Submitted:
05 December 2023
Posted:
07 December 2023
You are already at the latest version
Abstract
Hungary's first national park was created in 1973 in the Hortobágy area to protect Europe's largest contiguous steppe area with its flora and fauna. The Hortobágy National Park - the Puszta was inscribed on the UNESCO World Heritage List as a cultural landscape in 1999. The park's outstanding importance is due to the predominantly non-arboreal steppe vegetation, home to a unique bird fauna, and the alkaline and chernozem soils with a complex, mosaic-like spatial structure. In addition, the landscape of the Hortobágy has a pastoral history stretching back thousands of years. Several hypotheses have been put forward, which suggest that the alkaline soils and the habitats that cover them were formed as a result of human activities related to river regulation that began in the second half of the 19th century. However, palaeoecological and palaeobiological studies over the last 30-40 years have pointed to the natural origin of the alkaline steppes, dating back to the end of the Ice Age. For thousands of years human activities, in particular grazing by domestic animals, hardly influenced the natural evolution of the area. Drainage of marshy and flooded areas began in the 19th century, and the introduction of more and more intensive agriculture, had a significant impact on the landscape. This paper aims to describe the past natural development of this special alkaline steppe ecosystem, with particular reference to the impacts of past and present human activities, including conservation measures.
Keywords:
undisturbed core sequence
; Holocene and Pleistocene paleobotanical data
; salty environment
; alkalinization
; Hortobágy National Park
; Carpathian Basin
1. Introduction
One of the most important cornerstones of the management system of an IUCN cate-gory II national park is the identification of its “original” vegetation, where original means its natural flora and fauna prior to significant changes caused by humans at the landscape level, such as cutting down and fragmenting continuous forests and turning natural grasslands to arable land. The first and still the largest national park of Hungary was established 50 years ago in the eastern part of the country, in the Great Hungarian Plain. Most of the park area is dominated by different kinds of alkaline soils covered by mosaics of grassland-wetland vegetation complexes (Video S1). According to the prevailing academic views at that time, there were two major human activities that significantly changed the original character of the flat landscape of the Hortobágy region. The first one was supposed to be, like in other lowland areas of Europe, the cutting down of forests to create pastures for grazing domestic animals, mostly cattle and sheep. In the first and still the only monograph of the Hortobágy National Park [1] the secondary character of the alkaline grassland-wetland mosaics was assumed as a starting point for the future management of the grassland-wetland mosaics of the Park. The potential vegetation map published in this monograph by Jakucs [2] suggested the dominance of hard and softwood gallery forests enclosing ancient alkaline grassland patches during the Early Neolithic period in the Hortobágy region. However, according to a recent critical source analysis, the presence of continuous forests in the Hortobágy region was assumed on the basis of the misinterpretation of a royal charter issued in the mid-15th century [3].
The second significant human intervention that was supposed to play an important role in the significant extension alkaline areas was the landscape scale change of the water regime of the Hortobágy by the major river regulation and drainage works started in the mid-19th century. This assumption is contradicted by descriptions of naturalists, such as Kitaibel [4] and Townson [5], indicating the presence of alkaline soils and vegetation in large areas before the start of hydroregulations in the Hortobágy region. These observations are also supported by the relevant map sheets and their description of the first military mapping survey of Hungary [6], undertaken in the second half of the 18th century. In addition, the presence of endemic plant species in the Hortobágy National Park, which occur exclusively in alkaline habitats, also indicates a much earlier appearance of such vegetation in the region. The most obvious representative of such species is Plantago schwarzenbergiana (Figure 1), which inhabits alkaline habitats in Hungary, Romania and Ukraine and is abundant in the southwestern part of the Hortobágy National Park called Kunkápolnás marsh, the study area of the present paper (Figure 2).
The CORINE Land Cover GIS database of the European Union (2018, 100-meter resolution) indicates the dominance of natural grasslands in the Hortobágy National Park (Figure 2) and even with the relatively low resolution, places covered by the typical alkaline plant-less or single-plant association Camphorosmetum annue are distinguished as “sparsely vegetated areas”.
According to an alternative view, the process of alkalinization dates back much further in natural landscape evolution, and the contemporary measures regulating the water regime could only extend the coverage of the already existing alkaline areas of primary, natural origin. First of all the geological, hydrogeological, geomorphological and climatic basis of the alkalinization process had to be clarified, which required the collection and analysis of all the data and information regarding the origin of sodium salts, the chemical processes and climatic characteristics responsible for their accumulation in the close to surface soil levels, as well as the role of the unique geomorphology in the development of the grassland-wetland mosaic landscape of the Hortobágy region [7]. Geological and paleontological evidence suggests that salt accumulation may have been caused by specific climatic and environmental conditions [8].
Although one of the most important questions about the historicity of salt accumulation near the surface and accumulating in soils is whether the environment adapted to excess sodium developed before or after hydroregulation, travelers’ descriptions and the sporadic research data from the second half of the 19th century before river regulation [3,9,10] indicate that salt accumulation in soils and a biota adapted to the saline environment had already developed in the northern part of the Transdanubian region before the hydroregulation measures [3,11,12,13,14,15,16]. One of the major problems in understanding the historical aspect of alkalinisation in the Carpathian Basin is that the source of alkalinisation has not been correctly identified by researchers. In addition, theoretical considerations based on these foundations have completely excluded the possibility of alkalinisation during the glacial periods, stating that the environmental conditions in the Hungarian Lowlands were not favourable for the process [17], hence the presence of glacial alkaline deposits, or signs of prehistoric alkalization were not assumed and searched for in the Great Hungarian Plain until 1988, when a multi-proxy paleoecological study of an undisturbed core series from the eastern margin of the Hortobágy, which was obtained by multiproxy paleoecology, succeeded in identifying an alkaline paleosol horizon dated between 30-40 kys beneath the glacial loess deposits [18,19,20,21,22]. Subsequent analyses revealed the presence of minerals typical of alkaline soils, such as gypsum, polyaluminates and amorphous silica gel, supporting the assumptions established in the field [18,21,22,23]. These data provided consistent evidence that conditions favourable to alkalisation may have developed during the last glacial cycle dated to MIS 3 as part of an intense brief interstadial warming, the Dansgaard-Oeschger cycle [18]. Cores taken from Bronze Age burial mounds also revealed the presence of buried chernozem and alkaline soils in the study area in the Early Holocene [18,24].
Since previous data and studies generally included only a part of the salt landscape development over time [8,23,25,26] we searched for a region for sampling where the whole sequence of changes, the complete evolution of the salt landscape could be captured. This place was found in the Kunkápolnás Marshland area, in the territory of Kunmadaras town.
2. Materials and Methods
Alkaline habitats cover an area of ca. 10,000 km2 in several parts of the Great Hungarian Plain including the Danube-Tisza and the Körös-Maros interfluves. The area of Hortobágy is the largest coherent occurrence of these habitat types in Europe, covering an area of ca. 2300 km2 (Figure 3). The first national park in the Carpathian Basin was established here in 1973 and was followed by several international designations, such as the UNESCO Biosphere Reserve, Ramsar Convention and the Natura 2000 network of the European Union. In 1999, the Hortobágy National Park – the Puszta was inscribed on the World Heritage List of UNESCO as a cultural landscape, as it maintains intact and visible traces of its traditional land-use forms over several thousand years, and illustrates the harmonious interaction between people and nature.
In the northern part of the Trans-Tisza region, in the center of the salt build-up region, species-rich halophilous vegetation developed both in dry and marshy areas [37]. In the northern part of the Trans-Tisza region, in the center of the salt build-up region, species-rich halophilous vegetation developed both in dry and marshy areas [37]. Levels of salt accumulation were detectable in the higher, drier, predominantly grassy levels of the earth pyramid layers of a Yamnaja culture burial mound, and in the deepest areas (meadows) of the studied region (Figure 4). Since the target area of sampling was a bombing range until August 1991, and Hungary was under Soviet-Russian military occupation (1956-1991), no correct map of the area could be made, that is why a digital elevation model of the drilling site was prepared at first (Figure 5).
The undisturbed core drilling site was located at the edge of the Kunkápolnás marsh complex and in the middle of a backfilled embankment across one of the paleochannels of the swamp. The embankment was constructed in 1958 and the surrounding area was used as a bombing range and had human disturbed surfaces, in particular thousands of bomb-craters. However, the embankment protected the underlying stratigraphic sequence, so we could use the most complete data set possible from the sedimentary assemblage accumulated up to the beginning of the Neolithic for the palaeoecological study. We also used our drone images to map the morphological evolution of the area and to show the development of the vegetation. The drone images were taken at an average altitude of 500 m and with a DJI Mavic 2 Pro drone and a Hasselblad L1D-20c camera. The sampling sites correspond to abandoned and infilled paleochannels fringing the open vegetation of alkaline grasslands (Figure 6).
Samples were taken in winter to avoid possible pollen contamination and they were subjected to sedimentological, geochemical, palynological, malacological and plant macrofossil analyses, and 14C chronological analyses [8,23,26,39]. The independent chronology established suggests that paleoenvironmental changes can be dated back to 50,000 cal BP years.
In the Hortobágy region, undisturbed sediment sequences were sampled from 9 different sites (Figure 4), including the 10 m long core of the Kunmadaras-Kunkápolnás marsh (marked as no. 1 in Figure 3, and no. 2 in Figure 4) using a special double-walled core head with a diameter of 10 cm. The main lithostratigraphic characteristics of the sediment sequence were determined and analysed. For the description of the cores, the internationally accepted system and symbols developed by Troels-Smith for unconsolidated sediments were used [40]. For the description of the sediment sequence and the development of the figures, the protocol was followed [41,42].
The organic matter (OM, LOI500), inorganic matter (IM, LOIres) and carbonate [CC, LOI900] contents of the samples were determined by the loss on ignition method [43]. The core was sampled at 2 cm sampling intervals, giving a total of 501 samples. The bulk samples were subjected to magnetic susceptibility testing [44]. The magnetic susceptibility of the samples was measured at 2 kHz using a Bartington MS2 magnetic susceptibility meter with MS2E high-resolution sensor [45,46]. Each sample was measured three times, and the average values of magnetic susceptibility were calculated and reported. Each sample was measured three times and the average values of magnetic susceptibility were calculated and reported. Grain size data of pretreated sediment core samples were obtained at 2 cm (501 samples) intervals for 42 grain size classes by laser diffraction using the OMEC Easysizer20 laser grain size analyser [47].
The concentrations of selected major and trace elements were determined by flame and graphite furnace atomic absorption spectroscopy. Radiocarbon dates of the sequences were obtained by both bulk and AMS (accelerator mass spectrometry) analyses. The radiocarbon ages of twenty-one samples were analysed at the Nuclear Re-search Centre of the Hungarian Academy of Sciences (Debrecen, Hungary) and the Direct Radiocarbon Laboratory (D-AMS, USA) in Seattle. Sample preparation and measurement protocols are described in detail by Hertelendi et al. [48,49] and Molnár et al. [50]. Prior to graphitization, samples were pretreated with weak acid (2% HCl) to remove surface impurities and carbonate coatings. Raw dates were calibrated using the Intcal20 calibration curve [51], using atmospheric data of Stuiver et al. [52] The original dates (14C) are indicated as uncal BP, while, the calibrated dates are indicated as cal BC and cal BP. Age-depth modelling and the estimation of the sedimentation rate (accrate.depth) were conducted using rbacon 2.5.8 [53] in RStudio [54] Build 461, and the IntCal20 calibration curve [55].
The cores were also subsampled at 2-cm/4-cm intervals for pollen analysis. Samples of 1 cm3 were obtained using a volumetric sampler and processed for pollen analysis [56]. Some pollen samples were analysed using the Zólyomi-Erdtman ZnCl2 method, which is the most commonly used method in Hungary [57], as this method gives better results than others for oxbow lake sediments [58]. A known amount of exotic pollen was added to each sample to determine the concentration of identified pollen grains [59]. To ensure a statistically manageable sample size, at least 300 grains per sample were counted (excluding exotics) [60]. Charcoal abundance was determined by the point count method [61]. Tablets with a known Lycopodium spore content (from Lund University, Sweden) were added to each sample to calculate pollen concentrations and accumulation rates. Pollen types were identified and modified according to Moore et al. [62], Beug [63] and Kozáková and Pokorny [64], supplemented by examination of photographs in Reille [65,66,67] and of reference material held in the Hungarian Geological Institute in Budapest. The analysis of local pollen zones and the statistical interpretations have been carried out by the software package of Keith David Benneth [68,69]. For macrobotanical studies, QLCMA analyses [70,71] were used. For the quarter-malacological analyses the methods, assessments and recent distribution data of Ložek [72], Sparks [73], Alexandrowicz [74,75], Krolopp [76,77,78] and Welter-Schultes [79] were applied, and the samples were pooled at 16 cm intervals to achieve a minimum of 100 per sample. The overall study procedure was based on the approach of Birks & Birks [80]. The sedimentological, geochemical, pollen, macrobotanical and quarter-malacological material, as well as the geochronological results were used to reconstruct local and regional evolutionary events over the last 50,000 years.
It must also be acknowledged that watersheds in floodplains subject to recurrent flooding receive large amounts of so-called "alien" pollen from distant areas, which greatly distorts the final pollen spectrum [81,82]. Consequently, these paleochannels are far from ideal pollen traps. The extent of "pollen pollution" is highly dependent on the depth and extent of flooding and the vegetation of the flooded area, which can distort the reconstruction of local and regional vegetation. To control and limit the potential bias as much as possible, our work used the analysis of plant macrofossils that provide information on vegetation that has been destroyed and preserved in situ. In this way, elements of the once in situ flora could be separated from potential regional and extra-regional elements. Paleo-vegetation can be reconstructed from pollen data using several approaches. For our purposes, the key question is to assess the extent to which the surrounding landscape and region is occupied by forest-steppe or steppe, as opposed to closed forest [33]. In our work the so-called biomization method [83], was used, complemented by an indicator taxa approach to infer the potential local presence of steppe [33,84].
According to the biomization approach [83] steppe indicator pollen taxa are predominantly composed of herbaceous taxa typical of steppe grasslands. Although their occurrence was used as further evidence for the local presence of open stands, such conclusions should be drawn with caution. Many steppe indicator taxa (herbs) are insect or self-pollinated species and produce relatively small amounts of pollen (e.g. Allium, Astragalus, Euphoria, Verbascum) and thus under-represented in the pollen spectra. Other steppe indicators are wind-pollinated and produce abundant pollen (e.g. Artemisia, Gramineae, Che-nopidaceae), and are therefore over-represented. According to Beug [63], Kozáková & Pokorný [64] and Magyari et al. [33], the following steppe indicator pollen taxa were identified in the core sequence of the analyzed paleochannels: Ajuga, Allium, Compositae (including Artemisia, Aster-type species and representatives of the subfamily Cichorioideae), Caryophyllaceae (including undetermined and Dianthus-type species), Chenopodiaceae (Atriplex, Kochia), Euphorbia, Gramineae, Helianthemum, Inula, Matricaria-type species (including Achillea, Anthemis, Matricaria), Plantago lanceolata, Plantago major/P. media, Thalictrum, Astragalus, Trifolium pratense type species, Trifolium repens type species and Verbascum. The ultimate aim of our work was to provide a reliable reconstruction of vegetation development of Hortobágy based on the study of local catchment basins [25,26,30,85].
Recently, attempts have been made to extend the pollen results of oxbow lakes located in the distant floodplain of the Tisza (ca. 60 km) to the Hortobágy area [33]. These distance inferences are rather ambiguous, partly because of the taphonomic problems mentioned above, and partly because the present floodplain of the Tisza is much younger (15-18 kys) and has a morphological and geological evolution that is quite different from that of the Hortobágy [86].
3. Results
3.1. Geochronological Results
The calibration of radiocarbon ages dates back to the age of the bedrock sand to 50,000 cal BP years (Figure 7). The age of the top of the profile at 10 cm has also been slightly modified thanks to the new calibration from 403 ± 17 uncal BP years to 35,696 ± 297 uncal BP (850 cm). Thus, the Kunkápolnás 1000 cm section captures the paleoecological changes from approximately 400 years to 50,000 years in the 21 radiocarbon data (Table 1). We focused mainly on the geochronological delineation of the LGM, MIS 2, MIS 3 development in the eastern Carpathian Basin and it did not address especially the Late Glacial / post-Glacial transition, which is the main issue of this paper. The 100 cm (10 m) long core sequence taken near Kunkápolnás (Figure 7) provides us with information about the paleoenvironmental and paleovegetational changes of the study area during the past 50,000 years (Figure 7 and Figure 8).
The results obtained show that the paleochannel studied was characterised by a relatively steady and slow accumulation of clayey silt (As2Ag2), reflecting natural flood cycles over the last 50 years. Evidence of changes in sedimentation, associated with an increase in organic and clay content, is limited to the upper part of the past 5-6 kys. This suggests that relatively uniform sedimentological processes have prevailed over most of the evolution of the channel, which is advantageous from a palaeoecological point of view, as the fluctuations and differences observed in the pollen and macrobotanical spectra reflect changes independent of changing geological processes (changing erosion base, selective pollen accumulation and retention). Sedimentation rates showed relatively uniformly low values at the minimum, maximum and mean (Figure 8), but some faster rates were also identified. The acceleration of sedimentation rates during the glacial period is associated with the acceleration of loess accumulation phases observed in the Carpathian Basin [87,88] (Figure 8). The increase in clay and organic matter content recorded in the upper part of the sequence may indicate anthropogenic disturbances in the basin environment, which may be attributed to the emergence of food-producing cultures [89], as this level appears to coincide with the appearance of Late Bronze Age - Early Bronze Age pit grave culture representatives in the study area [24]. Representatives of this culture are characterized by extensive animal husbandry and the construction of earth burial mounds (kurgans). One of these mounds is located 650 meters southwest of the studied core of Kunkápolnás, and others associated with this culture have also been identified scattered within a radius of about 1 km from other investigated profiles in the Hortobágy area.
