Submitted:
04 December 2023
Posted:
06 December 2023
You are already at the latest version
Abstract
Ipomoea batatas L. (IBL) has gained significant popularity as a complementary therapy or herbal medicine in the treatment of anti-diabetes. The objective of this review is to examine the mechanism of action of flavonoid compounds found in IBL that can activate GLP-1 as an anti-diabetic agent. The review article refers to PRISMA guidelines. The literature search was conducted using electronic databases such as Crossref, Pubmed, Scopus, and Science Direct. The search query was based on specific keywords, including Ipomoea batatas OR sweet potato AND anti-diabetic OR hypoglycemic. A total of 1055 articles were found, but only 32 articles were selected for further review based on inclusion and exclusion criteria. IBL contains various compounds, including phenolic acid, flavonols, flavanols, flavones, and anthocyanins, which exhibit activity against anti-diabetes. Flavonols, flavonols, and flavones belong to a group of flavonoids that possess the ability to form complexes with AlCl3 and Ca2+. Retention of Ca2+ within intracellular L cells, resulting in the release of GLP-1. Flavonols, flavones, and flavone groups have been found to strongly interact with DPP-IV, which inhibits the degradation of GLP-1. This mechanism effectively prolongs the half-life of GLP-1 in the systemic system, thereby contributing to the anti-diabetic activity of IBL
Keywords:
Antidiabetic
; GLP-1
; Ipomoea batatas L. flavonoid
; pharmacology
1. Introduction
Globally, the prevalence of diabetes has been reported to increase due to aging populations, population expansion, and increasing age-specific prevalences [1]. The World Health Organization predicts that diabetes will be the seventh leading cause of death worldwide by 2030, as the incidence of the disease has increased significantly in the recent years. According to the most recent projections of the International Diabetes Federation, in 2015 there were 415 million cases of diabetes worldwide, and by 2040, the number is predicted to reach 642 million [2].
A total of 1055 publications have been identified to examine/report the potential of Ipomoea batatas L. (IBL) as an altenative method for diabetes treatment (Figure 2). These publications are categorized into 12 distinct research clusters, which are distinguished by the size of their nodes. The majority of the extensive cluster focuses on Ipomoea batatas L. as an antidiabetic and antioxidant, while a smaller portion delves with only one group reporting into the mechanism of Glucagon-like peptide-1 (GLP-1) on IBL.
All published articles searched for our review mentione that IBL contains various active compounds, including flavonoids, anthocyanins, phenolic acids, caffeoyl derivatives, triterpenoids, and alkaloids. Each compound is still discussable regarding its pharmacological mechanism. Recent reports indicate that flavonoid compounds have demonstrated to have efficacy against DM. The occurrence effect of flavonoid consumption to reduce DM is signaled by several signaling pathways, namely glucose transporter, liver enzymatic, tyrosine kinase inhibition, AMPK, PPARγ, and NF-kB [3]. Currently, 27 IBL cultivars have been identified to have anti-diabetic mechanisms. These compounds have been shown to exert pharmacological effects at various sites of action, including the liver, pancreas, skeletal muscle, and adipose tissue [4]. The pharmacological mechanism of IBL in the treatment of diabetes involves multi-chemical and multi-pharmacological sites.
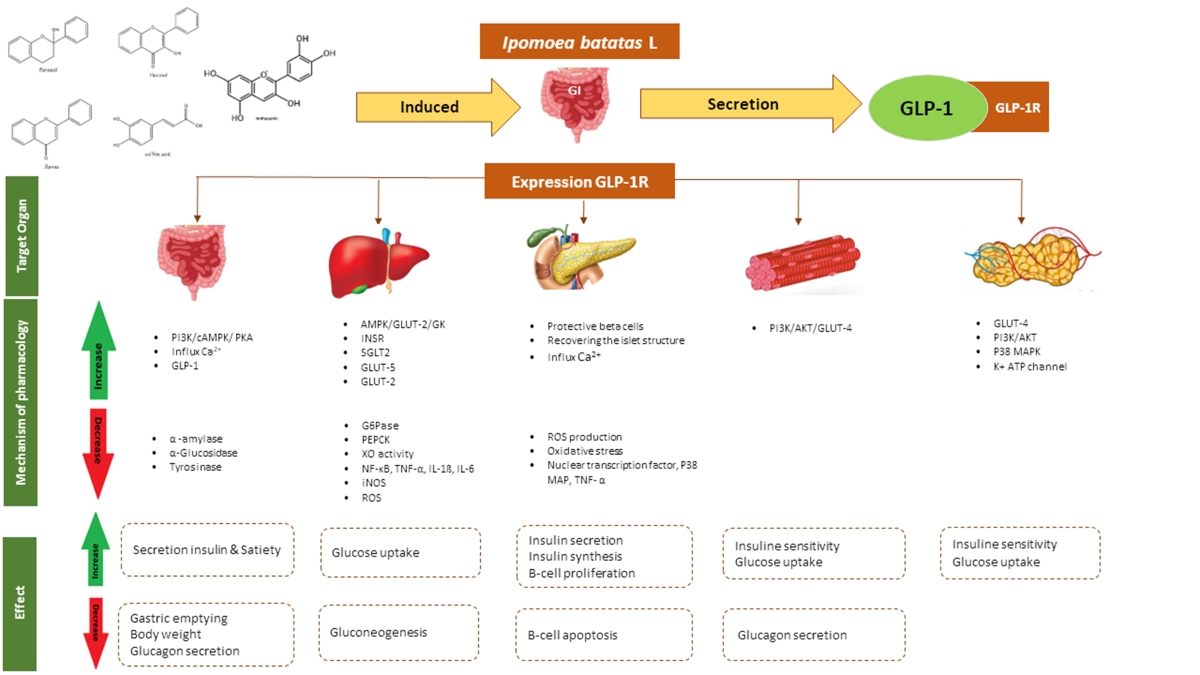
The compounds that have been identified are predicted to work in the gastrointestinal (GI) tract by α-glucosidase inhibition through degradation of polysaccharides into monosaccharides and secretion of GLP-1. It can lead to a reduction in gastric emptying, a decrease in gastrointestinal motility, and an increase in insulin secretion. [5,6]. These findings also exhibit an impact on the reduction of cholesterol and blood pressure, both of which are recognized as risk factors for DM based on the resultant result.
A study conducted by Nagamine et al. in 2014 found that IBL extract has the ability to induce the release of GLP-1, thereby making it an effective anti-diabetic agent. In this review article, we will analyze and discuss the potential and mechanism of action of flavonoid compounds present in IBL that can activate the GLP-1.
2. Materials and Methods
2.1. Search Strategy
The search approach was carried out in accordance with previously stated Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) criteria [7]. A systematic review was conducted to find out relevant articles on the therapeutic potential of IBL in anti-diabetic treatment. The literature search was performed extensively across several selected databases such as Crossref, Pubmed, Scopus, and Science Direct. The set of keywords included (1) Ipomoea batatas OR sweet potato AND (2) anti-diabetic OR hypoglycemic.
2.2. Inclusion Criteria
We selected research articles that focused on the effects, anti-diabetic potentials, phytochemical compounds, and signaling mechanisms of Ipomoea batatas. These articles were based on in vitro and in vivo studies. The chosen articles had to be written in English and include abstracts. The selected studies evaluated at least three essential measures: (1) Ipomoea batatas, (2) phytochemical compounds, and (3) signaling mechanisms involved.
2.3. Exclusion Criteria
Conference papers, thesis dissertations, review articles, papers published for conferences, manuscripts without abstracts, and manuscripts that did not adhere to the mentioned inclusion requirements were all excluded. Studies that examined Ipomoea batatas in relation to other diseases were additionally excluded from this analysis.
2.4. Data Extraction and Management
The article that were chosen to be included in this study were complied using Zotero, a reference manager. The publications that satisfied the requirements for inclusion were then examined. Information gathered included (1) type/cultivar, (2) part of the plant, (3) Identified compound, (4) predicted bioactive compound, (5) site of action, (6) mechanism pharmacology of anti-diabetic activity of Ipomoea batatas.
2.5. Strategy for Data Extraction
The results of this literature review focus on in vitro and in vivo studies of I. batatas in type 2 diabetes. Section 3 and 4 describes the results of reports regarding the phytochemicals involved. In Section 5 and Section 6, an analysis of the findings is provided, considering the sites of action and pharmacological mechanisms involved in the treatment of diabetes.
3. Results
The Literature Search
The literature search identified 1055 articles relevant to the topic (Figure 1). Duplications detected and removed were 44 articles. Based on screening titles and abstracts, 865 articles were removed. Then, the results of the screening of the inclusion criteria mentioned above, 125 articles were further excluded. A total of 32 appropriate articles will were reviewed in more depth. Successful data extraction is shown in the flow diagram displayed in Figure 1.
Figure 1.
Flow chart identification and screening literature search.
