
Submitted:
09 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
Numerous types of oligonucleotide modifications have been developed since automated synthesis of DNA/RNA became a common instrument in the creation of synthetic oligonucleotides. Despite the growing number of types of oligonucleotide modifications under development, only a few of them, and moreover, their combinations have been studied wide enough in terms of their in-fluence on the properties of corresponding NA constructions. In the present study, a number of oligonucleotides with combinations of 3′ end lipophilic (single cholesteryl or pair of dodecyl residues) and phosphate backbone modifications were synthesized. The influence of the combi-nation of used lipophilic groups with phosphate modifications of various nature and different position on the efficiency of cell penetration was evaluated. Obtained results indicate that even a couple of phosphate modifications are able to affect a set of oligonucleotide properties in a complex manner and can remarkably change cellular uptake. These data clearly show that the strategy of using different patterns of modification combinations has great potential for the ra-tional design of oligonucleotide structures with desired predefined properties.
Keywords:
lipophilic oligonucleotides
; phosphate modifications
; phosphoryl guanidine
; triazinyl phosphoramidate
; conjugates
; delivery
1. Introduction
At present, modified oligonucleotides are finding new applications in a broad range of scientific and technological fields including the creation of unique therapeutics and diagnostics [1,2,3,4,5,6,7,8,9,10,11,12]. For example, 18 drugs based on therapeutic nucleic acids (NA) have already been officially approved for use in the treatment of various human diseases, and all of them contain chemical modifications [1]. Modifications in oligonucleotide structure provide the enhanced therapeutic effect of such drugs due to improving some of their most important properties, e.g. efficiency of penetration through the cell membrane [13,14,15,16,17,18]. The absence of a universal solution within the choice of type of oligonucleotide modification for various tasks has led to the intensive development of new NA modifications. Despite the growing number of types of oligonucleotide modifications under development, only a few of them and moreover their combinations were studied wide enough in terms of their diverse influence on the properties of corresponding NA constructions.
Various phosphate modifications could potentially be used to study the complex impact of backbone modifications and their combinations on oligonucleotide properties. One of the most promising approaches to obtain NA derivatives is based on altering the protocol of the phosphite-triester oxidation stage during solid-phase synthesis. For instance, phosphorothioates being the most popular type of phosphate-modified oligonucleotides can be synthesized in this way [19,20,21,22,23]. This modification could be applied for preparing oligonucleotides with even fully modified sugar-phosphate backbone. Phosphorothioate derivatives possess much more stability to different nucleases [19,20] and improved cellular uptake as well as biodistribution due to high affinity to proteins compared to unmodified oligonucleotides [21,22]. Considering the chemical approach of phosphorothioate modification synthesis, there is a crucial drawback since only one representative of this class of altered phosphate group could be obtained using the developed sulfurizing reagents [23] which doesn’t mean to vary functional groups in its structure. Over the past decade, several chemical approaches utilizing alternative oxidation step to obtain novel structures of modified phosphate group which allow one not only to change the nature of phosphate backbone, but also to apply chemistry of its introduction for varying desirable functional moieties, have been proposed [24,25,26,27,28,29,30,31,32,33]. Particularly, we have adopted the method of use of electron-deficient azides in the Staudinger reaction during oxidation step for the efficient obtaining various phosphate derivatives [34,35,36,37,38,39]. We have also demonstrated that different representatives of the developed phosphoryl guanidine and triazinyl phosphoramidate modifications can be applied for synthesis of lipophilic oligonucleotides with enhanced level of intracellular accumulation [37].
The compatibility of newly developed methods for incorporation of phosphate modifications [29,38,39] with standard protocol of automated synthesis, as well as the possibility to combine them with many types of popular phosphoramidite monomers and modifiers, makes them a prospective tool for creating oligonucleotides with complex sugar-phosphate backbones, which have recently come into use. For example, the improvement of cell delivery and other beneficial properties of therapeutic oligonucleotides such as biodistribution, target binding, etc., for antisense [40,41,42,43,44,45,46], siRNA [47,48,49,50,51,52,53] and oligonucleotides with other types of action [54] could be achieved with different combinations of backbone modifications including altered phosphate groups.
One of the recent examples where the positive effect of a combination of modifications has been shown is the number of studies about properties of phosphoryl guanidine (PG) oligonucleotides. Despite early published data observed that introduction of 1,3-dimethylimidazolidin-2-iylidene phosphoramidate (DMI) groups, the most common representative of PG class, into the structure of native oligonucleotide did not lead to detectable increase in level of cellular uptake [55], later it was shown that incorporation of such modifications into phosphorothioate oligonucleotide may result in more than two-fold increasing level of intracellular accumulation [45]. This example demonstrates that combination of different modifications may provide synergistic improvements of desired oligonucleotide properties.
In this study, we have chosen the oligonucleotide containing lipophilic dodecyl residues as the initial NA system with proven significant level of cellular uptake [37], and then introduced in its backbone structure two adjacent DMI groups. Several parameters of the present system were varied in order to evaluate their influence on intracellular accumulation of corresponding oligonucleotides.
2. Results
2.1. Design and synthesis of lipophilic phosphate-modified oligonucleotides
The 5′-fluorescein labeled oligonucleotide with a randomized sequence, which was previously used in intracellular delivery study [37], was chosen as the primary system for evaluation of the impact of DMI groups on the efficiency of intracellular accumulation. This oligonucleotide contains two dodecyl residues introduced through 3′-end internucleotide triazinyl phosphoramidate (TPA) modification (oligonucleotide A, Table 1). The two neighboring DMI modifications were introduced on the opposite to TPA part of sequence (oligonucleotide AX, Table 1). The resulting oligonucleotide has a gapmer-like structure, where phosphate-modified regions of oligonucleotide flank unmodified middle part.
In order to evaluate the influence of several structure-related factors on cell penetration, namely the type of lipophilic modification, the nucleotide sequence, the position and number of DMI groups in the oligonucleotide structure, as well as the nature of phosphate backbone modification, the additional set of oligonucleotides was synthesized (Table 1).
The cholesterol residue being well-known and widely used as the delivery enhancing agent [56,57,58,59,60,61] was selected as the alternative lipophilic group and was introduced applying corresponding commercially available solid support in 3′-end of oligonucleotide (H) and its DMI-containing analog (HX).
To investigate the potential dependence of changes in cell penetration efficiency upon DMI groups introduction in oligonucleotide backbone on nucleotide sequence, the oligonucleotide pair B and BX containing lipophilic TPA modification similar to A and AX pair but with different sequence was synthesized (Table 1). It should be noted that two different, but both randomized sequences without any known cellular target have been chosen consciously to avoid their potential biological activity that may interfere with toxicity or survival of cells and evaluate a distinct effect of oligonucleotide delivery efficiency.
For the determination of the possible effect of the position of DMI groups on cellular uptake, in addition to BX with 5′-end localization of these modifications, oligonucleotides also containing two neighboring DMI groups in the middle part of the sequence (BX′, Table 1) and in 3′-end region directly adjacent to lipophilic TPA modification (BX’’, Table 1) were synthesized. Additionally, for evaluating the impact of varying the number of DMI groups in the structure of oligonucleotide A, a set of derivatives bearing 4, 6, 8 and 10 sequential DMI modifications starting from 5′ end region was obtained (AX2, AX3, AX4, AX5, Table 1).
Besides changing lipophilic part of oligonucleotides, their sequences, position and number of DMI groups, the type of introduced phosphate modifications was also varied. Di(pyrrolidine-1-yl)methylene phosphoramidate, another representative of the phosphoryl guanidine modification class [35], was used as a bulkier and more hydrophobic alternative to the DMI group and was introduced into the corresponding oligonucleotide (BY). Methanesulfonyl phosphoramidate modification being the simplest representative of alkanesulfonyl phosphoramidate modification class [62], which nowadays finds new applications [24,27], as well as common phosphorothioate modification were also used for altering oligonucleotide backbone (BM and BS, respectively, Table 1). It should be noted that all types of considered PN-containing modifications (TPA, DMI, di(pyrrolidine-1-yl)methylene phosphoramidate and methanesulfonyl phosphoramidate) were introduced in oligonucleotide backbone using a single approach based on the Staudinger reaction involving electron-deficient azides during alternative oxidation step. Thus, Staudinger reaction in solid phase phosphoramidite DNA/RNA synthesis could be considered as a single platform for creating wide range of phosphate-modified oligonucleotides.
All oligonucleotides were purified using reversed-phase HPLC and identified using ESI mass spectrometry. The HPLC profiles of the reaction mixtures of the modified oligonucleotides (Figure S1), the data of the mass spectrometry analysis (Figure S2), and the results of the thermal denaturation experiment of the studied oligonucleotides with a complementary unmodified oligonucleotide (Table S1) are provided in “Supplementary Materials”. To sum up, it was shown that the applied methods for synthesis of lipophilic phosphate-modified oligonucleotides possess high efficiency, allowing different types of phosphate modifications to be combined.
2.2. Dynamic light scattering (DLS) analysis of supramolecular complexes of lipophilic phosphate-modified oligonucleotides
Previously, the possibility of forming various supramolecular complexes with dodecyl-containing oligonucleotides was demonstrated [63]. Taking this into consideration, we decided to characterize solution of obtained oligonucleotides by DLS measurements under conditions equal to those of transfection [37] as the most relevant for identification and correct interpretation of the influence of various parameters of oligonucleotide system on cellular uptake. It was shown that in selected conditions (serum-free DMEM medium, 2 h, 10 measurements) oligonucleotide A containing a lipophilic TPA modification can form stable supramolecular complexes. The obtained data reliably differ from those for DMEM or DMEM with control unmodified oligonucleotide (Figure S3). All synthetized lipophilic oligonucleotides were also analyzed by DLS method in order to evaluate influence of introduced phosphate modifications on possibility of formation supramolecular complexes over the same experiment conditions (Figure 1).