3.2. Sedimentological Results
Between 50,000 and 25,000 years (1,000-500 cm), the sand content indicates fluvial sedimentation (Figure 9). The bedrock was dominated by the medium sand fraction (Figure 9), the layer was slightly cross-bedded in the undisturbed core layer and fluvial Valvata piscinalis shells were also found. It can be concluded that the analysed bed of the Kunkápolnás swamp (Ecse Basin) was formed by the development of a riverbed. During the first 25,000 years, carbonated river sediment rich in sand, and poor in finer-grained fractions accumulated in the gradually disconnected, 50,000 years old riverbed (Figure 9). The development of the river sediment is completely distinct from the sediments accumulated in the Tisza riverbed in terms of the grain composition (Figure 9 and Figure 10) and geochemical parameters (Figure 11), which are characteristic of the Sajó and Hernád rivers [86]. From about 25/27 thousand years onwards, the nature of sedimentation has fundamentally changed (Figure 9, Figure 10 and Figure 11) and sediment with a finer grain size composition has accumulated in the section. The changes in sediment composition (Figure 9, Figure 10 and Figure 11) indicate that the fluvial sedimentary phase has ended and that the cutoff meander phase dominated the bed formed for most of the years during MIS 3. It is likely that sandy sediment accumulated in the basin, which developed in the cutoff riverbed only during major floods and that it was the floodplain loess-like sediment that accumulated on the bank along the developed riverbed [90] that was washed into the basin of the oxbow lake. However, it should be noted that at this time extensive soil formation took place in the region and that has led to the formation of fossil saline soils by now [18,19,21,22,23].
Since due to the continuous military activity the area was excluded from the comprehensive geological mapping of the Hungarian lowlands [93,94,95,96,97], our undisturbed core drilling analyses can only suggest that a saline soil level could have developed in the vicinity of the riverbed, which was transformed into a sediment basin. The material of the eolic sedimentation [18,23,87,97,98,99] that developed in the region could have accumulated in the basin of the oxbow lake until 12800/13000 cal BP, when the eolic sedimentation in the Carpathian Basin came to an end [100], meaning that at the end of the Ice Age polygenetic alluviation took place in the oxbow lake, which evolved over 27/25 thousand years ago. This heterogenetic sedimentation is reflected in the highly heterogeneous grain composition from the clay to the fine sand fraction, and also in the rhythmic changes of the water-soluble element (Ca, Mg, Na and K) content (Figure 9, Figure 10 and Figure 11). The rhythmic changes were completed in the Late Glacial period of the Ice Age, and we can expect a steadily increasing clay, fine rock flour and organic matter content during the Holocene.
The increase in organic matter and clay content became more pronounced from about 5000/5200 cal BP, when livestock-keeping communities colonized the area (Pit Grave culture = Yamnaja = Kurgan culture). In this period, due to the human-induced soil erosion around the basin a sharp increase in organic matter and clay content can be detected in the sediment column of the Ecse Basin. Today, the riverbed is characterized by an alkaline marshy environment.
3.3. Results of the Pollen Analysis
The entire pollen sequence, both at the end of the Ice Age and during the Holocene, is dominated by herbaceous taxa, above all grasses (Poaceae), wormwood (Artemisia) and goosefoot species (Chenopodiaceae) (Figure 12), but also by Achillea taxa. The cumulative proportion of arboreal species (Arbor Pollen) exceeded 60% only in two glacial levels (43-46,000 and 25-28,000 years). Both glacial forest levels were dominated by the subgenus Pinus (Figure 12), as shown by anthracological analyses of charred trees recovered from fossil soils [101], and both glacial levels were associated with a fossil soil formation [21], when the proportion of vegetation cover, especially coniferous trees, increased in the region.
A characteristic feature of the pollen section is that the alluvial-fluvial influence may have been present from 50,000 years until the end of the Ice Age (12,800 years). This suggests [81,82] that the pollen composition may have reflected a more regional relationship during the glacial period, irrespective of the diameter of the studied basin [102]. However, the basin must have acted as a local pollen trap over the last 12,800 years. Yet the Holocene pollen assemblage was dominated by herbaceous taxa, with an overall proportion of more than 40-45% in all samples. The share of broad-leaved tree pollen varied between 8-16% in total, and the most characteristic broad-leaved taxon in the section was the eurytopic willow (Salix), a softwood gallery forest element (Figure 12).
The analysis of the pollen sequence and studies from the Eurasian forest-steppe-steppe environment [103,104,105] suggest that steppe to maximum boreal forest-steppe vegetation [33,106] might have stabilised in the study area during the glacial period. Although the presence of marshy vegetation became more abundant (Figure 12) in the Holocene, herbaceous species continued to dominate and a predominantly temperate steppe cover developed and persisted to the present day in the study area. Fluctuations in the pollen composition indicate cooler and milder climatic phases in the sequence [107,108,109,110,111,112,113,114,115]. The cooling phases can be synchronised with the increased dominance of grasses (Poaceae) and in general the Non-Arbor Pollen [113,116,117] and in parallel with this trend a decrease of the Arbor Pollen could be detected (Figure 12). Glacial warmings are indicated by an increasing dominance of coniferous pollen. The Holocene period is reflected in a marked increase in broad-leaved pollen and charcoal abundance (Figure 12). According to the changes in the pollen composition, the vegetation of the Hortobágy area have similarities with that of the Eurasian forest-steppe belt [118].
As the basin is of fluvial origin, the pollen composition may have been influenced by a fluvial inflow [81,82] and one has to consider the pollen input by the late glacial flooding of the Tisza river [119] into the already marsh-dominated former riverbed [33], our findings are based primarily on the AP/NAP pollen ratio. The relevant literature [33,120,121,122] clearly shows that during the warming periods, including the Holocene, when AP occurs at 50-60%, forest-steppe vegetation stabilised in the study area. Although the pollen composition is described to be consistent with Eurasian forest-steppe, the current climate analysis of the Carpathian Basin [123] suggests that the development of a basin effect (rain shadowing) caused by the surrounding mountain range, rather than the influence of the Eurasian continental climate zone.
3.4. Macrobotanical Results
From the core sequence, 2516 macrobotanical remains were recovered, the distribution of which is shown in Figure 13. Minimal plant remains were found in the bed of the section, corresponding to riverine infilling between 50,000 and 17,500 years (1000-350 cm). The plant remains at this level were dominated by Juncus roots, indeterminate monocots, and other indeterminate plant fragments that could easily have leached from the river bank into the Late Glacial river system and accumulated with the leached sediment in the abandoned, infilling riverbed at the study site. The bed was characterised by low vegetation cover with highly fluctuating water levels. Among the macrofossils, roots of a sedge species (Juncus sp.) were found in the largest quantities.
At 17,500 years, after the Last Glacial Maximum (LGM), in addition to the taxa that occupied the riverbed and the formed oxbow lake, oogonites (gyrogonites) of Chara taxa, above all Nitella cf. gracilis and Chara vulgaris, appeared in significant numbers. As a result, we can reconstruct the formation of a mesotrophic oxbow lake [124], poor in phosphate and organic matter, from 17,500 years ago and persisting until the beginning of the Holocene (12,000-11,500 years ago). In northern Europe, this Chara Lake stage is generally typical of the beginning of the Holocene, however, in the case of the sedimentary deposits in the Carpathian Basin dating back to the end of the Ice Age, such as the Kolon-tó near Izsák (Hungary) [125], this stage had already developed in the final part of the Ice Age. In the Chara Lake stage, around 15,000 years ago, terrestrial taxa, including those indicating a dry or periodically dry alkaline environment (Trifolium repens and Elatine triandra seeds), appeared (Figure 13). The presence of elements indicating salinity suggests that alkalinisation may have occurred as early as the end of the glacial period in the study area. Habitats typical of the alkaline environment may have developed along this mesotrophic oxbow lake, at the boundary between the loess grassland indicated by Trifolium repens seeds, and waterside mudflats, where groundwater fluctuations could be (and still are) most intense [126].
During the transition period between the end of the Ice Age and the beginning of the Holocene (between 12,800 and 11,300 cal BP), the studied floodplain was silted up, and the seasonal cyclical groundwater level fluctuations may have been amplified, which appears justified by the presence of Elatine hungarica seeds in the samples, indicating marked alkalinisation, as this species can stay in an anabiotic stage for several years and spread during favourable periods due to shallow water cover. However, no taxa indicating deeper or permanent water cover were found in the samples. The water level could be very low, a few centimeters at most, and the bed seasonally dried up. The constant presence of Typha species indicates a gradually warming climate. The vegetation of the marsh was poor, with the occurrence of the common water-plantain (Alisma plantago-aquatica), a few sand cinquefoil (Potentilla supina) and water fern (Salvinia natans). Mollusc and ostracod shells were negligible. Of the mosses, Amblystegium serpens was found in very small numbers, often living on woody debris, but at this site it is more likely to have been living on the decaying stems of some aquatic plant (e.g. lakeshore bulrush).
Later in the Holocene (7500-5000 cal BP years), higher water levels are indicated by the appearance of the white water lily (Nymphaea alba), common frogbit (Hydrocharis morsus-ranae), water fern (Salvinia natans) and duckweed species (Lemna sp.). Occasionally, the spiny naiad (Najas marina), typical of carbonate-rich waters, also appears, as well as the common bladderwort (Utricularia vulgaris) and water crowfoot species (Batrachium sp.). In open water, the presence of bryozoans and sponges, which are necessary for their colonisation, is also indicative of a more dense vegetation. Cladocerans and ostracods also appeared in open water with the most typical species of Daphnia pulex, Cerodaphnia sp. and Simocephalus vetulus. Juncus has been replaced by Typha in the riparian zone, while the common water plantain was found abundant in the marshy vegetation, with the presence of mint species (Mentha sp.) and the fine-leaved water dropwort (Oenanthe aquatica). It can be assumed that the marsh may have been periodically filled with water and that a typical Holocene aquatic-riparian zonation developed without any productive human influence.
From 5000 cal BP years the same indicator elements appeared, but in a different proportion than before. Mudflat communities spread and the amount of mollusc and ostracod shells increased, while the abundance of carophytes decreased. The amount of fly ash is the highest in the section, indicating a more intensive land-use. This change is fully associated with the appearance of Pit grave culture communities, as indicated by the nearby Ecse mound (kurgan). In parallel with the emergence of pastoral communities of the Pit grave culture, human communities engaged in productive farming (livestock keeping) also appeared in the landscape of the Hortobágy. Species indicative of seasonally drying mudflats are permanently present in the samples, such as the Hungarian waterwort (Elatine hungarica), the three-stamen waterwort (E. triandra), the sand cinquefoil (Potentilla supina), the dwarf clubrush (Schoenoplectus supinus) or the white clover (Trifolium repens). This community is very typical of regularly drying up or only periodically refilled beds. The occurrence of saltbush species (Atriplex hastata/saggitata) also indicates the development of silty, possibly alkaline soils.
3.5. Malacological Results
Samples were taken and processed at 8 cm but were evaluated aggregated at 16 cm to reach a statistical minimum of 100 individuals per sample [78]. More than 7,800 specimens of 27 molluscan taxa were retrieved from the sequence. Up to the LGM level, i.e. till 23,000 years, the malacological material (Figure 14) is dominated by taxa preferring flowing water (rheophilous group), and then, in the transitional period of the Ice Age and the Holocene, by aquatic Mollusca that require more water cover, but are less sensitive to water quality, and belong to the catholic group according to Sparks [73]. Direct fluvial recharge probably occurred in the area until 23,000 years, after which the proportion of elements indicating permanent water cover became dominant, i.e. fluvial recharge became more distant [21], but the area may have received significant additional water through rhythmic flooding after the development of the LGM level. At the same time, the slum group, which also tolerated intermittent water cover, also appeared in the sequence (Figure 14), but only in the second half of the Holocene, during the last 5000 years, when they became dominant within the malacofauna, suggesting a tripartite subdivision of the aquatic fauna composition within the sequence. The bedrock of the sequence indicates riverine recharge between 50,000 and 23,000 years ago, after which pond species with permanent water cover (Bithynia tentaculata, Anisus vorticulus, Gyraulus albus) dominated, but the members of the slum group also appeared, reflecting cyclical water level fluctuations.
The water supply in the second half of the Holocene was cyclical, and the studied river basin may have been periodically drying out during this period. Fauna elements living in a riparian environment were present throughout the sequence, but their proportion increased only in this last phase. The mesophilous and xerophilous taxa became dominant in the Holocene as well, but in the glacial period, between 29,000 and 24,000 years, such species (e.g. Cochlicopa lubrica, Chondrula tridens) also appeared in the sequence during the intensification of soil formation in the study area (Figure 14). During the glacial period, the proportion of cold-resistant elements (Lymnaea glabra, Valvata pulchella, Succinea oblonga and Trochulus hispidus) was highly significant. These taxa coexisted with thermophilous elements during the glacial/Holocene transition and at the beginning of the Holocene, before disappearing from the sequence in the early Holocene.
Cepaea vindobonensis, a character species of the Pannonian forest-steppe [127], appeared at the beginning of the Holocene, indicating the spread of Pannonian forest-steppe vegetation. At this time the number of species in the malacofauna increased, and beyond the appearance of 9-10 taxa at the end of the glacial period, species numbers exceeded 10 taxa per sample during the Pleistocene/Holocene transition and the early Holocene. This increase in species may have been due to the survival of glacial species that did not become extinct, whereas dispersing elements had already appeared in the section during the Holocene (Figure 14 and Figure 15). From the end of the Early Holocene (7500 cal BP) onwards, species abundance declined sharply, and the terrestrial fauna became dominated by thermophilous, mesophilous-xerophilous elements in the steppe-like environment evolved during the last 7500 years, which was certainly dry for part of the year. This mosaic environment may have stabilized after 5000 cal BP years, in the second half of the Holocene, when the number of individuals has doubled to more than 200 individuals per sample.
In the second half of the Holocene, the proportion of the group associated with a marshy environment also increased, along with the maximum of aquatic species indicating temporary water cover, and then the vegetation of the Kunkápolnás marsh may have stabilised for the last 5000 years in the study area.
4. Discussion
A complete fluvial cycle has been revealed [86,128,129,130,131,132] in the studied riverbed of the so-called Ecse basin) at the edge of the Kunkápolnás marsh complex, evolved from the carbonate fluvial sand sediment of the bedrock that formed about 50,000 years ago to the Holocene organic-rich clayey rock silt (marsh) sedimentary layer. Trends in sedimentation parameters can be synchronised with the accumulation of major sediment layers, changes in the sediment-forming environment and the climatic cycles of the past 50,000 years [107,108,109,110,112,116,117].
Pollen analysis was carried out on the entire undisturbed core section, results of which are completely different from those of the previously published pollen studies in Hungary [27,28,29,30,31,32,33,34,36,133,134]. This became particularly obvious when based on the biomization approach [83,103] the pollen composition of the undisturbed core drilling was compared with the pollen composition of the recent Eurasian biomes (Figure 16) and with the Arbor Pollen from pollen cores in the Carpathian Basin (Figure 17). These results show that a boreal forest-steppe with a dominance of Pinus diploxylon pollen was established in the glacial period in the studied Kunkápolnás region. Previous pollen analyses have shown that Pinus-dominated taiga forest patches developed along former living river branches [29] and the pollen composition (Figure 16) shows that the pollen composition of the Kunkápolnás is typical of the recent Eastern and Western Eurasian forest-steppe/steppe boundary. Results of the pollen, macrobotanical and malacological data suggest that the first patches of alkaline vegetation were established during the cold maximum of the glacial period (Figure 12, Figure 13, Figure 14 and Figure 15) in the study area.
However, general alkalinization and drier steppe phase became widespread in the region with the gradual warming of the climate from the Late Glacial about 12,000-13,000 years, together with the process that resulted in the dominance of Matricaria pollen type, Chenopodiaceae, Artemisia pollen (Figure 12). It was also the time when plant remains (Elatina taxa) typical of alkaline vegetation and drier loess steppes appeared [7]. Then, at the beginning of the Holocene warming, from about 11,000 years onwards, a change in the dominance of pollen and the appearance of vegetation remains typical of drier loess-steppe environments (Trifolium repens, Atriplex) resulted in the expansion of dry steppe and alkaline marsh patches. Based on the analysis of the radiocarbon-dated pollen, macrobotanical and malacological material, a mosaic habitat complex of alkaline marshes and steppes, without any human influence was established in the studied region at the end of the Ice Age, during the turn of the late glacial and the early Holocene. It appears that in contrast to the mountain rims and hill regions of the Carpathian Basin [32,34], no forest phase was established in the area at the beginning of the Early Holocene, but a mosaic vegetation structure of a forest-steppe developed, where trees occurred only at the margins of the former watercourses, while their cut and transformed beds gradually filled up with sediments. These data are in good agreement with the previously reported paleoecological data on the mosaic environmental structure of the Carpathian Basin [39]. However, it seems that local environmental factors (micromorphology, alkaline soil, morphology and groundwater) were extremely strong in the Kunkápolnás region, amplifying the essentially climate-driven alkalinization process (Figure 18), and therefore the alkaline patches in the Pannonian forest-steppe region were formed on a regional scale [123,136,137,138] under the influence of locally evolved edaphic factors.
The first pastoralist cultures (Pit Grave culture = Yamnaya = Kurgan culture), which appeared around 5000 years ago, only reinforced ongoing natural processes [24,139] but did not fundamentally transform the vegetation of the Kunkápolnás region. Similarly, there were no significant changes in the landscape character of the region over the following millennia, when the land management by domestic animals gradually increased and eventually took over completely the habitat management, i.e. grazing role of large ungulates, such as the aurochs and the European bison, which became extinct during the Holocene [140].