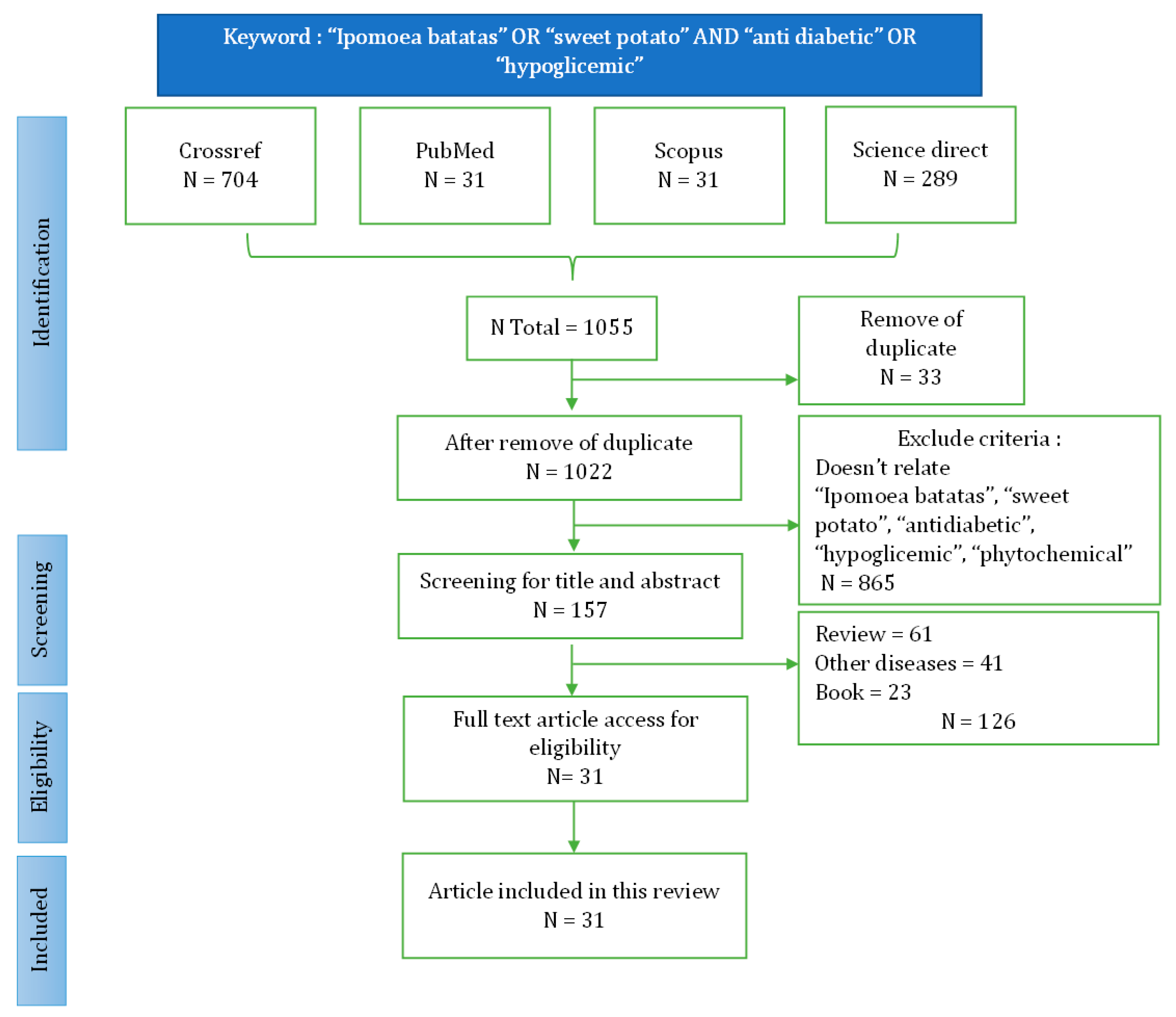
Figure 2.
Keyword co-occurence network.
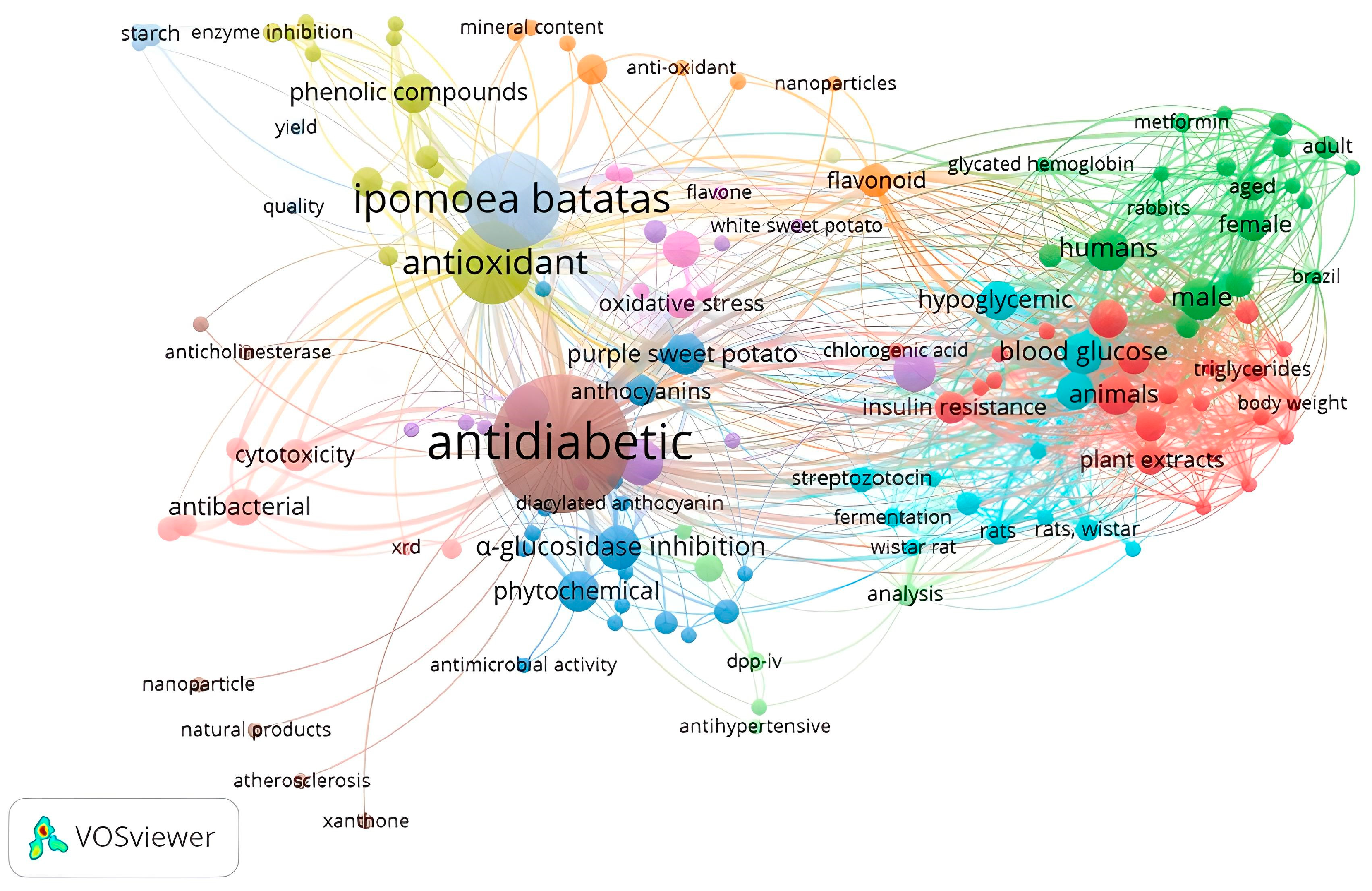

Table 1.
Type/cultivar of Ipomoea batatas, predictive bioactive compound, site of action activity, and pharmacology mechanism.
Table 1.
Type/cultivar of Ipomoea batatas, predictive bioactive compound, site of action activity, and pharmacology mechanism.
