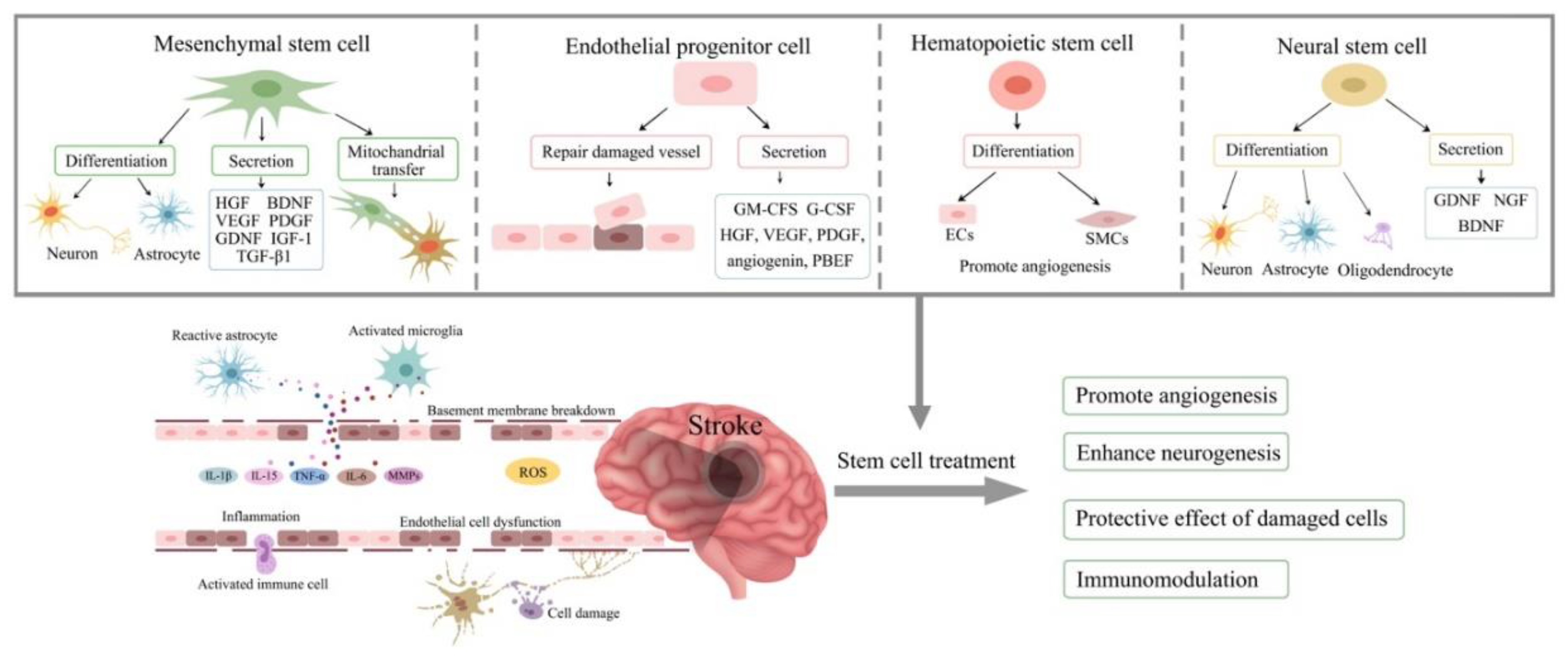
5. Mesenchymal Stem Cells
MSCs are multipotent adult stem cells with the ability to differentiate into various cell types within mesodermal lineage including bone cells, cartilage, muscle cells and skin cells. Despite their limited capacity to differentiate, evidence exist regarding trans-differentiation along ectodermal lineage into neural cells and along endodermal lineage into hepatocytes [40]. MSCs are isolated from bone marrow (BM), adipose tissue, Wharton’s Jelly (WJ) in umbilical tissue, amniotic fluid and dental pulp [41]. Since differentiation capacity decreases with age, WJ-derived MSCs show greater potential to differentiate than MSCs derive from other sources. Even so, BM-derived MSCs and adipose tissue-derived cells are most utilised in stem cell therapy due to their plasticity, availability, and immunomodulatory properties compared to other sources. Their lack of major histocompatibility complex (MHC)-I and MHC-II antigens allow for allogenic administration without the risk of transplant-induced teratoma formations [42,43].
The paracrine signalling of the MSCs secretome induce behavioural, mechanical and chemical changes in adjacent cells. These changes result in angiogenic, neovascular and anti-inflammatory effects. The secretome of MSCs also contains factors responsible for directing the fate of other stem cells [44,45,46,47]. The secretome of MSCs includes growth factors, cytokines, chemokines and various anti-inflammatory agents including vascular endothelial growth factor (VEGF), brain-derived neurotrophic factor (BDNF), basic fibroblast growth factor (bFGF), hepatocyte growth factor (HGF), insulin-like growth factor (IGF-1), platelet-derived growth factor (PDGF), transforming growth factor beta-1 (TGF-β1), osteopontin (OPN) and interleukin-6 (IL-6) [41,48,49]. Extracellular vesicles play an important role in transferring these molecules between neighbour cells, allowing for the coordination of regenerative mechanisms and cellular migration, proliferation and homing to the site of injury [50].
Homing of MSCs, and stem cells in general, to the site of injury is an important step in tissue regeneration. Increased availability of OPN after tissue injury is one of the key factors that regulates both MSC homing and migration. OPN mitigates stress-imposed alterations on cellular morphology by suppressing actin stress fibre formation which in turn allows dynamic movement and relocation in that integrin β-1, FAK and ERK pathways appear to play a role [51]. Activation of the stromal-derived factor-1 (SDF-1)/CXC chemokine receptor-4 (CXCR4) pathway, on the other hand, has been implicated in suppression of MSC migration [49].
There is evidence that the anti-inflammatory activities of MSCs is mediated through mechanisms involving VEGF signalling and concomitant reductions in the expression of tumour necrosis factor-α (TNF-α) and transcription factor NF-κB [52]. Indeed, TNF-α modulates the composition of MSC secretome [12,50,53] which may influence the endothelial cell migration, differentiation and proliferation and affect the extent of angiogenesis. It is assumed that bFGF, VEGF, TGF-β, HGF and IL-6 signalling are closely involved in these paracrine effects [54]. In support of these transplantation of adipose-derived MSCs to MCAo rats has been shown to promote angiogenesis and encourage behavioural recovery and transplantation of BM-MSC to cerebral infarcts has been shown to increase VEGF levels, resulting in ERK phosphorylation and repair of white matter damage to help cognitive recovery [55,56]. Furthermore, expression of VEGF and IGF-1 in ischaemic tissue support the regeneration of astrocytes, microglia and oligodendrocytes [57].
BM-MSC can also stimulate the release of BDNF in local parenchyma to activate the Akt/PI3K pathway which meditates cellular growth, proliferation, and angiogenesis [58]. BDNF’s role in neuroregeneration is further recognised in astrocytic Akt/mTOR signalling in the recruitment of additional astrocytes for nerve injury repair [59]. Co-administration of BM-MSCs with regulators of stem cell differentiation and migration, butylidenephthalide and sodium ferulate appear to enhance expression of astrocyte-derived VEGF and BDNF in vivo further supporting the role of MSCs in promoting angiogenesis [59]. Furthermore, by attenuating the degree of stroke-induced calcineurin (CaN) hyperactivation, MSCs rescue neurones undergoing apoptosis and improve neuronal activity in rodents [60]. Similarly, co-application of MSCs with erythropoietin (EPO) triggered neurogenesis and cellular proliferation along the lateral ventricles in rats subjected to transient MCAO [61]. It is of note that studies conducted with animal models of related other diseases like myocardial infarction also provide evidence for the regenerative properties of MSCs [62,63].
MSC-induced neuroplasticity has also been observed in clinical settings where injection of BM-MSC through IV route led to an increase in the number of cluster activations in Brodmann areas BA4 and BA6 and improved clinical outcome as evidenced by Barthel Index (BI) and Fugl-Meyer (FM) scores [64]. A randomised clinical trial also pinpoints the neuroplastic effects of MSCs in recovering motor function in ischaemic stroke patients [65]. The study was consistent with pre-clinical and clinical trials documenting that the IV administration of autologous BM-MSCs is safe and feasible without the precipitation of tumours or related adverse events (AEs). The trial provided further evidence for the paracrine action of MSCs associated with improvements in behavioural and motor abilities and an increase in task-related primary motor cortex (MI) activities. Furthermore, observations of behavioural and physiological improvements two years after treatment, the final point of assessment in the study, demonstrate long-term benefit in these patients and propose MSC therapy as a chronic management strategy for ischaemic stroke [65].
Accumulating recent evidence indicate that transplantation of MSCs through different routes is safe and efficacious in improving patients’ functional outcome [66]. Indeed, while intracerebral administration of BM-MSC attenuated disease severity and improved outcome as evidenced by changes in National Institutes of Health Stroke Scale (NIHSS) and fine motor scores [67], intra-arterial infusion of MSCs during subacute phase of the disease was coupled to a better clinical outcome, defined by modified Rankin Scale (mRS) score < 2, in treatment arm versus control group [68]. Patient improvements have also been witnessed in clinical settings assessing the efficacy of NSC and MSC co-transplantation, where treatment conferred the refinement of speech, balance and muscular control [69,70]. The Table 1 summarized details of the clinical trials indicating the safety of treatments with MSC. Taken together, these studies suggest that larger trials of bigger sample sizes with longer, more extensive follow-ups are necessary in order to comment on treatment efficacy reliably.
Another mechanism involved in the regenerative role of MSCs is rather unique in that MSCs can rescue cells injured due to mitochondrial dysfunction via mitochondrial transfer. The presence of tunnelling nanotubes transferring mitochondria from MSCs to damaged H9c2 cardiomyocytes to restore mitochondrial function has been shown in an in vitro ischaemia/reperfusion model by fluorescent microscopy [71]. Metabolic benefits relating to MSC mitochondrial transfer has also been reported in vivo and in vitro settings in response to oxidative stress [72]. Similar results also show growing evidence favouring MSC use in the treatment of stroke [73].
To abet their regenerative properties, MSCs can be primed or pre-conditioned which involves preparing cells for a specific purpose, including lineage-specific differentiation, through either epigenetic and morphological modifications or manipulation of cell culture environment [74]. Studies comparing MSC efficacy before and after priming attribute an important role to cell priming in achieving higher therapeutic efficacy [74,75].
6. Endothelial Progenitor Cells (EPCs)
EPCs are circulating stem cells of endothelial origin. They migrate and accumulate in areas of vascular injury to help repair damaged vasculature through both neovascularisation and vascular remodelling. Due their ability to detect and replace the damaged cerebral endothelial cells and restore BBB integrity by differentiating into mature endothelial cells, they are regarded as an important therapeutic for the management of ischaemic stroke [76,77]. Insufficient number and dysfunction of EPCs impairs vascular homeostatic and accelerates vascular disease [78,79]. EPCs are released into circulation by bone marrow in response to an ischaemic injury. They are isolated from mononuclear cell (MNC) population through use of specific antigens targeting endothelial cell maturity (e.g. KDR+), immaturity (e.g. CD133+) and stemness (e.g. CD34+) amongst a non-haematopoietic cell (CD45-) population [80]. To obtain cells that can be used for therapeutic purposes, MNCs are cultivated using specific endothelial cell media supplemented with a range of factors, including fibroblast growth factor (FGF), VEGF, insulin-like growth factor (IGF), hydrocortisone, ascorbic acid and heparin [81,82,83]. Exogenous addition of EPCs repairs the integrity of an in vitro BBB model under OGD condition and attenuates ischaemia-evoked oxidative stress and apoptosis of endothelial cell [76,84,85]. Beside administering the stem cells directly, the condition media of outgrowth endothelial cells were found to negate the deleterious effects of TNF-α on BBB function in an in vitro tri-culture model of BBB [86]. EPCs are able to secret a wide range of substances to regulate angiogenesis, migration, proliferation and antipoptosis, including angiogenin, HGF, PDGF, VEGF and PBEF [87]. Clinical study indicated that post-stroke patients have elevated levels of TNF-α and in vitro study determined that TNF-α significantly impair the function and integrity of BBB [86,88]. Aside from their therapeutic potential, EPCs can also be used as diagnostic and prognostic biomarkers. Significantly higher numbers of circulation EPCs were found in patients in the acute and subacute phase after ischaemic stroke, compared to healthy volunteers, indicated that the EPCs number can be used to monitor the recovery/progression of ischaemic stroke. However, no correlation was determined between EPC counts and patients’ outcome [89]. However, recent observation of a close correlation between baseline CD34+KDR+ and CD133+KDR+ counts and the outcome of stroke supports the idea that these particular EPC subtypes as potential prognostic markers for ischaemic stroke [90]. Low EPC concentrations have been reported in chronic stroke patients and those with cardiovascular risk factors [91].