Significant negative changes have been brought about to the natural areas of the wider Hortobágy region by the river management and agricultural intensification interventions of the last two centuries, including the drainage of marshes, irrigation of pastures, and creation of fishponds and rice fields. The Hortobágy National Park, Hungary’s largest protected area, was established in 1973 in the central part of the region, which has been relatively little affected by these interventions, and where the natural alkaline grassland-wetland complexes continue to dominate the landscape to this day (Figure 2). The foundation of the Hortobágy National Park 50 years ago put a halt to these negative processes. Since then the site management organization, the Hortobágy National Park Directorate initiated and implemented several habitat restoration projects aiming at the preservation and restoration of the degraded natural vegetation mosaics. As a result of these consequent conservation efforts water supply systems for altogether 5000 hectares of marshes have been established and more than 1000 kilometers of disused channels, dykes and ditches were eliminated in the already 80,000 ha large area of the National Park. These already implemented landscape-scale conservation measures together with the recently planned restoration of the water regime of the central part of the Hortobágy area will hopefully enable the conservation of this unique habitat complex mosaic structure, along with the diverse fauna it hosts for future generations.
In light of the recent scientific results providing evidence on the primary, natural origin of the alkaline grassland-wetland complexes of the site, the Hungarian competent authorities might consider the nomination of the Hortobágy National Park – the Puszta property on the World Heritage List under the following natural criteria as well.
Criterion (vii): the flat landscape of the Hortobágy National Park is an area of exceptional natural beauty, representing the highest scenic quality, with pleasing and dramatic patterns and combinations of landscape features, which give it a distinctive character, including aesthetic qualities and topographic and visual unity.
Criterion (viii): the site is an outstanding example that represents the natural landscape and vegetation development of the Late Quaternary stage of Earth’s history, including significant ongoing geological processes in the development of landforms and significant geomorphic features.
Criterion (x): the Hortobágy National Park contains the most significant natural habitats for in-situ conservation of biological diversity of the temperate steppe zone, including those containing threatened species of outstanding universal value from the point of view of science or conservation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Video S1: Grassland-wetland mosaic habitats of the alkaline steppe in the Hortobágy National Park, Hungary.
Acknowledgments
The authors very much appreciate the generous support by the Hortobágy National Park Directorate. Special thanks to Attila Szilágyi for the drone image and video, and to Balázs Lesku for the photo of Plantago schwarzenbergiana.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Béres, A.; Bodó I.; Jakuts, P. Hortobágy - A nomád Pusztától a Nemzeti Parkig; Kovács, G., Salamon, F.; Ed.; Natura: Budapest, 1976; p. 351.
- Jakucs, P. A Hortobágy növényvilága. In Hortobágy - a nomád Pusztától a Nemzeti Parkig, Kovács, G., Salamon, F., Ed.; Natura: Budapest, 1976; pp. 38-56.
- Molnár, Z. A Hortobágy hazánk egyik legősibb növényzetű tája. In Válogatás az MTA Ökológiai és Botanikai Kutatóintézet kutatási eredményeiből, ÖBKI Műhelyfüzetek 2, Török, K., Keve, K.T., Kertész, M., Ed.; MTA Ökológiai és Botanikai Kutatóintézet: Vácrátót, Hungary, 2009; pp. 143-148.
- Lőkös, L. Diaria Itinerum Pauli Kitaibelii III; Természettudományi Múzeum: Budapest, 2001.
- Townson, R. Travels in Hungary, with a short account of Vienna in the year 1793; G G & J Robinson: London, UK, 1797; p. 506.
- Pók, J. Szabolcs vármegye katonai leírása 1782-1785; Nyíregyháza, 1994; Volume II, p. 128.
- Sümegi, P.; Szilágyi, G.; Gulyás, S.; Jakab, G.; Molnár, A. The Late Quaternary Paleoecology and Environmental History of the Hortobágy, a unique Mosaic Alkaline Steppe from the Heart of the Carpathian Basin, Central Europe. In Steppe Ecosystems Biological Diversity, Management and Restoration, Prieto, M.B.M., Diaz, T.B., Ed.; Nova Publishers: New York, USA, 2013; pp. 165-194.
- Sümegi, P.; Molnár, A.; Szilágyi, G. Szikesedés a Hortobágyon. Természet Világa 2000, 131, 213-216.
- Dapsy, L. Tiszaszabályozás befolyása a magyar talajra. Természettudományi Közlöny 1869, 1, 97-108.
- Boros, E.; Bíró, Cs. A Duna-Tisza-közi szikes tavak ökológiai állapot változásai a XVIII-XX. századokban (Ecological changes in natron lakes of the Danube-Tisza Interfluve between the 18th and 20th centuries). Acta Biologica Debrecina Oecologica Hungarica 1999, 9, 81-105.
- Molnár, Z. A Pitvarosi-puszták és környékük vegetáció- és tájtörténete a Középkortól napjainkig. Natura Bekesiensis 1996, 2, 65-97.
- Molnár, Z. Vegetation history of the Kardoskút area (S.E. Hungary) II: The lake Fehér-tó in the last 200 years. Tiscia 1997, 30, 27-34.
- Molnár, Z. Tájtörténeti adatok a hazai szikesek növényzetének ismeretéhez. In Ohattól Farkas-szigetig. Ökológiai kultúra – ökológiai nevelés, Tóth, A., Ed.; Alföldkutatásért Alapítvány: Budapes-Kisújszállás, 2003; pp. 71-95.
- Molnár, Z. Történeti tájökológiai kutatások az Alföldön. University of Pécs, Pécs, Hungary, 2007.
- Molnár, Z. A Duna – Tisza köze és a Tiszántúl fontosabb vegetáció típusainak holocén kori története: irodalmi értékelés egy vegetációkutató szemszögéből. Kanitzia 2008, 16, 93-118.
- Molnár, Z.; Bíró, M. Vegetation history of the Kardoskút area (SE Hungary) I: History of the steppes from the Middle Ages to the present. Tiscia 1997, 30, 15-25.
- Miháltz, I. A Duna-Tisza csatorna geológiai viszonyainak tanulmányozása. In A Duna-Tisza csatorna, Lampl, H., Hallósy F., Ed.; Egyetemi Nyomda: Budapest, Hungary, 1947.
- Sümegi, P. Hajdúság felső-pleisztocén fejlődéstörténete finomrétegtani (üledékföldtani, őslénytani, geokémiai) vizsgálatok alapján. Debreceni Egyetem, Debrecen, 1989.
- Sümegi, P. A negyedidőszak földtanának és őskörnyezettanának alapjai; JATEPress: Szeged, Hungary, 2001; p. 262.
- Sümegi, P. Régészeti geológia és történeti ökológia alapjai; JATEPress: Szeged, Hungary, 2003; p. 224.
- Szöőr, G.; Sümegi, P.; Balázs, É. Sedimentological and geochemical facies analysis Upper Pleistocene fossil soil zones discovered in the Hajdúság region, NE Hungary. In Quaternary environment in Hungary, Pécsi, M., Schweitzer, F., Ed.; Studies in geography in Hungary; Akadémiai Kiadó: Budapest, Hungary, 1991.
- Szöőr, G.; Sümegi, P.; Balázs, É. A Hajdúság területén feltárt felső pleisztocén fosszilis talajok szedimentológiai és geokémiai fácieselemzése. In Fáciesanalitikai, paleobiogeokémiai és paleoökológiai kutatások, Szöőr, G., Ed.; MTA Debreceni Bizottsága: Debrecen, Hungary, 1992; pp. 81-91.
- Sümegi, P. The Process of Sodification on Hortobágy in Space and Time according to geopedological investigation. In Hydro-Petro-Geology and Hungary. A field trip across the country. Excursion guide.; Magyarhoni Földtani Társulat: Budapest, Hungary, 1997; pp. 237-242.
- Sümegi, P.; Szilágyi, G. A quarter-malacological inventory of Hungarian kurgans. In Kurgan Studies: An environmental and archaeological multiproxy study of burial mounds in the Eurasian steppe zone, Pető, Á., Barczi, A., Ed.; Archeopress, British Archaeological Reports 2238: Oxford, UK, 2011.
- Sümegi, P.; Bodor, E.; Törőcsik, T. The origins of sodification in the Hortobágy region in the light of the palaeoenvironmental studies at Zám–Halasfenék. In Environmental Archaeology in North-Eastern Hungary, Gál, E., Juhász, I., Sümegi, P., Ed.; Varia Archaeologica Hungarica; MTA Régészeti Intézet: Budapest, Hungary, 2005.
- Sümegi, P.; Bodor, E.; Törőcsik, T. A hortobágyi szikesedés eredete. In Táj, környezet és társadalom. Ünnepi tanulmányok Keveiné Bárány Ilona professzor asszony tiszteletére, Kiss, A., Mezősi, G., Sümeghy, Z., Ed.; Szegedi Tudományegyetem: Szeged, Hungary, 2006; pp. 633-641.
- Sümegi, P.; Molnár, M.; Jakab, G.; Persaits, G.; Majkut, P.; Páll, D.G.; Gulyás, S.; Jull, A.J.T.; Törőcsik, T. Radiocarbon-dated paleoenvironmental changes on a lake and peat sediment sequence from the central part of the Great Hungarian Plain (Central Europe) during the last 25.000 years. Radiocarbon 2011, 52, 85-97. [CrossRef]
- Willis, K.J. Impact of the early Neolithic Körös culture on the landscape: evidence from palaeoecological investigations of Kiri-tó. In The Early Neolithic on the Great Hungarian Plain: investigations of the Körös culture site of Ecsegfalva 23, Co. Békés, Whittle, A., Ed.; Varia Archaeologica Hungarica; Budapest, Hungary, 2007; pp. 83-99.
- Sümegi, P.; Magyari, E.; Daniel, P.; Hertelendi, E.; Rudner, E. A kardoskúti Fehér-tó negyedidőszaki fejlődéstörténetének rekonstrukciója. Földtani Közlöny 1999, 129, 479-519.
- Sümegi, P.; Persaits, G.; Gulyás, S. Woodland-Grassland Ecotonal Shifts in Environmental Mosaics: Lessons Learnt from the Environmental History of the Carpathian Basin (Central Europe) During the Holocene and the Last Ice Age Based on Investigation of Paleobotanical and Mollusk Remains. In Ecotones Between Forest and Grassland, Myster, R.W., Ed.; Springer Press: New York, USA, 2012; pp. 17-57. [CrossRef]
- Jakab, G.; Sümegi, P.; Magyari, E. New Quantative Method for the Paleobotanical Description of Late Quaternary Organic Sediments (Mire-Development Pathway and Paleoclimatic Records from Southern Hungary). Acta Geologica Hungarica 2004, 47, 373-409. [CrossRef]
- Willis, K.J.; Braun, M.; Sümegi, P.; Tóth, A. Does soil change cause vegetation change or vice-versa? Ecology 1997, 78, 740-750. [CrossRef]
- Magyari, E.; Chapman, J.C.; Passmore, D. G.; Allen, J. R. M.; Huntley, J.P.; Huntley, B. Holocene persistence of wooded steppe in the Great Hungarian Plain. Journal of Biogeography 2010, 37, 915-935. [CrossRef]
- Willis, K.J.; Sümegi, P.; Braun, M.; Tóth A. . The Late Quaternary environmental history of Bátorliget, N.E. Hungary. Palaeogeography, Palaeoclimatology, Palaeoecology 1995, 118, 25-47. [CrossRef]
- Sümegi, P.; Törőcsik, T. Magyarország növényzetének története. Rosalia 2010, 1, 11-13.
- Gardner, A.R. The impact of Neolithic agriculture on the environments of south-east Europe. University of Cambridge, Cambridge, 1999.
- Borhidi, A. Magyarország növénytársulásai; Akadémiai Kiadó: Budapest, Hungary, 2007.
- Ábrányi, K.; Bokor, J.; Timon, Á.; Radó, A.; Gaál, J.; Wargha, S. A Pallas Nagy Lexikona; Pallas Irodalmi és Nyomdai Részvénytársaság - Révai Testvérek: Budapest, 1895.
- Sümegi, P. Loess and Upper Paleolithic environment in Hungary; Aurea Kiadó: Nagykovácsi, Hungary, 2005; p. 312.
- Troels-Smith, J. Karakterisering af lose jordater (Characterisation of Unconsolidated Sediments). Danmarks Geologiske Undersogelse 1955, 3, 39-73. [CrossRef]
- Vári, T.Z.; Molnár, D.; Sümegi, P.; Sümegi, B.P.; Törőcsik, T.; Szakál, E.; Benyó-Korcsmáros, R. Holocene environmental history of the Bottomless Lake (Tăul fără fund) sphagnum peat bog in Bǎgǎu, Romania. Studia Quaternaria 2020, 37(2), 69-77.
- Vári, T.Z.; Gulyás, S.; Sümegi, P. Reconstructing the Paleoenvironmental Evolution of Lake Kolon (Hungary) through Integrated Geochemical and Sedimentological Analyses of Quaternary Sediments. Quaternary 2023, 6(3), 39. [CrossRef]
- Dean, W.E. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition; comparison with other methods. Journal of the Sedimentary Research 1974, 44, 242-248. [CrossRef]
- Oldfield, F.; Thompson, R.; Barber, K.E. Changing atmospheric fallout of magnetic particles recorded in recent ombrotrophic peat sections. Science 1978, 199, 679-680. [CrossRef]
- Dearing, J. Environmental Magnetic Susceptibility: Using the Bartington MS2 System; Chi Publishing: Keniloworth, UK, 1994.
- Xu, X.W.; Qiang, X.K.; Fu, C.F.; Zhao, H.; Chen, T.; Sun, Y.F. Characteristics of frequency-dependent magnetic susceptibility in Bartington MS2 and Kappabridge MFK1-FA, and its application in loess-paleosol, red clay and lacustrine sediments. Chinese Journal of Geophysics 2012, 55(1), 197-206.
- Njalsson, T.; Novosselov, I. Design and optimization of a compact low-cost optical particle sizer. Journal of Aerosol Science 2018, 119, 1-12. [CrossRef]
- Hertelendi, E.; Csongor, É.; Záborszky, L.; Molnár, I.; Gál, I.; Győrffy, M.; Nagy, S. Counting system for high precision C-14 dating. Radiocarbon 1989, 32, 399-408. [CrossRef]
- Hertelendi, E.; Sümegi, P.; Szöőr, Gy. Geochronologic and paleoclimatic characterization of Quaternary sediments in the Great Hungarian Plain. Radiocarbon 1992, 34, 833-839. [CrossRef]
- Molnár, M.; Janovics, R.; Major, I.; Orsovszki, J.; Gönczi, R.; Veres, M.; Leonard, A.G.; Castle, S.M.; Lange, T.E.; Wacker, L.; Hajdas, I.; Jull, A.J.T. Status Report of the New AMS 14C Sample Preparation Lab of the Hertelendi Laboratory of Environmental Studies (Debrecen; Hungary). Radiocarbon 2013, 55, 665-676. [CrossRef]
- Reimer, P.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; Grootes, P.M.; Guilderson, T.P.; Hajdas, I.; Heaton, T..J.; Hogg, A.G.; Hughen, K.A.; Kromer, B.; Manning, S.W.; Muscheler, R.; Palmer, J.G.; Pearson, C.; van der Plicht, J.; Reimer, R.W.; Richards, D.A.; Scott, E.M.; Southon, J.R.; Turney, C.S.M.; Wacker, L.; Adolphi, F.; Büntgen, U.; Capano, M.; Fahrni, S.; Fogtmann-Schulz, A.; Friedrich, R.; Köhler, P.; Kudsk, S.; Miyake, F.; Olsen, J.; Reinig, F.; Sakamoto, M.; Sookdeo, A.; Talamo, S. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725-757. [CrossRef]
- Stuiver, M.; Reimer, P.J.; Braziunas, T.F. High-precision radiocarbon age calibration for terrestrial and marine samples. Radiocarbon 1998, 40, 1127-1151. [CrossRef]
- RBacon. Age-Depth Modelling using Bayesian Statistics. Available online: https://cran.r-project.org/web/packages/rbacon/index.html (accessed on 29 August 2023).
- RStudio Desktop—Posit. Available online: (accessed on 29 August 2023).
- Blaauw, M.; Christen, J.A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Analysis 2011, 6(3), 457-474. [CrossRef]
- Berglund, B.E.; Ralska-Jasiewiczowa, M. Pollen analysis and pollen diagrams. In Handbook of Holocene Palaeoecology and Palaeohydrology, Berglund, B.E., Ed.; J. Wiley and Sons Ltd.: Chichester-Toronto, 1986.
- Zólyomi, B. Magyarország növénytakarójának fejlődéstörténete az utolsó jégkorszaktól. MTA Biológiai Osztályának Közleményei 1952, 1, 491-544.
- Magyari, E.K. Climatic versus human modification of the Late Quaternary vegetation in Eastern Hungary. Debrecen University, Debrecen, 2002.
- Stockmarr, J. Tablets with spores used in absolute pollen analysis. Pollen et Spores 1971, 13, 614-621.
- Maher, L.J.J. Nomograms for computing 0.95 confidence limits of pollen data. Review of Palaeobotany and Palynology 1972, 13, 85-93. [CrossRef]
- Clark, R.L. Point count estimation of charcoal in pollen preparations and thin sections of sediments. Pollen et Spores 1982, 24, 523-535.
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell Scientific Publications: Oxford, UK, 1991.
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete; Pfeil: München, Germany, 2004.
- Kozáková, R.; Pokorný, P. Dynamics of the biotopes at the edge of a medieval town: pollen analysis of Vltava river sediments in Prague, Czech Republic. Preslia 2007, 79, 259-281.
- Reille, M. Pollen et spores d'europe et d'afrique du nord; Laboratoire de Botanique historique et Palynologie: Marseille, 1992; p. 543.