| No | Type/cultivar | Part of plant | Identified compound | Predictive Bioactive compound | Site of action | Mechanism pharmacology | Reference |
| [8]1 | IBL from cultivar Simon No.1(Beijing, China) | Leaves |
|
- | Pancreas |
|
[8] |
| - | Liver |
|
|||||
| - | Muscle |
|
|||||
| Leaves | - |
|
Gastrointestinal |
|
[9] | ||
| Pancreas |
|
||||||
| 2 | Purple Ipomoea batatas were collected in Luzhu District, Taoyuan City, Taiwan |
Leaves |
|
Adipose |
|
[10] | |
| 3 | Ipomoea batatas were purchased from the local market India | Leaves |
|
Gastrointestinal |
|
[11] | |
| 4 | Purple Ipomoea batatas were collected from Aan Village, Klungkung Regency, Bali Province, Indonesia |
Leaves |
|
Pancreas |
|
[12] | |
| 5 | Fresh orange-fleshed SPL (Jishu No. 16) collected from the farm in Yichun | Leaves |
|
|
Pancreas |
|
[13] |
|
Gastrointestinal |
|
|||||
| 6 |
Ipomoea batatas leaves were obtained by local farmer (Hebei province) in autumn |
Leaves |
|
Pancreas |
|
[14] | |
| 7 | Ipomoea batatas was cultivated in Slatina (central Croatia). | Leaves |
|
Pancreas |
|
[15] | |
| 8 | Ipomoea batatas L. collected in October 2015 in Anguillara Veneta (Northern Italy) | Leaves |
|
|
Gastrointestinal |
|
[16] |
| Pancreas |
|
||||||
| 9 | Fresh leaves of Ipomoea batatas (family of clones B 00593) were harvested in July from the Bandungan plantation area in Central Java Indonesia. |
Leaves |
|
Pancreas |
|
[17] | |
| 10 | ‘Suioh,’ a IBL cultivar developed in Kumamoto prefecture, Japan | Leaves |
|
|
Gastrointestinal |
|
[18] |
| 11 | Purple IBL | - |
|
|
Gastrointestinal |
|
[19] |
| 12 | ‘Bophelo’ Orange-fleshed IBL cultivar | Tubers and leaves |
|
|
Pancreas |
|
[20] |
| Muscle |
|
||||||
| 13 | White potato Tainung No.10 | Tubers and leaves |
|
Muscle |
|
[21] | |
| 14 | Purple IBL (Cultivar Eshu No.12) were obtained from the Institute of Food Crops, Hubei Academy of Agricultural Sciences | Tubers |
|
|
Pancreas |
|
[22] |
| Liver |
|
||||||
| 15 | Purple IBL powder (cultivar Eshu No. 8) | Tubers | Diacylated anthocyanins |
|
Liver |
Enhancing secretion and sensitivity insulin eludidate mechanism:
|
[23] |
| 16 |
I. batatas (Linn.) Lam were purchased from Western Research Farm, National Root Crop Research Institute, Umudike, Abia state |
Tubers |
Flavonoid Terpenoid Tannin Phenol |
Pancreas |
|
[24] | |
| Gastrointestinal |
|
||||||
| Adipose |
|
||||||
| 17 | Purple IBL cv. Ayamurasaki were obtained from the Kyushu National Agricultural Experiment Station in Miyazaki prefecture (Japan) |
Tubers | Peonidin 3-O-[2-O-(6-O-E- feruloyl-β-D-glucopyranosyl)-6-O-E-caffeoyl-β-D-glucopyranoside]-5-O-β-D-glucopyranoside | Gastrointestinal |
|
[25] | |
| 18 | Korean red skin IBL (Ib 1) and Korean pumpkin IBL (Ib 2) purchased from the market in Goyang, Republic of Korea |
Peel-off tuber |
|
Gastrointestinal |
|
[26] | |
| 19 | White IBL (Caiapo) | Tubers |
|
Gastrointestinal |
|
[27] | |
| Adipose |
|
[28] | |||||
| 20 | White-skinned sweet potato (WSSP) was purchased from the Kagawa, Japan, Prefectural Cooperative |
Tubers |
|
Adipose |
|
[29] | |
|
|
Muscle |
|
[30] | |||
|
Liver |
|
|||||
| 21 | Korean purple IBL (Shinzami, Saeungbone9, Saeungyae33, Gyebone108, Gyeyae2469, and Gyeyae2258) |
Tubers |
|
|
Liver |
|
[31] |
| 22 | Color-fleshed potatoes (Sinjami and Sinhwangmi) |
Tubers |
|
|
Adipose |
|
[32] |
| 23 | Ipomoea batatas was grown in Kagawa Prefecture, Japan | Tubers |
|
Pancreas |
|
[33] | |
| 24 | White-skinned sweet potato (WSSP) | Tubers |
|
Liver |
|
[34] | |
| 25 | Purple IBL Antin-3 cultivar from the BALITKABI Malang |
Tubers |
|
Pancreas |
|
[35] | |
| 26 | White-skinned sweet potatoes (WSSP) from the local market Faisalabad (Pakistan) | Tubers | Carotenoid | Pancreas |
|
[36] | |
|
Liver |
|
[37] | ||||
| 27 | Purple IBL from Padang, West Sumatra, Indonesia | Tubers |
|
Gastrointestinal |
|
[38] |
4. Discussion
4.1. Type or cultivar
IBL have several varieties. These varieties are differentiated based on tuber color, skin color, leaf color, texture, and size. The number of cultivars identified in this journal is 27, with several variations, such as orange [13,20], purple [10,12,19,25,31,35,39], and white IBL [8,9,21,28,29,30,40,41]. Differences in cultivars will affect the phytochemical contents and their anti-diabetic activity.
4.2. Parts of plant and phytochemical identified of IBL
Many reports mentioned that parts of IBL used as anti-diabetic include leaves, tubers and tuber skin. Each part has different chemical compositions. In purple IBL tubers, the anthocyanin content commonly used as a marker is higher than in its leaves. The concentration of anthocyanins is also greater in purple IBL compared to white or orange IBL. Phenolic acid content such as chlorogenic acid, caffeic acid, 3,4,5-triCQA, 4,5-diCQA, 3,5-diCQA, caffeoyl acid derivative [8,9,13,18], flavonoid group such as cyanidin-3- glucoside, cyanidin-3,5-glucoside, cyanidin-3-rutinoside, peonidin-3-glucoside, cyanidin 3-caffeoyl-p-hydroxybenzolsophoroside-5-glucoside, peonidin 3-caffeoyl-p hydroxybenzoyl sophoroside-5-glucoside, Peonidin 3- O-[2-O-(6-O-E- feruloyl-β-D-glucopyranosyl)-6-O-E-caffeoyl-β-D-glucopyranoside]-5-O-β-D-glucopyranoside [19,25], quercetin, epicatechin, protocatechualdehyde, rutin, kaemferol, isoquercitrin and jaceosidin have been identified in such type of potato. They directly serve as effective anti-diabetic agents [8,9,20,33]. Flavonol, a subclass of flavonoids, is extensively found in various natural sources. The flavonols, such as quercetin and epicacthecin, have demonstrated their potential to enhance GLP-1 secretion in tissue culture of GLUTag cells [42]. Groud triterpenoid such as trans-N-(p-coumaroyl) tyramine, trans-N-feruloyltyramine, cis-N-feruloyltyramine, and 7-hydroxy-5-methoxycoumarin and alkaloid group such as Indole-3-carboxaldehyde also have potential as antidiabetic agents [13].
4.3.1. Site of Action
GLP-1 is produced in the intestine through posttranslational processing of proglucagon. The L-cells, primarily found in the ileum and colon, are a type of open-type epithelial cells that directly interact with nutrients in the intestinal lumen. GLP-1 in the circulation is rapidly increased by nutrients such as carbohydrates, fats, proteins, and dietary fibre. Glucose is taken up through GLUT2, fructose through GLUT5, and short-chain fatty acids (SCFAs) are absorbed and metabolized intracellularly. The secretion of GLP-1 in intestinal L-cells is induced by the closure of KATP-channels, which is a result of carbohydrate uptake through SGLT1, GLUT2, and GLUT5. This leads to cell membrane depolarization through intracellular metabolism, resulting in the production of ATP and the closure of ATP-sensitive K channels (KATP), as well as the opening of voltage-gated Ca2+-channels. Additionally, the uptake of free amino acids through coupled transport with Na+ and the uptake of peptides through PepT1 also contribute to depolarization and the opening of voltage-gated Ca2+-channels. These pathways stimulate GLP-1 secretion by activating voltage-gated Ca2+-channels (V-type). Furthermore, the further depolarization caused by the uptake of extracellular Ca and the Ca-induced mobilization of Ca from intracellular stores collectively activate the exocytotic machinery, leading to the secretion of GLP-1 [43,44]. The intracellular overexpression of GLP-1 leads to its distribution throughout the systemic system, binding to the GLP-1R receptor in various organ tissues including the GI tract, pancreas, liver, skeletal muscle, and adipose tissue. This binding of GLP-1 reduces glucose production in the liver and enhances glucose uptake in adipose tissue and muscle [45,46].
