
Submitted:
09 November 2023
Posted:
14 November 2023
You are already at the latest version
Abstract
West Nile virus (WNV) is an arbovirus mainly transmitted by Culex spp., which causes a worldwide zoonotic disease. This pathogen is endemically maintained in a life cycle with birds acting as reservoirs and humans and horses as accidental and dead-end hosts. Sporadic WNV outbreaks have been reported in Europe and the potential impact of WNV infection on populations of threatened or endangered birds of prey is considerable. Surveillance programs are needed for early detection of this virus. All four species of vultures present in Europe are considered protected species. As scavengers, vultures are at the top of the food chain, and can be susceptible to and negatively affected by pathogens like WNV. In a conservation perspective, the impact of WNV in European vultures, alone or concomitantly with other factors, should be addressed. This review of documented cases can be considered a state point.
Keywords:
epidemiology
; Europe
; scavengers
; vultures
; West Nile virus
; zoonotic
1. Introduction
Vultures are the biggest European bird scavengers, performing an essential role in the environment. Information and awareness-raising are needed to change the paradigm of the bad reputation they are given. Vultures provide an important service to the ecosystem by cleaning up and recycling carcasses of dead animals. They can quickly eliminate large amounts of flesh, in different stages of decomposition, that may potentially host pathogenic microorganisms, removing this potential source on infection from the environment. Furthermore, thanks to its extremely acidic gastric pH and stable intestinal microbiome with remarkable antimicrobial activity, can neutralize pathogens that pass through their gastrointestinal tract, limiting the spread of diseases [1,2,3,4]. These majestic group of raptors also provide great socioeconomic value to local communities. Birdwatching of scavenging birds is yielding economic benefits to rural economies within developing regions [5].
Four species of vulture are found in Europe: bearded vulture (Gypaetus barbatus), cinereous vulture (Aegypius monachus), griffon vulture (Gyps fulvus) and Egyptian vulture (Neophron percnopterus). Two-hundred years ago, these species of vultures were among the most common breeding bird species in the mountains of central and southern Europe. Yet, the decreasing availability of food, coupled with habitat loss, persecution, and poisoning, made vultures disappear from most of their European range, with populations considerably smaller and increasingly isolated by the 1960s. Many conservation efforts have been and are being made, so European vulture populations are steadily recovering.
In 2007 Egyptian vulture was declared globally ‘Endangered’ by the International Union for the Conservation of Nature (IUCN) Red List [6]. Most of its European population is found on the Iberian Peninsula with an estimated 1,300–1,500 pairs. This species is a long-distance migratory bird of prey, spending its winters in sub-Saharan Africa and returning from March to reproduce in Europe (Figure 1). It faces many threats, highlighting electrocution and poisoning.
The bearded vulture’s population crashed at the beginning of the 20th century and was driven to extinction throughout most of its former range. Nowadays, it is the rarest vulture in Europe, and its presence is limited to the Alps, Pyrenees, and Andalusia, with isolated populations in Crete and Corsica islands (Figure 2). It is considered ‘Near Threatened’ by IUCN Red List [6]. This vulture has a unique feeding habit: its diet consists of a huge percentage of bleached carcass bones [7]. In the wild, they rub themselves with ferric oxides, which turns their plumage rusty orange. Major threats are changes in farming practices and direct persecution [8,9,10].
Griffon vultures are the most social of European vultures, with gregarious and competitive habits, breeding in large colonies. They feed in groups and can penetrate animal carcasses to feed on the softer tissues, such as muscles and viscera [11,12]. Following a decline in the 20th century, as a result of wildlife poisoning, hunting and decreasing of food supplies [13], in recent years the species has significantly increased in some areas, particularly in France and in the Iberian Peninsula (Figure 3), and has an extremely large range in Europe, being now considered as ‘Least concern’ by IUCN [6,11].
The cinereous vulture is the biggest raptor in Europe, with a wingspan almost reaching 3 meters. Its conservation status is listed as ‘Near Threatened’ by IUCN’s red list and as ‘Critically Endangered’ in Portugal [6,14]. Over the last century, the population of this species has decreased across its distribution area in Europe, and it is now extinct in many European countries (namely in Italy, Austria, Poland, Slovakia, and Romania) (Figure 4) [6]. Major threats to the viability of the species include habitat destruction by increasing forest fires, illegal use of poisons, limited food availability due to health restrictions and reduced wild herbivore populations, consumption of food resources contaminated by veterinary drugs or lead (from hunting ammunition), human disturbance during the breeding season and death by collision with/electrocution on power lines [14].
The most frequently reported cause of free-living vulture death and disease worldwide (Table 1) was due to toxic agents, with special emphasis to lead and pesticides. Regarding traumatic causes of morbidity and mortality, the second highest prevalence, the most reported are of anthropogenic origin: collision with and electrocution by power lines, gunshot, direct persecution, among others [15]. It is not uncommon that the main cause of vulture admission at wildlife rehabilitation centers and the cause of subsequent death, when it occurs, remains unknown [16].
Vultures may be considered resistant to certain microorganisms, such as Bacillus anthracis [17] or Clostridium botulinum, due to the fact of having naturally-occurring antibodies against their toxins [18]; however, on the other hand, vultures can be susceptible to and negatively affected by other pathogens. Infectious disease effects on their hosts have not been thoroughly investigated for most vulture species, obligate avian scavengers. Some pathogens should be seen as a potential threat to vulture conservation because they can cause disease in individual birds, and potentially jeopardize vultures’ health when associated with other menaces, such as contaminants and intentional poisoning [5].
2. West Nile Virus
West Nile virus (WNV), lying within the genus Flavivirus and family Flaviviridae, is the most widespread arbovirus [19,20]. The cycle of WNV is maintained between birds (as reservoirs) and mainly mosquitos of the genus Culex (as vectors). When infected mosquitoes bite mammals, spread occurs, and horses and humans can be accidental dead-end hosts, along with other mammals [21,22]. WNV has recently emerged as a major public health concern in Europe. The infection numbers recently raised in European countries, while some remarkable socio-economic and environmental changes were noticed, including an economic crisis (in part due to the COVID-19 pandemic) and very high temperatures registered [23,24].
WNV strains have been grouped into nine genetic lineages, by phylogenetic analysis, and lineage 1 and 2 are the most important [25,26]. Both lineages 1 and 2 have been shown to be able to cause severe disease in birds, horses and humans [27,28,29].
In humans, the severity of infections can range from asymptomatic to serious or fatal hemorrhagic fever or neurological disease. Most people (around 80%) with WNV infections remain asymptomatic. Of those who develop symptoms and clinical signs, the most common presentation is febrile illness (fever) accompanied by an acute syndrome, and less than 1% develop encephalitis or other forms of neuroinvasive disease [30,31,32].
As the key vertebrate hosts in WNV transmission cycle, avian species are the focus of surveillance worldwide [33]. The information on the role of the birds of prey as reservoirs, spreaders, or sentinels of WNV is not yet clearly described, and specially the vulture species influence on the epidemiology of WNV remains scarce [34,35].
For birds of prey, the pathogen is usually detected in the carcasses of birds found dead or moribund through wildlife surveillance programs or raptors admitted to wildlife rehabilitation centers [36]. Three modes of WNV infection may be considered: mosquito-borne transmission, contact transmission and oral transmission [34]. Variations in the prevalence of WNV infection between different species and populations of birds of prey can be explained by differences in exposure, based on infection prevalences of prey species, differences in roosting habitat and exposure to ornithophilic mosquito [37,38,39].
WNV infection in raptors appears to act like a multi-systemic disease, affecting different organs depending on the host species and also individuals. The disease can be fatal in most raptors, with an acute onset of illness a few days after infection occurs [40]. On the other hand, the infection can also originate a debilitating and chronic disease, predisposing the affected animals to concurrent pathological conditions. In these cases, unspecific signs often occur, like depression, dehydration and severe weight loss [39,41,42]. WNV is neurotropic and, consequently, neurological signs are a very common outcome of infection, such as alterations in the mental status, head tilt, tremors, leg paresis or paralysis, among others. Ocular disorders often occur as well. [27,43,44,45]. The most common abnormalities in haematology screening are anaemia and leukocytosis [46], accompanied by splenomegaly, which can be seen in the ventrodorsal and laterolateral radiographic projections [47]. Long-term sequelae have been detected in raptors, among which reccurrence of neurologic signs, feather pulp abnormalities and abnormal molt can persist over the long term, and may have a negative impact on the longevity of these species [35,40].
