
Submitted:
21 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
In livestock, brucellosis is mainly an asymptomatic disease except for abortion, therefore two serological tests are used for diagnosis as no single test is suitable. Abattoir samples enable a combination of culture, molecular and serological tests to detect brucellosis. This study assessed Brucella specific PCR (ITS-PCR) to detect brucellosis and to conduct molecular characterization of Brucella spp. isolated from PCR-positive livestock (n=565) slaughtered at abattoirs and the appropriate sample tissue(s). ITS-PCR detected Brucella DNA in 33.6% cattle, 14.5% sheep and 4.7% pig tissues. Impure Brucella cultures from PCR-positive tissues were 43.6% (44/94) cattle, 51.7% (15/29) sheep, and 50% (2/4) pigs with predominantly B. abortus identification with AMOS-PCR and low isolation of mixed B. abortus and B. melitensis in all species. In cattle 33% of isolates were from lymph nodes while in sheep 38.0% were from liver and kidney and only from tonsils in pigs (2/4). Brucella infections identified with AMOS-PCR were present in seropositive and mainly seronegative (75.6-100%) livestock with the potential to cause brucellosis during pregnancy or breeding. This study demonstrated the value of the polyphasic approach, especially with chronic infections and the potential risk of these asymptomatic animals.
Keywords:
Brucellosis
; Livestock
; Tissue samples
; culture positive
; AMOS-PCR
; Brucella abortus
; Brucella melitensis
; South Africa
1. Introduction
Brucellosis is a highly contagious zoonotic infection of humans, domestic and marine animals [1]. The Brucella species are facultative intracellular gram-negative, non-spore forming, cocco-bacilli bacteria [2,3,4] causing the disease called brucellosis. In animals, Brucella invades the host without any clinical symptoms, resulting and progressing to acute infection only when the bacteria replicate actively within the macrophages and other replication sites [4]. The infection in animals remains asymptomatic in most cases, or until the first pathological signs/symptoms appear [4]. Chronic infections occur when the bacterial load decreases after plateauing, with sporadic clinical symptoms when the infection localizes in the reproductive system of sexually mature animals, resulting in sterility in males and placentitis as well as abortion in females during pregnancies. It continues to spread amongst animals in the herd [5,6,7,8]. Infection is described as mostly self-limiting [8] due to low activation of the phagocytosis and other host defences related to innate immunity [9,10]. After the initial phase of the illness has passed, the majority of brucellosis symptoms are not pathognomonic, and the organism can persist over time in the mammary glands and supramammary lymphatic nodes of 80% of infected animals [11]. Brucella replicates extensively in the endoplasmic reticulum (ER) compartment within the host cells [12]. The host cells' specialized compartment where intracellular pathogens reside, prevents antibiotics from reaching them, thus, affecting the efficacy of current therapies [12]. The association of Brucella within the host cell ER provides optimal bacterial growth conditions and replications in organs such as spleen lymph nodes, liver, bone marrow, epididymis and placenta that is rich in reticuloendothelial cells [8]. In the chicken embryo model, the replication of Brucella abortus spreads to all tissue, with the liver and spleen being the most severely infected [13].
Gonzalez-Espinozo et al. [8] reviewed investigations to improve culture other than blood such as bone marrow aspirates, liver, and lymph nodes base on the rational to obtain specimens from macrophage-rich tissues where Brucella organisms multiply and concentrate that may increase bacterial recovery. Culture from these tissues remains far from resolved, due to its non-specific signs and symptoms, that are comparable to other febrile diseases, its slow growth rate on culture, and the complexity of its sero-detection, brucellosis remains difficult to diagnose [14,15]. The sensitivity of culture depends on the disease stage, Brucella spp., culture medium and technique being used, quality of circulating bacteria, and the number of contaminants present in the sample [16]. The skin, hair, limbs, blood, stomach, gut contents, bile, and other excretions of the animal as well as the facilities, can all contaminate the sample taken from the carcasses of the animals throughout the slaughter process in the abattoirs. [17]. It is crucial to minimize any surface contamination occurring in the abattoirs during the slaughtering process, using a hazard analysis critical control point (HACCP) plan, in order to effectively handle and regulate the microbiological hazards connected with meat products [18].
Several molecular and high-resolution phenotypic assays that allow the differentiation of Brucella spp., the biovars, and the traceability of the source have been published [19,20]. However, only the gold standard (culture) is capable of reliably diagnosing brucellosis [21,22]. Brucella isolates take up to 4-7 days for growth in the laboratory, however, an increased number of contaminants including fungi and bacteria are usually observed, resulting in the regular contamination of culture plates and the decreased sensitivity of bacteriological diagnosis [23]. The most common sample for brucellosis diagnosis is serum for serological tests which is not appropriate sample for culture. Investigation at abattoirs where various sample types can be collected offers a polyphasic approach. Serological tests such as Rose Bengal Test (RBT) is used as a screening test with high sensitivity and low specificity resulting in false positives. These can be confirmed through Compliment Fixation Test (CFT) and/or indirect enzyme-linked immunosorbent assay (iELISA) which both have high specificity, but the CFT is less sensitive than iELISA resulting in false negative results [24]. Modern molecular approaches are currently not widely implemented in low-income nations where brucellosis is endemic in livestock [14,25]. This may be due to resource constraints. However, Brucella genus specific PCR assays (conventional and real-time) such as 16-23S ribosomal DNA interspacer (ITS) region, bcsp31 and IS711-based assays have been used as well as multiplex PCR assays namely AMOS- PCR for B. abortus bv 1,2 and 4, B. meltensis bv1-3, B. ovis and B. suis bv 1 and Bruce-ladder PCR assay that identify all Brucella spp. [26].
The Brucella genus currently consists of twelve species of which four species are pathogenic to humans [27]. Brucella melitensis and B. abortus commonly infect small ruminants and cattle, respectively, leading to abortions and infertility and thus resulting in significant economic losses [28]. Five Brucella spp. have been discovered in wildlife and marine mammals, while four additional distinct strains have been discovered in rodents, frogs, baboons, and humans [29]. The most pathogenic species for human brucellosis is B. melitensis, followed by B. suis, and then B. abortus [29]. Brucella ceti, B. inopinata and B. canis (rarely) are also known to cause human brucellosis [29].
Brucellosis infected animals are the primary cause of human brucellosis, a persistent illness that can have serious side effects if neglected [30]. Despite brucellosis being a notifiable disease in many countries, official statistics do not accurately reflect the number of cases that are reported each year [31]. Most developing countries in Africa have listed brucellosis as an under-reported endemic infection, this is due to the limited number of studies and the lack of epidemiological data [25]. In South Africa, B. abortus and B. melitensis have been reported in humans, cattle, sheep, and goats at the turn of the century [32,33]. Cattle population contribute to the majority of the income in South Africa and thus bovine brucellosis has a significant negative economic impact on the country's dairy and beef industries [34]. A nationwide bovine brucellosis eradication program has existed in South Africa from 1979 [35] that includes the vaccination of heifers and test and slaughter of high-risk bovine such as dairy and export. The scheme is voluntary for other animal owners and depends on the resources and willingness of the owners [36]. Most owners are aware that a positive test results in quarantine and a test and slaughtering regime which limits participation and knowledge of brucellosis seroprevalence amongst livestock in SA. Despite the scheme, bovine brucellosis seroprevalence has increase from 3.74% to 9.18% based on retrospective results reported in SA from 2007-2015 [37]. The aim of this study was to use a polyphasic approach to I) screen tissue samples using Brucella ITS-PCR for Brucella DNA; (II) culture Brucella from ITS-PCR positive tissues using selective medium; (III) assess the most appropriate sample type (lymph nodes, spleen, kidney, liver and tonsils (latter only from cattle and pigs) to isolate Brucella; and (IV) characterize Brucella spp. from culture using AMOS-PCR assay from seropositive and sero-negative livestock (cattle, sheep, and pigs) slaughtered at abattoirs in the Eastern Cape province, SA.
2. Materials and Methods
2.1. Description of the Study Area
This study was based on voluntary participation from abattoirs in the Eastern Cape Province, South Africa. The Brucella isolates characterized in this study were recovered from cattle, pigs, and sheep (lymph nodes, liver, spleen, kidney, and tonsils (latter from cattle and pigs) collected from the abattoirs. The Eastern Cape (at 168,966 km2) has the largest percentage of livestock in the country [38] and stretches along the Indian Ocean between Western Cape and KwaZulu-Natal province. The collection of samples was from five abattoirs in Eastern Cape Province, but the livestock slaughtered were not only from the Eastern Cape but included livestock transported from bordering provinces such as KwaZulu-Natal and Free State as these provinces do not have any movement control. Western Cape Province is the only province enforcing movement control of foot and mouth disease susceptible animals into and out of the province in South Africa [39].