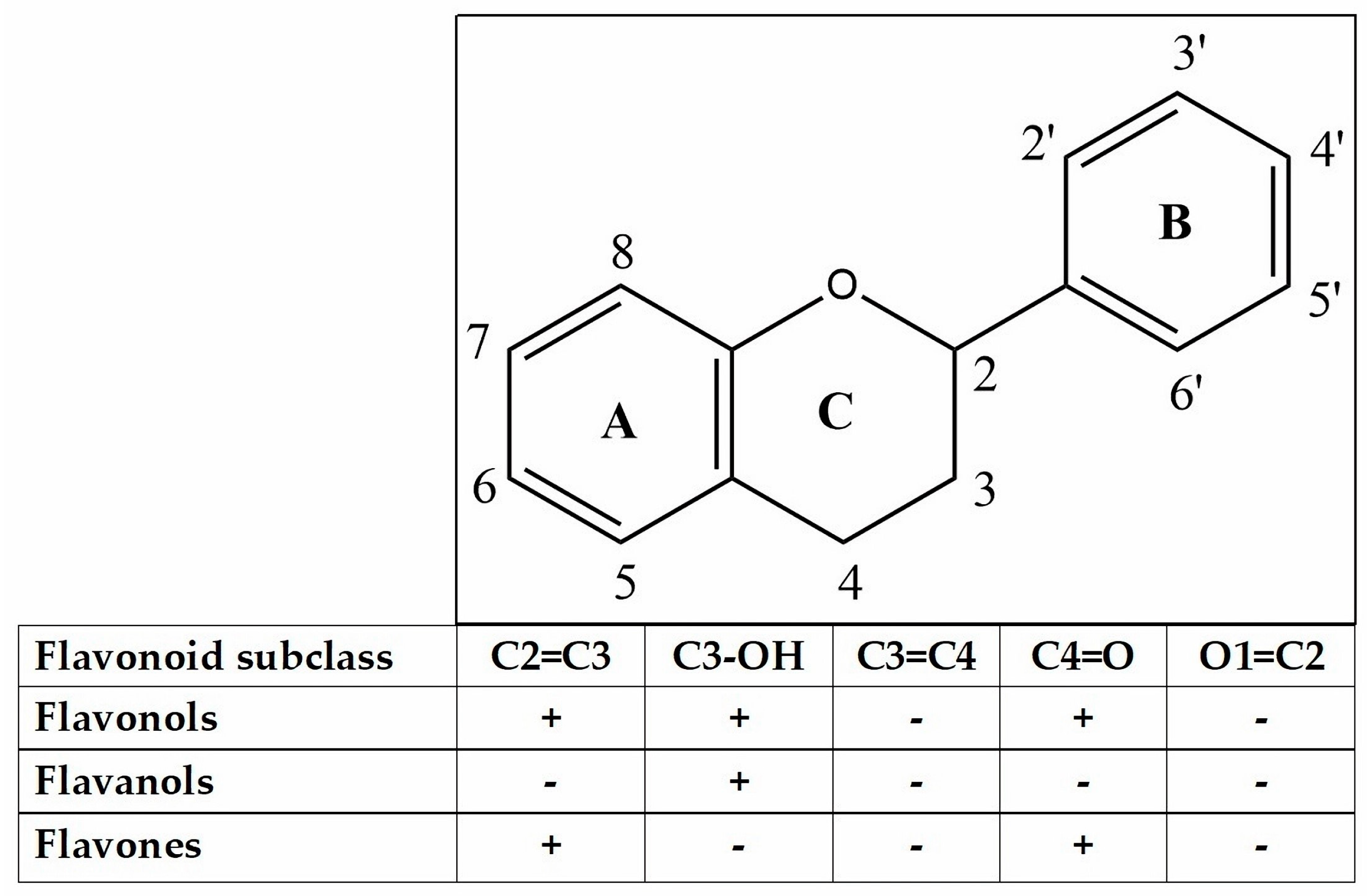
Various flavonoid compounds, including hispidulin, epicatechin, quercetin, cyanidin-3-glycosides, 5,7-dihydroxy-6-4-dimethoxyfavanone, and homoesperetin-7-rutinoside, have demonstrated their ability to enhance GLP-1 secretion both in vitro and in vivo. Epicatechin, cyanidin-3-glycosides and hispidulin have been reported to stimulate GLP-1 in GLUTag cells. Homoesperetin-7-rutinoside has also exhibited stimulation through molecular docking [42,47,48]. The chelation of Ca2+ by quercetin, similar to the chelation of AlCl3, has been documented in various studies. Numerous reports in the literature have detailed the formation of complexes between quercetin and metals, with complexation sites located at the ortho positions O3/O4, O4/O5, and O3'/O4' (Figure 3) [49]. Compounds belonging to the flavonol, flavanol, and flavones group exhibit activity against AlCl3, resulting in a yellow color change. It is anticipated that these compounds will operate via a similar mechanism against Ca2+. These compounds have the ability to prolong the half-life of GLP-1, leading to an increase in insulin secretion and a shift in glucose uptake from systemic to cellular in the form of glycogen [50]. Furthermore, the A or B rings of flavonoids are capable of interacting with AlCl3 through their ortho-dihydroxyl groups, resulting in the formation of complexes that are susceptible to acid [51].
The development and progression of diabetes and its complications are largely attributed to increased oxidative stress. Pancreatic islets are particularly susceptible to oxidative damage due to their low expression levels of antioxidant enzymes. Biomarkers such as MDA are used to assess oxidative stress, with increased levels indicating higher levels of lipid peroxidation. GLP-1 has been found to decrease oxidative stress in diabetes through receptor-mediated activation of cAMP, PI3K, and PKC pathways, as well as activation of Nrf-2, which in turn increases antioxidant capacity. Conversely, oxidative stress can be reduced by suppressing ROS through radical scavenging and chelating mechanisms, thus protecting pancreatic beta cells [52].
Figure 3.
Flavonoid skeleton.
4.3.2. Gastrointestinal Tract
- Regulation of carbohydrate metabolism
Inhibition of α-glucosidase influences the capability of the small intestine to inhibit absorption of carbohydrates. This particular enzyme inhibits the conversion of complex carbohydrates and is unable to be assimilated into simple carbohydrates [53]. IC50 values of all compound have been investigated to be in the range of 4.46 µM to 64.14 µM which were observed in various studies on the efficacy of α-glucosidase inhibitors, in comparison to acarbose. Ethyl caffeate has the ability to act as a stronger α-glucosidase inhibitor (app. 6.77 times higher than acarbose). Flavonoid compounds such as kaempferol, quercetin, hyperoside, isoquercitrin and routine have a strong α-glucosidase inhibitory activity [9]. Trans-N-(p-coumaroyl)tyramine and 3,4,5-triCQA respectively have IC50 values of 4.46 µM and 4.61 µM, considered as the highest activity, followed by trans-N-feruloyltyramine and cis-N-feruloyltyramine with respective IC50 values of 9.04 µM and 14.35 µM. The activity of these compounds surpasses that of acarbose and their IC50 value were 37.9, 36.6, 18.7, and 11.8 fold stronger than acarbose (IC50 168.95 µM), respectively. Quercetin-3-O-glucosidase and 7-Hydroxy-5-methoxycoumarin with respective IC50 values of 22.38 ±1.73 µM and 64.14 ±9.23 µM also have good activity [13]. Chlorogenic acid is also predicted to have inhibition activities on α-glucosidase and tyrosinase. The docking results also support that Chlorogenic acid binds to the active site of α-glucosidase and can act in a reversible competitive manner through hydrogen bonds [16]. Peonidin 3-O-[2-O-(6-O-E-feruloyl-β-D-glucopyranosyl)-6-O-E-caffeoyl-β-D-glucopyranoside]-5-O-β-D glucopyranoside which showed a potent maltase inhibitory activity with an IC50 value of 200 µM is preferable to sucrase inhibition [25]. The aqueous fraction contain an acidic glycoprotein (IC50 53 µg/mL) that exhibits antidiabetic properties and is predicted to have a mechanism of α-glicosidase inhibition is a significant breakthrough in the treatment of diabetes mellitus. This finding is particularly noteworthy when compared to acarbose [54].
α-amylase in ethyl caffeic has 13.1 stronger activity then acarbose. This is possibly due to contribution of the OH group of the ethyl caffeic binding to the enzyme. Chlorogenic acid inhibits α-amylase in a mixed-type manner, by binding through hydrogen bonds, to amino acid residues near the active site and changing the secondary structure of the enzyme's protein [9]. Cyanidin-3-glucoside, cyanidin-3-rutinoside, cyanidin-3,5-glucoside, and peonidin-3-glucoside with IC50 values of 0.024 ± 0.003, 0.040 ± 0.007, 0.031 ± 0.007, 0.075 ± 0.007, respectively against porcine pancreatic α-amylase inhibition. These four anthocyanin compounds were proven in silico, the active side of the compound was thought to be mediated by binding to the side chain of GLU233. Cyanidin-3-glucoside showed the highest inhibitory activity with a Ki value of 0.0014 mM, followed by cyanidin-3-rutinoside, cyanidin-3,5-glucoside, and peonidin-3-glucoside with Ki values of 0.019, 0.020, 0.045 nM respectively. GLU233 was found to be the common key side chain for imparting the inhibition activity [19]. Each compound induces absorption that exclusively takes place as fiber and is subsequently eliminated through the gastrointestinal tract.
- b.
- Increase insulin secretion
GLP-1 is an incretin hormone released in intestinal endocrine L cells that respond to incoming foods. Enhancement of GLP-secretion in the gastrointestinal tract leads to an increase in insulin secretion, a decrease in GI motility, and a delay in gastric emptying. Various studies have identified compounds belonging to the phenolic acid and flavonoid groups that exhibit the potential to augment GLP-1 secretion [5,6].
3,4,5-triCQA is the most potent CQA derivative, which increases GLP-1 secretion around 10-fold compared to the sulfonylure control tested on GLUTag cells. The same effect was also produced the in vivo testing, that the treatment group produced more GLP-1 secretion compared to the sulfonylurea control group. Therefore, GLP-1 can regulate glycaemic homeostasis without the risk of hypoglycaemia. The elevation of cAMP concentration may lead to the stimulation of GLP1 secretion in both in the vivo model and a cell line. The observed increases in GLP-1 secretion in L cells were caused by the activation of PKA and cAMP in vitro model using GLUTag cells [18]. Cyanidin 3-O-glucoside and epicatechin function through the activation of cAMP/PKA and ERK ½ pathways. Ca2+ chelation's mechanism of action leads to an elevation in intracellular Ca2+ concentration, consequently resulting in an augmentation of GLP-1 secretion. The presence of a 3'4' catechol group in the B ring appears to be a key chemical structural element that supports this activity [42], Hispidulin has also been identified as an effective anti-diabetic agent. Notably, the study demonstrates that hispidulin treatment leads to an increase in intracellular cAMP levels in L-cells [48].