EPCs in culture produce two distinct types of cells: early-EPCs (eEPCs) and outgrowth ECs (OECs) or endothelial colony-forming cells (ECFCs). eEPCs represent an immature (CD133+) population of EPCs, with little proliferative capacity, appearing early in culture (three to four days). In contrast, OECs appear late in culture (two to four weeks) and demonstrate maturity and commitment to differentiation [92]. eEPCs and OECs can also be distinguished by their different morphology in that while eEPCs show a spindle-shaped morphology, OECs manifest the classical endothelial phenotype of cobblestone morphology [80,87]. Moreover, eEPCs express haematopietic markers CD45 and CD14 while OECs express endothelial markers, CD31 CD146 CD105 and stemness marker CD34 [93,94].
A variety of agents, including VEGF, NO, EPO, SDF-1 and active MMP-9 regulate the mobilisation of EPCs from BM into circulation [95,96]. VEGF, a key mediator of angiogenesis, stimulates EC proliferation, migration and tube formation, eventually giving rise to new blood vessels and capillary networks [97,98,99]. In vivo and in vitro experiments using rat spleen-derived EPCs have documented that connexin-43 plays a pivotal role in EPC differentiation while VEGF promotes EPC proliferation and vascular repair [99]. VEGF meditation of angiogenesis explains just one of the mechanisms by which EPCs promote neovascularisation [100].
The trafficking of EPCs, co-ordinated by SDF-1, is supported by results of another in vivo investigation looking at the relationship between hypoxia-inducible factor-1 (HIF-1) and SDF-1 in ischaemic mice [29]. The study concluded that HIF-1 directly regulated SDF-1 gene expression in ischaemic tissue and that the migration and adhesion of EPCs to sites of injury was supported via CXCR4 and SDF-1 binding. A different study exploring the dynamics of EPC-induced vascular remodelling and angiogenesis provide evidence for IL-6 which activate gp80/130 signalling pathways, including downstream ERK1/2 phosphorylation [101]. Most importantly, this study confirmed that IL-6 stimulates EPC proliferation, movement, and adhesion. Manipulation of ischaemic matrigel models confirm this relationship where IL-6 influence was credited with significantly promoting EPC capillary-like tube formations [101].
Nitric oxide generated in endothelial cells by endothelial nitric oxide synthase (eNOS) is another important molecule that co-ordinates EPC proliferation and migration and inhibits apoptosis and platelet aggregation [102,103]. Observation of an impaired ischaemia-induced neovascularisation in eNOS-deficient mice bestows a key role on NO in mobilising EPCs. By inducing phosphorylation of eNOS, VEGF plays an important role in stimulating NO production, a relationship confirmed by increases in peripheral EPC count in normal mice after VEGF administration, but not in eNOS deficient mice [104]. This study also shows that exogenous EPC delivery improves limb function and recovery in “healthy” mice in that NO-mediated activation of MMP-9 appears to be critical [105]. Activated MMP-9 promotes the transformation of insoluble membrane-bound Kit ligand into its soluble arrangement (sKitL) allowing for movement and mobilisation of EPCs from BM into circulation [95].
Another pathway linked to the neovascular effects of EPCs is Notch1, a transmembrane receptor. Notch1 and its ligand Jagged1 has been implicated in post-ischaemic neovascularisation in both experimental and clinical stroke where increases in the expression of activated Notch1 (Notch intracellular domain or NICD) in peri-infarct endothelial cells are coupled to the level of angiogenesis [106]. Neo-angiogenesis occurs by the proliferative sprouting of endothelial tip cells, followed and stabilised by endothelial stalk cells. Notch1 signalling co-ordinates this motility between tip and stalk cells and possibly directs arterial EC differentiation [107]. This relationship is supported by suppression of tumour growth via inhibition of Notch signalling [108]. NICD may also regulate transcription and allow physical cellular changes to take place during angiogenesis [107]. Defects in EPC mobilisation contribute to neovascularisation impairment in ischaemic tissue, again supporting the association between EPCs and their role in vascular remodelling [105].
Whilst there are many pre-clinical studies scrutinising the safety and efficacy of EPC treatment in stroke, only few clinical studies investigate the therapeutic efficacy of autologous EPCs in stroke patients. For instance, a recent phase I/IIa study examining the effects of autologous EPC transplantation in eighteen acute ischaemic stroke patients has reported this approach to be safe with no sign of any increased tumorigenicity or any other adverse events over the four-year follow-up period. Though no improvements in neurological outcomes were measured, the lower incidence of adverse reactions suggests a certain degree of efficacy in improving the quality of life. Pre-conditioning of treatment may be considered here, where upregulation of CXCR4 could become a therapeutic target in enhancing EPC efficacy [109]. Other attempts to increase HIF-1 and VEGF also concluded that the overexpression of such genes augmented neovascularisation, proving therapeutic benefit as a result of EPC priming [28,96].
7. Haematopoietic Stem Cells
HSCs are multipotent, tissue-specific stem cells able to give rise to all functional blood cell types, including leukocytes, erythrocytes and thrombocytes. HSCs present treatment possibilities as their supplementation encourages the recovery of diseased tissue by restoring blood and oxygen flow. Regeneration of ischaemic cells is facilitated by HSC differentiation (haematopoiesis), a process regulated by several hormones and cytokines, namely EPO, IL-3, granulocyte colony-stimulating factor (G-CSF) and macrophage colony-stimulating factor (M-CSF) [110]. CD34, though a surface marker expressed by other cells, is generally understood to represent hematopoietic stem and hematopoietic progenitor cells [111]. CD45 is another notable marker of HSCs [112]. HSCs can be obtained from BM, peripheral blood and umbilical cord blood. Where engrafting HSCs from BM-MNCs allows for faster expansion, they express higher human leukocyte antigen (HLA) levels creating problems with rejection during allogeneic therapy. Conversely, cord cells are more immature and flexible in HLA matching but are lower in concentration [113].
The human adult produces over two hundred billion red blood cells per day [114]. With such a high turnover rate, the proliferative abilities of stem cells are most vitally exercised here where cell fate, regarding self-renewal or differentiation, is determined by gene expression and regulated by transcriptional factors [115,116]. Regulators between the two pathways are not distinct or separate, with factors able to influence cell fate down either route. However, some lineage-specific growth factors, such as G-CSF, M-CSF and EPO are categorical in directing HSCs down their respective pathways [117]. At high concentrations, GATA-1 suppresses the HSC exosome complex, consequently arresting early erythroblast proliferation and thus allowing for their maturation [118]. Conversely, GATA-1 downregulation is an important molecular cue in terminal erythroid maturation [119].
The Wnt and Notch pathways are other regulators of haematopoietic cell fate. Both Wnt and Notch receptors are widely expressed throughout the haematopoietic system and are critical in co-ordinating the development of leukocytes and their divisions. Wnt3a and Notch signalling promote early T-cell differentiation in human umbilical cord (hUCB) blood stem cells. Conversely, inhibition of Wnt in the presence of Notch instead directs HSCs to give rise to natural killer cells [120]. It is necessary to understand the signalling that occurs during HSC differentiation as it allows better identification of potential targets to enhance therapeutic efficacy of stem cell application in future trials.
Aside from transcriptional signalling, external situations also drive haematopoietic cell fate. For example, erythropoiesis occurs when HIF is activated under oxidative stress [121]. A study with MCAO rats provides insight into this relationship [122] where rats intracerebrally receive a culture of hypoxia-exposed (3% O2) HSCs displayed significantly better neurological outcomes compared to those received normoxia-exposed (20% O2) or no treatment at all. This study also showed the role of exchange protein Epac1 in regulating the HIF/MMP pathway, with evidence connecting this communication to the promotion of neural progenitor cell (NPC) homing, aiding cerebral neuroplasticity. These results confirm previous findings documenting Epac1 action to enhance MMP activity and promote neovascularisation through the integrin-mediated adhesion of circulating HSCs to endothelial layers [123]. CD45+ bone marrow mononuclear cells (BMMNCs) were shown to differentiate into endothelial cells and smooth muscle cells to promote angiogenesis in an ischaemic stroke rat model [124]. Other pre-clinical studies have also reported enhanced angiogenesis as a result of HSC treatment [125,126].
Analysis of an “open-labelled nonrandomised phase I/II trial” investigating the safety of G-CSF treatment in patients with acute ischaemic stroke allowed for broader observations of HSC treatment. Subcutaneous G-CSF administration was deemed safe and feasible; however, the small cohort could not infer a clear dose-response relationship between G-CSF and related HSC mobilisation. The improvements in neurocognitive performance, long-term memory and attention observed in this study may derive from the adjunctive contribution of IV thrombolysis present in this study [127].
The first clinical trial measuring IA autologous CD34+ stem cell delivery in human ischaemic stroke patients found intervention was well-tolerated by all participants (primary outcome measure) with improvements in clinical function characterised by a significant decrease in mean NIHSS score from 10.40 to 2.20 (95% CI; 3.69–12.71; p = .007) [128]. In addition, reductions in lesion volume were also observed, furthering evidence of neuroprotection. A separate trial into the intrathecal delivery of CD34+ stem cells also deemed safe and did not lead to any allergic or immunological AEs (Wang et al., 2013). Improvements in neurological scores, as assessed by NIHSS and BI, were again observed. These promising results and that of other studies [129,130,131,132], prompted STROKE34 (EU clinical trial registration: 2017-002456-88), a randomised controlled phase IIa trial of IA CD34+ cells in acute ischaemic stroke [133]. Its primary outcome looks to measure infarct volume by magnetic resonance imaging (mRI) at three months, with secondary outcomes looking into the safety and efficacy of treatment. The results of the trial are expected to shed some light on the direction of future studies with HSC in the field of stroke medicine.
Finally, in an attempt to augment therapeutic benefit, one study explored the idea of genetic editing of HLA expression in HSCs [134]. Downregulation of HLA-A lifts restrictions on current HSC-based therapies as the risk of rejection for allogenic transplantation is reduced. This increases the suitability of treatment in cases where the patient’s HLA profile is under-represented in the current donor pool.
8. Neural Stem Cells
NSCs are undifferentiated stem cells of the CNS. They are multipotent stem cells able to self-renew and proliferate, give rise to different cell types and differentiate into the three cell types of neural lineage, neurones, astrocytes, and oligodendrocytes [135]. Neurones, simply, are electrically excitable cells that synaptically transmit signals throughout the body [136]. Glial cells support and define these communications and are categorised by their functions; astrocytes maintain an appropriate chemical environment for brain functionality and oligodendrocytes are responsible for myelination [137]. NSCs are sometimes referred to in literature as “NPCs”, “neural precursor cells” or “radial glia” of which terminology is used interchangeably and tends to be a difference of semantics. For clarity, this review distinguishes NSCs from their progenitors by the differences in their capacity to proliferate and differentiate. Where NSCs can infinitely divide, NPCs are slightly more specialised with a limited number of replication cycles. NPCs also cannot give rise to non-neural cells present in the CNS, such as immune cells, whereas NSCs can [135].
NSCs originate from neuroectodermal tissue of the neural plate and are primarily found in the ventricular-subventricular zone (V-SVZ) of the walls of the lateral ventricles and the subgranular zone (SGZ) of the dentate nucleus [138,139]. NSCs are isolated by enzymatic digestion of these locations [140] and quantified either in vitro by Reynolds and Weiss’ method of Neurosphere Assay or by using a more recently developed collagen-based assay, Neural Colony Forming Cell (NCFC) Assay [141]. NCFC Assays are now more commonly used as they are efficient in multiplying NSC count and can also discriminate between NSC and NPC populations by analysing the sizes of the colonies, representative of their proliferative abilities, the assay produces [142].
Neurogenesis is the growth and development of neuronal tissue and occurs both prenatally and in adults. It is the process by which NSCs develop into either neurones or glial cells (gliogenesis) and is influenced by both internal and external factors. Extrinsic factors in the local microenvironment of the SVZ and SGZ determine the lineage of NSCs, with soluble factors and transcriptional factors controlling intracellular signalling cascades such as the Notch-Hes1 pathway [143,144,145]. Activation of such pathways, triggered by oxidative pressures, decide whether NSCs will transform into astrocytes and oligodendrocytes or differentiate into neurones. As discussed before, MSCs can also induce neurogenesis. bFGF and EGF initiate the self-differentiation of MSCs into NPCs [46] or direct the differentiation of already established NSCs into astrocytes or neurones [44].