2.2. Study Design and Sample Size
The abattoirs recruited for this study included both high throughput and low throughput abattoirs. The target animal population was livestock (sheep, cattle, and pigs) from apparently healthy animals with unknown Brucella status. During the abattoir visits, blood (serum) and tissue (kidney, spleen, liver, tonsils, and lymph nodes) samples were collected from corresponding animals. For this study, tissue samples were collected from 565 animals, made up of 280 cattle, 200 sheep, and 85 pigs. This number is not representative of the population ratio of 12.7 million cattle, 22.3 million sheep and 1.4 million pigs in South Africa (https://www.agriseta.co.za/wpcontent/uploads/2021/02/Agriseta_Red_Meat_SSSP_DIGITAL.pdf) as the number of species sampled depended on the number of animals slaughtered at the abattoir on the day of collection. Samples were collected in a sterile plastic bag and stored at -20°C at the University of Pretoria, Department of Veterinary Tropical Diseases Biosafety Level 2+ laboratory prior to processing.
2.3. Samples Collection Procedure
An opportunistic sampling procedure was followed for the collection of the samples. Multiple animal species are slaughtered in these selected abattoirs on any given day. Animals were sampled consecutively from within a randomly selected subset of a single species. That is, for every species, the daily quota of animals was sampled one after the other to ensure accurate sampling and assignation of samples per species. Our approach was carried out in three steps: (I). Planning: The relevant information was requested from the abattoir managers regarding the animals and herd information, this included the age, sex, vaccination status, location, and owners or seller of the animals. However, the animal, herd and vaccination information were not available, while only the abattoir and regional veterinary services are allowed access to the location and owner information which must be requested through official procedures by the regional state veterinary office which were obtained. (II). Sample collection: The selected abattoirs in the Eastern Cape Province are located more than 100 kms apart, except for two which are located within the same region. We aimed to collect samples from herds as Brucella infection is a known herd disease [40]. Upon slaughtering of the animals by the butchers, the animals were immediately eviscerated, and all the organs were removed from the carcasses (Figure 1A and B). To avoid animal to animal contamination, the knifes were cleaned with boiling hot water between uses. Approximately 100 g of each tissue was excised (Figure 1C). The sample collection procedure was lengthy, since it also included a cursory meat inspection by our team and the abattoir meat inspector (Figure 1 D-F). Therefore, only one abattoir could be sampled per day. The tissues were stored in a clearly labelled sterile plastic bag followed by ~4°C cold chain in the abattoir. (III). Packing and transportation: The samples were stored in a -20 freezer prior to transportation to the University of Pretoria, Department of Veterinary Tropical Diseases, South Africa in triple layer packaging for processing in accordance with the National Road Traffic Act, 1996 (Act No. 93 of 1996).
2.4. Sample Processing
The excised tissues were processed according to set laboratory protocols in a bio-safety level (BSL) 2 plus laboratory. The kidney, spleen, liver, tonsils, and lymph nodes were examined for lesions and calcification. A cubic centimeter of healthy-looking tissue was dissected with a sterile surgical blade and aliquoted into two separate cryovials. These matching tubes were submitted for direct DNA extraction, PCR, and microbial isolation, respectively. The serological test results have been determined by [41] using serum samples were subjected to the RBT from Onderstepoort Biological Products, SA, CFT (conducted at Onderstepoort Veterinary Institute laboratory where the test is SANAS accredited for bovine, but not sheep and pigs), and the iELISA (IDVet, France) as per manufacturer recommendations.
2.5. Genomic DNA Extraction
DNA was extracted directly from all the tissue samples for Brucella spp. screening. This was done using the Pure-Link Genomic DNA Kit (tissue protocol) according to the instructions of the manufacturer (Thermo Fisher Scientific, USA).
2.6. Brucella Genus PCR Screening Using ITS
DNA amplification for detection of the target Brucella gene using genus-specific 16S-23S rRNA interspacer region (ITS) primers (ITS66: ACATAGATCGCAGGCCAGTCA and ITS279: AGATACCGACGCAAACGCTAC) were used for the detection of Brucella DNA in the tissues [42]. During culturing, colonies can be screen with ITS-PCR to detect Brucella colonies. Briefly, a PCR master mix of 12 μl was prepared as follows: 6.5 μl Dream Taq polymerase, 0.3 μl (0.2 μm) Forward primer, 0. 3μl reverse primer (0.2 μm) and 4.9 μl of nuclease-free water (Thermo Fisher Scientific, South Africa). From each sample, 3 μl of DNA was used in a 15 μl PCR reaction. The mix was amplified on a thermal cycler (Veriti, Thermo Fisher Scientific, USA) with a heated lid, preheated to 105°C. The PCR cycling condition consisted of 95°C for 3 minutes, followed by 35 cycles of 95°C for 1 minute, 60°C for 2 minutes, 72°C for 2 minutes and a final extension of 72°C for 5 minutes. The target DNA has a product size of 214 bp determined with agarose electrophoresis. The positive controls for used were B. abortus bv 1 strain (BCCN R4) and B. melitensis Rev 1 (Onderstepoort Biological Products, SA). The amplified products were examined by electrophoresis in a 2% agarose gel (agarose LE, Lasec) and stained with ethidium bromide (0.03μl/ml). The gel ran at 120 volts for 1 hour. The gel was documented under UV light by a molecular imager (Bio-rad, ChemiDocTM XRS, USA).
2.7. Sample Preparations and Brucella Culture
Each tissue was homogenized with 1 mL of ddH2O in a Precellys 24 lysis and tissue homogeniser (Bertin technologies, France). About 200 μl of the tissue homogenate from pre-screened Brucella ITS-PCR positive tissues were inoculated onto the modified CITA medium [43] and incubated at 37°C with 5.0% CO2 for 5-14 days, including subculturing for purification where necessary. Culture plates were considered negative and discarded following 14 days of incubation with no growth observed.
2.8. Bacteriological Examination
ITS-PCR positive tissues were cultured on modified CITA medium. Brucella suspected isolates were selected base on their morphology on the CITA medium, Gram staining and modified Ziehl-Neelsen stain [44]. DNA was extracted from all the Brucella suspected (Gram negative and modified Ziehl-Neelsen) isolates for molecular characterization using the Pure-Link Genomic DNA Kit (gram negative protocol). During purification suspect Brucella single colonies were transferred to CITA medium and screened using staining and/or ITS-PCR. Fast growing bacteria kept on overgrowing slow growing Brucella colonies observed with Gram staining and ITS-PCR. Gram-negative fast-growing isolates were selected on the culture plates and submitted for genomic DNA extraction (Pure-Link Genomic DNA Kit; section 2.5) and 16S sequencing (see section 2.9). These organisms grew on modified CITA medium in the presence of antibiotics (natamycin, nitrofurantoin, amphotericin B, colistin, nystatin and vancomycin). On culture, Brucella spp. isolates were overgrown by fast growing organisms thus resulting in impure isolation of Brucella. AMOS-PCR assay was used to identify Brucella spp. from DNA extracted (Pure-Link Genomic DNA Kit; section 2.5) from impure Brucella cultures isolates from livestock tissues (see section 2. 10) .
2.9. Identification of Fast-Growing Contaminants
The metagenomic analysis of full length 16S gene amplicons were conducted by Inqaba biotec, SA. Isolated DNA Samples were sequenced on the Sequel system by PacBio (www.pacb.com). Raw sub-reads were processed through the SMRTlink (v11.0) Circular Consensus Sequences (CCS) algorithm to produce highly accurate reads (>QV40). These highly accurate reads were processed through DADA2 (https://benjjneb.github.io/dada2/index.html) and qiime2 (https://docs.qiime2.org/2021.11/) for quality control assessment and taxonomic classification, respectively.
2.10. AMOS-PCR and Bruce-Ladder PCR Assays
The multiplex AMOS PCR include species specific primers, B. abortus (F-GAC GAA CGG AAT TTT TCC AAT CCC), B. melitensis (F-AAA TCG CGT CCT TGC TGG TCT GA), B. ovis (F-CGG GTT CTG GCA CCA TCG TCG GG), B. suis (F-GCG CGG TTT TCT GAA GGT GGT TCA) and reverse primer IS711 (R-TGC CGA TCA CTT AAG GGC CTT CAT) as described [22]. Four species-specific forward primers were used at a final concentration of 0.1 μM with 0.2 μM reverse primer IS711. PCR cycling condition consisted of an initial denaturation at 95°C for 5 minutes followed by 35 cycles of 95°C for 1 minute, 55.5°C for 2 minutes, 72°C for 2 minutes and a final extension step at 72°C for 10 minutes. Specific amplicon sizes were determines using agarose electrophoresis.
As described by [19,45], a multiplex Bruce-ladder PCR experiment was performed to identify and distinguish between vaccine strains and field isolates of Brucella spp. The positive controls for used were B. abortus bv 1 strain (REF 544, BCCN R4), B. abortus S19 (Design Biologix, SA) and B. melitensis Rev 1 (Onderstepoort Biological Products, SA). The amplified products were examined by electrophoresis in a 2% agarose gel (agarose LE, Lasec) and stained with ethidium bromide (0.03 μl/ml). The gel ran at 120 volts for 1 hour. The gel was documented under UV light by a molecular imager (Bio-rad, ChemiDocTM XRS).