The results of DLS experiment showed that oligonucleotides A and B both form stable supramolecular complexes with hydrodynamic diameters of about 10 nm. Moreover, the introduction of DMI groups in 5′ end region (AX and BX) didn’t lead to any significant changes in complex formation (Figure 1). The oligonucleotides H and HX bearing cholesterol moiety instead of dodecyl-containing TPA modification were shown to form number of irregular or unstable complexes with a wide range of hydrodynamic diameters, independent of the presence of DMI modifications. Consecutive changing of the position of pair of neighboring DMI groups from 5′ to 3′ end region of oligonucleotide structure (BX, BX’ and BX” respectively) led to sequential decrease in the efficiency of complex formations. However, sequential increase in the number of introduced DMI groups in the same direction (5′ → 3′ end region) did not result in any regularities within complexes formation. Thus, oligonucleotide AX2, containing four DMI modifications forms stable complexes with hydrodynamic diameters more than 100 nm, at the same time oligonucleotides AX3 (six DMI groups) and AX5 (ten DMI groups) assemble in complexes similar to those formed by oligonucleotide AX (two DMI groups). Oligonucleotide AX4 (eight DMI groups) is capable of forming both types of complexes with hydrodynamic diameters of 10 and more than 100 nm. Interestingly, the nature of even two introduced phosphate modifications (oligonucleotides BX, BY, BM and BS) also can influence the formation of supramolecular structures. Thus, the introduction of DMI (BX) or phosphorothioate (BS) modifications in oligonucleotide backbone didn’t lead to significant changes in complex formation, whereas the incorporation of bulkier representative of the phosphoryl guanidine class (BY) as well as methanesulfonyl phosphoramidate modifications (BM) resulted in formation of less stable complexes. All of the abovementioned points out unobvious manner of influence of modifications and their combinations on the properties of the resulting oligonucleotide structures.
2.3. Study of the efficiency of intracellular accumulation of phosphate-modified oligonucleotides by flow cytometry and confocal microscopy
For quantitative assessment of the efficiency of intracellular accumulation of oligonucleotides with different number of DMI groups transfection was carried out for 4 hours in a serum-free DMEM medium with 1 or 5 μM concentration of oligonucleotides on the HEK293T cell line, which proved to be a convenient model to study the cellular uptake of NA derivatives [34,36,37,64,65]. Transfected cells were analyzed by flow cytometry (Figure 2). To evaluate the transfection efficiency, two parameters were used: the percentage of fluorescent cells (Figure 2A) and the relative fluorescence intensity (Figure 2B). The untreated cells served as a negative control. All of the modified oligonucleotides are accumulated in almost 100% of the cells under the experiment conditions, and hence their mean fluorescent intensity can be compared directly.
One of the most unexpected result of the experiment consists in the manner of cellular uptake of oligonucleotides bearing several DMI groups. Initial introduction of DMI groups in 5′ end region (Figure 2, oligonucleotides A and AX) led to significant rise in intracellular accumulation level. However, sequentially increasing number of DMI groups resulted in gradually decreases uptake efficiency of corresponding multiple modified oligonucleotide derivatives, despite the growth of the overall hydrophobicity (Figure 2, AX – AX5).
Taking into account significant differences in a sugar-phosphate backbone of oligonucleotides with various number of DMI groups as well as their differed cellular uptake levels, we have decided to compare kinetics of intracellular accumulation of oligonucleotides with two (AX) and ten (AX5) DMI groups.
Transfection was carried out for 1, 2, 4, 8 and 24 hours in a serum-free medium or complete DMEM using 5 μM concentration of oligonucleotides on the HEK293T cell line (Figure 3A). It was shown that in early time points, e.g. prior to 8 hours, oligonucleotide AX accumulates more efficiently comparing to AX5 in both types of medium. However, at point of 24 hours the intracellular accumulation levels of AX and AX5 were nearly equal in serum-free transfection conditions, and in the presence of serum AX5 demonstrated even higher level of accumulation.
To investigate the ability of studied oligonucleotides to stay inside the cells, it was decided to evaluate the intracellular oligonucleotide quantity level over time after 4 hours of transfection with subsequent removing the oligonucleotide solutions and medium changing (Figure 3B). It was shown that at point of 4h after the end of transfection derivatives AX and AX5 demonstrated nearly equal level and at point of 20h AX5 possessed higher level of intracellular presence in both type of medium. It can be concluded that AX5 accumulates slower but persists longer than AX once inside of the cells.
The presented results of confocal microscopy indicate that intercellular distribution for oligonucleotide AX didn’t significantly differ from this previously shown for initial oligonucleotide system (A) [37]: FAM-labeled oligonucleotides were distributed throughout the cytoplasm in both disassembled and aggregated forms without attaching to the inner side of the cell membrane, no co-localization with nuclei was revealed (Figure 3C, AX). Distribution of oligonucleotide AX5 was similar to that for AX, but with prevail of disassembled form (Figure 3C, AX5), despite both of them are capable of forming supramolecular complexes with the same hydrodynamic diameter under the transfection conditions (Figure 1, AX and AX5).
Efficiency of intracellular accumulation of other lipophilic derivatives were analyzed by flow cytometry in a serum-free DMEM medium for 4 h of incubation and 5 μM concentration of oligonucleotides. Unmodified oligonucleotide (C) was analyzed both individually as a negative control and with the commercially available delivery agent Lipofectamine 2000 (LF) as a positive control (Figure 4).
Analysis of the flow cytometry data showed that the other studied factors besides the number of DMI groups (Figure 4) are able to remarkably influence the accumulation efficiency of the investigated lipophilic phosphate-modified oligonucleotides. Dodecyl- and cholesterol-containing conjugates (Figure 4B, oligonucleotides A, B and H) have an increased cellular uptake, as it was shown previously [34,36,37,58]. The contribution of not only the different nature of the lipophilic part (A vs. H) but also of the nucleotide sequence (A vs. B) to the delivery efficiency was observed with fluorescence signal differences up to several times (Figure 4B).
One of the important findings is a remarkable increase in intracellular accumulation in all studied oligonucleotides upon introduction of phosphate modifications in the 5′ end region, regardless of the nucleotide sequence (A/AX vs. B/BX), the lipophilic part of the conjugate (A/AX and H/HX) or the nature of phosphate modification (B/BX/BY/BM/BS) (Figure 4B,D). Despite the relatively small contribution of phosphate modifications in total hydrophobicity of oligonucleotides containing the 3′ end region lipophilic TPA modification, adding just two groups was shown to significantly improve their delivery efficiency. It should be noted that the contrast in observed effect diminishes with introduction of DMI groups in a cholesterol-containing oligonucleotide (Figure 4B, A/AX vs. H/HX,). Another important finding is that the efficiency of delivery is also strongly influenced by relative position of DMI modifications and the lipophilic TPA group in oligonucleotide sequence (oligonucleotides BX, BX’ and BX”): as these groups gradually converge, the level of intracellular accumulation decreases (Figure 4B).
In addition to cellular uptake studies by flow cytometry, the resulting lipophilic derivatives containing TPA group and different phosphate backbone modifications (B/BX/BS/BM/BY) were analyzed by confocal microscopy in order to reliably determine accumulation within HEK293T cells. Transfection was carried out under incubation conditions for 4 hours in serum-free medium (DMEM) and an oligonucleotide concentration of 5 µM (Figure 5).
Confocal microscopy demonstrated that intracellular distribution of lipophilic B oligonucleotide was characterized by mostly diffuse disposition in the cytoplasm with a small amount of aggregated forms and without co-localization with nuclei (Figure 5B). Introduction of two phosphate modifications of various types (BX, BY, BM, BS) in the 5′-end of the oligonucleotide resulted in significant increase in the number of aggregated oligonucleotide forms. Among studied lipophilic phosphate-modified oligonucleotides, BM oligonucleotide was characterized by more efficient accumulation but in more co-localized aggregated form compared to other studied derivatives including AX5 (Figure 5, BM, Figure 3C, AX5).
3. Discussion
The use of different combinations of modifications in the design of synthetic oligonucleotide structures has become a popular trend in recent years. Modern oligonucleotide therapeutic candidates contain not only multiple modifications of the same type but also various combinations of modifications. The design principles of such complex modified oligonucleotides are becoming the basis for the creation of new generation of antisense and siRNA agents.
Phosphate modifications represent a versatile tool for creating modified oligonucleotides. One of the most convenient approaches for obtaining phosphate-modified oligonucleotides is based on the alternative oxidation step in protocol of standard solid-phase phosphoramidite method of synthesis. For example, phosphorothioate modifications, which can be obtained by this approach, are widely used in creating various NA constructions including complex oligonucleotide structures with combinations of different modifications [1,2,4,7,45]. Previously we developed another way of alternative oxidation step realization, based on the application of the Staudinger reaction involving electron-deficient azides. It allows one to introduce a wide range of representatives of different classes of phosphate modifications into the obtained sequences without significant changes in the protocols of oligonucleotide synthesis. Moreover, the approach is suitable for introducing various combination of phosphate modifications using a single platform to obtain complex modified structures, which was demonstrated in current study.
In the present work, dodecyl-containing triazinyl phosphoramidate was used as the lipophilic modification providing enhanced cellular accumulation and combined with other types of phosphate modifications in a backbone of synthetized oligonucleotides applying the developed single platform based on the Staudinger reaction. One of the important findings was two-fold increase of cellular uptake upon the introduction of two DMI groups at 5′ end region of oligonucleotide structure (A to AX), despite only a slight increase in overall hydrophobicity. Subsequently, additional parameters of studied system were examined for their influence on intracellular accumulation.
The introduction of DMI groups was found to have a positive effect regardless of the oligonucleotide sequence (A to AX and B to BX). At the same time, replacement of the lipophilic part with cholesterol residue led to the preservation of the positive effect upon introduction of DMI group, however it became less pronounced (A to AX and H to HX). Changing the position of DMI groups in the oligonucleotide structure had a major impact on the studied properties: gradual convergence of DMI and lipophilic moiety position led to decreased cellular uptake as well as to lowered supramolecular complex stability (BX, BX’ and BX”).