Another clinically useful strategy is to elongate GLP-1 by inhibiting DPP-4. Flavonoid as flavonol, flavanol, and flavone (Figure 3) among which myricetin, hyperoside, narcissoside, cyanidin 3-O-glucoside, and isoliquiritigenin showed higher inhibitory activities in a concentration-dependent manner. An analysis of the structure-activity relationship indicated that the introduction of hydroxyl groups to C3', C4', and C6 of the flavonoid structure was advantageous in enhancing the inhibitory efficacy against DPP-4. However, hydroxylation at position 3 of the ring C in the flavonoid structure was found to be unfavorable for inhibition. Additionally, the methylation of the hydroxyl groups at C3', C4', and C7 of the flavonoid structure tended to decrease the inhibitory activity against DPP-4 enzyme. Furthermore, the presence of a 2,3-double bond and a 4-carbonyl group on ring C of the flavonoid structure was deemed essential for the inhibitory effect [55].
4.3.3. Pancreas
- Inhibit apoptosis beta cell and recovering the islet structure through protective cell beta
Pancreatic beta cells play an important role in maintaining glucose homeostasis and as the main source of insulin production. These cells are responsible for the synthesis, storage and release of insulin [56]. The research findings indicated that the IC50 value for total compound fell within the range of 9.69 ± 0.03 µM to IC50 125 ± 0 µM for ascorbic acid. This particular range demonstrates a significant efficacy in conferring natural antioxidant properties to the compound. An elevation in ROS is the sole factor responsible for harm to beta cells. Antioxidants in abundance are believed to have the potential to diminish ROS, leading to the recovery of islet beta cells. This recovery process will enhance the insulin secretion process through protective cell beta.
Giving SPLP (sweet potato leave phenol) leaf extract from Beijing, to T2DM mice for 4 weeks showed recovery of pancreatic tissue with increased area and complete islet structure, increased mass and clear borders. Giving SPLP treatment is indicated to inhibit beta cell apoptosis. It is suspected that phenolic compounds such as 1-caffeoylquinic acid, caffeac acid, 3,4,5-tricaffeoylquinic acid (3,4,5-triCQA) and chlorogenic acid modulate beta cell regeneration. The radical scavenging activity of 3,4,5-triCQA has an IC50 value of 10.8 times stronger than the ascorbic acid control. Then other compounds that are stronger than acorbic acid are 4,5-diCQA > 3,5-diCQA > 3,4-diCQA > Caffeic acid ethyl ester > Caffeic acid. Other studies reported antioxidant activity with ABTS, that Ethyl caffeate and 3,4,5-CQA were reported to have good activity with IC50 values of similar to ascorbic acid. Other anti-oxidant activity values from the highest to the lowest activity are 4,5-CQA >3-CQA=5-CQA=CA=4-CQA>3,5-CQA>1-CQA>3,4-CQA>esculin>7-hydroxycoumarin.
Protocatechualdehyde showed the best antioxidant activity with antioxidant activity of 2.32 higher than that of ascorbic acid. For the flavonoid group, quercetin showed the best antioxidant activity, namely 4.14 greater than ascorbic acid. Then kaemferol and jaceosidin showed good ABTS radical scavenging activity and were almost equivalent to ascorbic acid. The radical scavenging test with DPPH showed that protocatechualdehyde showed the best DPPH activity which was 2.69 higher than ascorbic acid. Then proceed with Ethyl caffeate and 3,4,5-CQA which have good DPPH activity. Another order of DPPH antioxidant activity from high to low is CA>4,5-CQA>3-CQA>1-CQA=5-CQA=3,5-CQA>4-CQA=3,4-CQA>7-hydroxycoumarin>esculin. Quercetin has DPPH radical scavenging activity of 1.8 higher compared to ascorbic acid. Isoquercitrin, hyperoside, kaempferol and routine showed good activity. The high DPPH radical scavenging capacity of CA is probably related to the 1,2-phenolic diol group and to conjugation with the C=C and C=O bonds of Caffeic acid. Another mechanism using the FRAP method is that protocatechualdehyde has 2.22 stronger activity than ascorbic acid. Ethyl caffeate, caffeac acid, 3,4,5-CQA and 4,5-CQA also have good activity. For flavonoid compounds, namely quercetin produces a stronger reducing power activity of 1.54 compared to ascorbic acid. Hyperoside, isoquercitrin, kaempferol and routine showed good FRAP capacity [8,13,57]. These IC50 values of SPLP were consistent with a previous report, in that they had stronger anti-oxidant activity than polyphenols from tea and grape seed [58].
The significant increase in the enzymatic antioxidants SOD and GSH-Px will correlate with antioxidant activity to reduce ROS. A decrease in ROS will cause the recovery of pancreatic beta cells. The compound suspected in this process is protein-bound anthocyanin from tuber. Anthocyanins that have been identified from purple IBL and possibly bound to protein are cyanidin-3-sophoroside-5-glucoside and peonidin-3-sophoroside-5-glucoside [12,22]. Other studies report that the antioxidant activity of Cyanidin 3-caffeoyl-p-hydroxybenzoyl-sophoriside-5-glucoside is the highest compared to other anthocyanin compounds when tested using ascorbic acid using the DPPH and ABTS methods. Furthermore, other antioxidant activities followed from peonidin 3-caffeoyl sophoroside-5-glucoside>peonidin3-(6''-caffeoyl-6''-feruoyl sophoroside)-5-glucoside>peonidin3-caffeoyl-p-hydroxybenzoyl-sophoroside- 5-glucoside [31].
Tuber extract and orange-flesh IBL leaves as beta cell protectors can reduce lipid peroxidation through a radical scavenging mechanism and the results of measuring antioxidant activity with FRAP and TEAC can significantly reduce ROS. The antioxidant capacity values using FRAP were respectively 299.8 ± 2.5 and 296.9 ± 7.4 (µM AAE/mg protein) and testing using the TEAC method were 127.9 ± 2.10 and 126.3 ± 2.51 (µM TE/mg Protein). The results of this test are greater than standard ascorbic acid, namely 271.0 ± 4.17 (µM AAE/mg protein and 107.2 ± 1.68 (µM TE/mg protein). Compounds predicted to be contained in the extract that have radical scanning activity are caffeic acid, hyperoside, protocatechuic acid , quercetin, routine, vanillic acid [20]. Caffeic acid is reported to be a compound with potent antioxidant activity [59,60]. MAE (Microwave Assisted Extraction) leaves of purple IBL cultivar antin-3 which were predicted to contain the anthocyanin group produced an IC50 value of 61.91 ± 1.11 ppm [61].
This compound works directly in increasing insulin secretion through repair of islet beta cells. This compound leads to scavenges ROS and increases the AMP/ATP ratio in beta cells. Change in the AMP/ATP ratio activates mitochondrial target, induces mitogenesis and stimulates insulin secretion [62].
- b.
- Suppression anti-inflammatory pathway
Compounds identified from Bandungan, Java, Indonesia, were anthocyanin, catechin, quercetin, proanthocyanin, and caffeic acid. These compounds are predicted to work in the pancreas by inhibiting anti-inflammatory mechanisms [17]. Administration of leaf extract at a dosage of 2.5 g/kgBW for a duration of 14 days has been shown to result in a 50% increase in pancreatic islet cells when compared to the control group. In contrast, administration of caiapo at a dosage of 5g/KgBW for a period of 8 weeks has been found to significantly increase beta cell mass by two-fold when compared to untreated diabetic control subjects (p<0.05). These findings suggest that higher dosages of the extract may lead to a greater recovery of islet beta cells. It is worth noting that the extract is believed to contain quercetin, chlorogenic acid, caffeic acids, and their derivatives. The anti-inflammatory properties of these compounds are thought to be responsible for the reduction in inflammation by suppressing inflammatory mediators. Quercetin is known to have capability to inhibit tyrosine kinase activity, which is shown to be anti-diabetic. Regulation of quercetin effects through inhibition of NFκB activation of beta cells also helps to improve glucose-stimulated insulin secretion [12,63,64]. Apart from that, the content of chlorogenic acid, caffeic acids, and their derivatives is thought to be able to inhibit the JNK, P38 MAP, and NF-κB pathways and is also associated with various inflammatory mediators such as TNF-α, IL-6, and CRP. It has also been reported that inhibiting oxidative stress may also induce hypoglycemic effect [33].