2.11. Statistical Analysis
Descriptive analysis was used to determine the frequency (percentage) of Brucella PCR positivity among the different variables (abattoir, throughput, animal species, sex of animal). Univariate analysis using the chi-squared or Fisher’s exact test were used to determine association between each of the four variables and PCR positivity. This was followed by multivariable analysis using Generalised Linear models with a stepwise backward elimination procedure and Akaike Information Criteria, to determine the risk factors for Brucella infection. Data analyses were perfumed using R statistical software version 4.21 [46] at 0.05 level of significance.
2.12. Ethical Considerations
The approvals from the Research, and Animal Ethics Committees of the University of Pretoria (Ref: REC 028-22), and the Eastern Cape Department of Agriculture and Rural Development were obtained prior to the commencement of the study. Section 20 of the Animal Diseases Act, (Act No. 35 of 1984) approval was obtained from Department of Agriculture, Land Reform and Rural Development (DALRRD). Appropriate health and safety precautions with risk assessments were followed throughout collection and processing of the samples.
The approvals from the Research, and Animal Ethics Committees of the University of Pretoria (Ref: REC 028-22), and the Eastern Cape Department of Agriculture and Rural Development were obtained prior to the commencement of the study. Section 20 of the Animal Diseases Act, (Act No. 35 of 1984) approval was obtained from Department of Agriculture, Land Reform and Rural Development (DALRRD). Appropriate health and safety precautions with risk assessments were followed throughout collection and processing of the samples.
3. Results
3.1. Identification of Brucella spp. Directly from the Tissues Using 16S-23S Ribosomal DNA Interspacer Region (ITS) PCR Assay
Of the tissue samples from the 280 slaughtered cattle tested using the Brucella ITS-PCR consisting of 214 bp product for the screening of Brucella DNA, the frequency of detection was 33.57% (94/280) (Supplementary Figure 1) Of the 200 slaughtered sheep tested using the ITS-PCR, the frequency of detection was 14.5% (29/200). Of the tissue samples from the 85 slaughtered pigs tested using the ITS-PCR, the frequency of detection was 4.71% (4/85).
3.2. Identification of Gram-Negative Isolates Using Gram Staining
Tissues from Brucella ITS PCR positive animals (127/565) were included in culturing, after which; round, smooth margin, translucent, yellowish-white coloured colonies on modified CITA medium were examined using microscopy and staining. Of the 94 cattle tissues that tested positive on ITS-PCR, 41 Brucella isolates were identified based on Gram-negative coccobacilli on Gram's staining and positive for modified Ziehl Neelsen staining. Fifteen [15] Brucella suspect culture from 29 ITS-PCR positive sheep tissues were identified using microscopy. Additionally, 2 Brucella cultures were observed from 4 ITS-PCR pig tissues based on microscopy. Brucella colonies were further subjected to several rounds of streaking and dilution to purify the colonies. Additional fast-growing gram-negative bacteria were also observed on culture. Spingomonas was identified among other bacteria, this has identical antibiogram thus making it impossible to select and purify Brucella from this faster growing contaminant using antibiotics. None of the isolations could be purified and remain impure isolates, which we identified with AMOS-PCR.
3.3. Characterisation of Brucella spp. Using AMOS PCR Assay and Seropositivity
Of the 41/94 (43.6%) Brucella suspect isolates observed on microscopy from ITS-PCR positive cattle tissues, AMOS-PCR characterised 38 as B. abortus and a mixed infection of both B. abortus and B. melitensis were observed in 3 cattle (Table 1). From the 15/29 (51.7%) Brucella suspect isolates from ITS-PCR positive sheep tissues, AMOS-PCR characterise 11 as B. abortus and a mixed infection of both B. abortus and B. melitensis were observed in 4 sheep (Figure 2A). Of the 2/4, (50%) Brucella suspect isolates from ITS-PCR positive pig tissues, AMOS-PCR characterised 1 as B. abortus and 1 as a mixed infection of B. abortus and B. melitensis (Table 1, supplementary data). The single-plex AMOS PCR was used to separate and confirm the mixed infection of B. abortus and B. melitensis (Figure 2 B & C). Using the AMOS-PCR and Bruce-ladder PCR assays, the isolates were distinguished from the vaccine strain (Supplementary Figure 2).
Brucella spp. identified with AMOS-PCR identified 14.6% (41/280), 7.3% (15/200) and 2.4% (2/85) from cattle, sheep and pig tissue collected from Eastern Cape abattoirs (Table 1). Seropsotivity based on one or more serological tests (RBT, CFT and/or iELISA of Brucella infected animals identified with AMOS-PCR consisted of 24.4% (10/41) cattle, 13.3% (2/15) sheep and no pigs (Table 1). See Table 1 for the animals that were AMOS-PCR Brucella spp. infected and seronegative.
3.4. Brucella Isolation amongst Livestock Stratified by Tissue
Brucella isolation from ITS-PCR positive tissues and identified with AMOS-PCR stratified by cattle tissue were 33.0% (31/94) from lymph nodes, 26.6% (25/94) from liver, 21.3% (20/94) from spleen, 20.2% (20/94) from kidney and 10.6% (10/92) from tonsils. With sheep tissues AMOS-PCR Brucella isolates were 37.9% (11/29) in liver and kidney, 34.5% (10/29) in spleen and 27.6% (8/29) from lymph nodes. No tonsils samples were collected from sheep as the abattoirs sell the head intact. Brucella isolates identified with AMOS-PCR from pigs were isolated from tonsils (50%, 2/4) (Table 1).
3.5. Association between Brucella ITS-PCR Positivity and Predictor Variables
Three (abattoir, throughput, and animal species) of the four variables analyzed in univariate analyses showed statistical significance (p≤0.05) (Table 2). The four variables regardless of p value were included in a multivariable logistic regression model. After multivariable analysis that followed a backward stepwise elimination procedure, only three variables (sex, species, abattoir) out of the four comprised the final regression model (Table 3). The abattoir factor was a significant determinant for positivity amongst the specimens from different animal species. With abattoir B as the reference level, animals in abattoir D (39.1%; OR=7.0, p=0.00014), abattoir E (41.7%; OR=5.13, p<0.0001), abattoir A (38.0; OR=4.9, p<0.0001) were more likely to be PCR positive for Brucella spp., while abattoir C (15.6%, OR=0.91, p=0.85) had similar positivity rate (Table 2 and Table 3).
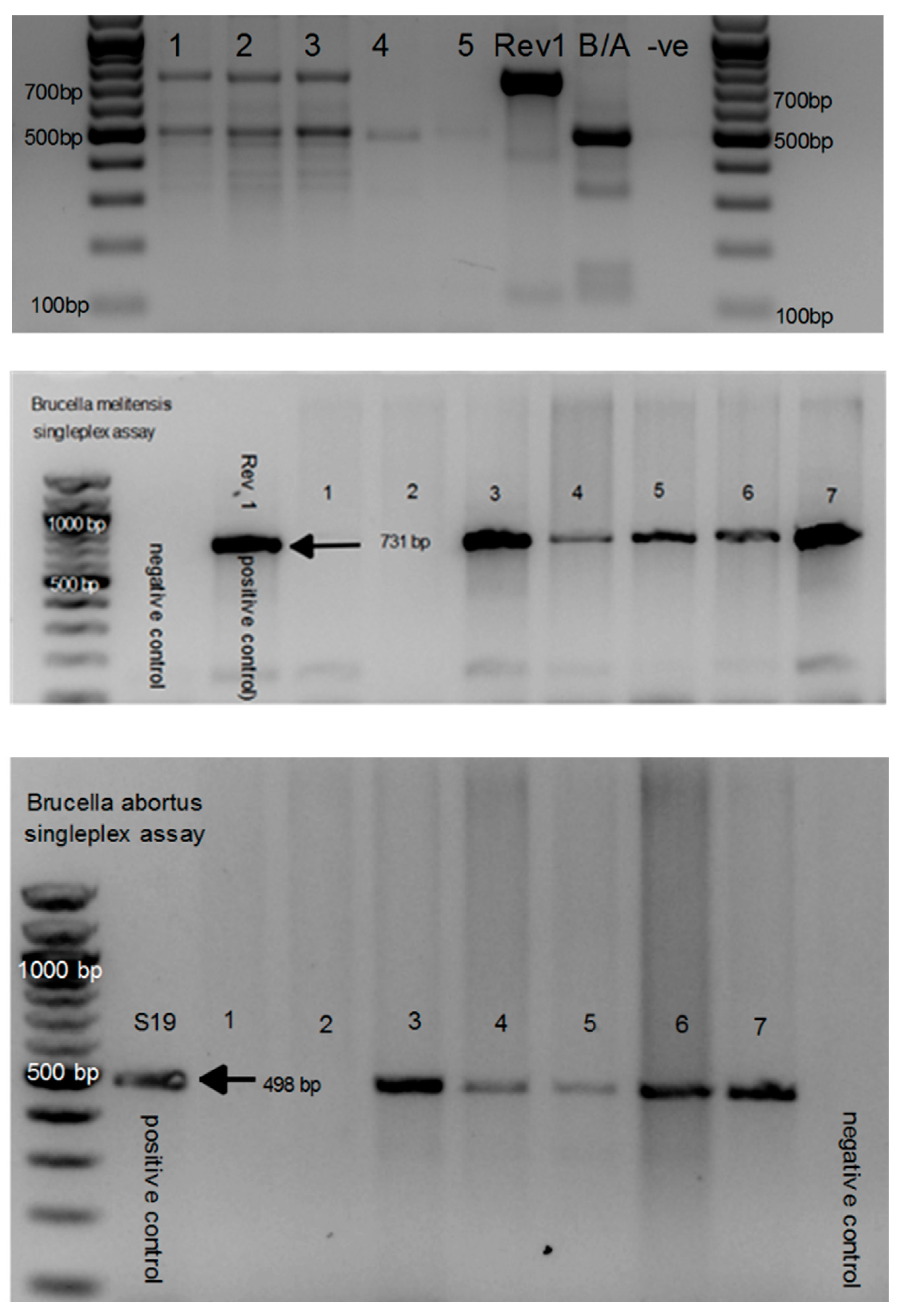
Figure 2.