By replacing necrotic neurones and positively influencing neuroregenerative pathways adversely affected by ischaemia, NSCs, through neurogenesis, present an exciting therapeutic option. Migration and differentiation of NSCs into mature neurons have been shown to restore cerebral homeostasis in MCAO rats [146]. Other therapeutic actions of NSCs such as those including the modulation of immunomodulatory response, reorganisation of neuronal pathways and angiogenesis somewhat resemble that of MSCs. The immunomodulatory properties of NSCs are supported by a marked attenuation in BBB damage, reduced cytokine production and expression of pro-inflammatory markers IL-6 and TNF-α observed in acute stroke mice injected with a mixture of human-induced pluripotent stem cells (iPSCs) and NSCs in hippocampus [147]. Behavioural improvements observed in these mice were comparable to those noted by other studies [148,149]. In another study improvements in behavioural scores of NSC-treated mice appeared to correlate with the extent of angiogenesis and reduction in infarct volumes [150]. The neuroprotective effects of NSCs are further recognised in a study of pre-conditioned cells in magnetically targeted MCAO rats [151]. The study found that compared to the other treatment groups, the pre-conditioned NSCs demonstrated better migration and differentiation capacity as well as comparable improvements in neurological function. Taken together these studies revealed a range of NSC-related benefits ranging from cell replacement to enhanced vascularisation thereby proving the ability of NSC to induce improvements in functional outcomes and neuronal reorganisation.
Similar to translational studies, treatments with NSCs have led to improved mental status, limb strengthening and speech recovery in clinical settings. These were inevitably associated with marked improvements in overall quality of life compared to the control group and functional benefits further manifested in improvements in functional status [152].
The Pilot Investigation of Stem Cells in Stroke (PISCES) trials are a collection of clinical studies looking at NSC treatment for ischaemic stroke. In response to a successful pre-clinical trial in which CTX-DP (a manufactured product as the suspension composed of CTX0E03 cells at a concentration of 5 × 104 cells/μl) yielded sensorimotor improvements in MCAO rats, an outsetting phase-I, open-label, dose-escalation study into the safety and tolerability of CTX-DP was conducted in human stroke patients [153]. The trial was thorough in its endeavours, analysing eleven men at a range of doses (three patients receiving two million CTX0E03 NSCs; three other patients receiving five million; three others receiving ten million; two others receiving twenty million) at a mean time of twenty-nine months (range from 6-60 months) after stroke onset. The primary endpoint was safety, measuring the emergence of any serious AEs to which no treatment-related adversities were found thus proving safety up to twenty million CTX0E03 NSCs. Though changes in NIHSS and mRS scores suggested some degree of neurological and functional improvement, the trial concluded that the small patient population and nature of its design limited the reliability of these conclusions, precluding further investigations.
PISCES-2 also reflected this requisite for additional research [154]. Intracerebral implantation of CTX0E03 NSCs (dose twenty million) was deemed feasible and safe, and improvements in Action Research Arm Test’s (ARAT, an evaluative measure to assess limb function among individuals with cortical damage) were seen in a total of four patients (17%) by twelve months after implantation. It was noted these improvements were only found by those who initially demonstrated residual upper limb control and not by anyone with absent upper limb movement at baseline. PISCES-3 (trial registration: NCT03629275) began in August 2018 and was the natural continuation of PISCES-2. Unfortunately, PISCES-3 trial was terminated due to the COVID-19 pandemic. Whilst few other clinical trials are assessing NSCs, there is evidence in other areas to prove the transplantation of neurones themselves is safe and feasible [155,156]. However, no clinical benefit was found in these cases, perhaps due to the therapeutic limitations of mature neuronal cells.

Table 1.
Translational and clinical studies employing stem cells as therapeutics.
| Cell type | Dose | Route of Administration | Timing of treatment (post model onset) | Participants | Outcome Assessment | References |
|---|---|---|---|---|---|---|
| Preclinical research | ||||||
| AD-MSC | 2×106 | IV | 2 - 7d | 44 MCAO rats | Safe; improved sensorimotor function | [56] |
| BM-MSC | 2×106 |
IV & IP | 3 hrs- 7 d | 36 MCAO rats | Significantly improved neurological function | [59] |
| BM-MNCs | 3×106 |
IV | 4 d | 71 rats | Improved cognitive function | [55] |
| MSCs | 1×105 |
IA | 6 hrs | MCAO rats | Improved functional outcome | [60] |
| BM-MSCs | 1−2×106 |
IV | 72 hrs | T2DM-MCAO rats | Improved functional outcome | [63] |
| MSCs &EPO | 2×106 |
IV | 24 hrs | Focal ischaemic rats | Increased neurogenesis | [61] |
| EPCs | 4×106 |
IV | ND | Hind limb ischaemic rats | Improved limb function | [105] |
| EPCs | 4×106 |
IV | 24 hrs | MCAO rats | Improved functional outcome | [157] |
| ECFC & EPO | 5×106 |
IV & IP | 24 -72 hrs | Focal ischaemic rats | Improved neurological function | [158] |
| ECFCs | 1×106 |
IA | 72 hrs | Focal ischaemic mice | Improved neurological function | [159] |
| hUCB-HSCs | 1×106 |
Intracerebral | 7 d | MCAO rats | Improved neurological function | [122] |
| iPSC-NSCs | 1×106 |
Intra-striatal | 7-14 d | 15 MCAO mice | Improved behavioural and sensorimotor function | [146] |
| iPSC-NSCs | 1×105 |
Intra-hippocampal | 24 hrs | MCAO mice | Improved neurological function | [147] |
| NSCs | 1×105 |
Intracerebral | 24 hrs | MCAO mice | Behavioural improvement | [148] |
| NSCs | 3×106 |
IV | 6 hrs | 15 MCAO rats | Neuroprotective effects | [149] |
| NSCs | 1.2×105 |
Intracerebral | 24 hrs | 7 MCAO rats | Increased vascularisation | [150] |
| NSCs | 4×106 |
IV | 24 hrs | 23 MCAO rate | Improved neurological function | [151] |
| HSCs | 𝑁𝐷 |
IV | 48 hrs | Transient ischaemic mice | Increased neovascularisation | [125] |
| OECs | 4×106 | IV | 24 hrs | MCAO mice | Decreased brain oedema volume | [76] |
| Clinical trials | ||||||
| Acute phase | ||||||
| BM-MNCs | 4−6×108 |
IV | 24-72 hrs | 10 | Safe; clinical improvements | [130] |
| HSCs | 2.5−10 µ𝑔/𝑘𝑔 | Subcutaneous injection | 12 hrs | 20 | Safe; neuropsychological improvements | [127] |
| Sub-acute phase | ||||||
| HSCs | 5.1×107−6×108 |
IA | 3-7 d | 20 | Safe; clinical improvements | [160] |
| HSCs | 1.59×108 |
IA | 5-9 d | 10 | Feasible and safe | [131] |
| HSCs | 1×108 |
IA | 7 d | 5 | Safe; significant clinical improvements | [128] |
| BM-ALDHbr stem cells | 1.6×105−7.5×107 |
IA | 11-19 d | 29 | Safe | [31] |
| UC stem cells | 5×106−5×107/𝑘𝑔 |
IV | 3-10 d | 10 | Safe and feasible | [161] |
| HSCs | 6.1×108 |
IA | 8-15 d | 10 | Safe; good clinical outcome | [68] |
| HSCs | 2.8×108 |
IV | 18 d | 58 | Safe | [132] |
| HSCs | 3×107 |
IA | 9 d | 1 | Feasible | [129] |
| MSC | 1×108 |
IV | ND | 5 | Safe | [162] |
| Chronic phase | ||||||
| BM-MSCs | 1×108 (𝑛=10) 3×108 (𝑛=10) |
IV | 1 m | 16 | Safe; behavioural and physiological improvements | [65] |
| MSCs &NSCs | 0.5−6×106/𝑘𝑔 | IV & intracistern | <1 wk-2 yrs | 6 | Safe | [70] |
| MSCs | 0.5−1.5×106/𝑘𝑔 |
IV | >6 m | 36 | Safe; behavioural improvements | [163] |
| EPCs | 5×106/𝑘g |
IV | 4-6 wks | 18 | Improved long-term safety | [164] |
| NSCs (CTX-DP) | 0.2−2×107 |
Ipsilateral putamen injection | 6-60 m | 11 | Safe; improved neurological function | [153] |
| NSCs | 2×107 |
Intracerebral | 2-13 m | 23 | Improvements in upper limb function | [154] |
| BM-MSCs | 2.5×106−1×107 |
Intracerebral | 6-60 m | 18 | Safe; significant clinical improvements | [67] |
| HSCs | 0.8−3.3×107 |
Intrathecal | 1-7 yrs | 8 | Safe; improved clinical neurological function | [165] |
| Neurones | 0.5−1×107 |
ND | 6 m-6 yrs | 26 | Safe and feasible | [155,156] |
| US-MSCs | 2×107 |
IA | <3 m | 3 | Safe; improved neurological function | [166] |
| NSCs & HSCs | 2×108 |
Intracerebral | ND | 10 | Safe; functional improvements | [152] |
| BM-MSCs/HSCs | 5−6×107 |
IV | 3 m-2 yrs | 20 | Safe; significant functional improvements | [64] |
| NPCs&UC-MSCS | 0.2−2.3×107 |
Intraparenchymal | 6 m-20 yrs | 10 | Safe; functional improvements | [69] |
| MSC | 0.2−2.3×108 | IV | <6 m | 12 | Safe and feasible | [66] |
AD, adipose tissue; MSC, mesenchymal stem cell; MCAO, middle cerebral artery occlusion; BM, bone marrow; EPO, erythropoietin; ECFC, endothelial colony forming cell; hUCB,human umbilical cord blood; iPSC, induced pluripotent stem cell; MNCs, mononuclear cells; NPCs, neural progenitor cells.
9. Route, Dose and Timing of Treatment
9.1. Route
The main routes for treatment are IV, IA and intracerebral administration. Both the pre-clinical and clinical studies show the IV route as the most preferred route due largely to ease of use and its non-invasive nature. However, the IV treatment poses issues with engraftment and thereaputic efficacy due to clearance of most cells by the lungs and liver during circulation [167,168]. Furthermore, chemotaxis signalling subsides over time which suggests IV administration may be more suitable in treating acute and subacute stroke where the levels of inflammatory biomarkers are at their highest, compared to chronic cases. Even so, studies demonstrating the efficacy of IV administration in chronic settings also exist, supporting the fundamental property of stem cells in their ability to home to sites of injury [163]. Additional references for this can be found in current Table 1.
IA administration is similar in technique as a minimally invasive and straightforward procedure. It is argued the IA route is more efficient than IV transport as this route does not lead to excessive cell trapping. While some studies comparing routes of stem cell delivery favour the IA route over IV administration [169], others report that there is no real difference in efficiency, with both routes revealing similar biodistribution rates and comparable functional outcomes [170,171,172].
Another route mentioned is the intracerebral route. The direct administration to site of injury eliminates the need to rely on chemical paracrine signalling in directing stem cell migration, allowing for smaller dose deliveries. This also, in theory, makes it a better option for chronic stroke patients (where the homing of stem cells may be weaker due to the absence of inflammatory mediators attracting as such) to maximise stem cell transfer; however, not all targets are physically accessible. Its neuronal nature has been associated therapeutic benefits, however, its intrusiveness increases the risk of adversities, including infection and haemorrhage [173].
Other routes include intrathecal and intraperitoneal administration. However, little is known about the efficacy and overall suitability of these routes due to limited availability of studies employing them. When organising management strategies for ischaemic stroke patients, concerns of safety, the stage of stroke, and other practical measures must be considered. Although there is little evidence comparing the effectiveness of routes at different stages, it is reasonable to think that IV and IA routes may prove greater therapeutic effect in acute and subacute phase of the disease [68,128], whereas IC administration may be better suited for patients in chronic phase of the disease [67,153,154]. Figure 2.