Influence of the nature and number of phosphate modifications on the properties of NA-constructions turned out to be the least predictable. For example, oligonucleotide BY was shown to have a lower transfection efficiency than BX, even though the phosphoryl guanidine modification in BY structure possesses more hydrophobicity than BX. The gradual increase of the number of introduced DMI groups in dodecyl-containing oligonucleotide (AX to AX5) has also resulted in lowered levels of cellular uptake, despite the growth of overall hydrophobicity, but enhance the retention rate of oligonucleotides inside the cells. The possibility of forming supramolecular complexes also differs for oligonucleotides BX and BY in non-obvious way as well as for derivatives with the other types of altered phosphate (BM, BS) or different number of introduced DMI groups (AX to AX5).
The obtained results demonstrate the complex and unobvious influence of the phosphate modifications in oligonucleotide backbone on the properties of the synthesized lipophilic derivatives. Revealing such effects and their systematization is important from the viewpoint of rational design of oligonucleotides containing combinations of modifications and indicates the relevance of the search for various effective modification patterns.
4. Materials and Methods
4.1. Oligonucleotide synthesis
The standard phosphoramidite solid-phase synthesis of all modified and unmodified oligonucleotides containing phosphodiester linkages was carried out on an ASM-800 DNA/RNA synthesizer (Biosset, Novosibirsk, Russia). Oligonucleotides were synthesized at the 0.4 µmol scale, using standard commercial 2-cyanoethyl deoxynucleoside phosphoramidites and CPG solid supports (Glen Research, San Diego, CA, USA).
The insertion of triazinyl phoshporamidate (TPA) modification bearing two dodecyl residues in appropriate oligonucleotide structures using triazine modifier during the modified protocol of oxidation step was performed as described in [34].
The insertion of DMI modification in appropriate oligonucleotide structures during the modified oxidation step was performed using commercial 2-azido-1,3-dimethylimidazolidinium hexafluorophosphate (TCI, Tokyo, Japan) for obtaining 1,3-dimethylimidazolidin-2-iylidene phosphoramidate structure (DMI) as described in [38].
The synthesis of pyrrolidine phosphoryl guanidine modifier (azidodipyrrolidinocarbenium hexafluorophosphate) was performed as described in [35].
The synthesis of methanesulfonyl azide was performed as described in [66].
For introduction of methanesulfonyl phoshoramidate (oligonucleotide BM) and pyrrolidine-containing phosphoryl guanidine (oligonucleotide BY) modifications, the following procedure was performed. The corresponding monomer was coupled according to a special protocol including standard coupling step followed by the oxidation step. At this step the Staudinger reaction was performed. Solution of the corresponding azide in acetonitrile was pumped through a column portionwise. Next, the capping and deblocking stages were performed, and all the following procedures were performed according to the standard protocol of automatic solid-phase phosphoramidite synthesis.
Conditions of the Staudinger reaction were the following: for the methanesulfonyl phoshoramidate group obtaining (oligonucleotide BM), 0.25 M solution of methanesulfonyl azide in acetonitrile was used, reaction was proceeding for 1 h; for the pyrrolidine-containing phosphoryl guanidine group obtaining (oligonucleotide BY), 0.25 M solution of azidodipyrrolidinocarbenium hexafluorophosphate in acetonitrile was used, reaction was proceeding for 1 h.
The insertion of phosphorothioate modification (PS) in appropriate oligonucleotide structures was performed using 3-((dimethylaminomethylidene)amino)-3H-1,2,4-dithiazole-3-thione (DDTT, Sulfurizing Reagent II) from Glen Research (San Diego, CA, USA) according to the manufacturer’s protocol.
For the introduction of cholesterol moiety at 3′ end of the oligonucleotide sequence, the corresponding CPG (Primetech, Minsk, Belarus) was used according to manufacturer’s protocol.
For the introduction of the 6-carboxyfluoresceine (FAM) residue at the 5′ end of the oligonucleotide sequence, the corresponding phosphoramidite (Lumiprobe, Moscow, Russia) was used according to the manufacturer’s protocols. The CPG cleavage of FAM-containing oligonucleotides was first performed in aqueous ammonia (30% m/v, 15 min, 55°C), and then aqueous methylamine (40% m/v, 15 min, 55°C or 1 day, 25°C for BS oligonucleotide) was added equally to complete the process.
4.2. Oligonucleotide Purification and Identification
A quality assessment of oligonucleotide synthesis was performed through reverse phase HPLC analysis on a Millichrom A02 system using a ProntoSIL-120-5-C18 column 2 × 75 mm (Econova, Novosibirsk, Russia) in a linear gradient of acetonitrile 0–50% or 0–90% in 20 mM triethylammonium acetate, pH 7.0, at a flow rate of 200 µL/min, and with detection on 260, 280, and 300 nm wavelengths.
Modified oligonucleotides were isolated through reversed-phase HPLC on an Agilent 1200 HPLC system using a Zorbax SB-C18 5 mm column 4.6 × 150 mm (Santa Clara, CA, USA) in a linear gradient of acetonitrile 0–50% or 0–90% in 20 mM triethylammonium acetate, pH 7.0, at a flow rate of 2 mL/min, and with detection on 260, 280, 300 and 500 nm wavelengths. Fractions containing the desired product were collected, evaporated several times with a 1:1 EtOH/H2O mixture, concentrated in vacuo, and precipitated by 1 mL of 2% LiClO4 in acetone. After centrifugation at 14,500 rpm for 2 min, washing with acetone, and drying in air for 20 min at 40°C, the oligonucleotide precipitates were dissolved in 0.1 mL of deionized water and stored at −20°C.
Molecular masses of oligonucleotides were confirmed by LC-MS/MS ESI-MS on an Agilent G6410A mass spectrometer (Santa Clara, CA, USA) in a negative-ion detection mode. The samples were prepared by dissolving oligonucleotides in 20 mM triethylammonium acetate in 60% aq. Acetonitrile at a concentration of 0.1 mM in 10 µL of the sample. Analysis was carried out using 80% aq. Acetonitrile as an eluent at a flow rate of 0.1 mL/min and using standard device settings. Molecular masses were calculated from the experimental m/z values obtained for each sample.
4.3. Characterization of modified oligonucleotides by DLS
The size distributions of supramolecular complexes of modified oligonucleotides were determined by dynamic light scattering technique using a Zetasizer Nano-ZS (Malvern Panalytical Ltd., Malvern, UK) at 25°C. The oligonucleotides were dissolved in DMEM medium (SigmaAldrich, St. Louis, MO, USA) to 5 μM concentration and then analysis of the size was conducted for 2 hours with a period of 10 minutes between measurements.
4.4. Analysis of Intracellular Accumulation of Oligonucleotides
HEK293T cells were seeded at a density 75 000 – 150 000 cells in complete DMEM medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (HyClone, GE Healthcare, Chicago, IL, USA) and 1% antibiotic/antimycotic solution (MP Biomedicals, Santa Ana, CA, USA)) in wells of a 24-well plates and incubated under standard conditions (37°C, humidified atmosphere containing 5% CO2) overnight for cell attachment. The next day, the culture medium was replaced with a serum- and antibiotic-free DMEM medium supplemented with 1 or 5 µM of the lipophilic phosphate-modified oligonucleotides, and the cells were transfected for 4 h in standard conditions. Cells were washed with PBS, trypsinized, quenched with complete DMEM medium, pelleted, washed in PBS, and fixed in 2% formaldehyde in PBS. The intracellular accumulation of the oligonucleotides was examined through flow cytometry using a NovoCyte flow cytometer (ACEA Biosciences, Santa Clara, CA, USA). Flow cytometry data were processed with NovoExpress software v. 1.1.0 (ACEA Biosciences, Santa Clara, CA, USA). All experiments were run in triplicate for statistical analysis. Transfection efficiency was characterized by the percentage of fluorescent-positive cells and relative fluorescence intensity cells in a sample.
In the kinetics experiment, HEK293T cells were transfected with 5 µM of oligonucleotides in the absence (–FBS) or the presence (+FBS) of 10% FBS in culture medium. Analysis of cell uptake of oligonucleotides was performed 1, 2, 4, 8 and 24 h after start of transfection. In the wash-out regimen, transfection of HEK293T cells with oligonucleotides was performed in –FBS or +FBS conditions for 4 h followed by refreshing with complete DMEM medium and incubated under standard conditions until analysis (8 and 24 h after start of transfection).
4.5. Confocal microscopy
HEK293T cells were seeded on coverslips (Marienfeld, Lauda-Königshofen, Germany) in 24-well plates at a density of 150 000 cells/well in complete DMEM medium and incubated overnight under SC to adhere. The next day, HEK293T cells were incubated in serum- and antibiotics-free DMEM medium in the presence of 5 µM of the respective lipophilic phosphate-modified oligonucleotides under SC for 4 h. After incubation, coverslips with cells were washed twice with PBS, fixed with 4% formaldehyde in PBS buffer (15 min, 37°C), and washed twice with PBS. The cells were stained with DAPI solution (1:100 in PBS, Thermo Fisher Scientific, Waltham, MA, USA) at room temperature in the dark for 10 min. After staining, the cells were washed twice with PBS and placed on slides in a drop of Fluoromount-G® (SouthernBiotech, Birmingham, USA). Mounted samples were allowed to cure on a flat, dry surface for 18–24 h at room temperature in the dark. Intracellular localization of lipophilic phosphate-modified oligonucleotides was assessed by confocal fluorescent microscopy on an LSM710 (Zeiss, Oberkochen, Germany) using a Plan-Apochromat 63x/1.40 Oil DIC M27 objective. Analysis of intracellular accumulation of lipophilic phosphate-modified oligonucleotides was performed using ZEN software (Zeiss, Oberkochen, Germany). Confocal microscopic analysis was performed in three channels (blue, green, brightfield). Fluorescence in the blue channel corresponded to DAPI (nuclei staining); the green channel corresponded to fluorescence of lipophilic phosphate-modified oligonucleotides labeled with FAM, and the brightfierld channel corresponded cell morphology visualization.