Amplification of Brucella isolates from sheep tissues using AMOS-PCR with 498 bp amplifying B. abortus target region and 731 bp amplified B. melitensis target region. A. Multi-plex AMOS-PCR with are mixed infection of both B. abortus and B. melitensis isolated from the kidneys in lanes 1-3; B. abortus isolated from the liver in lane 4; negative control in lane 5 and 9 (-ve); B. melitensis Rev 1 and B. abortus positive controls in lane 6 and 7. B: Single-plex B. abortus specific primer of AMOS-PCR with negative water control and 731 bp B. melitensis PCR product using B. melitensis Rev 1 positive control; lanes 1-7 included tissues that were AMOS negative in lanes 1 and 2; Lane 3-7 included mixed B. abortus and B. melitensis sheep isolates from 4 sheep (with the same animals repeated in lanes 3 and 7). C: Single-plex B. abortus specific primer of AMOS-PCR with negative water control and 498 bp B. abortus PCR product using B. abortus S19 positive control; lanes 1-7 included tissues that were AMOS negative in lanes 1 and 2; Lane 3-7 included mixed B. abortus and B. melitensis sheep isolates from 4 sheep (with the same animals repeated in lanes 3 and 7).
Figure 2.
Amplification of Brucella isolates from sheep tissues using AMOS-PCR with 498 bp amplifying B. abortus target region and 731 bp amplified B. melitensis target region. A. Multi-plex AMOS-PCR with are mixed infection of both B. abortus and B. melitensis isolated from the kidneys in lanes 1-3; B. abortus isolated from the liver in lane 4; negative control in lane 5 and 9 (-ve); B. melitensis Rev 1 and B. abortus positive controls in lane 6 and 7. B: Single-plex B. abortus specific primer of AMOS-PCR with negative water control and 731 bp B. melitensis PCR product using B. melitensis Rev 1 positive control; lanes 1-7 included tissues that were AMOS negative in lanes 1 and 2; Lane 3-7 included mixed B. abortus and B. melitensis sheep isolates from 4 sheep (with the same animals repeated in lanes 3 and 7). C: Single-plex B. abortus specific primer of AMOS-PCR with negative water control and 498 bp B. abortus PCR product using B. abortus S19 positive control; lanes 1-7 included tissues that were AMOS negative in lanes 1 and 2; Lane 3-7 included mixed B. abortus and B. melitensis sheep isolates from 4 sheep (with the same animals repeated in lanes 3 and 7).
3.6. Sequence Identification of Additional Gram-Negative and Positive Isolates from Culture
Faster growing contaminants were a recurring hindrance to obtaining pure Brucella isolates. To identify the contaminants and in doing so, improve the selective media, isolates were sequenced. The following isolates were identified by nucleotide identity using QIIME2. Proteus vulgaris (21%), Cuktibacterium acnes (3%), Brevundiminas terrae, Brevundimonas naejangsanensis (20%), Serratia nematodiphila (3%) and Serratia marcescens (24%) were identified on culture from the livestock tissue samples (Figure 3).
4. Discussion
This study used samples available from abattoirs to investigate brucellosis which allowed a polyphasic approach and thus serology, molecular and bacteriology detection while most studies only use serology, and few continue to obtain Brucella culture isolates. Brucella specific PCR on tissues from livestock followed by culture and AMOS-PCR identification detected mainly B. abortus with a few mixed infections of B. melitensis in 14.6% (41/280) cattle, 7.5% (15/200) sheep and 2.4% (2/85) pig tissues collected from Eastern Cape Province abattoirs. This study demonstrated the value of the polyphasic approach, especially to identify the potential risk of brucellosis in asymptomatic animal with possible chronic infections.
This study isolated Brucella spp. from the liver, spleen, kidney, lymph nodes (mesenteric and mandibular) and tonsils of apparently healthy livestock from the abattoirs in the Eastern Cape Province. Tissue samples from livestock including liver, spleen, kidneys, lungs, and lymph nodes have previously been processed for the isolation Brucella spp. [8,47]. In this study, of the 58 AMOS-PCR identified Brucella isolates, 19.0% (11/58) were seropositive using either RBT, CFT or and iELISA [41] with the majority being seronegative. The isolation of Brucella spp. from seronegative animals (see supplementary data) may be an indication of chronic infection in the animals [48], with these asymptomatic animals posing a risk to spread the pathogen once they become pregnant or during breeding as Brucella will then start to replicate . Disease surveillance from live animals using serological tests is limiting and cannot detect latent or chronic infected animals and thus show the value of the sample availability combined with molecular methods at abattoirs to determine this risk to contribute to disease spread and spillover [49].
Although the culture technique is not a sensitive procedure, bacterial isolation is considered as the gold standard for diagnosing Brucella spp. in human and animals [50]. Aborted tissues from a B. abortus abortion episodic yield more than 1014 microbial organisms, which constitutes 105 times the presumed infectious dosage of heifers vaccinated with S19 [51]. Hence, increase isolation of Brucella spp. on culture has been reported when sampling from aborted materials and vaginal swaps [52], as compared to tissues from asymptomatic and apparently healthy animals. Thus, Brucella specific PCR was used for tissue screening before attempting low sensitivity isolation especially as the samples were collected from asymptomatic livestock. However, tissues were collected from organs with ER cells such as spleen, liver, kidney, and lymph nodes as these are macrophage-rich tissues where Brucella organisms multiply and concentrate and thus increase culture sensitivity [8] and can only be collected from dead animals [23]. The Brucella spp. isolation frequency of cattle was higher in lymph nodes (31/94), followed by liver (25/94) and spleen (20/94). Whereas the frequency of isolation of Brucella spp. in sheep was higher in liver (11/29) and kidney (11/29), followed by spleen (10/29). Brucella isolates were only recovered from tonsils from tonsils (2/4) in pigs. Brucella spp. were isolated and detected with AMOS-PCR from the lymph nodes, liver, spleen, and kidney samples from animals showing no clinical signs of brucellosis infection. Thus, suggesting that the above-mentioned tissues may be utilised for brucellosis screening purposes and diagnostics in slaughtered abattoir animals. This study also highlights improved assessment standards and procedures that may result from routine sampling, such as obtaining tonsils from monogastric animals and liver, kidneys, and spleen from ruminants.
This study further shows the presence of fast-growing contaminants that made isolation of low concentration Brucella in asymptomatic tissue impossible despite various attempts. Brucella organisms’ isolation from vaginal secretions, placenta, fetal tissues, milk, and semen from animals are normally impaired by contaminants with short generation times that overgrown slow growing brucellae even on selected media [8]. The presence of other fast-growing, gram-negative bacteria on culture affects the growth of Brucella spp. through competitive inhibition thus resulting in impure/contaminated isolates. The present study reports isolation of other pathogenic organisms such as Proteus Vulgaris, Serratia marcescens and Brevundimonas naejangsanensis. Proteus vulgaris has been reported as a zoonotic infection which is mainly known for causing wound and urinary infections in humans [53]. Previous researchers have reported S. marcescens as a common cause of mastitis and early abortions in cows [54,55]. B. naejangsanensis is an environmental gram-negative bacterium which has been isolated from the soil [56]. The risk of zoonotic disease is increased by the isolation of potentially harmful foodborne such as Brucella spp. and P. vulgaris [57], from apparently healthy abattoir livestock. Microbial contamination of the abattoir meat may occur during the exsanguination process, particularly if a sterile environment is not maintained [58]. Based on our study, it was observed that the butcher only washes the knife to remove the excessive amount of blood and not to avoid contamination from one animal to the other. It was also observed that the operators clean/spray the floors frequently for blood removal, however this process allows contaminated water/blood to splash onto the meat. According to [58], they reported the blood removal procedure on the floor as unhygienic. To reduce these contaminants, it is advisable to surface sterilize the tissues before culturing which can reduce these contaminants.