9.2. Dose
Despite investigation of a wide range of cell concentrations in various clinical and pre-clinical studies, the optimal dose for an effective therapy after a cerebral ischaemic event continues to be a matter of debate. Lack of AEs at all doses tested negates the concerns regarding the numbers of stem cells to be administered and suggests consideration of reported efficacy of cells at a particular dose for a particular stem cell type. Though no clinical studies specifically evaluate the differences in stem cell efficacy at different concentrations, several studies comment on safety over a range of cell doses. In the studies discussed in this paper, doses of cells administered varied from 0.5 x 105 cells/kg to 6.1 x 108 cells [68,163]. In clinical trials, higher cell doses appear generally to associate with better outcomes and lower transplant-related mortalities. However, as alluded above, it is impossible to draw a conclusion about the relationship between treatment concentrations and efficacy [65,67,68,128]. Functional and neurological improvements in pre-clinical trials were observed throughout the range of 1 x 105 - 5 x 106 cells/kg, suggesting an optimal dose may lie within [60,126,148,158].
The optimal dose of stem cell treatments is likely to be dependent on the the cell types and administration routes. It is important to remember that certain routes, notably IV injection will lead to major losses in stem cell numbers due to their trapping by the lungs [174]. Therefore, higher dose and repeated injection may be necessary while using these routes [175].
9.3. Timing
One of the biggest arguments for investing time and resources into stem cell research is the hope that emerging treatment option(s) will demonstrate a larger therapeutic window than current timely limitations. The short life span of rodents is an issue when considering long-term intervention, explaining why pre-clinical studies fail to produce data on optimal treatment timing. Clinical trials, however, can evaluate the safety, feasibility and efficacy of treatments with stem cells over a significant period of time. In addition, the time of administration vary significantly in clinical studies ranging from twelve hours to twenty years, where safety is confirmed throughout [69,127].
There is little clinical evidence as to the application of stem cells during the hyperacute phase of stroke, so it is difficult to establish a consensus on the optimal timing of treatment in the immediate aftermath of stroke. In contrast, several clinical studies with acute stroke patients exist. They unanimously show that patients received stem cells 7-72 hours after stroke onset displayed better neurological outcomes [127,130]. A multi-centre phase 2 clinical trial showed improved outcomes in acute ischaemic stroke patients who received intravenous multipotent adult progenitor cells within the first 36 hours of stroke, suggesting greater benefits of early interventions with stem cells [176]. Clinical improvements with stem cell therapy are also commonly observed during the chronic phase of stroke, even up to 5 years after stroke onset [67,153,154,163]. Pre-clinical studies, which solely investigate the efficacy of stem cell reatment in hyperacute or acute stroke models, replicate these findings, report improved outcomes and strengthen the argument that earlier treatments provide greater benefits. Even so, the current evidence do not strongly associate the degree of therapeutic efficacy with the timing of treatment and imply a need for future studies.
10. Discussion and Conclusion
This paper set out to discuss the potential role of different type of stem cells as therapeutics for ischaemic stroke. Pre-clinical and clinical data analysed throughout the text provide insight into the current position of treatments with stem cells in ischaemic stroke. Evaluation of regenerative processes such as angiogenesis, neovascularisation, neurogenesis and erythropoiesis explain the ways in which stem cells act through differentiation and proliferation to recover ischaemic tissue. In addition, analysis of the (paracrine) signalling that direct these activities further help guide our understanding of stem cells as therapeutics.
In light of the existing literature, it is safe to conclude that stem cell-based treatments are safe. Indeed, safety and feasibility are conclusively measured in all the relevant studies cited in this review and it is evident that intervention with stem cells for ischaemic stroke does not induce any major AEs. This is an important feature, especially when considering current treatment strategies associated with few complications, such as haemorrhage or infection.
In terms of therapeutic efficacy of stem cell application, majority of the pre-clinical studies yielded positive results ranging from enhanced angiogenesis to improved sensorimotor functions. Similarly, a number of clinical studies has also reported clinical improvements [65,68,127] while others failed to document any neurological or functional benefit [31,70,131,164]. This may in part be due to the fact that in pre-clinical studies acute interventions aiming to rescue ischaemic tissue present fewer challenges (with chemotaxic signalling and viability of penumbra are at their highest) than late interventions in chronic stroke in clinical settings. It could also be due to the design limitations of early phase trials, where research is restricted in its endeavours (Phases of Clinical Trials, 2019). Phase III and later trials may properly abet investigations into treatment efficacy with more time, resources, and larger patient cohorts. Trials like “Umbilical cord-derived Mesenchymal Stem Cells for Ischaemic Stroke (UMSIS; NCT04811651): a Prospective, Double-blinded, Randomized Controlled, Pilot Study” may provide insight on the direction of future clinical trials. With an estimated enrolment of two hundred participants, UMSIS is one of the largest trials in the area. It is a quadruple masked, randomised parallel assignment monitoring the effects of IV injected umbilical cord-MSCs (1 x 108 cells). The trial evaluates functional improvements by primarily comparing mRS scores before and after treatment. It will also examine other clinical measures such as changes in FM scale and NIHSS score.
Methods and ideas surrounding treatment priming have also gained attention in regenerative stroke research. Another option for future practice may involve mixed cell approaches, whereby conjunctive therapy using multiple stem cell types promises to target a range of pathologies [177,178]. Research may also benefit from investigations into the parameters of route, dose and timing of administration to create a standard strategy for stem cell treatment, providing an interventional framework available for case-to-case manipulation.
Due to a gap in literature, it is difficult to reflect on the other elements of treatment, such as cost-effectiveness and the practicalities of cell proliferation. Ethics is another important consideration of stem cell research. Though the use of adult and cord blood stem cells are less topical than embryonic sources, they still raise regulatory concerns over genetic manipulation [179]. Albeit somewhat crucial for successful allogeneic therapy [163], genetic editing may not completely eradicate the risk of rejection.
Pre-clinical or translational studies constitute important prerequisites for proving or disproving the therapeutic action or capacity of any given agent. They provide preliminary data on the desired biological effect (efficacy) and associated toxicities (safety) of drugs which ultimately inform the design of subsequent clinical research [180]. The majority of pre-clinical studies investigating the impact of stem cell therapy for stroke utilises a rodent model of human ischaemic stroke, achieved by temporary or permanent occlusion of middle cerebral artery (MCAO) [181,182,183]. Damage occurring in the cortex and subcortical structures like the thalamus and striatum in this model reflect the pathology observed in clinical settings. Following clinical trials with stem cells in ischaemic stroke have shown the safety and effectiveness of this approach in humans [65,68,161,164]. However, for the progression of clinical studies into later phase trials (phase III onwards) treatment safety must be confirmed.
In conclusion, stem cell treatment presents possibilities for patients with all types of ischaemic stroke. With evidence of safety and efficacy measured in patients with acute, subacute and chronic disease, therapeutic interventions appear to be promising for patients at every stage of disease (Table 1). However, further clinical research is necessary to standardise the treatment regimens.
References
- Brine, S. New figures show larger proportion of strokes in the middle aged. 2018. Available online: https://www.gov.uk/government/news/new-figures-show-larger-proportion-of-strokes-in-the-middle-aged (accessed on 22 October 2023).
- Clark, W.M.; Albers, G.W.; Madden, K.P.; Hamilton, S. The rtPA (alteplase) 0- to 6-hour acute stroke trial, part A (A0276g): results of a double-blind, placebo-controlled, multicenter study. Thromblytic therapy in acute ischemic stroke study investigators. Stroke 2000, 31, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Del Zoppo, G.J.; Saver, J.L.; Jauch, E.C.; Adams, H.P., Jr.; American Heart Association Stroke, C. Expansion of the time window for treatment of acute ischemic stroke with intravenous tissue plasminogen activator: a science advisory from the American Heart Association/American Stroke Association. Stroke 2009, 40, 2945–2948. [Google Scholar] [CrossRef] [PubMed]
- Broocks, G.; Kniep, H.; Kemmling, A.; Flottmann, F.; Nawabi, J.; Elsayed, S.; Schön, G.; Thomalla, G.; Fiehler, J.; Hanning, U. Effect of intravenous alteplase on ischaemic lesion water homeostasis. European Journal of Neurology 2020, 27, 376–383. [Google Scholar] [CrossRef]
- Nogueira, R.G.; Jadhav, A.P.; Haussen, D.C.; Bonafe, A.; Budzik, R.F.; Bhuva, P.; Yavagal, D.R.; Ribo, M.; Cognard, C.; Hanel, R.A.; et al. Thrombectomy 6 to 24 Hours after Stroke with a Mismatch between Deficit and Infarct. N Engl J Med 2018, 378, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Powers, W.J.; Rabinstein, A.A.; Ackerson, T.; Adeoye, O.M.; Bambakidis, N.C.; Becker, K.; Biller, J.; Brown, M.; Demaerschalk, B.M.; Hoh, B.; et al. 2018 Guidelines for the Early Management of Patients With Acute Ischemic Stroke: A Guideline for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke 2018, 49, e46–e110. [Google Scholar] [CrossRef] [PubMed]
- Wheaton, W.W.; Chandel, N.S. Hypoxia. 2. Hypoxia regulates cellular metabolism. American Journal of Physiology-Cell Physiology 2011, 300, C385–C393. [Google Scholar] [CrossRef] [PubMed]
- Losenkova, K.; Zuccarini, M.; Helenius, M.; Jacquemet, G.; Gerasimovskaya, E.; Tallgren, C.; Jalkanen, S.; Yegutkin, G.G. Endothelial cells cope with hypoxia-induced depletion of ATP via activation of cellular purine turnover and phosphotransfer networks. Biochim Biophys Acta Mol Basis Dis 2018, 1864, 1804–1815. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, L.; Cataldi, M.; Pignataro, G.; Secondo, A.; Molinaro, P. Glutamate-independent calcium toxicity: introduction. Stroke 2007, 38, 661–664. [Google Scholar] [CrossRef] [PubMed]
- Bano, D.; Nicotera, P. Ca<sup>2+</sup> Signals and Neuronal Death in Brain Ischemia. Stroke 2007, 38, 674–676. [Google Scholar] [CrossRef]
- Garcia, J.H.; Liu, K.F.; Yoshida, Y.; Lian, J.; Chen, S.; del Zoppo, G.J. Influx of leukocytes and platelets in an evolving brain infarct (Wistar rat). Am J Pathol 1994, 144, 188–199. [Google Scholar]
- Gursoy-Ozdemir, Y.; Can, A.; Dalkara, T. Reperfusion-induced oxidative/nitrative injury to neurovascular unit after focal cerebral ischemia. Stroke 2004, 35, 1449–1453. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.L.; Srivastava, K.; Sprigg, N.; Bath, P.M.; Bayraktutan, U. Inhibition of Rho-kinase protects cerebral barrier from ischaemia-evoked injury through modulations of endothelial cell oxidative stress and tight junctions. J Neurochem 2014, 129, 816–826. [Google Scholar] [CrossRef]