WNV infections can be diagnosed by isolating the virus, detecting viral antigens or RNA, or using serological methods. Research on avian immunity has focused on humoral immunity and there is yet a lack of information on cellular immunity. Humoral immunity against WNV is determined by the presence of antibodies in the blood of animals. This can be measured by a range of serological assays, being the virus neutralization test (VNT) considered the gold standard in flaviviruses serology, which detects serum neutralizing antibodies and more accurately detects protective antibodies, being more specific than other serological techniques [34]. The main alternative is the capture enzyme-linked immunoassay (cELISA) that detects antibodies directed against the virus envelope protein. There are class-specific ELISAs for the detection of immunoglobulin (Ig) Y (the avian equivalent of IgG in mammals) or IgM against WNV. In adult birds, antibodies are developed following exposure and infection with WNV [34,48]. In juveniles, presence of maternal antibodies can be detected [49]. The long-term stability of antibodies confirmed in several species of raptors suggests that humoral immunity to WNV may be long-lasting in most individuals that survive infection [50]. That is why positive serological results from ELISA suggest that a bird has been exposed to WNV, but generally do not indicate when the infection occurred [51]. It is highly suggested to use VNTs to confirm WNV in all positive and doubtful samples detected by ELISA, in order to increase the accuracy of estimated seroprevalences in wild birds. The use of VNT will be especially important in areas where co-circulation of related flaviviruses exists [48].
The molecular diagnosis of WNV infection can be made using specific RT-PCR. Fresh brain samples, cloacal, and choanal swabs can be tested for this virus, as well as other organs where the virus is commonly found in raptors (heart, spleen, liver, kidney, adrenal gland and intestine. Immunohistochemistry (IHC)also can be useful to detect WNV antigens in affected tissues from live or dead birds [42,45,52,53].
There is yet no specific treatment for WNV infection in raptors. In spite of the supportive treatment and anti-inflammatory drugs, to minimize the effects caused by infection, the majority of affected birds end up dying [54,55]. There are still no vaccines specifically available for use in birds [34,41,43] or humans [22].
2.1. West Nile Virus in vultures
We have reviewed the scientific literature retrieved from PubMed and ScienceDirect databases, using general searches with the following terms: ‘West Nile Virus’ AND ‘Vultures’. We performed other searches using different combinations of the following relevant terms: ‘Scaveng* bird’ AND ‘flaviviruses’. We filtered our search based on the geographical location where the studies were performed, focusing on Europe and European species of vultures (Table 1). Eight reports were found that reported WNV infections in European vulture species, in Russia, Serbia, Austria and Spain, plus one in Iran.
Between 2008 and 2009 outbreaks of WNV infection occurred in central Europe, with an unexpected spread of a lineage 2 WNV strain. In 2011, a study from Austria described a WNV outbreak in birds of prey in 2008 and 2009 in the eastern part of the country. Samples were collected from birds nearby the location of the first detected infections and from poultry houses aviaries at the Research Institute of Wildlife Ecology in Vienna. ELISA positive results were found in 10 bearded vultures (G. barbatus) among other species of birds. 66% of the positive ELISA reactions could be confirmed with VNT, including two out of three vultures tested [56]. One wild bearded vulture was confirmed by RT-PCR and IHC as positive for WNV, also in Austria [57].
In 2011 the first report of flaviviruses circulation in Egyptian vulture (N. percnopterus) was published from Spain. Between 2006 and 2009 serum samples were obtained from several bird species in Andalusia, southern Spain, among free-ranging and captive animals held in rehabilitation centres. Two cinereous vultures (A. monachus) and two Egyptian vultures tested positive on cELISA for WNV. As the used ELISA kit has been designed to detect antibodies directed against the envelope protein (pr-E) of WNV, which contains an epitope common to Japanese encephalitis virus (JEV) antigen, the VNT was further performed as a confirmation and more specific test for the positive samples on ELISA [58].
In 2017, WNV was detected in one bearded vulture (G. barbatus) in Lleida, northeastern Spain. The animal had been admitted at a local wildlife rehabilitation centre, and evidenced neurological signs. Serum samples were analysed and the results were positive on cELISA and negative on serum neutralization test (SNT). One month later antibodies against WNV were detected on SNT. Thirteen additional vultures of the same species tested positive by cELISA, from which two were confirmed by SNT (titers between 1/20 and 1/40) [59].
Among samples collected in 2017-2018, two Egyptian vultures published (N. percnopterus) tested positive with micro-VNT, after a positive result on ELISA in Iran [60]. This species is estival migratory in the European continent, and migrates to the African Sahelian region in the winter [61]. Birds from western Europe usually winter in western Africa, and birds from eastern Europe winter in eastern Africa or the Arabian Peninsula [62]. They are the same birds that return to Europe, a reason for mentioning this study, despite the fact of being based on another continent.
From October 2017 to December 2019, almost 400 wild birds were sampled in Cáceres and Badajoz provinces, western Spain, in collaboration with two local rehabilitation centres. Analysis by RT-PCR in sick birds confirmed the presence of WNV lineage 1 RNA in a griffon vulture (G. fulvus) and specific antibodies against WNV in juvenile birds were detected in specimens of griffon vulture and cinereous vulture [63].
In the period of 2018 and 2022 in Serbia, 25 dead griffon vultures from the wild were submitted to macroscopical and histopathological examination, followed by different diagnostic screening. Infectious agents were detected in 4/25 animals, confirming a West Nile virus infection with lymphoplasmacytic encephalitis by PCR [64].
WNV emerged in the United States of America (USA) in 1999 [65] and has since spread throughout much of the American continent. WNV was identified as the primary cause of death in one nestling wild California condor (Gymnogyps californianus). The infection was diagnosed by IHC on heart in conjunction with compatible lesions. In the USA, WNV infection is an emerging mortality factor for young, wild-hatched birds and vaccination is required at nest sites where WNV is prevalent [66]. Other New World vulture species such as the black vulture (Coragyps atratus) and Turkey vulture (Cathartes aura) were confirmed to WNV infection [46,50,67], as well as the Lappet-faced vulture (Torgos tracheliotus), an Old-World vulture not present in Europe [55].

Table 2.
Diagnosed cases of WNV infection in European vultures.
| Species | WNV diagnosis | Geographical area | Year | Study | |
|---|---|---|---|---|---|
|
Gypaetus barbatus |
ELISA; VNT |
Austria |
NM |
[56] |
|
|
Gypaetus barbatus |
RT-PCR |
Austria |
2008 |
[57] |
|
|
Aegypius monachus Neophron percnopterus |
ELISA | Spain | 2006-2009 | [58] | |
|
Gypaetus barbatus |
ELISA; SNT |
Spain |
2017 |
[59] |
|
| Neophron percnopterus | ELISA; MNT | Iran | 2017-2018 | [60] | |
|
Aegypius monachus Gyps fulvus |
ELISA; VNT; RT-PCR |
Spain | 2017-2019 | [63] | |
|
Gyps fulvus |
RT-PCR | Serbia | 2018-2022 | [64] |
ELISA – enzyme-linked immunosorbent assay; MNT – microneutralization test; NM – not mentioned; RT-PCR – reverse transcription polymerase chain reaction; SNT – serum neutralization test VNT – virus neutralization test.
3. Discussion
Zoonotic emerging infectious diseases represent a raising and important threat to public health, and vector-borne diseases were responsible for almost a quarter of the documented emerging infectious diseases events in the first decade of the 21st century [68].