- Reille, M. Pollen et Spores d'Europe et d'Afrique du Nord, Supplément 1; Laboratoire de Botanique historique et Palynologie: Marseille, 1995; p. 331.
- Reille, M. Pollen et spores d'Europe et d'Afrique du Nord: Supplément 2; Laboratoire de botanique historique et palynologie: Marseille, 1998; p. 521.
- Bennett, K.D. Psimpoll manual. 2005.
- Birks, H.J.B.; Gordon, A.D. Numerical methods in Quaternary pollen analysis; Academic Press: London, UK, 1985.
- Barber, K.E.; Chambers, F. M.; Maddy, D.; Brew, J. A sensitive high resolution record of the Holocene climatic change from a raised bog in northern England. The Holocene 1994, 4, 198-205. [CrossRef]
- Sugita, S. Pollen representation of vegetation in Quaternary sediments: theory and method in patchy vegetation. Journal of Ecology 1994, 82, 881-897. [CrossRef]
- Ložek, V. Quartärmollusken der Tschechoslowakei. Rozpravy Ústredniho ústavu geologického 1964, 31, 1-374.
- Sparks, B.W. Non-marine Mollusca and Quaternary ecology. The Journal of Animal Ecology 1964, 33, 87-98. [CrossRef]
- Alexandrowicz, W.P. Molluscan assemblages of Late Glacial and Holocene calcareous tufa in Southern Poland. Folia Quaternaria 2004, 75, 3-309.
- Alexandrowicz, W.P. Malacological sequence of Weichselian (MIS 5-2) loess series from a profile in Grodzisko Dolne (southern Poland) and its palaeogeographic significance. Quaternary International 2014, 319, 109-118. [CrossRef]
- Krolopp, E. Mollusc fauna of the sedimentary formations of the Quaternary period, Hungary. Acta Geologica Hungarica 1965, 9, 153-160.
- Krolopp, E. Quaternary malacology in Hungary. Földrajzi Közlemények 1973, 21, 161-171.
- Krolopp, E. Biostratigraphic division of Hungarian Pleistocene Formations according to their Mollusc fauna. Acta Geologica Hungarica 1983, 26, 62-89.
- Welter-Schultes, F. European non-marine mollusc, a guide for species identification; Planet Poster Edition: Göttingen, 2012; p. 760.
- Birks, H.J.B.; Birks, H.H. Quaternary paleoecology; Edward Arnold: London, 1980; p. 289.
- Fall, P. Pollen taphonomy in a canyon stream. Quaternary Research 1987, 28(3), 393-406. [CrossRef]
- Hall, S.A. Pollen analysis and paleoecology of alluvium. 31(3) 1989, 435-438. [CrossRef]
- Prentice, I.C.; Guiot, J.; Huntley, B.; Jolly, D.; Cheddadi, R. Reconstructing biomes from palaeoecological data: a general method and its application to European pollen data at 0 and 6 ka. Climate Dynamics 1996, 12, 185-194. [CrossRef]
- Magyari, E.K. Late quaternary vegetation history in the Hortobágy steppe and Middle Tisza floodplain, NE Hungary. Studia Botanica Hungarica 2011, 42, 185-203.
- Sümegi, P.; Szilágyi, G. A Hortobágy új felszínfejlődési modellje és a szikesedés eredete. Acta Biologica Debrecina 2010, 22, 37-36.
- Timár, G.; Sümegi, P.; Horváth, F. Late Quaternary dynamics of Tisza River: Evidence of climatic and tectonic controls. Tectonophysics 2005, 410, 97-110. [CrossRef]
- Sümegi, P.; Gulyás, S.; Molnár, D.; Sümegi, B.P.; Almond, P.C.; Vandenberghe, J.; Liping Zhou; Pál-Molnár; E.; Törőcsik, T.; Hao, Q.; Smalley, I.; Molnár, M.; Marsi, I. New chronology of the best developed loess/paleosol sequence of Hungary capturing the past 1.1 ma: Implications for correlation and proposed pan Eurasian stratigraphic schemes. Quaternary Sciences Reviews 2018, 191, 144-166. [CrossRef]
- Sümegi, P.; Molnár, D.; Náfrádi, K.; Makó, L.; Cseh, P.; Törőcsik, T.; Molnár, M.; Zhou, L. Vegetation and land snail-based reconstruction of the palaeocological changes in the forest steppe eco-region of the Carpathian Basin during last glacial warming. Global Ecology and Conservation 2022, 33, e01976. [CrossRef]
- Willis, K.J.; Sümegi, P.; Braun, M.; Bennett, K.D.; Tóth, A. Prehistoric land degradation in Hungary: who, how and why? Antiquity 1998, 72, 101-113. [CrossRef]
- Sümegi, P.; Náfrádi, K.; Molnár, D.; Sávai, S. Results of paleoecological studies in the loess region of Szeged-Öthalom (SE Hungary). Quaternary International 2015, 372, 66-78. [CrossRef]
- Dániel, P. Methods of the five-step extraction-digestion method. In The geohistory of Bátorliget Marshland, Sümegi, P., Gulyás, S., Ed.; Archaeolingua Press: Budapest, Hungary, 2004.
- Dániel, P.; Kovács, B.; Győri, Z.; Sümegi, P. A Combined Sequential Extraction Method for Analysis of Ions Bounded to Mineral Component. In Proceedings of the Workshop of the 4th Soil and Sediment Contaminant Analysis, Lausanne, Switzerland, 1996.
- Rónai, A. A síkvidékek földtani kutatásának jelentősége. Földtani Intézet Évi Jelentése 1961-ről 1964, 5-17.
- Rónai, A. Negyedkori üledékképződés és éghajlattörténet az Alföld Medencéjében: Quartärsedimentation und Klimageschichte im Becken der ungarischen Tiefebene (Alföld). Magyar Állami Földtani Intézet évkönyve 56. köt. 1. füz.; Műszaki Könyvkiadó: Budapest, Hungary, 1972; p. 421.
- Rónai, A. Negyedidőszaki kéregmozgások a Magyar medencében. Földtani Közlöny 1977, 107, 431-436.
- Rónai, A. Komplex síkvidéki földtani kutatások és agrogeológiai kapcsolataik. MTA X. Osztály Közleményei 1982, 15, 183-188.
- Rónai, A. Az Alföld földtana. Acta Geologica Hungarica 1985, 21, 1-445.
- Lehmkuhl, F.; Bösken, J.; Hošek, J.; Sprafke, T.; Marković, S.B.; Obreht, I.; Hambach, U.; Sümegi, P.; Lindner, H. Loess distribution and related Quaternary sediments in the Carpathian Basin. Journal of Maps 2018, 14, 673-682. [CrossRef]
- Lehmkuhl, F.; Bösken, J.; Pötter, S.; Sprafke, T.; Schulte, P.; Jary, Z.; Antoine, P.; Wacha, L.; Wolf, D.; Zerboni, A.; Hošek, J.; Marković, S.B.; Obreht, I.; Sümegi, P.; Veres, D.; Boemke, B.; Schaubert, V.; Viehweger, J.; Hambach, U. Loess landscapes of Europe – mapping, geomorphology and zonal differentiation. Earth-Science Reviews 2021, 211, 1-82. [CrossRef]
- Sümegi, P.; Gulyás, S. (eds.). The geohistory of Bátorliget Marshland; Archaeolingua Press: Budapest, 2004; p. 353.
- Willis, K.J.; Rudner, E.; Sümegi, P. The full-glacial forests of central and southeastern Europe: Evidence from Hungarian palaeoecological records. Quaternary Research 2000, 53, 203-213. [CrossRef]
- Jacobson, G.L.; Bradshaw, R.H.W. The selection of sites for palaeovegetational studies. Quaternary Research 1981, 16, 80-96. [CrossRef]
- Prentice, I.C. Pollen representation, source area, and basin size: toward a unified theory of pollen analysis. Quaternary Research 1985, 23, 76-86. [CrossRef]
- Prentice, I.C.; Webb III, T. BIOME 6000: reconstructing global mid-Holocene vegetation patterns from palaeoecological records. Journal of Biogeography 1988, 25, 997-1005. [CrossRef]
- Prentice, I.C.; Cramer W., Harrison; S.P.; Leemans, R.; Monserud, R.A.; Solomon, A.M. A global biome model based on plant physiology and dominance, soil properties and climate. Journal of Biogeography 1992, 19, 1177-1134. [CrossRef]
- Magyari, E.K.; Kuneš, P.; Jakab, G.; Sümegi, P.; Pelánková, B.; Schäbitz, F.; Braun, M.; Chytrý, M. Late Pleniglacial vegetation in eastern-central Europe: are there modern analogues in Siberia? Quaternary Science Reviews 2014, 95, 60-79. [CrossRef]
- Buiron, D.; Stenni, B.; Chappellaz, J.; Landais, A.; Baumgartner, M.; Bonazza, M.; Capronc, E.; Frezzotti, M.; Kageyama, M.; Lemieux-Dudon, B.; Masson-Delmotte, B.; Parrenin, F.; Schilt, A.; Selmo, E.; Severi, M.; Swingedouwc, D.; Udisti, R. Regional imprints of millennial variability during the MIS 3 period around Antarctica. Quaternary Science Reviews 2012, 48, 99-112. [CrossRef]
- Bond, G.C.; Heinrich, H.; Broecker, W.; Labeyrie, L.; McManus, J.; Andrews, J.; Huon, S.; Jantschik, R.; Clasen, S.; Simet, C.; Tedesco, K.; Klas, M.; Bonani, G.; Ivy, S. Evidence for massive discharges of icebergs into the North Atlantic ocean during the last glacial. Nature 1992, 360, 245-249. [CrossRef]
- Genty, D.; Blamart, D.; Ouahdi, R.; Gilmour, M.; Baker, A.; Jouzel, J.; Van Exter, S. Precise dating of Dansgaard-Oeschger climate oscillations in Western Europe from stalagmite data. Nature 2003, 421, 833-837. [CrossRef]
- Grimm, E.C.; Jacobson, G.L.; Watts, W.A.; Hansen, B.C.S.; Maasch, K.A. A 50,000-Year Record of Climate Oscillations from Florida and Its Temporal Correlation with the Heinrich Events. Science 1993, 261, 198-200. [CrossRef]
- Grimm, E.C.; Watts, W.A.; Jacobson, G.L.; Hansen, B.C.S.; Almquist, H.R.; Dieffenbacher-Krall, A.C. Evidence for warm wet Heinrich events in Florida. Quaternary Science Reviews 2006, 25, 2197–2211. [CrossRef]
- Grootes, P.M.; Stuiver, M.; White, J.W.C.; Johnsen, S.J.; Jouzel, J. Comparison of oxygen isotope records from the GISP2 and GRIP Greenland ice cores. Nature 1993, 366, 552-554. [CrossRef]
- Svensson, A.; Andersen, K.K.; Bigler, M.; Clausen, H.B.; Dahl-Jensen, D.; Davies, S.M.; Johnsen, S.J.; Muscheler, R.; Parrenin, F.; Rasmussen, S.O. A 60 000 year Greenland stratigraphic ice core chronology. Climate of the Past 2008, 4, 47-57. [CrossRef]
- Timmermann, A.; Menviel, L.; Okumura, Y.; Schilla, A.; Merkel, U.; Timm, O.; Hu, A.; Otto-Bliesner, B.; Schulz, M. Towards a quantitative understanding ofmillennial-scale Antarctic warming events. Quaternary Science Reviews 2010, 29, 74-85. [CrossRef]
- Yiou, P.; Jouzel, J.; Johnsen, S.; Rögnvaldsson, Ö.E. Rapid oscillations in Vostok and GRIP Ice Cores. Geophyical Research Letters 1995, 22, 2179–2182. [CrossRef]
- Björck, S.; Walker, M.J.C.; Cwynar, L.C.; Johnsen, S.; Knudsen, K.L.; Lowe, J.J.; Wohlfarth, B. and INTIMATE Members. An event stratigraphy for the Last Termination in the North Atlantic region based on the Greenland ice-core record: A proposal by the INTIMATE group. Journal of Quaternary Science 1998, 13, 282-292. [CrossRef]
- Walker, M.J.C.; Björck, S.; Lowe, J.J.; Cwynar, L.C.; Johnsen, S.; Knudsen, K.L.; Wohlfarth, B. and INTIMATE GROUP. Isotopic “events” in the GRIP ice core: A stratotype for the Late Pleistocene. Quaternary Science Reviews 1999, 18, 1143–1150. [CrossRef]
- Kremenetski, K.V. Steppe and forest-steppe belt of Eurasia: Holocene environmental history. In Prehistoric steppe adaptation and the horse, Levine, M., Renfrew, C., Boyle, K., Ed.; McDonald Institute for Archaeological Research, University of Cambridge: Cambridge, 2003; pp. 11-27.
- Tímár, G.; Gábris, G. Estimation of water conductivity of the natural flood channels on the Tisza flood-plain, the Great Hungarian Plain. Geomorphology 2008, 98(3-4), 250-261. [CrossRef]
- Allen, J.R.M.; Watts, W.A.; Huntley, B. Weichselian palynostratigraphy, palaeovegetation and palaeoenvironment: the record from Lago Grande di Monticchio, southern Italy. Quaternary International 2000, 73-74, 91–110. [CrossRef]
- Tarasov, P.E.; Webb, T.III; Andreev, A.A.; Afanaseva, N.B.; Berezina, N.A.; Bezusko, L.G.; Blyakharchuk, T.A.; Bolikhovskaya, N.S.; Cheddadi, R.; Chernavskaya, M.M.; Chernova, G.M.; Dorofeyuk, N.I.; Dirksen, V.G.; Elina, G.A.; Filimonova, L.V.; Glebov, F.Z.; Guiot, J.; Gunova, V.S.; Harrison, S.P.; Jolly, D.; Khomutova, V.I.; Kvavadze, E.V.; Osipova, I.M.; Panova, N.K.; Prentice, I.C.; Saarse, L.; Sevastyanov, D.V.; Volkova, V.S.; Zernitskaya, V.P. Present-day and mid-Holocene biomes reconstructed from pollen and plant macrofossil data from the Former Soviet Union and Mongolia. Journal of Biogeography 1998, 25, 1029-1053. [CrossRef]
- Tarasov, P.E.; Volkova, V.S.; Webb, T.III; Guiot, J.; Andreev, A.A.; Bezusko, L.G.; Bezusko, T.V.; Bykova, G.V.; Dorofeyuk, N.I.; Kvavadze, E.V.; Osipova, I.M.; Panova, N.K.; Sevastyanov, D.V. Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from northern Eurasia. Journal of Biogeography 2000, 27, 609-620. [CrossRef]
- Szelepcsényi, Z.; Breuer, H.; Kis, A.; Pongrácz, R.; Sümegi, P. Assessment of projected climate change in the Carpathian Region using the Holdridge life zone system. Theoretical and Applied Climatology 2018, 131, 593-610. [CrossRef]
- Pełechata, A.; Pełechaty, M.; Pukacz, A. Factors influencing cyanobacteria community structure in Chara-lakes. Ecological Indicators 2016, 71, 477-490. [CrossRef]
- Vári, T.Z.; Pál-Molnár, R.; Sümegi P. Reconstructing the paleoenvironmental Evolution of Lake Kolon (Hungary) through Malacological, Macrobotanical and Pollen Analyses. Diversity 2023, This volume. [CrossRef]
- Várallyay, G. Soil water problems in Hungary. Agrokémia és Talajtan 1989, 38(3-4), 577-595.
- Bába, K. History of the investigation of the terrestrial snails of the Great Hungarian Plain and its present situation - II. Tiscia 1983, 18, 83-95.
- Molnár, B. Pliocén és pleisztocén lehordási területek az Alföldön. Földtani Közlöny 1960, 89, 403-413.
- Molnár, B. A magyarországi folyók homoküledékeinek nehézásvány-összetétel vizsgálata. Hidrológiai Közlöny 1964, 44, 347-355.
- Molnár, B. Changes in Area and Directions of Stream Erosion in the Eastern Part of the Hungarian Basin (Great Plain) during the Pliocene and Pleistocene. Acta Minerologica et Petrographica 1965, 17, 39-52.
- Molnár, B. A Dél-Alföld feltöltődésének ritmusai és vízföldtani jelentőségük. Hidrológiai Közlöny 1967, 47, 537-552.
- Molnár, B. Az Alföld harmadidőszak-végi és negyedkori feltöltődési ciklusai. Földtani Közlöny 1973, 103, 294-310.
- Magyari, E.; Jakab, G.; Rudner, E.; Sümegi, P. Palynological and plant macrofossil data on Late Pleistocen short term climatic osscilations in Northeast Hungary. Acta Palaeobotanica Supplement 1999, 2, 491-502.
- Sümegi, P. Reconstruction of flora, soil and landscape evolution, and human impact on the Bereg Plain from late, glacial up to the present, based on palaeoecological analysis. In The Upper Tisa Valley. Tiscia Monograph Series, Hamar, J., Sárkány-Kiss, A., Ed.; Szeged, 1999; pp. 173-204.
- Peterson, G.M. Recent pollen spectra and zonal vegetation in the western USSR. Quaternary Science Reviews 1983, 2(4), 281-321. [CrossRef]
- Szelepcsényi, Z.; Breuer, H.; Ács, F.; Kozma, I. Biofizikai klímaklasszifikációk (1. rész: a módszerek bemutatása). Légkör 2009a, 54, 21-26.
- Szelepcsényi, Z.; Breuer, H.; Ács, F.; Kozma, I. Biofizikai klímaklasszifikációk (2. rész: magyarországi alkalmazások). Légkör 2009b, 54, 21-26.