- Rakkar, K.; Bayraktutan, U. Increases in intracellular calcium perturb blood-brain barrier via protein kinase C-alpha and apoptosis. Biochim Biophys Acta 2016, 1862, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Shin, D.; Kim, J.-H.; Shin, Y.-J.; Rajanikant, G.K.; Majid, A.; Baek, S.-H.; Bae, O.-N. Role of Autophagy in Endothelial Damage and Blood–Brain Barrier Disruption in Ischemic Stroke. Stroke 2018, 49, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.X.; Li, Y.J.; Shi, K.; Wood, K.; Ducruet, A.F.; Liu, Q. IL (Interleukin)-15 Bridges Astrocyte-Microglia Crosstalk and Exacerbates Brain Injury Following Intracerebral Hemorrhage. Stroke 2020, 51, 967–974. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Yang, G.Y.; Zhang, Z. Crosstalk of Astrocytes and Other Cells during Ischemic Stroke. Life (Basel) 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Broughton, B.R.; Reutens, D.C.; Sobey, C.G. Apoptotic mechanisms after cerebral ischemia. Stroke 2009, 40, e331–339. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicologic Pathology 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Skulachev, V.P. Bioenergetic aspects of apoptosis, necrosis and mitoptosis. Apoptosis 2006, 11, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol Cancer 2015, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Jun-Long, H.; Yi, L.; Bao-Lian, Z.; Jia-Si, L.; Ning, Z.; Zhou-Heng, Y.; Xue-Jun, S.; Wen-Wu, L. Necroptosis Signaling Pathways in Stroke: From Mechanisms to Therapies. Curr Neuropharmacol 2018, 16, 1327–1339. [Google Scholar] [CrossRef]
- Sekerdag, E.; Solaroglu, I.; Gursoy-Ozdemir, Y. Cell Death Mechanisms in Stroke and Novel Molecular and Cellular Treatment Options. Current Neuropharmacology 2018, 16, 1396–1415. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Sun, Y.-Y.; Liu, K.-Y. Autophagy and inflammation in ischemic stroke. Neural Regeneration Research 2020, 15, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Nian, K.; Harding, I.C.; Herman, I.M.; Ebong, E.E. Blood-Brain Barrier Damage in Ischemic Stroke and Its Regulation by Endothelial Mechanotransduction. Frontiers in Physiology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Winkler, L.; Blasig, R.; Breitkreuz-Korff, O.; Berndt, P.; Dithmer, S.; Helms, H.C.; Puchkov, D.; Devraj, K.; Kaya, M.; Qin, Z.; et al. Tight junctions in the blood–brain barrier promote edema formation and infarct size in stroke – Ambivalent effects of sealing proteins. Journal of Cerebral Blood Flow & Metabolism 2021, 41, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Nakano-Doi, A.; Sakuma, R.; Matsuyama, T.; Nakagomi, T. Ischemic stroke activates the VE-cadherin promoter and increases VE-cadherin expression in adult mice. Histol Histopathol 2018, 33, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Kalka, C.; Masuda, H.; Takahashi, T.; Gordon, R.; Tepper, O.; Gravereaux, E.; Pieczek, A.; Iwaguro, H.; Hayashi, S.-I.; Isner, J.M.; et al. Vascular Endothelial Growth Factor<sub>165</sub> Gene Transfer Augments Circulating Endothelial Progenitor Cells in Human Subjects. Circulation Research 2000, 86, 1198–1202. [Google Scholar] [CrossRef] [PubMed]
- Ceradini, D.J.; Kulkarni, A.R.; Callaghan, M.J.; Tepper, O.M.; Bastidas, N.; Kleinman, M.E.; Capla, J.M.; Galiano, R.D.; Levine, J.P.; Gurtner, G.C. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nature Medicine 2004, 10, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, W.; Tripathi, D.; Ronaldson, P.T. Blood-brain barrier dysfunction in ischemic stroke: targeting tight junctions and transporters for vascular protection. American Journal of Physiology-Cell Physiology 2018, 315, C343–C356. [Google Scholar] [CrossRef] [PubMed]
- Savitz, S.I.; Yavagal, D.; Rappard, G.; Likosky, W.; Rutledge, N.; Graffagnino, C.; Alderazi, Y.; Elder, J.A.; Chen, P.R.; Budzik, R.F.; et al. A Phase 2 Randomized, Sham-Controlled Trial of Internal Carotid Artery Infusion of Autologous Bone Marrow–Derived ALD-401 Cells in Patients With Recent Stable Ischemic Stroke (RECOVER-Stroke). Circulation 2019, 139, 192–205. [Google Scholar] [CrossRef]
- Abdullah, Z.; Bayraktutan, U. Suppression of PKC-alpha attenuates TNF-alpha-evoked cerebral barrier breakdown via regulations of MMP-2 and plasminogen-plasmin system. Biochim Biophys Acta 2016, 1862, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Asahi, M.; Wang, X.; Mori, T.; Sumii, T.; Jung, J.C.; Moskowitz, M.A.; Fini, M.E.; Lo, E.H. Effects of matrix metalloproteinase-9 gene knock-out on the proteolysis of blood-brain barrier and white matter components after cerebral ischemia. J Neurosci 2001, 21, 7724–7732. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, M.; Takagi, Y.; Aoki, T.; Hayase, M.; Marumo, T.; Gomi, M.; Nishimura, M.; Kataoka, H.; Hashimoto, N.; Nozaki, K. Tissue inhibitor of metalloproteinases protect blood-brain barrier disruption in focal cerebral ischemia. J Cereb Blood Flow Metab 2008, 28, 1674–1685. [Google Scholar] [CrossRef]
- Ya, J.; Kadir, R.R.A.; Bayraktutan, U. Delay of endothelial cell senescence protects cerebral barrier against age-related dysfunction: role of senolytics and senomorphics. Tissue Barriers 2023, 11, 2103353. [Google Scholar] [CrossRef] [PubMed]
- Al Ahmad, A.; Gassmann, M.; Ogunshola, O.O. Maintaining blood-brain barrier integrity: pericytes perform better than astrocytes during prolonged oxygen deprivation. J Cell Physiol 2009, 218, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Nirwane, A.; Yao, Y. Basement membrane and blood-brain barrier. Stroke Vasc Neurol 2019, 4, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y. Basement membrane and stroke. Journal of Cerebral Blood Flow & Metabolism 2019, 39, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Yao, Y. Basement Membrane Changes in Ischemic Stroke. Stroke 2020, 51, 1344–1352. [Google Scholar] [CrossRef]
- KRABBE, C.; ZIMMER, J.; MEYER, M. Neural transdifferentiation of mesenchymal stem cells – a critical review. APMIS 2005, 113, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tang, H.; Zhu, J.; Zhang, J.H. Transplanting Mesenchymal Stem Cells for Treatment of Ischemic Stroke. Cell Transplantation 2018, 27, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Zomer, H.D.; Vidane, A.S.; Goncalves, N.N.; Ambrosio, C.E. Mesenchymal and induced pluripotent stem cells: general insights and clinical perspectives. Stem Cells Cloning 2015, 8, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Croft, A.P.; Przyborski, S.A. Mesenchymal stem cells expressing neural antigens instruct a neurogenic cell fate on neural stem cells. Experimental Neurology 2009, 216, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Lian, Q.; Zhang, Y.; Liang, X.; Gao, F.; Tse, H.-F. Directed Differentiation of Human-Induced Pluripotent Stem Cells to Mesenchymal Stem Cells. In Mesenchymal Stem Cells: Methods and Protocols; Gnecchi, M., Ed.; Springer New York: New York, NY, 2016; pp. 289–298. [Google Scholar]
- Khan, A.A.; Huat, T.J.; Al Mutery, A.; El-Serafi, A.T.; Kacem, H.H.; Abdallah, S.H.; Reza, M.F.; Abdullah, J.M.; Jaafar, H. Significant transcriptomic changes are associated with differentiation of bone marrow-derived mesenchymal stem cells into neural progenitor-like cells in the presence of bFGF and EGF. Cell & Bioscience 2020, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Kruminis-Kaszkiel, E.; Osowski, A.; Bejer-Oleńska, E.; Dziekoński, M.; Wojtkiewicz, J. Differentiation of Human Mesenchymal Stem Cells from Wharton’s Jelly Towards Neural Stem Cells Using a Feasible and Repeatable Protocol. Cells 2020, 9, 739. [Google Scholar] [CrossRef] [PubMed]
- Venkat, P.; Shen, Y.; Chopp, M.; Chen, J. Cell-based and pharmacological neurorestorative therapies for ischemic stroke. Neuropharmacology 2018, 134, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.-C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem Cells International 2020, 2020, 4356359. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Luo, Q.; Qin, J.; Shi, Y.; Yang, L.; Ju, B.; Song, G. Osteopontin Promotes Mesenchymal Stem Cell Migration and Lessens Cell Stiffness via Integrin β1, FAK, and ERK Pathways. Cell Biochemistry and Biophysics 2013, 65, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Gebhart, N.; Richelson, E.; Brott, T.G.; Meschia, J.F.; Zubair, A.C. Mechanism of mesenchymal stem cell-induced neuron recovery and anti-inflammation. Cytotherapy 2014, 16, 1336–1344. [Google Scholar] [CrossRef]
- Tai, L.; Saffery, N.S.; Chin, S.P.; Cheong, S.K. Secretome profile of TNF-alpha-induced human umbilical cord mesenchymal stem cells unveils biological processes relevant to skin wound healing. Regen Med 2023. [Google Scholar] [CrossRef] [PubMed]
- Pankajakshan, D.; Agrawal, D.K. Mesenchymal Stem Cell Paracrine Factors in Vascular Repair and Regeneration. J Biomed Technol Res 2014, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, X.; Jiang, C.; Yu, L.; Wang, M.; Han, W.; Liu, L.; Wang, J. Bone marrow mononuclear cell transplantation promotes therapeutic angiogenesis via upregulation of the VEGF–VEGFR2 signaling pathway in a rat model of vascular dementia. Behavioural Brain Research 2014, 265, 171–180. [Google Scholar] [CrossRef]
- Mu, J.; Bakreen, A.; Juntunen, M.; Korhonen, P.; Oinonen, E.; Cui, L.; Myllyniemi, M.; Zhao, S.; Miettinen, S.; Jolkkonen, J. Combined Adipose Tissue-Derived Mesenchymal Stem Cell Therapy and Rehabilitation in Experimental Stroke. Frontiers in Neurology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ruan, L.; Wang, B.; ZhuGe, Q.; Jin, K. Coupling of neurogenesis and angiogenesis after ischemic stroke. Brain Research 2015, 1623, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.J.; Tsai, S.K.; Hu, B.R.; Liou, D.Y.; Huang, S.L.; Huang, M.C.; Huang, W.C.; Cheng, H.; Huang, S.S. Recovery of neurological function of ischemic stroke by application of conditioned medium of bone marrow mesenchymal stem cells derived from normal and cerebral ischemia rats. J Biomed Sci 2014, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Zang, J.; Sha, M.; Zhang, C.; Ye, J.; Zhang, K.; Gao, J. Senescent hepatocyte secretion of matrix metalloproteinases is regulated by nuclear factor-kappaB signaling. Life Sci 2017, 191, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Saraf, J.; Sarmah, D.; Vats, K.; Kaur, H.; Pravalika, K.; Wanve, M.; Kalia, K.; Borah, A.; Dave, K.R.; Yavagal, D.R.; et al. Intra-arterial stem cell therapy modulates neuronal calcineurin and confers neuroprotection after ischemic stroke. Int J Neurosci 2019, 129, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Esneault, E.; Pacary, E.; Eddi, D.; Freret, T.; Tixier, E.; Toutain, J.; Touzani, O.; Schumann-Bard, P.; Petit, E.; Roussel, S.; et al. Combined Therapeutic Strategy Using Erythropoietin and Mesenchymal Stem Cells Potentiates Neurogenesis after Transient Focal Cerebral Ischemia in Rats. Journal of Cerebral Blood Flow & Metabolism 2008, 28, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Selvasandran, K.; Makhoul, G.; Jaiswal, P.K.; Jurakhan, R.; Li, L.; Ridwan, K.; Cecere, R. A Tumor Necrosis Factor-α and Hypoxia-Induced Secretome Therapy for Myocardial Repair. The Annals of Thoracic Surgery 2018, 105, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Venkat, P.; Zacharek, A.; Landschoot-Ward, J.; Wang, F.; Culmone, L.; Chen, Z.; Chopp, M.; Chen, J. Exosomes derived from bone marrow mesenchymal stem cells harvested from type two diabetes rats promotes neurorestorative effects after stroke in type two diabetes rats. Experimental Neurology 2020, 334, 113456. [Google Scholar] [CrossRef] [PubMed]