West Nile virus (WNV) was first reported in Uganda in 1937 [69] and was subsequently isolated from patients, birds, and mosquitoes from the early 1950s [70]. The virus became endemic in several parts of the African continent [71] and the first outbreak of WNV in Europe was reported in 1962 in the French Camargue region [72]. Since 1996, after a high rate of confirmed cases in Eastern Europe (Romania) [73] several outbreaks have been often reported in the European continent, with a noticeable seasonal pattern, during the warmer weather (July to October) [74]. Birds of prey are especially susceptible to WNV [34]. Upon the species and individuals, WNV infection can cause acute death, a fatal outcome several weeks after infection, or birds can eventually survive to chronic infections [40,54,55]. Either in Europe or North America, raptors are among the birds described as more frequently infected during WNV outbreaks [47,57]. Identifying target species is important for developing an efficient surveillance and monitoring program, a reason why targeting specific raptor species as disease sentinels may be beneficial.
The role of Palearctic-African migratory birds in the introduction of WNV to Europe has long been the subject of debate [19,20,75]. The suspicion that migratory birds could be the main introductory hosts of WNV in new regions was based on the following: the majority of outbreaks in temperate regions occurring during late summer or early fall, when migratory birds and mosquitoes coexist on a large scale [70,74,76]; the principal vectors from which the virus has been isolated are mainly ornithophilic mosquitoes [77,78]; circulating antibodies against WNV have been found in many migratory bird species, and long-distance migrants in Europe (in particular, species wintering south of the Sahara) are exposed during their migratory journeys and/or their winter stay in Africa to higher levels of WNV circulation (or a closely antigenically related Flavivirus), when compared with the levels found in their breeding grounds in [79] and, finally, migratory birds have been linked with transporting related viruses in the Western Hemisphere [80]. It is difficult to define a population geographically, especially for migratory birds WNV has been isolated from some actively migrating species of birds (e.g., the White stork (Ciconia ciconia) [81] and the Turtle dove (Streptopelia turtur) [76]. Migratory birds play an essential role in the long-distance movement of JEV serocomplex flaviviruses. When migrations occur between enzootic and areas free of WNV, birds that become infected prior to or during migration can actively transport the virus in their blood or tissues and infect mosquitoes and/or their predators [79].West Nile virus (WNV) poses a threat to endangered species around the world, such as Iberian imperial eagle (Aquila adalberti) [82]. Egyptian vultures (N. percnopterus) is a long-distance migratory species, as mentioned above [6], and therefore may be even more susceptible to contracting a WNV infection.
A successfully introduction of WNV in destination territories depends on the conditions for local maintenance, such as ecological key factors which promotes the virus maintenance and transmission, like presence and abundance of competent birds (hosts) and mosquitoes (vectors), and favorable abiotic conditions [83,84]. Temperature is the most frequently used environmental condition when it refers to WNV and/or its vectors, followed by precipitation [84]. Mediterranean savannahs, in Spanish known as ‘dehesas’ and in Portuguese ‘montados’, are found in regions with mild, rainy winters and very hot, dry summers occupied by an agro-silvopastoral landscape in the Southwest of the Iberian Peninsula. These areas are threatened by climate change and the abandonment of traditional uses, and are therefore protected under the European Habitats Directive [85,86]. Foraging griffon vultures from different populations located across Western Europe made long trips into these regions of the Iberian Peninsula, suggesting that these areas have a beneficial added value for griffon vultures and other avian species, also providing the main habitat for wintering bird species that come from Northerly latitudes [86,87]. According to Casades-Martí [83], this continental Mediterranean territory, with wildlife–livestock interaction is favorable to the circulation on WNV and other flaviviruses. Interactions between wild birds and farm animals are more likely to occur here, resulting in a higher probability of exposure to flaviviruses [88]. Besides mosquitoes are considered the main vectors of WNV, it has occasionally been isolated from other hematophagous arthropods, namely argasid and amblyommine ticks, [78]. Like other soaring bird species, griffon vultures perform long-scale movements [87,89]and are considered a partial migratory species, with juveniles (especially in the first year of live) crossing the Strait of Gibraltar to Africa during fall and returning to Europe in the following years [90].
The abundance of vectors is a relevant parameter for pathogen transmission. Those habitats more suitable to the expansion of Culex mosquitoes during peak times of WNV transmission represent the highest risk for the potential spillover of WNV into human populations [91].
In spite of some ticks (soft ticks) can be considered resident and sedentary [92], that does not mean they have a limited role in the circulation of infectious agents. Pathogens transmitted by these ticks can be spread into new areas taking advantage of the large foraging movements or migration of its hosts, namely griffon vultures [87,89,90]. Although ticks can be alternative vectors, a recent study in the Pyrenees (Northeastern Spain), flavivirus was detected in all seven vulture’s blood samples by the generic qRT-PCR, but all analyzed ticks were negative for flavivirus detection, which reinforces the potential involvement of other more common arthropod vectors, like mosquitoes, in the transmission of the virus [93,94].
Scavengers are susceptible to be infected by consuming WNV-infected carcasses [39,95]. WNV activity is far more likely to be detected in urban areas than in rural areas, suggesting that human density and associated factors should be considered when interpreting dead wild bird surveillance for WNV [95]. Long-lived avian scavengers are affected by the habitat where they live, being anthropized landscapes considered a more stressful habitat. Birds that live there generally present poor body condition and are more vulnerable to disease. Foraging in more anthropized areas can bring advantages and disadvantages in terms of energy balance and stress. In these territories, availability and predictability of food is likely higher, but data suggest, on the other hand, that the food quality is not so good, leading to a poor nutritional status [96], which contributes to higher levels of circulating glucocorticoids. Concurrent factors that lead to lower immunity must be taken in consideration, such as the risk of lead accumulation, ingested from hunting ammunition, or severe competition in a high density and challenging environment, especially in highly social species, like griffon vultures (G. fulvus), or less capable species, for example Egyptian vultures (N. percnopterus) [97]. It is known that WNV has killed many thousands of birds around the world, but it is difficult to measure the long-term impact of the disease on wildlife populations.
Deforestation, besides being a major cause of biodiversity loss by itself and causing a negative impact on human health [98,99], is linked to the emergence of zoonotic and vector-borne diseases, like WNV. Deforestation can increase contact between vectors and avian reservoirs. Forest loss may facilitate exchanges between human and zoonotic cycles as open areas favor the human presence and settlement [98]. The relationship between deforestation and the occurrence of zoonotic outbreaks has already been suggested [74,99], and should be further investigated for WNV.
4. Conclusion
There are relatively few reports of infectious diseases as a direct and primary cause of mortality in avian scavengers, and it has been a neglected topic of research so far. Given the current decline in scavenging bird populations, baseline information on exposure to infectious agents will be helpful for monitoring population health and investigating future disease-related epidemics, helping to guide conservation efforts. Besides, early detection of pathogens, like WNV, will allow the establishment of effective measures to prevent or mitigate the effect of the virus on human populations, as well as to protect susceptible endangered species. Mediterranean ‘montados’ hold a very important fraction of the European populations of some endangered avian scavengers such as cinereous and Egyptian vultures, as well as other endemic globally endangered top predators. The implementation of a collaborative international, holistic and multi-disciplinary One Health action is crucial to allow a more accurate risk assessment and an early response to West Nile virus and other emerging zoonotic pathogens. The presence of WNV in vultures may therefore have public health and wildlife conservation implications and deserves further investigation.
Author Contributions
Conceptualization, F.L. and A.C.; methodology, F.L. and A.C.; writing—original draft preparation, F.L.; writing—review and editing, F.L., L.C., A.M., M.M. and A.C.; All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Houston, D.C.; Cooper, J.E. The digestive tract of the whiteback griffon vulture and its role in disease transmission among wild ungulates. J. Wildl. Dis. 1975, 11, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Moleón, M.; Sánchez-Zapata, J.A.; Margalida, A.; Carrete, M.; Owen-Smith, N.; Donázar, J.A. Humans and scavengers: The evolution of interactions and ecosystem services, BioScience. 2014, 64, 394–403. [CrossRef]
- Blumstein, D.T.; Rangchi, T.N.; Briggs, T.; De Andrade, F.S.; Natterson-Horowitz, B. A systematic review of carrion eaters' Adaptations to Avoid Sickness. J. Wildl. Dis. 2017, 53, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Arbulu, S., Jiménez, J.J., Gútiez, L. et al. Evaluation of bacteriocinogenic activity, safety traits and biotechnological potential of fecal lactic acid bacteria (LAB), isolated from Griffon Vultures (Gyps fulvus subsp. fulvus). BMC Microbiol. 2016, 16, 228. [CrossRef]
- Plaza, P.; Blanco, G.; Lambertucci, S. Implications of bacterial, viral and mycotic microorganisms in vultures for wildlife conservation, ecosystem services and public health. Ibis 2020, 162, 1109–1124. [Google Scholar] [CrossRef]
- BirdLife International and Handbook of the Birds of the World 2021. The IUCN Red List of Threatened Species, Version 2022-2. Available online: https://www.iucnredlist.org/ (accessed on 27 September 2023).