4.3.4. Liver
- Improving insulin secretion and insulin sensitivity by reducing glucose synthesis
The compounds cyanidin-3-sophoroside-5-glucoside and peonidin-3-sophoroside-5-glucoside were the primary structures of other acylated anthocyanins that are predicted to have a crucial function in enhancing glucose absorption and raising insulin levels. According to studies, this compound can lower glycolysis through p-AMPK activity impairment. The treatment group given 200 mg/kg of Free Anthocyanin Compound of Sweet potato (FAC-PSP) extract containing 40.74 ± 2.88 mg cyanidin-3-glycoside/g for 4 weeks showed a substantial rise in p-AMPK expression levels. In the liver, the insulin-responsive glucose transporter GLUT2 is crucial for metabolism and glucose uptake. GLUT2 protein expression can enhance glucose uptake and utilization in the liver [39]. Cyanidin 3-caffeoyl-p-hydroxybenzolsophoroside-5-glucoside and Peonidin 3-(6’’-caffeoyl-6’’’-feruloyl sophoroside)-5-glucoside suppressing hepatic gluconeogenesis in HepG2 cell but the results of an in vivo research of cyanidin effects showed that oral treatments dramatically lowered fasting blood glucose from its initial high values at time 0 (186-205 mg/dL, respectively) [31].
According to earlier studies, it was found that blackcurrant extract, which consists of 45% anthocyanins and 82% total polyphenols, has potential to enhance plasma GLP-1 levels by approximately 30% and stimulate AMPK in the liver. The increased GLP-1 levels directly contribute to the reduction in hepatic glucose production and the suppression of hepatic expression of phosphoenol pyruvate carboxykinase (PEPCK) and glucose-6-phosphatase (G6Pase) [65,66]. The increase in blood glucose can be effectively prevented by inhibiting the expression of PEPCK and G6Pase, which suggests that gluconeogenesis will be attenuated and that the amount of glucose produced will subsequently decrease [67]. GLP-1 in liver reduces hepatic gluconeogenesis and increases glycogen formation, but there are some debates over whether these effects are mediated by GLP-1R in hepatocytes, or whether the effects may be indirectly mediated through CNS or insulin release. GLP-1 promotes glycogen synthesis and decreased gluconeogenesis in vitro through upregulation of glycogen synthase that occurs downstream of PI3K/PKB, PKC, and serine/threonine protein phosphatase 1, and also by reduced expression of gluconeogenetic enzyme phosphoenol pyruvate carboxykinase in rat hepatocytes [65].
The P13K/AKT pathway is one of the major insulin signaling pathways focused in the current research [68]. In addition to increasing insulin sensitivity and glucose metabolism, upregulating the PI3K/AKT/GSK-3ß signaling pathway also lowers dyslipidemia. The phenolic and flavonoid compounds in SPLP (sweet potato leaf polyphenols) in high dose 150mg/kgBB could reduce FBG (fasting blood glucose) for 4 weeks more effectively compred with low dose treatment. These result indicated that SPLP reduce the FBG in dose-dependent and time-dependet manner. The component of SPLP include predictive compound such as 1-caffeoylquinic acid, 3,4,5-tricaffeoylquinic acid (3,4,5-triCQA), chlorogenic acid, caffeac acid derivate, quercetin, isoquercitrin, hyperoside and rutin are responsible to regulate hepatic glycogen synthesis in the liver, which is insulin-mediated by upregulating PI3K/AKT and down-regulating GSK-3ß FOXO1 expression[8].
4.3.5. Muscle
- Enhancing the absorption of glucose, secretion, and insulin sensitivity
Insulin-stimulated glucose uptake in skeletal muscle plays a key role in the regulation of glucose metabolism and energy homeostasis. The target of the PI3K/Akt pathway is essential for the treatment of T2DM functions related to this signaling change pathways in the muscle. In the present study, it was predicted that arabinogalactan and epigallocatechin in WSPP would increase the expression of p-IR, p-Akt, and M-GLUT4. High doses of DM+30% WSP-Tuber and DM+5% WSP-Leave can significantly reduce fasting blood count glucose levels, improving fasting glucose tolerance [21]. Epigallocationchin has recently been demonstrated to enhance the secretion of GLP-1. By overexpressing GLP-1, it will be able to enter the systemic circulation and bind to the GLP-1R receptor in different tissues, including skeletal muscle. Consequently, this binding event leads to an elevation in the level of cAMP signaling through the Gs protein, thereby promoting AMPK phosphorylation. This, in turn, triggers glycogen synthesis and facilitates glucose uptake in skeletal muscle by facilitating the translocation of GLUT-4 from the intracellular depot to the sarcolemma [46,69].
Another fraction in the ≤10 kDA from WSSp also has a significant increase in AKT phosphorylation by insulin (Kinoshita et al., 2023). The expression of GLUT-4, NRF1 gene, and MERF2a was observed to increase after subjecting C2C12 skeletal muscle cells to tissue culture testing. This testing involved treating the cells with doses of 500 µg/mL of OSPT (orange sweet potato tubers) and 100 µg/mL of OSPL (orange sweet potato leaves) for a duration of 3 hours. Both the transcription factors MEF2a and NRF1 are crucial in controlling the expression of GLUT-4 and, ultimately, the metabolism of glucose absorption. It has been discovered that the expression and activity of the GLUT-4 gene are closely related to those of the nrf1 and mef2a genes, as well as to insulin sensitivity and the preservation of glucose homeostasis in skeletal muscle. Acc2 and cpt1 are essential regulators of mitochondrial fatty acid oxidation, and therefore, strategies that influence their expression will influence intracellular lipid levels and have therapeutic implications in controlling insulin resistance. Therefore, the increased expression of these genes in the treated cells suggests that a compound derived from aqueous-methanol extracts of orange-fleshed IBL that contains caffeaic acid, catechin, hysperoside, kaemferol, rutin, quercetin, protocatechui acid, isovanillic acid, and vanillic acid has the potential to increase insulin sensitivity [20].
4.3.6. Adiposa
- Increase glucose uptake and insulin secretion
Pure compound of quercetin 3-O-β-D sophoroside, quercetin, benzyl β-d-glucoside, 4-hydroxy-3-methoxybenzaldehyde, methyl decanoate has been proven to increase the expression of PI3K, AKT, and GLUT-4 phosphorylation tested on 3T3-L1 adipocytes in vitro tested at dose 0.01 mg/mL using western blot analysis. Activation of this gene will increase GLUT-4 translocation so that glucose uptake also increases [10]. Caiapo containing aglycoprotein 4g/day orally has been clinically tested on 30 patients and is effective in reducing HbA1c progressively in diabetes patients for 1-2 months when compared to the placebo group[28]. Other studies also reported that caffeic acid could increase insulin secretion and sensitivity by increasing adiponectin expression [29]. Adiponectin, an insulin-sensitizing hormone with antiapoptotic and anti-inflammatory effects, is made almost exclusively in adipose tissue. Adiponectin levels are significantly decreased in obesity and type 2 diabetes. Adiponectin treatment enhances insulin-stimulated glucose uptake via activation of AMPK in primary rat adipocytes. Adiponectin directly targets insulin receptor substrate-1 (IRS-1) and plays a crucial role in insulin mediation of glucose uptake in adipocytes [70,71].
5. Conclusions
IBL were reported to have variety and cultivar that contain chemicals with antidiabetic properties. Such chemical compounds include flavonols, flavanol, flavones, antochyanin, phenolic acid, and triterpenoid groups. IBL can be considered as a multi-chemical and multi-pharmacological sites since they function in multiple organs through various ways. GLP-1 therapy for DM will prove to be quite advantageous in the future due to its efficacious nature. Flavonols, flavones, and flavone groups exhibit a robust interaction with DPP-IV, resulting in the inhibition of GLP-1 degradation. This interaction effectively extends the halflife of GLP-1 within the systemic system. The group compunds plays a crucial role in increasing GLP-1 activity and exerting its anti-diabetic effects, which would subsequently strengthen insulin production and the uptake of glucose into cells as glycogen from the systemic circulation.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, N.K.S.M.D. and I.M.A.G.W.; methodology, N.K.S.M.D.; software, N.K.S.M.D.; validation, N.K.S.M.D., I.M.A.G.W. and D.M.W.; formal analysis, DMW.; investigation, M.R.S.; resources, N.K.S.M.D.; data curation, I.M.A.G.W.; writing—original draft preparation, N.K.S.M.D.; writing—review and editing, Y.R.; visualization, M.R.S; supervision, DMW.; project administration, N.K.S.M.D.. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data to support the finding of this study can be made available by the corresponding author upon request.