4.6. Statistical Analysis
Data were statistically processed using one-way ANOVA with Tukey post hoc testing. P < 0.05 was considered to be significant. GraphPad Prism v. 8.0.1 software (GraphPad Software Inc., San Diego, CA, USA) was used for statistical analysis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.S.K. and T.D.Z.; methodology, S.N.K. and O.V.M.; formal analysis, O.V.M. and S.A.Z.; investigation, S.A.Z., O.V.M., and T.D.Z.; writing—original draft preparation, M.S.K., T.D.Z. and S.A.Z.; writing—review and editing, S.N.K. and O.V.M.; supervision, M.S.K. and S.N.K.; funding acquisition, S.N.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant from the Russian Science Foundation 19-14-00204 and the Russian State-funded project for the Institute of Chemical Biology and Fundamental Medicine, Siberian Branch of the Russian Academy of Sciences (grant number 121031300042-1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data can be easily obtained and linked in the respective sections.
Acknowledgments
The authors gratefully acknowledge Albina V. Vladimirova (Institute of Chemical Biology and Fundamental Medicine SB RAS) for cell maintenance. Cell line HEK293Т was obtained from the Russian Cell Culture Collection (RCCC) Institute of Cytology RAS (St. Petersburg, Russia).
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Egli, M. and Manoharan, M. Chemistry, structure and function of approved oligonucleotide therapeutics. Nucleic Acids Res 2023, 51, 2529-2573. [CrossRef]
- Hall, J. Future directions for medicinal chemistry in the field of oligonucleotide therapeutics. RNA 2023, 29, 423-433. doi: 10.1261/rna.079511.122.
- Thakur, S.; Sinhari, A.; Jain, P.; Jadhav, H. R., A perspective on oligonucleotide therapy: Approaches to patient customization. Front Pharmacol 2022, 13, 1006304. [CrossRef]
- Crooke, S. T.; Witztum, J. L.; Bennett, C. F.; Baker, B. F., RNA-Targeted Therapeutics. Cell Metab 2018, 27 (4), 714-739. [CrossRef]
- Helm, J.; Schols, L.; Hauser, S., Towards Personalized Allele-Specific Antisense Oligonucleotide Therapies for Toxic Gain-of-Function Neurodegenerative Diseases. Pharmaceutics 2022, 14 (8). [CrossRef]
- Moumne, L.; Marie, A. C.; Crouvezier, N., Oligonucleotide Therapeutics: From Discovery and Development to Patentability. Pharmaceutics 2022, 14 (2). [CrossRef]
- Rinaldi, C.; Wood, M. J. A., Antisense oligonucleotides: the next frontier for treatment of neurological disorders. Nat Rev Neurol 2018, 14 (1), 9-21. [CrossRef]
- Roberts, T. C.; Langer, R.; Wood, M. J. A., Advances in oligonucleotide drug delivery. Nat Rev Drug Discov 2020, 19 (10), 673-694. [CrossRef]
- Smith, C. I. E.; Zain, R., Therapeutic Oligonucleotides: State of the Art. Annu Rev Pharmacol Toxicol 2019, 59, 605-630. [CrossRef]
- Benizri, S.; Gissot, A.; Martin, A.; Vialet, B.; Grinstaff, M. W.; Barthelemy, P., Bioconjugated Oligonucleotides: Recent Developments and Therapeutic Applications. Bioconjug Chem 2019, 30 (2), 366-383. [CrossRef]
- Quemener, A. M.; Centomo, M. L.; Sax, S. L.; Panella, R., Small Drugs, Huge Impact: The Extraordinary Impact of Antisense Oligonucleotides in Research and Drug Development. Molecules 2022, 27 (2). [CrossRef]
- Crooke, S. T.; Baker, B. F.; Crooke, R. M.; Liang, X. H., Antisense technology: an overview and prospectus. Nat Rev Drug Discov 2021, 20 (6), 427-453. [CrossRef]
- Østergaard, M.E., Jackson, M., Low, A., A, E.C., R, G.L., Peralta, R.Q., Yu, J., Kinberger, G.A., Dan, A., Carty, R. et al. Conjugation of hydrophobic moieties enhances potency of antisense oligonucleotides in the muscle of rodents and non-human primates. Nucleic Acids Res 2019, 47, 6045-6058. [CrossRef]
- Pollak, A.J., Zhao, L. and Crooke, S.T. Systematic Analysis of Chemical Modifications of Phosphorothioate Antisense Oligonucleotides that Modulate Their Innate Immune Response. Nucleic Acid Ther 2023, 33, 95-107. [CrossRef]
- Wolfrum, C., Shi, S., Jayaprakash, K.N., Jayaraman, M., Wang, G., Pandey, R.K., Rajeev, K.G., Nakayama, T., Charrise, K., Ndungo, E.M. et al. Mechanisms and optimization of in vivo delivery of lipophilic siRNAs. Nat Biotechnol 2007, 25, 1149-1157. [CrossRef]
- Osborn, M.F., Coles, A.H., Biscans, A., Haraszti, R.A., Roux, L., Davis, S., Ly, S., Echeverria, D., Hassler, M.R., Godinho, B. et al. Hydrophobicity drives the systemic distribution of lipid-conjugated siRNAs via lipid transport pathways. Nucleic Acids Res 2019, 47, 1070-1081. [CrossRef]
- Biscans, A., Coles, A., Haraszti, R., Echeverria, D., Hassler, M., Osborn, M. and Khvorova, A. Diverse lipid conjugates for functional extra-hepatic siRNA delivery in vivo. Nucleic Acids Res 2019, 47, 1082-1096. [CrossRef]
- Raouane, M., Desmaele, D., Urbinati, G., Massaad-Massade, L. and Couvreur, P. Lipid conjugated oligonucleotides: a useful strategy for delivery. Bioconjug Chem 2012, 23, 1091-1104. [CrossRef]
- Eckstein, F., Phosphorothioates, essential components of therapeutic oligonucleotides. Nucleic Acid Ther 2014, 24 (6), 374-87. [CrossRef]
- Kawasaki, A. M.; Casper, M. D.; Freier, S. M.; Lesnik, E. A.; Zounes, M. C.; Cummins, L. L.; Gonzalez, C.; Cook, P. D., Uniformly modified 2'-deoxy-2'-fluoro phosphorothioate oligonucleotides as nuclease-resistant antisense compounds with high affinity and specificity for RNA targets. J Med Chem 1993, 36 (7), 831-41. [CrossRef]
- Crooke, S. T.; Seth, P. P.; Vickers, T. A.; Liang, X. H., The Interaction of Phosphorothioate-Containing RNA Targeted Drugs with Proteins Is a Critical Determinant of the Therapeutic Effects of These Agents. J Am Chem Soc 2020, 142 (35), 14754-14771. [CrossRef]
- Crooke, S. T.; Vickers, T. A.; Liang, X. H., Phosphorothioate modified oligonucleotide-protein interactions. Nucleic Acids Res 2020, 48 (10), 5235-5253. [CrossRef]
- Guzaev, A. P, Reactivity of 3H-1,2,4-dithiazole-3-thiones and 3H-1,2-dithiole-3-thiones as sulfurizing agents for oligonucleotide synthesis. Tetrahedron Letters 2011, 52 (3), 434–437. [CrossRef]
- Miroshnichenko, S. K.; Patutina, O. A.; Burakova, E. A.; Chelobanov, B. P.; Fokina, A. A.; Vlassov, V. V.; Altman, S.; Zenkova, M. A.; Stetsenko, D. A., Mesyl phosphoramidate antisense oligonucleotides as an alternative to phosphorothioates with improved biochemical and biological properties. Proc Natl Acad Sci U S A 2019, 116 (4), 1229-1234. [CrossRef]
- Santorelli, A.; Gothelf, K. V., Conjugation of chemical handles and functional moieties to DNA during solid phase synthesis with sulfonyl azides. Nucleic Acids Res 2022, 50 (13), 7235-7246. [CrossRef]
- Hansen, R. A.; Marcher, A.; Pedersen, K. N.; Gothelf, K. V., Insertion of Chemical Handles into the Backbone of DNA during Solid-Phase Synthesis by Oxidative Coupling of Amines to Phosphites. Angew Chem Int Ed Engl 2023, 62 (26), e202305373. [CrossRef]
- Anderson, B. A.; Freestone, G. C.; Low, A.; De-Hoyos, C. L.; Iii, W. J. D.; Ostergaard, M. E.; Migawa, M. T.; Fazio, M.; Wan, W. B.; Berdeja, A.; Scandalis, E.; Burel, S. A.; Vickers, T. A.; Crooke, S. T.; Swayze, E. E.; Liang, X.; Seth, P. P., Towards next generation antisense oligonucleotides: mesylphosphoramidate modification improves therapeutic index and duration of effect of gapmer antisense oligonucleotides. Nucleic Acids Res 2021, 49 (16), 9026-9041. [CrossRef]
- Vlaho, D.; Fakhoury, J. F.; Damha, M. J., Structural Studies and Gene Silencing Activity of siRNAs Containing Cationic Phosphoramidate Linkages. Nucleic Acid Ther 2018, 28 (1), 34-43. [CrossRef]
- Stetsenko, D.; Chelobanov, B., Fokina, A., Burakova, E. Modified oligonucleotides activating RNAse H. International Patent No. WO2018156056A1, 30 August 2018.