- Szelepcsényi, Z.; Breuer, H.; Sümegi, P. The climate of Carpathian Region in the 20th century based on the original and modified Holdridge life zone system. Central European Journal of Geosciences 2014, 6, 293-307. [CrossRef]
- Szilágyi, G.; Náfrádi, K.; Sümegi, P. A preliminary chronological study to understand the construction phases of a Late Copper–Early Bronze Age kurgan (kunhalom). Central European Geology 2019, 62(3), 1-29. [CrossRef]
- Németh, A.; Bárány, A.; Csorba, G.; Magyari, E.; Pazonyi, P.; Pálfy, J. Holocene mammal extinctions in the Carpathian Basin: A review. Mammal Review 2017, 47(1), 38-52. [CrossRef]
Figure 1.
Plantago schwarzenbergiana in the alkaline grassland.

Figure 2.
CORINE Land Cover map of the Hortobágy National Park – the Puszta World Heritage property (2018).
Figure 2.
CORINE Land Cover map of the Hortobágy National Park – the Puszta World Heritage property (2018).
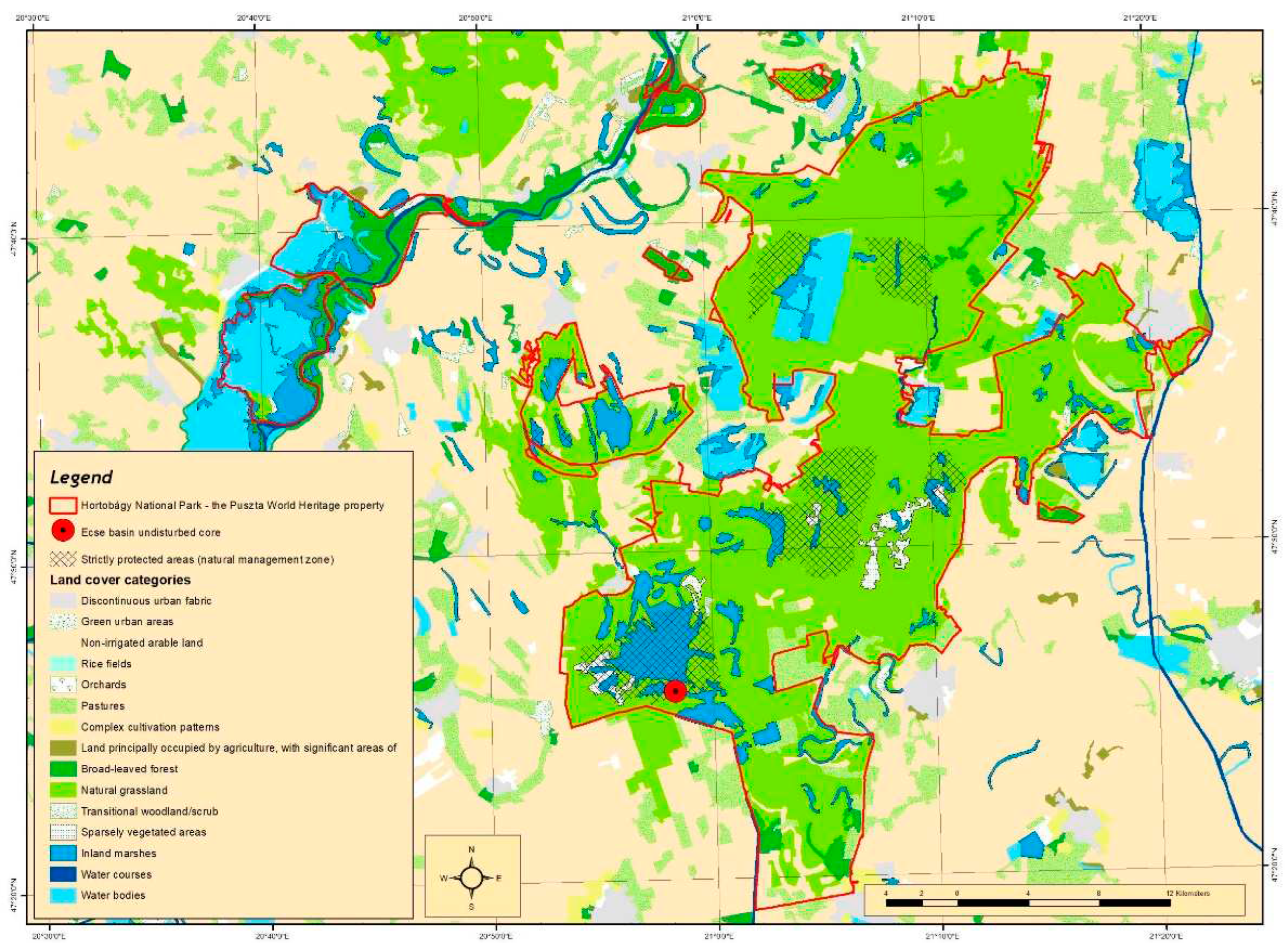
Figure 3.
Main area of salt accumulation in the northern part of the Trans-Tisza region, and the location of the undisturbed core points in the Carpathian Basin [27,28,29,30,31,32,33,34,35,36]. 1 = Kunkápolnás marsh; 2 = Halas Basin; 3 = Pap-ere; 4 = Fecske meadow; 5 = Lake Kolon; 6 = Lake Kiri; 7 = Lake Fehér; 8 = Pana-hát; 9 = Hajós; 10 = Kis-Mohos; 11 = Nagy-Mohos; 12 = Bátorliget fens; 13 = Nyírjes fen; 14 = Sarló-hát; 15 = Nádas Lake; 16 = Nyírjes Lake.
Figure 3.
Main area of salt accumulation in the northern part of the Trans-Tisza region, and the location of the undisturbed core points in the Carpathian Basin [27,28,29,30,31,32,33,34,35,36]. 1 = Kunkápolnás marsh; 2 = Halas Basin; 3 = Pap-ere; 4 = Fecske meadow; 5 = Lake Kolon; 6 = Lake Kiri; 7 = Lake Fehér; 8 = Pana-hát; 9 = Hajós; 10 = Kis-Mohos; 11 = Nagy-Mohos; 12 = Bátorliget fens; 13 = Nyírjes fen; 14 = Sarló-hát; 15 = Nádas Lake; 16 = Nyírjes Lake.
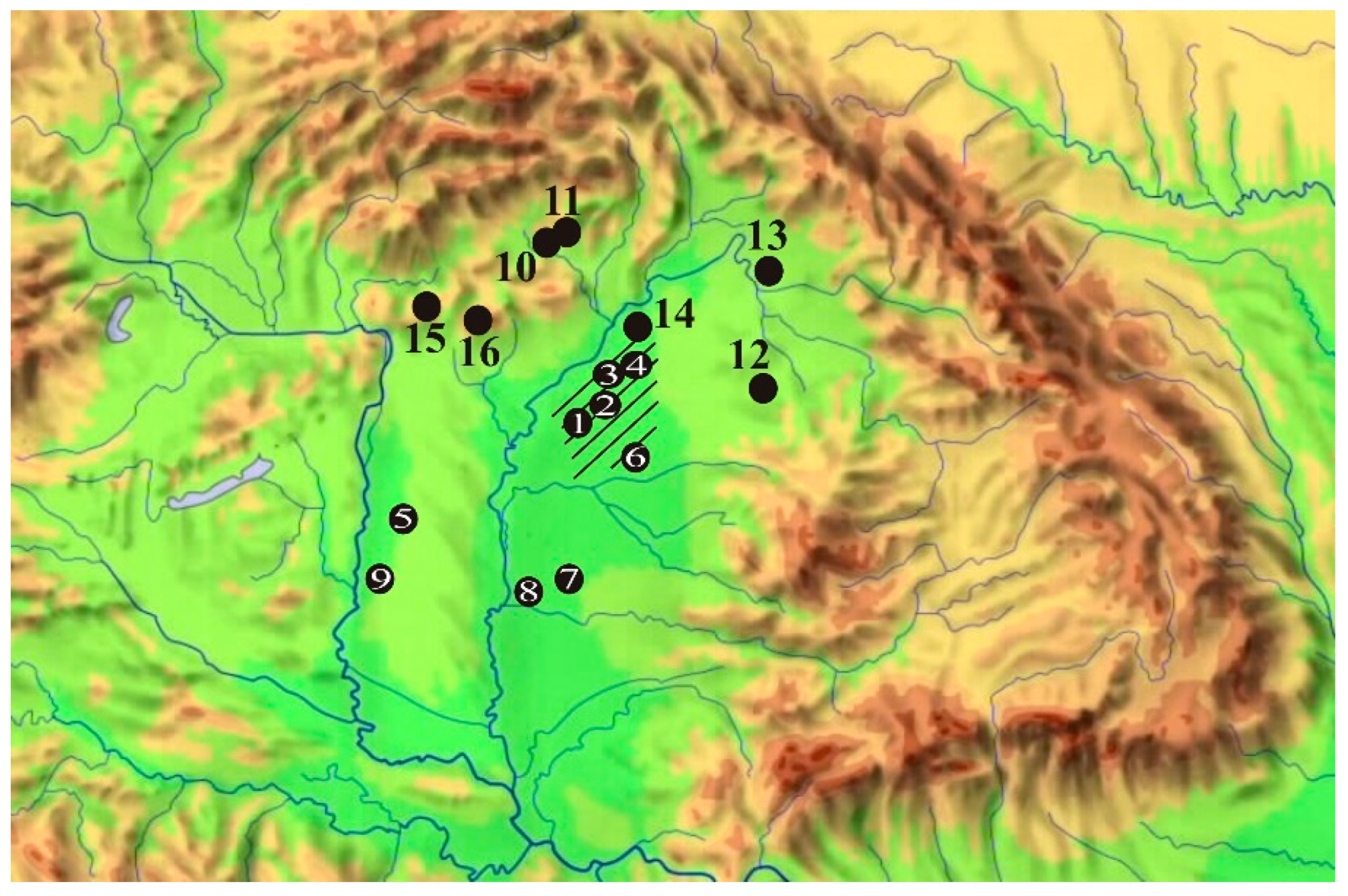
Figure 4.
Map of the wider Hortobágy region and surroundings [38] prepared before the major river regulations and creation of fishponds with the undisturbed core points. 1. Ecse mound near Kunmadaras (Yamnaja culture burial hill); 2 Kunkápolnás marsh near Kunmadaras with the analyzed paleochannel (Róna meadow); 3. Halas basin (paleochannel); 4. Faluvég mound (Yamnaja culture burial hill); 5. Kungyörgy Lake (paleochannel); 6. Szálka mound (Yamnaja culture burial hill); 7 = Fecske meadow (paleochannel); 8. = Pap-ere (paleochannel); 9. Csípő mound (Yamnaja culture burial hill).
Figure 4.
Map of the wider Hortobágy region and surroundings [38] prepared before the major river regulations and creation of fishponds with the undisturbed core points. 1. Ecse mound near Kunmadaras (Yamnaja culture burial hill); 2 Kunkápolnás marsh near Kunmadaras with the analyzed paleochannel (Róna meadow); 3. Halas basin (paleochannel); 4. Faluvég mound (Yamnaja culture burial hill); 5. Kungyörgy Lake (paleochannel); 6. Szálka mound (Yamnaja culture burial hill); 7 = Fecske meadow (paleochannel); 8. = Pap-ere (paleochannel); 9. Csípő mound (Yamnaja culture burial hill).
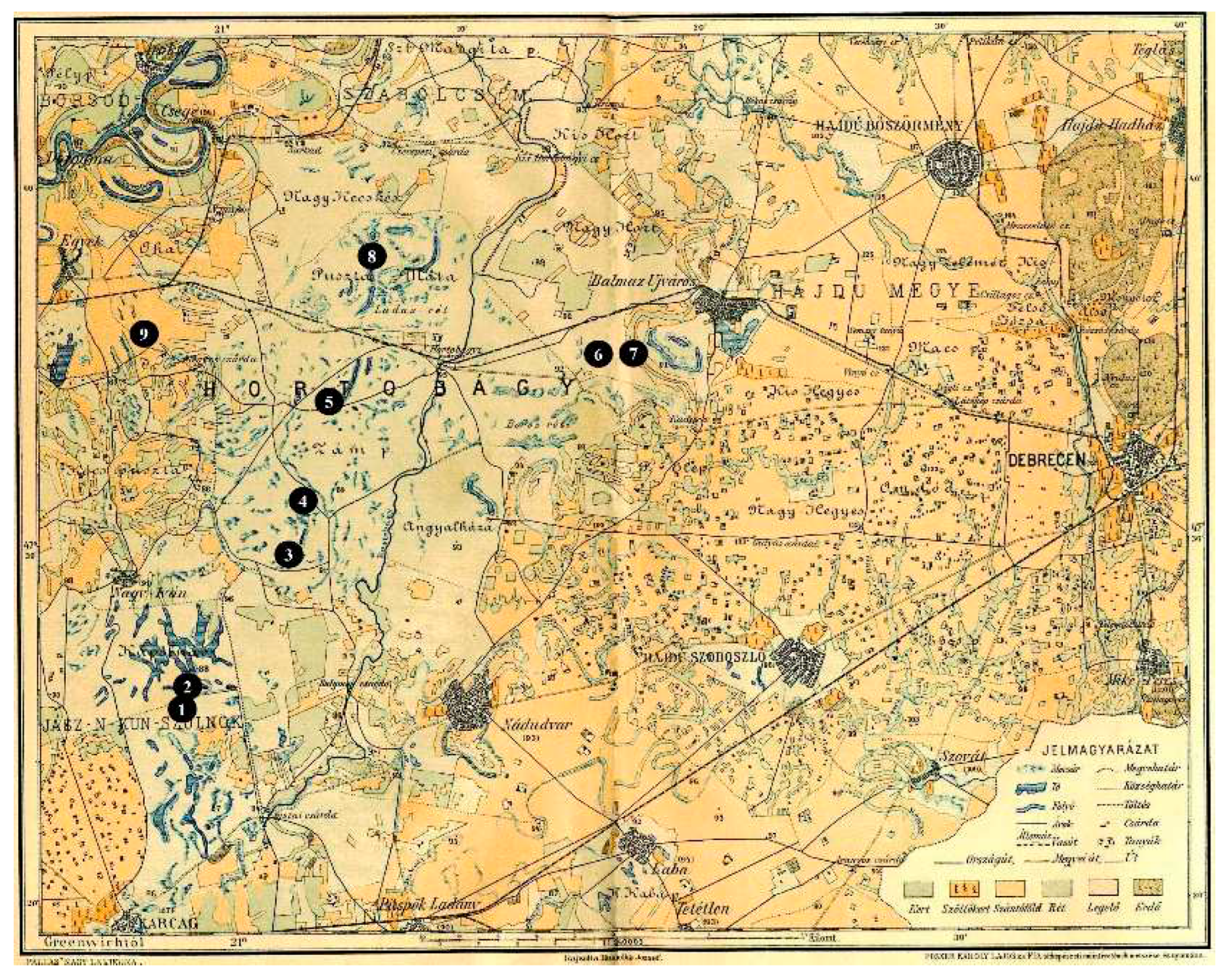
Figure 5.
Digital surface model of the study area ● undisturbed core point.
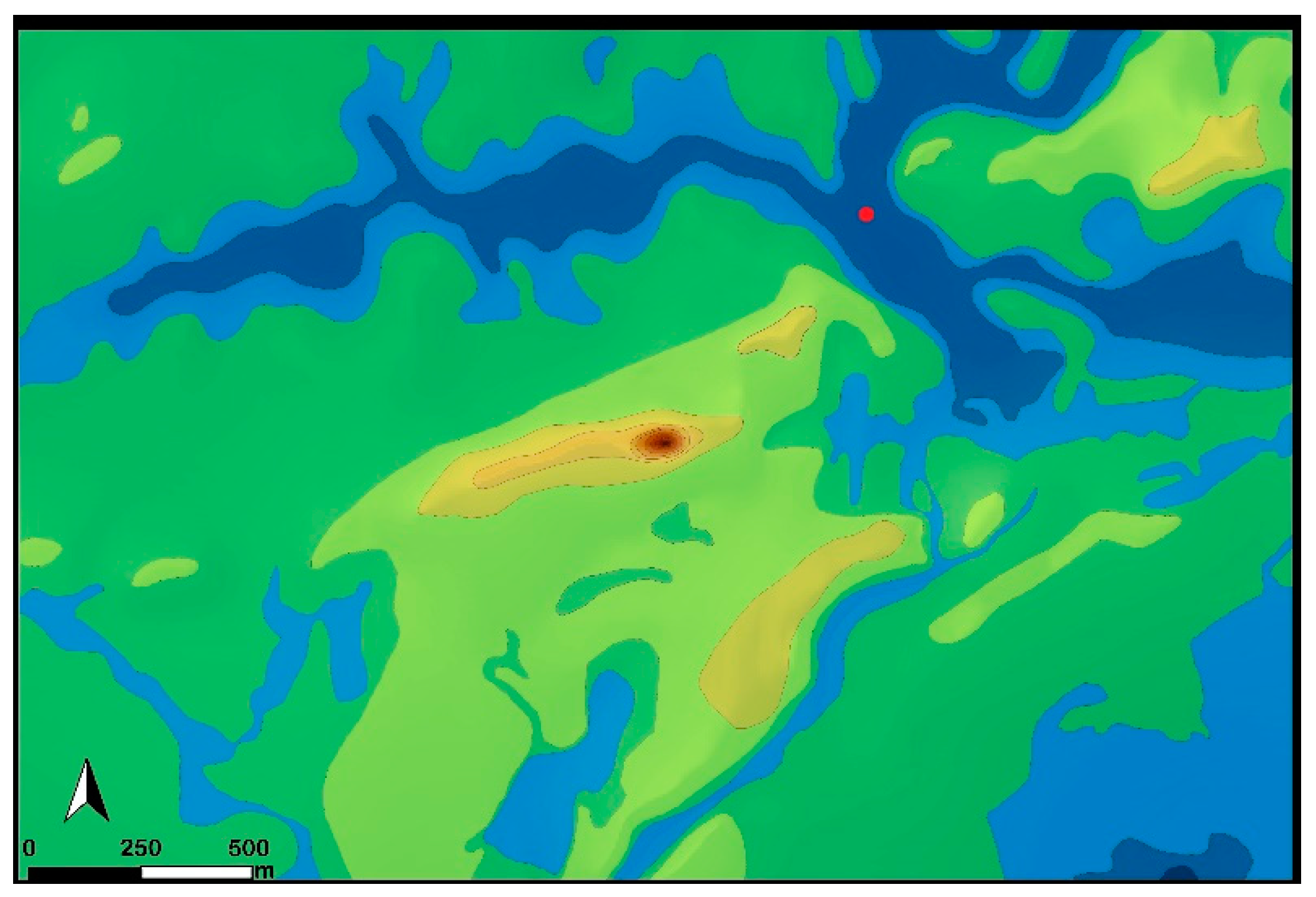
Figure 6.