In this study, on gross pathological examination, yellowish-white lesions, discoloration/bruises, abscesses, and cysts were observed on some cattle tissues. This included the mesenteric lymph nodes, skin, liver, and the spleen. The presence of lesions in the mesenteric lymph nodes can indicate Mycobacterium Tuberculosis Complex infection. As reported by a similar study conducted in the Eastern Cape abattoirs, the presence of nodular lesions was observed from 162 cattle lymph node samples with visible inflammation. Their study reported the isolation of Mycobacterium bovis and M. tuberculosis [59]. Feedlot cattle may develop liver abscesses as a result of vigorous grain-feeding programs, which are also influenced by a number of nutritional and management factors [60]. Our findings are in agreement with other studies which identified major causes of offal and carcass condemnation in the Eastern Cape abattoirs including tongue and spleen abscess, bruises, actinobacillosis, heart and kidney cysts, inflammatory conditions, and improper evisceration [61]. However, the underlying causes of the conditions remains unknown. Due to the tissue condemnation and decreased meat yield, the presence of pathological evidences on the tissue has a major economic impact on the animal industry [60] and increases the risks of zoonotic infections to humans.
Multivariable analyses showed that sheep (14.5%; OR=5.6, p=0.0043) and cattle (33.6%; OR=17.1, p<0.0001) were significantly more likely to be AMOS-PCR positive for Brucella species compared to pigs (4.7%). The current study reports the isolation of Brucella spp. from 43.6% (41/94) cattle, 51.7% (15/29) sheep and 50% (2/4) in pig samples using AMOS-PCR that only detects B. abortus bv 1, 2 and 4, B. meiltensis bv 1-3, B. ovis and B. suis bv 1. A similar study conducted in the Eastern Cape Province reported an increased isolation of Brucella spp. from cattle (62.3%) as compared to goats (25.4%) and sheep (12.3%) also using AMOS-PCR The current bovine brucellosis scheme includes a mandatory vaccination of heifers aged 4-8 months, serological testing, surveillance of high-risk farms, particularly dairy and breeding cattle with suspected or proven brucellosis infections. [62,63]. However, the participation of the farmers is voluntary and self-funded, thus negatively affecting the role and importance of early vaccination. None of the B. abortus isolates from the livestock tissue were S19 vaccine strain. This study indicates almost similar likelihood of Brucella positivity between male (21.1%) and female animals (23.9%) with an odds ratio between the two levels of 0.5. The abattoirs (except abattoir C), species and sex were a significant determinant for positivity in our study with a p≤0.05. The Brucella positivity in male animals may be due to high exposure of the bacteria or through consumption of milk from infected females. An increased positivity was observed from low throughout abattoirs (39.1%) as compared to high throughput abattoirs (21.8%). As reported by [64], an increased sero-positivity and isolation of Brucella spp. was also observed from low throughput abattoirs as compared to high throughput abattoirs in Gauteng Province. This may be that low throughput abattoirs receive animals from local community with animals from the same herd or animals grazing together thus increasing the possibility of transmission amongst each other.
Brucellosis is a controlled zoonotic infection in animals and a notifiable disease in humans in South Africa [65]. The infection is a major public health challenge, and still predominant as a neglected endemic zoonosis requiring proactive considerations in numerous communities worldwide [66]. Serological tests have detected brucellosis throughout SA in bovine, however brucellosis outbreaks have been reported mainly in the central and highveld regions [67]. The brucellosis scheme in SA is focused on bovine and from this study B. abortus was the dominate species detected with AMOS-PCR in Brucella infected animals. Brucella abortus was not only detected in bovine but sheep and pigs which indicate spillover to these species in SA. A previous study conducted in the Eastern Cape, reported the isolation of B. abortus in cattle, sheep, and goat, whereas the isolation of B. melitensis was observed in sheep and goats [68]. As reported by [64], the first case in SA of B. melitensis in cattle was isolated from abattoirs in Gauteng Province. The current study reports the isolation of B. melitensis from cattle in the Eastern Cape Province. Serological tests can not differentiate between Brucella species and brucellosis seropositive bovine are presumed to be B. abortus while seropositive sheep and goats are presumed to be infected with B. melitensis. Mixed B. abortus and B. melitensis infections were also detected in all livestock in this study and needs investigation. Despite the tremendous efforts of the SA government in eradication of the infection, an increased number of reports continue to indicate the presence of brucellosis in livestock in SA (64, 68). Surveillance schemes in countries where brucellosis has been eradicated focus on vaccination of livestock as well as test and slaughtering scheme of all the relevant species [69],unlike SA that focus only on high-risk bovines [70]. Brucellosis eradications takes decades and are a costly exercise [71]. In endemic countries such SA, serological tests will have its limitation due to chronic infected animals that will not detect infected animals as well as latency since the antibody level is below detection in these animals. However serological tests will identify some infected animals but the results in this study indicated that testing should be expanded to all bovine as proposed but to other livestock species as well especially sheep and goats to increase the detected of brucellosis.
5. Conclusion
This study has demonstrated the importance of multiple tests in the diagnosis and surveillance of brucellosis, as it is evidenced by the isolation and identification of B. abortus and B. melitensis from sero-positive but mainly sero-negative asymptomatic livestock. The use of only serological tests in chronic infected animals’ results in false negative results. This study demonstrated the value of the polyphasic approach using the molecular method in combination with samples from abattoirs, especially to identify the potential risk of brucellosis in asymptomatic animal with possible chronic infections. This study also emphasises refined evaluation criteria and processes could come from routine sampling, i.e., collecting liver, kidneys and spleen from ruminants and tonsils from monogastric. Abattoirs prove a valuable surveillance resource as the tissues are easily accessible post slaughter. The more data included from such sites would allow for a much clearer epidemiological picture of brucellosis in provinces across SA. This could in turn provide better data with which to plan targeted surveillance for both B. abortus and B. melitensis infections in livestock, to make effective management decisions against this devastating herd disease.
6. Limitations of the study
Isolation of Brucella spp. was recovered from the livestock samples, however due to increased growth of other fast growing gram-negative bacteria, impure cultures were observed. Bruce-ladder PCR assay requires a high concentrated Brucella DNA to amplify the multiple targets of this assay. Mixed infections of B. abortus and B. melitensis were observed in all the species (cattle, sheep, and pigs), however due to the confluent growth of contaminants, the mixed Brucella spp. could not be isolated separately. Further investigation which will be possible in a larger study is recommended. This could include surface sterilization of tissues to reduce the growth of the other organisms thus allowing the Brucella spp. to grow confluently.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Molecular and serological identification of Brucella spp. in livestock results are included in the supplementary data. Gel electrophoresis of Bruce-Ladder PCR amplification to differentiate the field strains has also been included in the supplementary materials.
Declaration of interest
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in the writing of this article, and therefore declare no conflict of interest.
Ethics statement
The approvals from the Research, and Animal Ethics Committees of the University of Pretoria (Ref: REC 028-22), and the Eastern Cape Department of Agriculture and Rural Development were obtained prior to the commencement of the study. Section 20 of the Animal Diseases Act, (Act No. 35 of 1984) approval was obtained from Department of Agriculture, Land Reform and Rural Development (DALRRD). Risk assessment and appropriate health and safety measures were taken during the sample collection and processing. The identity of the abattoir is kept private in accordance with the terms of the research ethics approval, participant permission, and agreement of inclusion for this study.
Role funding
The authors would like to thank the Department of Veterinary Tropical Diseases (AgriSETA), South Africa; Institute of Tropical Medicine Antwerp, Belgium (FA4 & FA5) and UNICEF for the research funding. The corresponding author would also like to acknowledge the NRF-DAAD scholarship fund.
Acknowledgments
The authors are grateful to the management and workers of the abattoirs in the Eastern Cape for their participation in the study. We also like to express our appreciation to Dr Sunday Ochai and students from the University of Fort Hare for assisting with the sample collection.
References
- Foster G, Osterman BS, Godfroid J, Jacques I, Cloeckaert A. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. International journal of systematic and evolutionary microbiology. 2007, 57, 2688–2693.
- Seleem MN, Boyle SM, Sriranganathan N. Brucellosis: a re-emerging zoonosis. Veterinary microbiology. 2010, 140, 392–398.
- Eiken, M. Studies on an anaerobic rod-shaped. Gram-negative microorganism: Bacteroides corro. 1958.
- Deng Y, Liu X, Duan K, Peng Q. Research progress on brucellosis. Current medicinal chemistry. 2019, 26, 5598–5608.
- Ilhan Z, Aksakal A, Ekin I, Gülhan T, Solmaz H, Erdenlig S. Comparison of culture and PCR for the detection of Brucella melitensis in blood and lymphoid tissues of serologically positive and negative slaughtered sheep. Letters in applied microbiology. 2008, 46, 301–306.