- Bhasin, A.; Padma Srivastava, M.V.; Mohanty, S.; Bhatia, R.; Kumaran, S.S.; Bose, S. Stem cell therapy: A clinical trial of stroke. Clinical Neurology and Neurosurgery 2013, 115, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Jaillard, A.; Hommel, M.; Moisan, A.; Zeffiro, T.A.; Favre-Wiki, I.M.; Barbieux-Guillot, M.; Vadot, W.; Marcel, S.; Lamalle, L.; Grand, S.; et al. Autologous Mesenchymal Stem Cells Improve Motor Recovery in Subacute Ischemic Stroke: a Randomized Clinical Trial. Translational Stroke Research 2020, 11, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Honmou, O.; Houkin, K.; Matsunaga, T.; Niitsu, Y.; Ishiai, S.; Onodera, R.; Waxman, S.G.; Kocsis, J.D. Intravenous administration of auto serum-expanded autologous mesenchymal stem cells in stroke. Brain 2011, 134, 1790–1807. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.K.; Kondziolka, D.; Wechsler, L.R.; Lunsford, L.D.; Coburn, M.L.; Billigen, J.B.; Kim, A.S.; Johnson, J.N.; Bates, D.; King, B.; et al. Clinical Outcomes of Transplanted Modified Bone Marrow-Derived Mesenchymal Stem Cells in Stroke: A Phase 1/2a Study. Stroke 2016, 47, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Gupta, V.; Khurana, D.; Sharma, R.R.; Khandelwal, N. Randomized Assessment of the Safety and Efficacy of Intra-Arterial Infusion of Autologous Stem Cells in Subacute Ischemic Stroke. American Journal of Neuroradiology 2018, 39, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xi, H.; Huang, H.; Zhang, F.; Liu, Y.; Chen, D.; Xiao, J. Multiple Cell Transplantation Based on an Intraparenchymal Approach for Patients with Chronic Phase Stroke. Cell Transplantation 2013, 22, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.-Y.; Huang, F.-J.; Zhao, M.; Xie, J.-H.; Shi, J.; Wang, J.; Lin, X.-Z.; Zuo, H.; Wang, Y.-L.; Geng, T.-C. A Two-Year Follow-Up Study of Cotransplantation with Neural Stem/Progenitor Cells and Mesenchymal Stromal Cells in Ischemic Stroke Patients. Cell Transplantation 2014, 23, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Hu, J.; Yan, Q.; Zhu, J.; Zhu, Z.; Chen, Y.; Sun, J.; Zhang, R. Bone marrow-derived mesenchymal stem cells rescue injured H9c2 cells via transferring intact mitochondria through tunneling nanotubes in an in vitro simulated ischemia/reperfusion model. Mol Med Rep 2016, 13, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Tseng, N.; Lambie, S.C.; Huynh, C.Q.; Sanford, B.; Patel, M.; Herson, P.S.; Ormond, D.R. Mitochondrial transfer from mesenchymal stem cells improves neuronal metabolism after oxidant injury in vitro: The role of Miro1. Journal of Cerebral Blood Flow & Metabolism 2021, 41, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ye, G.; Zhang, Y.-L.; He, H.-W.; Yu, B.-Q.; Hong, Y.-M.; You, W.; Li, X. Transfer of mitochondria from mesenchymal stem cells derived from induced pluripotent stem cells attenuates hypoxia-ischemia-induced mitochondrial dysfunction in PC12 cells. Neural Regeneration Research 2020, 15, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Noronha, N.d.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Research & Therapy 2019, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.J.; Wong, R.; Barrington, J.; Tamburrano, S.; Pinteaux, E.; Allan, S.M. Systemic conditioned medium treatment from interleukin-1 primed mesenchymal stem cells promotes recovery after stroke. Stem Cell Research & Therapy 2020, 11, 32. [Google Scholar] [CrossRef]
- Kadir, R.R.A.; Alwjwaj, M.; Bayraktutan, U. Treatment with outgrowth endothelial cells protects cerebral barrier against ischemic injury. Cytotherapy 2022, 24, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Bayraktutan, U. Endothelium, endothelial progenitor cells and stroke. J Neurol Clin Neurosci 2017, 1, 21–22. [Google Scholar]
- Williamson, K.; Stringer, S.E.; Alexander, M.Y. Endothelial progenitor cells enter the aging arena. Front Physiol 2012, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Ya, J.; Bayraktutan, U. Vascular Ageing: Mechanisms, Risk Factors, and Treatment Strategies. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Rakkar, K.; Othman, O.; Sprigg, N.; Bath, P.; Bayraktutan, U. Endothelial progenitor cells, potential biomarkers for diagnosis and prognosis of ischemic stroke: protocol for an observational case-control study. Neural Regeneration Research 2020, 15, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Urbich, C.; Dimmeler, S. Endothelial progenitor cells: characterization and role in vascular biology. Circ Res 2004, 95, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.-j.; Yang, J.; Sun, S.-l.; Zhang, R.; Xu, Y.-m. Endothelial Progenitor Cells’ Classification and Application in Neurological Diseases. Tissue Engineering and Regenerative Medicine 2017, 14, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Bayraktutan, U. Endothelial progenitor cells: Potential novel therapeutics for ischaemic stroke. Pharmacological Research 2019, 144, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Kadir, R.R.A.; Alwjwaj, M.; Bayraktutan, U. Protein kinase C-beta distinctly regulates blood-brain barrier-forming capacity of Brain Microvascular endothelial cells and outgrowth endothelial cells. Metab Brain Dis 2022, 37, 1815–1827. [Google Scholar] [CrossRef] [PubMed]
- Alwjwaj, M.; Kadir, R.R.A.; Bayraktutan, U. Outgrowth endothelial progenitor cells restore cerebral barrier function following ischaemic damage: The impact of NOX2 inhibition. Eur J Neurosci 2022, 55, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Kadir, R.R.A.; Alwjwaj, M.; Rakkar, K.; Othman, O.A.; Sprigg, N.; Bath, P.M.; Bayraktutan, U. Outgrowth Endothelial Cell Conditioned Medium Negates TNF-alpha-Evoked Cerebral Barrier Damage: A Reverse Translational Research to Explore Mechanisms. Stem Cell Rev Rep 2023, 19, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Alwjwaj, M.; Kadir, R.R.A.; Bayraktutan, U. The secretome of endothelial progenitor cells: a potential therapeutic strategy for ischemic stroke. Neural Regen Res 2021, 16, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, Z.; Rakkar, K.; Bath, P.M.; Bayraktutan, U. Inhibition of TNF-alpha protects in vitro brain barrier from ischaemic damage. Mol Cell Neurosci 2015, 69, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Rakkar, K.; Othman, O.A.; Sprigg, N.; Bath, P.M.; Bayraktutan, U. Evaluation of Endothelial Progenitor Cell Characteristics as Clinical Biomarkers for Elderly Patients with Ischaemic Stroke. Stem Cell Rev Rep 2023, 19, 1856–1869. [Google Scholar] [CrossRef] [PubMed]
- Kadir, R.R.A.; Rakkar, K.; Othman, O.A.; Sprigg, N.; Bath, P.M.; Bayraktutan, U. Analysis of endothelial progenitor cell subtypes as clinical biomarkers for elderly patients with ischaemic stroke. Scientific Reports 2023, 13, 21843. [Google Scholar] [CrossRef] [PubMed]
- Aragona, C.O.; Imbalzano, E.; Mamone, F.; Cairo, V.; Lo Gullo, A.; D’Ascola, A.; Sardo, M.A.; Scuruchi, M.; Basile, G.; Saitta, A.; et al. Endothelial Progenitor Cells for Diagnosis and Prognosis in Cardiovascular Disease. Stem Cells International 2016, 2016, 8043792. [Google Scholar] [CrossRef]
- Shmelkov, S.V.; Butler, J.M.; Hooper, A.T.; Hormigo, A.; Kushner, J.; Milde, T.; St Clair, R.; Baljevic, M.; White, I.; Jin, D.K.; et al. CD133 expression is not restricted to stem cells, and both CD133+ and CD133- metastatic colon cancer cells initiate tumors. J Clin Invest 2008, 118, 2111–2120. [Google Scholar] [CrossRef]
- Medina, R.J.; O’Neill, C.L.; Sweeney, M.; Guduric-Fuchs, J.; Gardiner, T.A.; Simpson, D.A.; Stitt, A.W. Molecular analysis of endothelial progenitor cell (EPC) subtypes reveals two distinct cell populations with different identities. BMC Med Genomics 2010, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Abdulkadir, R.R.; Alwjwaj, M.; Othman, O.A.; Rakkar, K.; Bayraktutan, U. Outgrowth endothelial cells form a functional cerebral barrier and restore its integrity after damage. Neural Regeneration Research 2020, 15, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Hristov, M.; Erl, W.; Weber, P.C. Endothelial Progenitor Cells. Arteriosclerosis, Thrombosis, and Vascular Biology 2003, 23, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Luo, C.; Cao, B.; Hu, H.; Wang, S.; Yue, H.; Chen, L.; Zhou, Z. Endothelial Progenitor Cells for Ischemic Stroke: Update on Basic Research and Application. Stem Cells International 2017, 2017, 2193432. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Chen, X. Vascular Endothelial Growth Factor as an Anti-Angiogenic Target for Cancer Therapy. Current Drug Targets 2010, 11, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, M. Vascular Endothelial Growth Factor (VEGF) and Its Receptor (VEGFR) Signaling in Angiogenesis:A Crucial Target for Anti- and Pro-Angiogenic Therapies. Genes & Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, H.; Xu, C.; Deng, M.; Song, M.; Yu, X.; Xu, S.; Zhao, X. VEGF promotes endothelial progenitor cell differentiation and vascular repair through connexin 43. Stem Cell Research & Therapy 2017, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- Hinds, J.J.G.a.M.T. Endothelial Outgrowth Cells: Function and Performance in Vascular Grafts. Tissue Engineering Part B: Reviews 2014, 20, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Ye, J.; Shen, F.; Zhu, Y.; Yeghiazarians, Y.; Zhu, W.; Chen, Y.; Lawton, M.T.; Young, W.L.; Yang, G.-Y. Interleukin-6 Stimulates Circulating Blood-Derived Endothelial Progenitor Cell Angiogenesis in vitro. Journal of Cerebral Blood Flow & Metabolism 2008, 28, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Münzel, T. Endothelial Nitric Oxide Synthase in Vascular Disease. Circulation 2006, 113, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Vodovotz, Y.; Tzeng, E.; Billiar, T.R. Nitric oxide, a protective molecule in the cardiovascular system. Nitric Oxide 2013, 35, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Feliers, D.; Chen, X.; Akis, N.; Choudhury, G.G.; Madaio, M.; Kasinath, B.S. VEGF regulation of endothelial nitric oxide synthase in glomerular endothelial cells. Kidney International 2005, 68, 1648–1659. [Google Scholar] [CrossRef] [PubMed]
- Aicher, A.; Heeschen, C.; Mildner-Rihm, C.; Urbich, C.; Ihling, C.; Technau-Ihling, K.; Zeiher, A.M.; Dimmeler, S. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nature Medicine 2003, 9, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Yao, Y.; Han, R.; Huang, Q.; Li, H.; Wang, B.; Li, S.; Li, M.; Mao, Y.; Mao, X.; et al. Cerebral ischemia induces angiogenesis in the peri-infarct regions via Notch1 signaling activation. Experimental Neurology 2018, 304, 30–40. [Google Scholar] [CrossRef]
- Akil, A.; Gutiérrez-García, A.K.; Guenter, R.; Rose, J.B.; Beck, A.W.; Chen, H.; Ren, B. Notch Signaling in Vascular Endothelial Cells, Angiogenesis, and Tumor Progression: An Update and Prospective. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.J.; Iruela-Arispe, M.L. NOTCH regulation of the endothelial cell phenotype. Current Opinion in Hematology 2018, 25, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, F.; Xia, W.-h.; Zhang, Y.-y.; Xu, S.-y.; Cheng, F.; Liu, X.; Zhang, X.-y.; Wang, S.-m.; Tao, J. CXCR4 gene transfer contributes to in vivo reendothelialization capacity of endothelial progenitor cells. Cardiovascular Research 2010, 88, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.R. Regulation of hematopoiesis. Yale J Biol Med 1990, 63, 371–380. [Google Scholar] [PubMed]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise Review: Evidence for CD34 as a Common Marker for Diverse Progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Ngo, N.; Patel, K.; Isaacson, P.G.; Naresh, K.N. Leucocyte common antigen (CD45) and CD5 positivity in an “undifferentiated” carcinoma: a potential diagnostic pitfall. Journal of Clinical Pathology 2007, 60, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Haspel, L.R.; Miller, B.K. Hematopoietic Stem Cells: Source Matters. Current Stem Cell Research & Therapy 2008, 3, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Britannica, T. Editors of Encyclopaedia. "organ." Encyclopedia Britannica, August 22, 2023. Available online: https://www.britannica.com/science/organ-biology (accessed on 22 October 2023).