Drone image of the studied area ● undisturbed core point.

Figure 7.
Age-depth modelling and the estimation of the sedimentation rate (accrate.depth) was conducted using rbacon 2.5.8 [53] in RStudio [54], and the IntCal20 calibration curve [51,55].
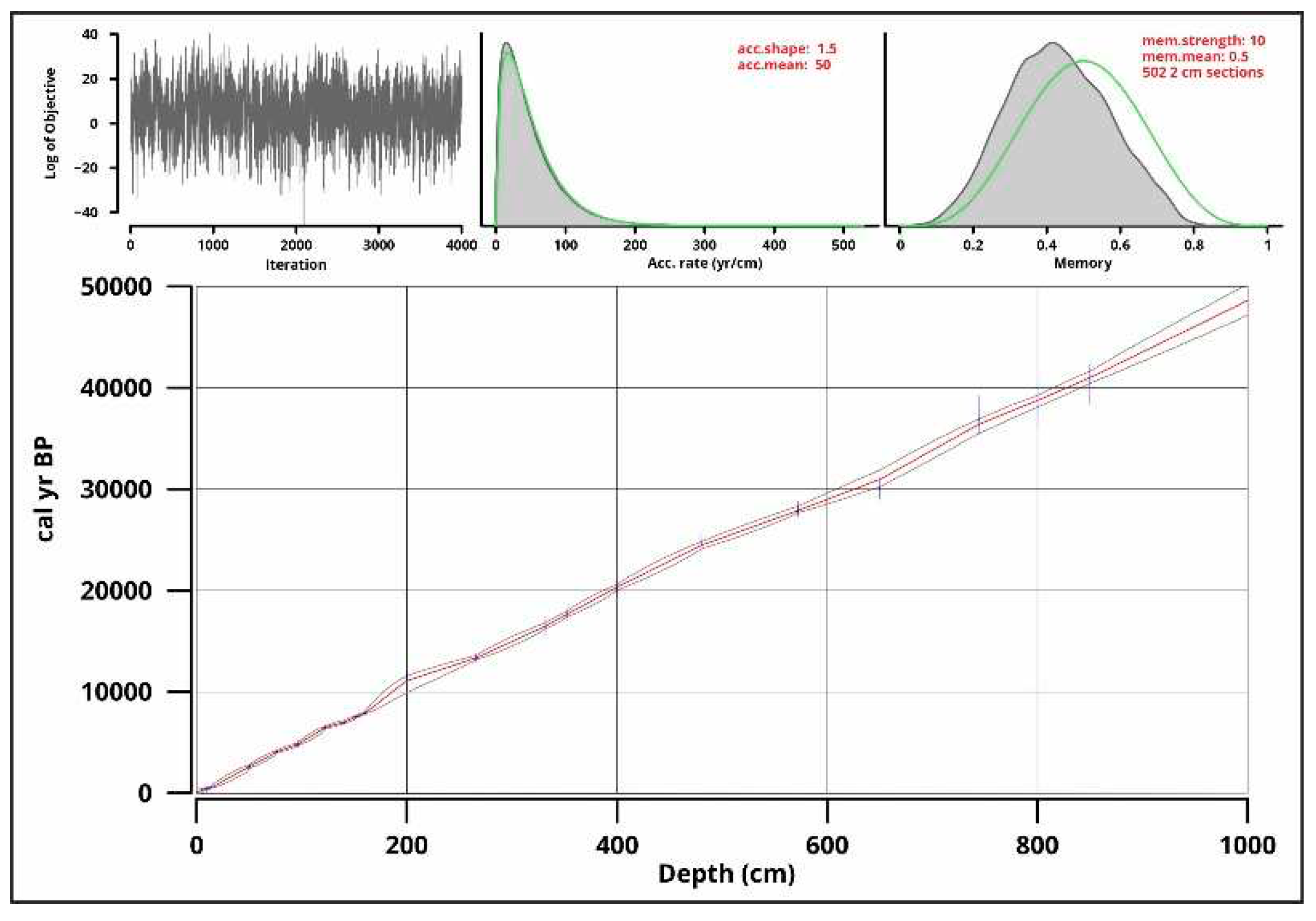
Figure 8.
Sedimentation rate (accrate.depth) made in RStudio [54] with rbacon [53] based on the results of the age-depth modeling.
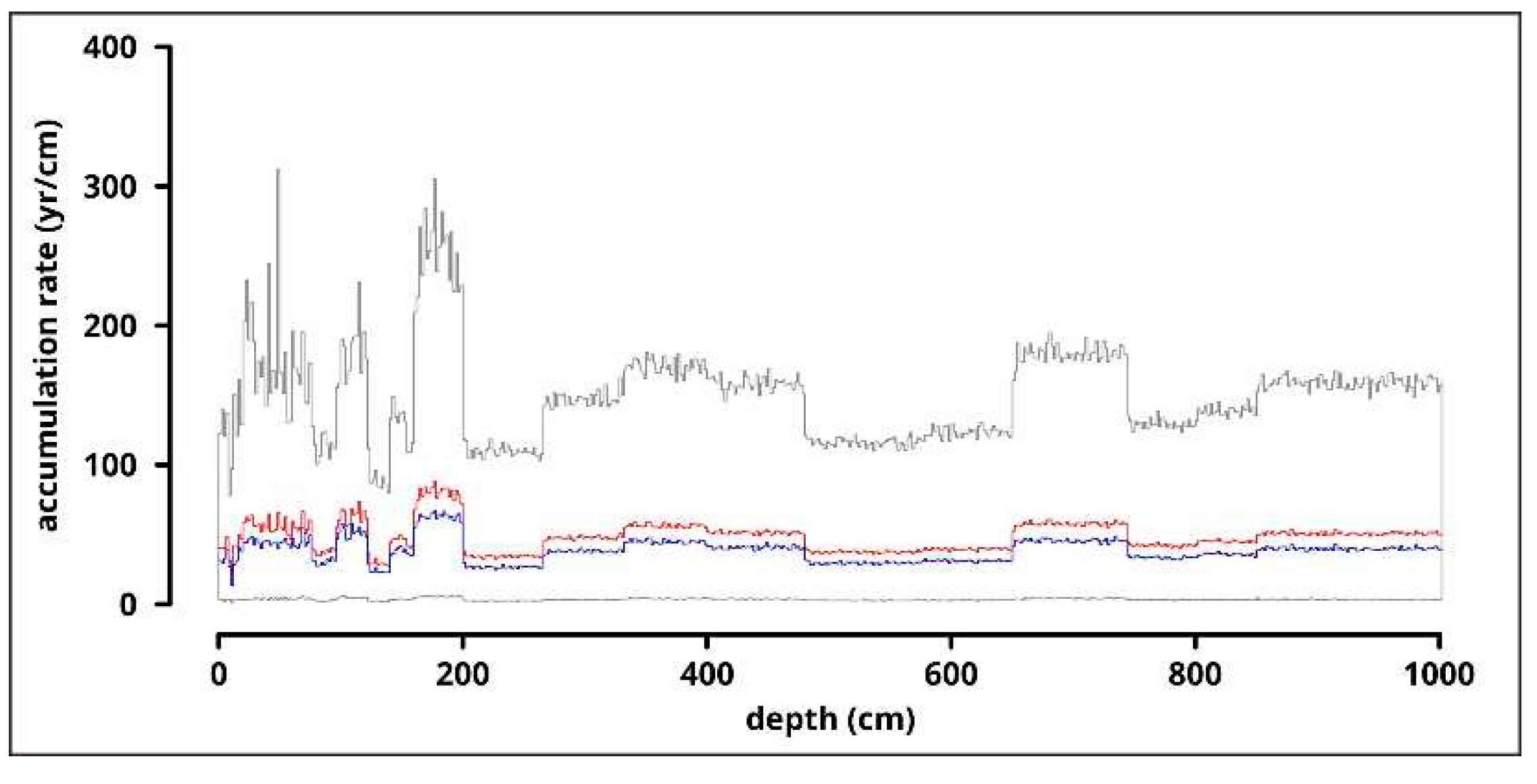
Figure 9.
Results of the grain size analysis: clay (<0.004 mm), silt (0.004-0.062 mm) and sand (0.062-0.5 mm) fractions in a percentage diagram and line diagrams of clay (<0.004 mm), very fine silt (0.004-0.008 mm), fine silt (0.008-0.016 mm), medium silt (0.016-0.031 mm), coarse silt (0.031-0.062 mm), very fine sand (0.062-0.125 mm), fine sand (0.125-0.25 mm) and medium sand (0.25-0.5 mm) fractions.
Figure 9.
Results of the grain size analysis: clay (<0.004 mm), silt (0.004-0.062 mm) and sand (0.062-0.5 mm) fractions in a percentage diagram and line diagrams of clay (<0.004 mm), very fine silt (0.004-0.008 mm), fine silt (0.008-0.016 mm), medium silt (0.016-0.031 mm), coarse silt (0.031-0.062 mm), very fine sand (0.062-0.125 mm), fine sand (0.125-0.25 mm) and medium sand (0.25-0.5 mm) fractions.
Figure 10.
Ternary diagram of clay, silt and sand grain sizes with Troels-Smith [40] sediment types based on the grain size analysis.
Figure 10.
Ternary diagram of clay, silt and sand grain sizes with Troels-Smith [40] sediment types based on the grain size analysis.
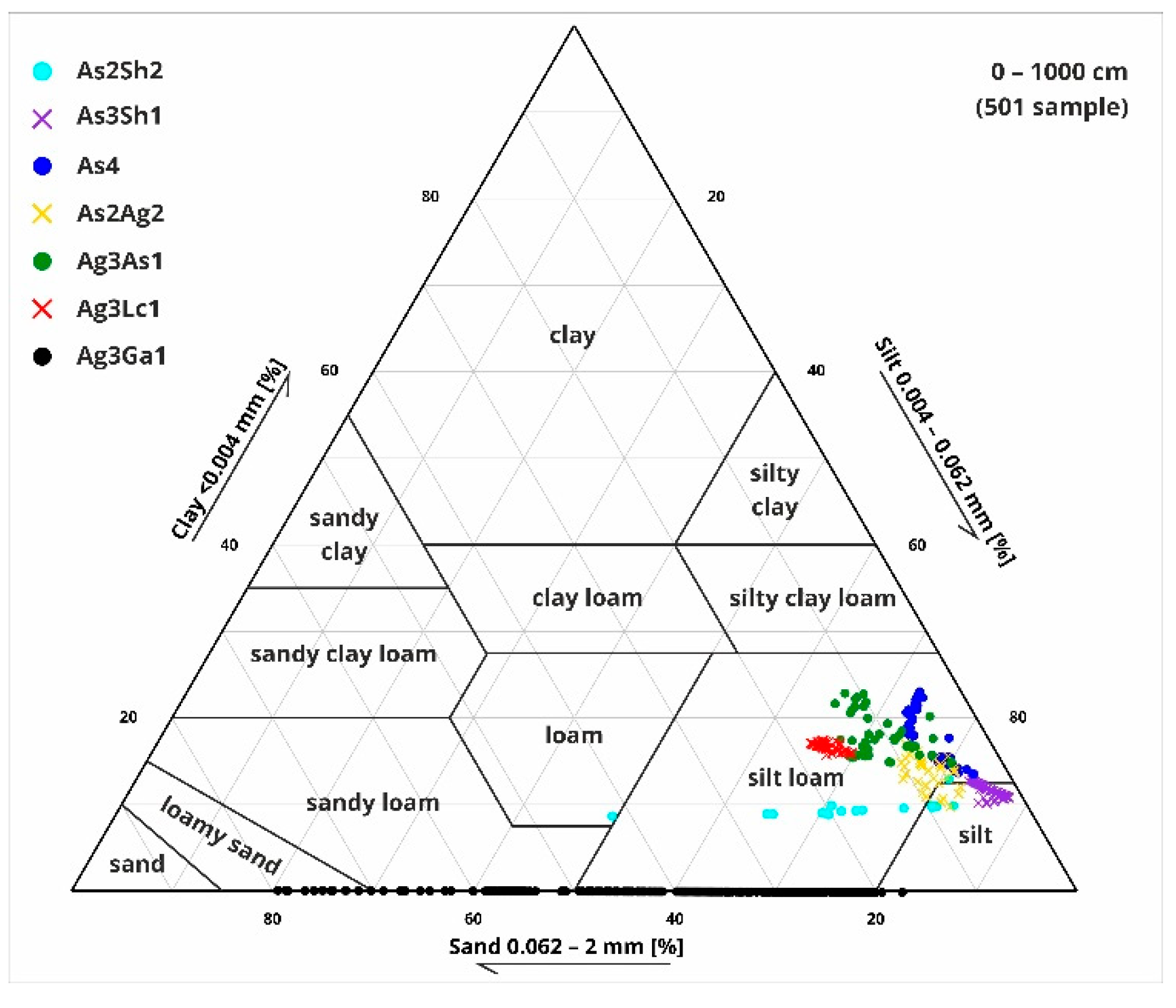
Figure 11.
Combined figure of (from left to right) the modeled median cal BP dates from the radiocarbon analysis; lithology profile based on grain size with Troels-Smith [40] sediment classification and depth (cm); results of the loss on ignition [43] with organic matter (OM), inorganic matter (IM) and carbonate content (CC); results of the geochemical [91,92] and magnetic susceptibility analysis [45].
Figure 11.
Combined figure of (from left to right) the modeled median cal BP dates from the radiocarbon analysis; lithology profile based on grain size with Troels-Smith [40] sediment classification and depth (cm); results of the loss on ignition [43] with organic matter (OM), inorganic matter (IM) and carbonate content (CC); results of the geochemical [91,92] and magnetic susceptibility analysis [45].
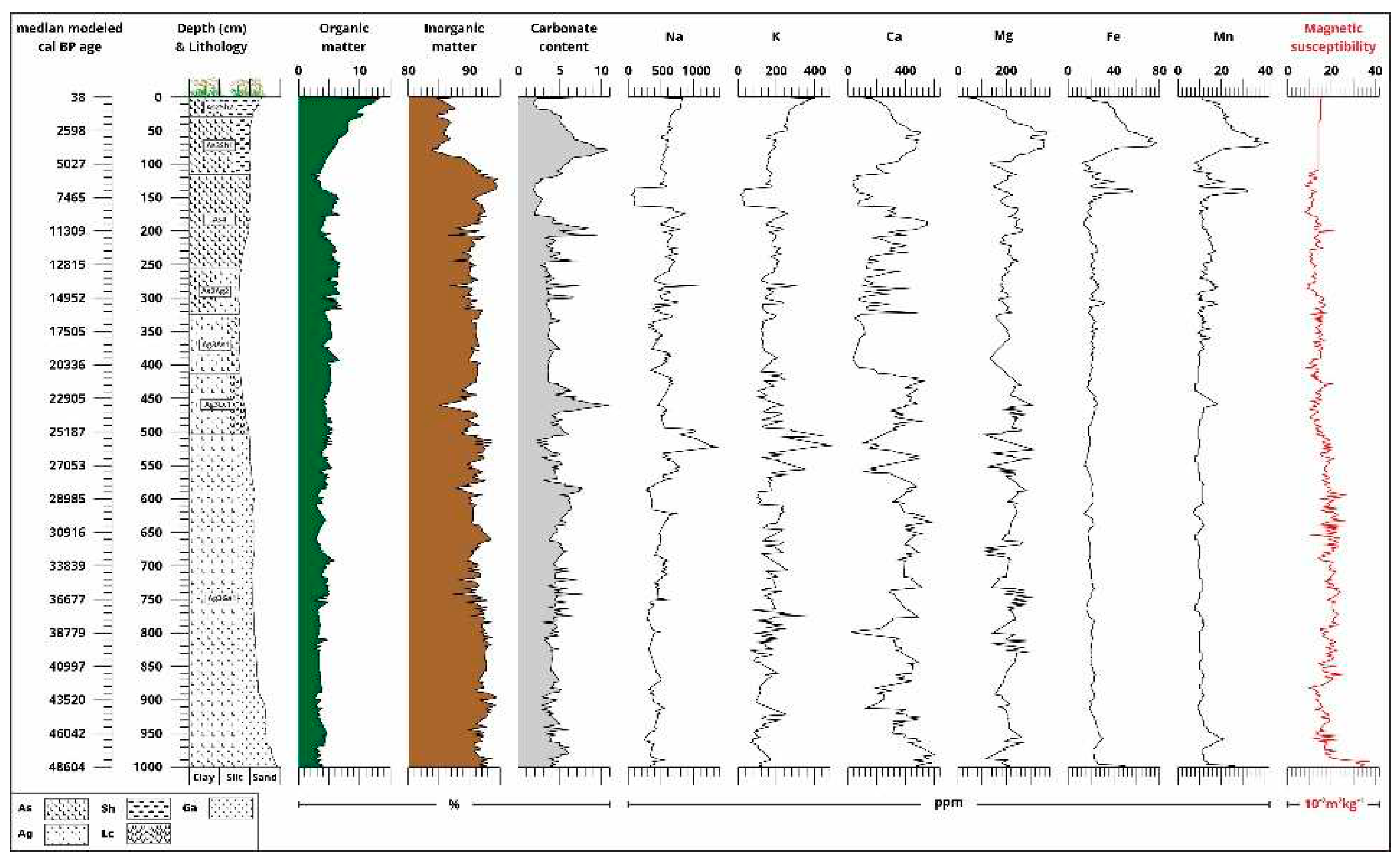
Figure 12.