- Hiraldo, F.; Delibes, M.; Calderón, J. El quebrantahuesos Gypaetus barbatus (L.). Sistemática, Taxonomía, Biología, Distribución y Protección. Monografías, 22. Instituto para la Conservación de la Naturaleza, Madrid, Spain. 1979.
- Angelov, I.; Hashim, I.; Oppel, S. Persistent electrocution mortality of Egyptian Vultures Neophron percnopterus over 28 years in East Africa. Bird Conserv. Int. 2013, 23, 1–6. [Google Scholar] [CrossRef]
- Thiollay, J-M. The decline of raptors in West Africa: long-term assessment and the role of protected areas. Ibis 2006, 148, 240-254. [CrossRef]
- Donázar, J.A.; Palacios, C.J.; Gangoso, L.; Ceballos, O.; González, M.J.; Hiraldo, F. Conservation status and limiting factors in the endangered population of Egyptian vulture (Neophron percnopterus) in the Canary Islands. Biol. Conserv. 2002, 107, 89–97. [Google Scholar] [CrossRef]
- Pirastru, M.; Mereu, P.; Manca, L.; Bebbere, D.; Naitana, S.; Leoni, G.G. Anthropogenic drivers leading to population decline and genetic preservation of the Eurasian griffon vulture (Gyps fulvus). Life 2021, 11, 1038. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Avizanda, A.; Jovani, R.; Carrete, M.; Donázar, J.A. Resource unpredictability promotes species diversity and coexistence in an avian scavenger guild: A field experiment. Ecology 2012, 93, 2570–2579. [Google Scholar] [CrossRef] [PubMed]
- Virani, M.Z.; Kendall, C.; Njoroge, P.; Thomsett, S. Major declines in the abundance of vultures and other scavenging raptors in and around the Masai Mara ecosystem, Kenya. Biol. Conserv. 2011, 144, 746–752. [Google Scholar] [CrossRef]
- Cabral, M.J.; Almeida, J.; Almeida, P.R.; Delliger, T.; Ferrand de Almeida, N.; Oliveira, M.E.; Palmeirim, J.M.; Queirós, A.I.; Rogado, L.; Santos-Reis M. Livro Vermelho dos Vertebrados de Portugal. Publisher: Instituto da Conservação da Natureza, Lisboa, Portugal, 2005; pp.215-216.
- Ives, A.M.; Brenn-White, M.; Buckley, J.Y.; Kendall, C.J.; Wilton, S.; Deem, S.L. A global review of causes of morbidity and mortality in free-living vultures. EcoHealth 2022, 19, 40–54. [Google Scholar] [CrossRef]
- Garcês, A.; Pires, I.; Sargo, R.; Sousa, L.; Prada, J.; Silva, F. Admission causes, morbidity, and outcomes in scavenger birds in the North of Portugal (2005–2022). Animals 2023, 13, 2093. [Google Scholar] [CrossRef] [PubMed]
- Hugh-Jones, M.E.; De Vos, V. Anthrax and wildlife. Rev Sci Tech. 2002, 21, 359–383. [Google Scholar] [CrossRef] [PubMed]
- Ohishi, I.; Sakaguchi, G.; Riemann, H.; Behymer, D.; Hurvell, B. Antibodies to Clostridium botulinum toxins in free-living birds and mammals. J Wildl Dis. 1979, 15, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Triana, L.M.; Jeffries, C.L.; Mansfield, K.L.; Carnell, G.; Fooks, A.R.; Johnson, N. Emergence of West Nile virus lineage 2 in Europe: a review on the introduction and spread of a mosquito-borne disease. Front Public Health. 2014, 2, 271. [Google Scholar] [CrossRef] [PubMed]
- May, F.J.; Davis, C.T.; Tesh, R.B.; Barrett, A.D. Phylogeography of West Nile virus: from the cradle of evolution in Africa to Eurasia, Australia, and the Americas. J Virol. 2011, 85, 2964–2974. [Google Scholar] [CrossRef] [PubMed]
- García-Carrasco, J.M.; Muñoz, A.R.; Olivero, J.; Segura, M.; Real, R. Mapping the risk for West Nile virus transmission, Africa. Emerg Infect Dis. 2022, 28, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Habarugira, G.; Suen, W.W.; Hobson-Peters, J.; Hall, R.A.; Bielefeldt-Ohmann, H. West Nile virus: an update on pathobiology, epidemiology, diagnostics, control and “One Health” implications. Pathogens 2020, 9, 589. [Google Scholar] [CrossRef]
- Watts, M.J.; Monteys, V.S.I.; Mortyn, P.G.; Kotsila, P. The rise of West Nile virus in Southern and Southeastern Europe: A spatial-temporal analysis investigating the combined effects of climate, land use and economic changes. One Health. 2021, 13, 100315. [Google Scholar] [CrossRef]
- Young, J.J.; Haussig, J.M.; Aberle, S.W.; Pervanidou, D.; Riccardo, F.; Sekulić, N.; Bakonyi, T.; Gossner, C.M. Epidemiology of human West Nile virus infections in the European Union and European Union enlargement countries, 2010 to 2018. Euro Surveill. 2021, 26, 2001095. [Google Scholar] [CrossRef]
- Pachler, K.; Lebl, K.; Berer, D.; Rudolf, I.; Hubalek, Z.; Nowotny, N. Putative new West Nile virus lineage in Uranotaenia unguiculata mosquitoes, Austria, 2013. Emerg Infect Dis. 2014, 20, 2119–2122. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Ebel, G.D.; Deubel, V.; Kerst, A.J.; Murri, S.; Meyer, R.; Bowen, M.; McKinney, N.; Morrill, W.E.; Crabtree, M.B.; Kramer, L.D.; Roehrig, J.T. Complete genome sequences and phylogenetic analysis of West Nile virus strains isolated from the United States, Europe, and the Middle East. Virology. 2002, 298, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Gamino, V.; Höfle, U. Pathology and tissue tropism of natural West Nile virus infection in birds: a review. Vet Res. 2013, 44, 39. [Google Scholar] [CrossRef] [PubMed]
- Savini, G.; Capelli, G.; Monaco, F.; Polci, A.; Russo, F.; Di Gennaro, A.; Marini, V.; Teodori, L.; Montarsi, F.; Pinoni, C.; Pisciella, M.; Terregino, C.; Marangon, S.; Capua, I.; Lelli, R. Evidence of West Nile virus lineage 2 circulation in Northern Italy. Vet Microbiol. 2012, 158, 267–73. [Google Scholar] [CrossRef] [PubMed]
- Valiakos, G.; Touloudi, A.; Iacovakis, C.; Athanasiou, L.; Birtsas, P.; Spyrou, V.; Billinis, C. Molecular detection and phylogenetic analysis of West Nile virus lineage 2 in sedentary wild birds (Eurasian magpie), Greece, 2010. Euro Surveill. 2011, 16, 19862. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J. Clinical manifestations and outcomes of West Nile virus infection. Viruses. 2014, 6, 606–623. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Diamond, M.S. Pathogenesis of West Nile Virus infection: a balance between virulence, innate and adaptive immunity, and viral evasion. J Virol. 2006, 80, 9349–9360. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile virus. Lancet Infect Dis. 2002, 2. 519-529. [CrossRef]