- Orkin, S.H.; Zon, L.I. Hematopoiesis: An Evolving Paradigm for Stem Cell Biology. Cell 2008, 132, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Mayani, H. The regulation of hematopoietic stem cell populations [version 1; peer review: 2 approved]. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.P.; Tipping, A.J.; Wang, Z.; Enver, T. GATA-2 mediated regulation of normal hematopoietic stem/progenitor cell function, myelodysplasia and myeloid leukemia. The International Journal of Biochemistry & Cell Biology 2012, 44, 457–460. [Google Scholar] [CrossRef] [PubMed]
- McIver, S.C.; Kang, Y.-A.; DeVilbiss, A.W.; O’Driscoll, C.A.; Ouellette, J.N.; Pope, N.J.; Camprecios, G.; Chang, C.-J.; Yang, D.; Bouhassira, E.E.; et al. The exosome complex establishes a barricade to erythroid maturation. Blood 2014, 124, 2285–2297. [Google Scholar] [CrossRef] [PubMed]
- Laura Gutiérrez, N.C. Luis Fernández-Calleja, Elena Karkoulia, John Strouboulis. Regulation of GATA1 levels in erythropoiesis. IUBMB Life 2020, 72, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, K.; Delaney, C.; Varnum-Finney, B.; Kohn, A.D.; Moon, R.T.; Bernstein, I.D. The Interaction of the Wnt and Notch Pathways Modulates Natural Killer Versus T Cell Differentiation. Stem Cells 2007, 25, 2488–2497. [Google Scholar] [CrossRef] [PubMed]
- Haase, V.H. Regulation of erythropoiesis by hypoxia-inducible factors. Blood Reviews 2013, 27, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Lee, H.-T.; Lee, S.-D.; Lee, W.; Cho, C.-W.C.; Lin, S.-Z.; Wang, H.-J.; Okano, H.; Su, C.-Y.; Yu, Y.-L.; et al. Role of HIF-1α-activated Epac1 on HSC-mediated neuroplasticity in stroke model. Neurobiology of Disease 2013, 58, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Lee, H.T.; Lee, S.D.; Lee, W.; Cho, C.W.; Lin, S.Z.; Wang, H.J.; Okano, H.; Su, C.Y.; Yu, Y.L.; et al. Role of HIF-1alpha-activated Epac1 on HSC-mediated neuroplasticity in stroke model. Neurobiol Dis 2013, 58, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, L.; Jiang, C.; Chen, M.; Ou, C.; Wang, J. Bone marrow mononuclear cells exert long-term neuroprotection in a rat model of ischemic stroke by promoting arteriogenesis and angiogenesis. Brain Behav Immun 2013, 34, 56–66. [Google Scholar] [CrossRef]
- Taguchi, A.; Soma, T.; Tanaka, H.; Kanda, T.; Nishimura, H.; Yoshikawa, H.; Tsukamoto, Y.; Iso, H.; Fujimori, Y.; Stern, D.M.; et al. Administration of CD34+ cells after stroke enhances neurogenesis via angiogenesis in a mouse model. J Clin Invest 2004, 114, 330–338. [Google Scholar] [CrossRef]
- Mathiyalagan, P.; Liang, Y.; Kim, D.; Misener, S.; Thorne, T.; Kamide, C.E.; Klyachko, E.; Losordo, D.W.; Hajjar, R.J.; Sahoo, S. Angiogenic Mechanisms of Human CD34<sup>+</sup> Stem Cell Exosomes in the Repair of Ischemic Hindlimb. Circulation Research 2017, 120, 1466–1476. [Google Scholar] [CrossRef]
- Boy, S.; Sauerbruch, S.; Kraemer, M.; Schormann, T.; Schlachetzki, F.; Schuierer, G.; Luerding, R.; Hennemann, B.; Orso, E.; Dabringhaus, A.; et al. Mobilisation of Hematopoietic CD34+ Precursor Cells in Patients with Acute Stroke Is Safe - Results of an Open-Labeled Non Randomized Phase I/II Trial. PLOS ONE 2011, 6, e23099. [Google Scholar] [CrossRef]
- Banerjee, S.; Bentley, P.; Hamady, M.; Marley, S.; Davis, J.; Shlebak, A.; Nicholls, J.; Williamson, D.A.; Jensen, S.L.; Gordon, M.; et al. Intra-Arterial Immunoselected CD34+ Stem Cells for Acute Ischemic Stroke. Stem Cells Translational Medicine 2014, 3, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Correa, P.L.; Mesquita, C.T.; Felix, R.M.; Azevedo, J.C.; Barbirato, G.B.; Falcão, C.H.; Gonzalez, C.; Mendonça, M.L.; Manfrim, A.; de Freitas, G.; et al. Assessment of Intra-arterial Injected Autologous Bone Marrow Mononuclear Cell Distribution by Radioactive Labeling in Acute Ischemic Stroke. Clinical Nuclear Medicine 2007, 32. [Google Scholar] [CrossRef] [PubMed]
- Savitz, S.I.; Misra, V.; Kasam, M.; Juneja, H.; Cox Jr, C.S.; Alderman, S.; Aisiku, I.; Kar, S.; Gee, A.; Grotta, J.C. Intravenous autologous bone marrow mononuclear cells for ischemic stroke. Annals of Neurology 2011, 70, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Moniche, F.; Gonzalez, A.; Gonzalez-Marcos, J.-R.; Carmona, M.; Piñero, P.; Espigado, I.; Garcia-Solis, D.; Cayuela, A.; Montaner, J.; Boada, C.; et al. Intra-Arterial Bone Marrow Mononuclear Cells in Ischemic Stroke. Stroke 2012, 43, 2242–2244. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Sharma, A.; Garg, A.; Mohanty, S.; Bhatnagar, S.; Johri, S.; Singh, K.K.; Nair, V.; Sarkar, R.S.; Gorthi, S.P.; et al. Intravenous Autologous Bone Marrow Mononuclear Stem Cell Therapy for Ischemic Stroke. Stroke 2014, 45, 3618–3624. [Google Scholar] [CrossRef] [PubMed]
- Sargento-Freitas, J.; Aday, S.; Nunes, C.; Cordeiro, M.; Gouveia, A.; Silva, F.; Machado, C.; Rodrigues, B.; Santo, G.C.; Ferreira, C.; et al. Endothelial progenitor cells enhance blood-brain barrier permeability in subacute stroke. Neurology 2018, 90, e127–e134. [Google Scholar] [CrossRef]
- Torikai, H.; Mi, T.; Gragert, L.; Maiers, M.; Najjar, A.; Ang, S.; Maiti, S.; Dai, J.; Switzer, K.C.; Huls, H.; et al. Genetic editing of HLA expression in hematopoietic stem cells to broaden their human application. Scientific Reports 2016, 6, 21757. [Google Scholar] [CrossRef] [PubMed]
- Kennea, N.L.; Mehmet, H. Neural stem cells. The Journal of Pathology 2002, 197, 536–550. [Google Scholar] [CrossRef]
- Ludwig PE, Reddy V, Varacallo M. Neuroanatomy, Neurons. [Updated 2023 Jul 24]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023 Jan-. Available online: https://www.ncbi.nlm.nih.gov/books/NBK441977/ (accessed on 22 October 2023).
- Purves D, Augustine GJ, Fitzpatrick D, et al., editors. Neuroscience. 2nd edition. Sunderland (MA): Sinauer Associates; 2001. Neuroglial Cells. Available online: https://www.ncbi.nlm.nih.gov/books/NBK10869/ (accessed on 22 October 2023).
- Martínez-Cerdeño, V.; Noctor, S.C. Neural Progenitor Cell Terminology. Frontiers in Neuroanatomy 2018, 12. [Google Scholar] [CrossRef] [PubMed]
- Obernier, K.; Alvarez-Buylla, A. Neural stem cells: origin, heterogeneity and regulation in the adult mammalian brain. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Binda, E.; Filippis, L.D.; Vescovi, A.L. Isolation of Neural Stem Cells from Neural Tissues Using the Neurosphere Technique. Current Protocols in Stem Cell Biology 2010, 15, 2D.6.1–2D.6.18. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, B.A.; Weiss, S. Generation of Neurons and Astrocytes from Isolated Cells of the Adult Mammalian Central Nervous System. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef]
- Louis, S.A.; Mak, C.K.H. Enumerating Stem Cell Frequency: Neural Colony Forming Cell Assay. In Neural Progenitor Cells: Methods and Protocols, Reynolds, B.A., Deleyrolle, L.P., Eds.; Humana Press: Totowa, NJ, 2013; pp. 117–132. [Google Scholar]
- Zhou, Z.-D.; Kumari, U.; Xiao, Z.-C.; Tan, E.-K. Notch as a molecular switch in neural stem cells. IUBMB Life 2010, 62, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Faigle, R.; Song, H. Signaling mechanisms regulating adult neural stem cells and neurogenesis. Biochimica et Biophysica Acta (BBA) - General Subjects 2013, 1830, 2435–2448. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.S.; Santos, A.K.; Vasconcellos, R.; Goulart, V.A.M.; Parreira, R.C.; Kihara, A.H.; Ulrich, H.; Resende, R.R. Neural stem cell differentiation into mature neurons: Mechanisms of regulation and biotechnological applications. Biotechnology Advances 2018, 36, 1946–1970. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Liao, W.; Feng, N.-H.; Lou, Y.-L.; Niu, X.; Zhang, A.-J.; Wang, Y.; Deng, Z.-F. Human induced pluripotent stem cell-derived neural stem cells survive, migrate, differentiate, and improve neurologic function in a rat model of middle cerebral artery occlusion. Stem Cell Research & Therapy 2013, 4, 73. [Google Scholar] [CrossRef] [PubMed]
- Eckert, A.; Huang, L.; Gonzalez, R.; Kim, H.-S.; Hamblin, M.H.; Lee, J.-P. Bystander Effect Fuels Human Induced Pluripotent Stem Cell-Derived Neural Stem Cells to Quickly Attenuate Early Stage Neurological Deficits After Stroke. Stem Cells Translational Medicine 2015, 4, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wong, S.; Snyder, E.Y.; Hamblin, M.H.; Lee, J.-P. Human neural stem cells rapidly ameliorate symptomatic inflammation in early-stage ischemic-reperfusion cerebral injury. Stem Cell Research & Therapy 2014, 5, 129. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Nagai, A.; Sheikh, A.M.; Mitaki, S.; Wakabayashi, K.; Kim, S.U.; Kobayashi, S.; Yamaguchi, S. A human neural stem cell line provides neuroprotection and improves neurological performance by early intervention of neuroinflammatory system. Brain Research 2016, 1631, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Lee, S.H.; Kim, S.U.; Yoon, B.W. Human neural stem cells promote proliferation of endogenous neural stem cells and enhance angiogenesis in ischemic rat brain. Neural Regen Res 2016, 11, 298–304. [Google Scholar] [CrossRef]