Pollen diagram of Kunkápolnás of the undisturbed core sequence (selected taxa, and summarized group).
Figure 12.
Pollen diagram of Kunkápolnás of the undisturbed core sequence (selected taxa, and summarized group).
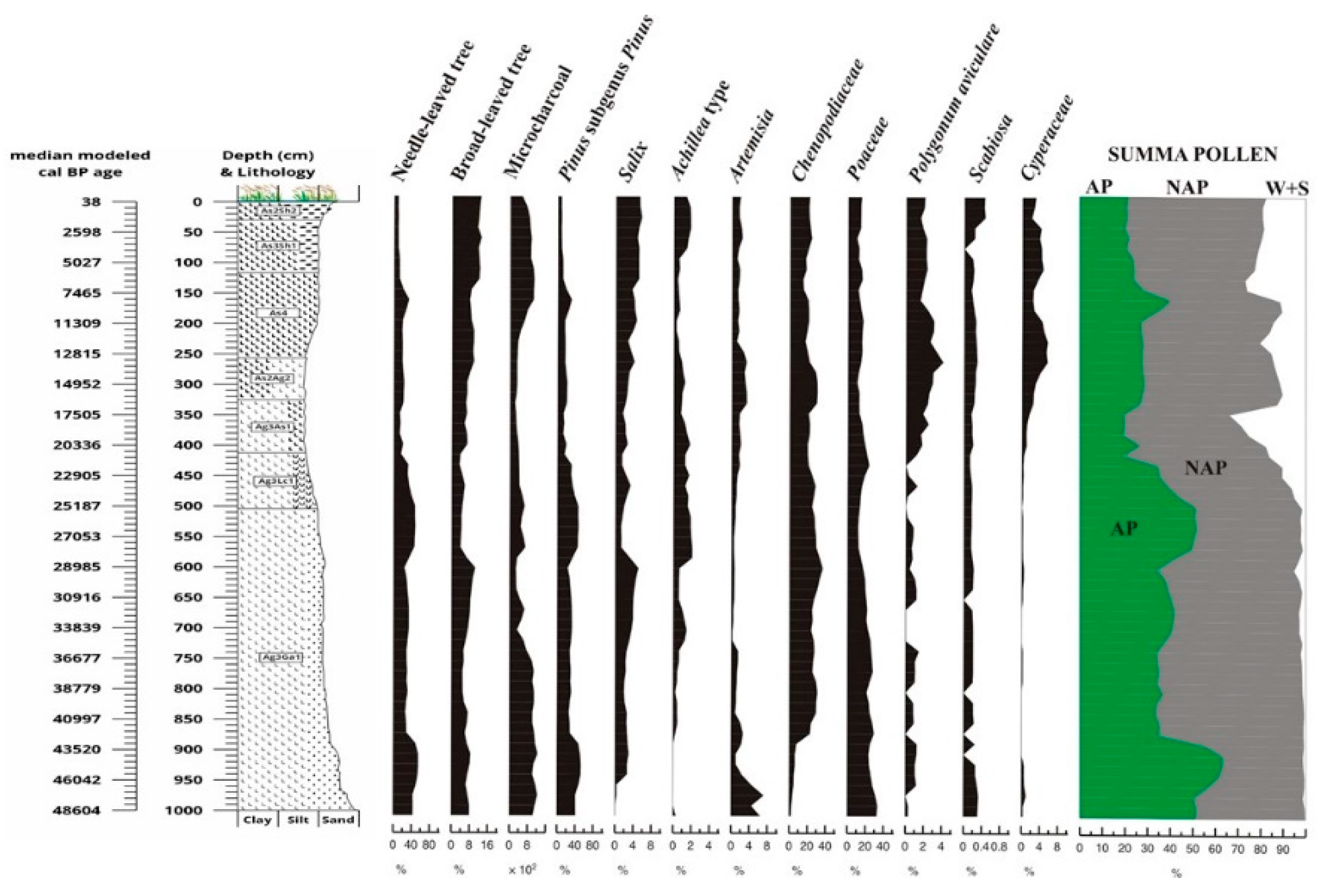
Figure 13.
Fauna and macrobotanical remains diagram of Kunkápolnás of the undisturbed core sequence.
Figure 13.
Fauna and macrobotanical remains diagram of Kunkápolnás of the undisturbed core sequence.
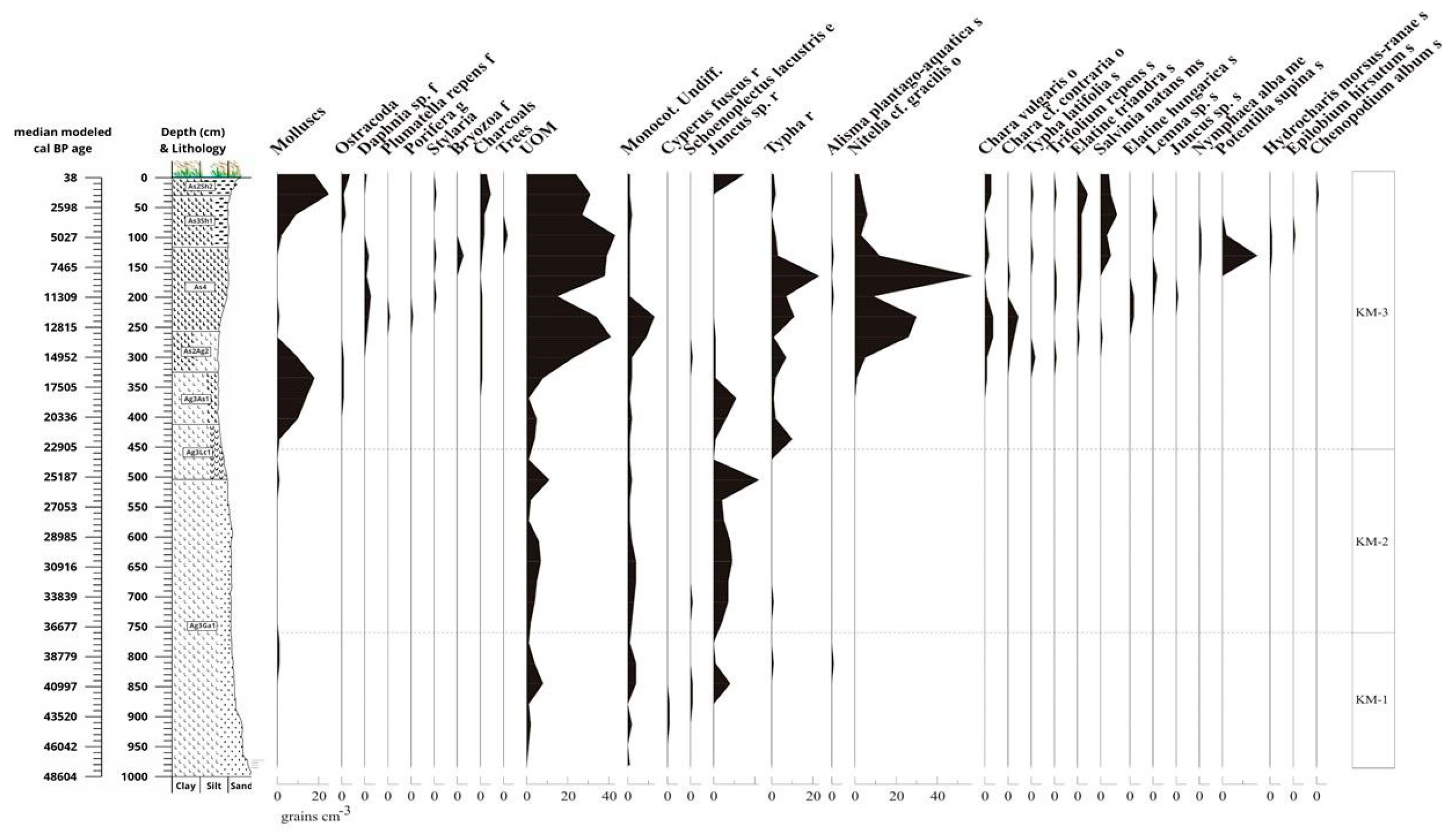
Figure 14.
Quartermalacological diagram 1: freshwater taxa in the Kunkápolnás undisturbed core sequence.
Figure 14.
Quartermalacological diagram 1: freshwater taxa in the Kunkápolnás undisturbed core sequence.
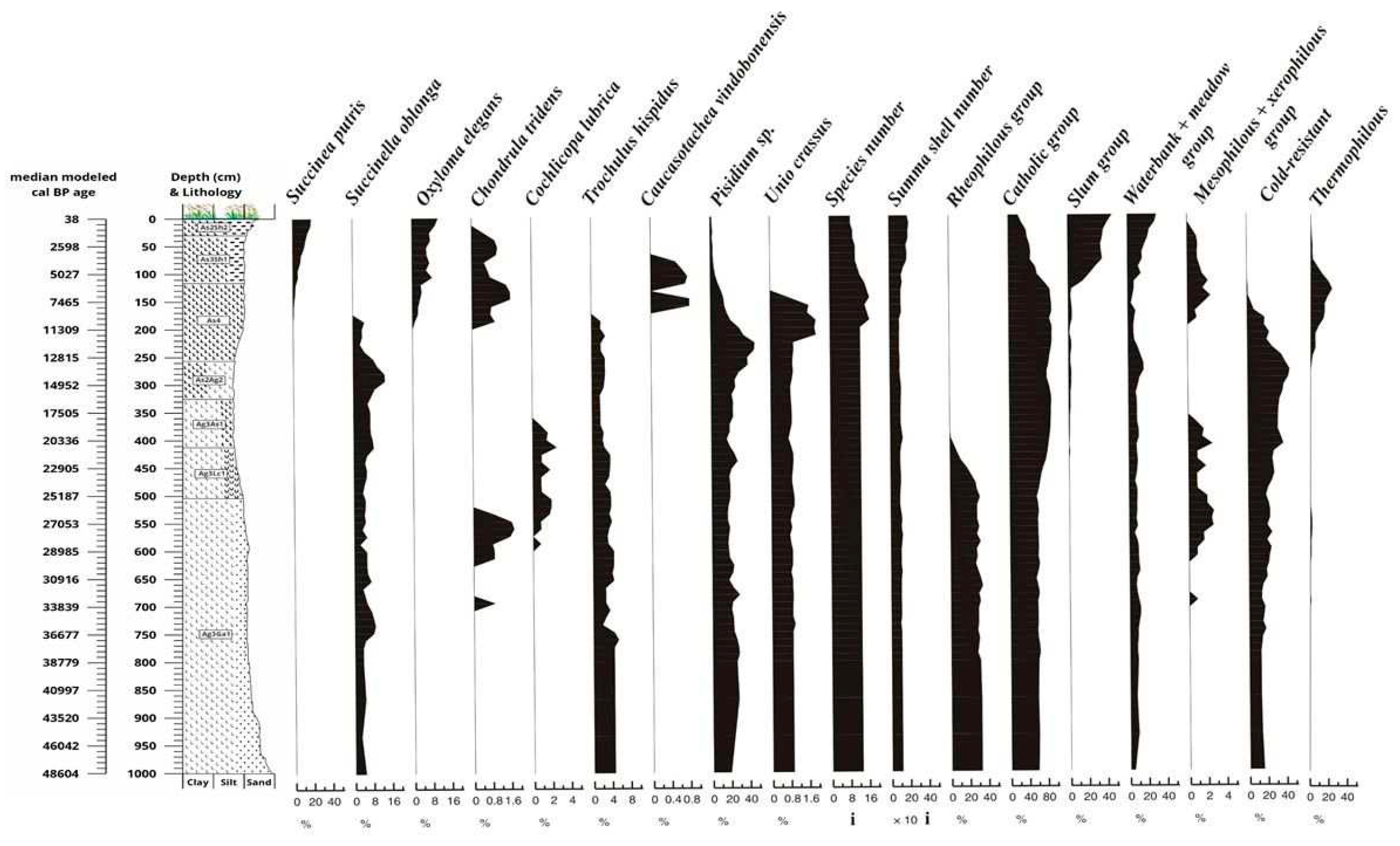
Figure 15.
Quartermalacological diagram 2: terrestrial taxa and quartermalacological based palaeoecological groups in the Kunkápolnás undisturbed core sequence.
Figure 15.
Quartermalacological diagram 2: terrestrial taxa and quartermalacological based palaeoecological groups in the Kunkápolnás undisturbed core sequence.
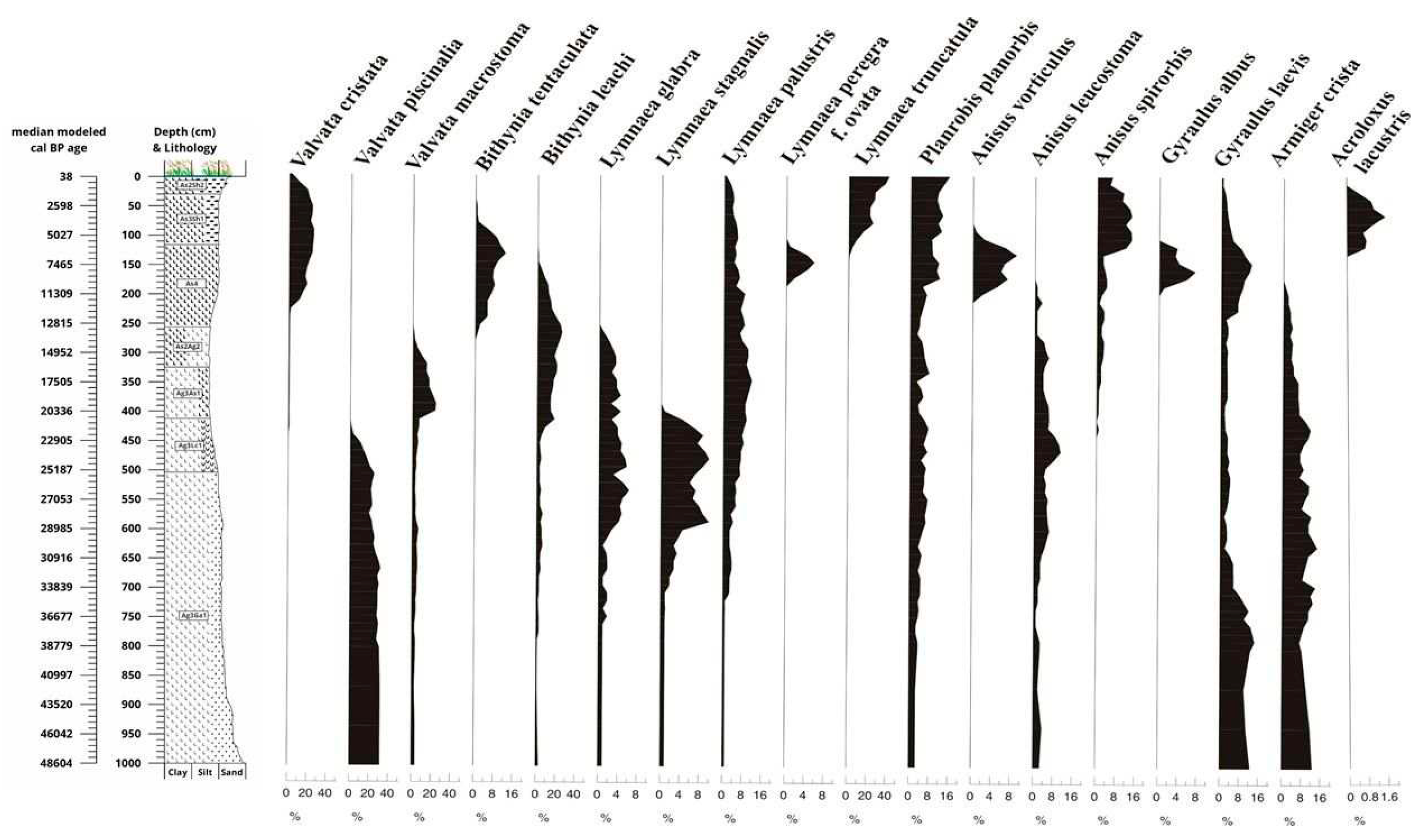
Figure 16.
Zone-average pollen frequencies of selected pollen types from Kunkápolnás marsh plotted alongside mean values of major pollen types in various vegetation zones of the former Soviet Union. Surface pollen spectra are redrawn from [135]. KKP-1 = pollen spectra from Dansgaard–Oschger (Greenland Interstadial) events in the sequence of the Kunkápolnás marsh; KKP-2 = pollen spectra from Heinrich (Greenland Stadial) events in the sequence of Kunkápolnás marsh; KKP-3 = pollen spectra from late-glacial age in the sequence of Kunkápolnás marsh; KKP-4 = pollen spectra from early postglacial time (Early Holocene Age) in the sequence of Kunkápolnás marsh; KKP-5 = pollen spectra from late postglacial time (Late Holocene Age) in the sequence of Kunkápolnás marsh.
Figure 16.
Zone-average pollen frequencies of selected pollen types from Kunkápolnás marsh plotted alongside mean values of major pollen types in various vegetation zones of the former Soviet Union. Surface pollen spectra are redrawn from [135]. KKP-1 = pollen spectra from Dansgaard–Oschger (Greenland Interstadial) events in the sequence of the Kunkápolnás marsh; KKP-2 = pollen spectra from Heinrich (Greenland Stadial) events in the sequence of Kunkápolnás marsh; KKP-3 = pollen spectra from late-glacial age in the sequence of Kunkápolnás marsh; KKP-4 = pollen spectra from early postglacial time (Early Holocene Age) in the sequence of Kunkápolnás marsh; KKP-5 = pollen spectra from late postglacial time (Late Holocene Age) in the sequence of Kunkápolnás marsh.