- van der Meulen, K.M.; Pensaert, M.B.; Nauwynck, H.J. West Nile virus in the vertebrate world. Arch Virol. 2005, 150, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Vidaña, B.; Busquets, N.; Napp, S.; Pérez-Ramírez, E.; Jiménez-Clavero, M.Á.; Johnson, N. The role of birds of prey in West Nile virus epidemiology. Vaccines (Basel). 2020, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Kratz, G.E.; Bates, R.; Scherpelz, J.A.; Bowen, R.A.; Komar, N. Clinical evaluation and outcomes of naturally acquired West Nile virus infection in raptors. J Zoo Wildl Med. 2009, 40, 51–63. [Google Scholar] [CrossRef]
- Kritzik, K.L.; Kratz, G.; Panella, N.A.; Burkhalter, K.; Clark, R.J.; Biggerstaff, B.J.; Komar, N. Determining raptor species and tissue sensitivity for improved West Nile virus surveillance. J Wildl Dis. 2018, 54, 528–533. [Google Scholar] [CrossRef]
- Jiménez-Clavero, M.Á. Animal viral diseases and global change: bluetongue and West Nile fever as paradigms. Front Genet. 2012, 3, 105. [Google Scholar] [CrossRef] [PubMed]
- Hull, J.; Hull, A.; Reisen, W.; Fang, Y.; Ernst, H. Variation of West Nile virus antibody prevalence in migrating and wintering hawks in central California. Condor 2006, 108, 435–439. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Gould, D.H.; Bowen, R.A.; Komar, N. Natural and experimental West Nile virus infection in five raptor species. J. Wildl. Dis. 2006, 42, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, F.; Fischer, D.; Fischer, L.; Maisch, H.; Risch, T.; Dreyer, S.; Sadeghi, B.; Geelhaar, D.; Grund, L.; Merz, S.; et al. Vaccination of zoo birds against West Nile Virus—A Field Study. Vaccines 2023, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Angenvoort, J.; Fischer, D.; Fast, C.; Eiden, M.; Rodriguez, A.V.; Revilla-Fernández, S.; Nowotny, N.; de la Fuente, J.G.; Lierz, M.; Groschup, M.H. Pathogenesis of West Nile virus lineage 1 and 2 in experimentally infected large falcons. Vet Microbiol. 2013, 161, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Jiménez de Oya, N.; Escribano-Romero, E.; Blázquez, A.B.; Martín-Acebes, M.A.; Saiz, J.C. Current progress of avian vaccines against West Nile virus. Vaccines (Basel). 2019, 7, 126. [Google Scholar] [CrossRef] [PubMed]
- D'Agostino, J.J.; Isaza, R. Clinical signs and results of specific diagnostic testing among captive birds housed at zoological institutions and infected with West Nile virus. J Am Vet Med Assoc. 2004, 224, 1640–1643. [Google Scholar] [CrossRef]
- Steele, K.E.; Linn, M.J.; Schoepp, R.J.; Komar, N.; Geisbert, T.W.; Manduca, R.M.; Calle, P.P.; Raphael, B.L.; Clippinger, T.L., Larsen, T.; Smith, J.; Lanciotti, R.S.; Panella, N.A.; McNamara, T.S. Pathology of fatal West Nile virus infections in native and exotic birds during the 1999 outbreak in New York City, New York. Vet Pathol. 2000, 37, 208–24. [CrossRef]
- Joyner, P.H.; Kelly, S.; Shreve, A.A.; Snead, S.E.; Sleeman, J.M.; Pettit, D.A. West Nile virus in raptors from Virginia during 2003: clinical, diagnostic, and epidemiologic findings. J. Wildl. Dis. 2006, 42, 335–344. [Google Scholar] [CrossRef]
- Saggese, M.D. West Nile virus in Neotropical raptors: should we be concerned? In Neotropical Raptors; Bildstein et al., Eds.; United States of America, 2007, pp. 149-173.
- Ferraguti, M.; De La Puente, J.M.; Soriguer, R.; Llorente, F.; Jiménez-Clavero, M.Á.; Figuerola, J. West Nile virus-neutralizing antibodies in wild birds from southern Spain. Epidemiol. Infect. 2016, 144, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.C.; Nemeth, N.M.; Edwards, E.; Bright, P.R.; Komar, N. Passive West Nile virus antibody transfer from maternal Eastern screech-owls (Megascops asio) to progeny. Avian Dis. 2006, 50, 454–455. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Kratz, G.E.; Bates, R.; Scherpelz, J.A.; Bowen, R.A.; Komar, N. Naturally induced humoral immunity to West Nile virus infection in raptors. Ecohealth. 2008, 5, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Marra, P.P.; Griffing, S.; Caffrey, C.; Kilpatrick, M.A.; McLean, R.; Brand, C.; Saito, E.; Dupuis, A.P.; Kramer, L.; Novak, R. West Nile virus and wildlife. BioScience. 2004, 54, 393–402. [Google Scholar] [CrossRef]
- Wünschmann, A.; Shivers, J.; Bender, J.; Carroll, L.; Fuller, S.; Saggese, M.; Van Wettere, A.; Redig, P. Pathologic and immunohistochemical findings in Goshawks (Accipiter gentilis) and Great Horned Owls (Bubo virginianus) naturally infected with West Nile virus. Avian Dis. 2005, 49, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, C.; Franca, M.; Uzal. F.; Anderson, M.; Barr, B.; Woods, L.; Moore, J.; Woolcock, P.; Shivaprasad, H.L. Pathology and immunohistochemical findings of West Nile virus infection in Psittaciformes. Vet Pathol. 2011, 48, 975–984. [CrossRef]
- Jones, M.P. Selected infectious diseases of birds of prey. J. Exot. Pet Med. 2006, 15, 5–17. [Google Scholar] [CrossRef]
- Phalen, D.N.; Dahlhausen, B. West Nile virus. Semin. Avian Exot. Pet Med. 2004, 13, 67–78. [Google Scholar] [CrossRef]
- Wodak, E.; Richter, S.; Bagó, Z.; Revilla-Fernández, S.; Weissenböck, H.; Nowotny, N.; Winter, P. Detection and molecular analysis of West Nile virus infections in birds of prey in the eastern part of Austria in 2008 and 2009. Vet. Microbiol. 2011, 149, 358–366. [Google Scholar] [CrossRef]
- Bakonyi, T.; Ferenczi, E.; Erdelyi, K.; Kutasi, O.; Csörgö, T.; Seidel, B.; Weissenböck, H.; Brugger, K.; Bán, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Busquets, N.; Napp, S.; Alba, A.; Zorrilla, I.; Villalba, R.; Arenas, A. Serosurvey of West Nile virus and other flaviviruses of the Japanese encephalitis antigenic complex in birds from Andalusia, southern Spain. Vector Borne Zoonotic Dis. 2011, 11, 1107–1113. [Google Scholar] [CrossRef]
- Busquets, N.; Laranjo-González, M.; Soler, M.; Nicolás, O.; Rivas, R.; Talavera, S.; Villalba, R.; San Miguel, E.; Torner, N.; Aranda, C.; Napp, S. Detection of West Nile virus lineage 2 in North-Eastern Spain (Catalonia). Transbound Emerg. Dis. 2019, 66, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, H.; Beck, C.; Lecollinet, S.; Monier, M.; Mousson, L.; Zakeri, S.; Raz, A.; Arzamani, K.; Nourani, L.; Dinparast-Djadid, N.; Failloux, A.B. Serological evidence of West Nile virus infection among birds and horses in some geographical locations of Iran. Vet. Med. Sci. 2021, 7, 204–209. [Google Scholar] [CrossRef] [PubMed]