- Poester F, Samartino L, Santos R. Pathogenesis and pathobiology of brucellosis in livestock. Rev Sci Tech. 2013, 32, 105–115.
- Megid J, Mathias LA, Robles C. Clinical manifestations of brucellosis in domestic animals and humans. The Open Veterinary Science Journal. 2010, 119–126.
- González-Espinoza G, Arce-Gorvel V, Mémet S, Gorvel J-P. Brucella: Reservoirs and niches in animals and humans. Pathogens. 2021, 10, 186.
- Levin BR, Baquero F, Ankomah PP, McCall IC. Phagocytes, antibiotics, and self-limiting bacterial infections. Trends in microbiology. 2017, 25, 878–892.
- Barquero-Calvo E, Chaves-Olarte E, Weiss DS, Guzman-Verri C, Chacon-Diaz C, Rucavado A, et al. Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PloS one. 2007, 2, e631.
- Rahman MS, Hahsin MFA, Ahasan MS, Her M, Kim JY, Kang SI, et al. Brucellosis in sheep and goat of Bogra and Mymensingh districts of Bangladesh. Korean Journal of Veterinary Research. 2011, 51, 277–280.
- Goswami A, Sharma PR, Agarwal R. Combatting intracellular pathogens using bacteriophage delivery. Critical Reviews in Microbiology. 2021, 47, 461–478.
- Detilleux P, Cheville N, Deyoe B. Pathogenesis of Brucella abortus in chicken embryos. Veterinary Pathology. 1988, 25, 138–146.
- Pappas G, Papadimitriou P, Akritidis N, Christou L, Tsianos EV. The new global map of human brucellosis. The Lancet infectious diseases. 2006, 6, 91–99.
- Shirima G, Fitzpatrick J, Kunda J, Mfinanga G, Kazwala R, Kambarage D, et al. The role of livestock keeping in human brucellosis trends in livestock keeping communities in Tanzania. Tanzania Journal of Health Research. 2010, 12, 203–207.
- Navarro E, Casao MA, Solera J. Diagnosis of human brucellosis using PCR. Expert Review of Molecular Diagnostics. 2004, 4, 115–123.
- Sofos, JN. Challenges to meat safety in the 21st century. Meat science. 2008, 78, 3–13. [Google Scholar] [CrossRef]
- Korsak N, Daube G, Ghafir Y, Chahed A, Jolly S, Vindevogel H. An efficient sampling technique used to detect four foodborne pathogens on pork and beef carcasses in nine Belgian abattoirs. Journal of Food Protection. 1998, 61, 535–541.
- Lopez-Goñi I, Garcia-Yoldi D, Marín C, De Miguel M, Munoz P, Blasco J, et al. Evaluation of a multiplex PCR assay (Bruce-ladder) for molecular typing of all Brucella species, including the vaccine strains. Journal of clinical microbiology. 2008, 46, 3484–3487.
- Le Flèche P, Jacques I, Grayon M, Al Dahouk S, Bouchon P, Denoeud F, et al. Evaluation and selection of tandem repeat loci for a Brucella MLVA typing assay. BMC microbiology. 2006, 6, 1–14.
- Probert WS, Schrader KN, Khuong NY, Bystrom SL, Graves MH. Real-time multiplex PCR assay for detection of Brucella spp., B. abortus, and B. melitensis. Journal of clinical microbiology. 2004, 42, 1290–1293.
- Bricker BJ, Halling SM. Differentiation of Brucella abortus bv. 1, 2, and 4, Brucella melitensis, Brucella ovis, and Brucella suis bv. 1 by PCR. Journal of clinical microbiology. 1994, 32, 2660–2666.
- De Miguel M, Marín CM, Muñoz PM, Dieste L, Grilló MJ, Blasco JM. Development of a selective culture medium for primary isolation of the main Brucella species. Journal of clinical microbiology. 2011, 49, 1458–1463.
- Nielsen, K. Diagnosis of brucellosis by serology. Veterinary microbiology. 2002, 90, 447–459. [Google Scholar] [CrossRef]
- McDermott JJ, Arimi S. Brucellosis in sub-Saharan Africa: epidemiology, control and impact. Veterinary microbiology. 2002, 90, 111–134.
- Yu WL, Nielsen K. Review of detection of Brucella spp. by polymerase chain reaction. Croatian medical journal. 2010, 51, 306–313.
- Barbuddhe SB, Vergis J, Rawool DB. Immunodetection of bacteria causing brucellosis. Methods in Microbiology. 47: Elsevier; 2020. p. 75-115.
- Shome R, Kalleshamurthy T, Rathore Y, Ramanjinappa KD, Skariah S, Nagaraj C, et al. Spatial sero-prevalence of brucellosis in small ruminants of India: Nationwide cross-sectional study for the year 2017–2018. Transboundary and Emerging Diseases. 2021, 68, 2199–2208.
- Al Dahouk S, Sprague L, Neubauer H. New developments in the diagnostic procedures for zoonotic brucellosis in humans. Rev Sci Tech. 2013, 32, 177–188.
- Zinsstag J SE, Solera J, Blasco JM, Moriyón I. Brucellosis. In: Palmer SR, Soulsby L, Torgeson PR, Brown DG. Handbook of zoonoses. Oxford, New York: Oxford University Press2011. p. 54-62 p.
- Musallam I, Abo-Shehada M, Omar M, Guitian J. Cross-sectional study of brucellosis in Jordan: prevalence, risk factors and spatial distribution in small ruminants and cattle. Preventive veterinary medicine. 2015, 118, 387–396.
- Schrire, L. Human brucellosis in South Africa. South African Medical Journal. 1962, 36, 342–349. [Google Scholar]
- Van Drimmelen, G. A urease test for characterizing Brucella strains. 1962.
- Tempia S, Mayet N, Gerstenberg C, Cloete A. Brucellosis knowledge, attitudes and practices of a South African communal cattle keeper group. Onderstepoort Journal of Veterinary Research. 2019, 86, 1–10.
- Govindasamy K, Etter EM, Geertsma P, Thompson PN. Progressive area elimination of bovine brucellosis, 2013–2018, in Gauteng Province, South Africa: Evaluation using laboratory test reports. Pathogens. 2021, 10, 1595.
- Simpson GJ, Marcotty T, Rouille E, Chilundo A, Letteson J-J, Godfroid J. Immunological response to Brucella abortus strain 19 vaccination of cattle in a communal area in South Africa. Journal of the South African Veterinary Association. 2018, 89, 1–7.
- Kolo FB, Adesiyun AA, Fasina FO, Potts A, Dogonyaro BB, Katsande CT, et al. A retrospective study (2007–2015) on brucellosis seropositivity in livestock in South Africa. Veterinary medicine and science. 2021, 7, 348–356.
- Goni S, Skenjana A, Nyangiwe N. The status of livestock production in communal farming areas of the Eastern Cape: A case of Majali Community, Peelton. Appl Anim Husb Rural Dev. 2018, 11, 34–40.
- Agriculture WCDo. Animal health and disease control 2023 (Available from: https://www.elsenburg.com/veterinary-services/animal-health-and-disease-control/.
- Langoni H, Ichihara SM, SILVA AVd, Pardo RB, Tonin FB, Mendonça LJP, et al. Isolation of Brucella spp from milk of brucellosis positive cows in São Paulo and Minas Gerais states. Brazilian Journal of Veterinary Research and Animal Science. 2000, 37, 444–448.
- Mazwi K, Kolo F, Jaja I, Byaruhanga C, Hassim A, Heerden Hv. Serological Evidence and Co-Exposure of Selected Infections Among Livestock Slaughtered at Eastern Cape Abattoirs in South Africa International Journal of Microbiology. 2023.
- Keid LB, Soares RM, Vasconcellos SA, Chiebao D, Salgado V, Megid J, et al. A polymerase chain reaction for detection of Brucella canis in vaginal swabs of naturally infected bitches. Theriogenology. 2007, 68, 1260–1270.
- Ledwaba MB, Matle I, Van Heerden H, Ndumnego OC, Gelaw AK. Investigating selective media for optimal isolation of Brucella spp. in South Africa. Onderstepoort Journal of Veterinary Research. 2020, 87, 1–9.
- Tripathi N, Sapra A. Gram staining. 2020.
- Weiner M, Iwaniak W, Szulowski K. Comparison of PCR-based AMOS, Bruce-Ladder and MLVA assays for typing of Brucella species. Bull Vet Inst Pulawy. 2011, 55, 625–630.
- Team RDC. R: A language and environment for statistical computing. (No Title). 2010.
- Hosein H, El-Nahass E, Rouby S, El-Nesr K. Detection of Brucella in tissues and in formalin-fixed paraffin-embedded (FFPE) specimens using PCR. Adv Anim Vet Sci. 2018, 6, 55–62.
- Spickler, AR. Brucellosis: Canine Brucellosis. The Center for Food Security and Public Health. 2018.