- Paul, S.; Roy, S.; Monfregola, L.; Shang, S.; Shoemaker, R.; Caruthers, M. H., Oxidative substitution of boranephosphonate diesters as a route to post-synthetically modified DNA. J Am Chem Soc 2015, 137 (9), 3253-64. [CrossRef]
- Roy, S.; Paul, S.; Roy, M.; Kundu, R.; Monfregola, L.; Caruthers, M. H., Pyridinium Boranephosphonate Modified DNA Oligonucleotides. J Org Chem 2017, 82 (3), 1420-1427. [CrossRef]
- Chatelain, G.; Meyer, A.; Morvan, F.; Vasseur, J. J.; Chaix, C., Electrochemical detection of nucleic acids using pentaferrocenyl phosphoramidate α-oligonucleotides. New Journal of Chemistry 2011, 35(4), 893-901. DOI . [CrossRef]
- Vlaho, D.; Damha, M. J., Synthesis of Chimeric Oligonucleotides Having Modified Internucleotide Linkages via an Automated H-Phosphonate/Phosphoramidite Approach. Curr Protoc Nucleic Acid Chem 2018, 73 (1), e53. [CrossRef]
- Kupryushkin, M. S., Zharkov, T. D., Ilina, E. S., Markov, O. V., Kochetkova, A. S., Akhmetova, M. M., ... & Khodyreva, S. N, Triazinylamidophosphate Oligonucleotides: Synthesis and Study of Their Interaction with Cells and DNA-Binding Proteins. Russian Journal of Bioorganic Chemistry 2021, 47 (3), 719-733. [CrossRef]
- Zhukov, S. A., Pyshnyi, D. V., & Kupryushkin, M. S, Synthesis of novel representatives of phosphoryl guanidine oligonucleotides. Russian Journal of Bioorganic Chemistry 2021, 47(2), 380-389. [CrossRef]
- Markov, O. V.; Filatov, A. V.; Kupryushkin, M. S.; Chernikov, I. V.; Patutina, O. A.; Strunov, A. A.; Chernolovskaya, E. L.; Vlassov, V. V.; Pyshnyi, D. V.; Zenkova, M. A., Transport Oligonucleotides-A Novel System for Intracellular Delivery of Antisense Therapeutics. Molecules 2020, 25 (16). [CrossRef]
- Zharkov, T. D.; Mironova, E. M.; Markov, O. V.; Zhukov, S. A.; Khodyreva, S. N.; Kupryushkin, M. S., Fork- and Comb-like Lipophilic Structures: Different Chemical Approaches to the Synthesis of Oligonucleotides with Multiple Dodecyl Residues. Int J Mol Sci 2023, 24 (19). [CrossRef]
- Stetsenko, D.; Kupryushkin, M.; Pyshnyi, D. Modified Oligonucleotides and Methods for Their Synthesis. International Patent No. WO2016028187A1, 22 June 2014.
- Kupryushkin, M.; Zharkov, T.; Dovydenko, I.; Markov, O. Chemical compound comprising a triazine group and method for producing same. International Patent No. WO2022081046A1, 21 April 2022.
- Craig, K.; Abrams, M.; Amiji, M., Recent preclinical and clinical advances in oligonucleotide conjugates. Expert Opin Drug Deliv 2018, 15 (6), 629-640. [CrossRef]
- Khvorova, A.; Watts, J. K., The chemical evolution of oligonucleotide therapies of clinical utility. Nat Biotechnol 2017, 35 (3), 238-248. [CrossRef]
- Hnedzko, D.; McGee, D. W.; Karamitas, Y. A.; Rozners, E., Sequence-selective recognition of double-stranded RNA and enhanced cellular uptake of cationic nucleobase and backbone-modified peptide nucleic acids. RNA 2017, 23 (1), 58-69. doi: 10.1261/rna.058362.116.
- Baker, Y. R.; Thorpe, C.; Chen, J.; Poller, L. M.; Cox, L.; Kumar, P.; Lim, W. F.; Lie, L.; McClorey, G.; Epple, S.; Singleton, D.; McDonough, M. A.; Hardwick, J. S.; Christensen, K. E.; Wood, M. J. A.; Hall, J. P.; El-Sagheer, A. H.; Brown, T., An LNA-amide modification that enhances the cell uptake and activity of phosphorothioate exon-skipping oligonucleotides. Nat Commun 2022, 13 (1), 4036. [CrossRef]
- Kandasamy, P.; Liu, Y.; Aduda, V.; Akare, S.; Alam, R.; Andreucci, A.; Boulay, D.; Bowman, K.; Byrne, M.; Cannon, M.; Chivatakarn, O.; Shelke, J. D.; Iwamoto, N.; Kawamoto, T.; Kumarasamy, J.; Lamore, S.; Lemaitre, M.; Lin, X.; Longo, K.; Looby, R.; Marappan, S.; Metterville, J.; Mohapatra, S.; Newman, B.; Paik, I. H.; Patil, S.; Purcell-Estabrook, E.; Shimizu, M.; Shum, P.; Standley, S.; Taborn, K.; Tripathi, S.; Yang, H.; Yin, Y.; Zhao, X.; Dale, E.; Vargeese, C., Impact of guanidine-containing backbone linkages on stereopure antisense oligonucleotides in the CNS. Nucleic Acids Res 2022, 50 (10), 5401-5423. [CrossRef]
- Kandasamy, P.; McClorey, G.; Shimizu, M.; Kothari, N.; Alam, R.; Iwamoto, N.; Kumarasamy, J.; Bommineni, G. R.; Bezigian, A.; Chivatakarn, O.; Butler, D. C. D.; Byrne, M.; Chwalenia, K.; Davies, K. E.; Desai, J.; Shelke, J. D.; Durbin, A. F.; Ellerington, R.; Edwards, B.; Godfrey, J.; Hoss, A.; Liu, F.; Longo, K.; Lu, G.; Marappan, S.; Oieni, J.; Paik, I. H.; Estabrook, E. P.; Shivalila, C.; Tischbein, M.; Kawamoto, T.; Rinaldi, C.; Rajao-Saraiva, J.; Tripathi, S.; Yang, H.; Yin, Y.; Zhao, X.; Zhou, C.; Zhang, J.; Apponi, L.; Wood, M. J. A.; Vargeese, C., Control of backbone chemistry and chirality boost oligonucleotide splice switching activity. Nucleic Acids Res 2022, 50 (10), 5443-5466. [CrossRef]
- Shen, W.; De Hoyos, C. L.; Migawa, M. T.; Vickers, T. A.; Sun, H.; Low, A.; Bell, T. A., 3rd; Rahdar, M.; Mukhopadhyay, S.; Hart, C. E.; Bell, M.; Riney, S.; Murray, S. F.; Greenlee, S.; Crooke, R. M.; Liang, X. H.; Seth, P. P.; Crooke, S. T., Chemical modification of PS-ASO therapeutics reduces cellular protein-binding and improves the therapeutic index. Nat Biotechnol 2019, 37 (6), 640-650. [CrossRef]
- Maguregui, A.; Abe, H., Developments in siRNA Modification and Ligand Conjugated Delivery To Enhance RNA Interference Ability. Chembiochem 2020, 21 (13), 1808-1815. [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X. J., Therapeutic siRNA: state of the art. Signal Transduct Target Ther 2020, 5 (1), 101. [CrossRef]
- Kenski, D. M.; Butora, G.; Willingham, A. T.; Cooper, A. J.; Fu, W.; Qi, N.; Soriano, F.; Davies, I. W.; Flanagan, W. M., siRNA-optimized Modifications for Enhanced In Vivo Activity. Mol Ther Nucleic Acids 2012, 1 (1), e5. [CrossRef]
- Ly, S.; Echeverria, D.; Sousa, J.; Khvorova, A., Single-Stranded Phosphorothioated Regions Enhance Cellular Uptake of Cholesterol-Conjugated siRNA but Not Silencing Efficacy. Mol Ther Nucleic Acids 2020, 21, 991-1005. [CrossRef]
- Watts, J. K.; Deleavey, G. F.; Damha, M. J., Chemically modified siRNA: tools and applications. Drug Discov Today 2008, 13 (19-20), 842-55. [CrossRef]