- García-Ripollés, C.; López-López, P.; Urios, V. First description of migration and wintering of adult Egyptian Vultures Neophron percnopterus tracked by GPS satellite telemetry. Bird study. 2010, 57, 261–265. [Google Scholar] [CrossRef]
- Phipps, W.L.; López-López, P.; Buechley, E.R.; Oppel, S.; Álvarez, E.; Arkumarev, V.; Bekmansurov, Rinur.; Berger-Tal, O.; Bermejo, A.; Bounas, A.; Alanís, I.C.; de la Puente, J.; Dobrev, V.; Duriez, O.; Efrat, R.; Fréchet, G.; García, J.; Galán, M.; García-Ripollés, C.; Gil, A.; Iglesias-Lebrija, J.J.; Jambas, J.; Karyakin, I.V.; Kobierzycki, E.; Kret, E.; Loercher, F.; Monteiro, A.; Morant, E.J.; Nikolov, S.C.; Pereira, J.; Peške, L.; Ponchon, C.; Realinho, E.; Saravia, V.; Sekercioğlu, C.H.; Skartsi, T.; Tavares, J.; Teodósio, J.; Urios, V.; Vallverdú, N. Spatial and temporal variability in migration of a soaring raptor across three continents. Front. Ecol. Evol. 2019, 7, 1-14. [CrossRef]
- Bravo-Barriga, D.; Aguilera-Sepúlveda, P.; Guerrero-Carvajal, F.; Llorente, F.; Reina, D.; Pérez-Martín, J.E.; Jiménez-Clavero, M.Á.; Frontera, E. West Nile and Usutu virus infections in wild birds admitted to rehabilitation centres in Extremadura, western Spain, 2017-2019. Vet. Microbiol. 2021, 255, 109020. [Google Scholar] [CrossRef] [PubMed]
- Marinković, D.; Nešić, V.; Davitkov, D.; Aničić, M. Causes of morbidity and mortality in European griffon vulture (Gyps fulvus) population in Serbia in the period of 2018–2022 – post-mortem findings. J. Comp. Pathol. 2023, 203, 52. [Google Scholar] [CrossRef]
- Nash, D.; Mostashari, F.; Fine, A.; Miller, J.; O’Leary, D.; Murray, K.; Huang, A.; Rosenberg, A.; Greenberg, A.; Sherman, M.; Wong, S.; Layton, M; 1999 West Nile Outbreak Response Working Group. The outbreak of West Nile virus infection in the New York City area in 1999. N Engl J Med. 2001, 344, 1807–1814. [CrossRef]
- Rideout, B.A.; Stalis, I.; Papendick, R.; Pessier, A.; Puschner, B.; Finkelstein, M.E.; Smith, D.R.; Johnson, M.; Mace, M.; Stroud, R.; Brandt, J.; Burnett, J.; Parish, C.; Petterson, J.; Witte, C.; Stringfield, C.; Orr, K.; Zuba, J.; Wallace, M.; Grantham, J. Patterns of mortality in free-ranging California Condors (Gymnogyps californianus). J. Wildl. Dis. 2012, 48, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Straub, M.H.; Kelly, T.R.; Rideout, B.A.; Eng, C.; Wynne, J.; Braun, J.; Johnson, C.K. Seroepidemiologic Survey of Potential Pathogens in Obligate and Facultative Scavenging Avian Species in California. PLoS One. 2015, 10, e0143018. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature. 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940, s1-20, 471–492. [Google Scholar] [CrossRef]
- Taylor, R.M.; Work, T.H.; Hurlbut, H.S.; Rizk, F. A study of the ecology of West Nile virus in Egypt. Am J Trop Med Hyg. 1956, 5, 579–620. [Google Scholar] [CrossRef]
- Mencattelli, G.; Ndione, M.H.D.; Rosà, R.; Marini, G.; Diagne, C.T.; Diagne, M.M.; Fall, G.; Faye, O.; Diallo, M.; Faye, O.; Savini, G.; Rizzoli, A. Epidemiology of West Nile virus in Africa: An underestimated threat. PLoS Negl Trop Dis. 2022, 16, e0010075. [Google Scholar] [CrossRef]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.C.; Panthier, R. Epidémiologie du virus West Nile: etude d'un foyer en Camargue. IV. La méningo-encéphalomyélite du cheval [Epidemiology of the West Nile virus: study of a focus in Camargue. IV. Meningo-encephalomyelitis of the horse]. Ann Inst Pasteur (Paris). 1970, 118, 239–247. [Google Scholar] [PubMed]
- Tsai, T.F.; Popovici, F.; Cernescu, C.; Campbell, G.L.; Nedelcu, N.I. West Nile encephalitis epidemic in Southeastern Romania. Lancet. 1998, 352, 767–771. [Google Scholar] [CrossRef] [PubMed]
- García-Carrasco, J.M.; Muñoz, A.R.; Olivero, J.; Figuerola, J.; Fa, J.E.; Real, R. Gone (and spread) with the birds: can chorotype analysis highlight the spread of West Nile virus within the Afro-Palaearctic flyway? One Health. 2023, 17, 100585. [Google Scholar] [CrossRef]
- Rappole, J.H., Derrickson, S.R.; Hubálek, Z. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerg Infect Dis. 2000, 6, 319–328. [CrossRef] [PubMed]
- Nir, Y.; Goldwasser, R.; Lasowski, Y.; Avivi, A. Isolation of arboviruses from wild birds in Israel. Am J Epidemiol. 1967, 86, 372–378. [Google Scholar] [CrossRef]
- Jupp, P.G. The ecology of West Nile virus in South Africa and the occurrence of outbreaks in humans. Ann N Y Acad Sci. 2001, 951, 143–152. [Google Scholar] [CrossRef]
- Hubálek, Z.; Halouzka, J. West Nile fever—a reemerging mosquito-borne viral disease in Europe. Emerg Infect Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef] [PubMed]
- López, G.; Jiménez-Clavero, M.A.; Tejedor, C.G.; Soriguer, R.; Figuerola, J. Prevalence of West Nile virus neutralizing antibodies in Spain is related to the behavior of migratory birds. Vector Borne Zoonotic Dis. 2008, 8, 615–621. [Google Scholar] [CrossRef]
- Lord, R.D.; Calisher, C.H. Further evidence of southward transport of arboviruses by migratory birds. Am J Epidemiol. 1970, 92, 73–78. [Google Scholar] [CrossRef]
- Malkinson, M.; Banet, C.; Weisman, Y.; Pokamunski, S.; King, R.; Drouet, M.T.; Deubel, V. Introduction of West Nile virus in the Middle East by migrating white storks. Emerg Infect Dis. 2002, 8, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Höfle, U.; Blanco, J.M.; Crespo, E.; Naranjo, V.; Jiménez-Clavero, M.A.; Sanchez, A.; de la Fuente, J.; Gortazar, C. West Nile virus in the endangered Spanish imperial eagle. Vet Microbiol. 2008, 129, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Casades-Martí, L.; Holgado-Martín, R.; Aguilera-Sepúlveda, P.; Llorente, F.; Pérez-Ramírez, E.; Jiménez-Clavero, M.Á.; Ruiz-Fons, F. Risk factors for exposure of wild birds to West Nile virus in a gradient of wildlife-livestock interaction. Pathogens. 2023, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Brugueras, S.; Fernández-Martínez, B.; Martínez-de la Puente, J.; Figuerola, J.; Porro, T.M.; Rius, C.; Larrauri, A.; Gómez-Barroso, D. Environmental drivers, climate change and emergent diseases transmitted by mosquitoes and their vectors in southern Europe: a systematic review. Environ Res. 2020, 191, 110038. [Google Scholar] [CrossRef] [PubMed]
- European Environment Agency (EUNIS). Available online: https://eunis.eea.europa.eu/habitats/393 (accessed on 23 October 2023).