- El-Diasty M, Wareth G, Melzer F, Mustafa S, Sprague LD, Neubauer H. Isolation of Brucella abortus and Brucella melitensis from seronegative cows is a serious impediment in brucellosis control. Veterinary Sciences. 2018, 5, 28.
- Nielsen K, Yu W. Serological diagnosis of brucellosis. Prilozi. 2010, 31, 65–89.
- Corner L, Alton G, McNichol L, Streeten T, Trueman K. An evaluation of the anamnestic test for brucellosis in cattle of the northern pastoral area. Australian Veterinary Journal. 1983, 60, 1–3.
- Tekle M, Legesse M, Edao BM, Ameni G, Mamo G. Isolation and identification of Brucella melitensis using bacteriological and molecular tools from aborted goats in the Afar region of north-eastern Ethiopia. BMC microbiology. 2019, 19, 1–6.
- Mendoza-Roldan JA, Noll Louzada-Flores V, Lekouch N, Khouchfi I, Annoscia G, Zatelli A, et al. Snakes and Souks: Zoonotic pathogens associated to reptiles in the Marrakech markets, Morocco. PLOS Neglected Tropical Diseases. 2023, 17, e0011431.
- Friman MJ, Eklund MH, Pitkälä AH, Rajala-Schultz PJ, Rantala MHJ. Description of two Serratia marcescens associated mastitis outbreaks in Finnish dairy farms and a review of literature. Acta Veterinaria Scandinavica. 2019, 61, 1–11.
- Das A, Paranjape V, Pitt T. Serratia marcescens infection associated with early abortion in cows and buffaloes. Epidemiology & Infection. 1988, 101, 143–149.
- Kang S-J, Choi N-S, Choi JH, Lee J-S, Yoon J-H, Song JJ. Brevundimonas naejangsanensis sp. nov., a proteolytic bacterium isolated from soil, and reclassification of Mycoplana bullata into the genus Brevundimonas as Brevundimonas bullata comb. nov. International journal of systematic and evolutionary microbiology. 2009, 59, 3155–3160.
- Riedel J, Halm U, Prause C, Vollrath F, Friedrich N, Weidel A, et al. Multilocular hepatic masses due to Enterobius vermicularis. Innere Medizin (Heidelberg, Germany). 2023.
- Wardhana, DK. Risk factors for bacterial contamination of bovine meat during slaughter in ten Indonesian abattoirs. Veterinary medicine international. 2019, 2019. [Google Scholar]
- Bhembe NL, Jaja IF, Nwodo UU, Okoh AI, Green E. Prevalence of tuberculous lymphadenitis in slaughtered cattle in Eastern Cape, South Africa. International Journal of Infectious Diseases. 2017, 61, 27–37.
- Nagaraja T, Lechtenberg KF. Liver abscesses in feedlot cattle. Veterinary Clinics of North America: Food Animal Practice. 2007, 23, 351–369.
- Jaja IF, Mushonga B, Green E, Muchenje V. Factors responsible for the post-slaughter loss of carcass and offal’s in abattoirs in South Africa. Acta tropica. 2018, 178, 303–310.
- Bosman, P. Scheme for the control and eventual eradication of bovine brucellosis (author's transl). Journal of the South African Veterinary Association. 1980, 51, 75–79. [Google Scholar] [PubMed]
- Matle I, Ledwaba B, Madiba K, Makhado L, Jambwa K, Ntushelo N. Characterisation of Brucella species and biovars in South Africa between 2008 and 2018 using laboratory diagnostic data. Veterinary Medicine and Science. 2021, 7, 1245–1253.
- Kolo FB, Adesiyun AA, Fasina FO, Katsande CT, Dogonyaro BB, Potts A, et al. Seroprevalence and characterization of Brucella species in cattle slaughtered at Gauteng abattoirs, South Africa. Veterinary Medicine and Science. 2019, 5, 545–555.
- Govindasamy, K. Human brucellosis in South Africa: A review for medical practitioners. South African Medical Journal. 2020, 110, 646–651. [Google Scholar] [PubMed]
- De Massis F, Zilli K, Di Donato G, Nuvoloni R, Pelini S, Sacchini L, et al. Distribution of Brucella field strains isolated from livestock, wildlife populations, and humans in Italy from 2007 to 2015. PloS one. 2019, 14, e0213689.
- Frean J, Cloete A, Rossouw J, Blumberg L.. Brucellosis in South Africa–A notifiable medical condition. NICD Communicable Diseases Communique. 2018, 16, 110–117.
- Caine L-A, Nwodo UU, Okoh AI, Green E. Molecular characterization of Brucella species in cattle, sheep and goats obtained from selected municipalities in the Eastern Cape, South Africa. Asian Pacific Journal of Tropical Disease. 2017, 7, 293–298.
- Godfroid J, Al Dahouk S, Pappas G, Roth F, Matope G, Muma J, et al. A “One Health” surveillance and control of brucellosis in developing countries: moving away from improvisation. Comparative immunology, microbiology and infectious diseases. 2013, 36, 241–248.
- Health WOfA. Manual of diagnostic tests and vaccines for terrestrial animals. Infect Bursal Dis. 2012;12:549-65.
- Zhang N, Huang D, Wu W, Liu J, Liang F, Zhou B, et al. Animal brucellosis control or eradication programs worldwide: a systematic review of experiences and lessons learned. Preventive veterinary medicine. 2018, 160, 105–115.
Figure 1.
Collection of samples during slaughtering process workflow and inspection in this study. A & B: Livestock in holding pens at the abattoirs. C. Processing of approximately 100 g of each tissue D. Atypical cattle lymph node with visible lesions. E. Cyst/abscess on a liver. F. Bruised carcass post slaughter.
Figure 1.
Collection of samples during slaughtering process workflow and inspection in this study. A & B: Livestock in holding pens at the abattoirs. C. Processing of approximately 100 g of each tissue D. Atypical cattle lymph node with visible lesions. E. Cyst/abscess on a liver. F. Bruised carcass post slaughter.
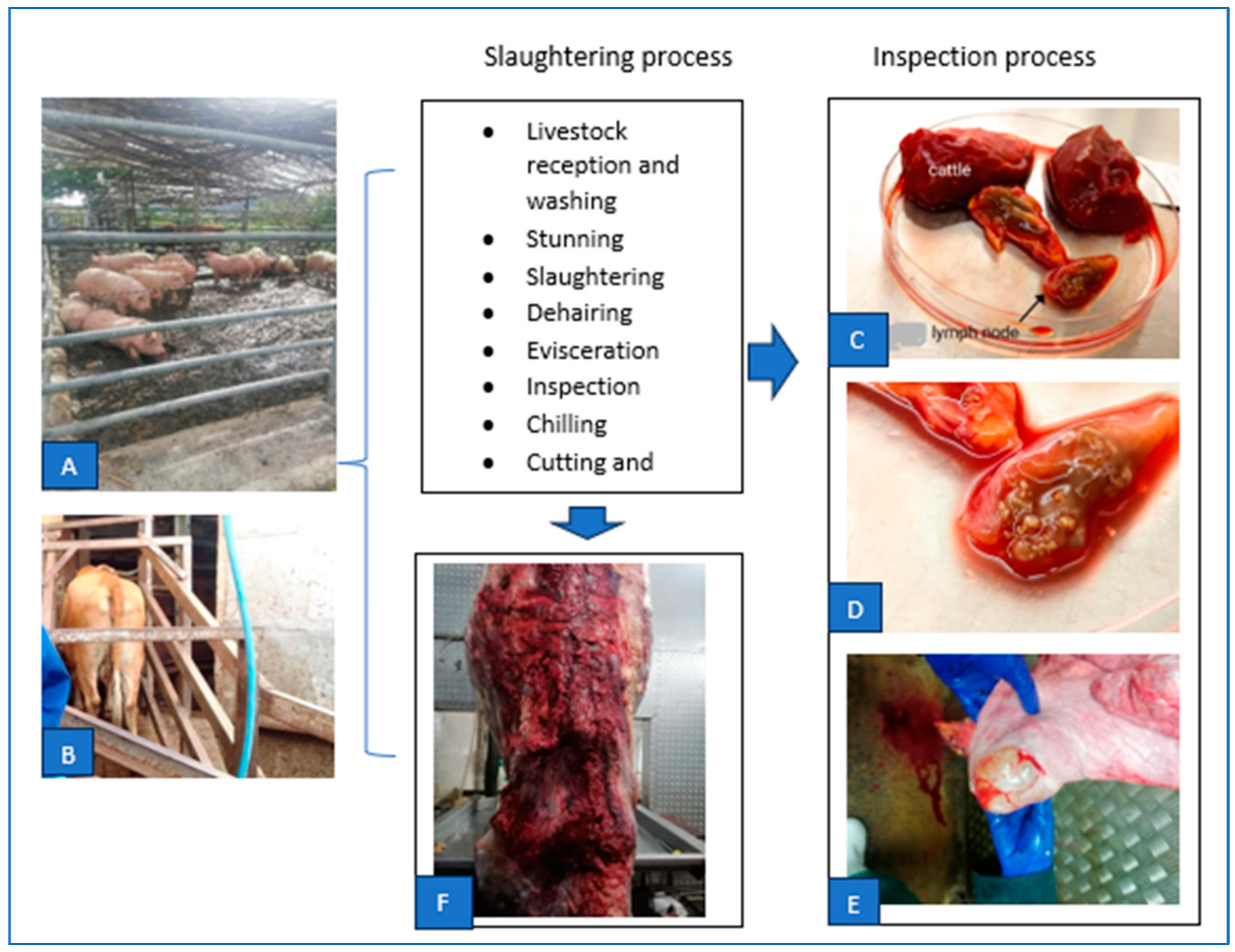
Figure 3.