- Pavlova, A. S.; Yakovleva, K. I.; Epanchitseva, A. V.; Kupryushkin, M. S.; Pyshnaya, I. A.; Pyshnyi, D. V.; Ryabchikova, E. I.; Dovydenko, I. S., An Influence of Modification with Phosphoryl Guanidine Combined with a 2'-O-Methyl or 2'-Fluoro Group on the Small-Interfering-RNA Effect. Int J Mol Sci 2021, 22 (18). [CrossRef]
- Liu, W.; Iwamoto, N.; Marappan, S.; Luu, K.; Tripathi, S.; Purcell-Estabrook, E.; Shelke, J. D.; Shah, H.; Lamattina, A.; Pan, Q.; Schrand, B.; Favaloro, F.; Bedekar, M.; Chatterjee, A.; Desai, J.; Kawamoto, T.; Lu, G.; Metterville, J.; Samaraweera, M.; Prakasha, P. S.; Yang, H.; Yin, Y.; Yu, H.; Giangrande, P. H.; Byrne, M.; Kandasamy, P.; Vargeese, C., Impact of stereopure chimeric backbone chemistries on the potency and durability of gene silencing by RNA interference. Nucleic Acids Res 2023, 51 (9), 4126-4147. [CrossRef]
- Monian, P.; Shivalila, C.; Lu, G.; Shimizu, M.; Boulay, D.; Bussow, K.; Byrne, M.; Bezigian, A.; Chatterjee, A.; Chew, D.; Desai, J.; Favaloro, F.; Godfrey, J.; Hoss, A.; Iwamoto, N.; Kawamoto, T.; Kumarasamy, J.; Lamattina, A.; Lindsey, A.; Liu, F.; Looby, R.; Marappan, S.; Metterville, J.; Murphy, R.; Rossi, J.; Pu, T.; Bhattarai, B.; Standley, S.; Tripathi, S.; Yang, H.; Yin, Y.; Yu, H.; Zhou, C.; Apponi, L. H.; Kandasamy, P.; Vargeese, C., Endogenous ADAR-mediated RNA editing in non-human primates using stereopure chemically modified oligonucleotides. Nat Biotechnol 2022, 40 (7), 1093-1102. [CrossRef]
- Kupryushkin, M. S.; Filatov, A. V.; Mironova, N. L.; Patutina, O. A.; Chernikov, I. V.; Chernolovskaya, E. L.; Zenkova, M. A.; Pyshnyi, D. V.; Stetsenko, D. A.; Altman, S.; Vlassov, V. V., Antisense oligonucleotide gapmers containing phosphoryl guanidine groups reverse MDR1-mediated multiple drug resistance of tumor cells. Mol Ther Nucleic Acids 2022, 27, 211-226. [CrossRef]
- Krutzfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K. G.; Tuschl, T.; Manoharan, M.; Stoffel, M., Silencing of microRNAs in vivo with 'antagomirs'. Nature 2005, 438 (7068), 685-9. [CrossRef]
- Wada, S.; Yasuhara, H.; Wada, F.; Sawamura, M.; Waki, R.; Yamamoto, T.; Harada-Shiba, M.; Obika, S., Evaluation of the effects of chemically different linkers on hepatic accumulations, cell tropism and gene silencing ability of cholesterol-conjugated antisense oligonucleotides. J Control Release 2016, 226, 57-65. [CrossRef]
- Chernikov, I. V.; Gladkikh, D. V.; Meschaninova, M. I.; Ven’yaminova, A. G.; Zenkova, M. A.; Vlassov, V. V.; Chernolovskaya, E. L., Cholesterol-Containing Nuclease-Resistant siRNA Accumulates in Tumors in a Carrier-free Mode and Silences MDR1 Gene. Mol Ther Nucleic Acids 2017, 6, 209-220. [CrossRef]
- LeDoan, T.; Etore, F.; Tenu, J. P.; Letourneux, Y.; Agrawal, S., Cell binding, uptake and cytosolic partition of HIV anti-gag phosphodiester oligonucleotides 3'-linked to cholesterol derivatives in macrophages. Bioorg Med Chem 1999, 7 (11), 2263-9. [CrossRef]
- Hwang, J.; Chang, C.; Kim, J. H.; Oh, C. T.; Lee, H. N.; Lee, C.; Oh, D.; Kim, B.; Hong, S. W.; Lee, D. K., Development of Cell-Penetrating Asymmetric Interfering RNA Targeting Connective Tissue Growth Factor. J Invest Dermatol 2016, 136 (11), 2305-2313. [CrossRef]
- Soutschek, J.; Akinc, A.; Bramlage, B.; Charisse, K.; Constien, R.; Donoghue, M.; Elbashir, S.; Geick, A.; Hadwiger, P.; Harborth, J.; John, M.; Kesavan, V.; Lavine, G.; Pandey, R. K.; Racie, T.; Rajeev, K. G.; Rohl, I.; Toudjarska, I.; Wang, G.; Wuschko, S.; Bumcrot, D.; Koteliansky, V.; Limmer, S.; Manoharan, M.; Vornlocher, H. P., Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature 2004, 432 (7014), 173-8. [CrossRef]
- Heindl, D.; Kessler, D.; Schube, A.; Thuer, W.; Giraut, A., Easy method for the synthesis of labeled oligonucleotides. Nucleic Acids Symp Ser (Oxf) 2008, (52), 405-6. [CrossRef]
- Pavlova, A.S.; Dovydenko, I.S.; Kupryushkin, M.S.; Grigor’eva, A.E.; Pyshnaya, I.A.; Pyshnyi, D.V.; Amphiphilic “like-a-brush” oligonucleotide conjugates with three dodecyl chains: Self-assembly features of novel scaffold compounds for nucleic acids de-livery. Nanomaterials 2020, 10, 1948. [CrossRef]
- Tanaka, Y.; Tanioku, Y.; Nakayama, T.; Aso, K.; Yamaguchi, T.; Kamada, H.; Obika, S., Synthesis of multivalent fatty acid-conjugated antisense oligonucleotides: Cell internalization, physical properties, and in vitro and in vivo activities. Bioorg Med Chem 2023, 81, 117192. [CrossRef]
- Gooding, M.; Malhotra, M.; Evans, J. C.; Darcy, R.; O'Driscoll, C. M., Oligonucleotide conjugates - Candidates for gene silencing therapeutics. Eur J Pharm Biopharm 2016, 107, 321-40. [CrossRef]
- Xing, Y.; Sheng, G.; Wang, J.; Lu, P.; Wang, Y., Preparation of triazoloindoles via tandem copper catalysis and their utility as alpha-imino rhodium carbene precursors. Org Lett 2014, 16 (4), 1244-7. [CrossRef]
Figure 1.
Number graphs of DLS experiment for the lipophilic oligonucleotides used in current study after 2h incubation in serum-free DMEM medium, 10 measurements. X axis is diameter size, nm; Y axis is number, percent.
Figure 1.
Number graphs of DLS experiment for the lipophilic oligonucleotides used in current study after 2h incubation in serum-free DMEM medium, 10 measurements. X axis is diameter size, nm; Y axis is number, percent.
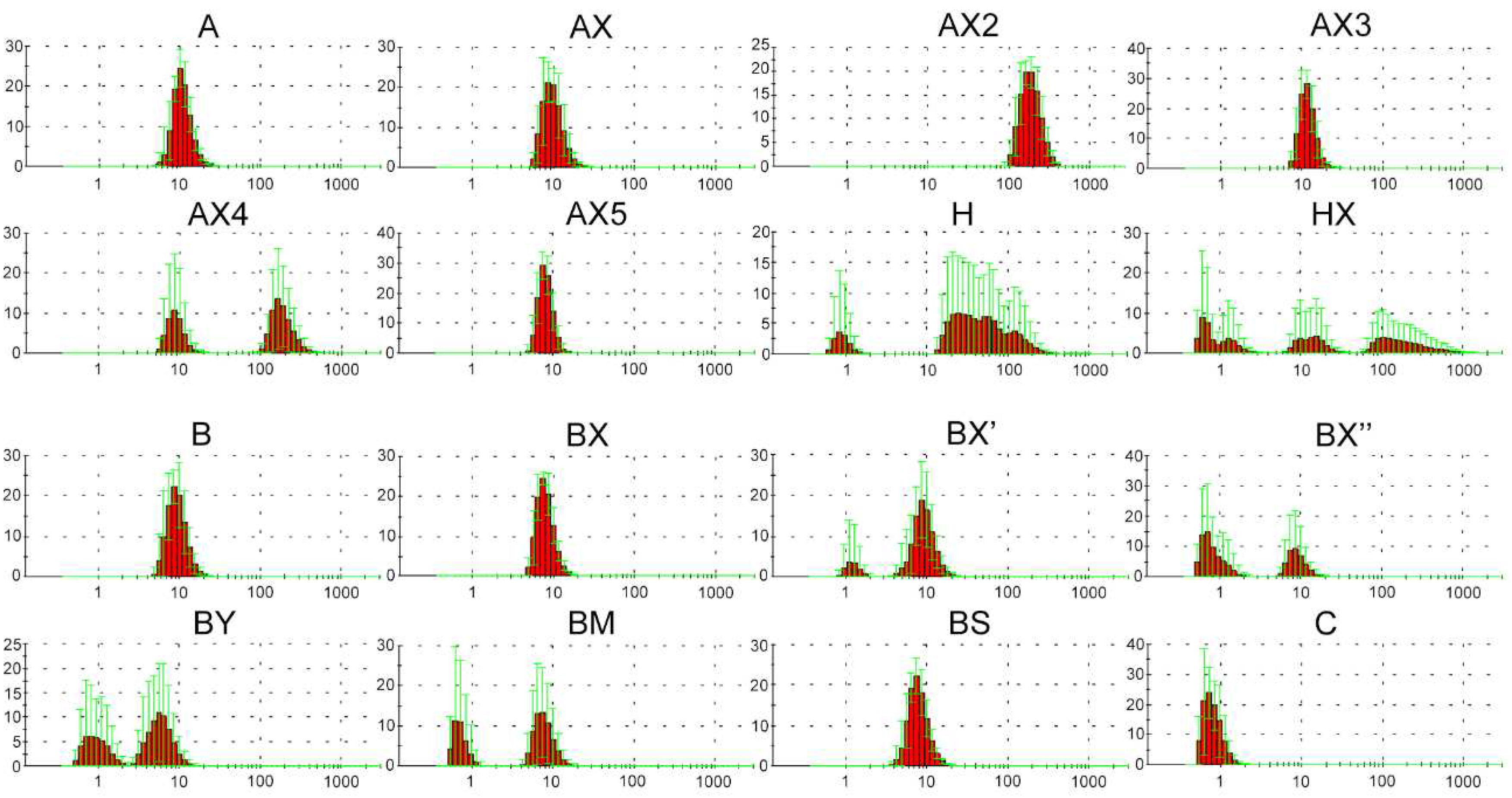
Figure 2.
Effects of the number of DMI modifications on the cellular uptake of FAM-labeled lipophilic phosphate-modified oligonucleotides. Analysis was performed in HEK293T cells 4 h post-transfection. w/o – untreated cells. Percentage of fluorescent cells (A) and relative fluorescence intensity (B) were measured by flow cytometry. Data are presented as MEAN±SD.
Figure 2.
Effects of the number of DMI modifications on the cellular uptake of FAM-labeled lipophilic phosphate-modified oligonucleotides. Analysis was performed in HEK293T cells 4 h post-transfection. w/o – untreated cells. Percentage of fluorescent cells (A) and relative fluorescence intensity (B) were measured by flow cytometry. Data are presented as MEAN±SD.
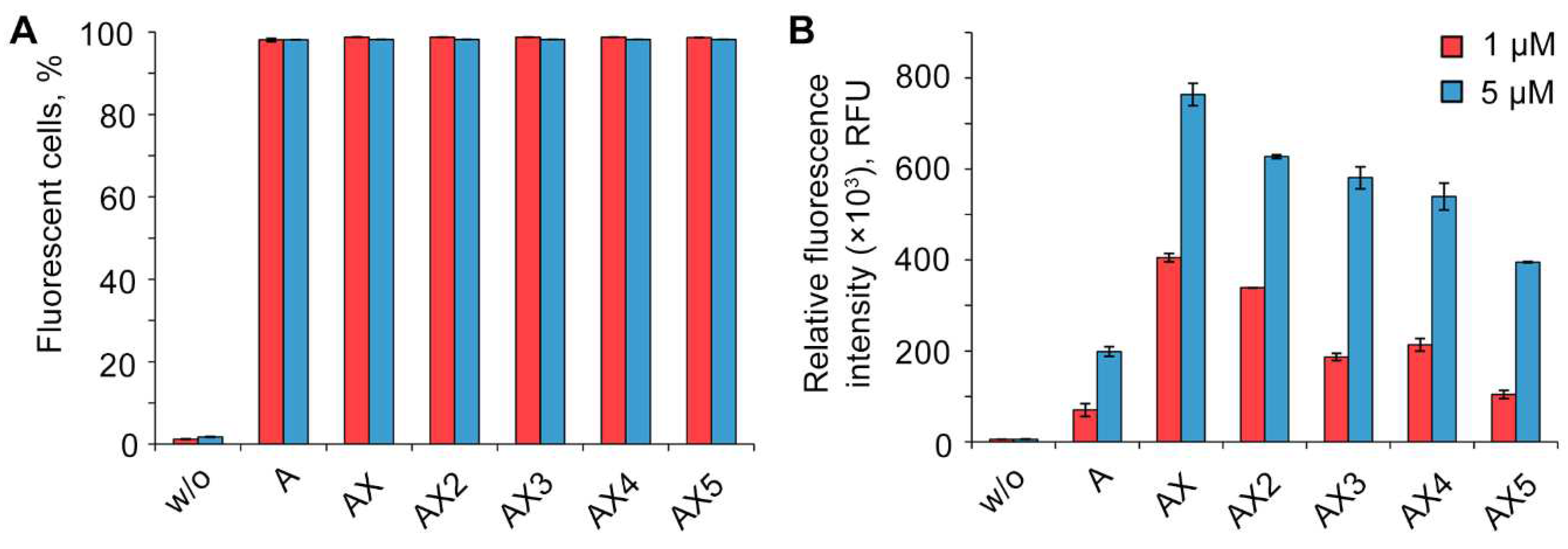
Figure 3.