- Díaz, M.; Campos, P.; Pulido, F.J. The Spanish dehesas: a diversity of land use and wildlife. In Farming and birds in Europe: The Common Agricultural Policy and its implications for bird conservation; Pain, D., Pienkowski, M., Eds.; Academic Press: London, England, 1997; pp. 178–209. [Google Scholar]
- Delgado-González, A.; Cortés-Avizanda, A.; Serrano, D.; Arrondo, E.; Duriez, O.; Margalida, A.; Carrete, M.; Oliva-Vidal, P.; Sourp, E.; García-Barón, I.; de La Riva, M.; Sánchez-Zapata, J.A.; Donázar, J.A. Apex scavengers from different European populations converge at threatened savannah landscapes. Sci Rep. 2022, 12, 2500. [Google Scholar] [CrossRef]
- Guerrero-Carvajal, F.; Bravo-Barriga, D.; Martín-Cuervo, M.; Aguilera-Sepúlveda, P.; Ferraguti, M.; Jiménez-Clavero, M.Á.; Llorente, F.; Alonso, J.M.; Frontera, E. Serological evidence of co-circulation of West Nile and Usutu viruses in equids from western Spain. Transbound Emerg Dis. 2021, 68, 1432–1444. [Google Scholar] [CrossRef]
- Morant, J.; Arrondo, E.; Sánchez-Zapata, J.A.; Donázar, J.A.; Cortés-Avizanda, A.; de La Riva, M.; Blanco, G.; Martínez, F.; Oltra, J.; Carrete, M.; Margalida, A.; Oliva-Vidal, P.; Martínez, J.M.; Serrano, D.; Pérez-García, J.M. Large-scale movement patterns in a social vulture are influenced by seasonality, sex, and breeding region. Ecol Evol. 2023, 13, e9817. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.; de Langarica, F.M.Z.G.; Molina, M.G. Spring migration of eurasian griffon vultures across the strait of Gibraltar: number, timing and age composition. Ardeola 2019, 66, 113–118. [Google Scholar] [CrossRef]
- Adelman, J.S.; Tokarz, R.E.; Euken, A.E.; Field, E.N.; Russell, M.C.; Smith, R.C. Relative influence of land use, mosquito abundance, and bird communities in defining West Nile virus infection rates in Culex mosquito populations. Insects. 2022, 13, 758. [Google Scholar] [CrossRef]
- Palomar, A.M.; Veiga, J.; Portillo, A.; Santibáñez, S.; Václav, R.; Santibáñez, P.; Oteo, J.A.; Valera, F. Novel genotypes of nidicolous argas ticks and their associated microorganisms from Spain. Front Vet Sci. 2021, 8, 637837. [Google Scholar] [CrossRef]
- Moraga-Fernández, A.; Oliva-Vidal, P.; Sánchez-Sánchez, M.; Muñoz-Hernández, C.; Martínez, J.M.; Margalida, A.; de la Fuente, J.; de Mera, I.G.F. Health risks associated with argasid ticks, transmitted pathogens, and blood parasites in Pyrenean griffon vulture (Gyps fulvus) nestlings. Eur J Wildl Res. 2023, 69, 112. [Google Scholar] [CrossRef]
- Blahove, M.R.; Carter, J.R. Flavivirus persistence in wildlife populations. Viruses 2021, 13, 2099. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.R.; Stallknecht, D.E.; Willis, J.; Conroy, M.J.; Davidson, W.R. Wild bird mortality and West Nile virus surveillance: biases associated with detection, reporting, and carcass persistence. J Wildl Dis. 2006, 42, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Cacho, I.M. Exposure and carriage of West Nile virus in feathered Iberian scavengers. Master’s thesis: Uppsala University, Uppsala. 2022.
- Gangoso, L.; Cortés-Avizanda, A.; Sergiel, A.; Pudifoot, B.; Miranda, F.; Muñoz, J.; Delgado-González, A.; Moleón, M.; Sánchez-Zapata, J.A.; Arrondo, E.; Donázar, J.A. Avian scavengers living in anthropized landscapes have shorter telomeres and higher levels of glucocorticoid hormones. Sci. Total Environ. 2021, 782, 146920. [Google Scholar] [CrossRef]
- FAO & UNEP. The State of the World’s Forests 2020. Forests, Biodiversity and People. FAO & UNEP, Rome, Italy, 2020; pp. 72-79.
- Morand, S.; Lajaunie, C. Outbreaks of vector-borne and zoonotic diseases are associated with changes in forest cover and oil palm expansion at global scale. Front Vet Sci. 2021, 8, 661063. [Google Scholar] [CrossRef]
Figure 1.
Distribution map of Egyptian vulture (Neophron percnopterus). Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Neophron percnopterus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
Figure 1.
Distribution map of Egyptian vulture (Neophron percnopterus). Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Neophron percnopterus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
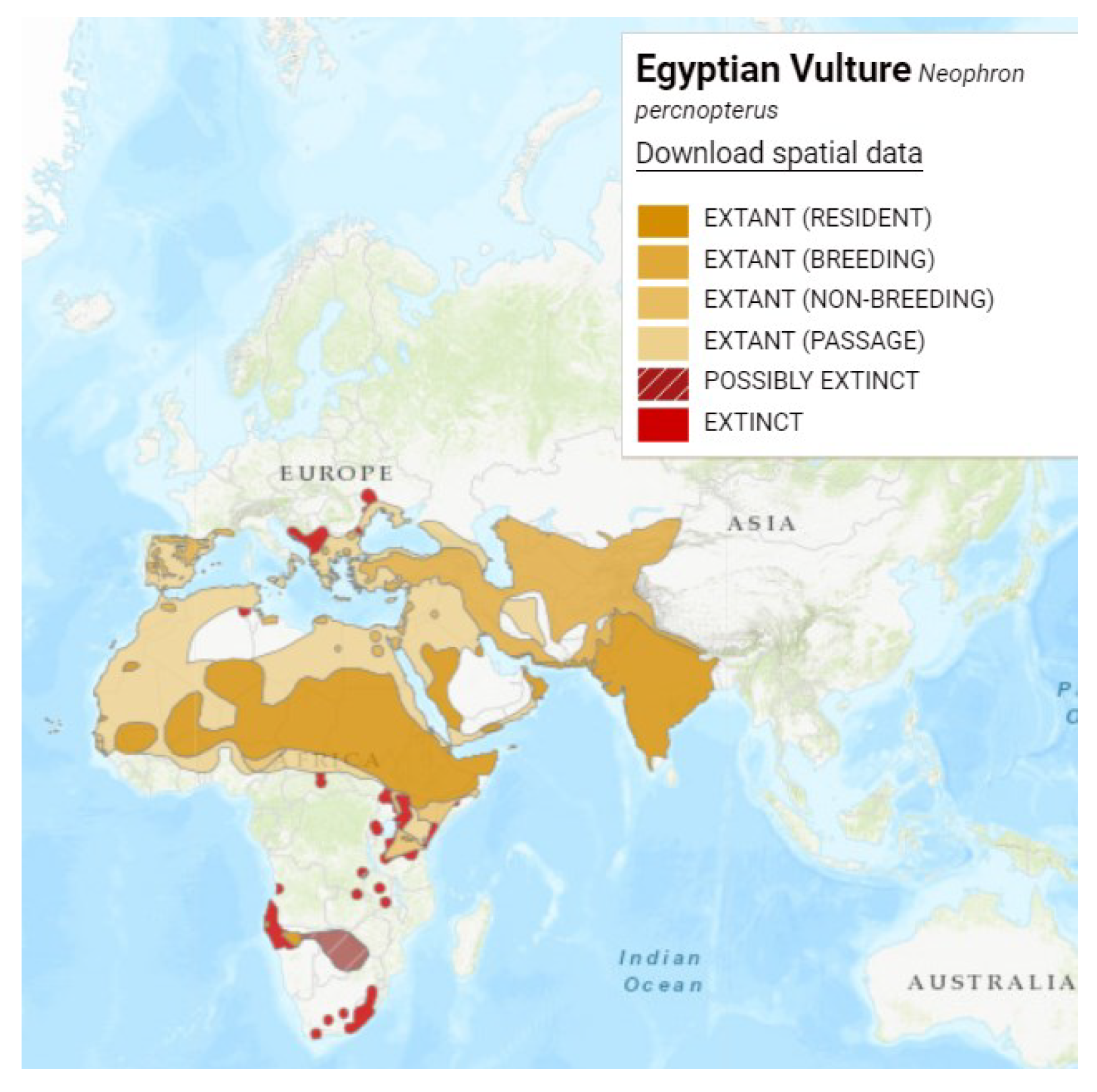
Figure 2.
Distribution map of bearded vulture (Gypaetus barbatus), Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Gypaetus barbatus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
Figure 2.