Sequencing identification of other bacterial organisms present in cultures from abattoir livestock tissues on modified CITA medium.
Figure 3.
Sequencing identification of other bacterial organisms present in cultures from abattoir livestock tissues on modified CITA medium.
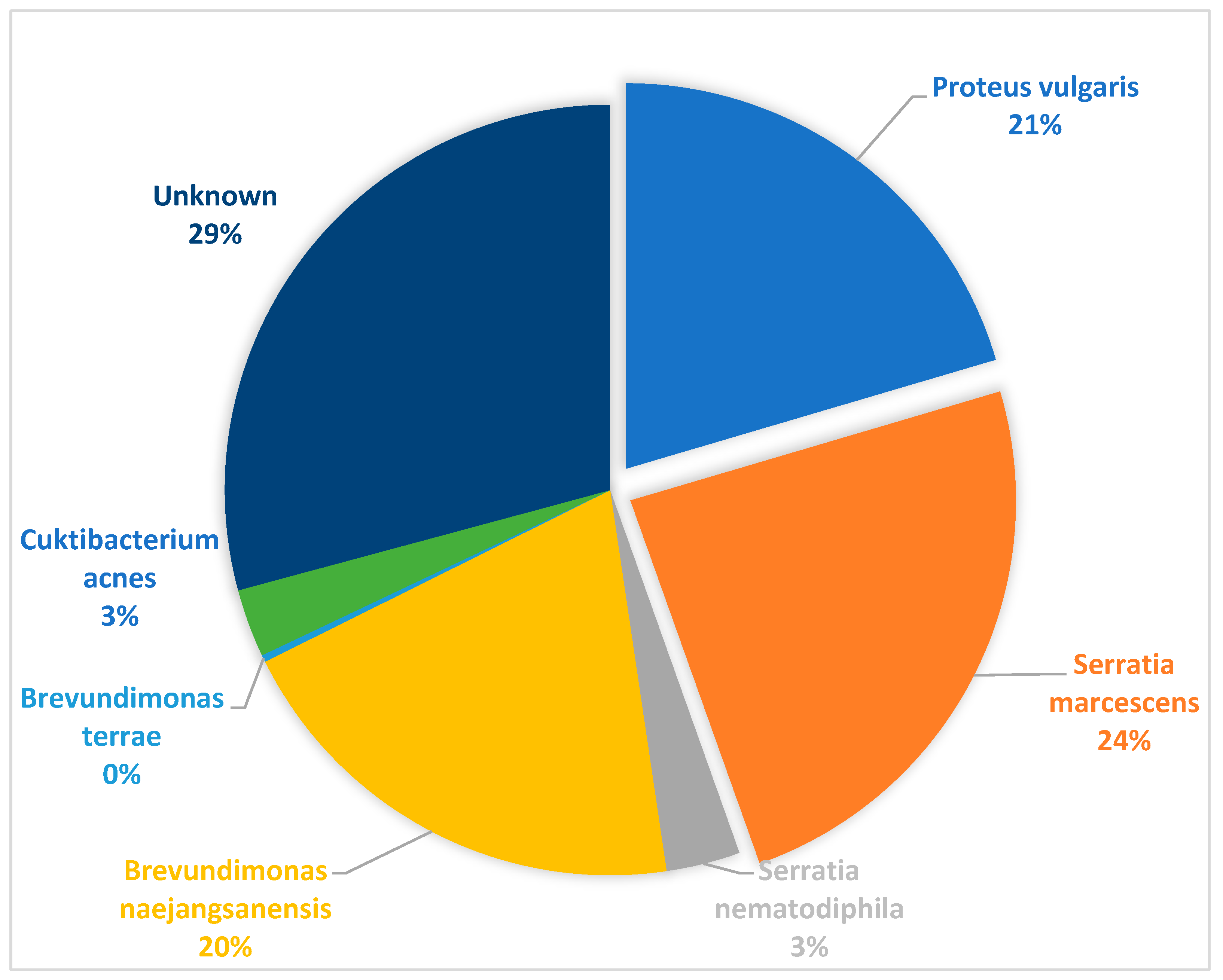

Table 1.
Brucellosis characterization of slaughtered livestock using 16-26S ribosomal interspacer region (ITS)-PCR, Brucella isolation identified with AMOS-PCR stratified by tissue and serological information using Rose Bengal test (RBT), complement fixation test (CFT) and iELISA (latter from [41].
Table 1.
Brucellosis characterization of slaughtered livestock using 16-26S ribosomal interspacer region (ITS)-PCR, Brucella isolation identified with AMOS-PCR stratified by tissue and serological information using Rose Bengal test (RBT), complement fixation test (CFT) and iELISA (latter from [41].
| Species | ITS-PCR positive animals (%) | Culture AMOS-PCR animals (%) |
Culture positive animals identified with AMOS-PCR from ITS-PCR positive tissue (%) |
Number positive tissues per animal species |
Sero-negative (RBT, CFT & iELISA) and culture positive animals |
Brucella culture and sero-positive animals | |||||||
| Liver | Spleen | Kidney | Lymph nodes | Tonsils | RBT | ELISA | RBT and iELISA | RBT, iELISA & CFT | |||||
| Cattle | 94/280 (33.6%) | 41/280 (14.6%) |
41/94 (43.6%) |
25/94 (26.6%) |
20/94 (21.3%) |
19/94 (20.2%) |
31/94 (33.0%) |
10/94 (10.6%) |
31/41 (76.6%) |
7/41 (17.1%) |
4/41 (9.8%) |
2/41 (4.9%) |
1/41 (2.4%) |
| Sheep | 29/200 (14.5%) |
15/200 (7.5%) |
15/29 (51.7%) |
11/29 (37.9%) |
10/29 (34.5%) |
11/29 (37.9%) |
8/29 (25.6%) |
- |
13/15 (86.7%) |
2/15 (13.3%) |
0/15 | 0/15 | 0/15 |
| Pigs | 4/85 (4.7%) |
2/85 (2.4%) |
2/4 (50.0%) |
0/4 | 0/4 | 0/4 | 0/4 | 2/4 50.0%) |
2/2 (100%) |
0/15 | 0/15 | 0/15 | 0/15 |
Table 2.
Descriptive and univariate analyses to determine the association between various factors and occurrence of Brucella spp. occurrence in the tissue was determined using ITS-PCR.
Table 2.
Descriptive and univariate analyses to determine the association between various factors and occurrence of Brucella spp. occurrence in the tissue was determined using ITS-PCR.
| Variable | Level | Number of animals positive for Brucella spp. (%) | p-value |
| Abattoir | <0.0001 | ||
| Abattoir A (n=50) | 19 (38.0) | ||
| Abattoir B (n=344) | 48 (14.2) | ||
| Abattoir C (n=45) | 7 (15.6) | ||
| Abattoir D (n=23) | 9 (39.1) | ||
| Abattoir E (n=103) | 43 (41.7) | ||
| Throughput | 0.05078 | ||
| High (n=542) | 118 (21.8) | ||
| Low (n=23) | 9 (39.1) | ||
| Animal species | <0.0001 | ||
| Cattle (n=280) | 94 (33.6) | ||
| Pig (n=85) | 4 (4.7) | ||
| Sheep (n=200) | 29 (14.5) | ||
| Sex | Female (n=276) | 66 (23.9) | 0.4245 |
| Male (n=289) | 61 (21.1) |
Table 3.
Multivariable analysis.
| Variable | Category | Odds ratio (CI) | p-value |
| Abattoir | Abattoir B (ref) | ||
| Abattoir A | 4.89 (2.26, 10.57) | <0.0001 | |
| Abattoir C | 0.91 (0.36, 2.30) | 0.8495 | |
| Abattoir D | 7.02 (2.57, 19.15) | 0.000142 | |
| Abattoir E | 5.13 (2.92, 8.99) | <0.0001 | |
| Species | |||
| Pig (ref) | |||
| Cattle | 17.09 (5.66, 51.61) | <0.0001 | |
| Sheep | 5.59 (1.71,18.29) | 0.0043 | |
| Sex | |||
| Male (ref) | |||
| Female | 0.54 (0.33, 0.89) | 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