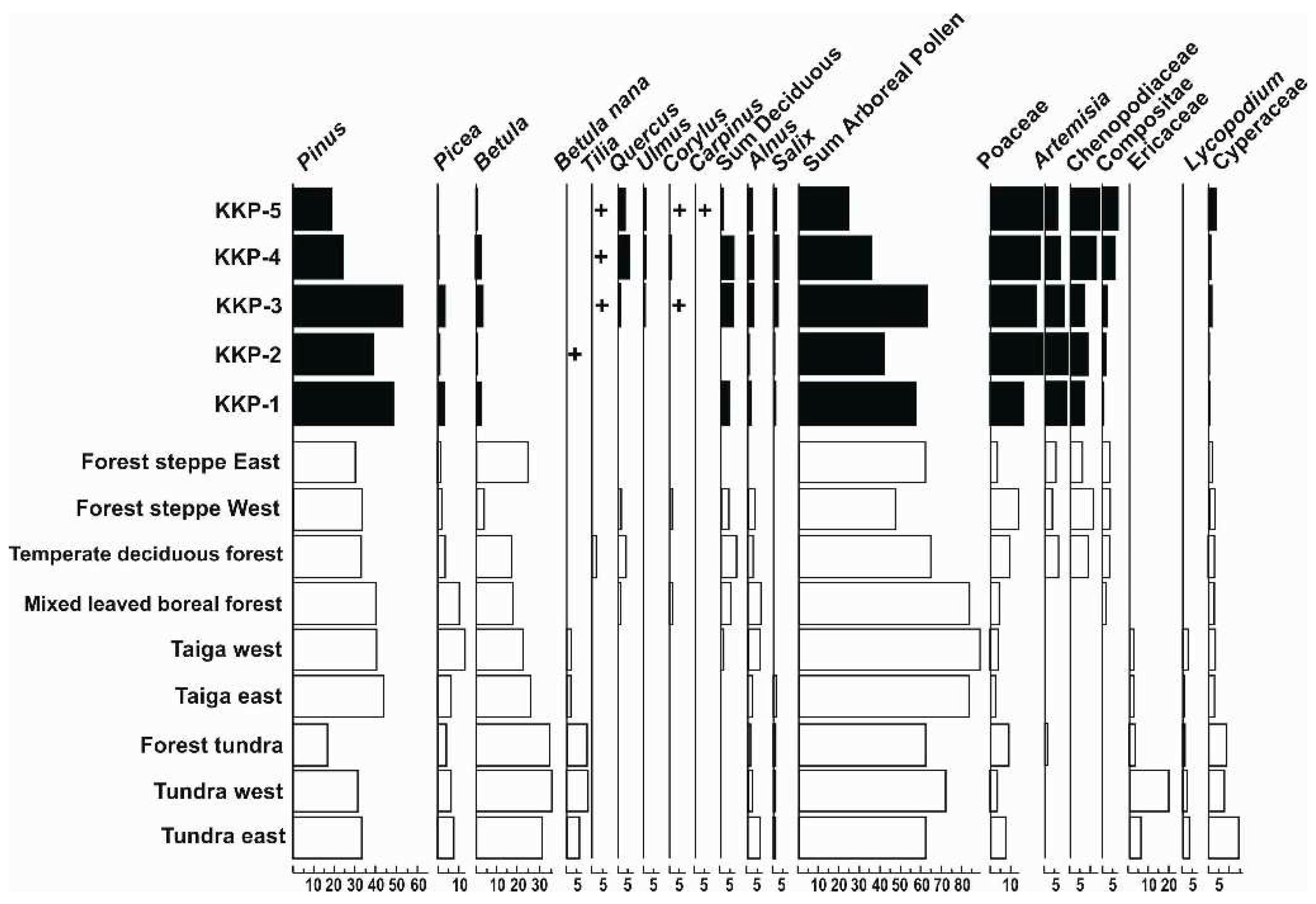
Figure 17.
Abundance of pollen of woody taxa in Holocene records from the Hortobágy and the Carpathian Basin. The range of values, as well as an indication of the most frequent value, is plotted for each site for 2 millennial intervals from 50,000 to 0 cal BP years. AP = Arboreal Pollen 1 = Kunmadaras, Kunkápolnás marsh; 2 = Hortobágy, Halas Basin; 3 = Hortobágy, Pap-ere; 4 = Balmazújváros, Fecske-rét; 5 = Izsák, Lake Kolon [27]; 6 = Ecsegfalva, Lake Kiri [28]; 7 = Kardoskút, Lake Fehér [29]; 8. Maroslele, Pana-hát [30]; 9.Hajós [31]; 10. Kelemér, Kis-Mohos [32]; 12. Bátorliget, fens [34]; 13. Csaroda, Nyíres fen [134]; 14. Tiszadob, Sarló-hát [33]; 15. Nagybárkány, Nádas Lake [35]; 16. Sirok, Nyírjes Lake [36].
Figure 17.
Abundance of pollen of woody taxa in Holocene records from the Hortobágy and the Carpathian Basin. The range of values, as well as an indication of the most frequent value, is plotted for each site for 2 millennial intervals from 50,000 to 0 cal BP years. AP = Arboreal Pollen 1 = Kunmadaras, Kunkápolnás marsh; 2 = Hortobágy, Halas Basin; 3 = Hortobágy, Pap-ere; 4 = Balmazújváros, Fecske-rét; 5 = Izsák, Lake Kolon [27]; 6 = Ecsegfalva, Lake Kiri [28]; 7 = Kardoskút, Lake Fehér [29]; 8. Maroslele, Pana-hát [30]; 9.Hajós [31]; 10. Kelemér, Kis-Mohos [32]; 12. Bátorliget, fens [34]; 13. Csaroda, Nyíres fen [134]; 14. Tiszadob, Sarló-hát [33]; 15. Nagybárkány, Nádas Lake [35]; 16. Sirok, Nyírjes Lake [36].
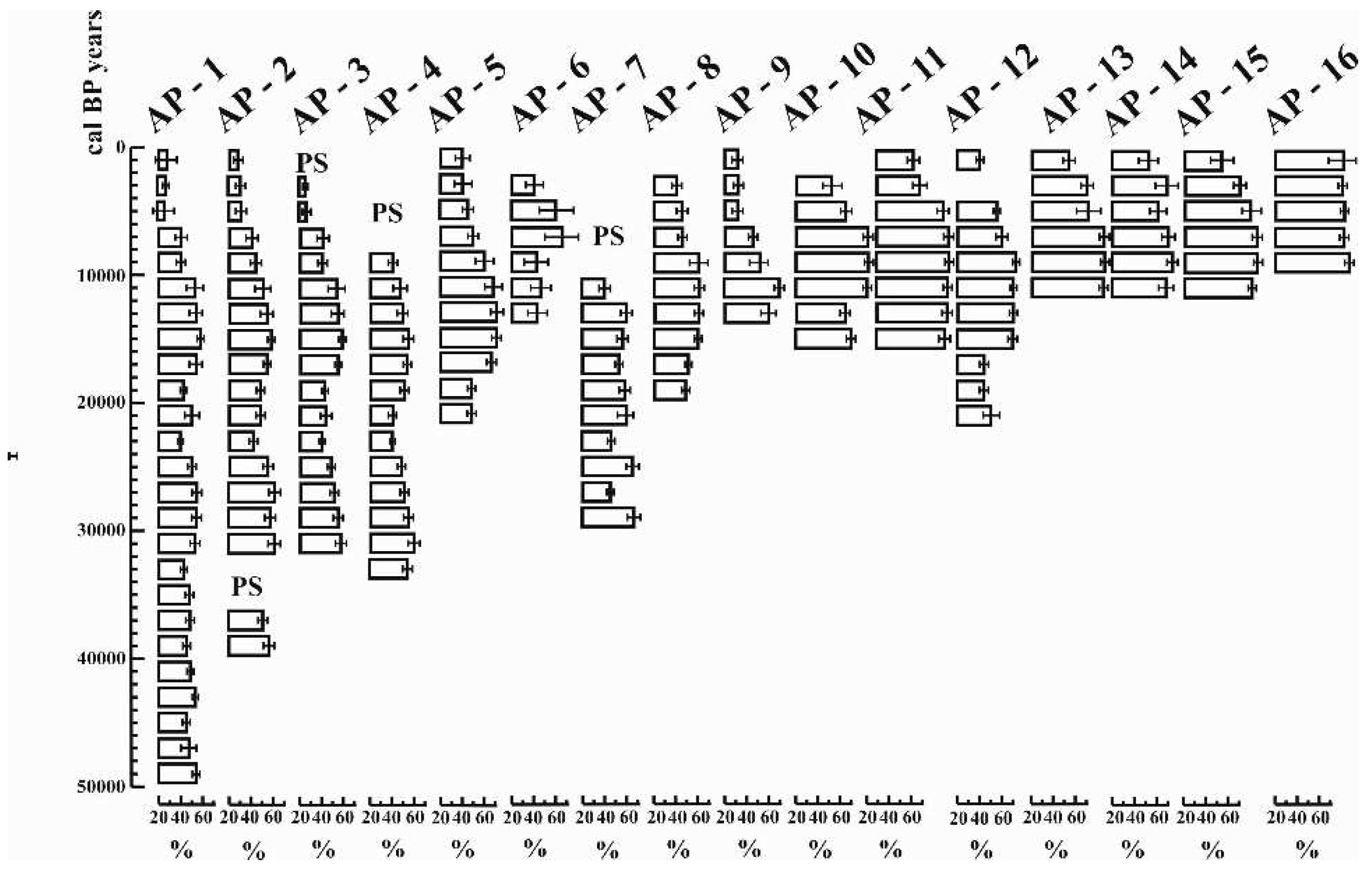
Figure 18.
The Holdrige type classification of the vegetation of the Carpathian Basin [123,136,137,138].
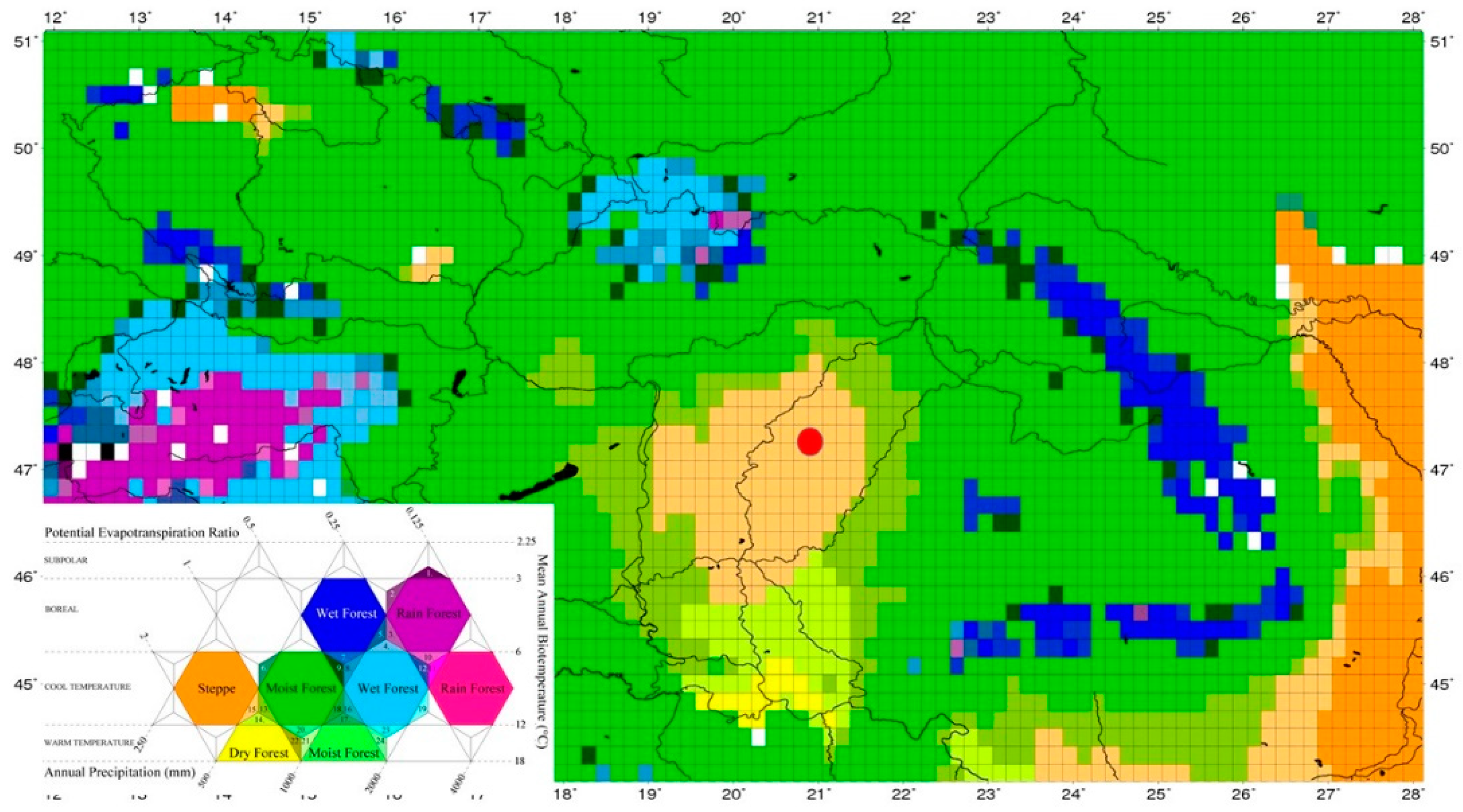

Table 1.
Twenty-one radiocarbon (AMS) data from undisturbed core sequence of the Kunkápolnás marsh in Hortobágy.
Table 1.
Twenty-one radiocarbon (AMS) data from undisturbed core sequence of the Kunkápolnás marsh in Hortobágy.
| cm | type of organic material | uncal BP | +/- | cal BP | +/- | cal BP interval | cal BC/AD | +/- | cal BC/AD interval | Code |
|---|---|---|---|---|---|---|---|---|---|---|
| (from 1950) | ||||||||||
| 10 | Planorbis shell | 403 | 17 | 422 | 84 | 339-506 | 1527 AD | 84 | 1444-1611 AD | DeA-130891 |
| 12 | Planorbis shell | 444 | 21 | 493 | 32 | 461-524 | 1448 AD | 22 | 1426-1469 AD | D-AMS21141 |
| 50 | Planorbis shell | 2524 | 23 | 2616 | 120 | 2496-2736 | 767 BC | 120 | 547 - 787 BC | D-AMS21152 |
| 76 | Planorbis shell | 3732 | 20 | 4169 | 82 | 3987-4151 | 2120 BC | 82 | 2038-2202 BC | DeA-130902 |
| 96 | Lymnaea shell | 4232 | 29 | 4753 | 104 | 4649-4857 | 2804 BC | 104 | 2700-2908 BC | D-AMS2113 |
| 122 | Unio shell | 5696 | 22 | 6479 | 73 | 6406-6552 | 4530 BC | 73 | 4457-4603 BC | DeA-130913 |
| 140 | Unio shell | 6065 | 26 | 6973 | 174 | 6799-7146 | 5023 BC | 174 | 4850-5197 BC | D-AMS21112 |
| 152 | Unio shell | 6698 | 30 | 7549 | 76 | 7504-7655 | 5628 BC | 73 | 5555-5701 BC | D-AMS21123 |
| 160 | Bithynia shell | 7067 | 29 | 7900 | 65 | 7836-7965 | 5949 BC | 67 | 5887-6016 BC | D-AMS21104 |
| 200 | Unio shell | 10,055 | 33 | 11,580 | 223 | 11,357-11,803 | 9629 BC | 225 | 9404-9854 BC | DeA-130914 |
| 266 | Pisidium shell | 11,417 | 52 | 13,292 | 118 | 13,174-13,411 | 11,343 BC | 118 | 11,225-11,462 BC | D-AMS21095 |
| 332 | Pisidium shell | 13,598 | 70 | 16,431 | 234 | 16,197-16,654 | 14,457 BC | 249 | 14,208-14,705 BC | D-AMS21086 |
| 352 | Cochlicopa shell | 14,474 | 58 | 17,645 | 233 | 17,412-17,878 | 15,696 BC | 233 | 15,463-15,929 BC | D-AMS21077 |
| 400 | Succinella shell | 16,847 | 78 | 20,342 | 191 | 20,151-20,535 | 18,262 BC | 159 | 18,203-18,521 BC | D-AMS21068 |
| 480 | Trochulus hispidus | 20,529 | 72 | 24,676 | 310 | 24,366-24,986 | 22,727 BC | 310 | 22,417-23,037 BC | DeA-130965 |
| 572 | Succinella shell | 23,725 | 85 | 27,851 | 157 | 27,694-28,008 | 25,902 BC | 157 | 25,745-26,059 BC | D-AMS21049 |
| 650 | Pinus microharcoal | 25,661 | 121 | 29,935 | 254 | 29,681-30,189 | 27,986 BC | 254 | 27,732-28,240 BC | DeA-131026 |
| 744 | Helicopsis shell | 32,535 | 175 | 36,845 | 481 | 36,364-37,325 | 34,896 BC | 481 | 34,415-35,376 BC | D-AMS210510 |
| 800 | Succinella shell | 33,433 | 232 | 38,308 | 853 | 37,455-39,161 | 36,359 BC | 853 | 35,506-37,212 BC | DeA-130977 |
| 850 | Trochulus shell | 35,696 | 297 | 40,740 | 583 | 40,158-41,323 | 38,791 BC | 583 | 38,209-39,374 BC | DeA-130988 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