Kinetics of intracellular accumulation of AX and AX5 oligonucleotides (5 µM) in HEK293T cells. Transfection was performed in serum-free (A) or serum-containing (B) medium. After 4 h of transfection, cells were washed with PBS and refreshed with culture medium in a wash-out regimen. Relative fluorescence intensity was measured by flow cytometry. Data are presented as MEAN±SD. (C) Intracellular distribution of AX and AX5 oligonucleotides (5 µM) in HEK293T cells 4 h post transfection. ODN – FAM-labeled oligodeoxynucleotides. Analysis was performed on a Zeiss LSM710 confocal microscope using a Plan-Apochromat 63x/1.40 Oil DIC M27 objective. ODN – FAM-labeled oligodeoxynucleotide. Nuclei were stained with DAPI. Scale bars 20 µm.
Figure 3.
Kinetics of intracellular accumulation of AX and AX5 oligonucleotides (5 µM) in HEK293T cells. Transfection was performed in serum-free (A) or serum-containing (B) medium. After 4 h of transfection, cells were washed with PBS and refreshed with culture medium in a wash-out regimen. Relative fluorescence intensity was measured by flow cytometry. Data are presented as MEAN±SD. (C) Intracellular distribution of AX and AX5 oligonucleotides (5 µM) in HEK293T cells 4 h post transfection. ODN – FAM-labeled oligodeoxynucleotides. Analysis was performed on a Zeiss LSM710 confocal microscope using a Plan-Apochromat 63x/1.40 Oil DIC M27 objective. ODN – FAM-labeled oligodeoxynucleotide. Nuclei were stained with DAPI. Scale bars 20 µm.
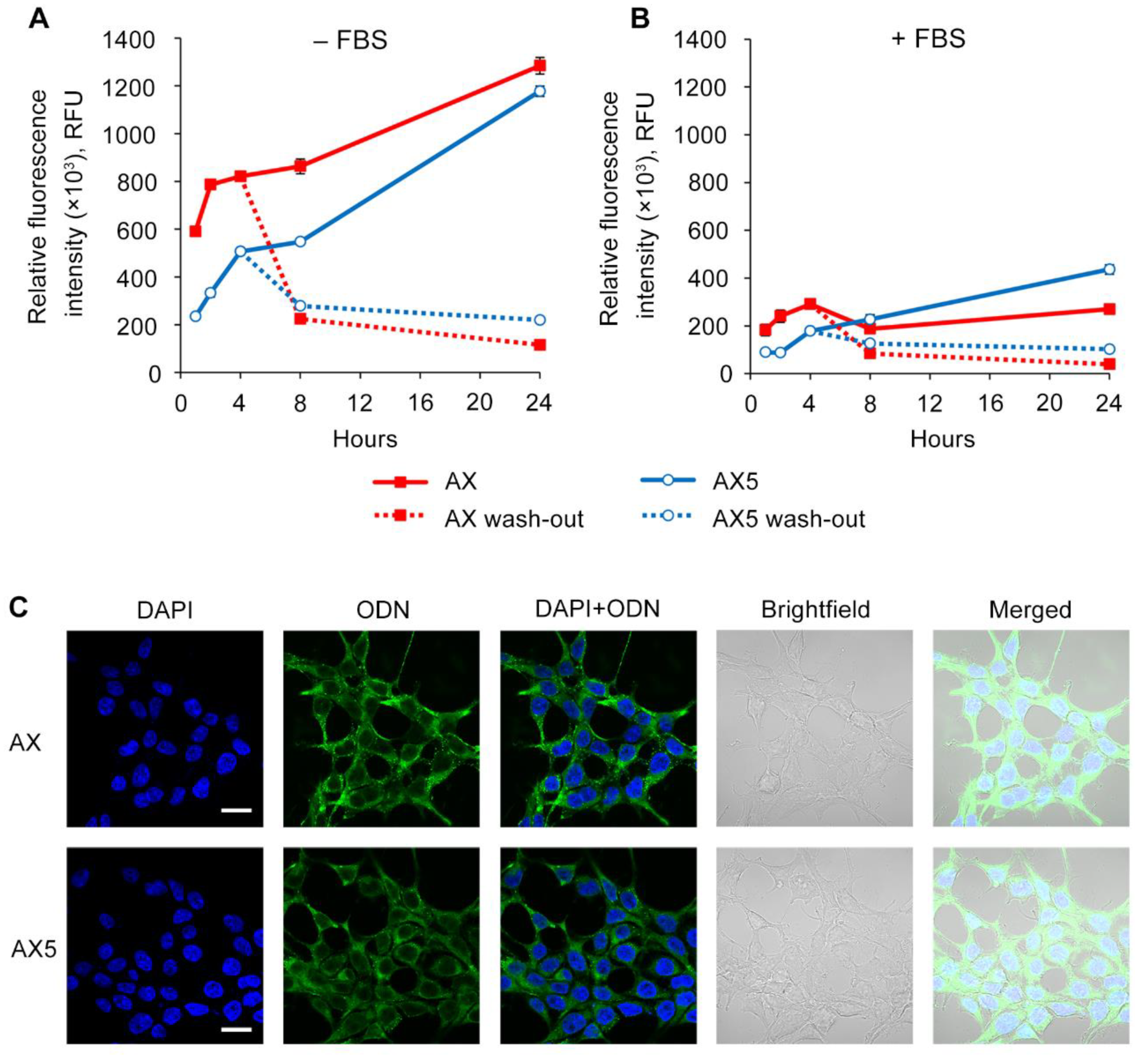
Figure 4.
Effects of oligonucleotide sequence, hydrophobic groups, type and location of phosphate modification on cellular uptake of lipophilic phosphate-modified oligonucleotides. Cellular accumulation of FAM-labeled lipophilic oligonucleotides (5 µM) in HEK293T cells was measured by flow cytometry 4 h post-transfection. w/o – untreated cells, LF – Lipofectamine 2000. (A), (C) – percentage of fluorescent cells; (B), (D) – relative fluorescence intensity. Data are presented as MEAN±SD.
Figure 4.
Effects of oligonucleotide sequence, hydrophobic groups, type and location of phosphate modification on cellular uptake of lipophilic phosphate-modified oligonucleotides. Cellular accumulation of FAM-labeled lipophilic oligonucleotides (5 µM) in HEK293T cells was measured by flow cytometry 4 h post-transfection. w/o – untreated cells, LF – Lipofectamine 2000. (A), (C) – percentage of fluorescent cells; (B), (D) – relative fluorescence intensity. Data are presented as MEAN±SD.
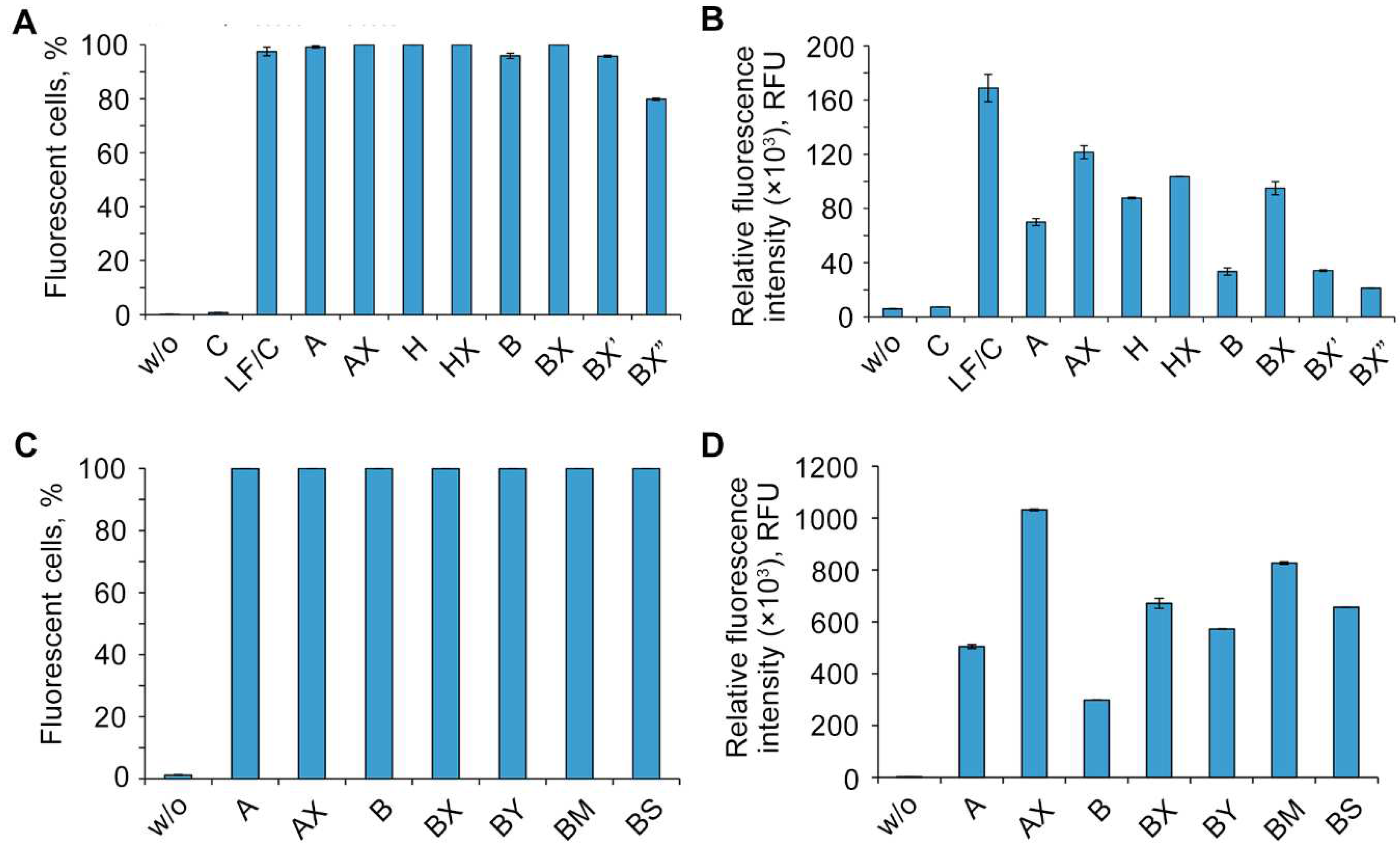
Figure 5.