- Song, M.; Kim, Y.-J.; Kim, Y.-H.; Roh, J.; Kim, E.-C.; Lee, H.J.; Kim, S.U.; Yoon, B.-W. Long-Term Effects of Magnetically Targeted Ferumoxide-Labeled Human Neural Stem Cells in Focal Cerebral Ischemia. Cell Transplantation 2015, 24, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, S.S.; Seledtsov, V.I.; Banul, N.V.; Poveshchenko, O.V.; Senyukov, V.V.; Astrakov, S.V.; Samarin, D.M.; Taraban, V.Y. Cell therapy of brain stroke. Bulletin of Experimental Biology and Medicine 2005, 139, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Kalladka, D.; Sinden, J.; Pollock, K.; Haig, C.; McLean, J.; Smith, W.; McConnachie, A.; Santosh, C.; Bath, P.M.; Dunn, L.; et al. Human neural stem cells in patients with chronic ischaemic stroke (PISCES): a phase 1, first-in-man study. The Lancet 2016, 388, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Muir, K.W.; Bulters, D.; Willmot, M.; Sprigg, N.; Dixit, A.; Ward, N.; Tyrrell, P.; Majid, A.; Dunn, L.; Bath, P.; et al. Intracerebral implantation of human neural stem cells and motor recovery after stroke: multicentre prospective single-arm study (PISCES-2). Journal of Neurology, Neurosurgery & Psychiatry 2020, 91, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Kondziolka, D.; Wechsler, L.; Goldstein, S.; Meltzer, C.; Thulborn, K.R.; Gebel, J.; Jannetta, P.; DeCesare, S.; Elder, E.M.; McGrogan, M.; et al. Transplantation of cultured human neuronal cells for patients with stroke. Neurology 2000, 55, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Kondziolka, D.; Steinberg, G.K.; Wechsler, L.; Meltzer, C.C.; Elder, E.; Gebel, J.; DeCesare, S.; Jovin, T.; Zafonte, R.; Lebowitz, J.; et al. Neurotransplantation for patients with subcortical motor stroke: a Phase 2 randomized trial. Journal of Neurosurgery 2005, 103, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Moubarik, C.; Guillet, B.; Youssef, B.; Codaccioni, J.-L.; Piercecchi, M.-D.; Sabatier, F.; Lionel, P.; Dou, L.; Foucault-Bertaud, A.; Velly, L.; et al. Transplanted Late Outgrowth Endothelial Progenitor Cells as Cell Therapy Product for Stroke. Stem Cell Reviews and Reports 2011, 7, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, L.; Bennis, Y.; Guillet, B.; Velly, L.; Garrigue, P.; Sabatier, F.; Dignat-George, F.; Bruder, N.; Pisano, P. Therapeutic benefit of a combined strategy using erythropoietin and endothelial progenitor cells after transient focal cerebral ischemia in rats. Neurological Research 2013, 35, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhao, Z.; Wang, C.; Wang, C.X.; Li, P.C.; Qian, C.; Teng, G.J. Bioluminescence imaging of transplanted human endothelial colony-forming cells in an ischemic mouse model. Brain Res 2016, 1642, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.A.G.; Martins, M.P.; Araújo, M.D.; Klamt, C.; Vedolin, L.; Garicochea, B.; Raupp, E.F.; Ammar, J.S.E.; Machado, D.C.; Da Costa, J.C.; et al. Intra-Arterial Infusion of Autologous Bone Marrow Mononuclear Cells in Patients with Moderate to Severe Middle Cerebral Artery Acute Ischemic Stroke. Cell Transplantation 2012, 21, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Laskowitz, D.T.; Bennett, E.R.; Durham, R.J.; Volpi, J.J.; Wiese, J.R.; Frankel, M.; Shpall, E.; Wilson, J.M.; Troy, J.; Kurtzberg, J. Allogeneic Umbilical Cord Blood Infusion for Adults with Ischemic Stroke: Clinical Outcomes from a Phase I Safety Study. Stem Cells Translational Medicine 2018, 7, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Bang, O.Y.; Lee, J.S.; Lee, P.H.; Lee, G. Autologous mesenchymal stem cell transplantation in stroke patients. Annals of Neurology 2005, 57, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.L.; Crawford, J.R.; Dib, N.; Verkh, L.; Tankovich, N.; Cramer, S.C. Phase I/II Study of Safety and Preliminary Efficacy of Intravenous Allogeneic Mesenchymal Stem Cells in Chronic Stroke. Stroke 2019, 50, 2835–2841. [Google Scholar] [CrossRef]
- Fang, J.; Guo, Y.; Tan, S.; Li, Z.; Xie, H.; Chen, P.; Wang, K.; He, Z.; He, P.; Ke, Y.; et al. Autologous Endothelial Progenitor Cells Transplantation for Acute Ischemic Stroke: A 4-Year Follow-Up Study. Stem Cells Transl Med 2019, 8, 14–21. [Google Scholar] [CrossRef]
- Wang, L.; Ji, H.; Li, M.; Zhou, J.; Bai, W.; Zhong, Z.; Li, N.; Zhu, D.; Zhang, Z.; Liu, Y.; et al. Intrathecal Administration of Autologous CD34 Positive Cells in Patients with Past Cerebral Infarction: A Safety Study. ISRN Neurology 2013, 2013, 128591. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhu, W.; Zhu, J.; Wu, L.; Xu, G.; Liu, X. Feasibility of Delivering Mesenchymal Stem Cells via Catheter to the Proximal End of the Lesion Artery in Patients with Stroke in the Territory of the Middle Cerebral Artery. Cell Transplantation 2013, 22, 2291–2298. [Google Scholar] [CrossRef]
- Wechsler, L.R.; Bates, D.; Stroemer, P.; Andrews-Zwilling, Y.S.; Aizman, I. Cell Therapy for Chronic Stroke. Stroke 2018, 49, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.W.; Kinder, H.A.; West, F.D. Neural stem cell therapy for stroke: A multimechanistic approach to restoring neurological function. Brain and Behavior 2019, 9, e01214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-l.; Xie, X.-f.; Xiong, Y.-q.; Liu, S.-m.; Hu, G.-z.; Cao, W.-f.; Wu, X.-m. Comparisons of the therapeutic effects of three different routes of bone marrow mesenchymal stem cell transplantation in cerebral ischemic rats. Brain Research 2018, 1680, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos-dos-Santos, A.; Rosado-de-Castro, P.H.; Lopes de Souza, S.A.; da Costa Silva, J.; Ramos, A.B.; Rodriguez de Freitas, G.; Barbosa da Fonseca, L.M.; Gutfilen, B.; Mendez-Otero, R. Intravenous and intra-arterial administration of bone marrow mononuclear cells after focal cerebral ischemia: Is there a difference in biodistribution and efficacy? Stem Cell Research 2012, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rosado-de-Castro, P.H.; Schmidt Fda, R.; Battistella, V.; Lopes de Souza, S.A.; Gutfilen, B.; Goldenberg, R.C.; Kasai-Brunswick, T.H.; Vairo, L.; Silva, R.M.; Wajnberg, E.; et al. Biodistribution of bone marrow mononuclear cells after intra-arterial or intravenous transplantation in subacute stroke patients. Regen Med 2013, 8, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Migliati, E.; Parsha, K.; Schaar, K.; Xi, X.; Aronowski, J.; Savitz, S.I. Intra-Arterial Delivery Is Not Superior to Intravenous Delivery of Autologous Bone Marrow Mononuclear Cells in Acute Ischemic Stroke. Stroke 2013, 44, 3463–3472. [Google Scholar] [CrossRef] [PubMed]
- Sussman, E.S.; Steinberg, G.K. A Focused Review of Clinical and Preclinical Studies of Cell-Based Therapies in Stroke. Neurosurgery 2017, 64. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.M.; Harting, M.T.; Jimenez, F.; Monzon-Posadas, W.O.; Xue, H.; Savitz, S.I.; Laine, G.A.; Cox, C.S., Jr. Pulmonary passage is a major obstacle for intravenous stem cell delivery: the pulmonary first-pass effect. Stem Cells Dev 2009, 18, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Li, F.; Dong, M.; Liang, W.H.; Lai, W.H.; Ho, W.I.; Wei, R.; Huang, Y.; Liao, S.Y.; Tse, H.F. Repeated intravenous administration of hiPSC-MSCs enhance the efficacy of cell-based therapy in tissue regeneration. Commun Biol 2022, 5, 867. [Google Scholar] [CrossRef] [PubMed]
- Hess, D.C.; Wechsler, L.R.; Clark, W.M.; Savitz, S.I.; Ford, G.A.; Chiu, D.; Yavagal, D.R.; Uchino, K.; Liebeskind, D.S.; Auchus, A.P.; et al. Safety and efficacy of multipotent adult progenitor cells in acute ischaemic stroke (MASTERS): a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Neurol 2017, 16, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Popescu, S.; Preda, M.B.; Marinescu, C.I.; Simionescu, M.; Burlacu, A. Dual Stem Cell Therapy Improves the Myocardial Recovery Post-Infarction through Reciprocal Modulation of Cell Functions. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, T.; Nakano-Doi, A.; Kubo, S.; Sawano, T.; Kuramoto, Y.; Yamahara, K.; Matsuyama, T.; Takagi, T.; Doe, N.; Yoshimura, S. Transplantation of Human Brain-Derived Ischemia-Induced Multipotent Stem Cells Ameliorates Neurological Dysfunction in Mice After Stroke. Stem Cells Transl Med 2023, 12, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Parham, L. Ethical Issues in Stem Cell Research. Endocrine Reviews 2009, 30, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Polson, A.G.; Fuji, R.N. The successes and limitations of preclinical studies in predicting the pharmacodynamics and safety of cell-surface-targeted biological agents in patients. British Journal of Pharmacology 2012, 166, 1600–1602. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Pan, J.; Li, Y.; Jiang, Y.; Zheng, H.; Shi, R.; Zhang, Q.; Liu, C.; Tian, H.; Zhang, Z.; et al. Extracellular vesicles from adipose-derived stem cells promote microglia M2 polarization and neurological recovery in a mouse model of transient middle cerebral artery occlusion. Stem Cell Res Ther 2022, 13, 21. [Google Scholar] [CrossRef]
- Tae-Hoon, L.; Yoon-Seok, L. Transplantation of mouse embryonic stem cell after middle cerebral artery occlusion. Acta Cir Bras 2012, 27, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Sarmah, D.; Kaur, H.; Chaudhary, A.; Mounica, K.L.; Kalia, K.; Borah, A.; Yavagal, D.R.; Bhattacharya, P. Post-stroke Impairment of the Blood-Brain Barrier and Perifocal Vasogenic Edema Is Alleviated by Endovascular Mesenchymal Stem Cell Administration: Modulation of the PKCdelta/MMP9/AQP4-Mediated Pathway. Mol Neurobiol 2022, 59, 2758–2775. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Different sources, routes and time windows for therapeutic stem cells. Table summarises the most commonly used stem cells obtained from adipose tissue, bone marrow, umbilical cord and laboratory reprogramming in clinical studies. Despite availability of different routes for administration, intravenous and intra-arterial routes remain the most preferred routes particularly during the acute phase of stroke.
Figure 2.
Different sources, routes and time windows for therapeutic stem cells. Table summarises the most commonly used stem cells obtained from adipose tissue, bone marrow, umbilical cord and laboratory reprogramming in clinical studies. Despite availability of different routes for administration, intravenous and intra-arterial routes remain the most preferred routes particularly during the acute phase of stroke.
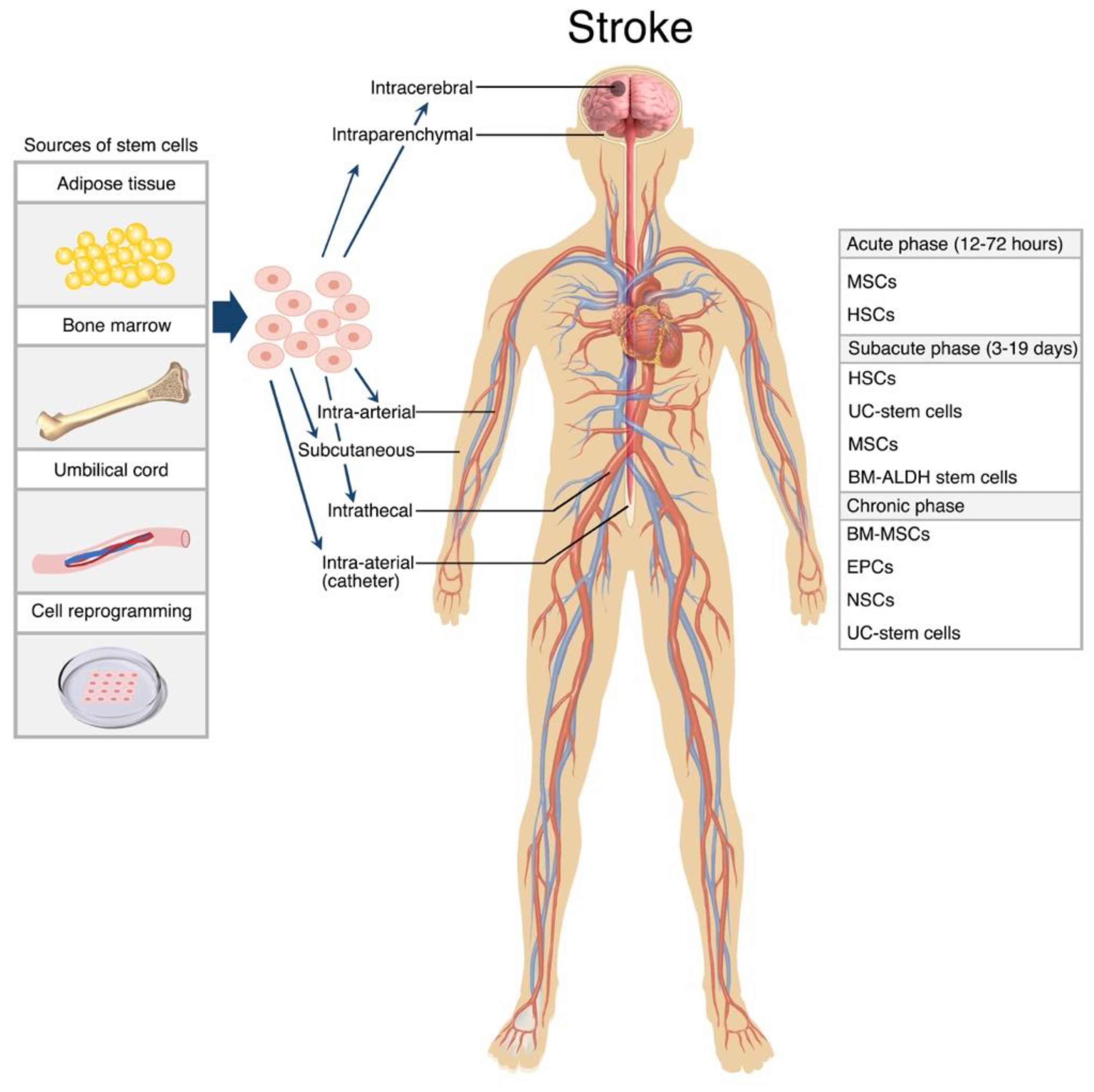
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