Acknowledgments
The authors express their gratitude to the Rector of Universitas Udayana, Indonesia for providing access to their facilities during the course of this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- G. Danaei et al. National, regional, and global trends in fasting plasma glucose and diabetes prevalence since 1980: Systematic analysis of health examination surveys and epidemiological studies with 370 country-years and 2·7 million participants. Lancet 2011, 378, 31–40. [Google Scholar] [CrossRef] [PubMed]
- K. Ogurtsova et al. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed]
- R. K. Al-Ishaq, M. Abotaleb, P. Kubatka, K. Kajo, and D. Büsselberg. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules, 2019, 9, 430. [Google Scholar] [CrossRef]
- C. I. S. Arisanti, I. M. A. G. Wirasuta, I. Musfiroh, E. H. K. Ikram, and M. Muchtaridi. Mechanism of Anti-Diabetic Activity from Sweet Potato (Ipomoea batatas): A Systematic Review. Foods, 2023, 12, 2810. [Google Scholar] [CrossRef] [PubMed]
- T. D. Müller et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab., 2019, 30, 72–130. [Google Scholar] [CrossRef] [PubMed]
- A. Andersen, A. S. Christensen, F. K. Knop, and T. Vilsbøll. Glucagon-like peptide 1 receptor agonists for the treatment of Type 2 diabetes. Ugeskr. Laeger 2022, 181, 202–210. [Google Scholar] [CrossRef]
- B. Hutton. et al. The PRISMA extension statement for reporting of systematic reviews incorporating network meta-analyses of health care interventions: Checklist and explanations. Ann. Intern. Med. 2015, 162, 777–784. [CrossRef]
- D.Luo, T. Mu, and H. Sun. Sweet potato (: Ipomoea batatas L.) leaf polyphenols ameliorate hyperglycemia in type 2 diabetes mellitus mice. Food Funct. 2021, 12, 4117–4131. [Google Scholar] [CrossRef]
- D. Luo, T. Mu, and H. Sun. Profiling of phenolic acids and flavonoids in sweet potato (Ipomoea batatas L.) leaves and evaluation of their anti-oxidant and hypoglycemic activities. Food Biosci., 2021, 39, 100801. [Google Scholar] [CrossRef]
- C. L. Lee et al. Characterization of secondary metabolites from purple Ipomoea batatas leaves and their effects on glucose uptake. Molecules, 2016, 21, 745. [Google Scholar] [CrossRef]
- G. Pal, S., A. S., Mishra, R. Maurya, and A. K. Srivastava. Antihyperglycemic and antidyslipidemic potential of ipomoea batatas leaves in validated diabetic animal models. Int. J. Pharm. Pharm. Sci. 2015, 7, 176–186, [Online]. Available: https://api.elsevier.com/content/abstract/scopus_id/84936852883. [Google Scholar]
- P. S. Yustiantara et al. Determination of TLC fingerprint biomarker of Ipomoea batatas (L.) Lam leaves extracted with ethanol and its potential as antihyperglycemic agent. Pharmacia 2021, 68, 907–917. [Google Scholar] [CrossRef]
- L. Zhang, Z.-C. Tu, T. Yuan, H. Wang, X. Xie, and Z.-F. Fu. Antioxidants and α-glucosidase inhibitors from Ipomoea batatas leaves identified by bioassay-guided approach and structure-activity relationships. Food chemistry 2016, 208, 61–67. [Google Scholar] [CrossRef] [PubMed]
- R. Zhao, Q. Li, L. Long, J. Li, R. Yang, and D. Gao, Antidiabetic activity of flavone from Ipomoea Batatas leaf in non-insulin dependent diabetic rats. 2007. [CrossRef]
- M. Zovko, R. Petlevski, Z. Kaloðera, and K. Plantak, Antioxidant and antidiabetic activity of leaves of Ipomoea batatas grown in continental Croatia. 2008. [CrossRef]
- G. Zengin et al. Chemical characterization, antioxidant properties, anti-inflammatory activity, and enzyme inhibition of Ipomoea batatas L. leaf extracts. Int. J. Food Prop. 2017, 20, 1907–1919. [Google Scholar] [CrossRef]
- D. Novrial, S. Soebowo, and P. Widjojo. Protective Effect of Ipomoea batatas L Leaves Extract on Histology of Pancreatic Langerhans Islet and Beta Cell Insulin Expression of Rats Induced by Streptozotocin. Molekul, 2020, 15, 48. [Google Scholar] [CrossRef]
- R. Nagamine et al. Dietary sweet potato (Ipomoea batatas L.) leaf extract attenuates hyperglycaemia by enhancing the secretion of glucagon-like peptide-1 (GLP-1). Food Funct. 2014, 5, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- X. Sui, Y. Zhang, and W. Zhou. In vitro and in silico studies of the inhibition activity of anthocyanins against porcine pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- T. Ayeleso, K. Ramachela, and E. Mukwevho. Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells. Molecules, 2018, 23, 2058. [Google Scholar] [CrossRef] [PubMed]
- C. K. Shih, C. M. Chen, V. Varga, L. C. Shih, P. R. Chen, and S. F. Lo. White sweet potato ameliorates hyperglycemia and regenerates pancreatic islets in diabetic mice. Food Nutr. Res., 2020, 64. [Google Scholar] [CrossRef]
- T. Jiang, X. Shuai, J. Li, N. Yang, S. Deng, L., Li, and J. He. Protein-Bound Anthocyanin Compounds of Purple Sweet Potato Ameliorate Hyperglycemia by Regulating Hepatic Glucose Metabolism in High-Fat Diet/Streptozotocin-Induced Diabetic Mice. J. Agric. Food Chem. 2020, 68, 1596–1608. [Google Scholar] [CrossRef]
- L. Shen, Y. Yang, J. Zhang, L. Feng, and Q. Zhou. Diacylated anthocyanins from purple sweet potato (Ipomoea batatas L.) attenuate hyperglycemia and hyperuricemia in mice induced by a high-fructose/high-fat diet. J. Zhejiang Univ. Sci. B 2023, 24, 587–601. [Google Scholar] [CrossRef]
- C. S. Okafor, C. Ezekwesili, N. Mbachu, K. C. Onyewuchi, and U. C. Ogbodo, Anti-diabetic Effects of the Aqueous and Ethanol Extracts of Ipomoea batatas Tubers on Alloxan Induced Diabetes in Wistar Albino Rats. 2022. [CrossRef]
- T. Matsui et al., Anti-hyperglycemic effect of diacylated anthocyanin derived from Ipomoea batatas cultivar Ayamurasaki can be achieved through the alpha-glucosidase inhibitory action., 2002, 50. [CrossRef]
- G. Das, J. K. Patra, N. Basavegowda, C. N. Vishnuprasad, and H.-S. H.-S. S. Shin, Comparative study on antidiabetic, cytotoxicity, antioxidant and antibacterial properties of biosynthesized silver nanoparticles using outer peels of two varieties of Ipomoea batatas (L.) Lam 2019, 14. [Google Scholar] [CrossRef]
- Y. Sakuramata, H. Oe, S. Kusano, and O. Aki, Effects of combination of Caiapo® with other plant-derived substance on anti-diabetic efficacy in KK-Ay mice. 2004. [CrossRef]
- B. Ludvik, B. Neuffer, and G. Pacini. Efficacy of Ipomoea batatas ( Caiapo ) on Diabetes Control in Type 2 Diabetic. Diabetes Care 2004, 27, 436–440. [Google Scholar] [CrossRef]
- S. Kusano, S. Tamasu, and S. Nakatsugawa. Effects of the White-Skinned Sweet Potato (Ipomoea batatas L.) on the Expression of Adipocytokine in Adipose Tissue of Genetic Type 2 Diabetic Mice. Food Sci. Technol. Res. 2005, 11, 369–372. [Google Scholar] [CrossRef]
- A. Kinoshita, T. Nagata, F. Furuya, M. Nishizawa, and E. Mukai. White-skinned sweet potato (Ipomoea batatas L.) acutely suppresses postprandial blood glucose elevation by improving insulin sensitivity in normal rats. Heliyon 2023, 9. [CrossRef]
- H. Jang, H. Kim, and S. Kim. In vitro and in vivo hypoglycemic effects of cyanidin 3-caffeoyl-p-hydroxybenzoylsophoroside-5-glucoside, an anthocyanin isolated from purple-fleshed sweet potato. Food Chem. 2019, 272, 688–693. [Google Scholar] [CrossRef]
- A. H.-J. Kim et al. Anti-obesity activity of anthocyanin and carotenoid extracts from color-fleshed sweet potatoes. J. Food Biochem., 2020, 44. [CrossRef]
- Niwa, T. Tajiri, and H. Higashino. Ipomoea batatas and Agarics blazei ameliorate diabetic disorders with therapeutic antioxidant potential in streptozotocin-induced diabetic rats. J. Clin. Biochem. Nutr., 2011. [CrossRef]
- N. Oki, S. Nonaka, and S. Ozaki. The effects of an arabinogalactan-protein from the white-skinned sweet potato (Ipomoea batatas L.) on blood glucose in spontaneous diabetic mice. Biosci. Biotechnol. Biochem. 2011, 75, 596–598. [Google Scholar] [CrossRef]
- L. A. Wicaksono, Yunianta, and T. D. Widyaningsih. Anthocyanin extraction from purple sweet potato cultivar antin-3 (Ipomoea batatas L.) using maceration, microwave assisted extraction, ultrasonic assisted extraction and their application as anti-hyperglycemic agents in alloxan-induced wistar rats. Int. J. PharmTech Res. 2016, 9, 181–192, [Online]. Available: https://api.elsevier.com/content/abstract/scopus_id/84963724810. [Google Scholar]