Intracellular distribution of lipophilic phosphate-modified oligonucleotides (5 µM) in HEK293T cells 4 h post transfection. ODN – FAM-labeled oligodeoxynucleotides. Analysis was performed on a Zeiss LSM710 confocal microscope using a Plan-Apochromat 63x/1.40 Oil DIC M27 objective. Cell nuclei were stained with DAPI. Scale bars 20 µm.
Figure 5.
Intracellular distribution of lipophilic phosphate-modified oligonucleotides (5 µM) in HEK293T cells 4 h post transfection. ODN – FAM-labeled oligodeoxynucleotides. Analysis was performed on a Zeiss LSM710 confocal microscope using a Plan-Apochromat 63x/1.40 Oil DIC M27 objective. Cell nuclei were stained with DAPI. Scale bars 20 µm.
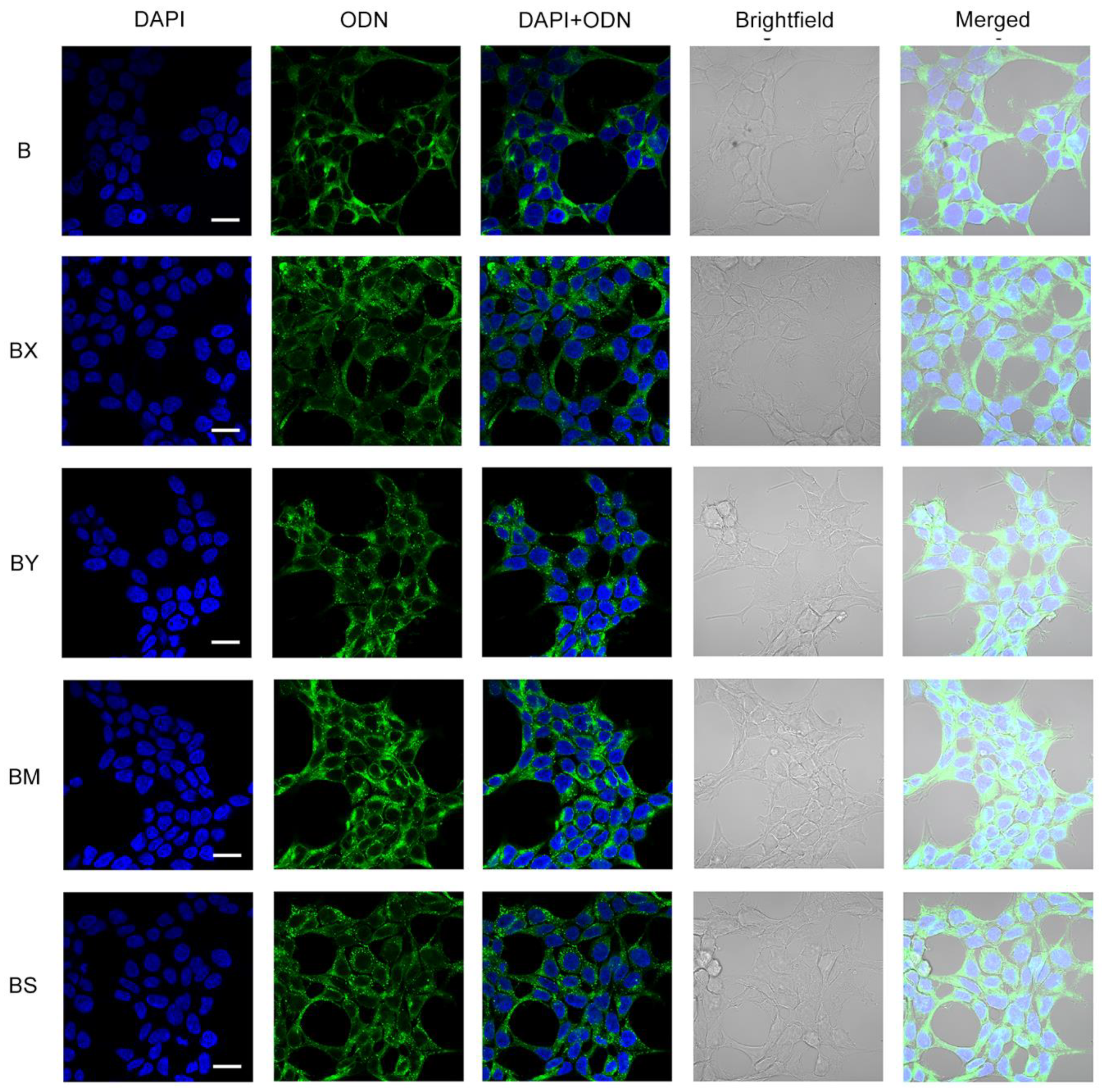

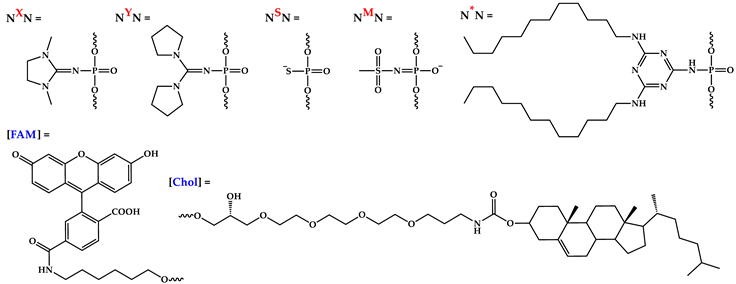
Table 1.
The list of modified oligonucleotides used in this study. [FAM] – 5′- terminal fluoresceine moiety, [Chol] – 3′-terminal cholesterol moiety. Internucleotide phosphate modifications: * – dodecyl-containing triazinyl phoshporamidate (TPA), X – 1,3-dimethylimidazolidin-2-iylidene phosphoramidate (DMI), Y – di(pyrrolidine-1-yl)methylene phosphoramidate, M – methanesulfonyl phosphoramidate, S – phosphorothioate.
Table 1.
The list of modified oligonucleotides used in this study. [FAM] – 5′- terminal fluoresceine moiety, [Chol] – 3′-terminal cholesterol moiety. Internucleotide phosphate modifications: * – dodecyl-containing triazinyl phoshporamidate (TPA), X – 1,3-dimethylimidazolidin-2-iylidene phosphoramidate (DMI), Y – di(pyrrolidine-1-yl)methylene phosphoramidate, M – methanesulfonyl phosphoramidate, S – phosphorothioate.
| Code | Sequence 5′→3′ | MS (calc / found) |
|---|---|---|
| A | 5′-[FAM]CTGACTATGAAGTAT*T-3′ | 5877.5 / 5876.5 |
| AX | 5′-[FAM]XCXTGACTATGAAGTAT*T-3′ | 6067.8 / 6067.6 |
| AX2 | 5′-[FAM] XCXTXGXACTATGAAGTAT*T-3′ | 6254.8 / 6253.8 |
| AX3 | 5′-[FAM] XCXTXGXAXCXTATGAAGTAT*T-3′ | 6444.9 / 6446.0 |
| AX4 | 5′-[FAM] XCXTXGXAXCXTXAXTGAAGTAT*T-3′ | 6635.1 / 6637.0 |
| AX5 | 5′-[FAM]XCXTXGXAXCXTXAXTXGXAAGTAT*T-3′ | 6825.3 / 6827.0 |
| H | 5′-[FAM]CTGACTATGAAGTATT[Chol]-3′ | 6188.7 / 6189.0 |
| HX | 5′-[FAM]XCXTGACTATGAAGTATT[Chol]-3′ | 6375.6 / 6376.5 |
| B | 5′-[FAM]AGTCTCGACTTGCTAT*T-3′ | 6130.4 / 6132.0 |
| BX | 5′-[FAM]XAXGTCTCGACTTGCTAT*T-3′ | 6320.6 / 6321.6 |
| BX’ | 5′-[FAM]AGTCTCGXAXCTTGCTAT*T-3′ | 6320.6 / 6322.0 |
| BX’’ | 5′-[FAM]AGTCTCGACTTGCTXAXT*T-3′ | 6320.6 / 6322.0 |
| BY | 5′-[FAM]YAYGTCTCGACTTGCTAT*T-3′ | 6428.7 / 6430.0 |
| BM | 5′-[FAM]MAMGTCTCGACTTGCTAT*T-3′ | 6284.4 / 6286.0 |
| BS | 5′-[FAM]SASGTCTCGACTTGCTAT*T-3′ | 6162.4 / 6163.5 |
| C | 5′-[FAM]AGTCTCGACTTGCTATT-3′ | 5686.0 / n.d. |
 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