- S. Kamal et al. Anti-diabetic activity of aqueous extract of Ipomoea batatas L. in alloxan induced diabetic Wistar rats and its effects on biochemical parameters in diabetic rats. Pak. J. Pharm. Sci. 2018, 31, 1539–1548, [Online]. Available: https://api.elsevier.com/content/abstract/scopus_id/85064116246. [Google Scholar]
- N. Akhtar, M. Akram, M. Daniyal, and S. Ahmad. Evaluation of antidiabetic activity of Ipomoea batatas L. extract in alloxan-induced diabetic rats. International Journal of Immunopathology and Pharmacology 2018, 32. [CrossRef]
- S. Nurdjanah,. I., S. Astuti, and N. Yuliana. Inhibition Activity of α-amylase by Crude Acidic Water Extract from Fresh Purple Sweet Potato (Ipomoea batatas L.) and its Modified Flours. Asian Journal of Scientific Research, 2020, 13, 190–196. [Google Scholar] [CrossRef]
- T. Jiang et al. Protein-Bound Anthocyanin Compounds of Purple Sweet Potato Ameliorate Hyperglycemia by Regulating Hepatic Glucose Metabolism in High-Fat Diet/Streptozotocin-Induced Diabetic Mice. J. Agric. Food Chem. 2020, 68, 1596–1608. [Google Scholar] [CrossRef]
- N. Akhtar et al. Anti-diabetic activity of aqueous extract of Ipomoea batatas L. in alloxan induced diabetic Wistar rats and its effects on biochemical parameters in diabetic rats. Food Sci. Biotechnol. 2021, 9, 1185–1193. [Google Scholar] [CrossRef]
- Y. Sakuramata, H. Ohe, and S. Kusano. Anti-diabetic effects of combination of white skinned sweet potato (Ipomoea batatas L.) with loquat leaf extract. J. Tradit. Med. 2004, 21, 237–240. [Google Scholar] [CrossRef]
- E. Cremonini, E. Daveri, A. Mastaloudis, and P. I. Oteiza. (-)-Epicatechin and Anthocyanins Modulate GLP-1 Metabolism: Evidence from C57BL/6J Mice and GLUTag Cells. J. Nutr. 2021, 151, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- A. P. Hjørne, I. M. Modvig, and J. J. Holst. The Sensory Mechanisms of Nutrient-Induced GLP-1 Secretion. Metabolites, 2022, 12, 420. [Google Scholar] [CrossRef] [PubMed]
- R. E. Kuhre, C. R. Frost, B. Svendsen, and J. J. Holst. Molecular mechanisms of glucose-stimulated GLP-1 secretion from perfused rat small intestine. Diabetes 2015, 64, 370–382. [Google Scholar] [CrossRef]
- Y. Jiang et al. GLP-1 improves adipocyte insulin sensitivity following induction of endoplasmic reticulum stress. Front. Pharmacol., 2018, 9, 1–10. [Google Scholar] [CrossRef]
- L. Wu et al. GLP-1 regulates exercise endurance and skeletal muscle remodeling via GLP-1R/AMPK pathway. Biochim. Biophys. Acta - Mol. Cell Res., 2022, 1869. [CrossRef]
- O. I. Omotuyi et al. Flavonoid-rich extract of Chromolaena odorata modulate circulating GLP-1 in Wistar rats: computational evaluation of TGR5 involvement. 3 Biotech 2018, 8. [CrossRef]
- A. W. Wang, Yao, H. Alkhalidy, J. Luo, E. Moomaw, A. N. P., and D. Liu. Flavone Hispidulin Stimulates Glucagon-Like peptide-1 secretion and Ameliorates Hyperglycemia in Streptozotocin-Induced Diabetic Mice. Mol Nutr Food Res 2020, 64, 1–26. [Google Scholar] [CrossRef]
- T. S. de Castilho, T. B. Matias, K. P. Nicolini, and J. Nicolini. Study of interaction between metal ions and quercetin. Food Sci. Hum. Wellness 2018, 7, 215–219. [Google Scholar] [CrossRef]
- L. Horáková. Flavonoids in prevention of diseases with respect to modulation of Ca-pump function. Interdiscip. Toxicol. 2011, 4, 114–124. [Google Scholar] [CrossRef]
- L. F. de Almeida et al. Phytochemical profile, in vitro activities, and toxicity of optimized Eugenia uniflora extracts. Bol. Latinoam. y del Caribe Plantas Med. y Aromat. 2023, 22, 130–144. [Google Scholar] [CrossRef]
- Y. S. Oh and H. S. Jun. Effects of glucagon-like peptide-1 on oxidative stress and Nrf2 signaling. Int. J. Mol. Sci. 2018, 19, 1–16. [Google Scholar] [CrossRef]
- E. Barber, M. J. Houghton, and G. Williamson. Flavonoids as human intestinal α-glucosidase inhibitors. Foods 2021, 10, 1939. [Google Scholar] [CrossRef]
- S. Pal, S. Gautam, A. Mishra, R. Maurya, and A. K. Srivastava. Antihyperglycemic and antidyslipidemic potential of ipomoea batatas leaves in validated diabetic animal models. Int. J. Pharm. Pharm. Sci. 2015, 7, 176–186. [Google Scholar]
- J. Pan et al. Inhibition of Dipeptidyl Peptidase-4 by Flavonoids: Structure–Activity Relationship, Kinetics and Interaction Mechanism. Front. Nutr., 2022, 9, 1–17. [Google Scholar] [CrossRef]
- A. M. Ackermann and M. Gannon. Molecular regulation of pancreatic β-cell mass development, maintenance, and expansion. J. Mol. Endocrinol., 2007, 38, 193–206. [Google Scholar] [CrossRef]
- I. M. A. G. Wirasuta. Chemical profiling of ecstasy recovered from around Jakarta by High Performance Thin Layer Chromatography (HPTLC)-densitometry. Egypt. J. Forensic Sci. 2012, 2, 97–104. [Google Scholar] [CrossRef]
- L. Xi, T. Mu, and H. Sun. Preparative purification of polyphenols from sweet potato (Ipomoea batatas L.) leaves by AB-8 macroporous resins. Food Chemistry, vol. 172. Elsevier BV, pp. 166–174, 2015. [CrossRef]
- D. K. Olivier, B. E. van Wyk, and F. R. van Heerden. The chemotaxonomic and medicinal significance of phenolic acids in Arctopus and Alepidea (Apiaceae subfamily Saniculoideae). Biochem. Syst. Ecol. 2008, 36, 724–729. [Google Scholar] [CrossRef]
- A. Masek, E. Chrzescijanska, and M. Latos. Determination of antioxidant activity of caffeic acid and p-coumaric acid by using electrochemical and spectrophotometric assays. Int. J. Electrochem. Sci. 2016, 11, 10644–10658. [Google Scholar] [CrossRef]
- L. A. Wicaksono, Yunianta, and T. D. Widyaningsih. Anthocyanin extraction from purple sweet potato cultivar antin-3 (Ipomoea batatas L.) using maceration, microwave assisted extraction, ultrasonic assisted extraction and their application as anti-hyperglycemic agents in alloxan-induced wistar rats. Int. J. PharmTech Res. 2016, 9, 181–192, [Online]. Available: https://api.elsevier.com/content/abstract/scopus_id/84963724810. [Google Scholar]
- R. Dhanya. Quercetin for managing type 2 diabetes and its complications, an insight into multitarget therapy. Biomed. Pharmacother. 2022, 146. [CrossRef]
- X. C. Y. L. Xiaoqian Dai, Ye Ding, Zhaofeng Zhang. Quercetin and quercitrin protect against cytokine-induced injuries in RINm5F β-cells via the mitochondrial pathway and NF-κB signaling. Inflamm. Insul. Prod. β-cells Mitochondrial apoptosis Quercetin Quercitrin 2012, 31, 265–271. [Google Scholar]
- I. M. A. G. Wirasuta, N. M. A. R. Dewi, K. D. Cahyadi, L. P. M. K. Dewi, N. M. W. Astuti, and I. N. K. Widjaja. Studying systematic errors on estimation decision, detection, and quantification limit on micro-TLC. Chromatographia, 2013, 76, 1261–1269. [Google Scholar] [CrossRef]
- S. Ben-Shlomo et al. Glucagon-like peptide-1 reduces hepatic lipogenesis via activation of AMP-activated protein kinase. J. Hepatol. 2011, 54, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- P. Solverson. Anthocyanin Bioactivity in Obesity and Diabetes : and Periphery. Cells 2020, 9, 1–21. [Google Scholar]
- T. Wang et al. Baicalin and its metabolites suppresses gluconeogenesis through activation of AMPK or AKT in insulin resistant HepG-2 cells. Eur. J. Med. Chem. 2017, 141, 92–100. [Google Scholar] [CrossRef] [PubMed]
- C. Chen, S. Tan, T. Ren, H. Wang, X. Dai, and H. Wang. Polyphenol from Rosaroxburghii Tratt Fruit Ameliorates the Symptoms of Diabetes by Activating the P13K/AKT Insulin Pathway in db/db Mice. Foods, 2022, 11, 636. [Google Scholar] [CrossRef]
- W. Y. Song, Y. Aihara, T. Hashimoto, K. Kanazawa, and M. Mizuno. (-)-Epigallocatechin-3-gallate induces secretion of anorexigenic gut hormones. Journal of Clinical Biochemistry and Nutrition 2015, 57, 164–169. [Google Scholar] [CrossRef]
- Y. Wang et al. Plasma adiponectin levels and type 2 diabetes risk: A nested case-control study in a Chinese population and an updated meta-analysis. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- H. Yanai and H. Yoshida. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression : Mechanisms and Perspectives. pp. 1–25, 2019. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