Distribution map of bearded vulture (Gypaetus barbatus), Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Gypaetus barbatus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
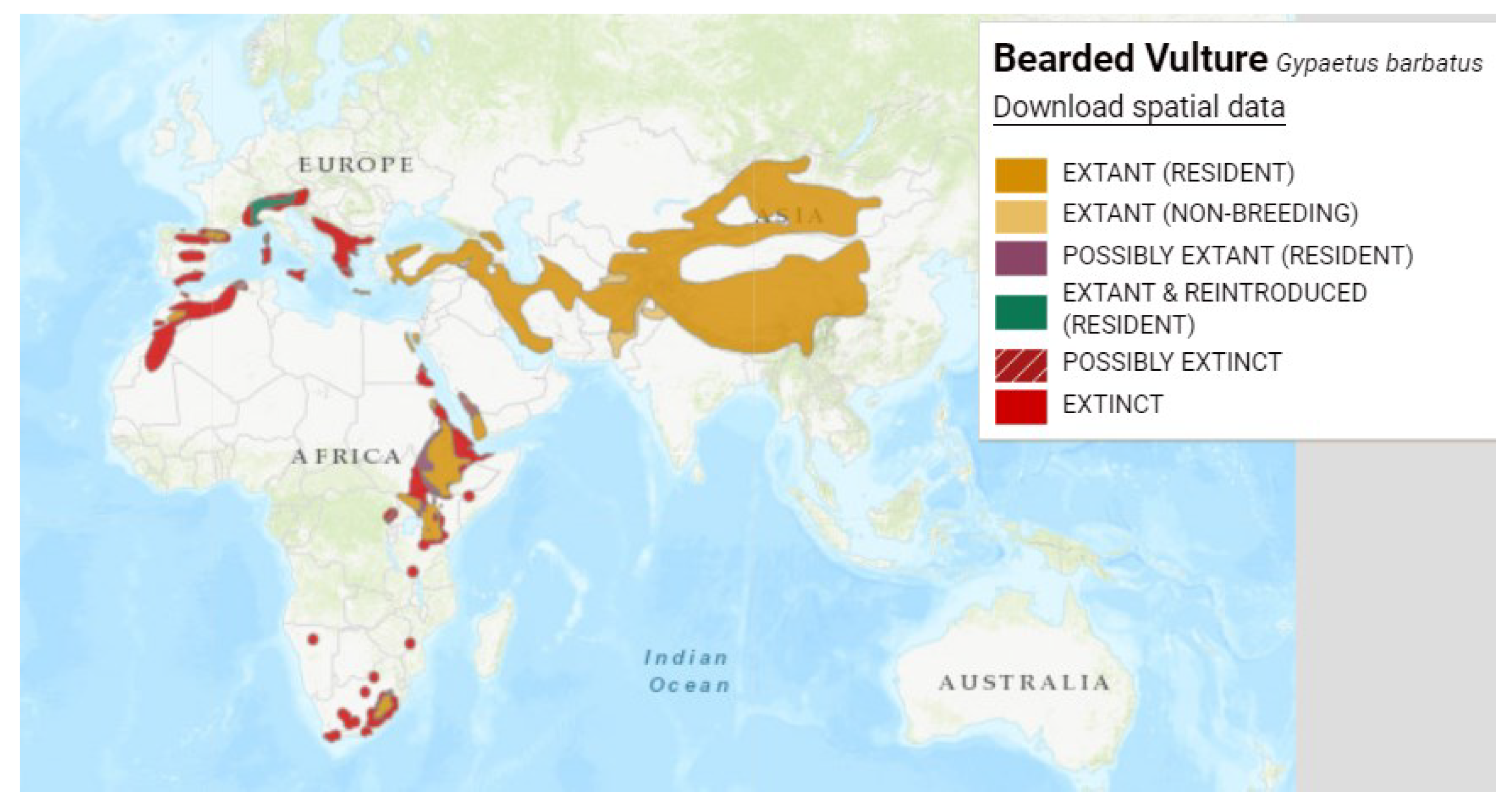
Figure 3.
Distribution map of griffon vulture (Gyps fulvus), Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Gyps fulvus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
Figure 3.
Distribution map of griffon vulture (Gyps fulvus), Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Gyps fulvus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
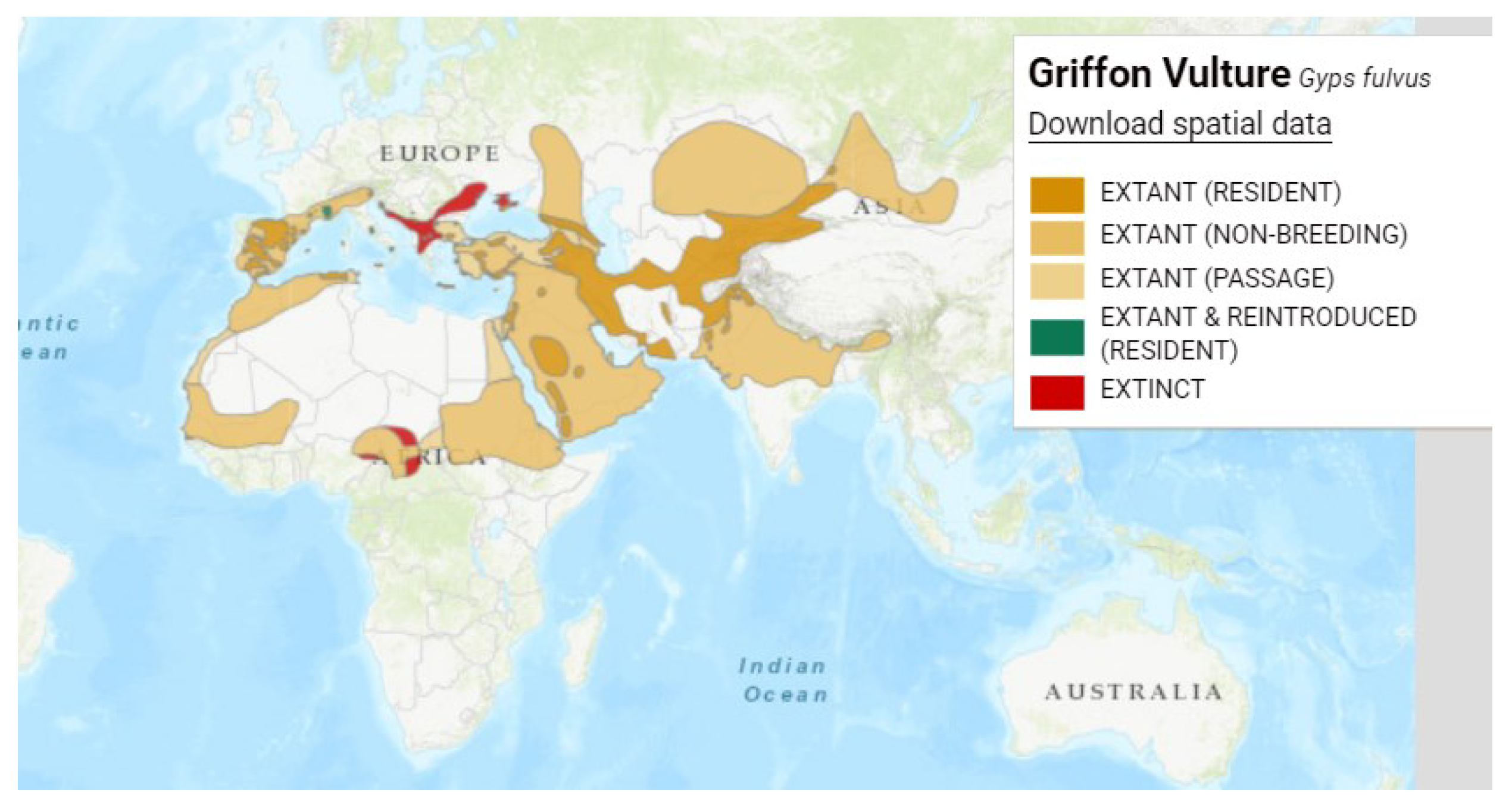
Figure 4.
Distribution map of cinereous vulture (Aegypius monachus), Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Aegypius monachus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
Figure 4.
Distribution map of cinereous vulture (Aegypius monachus), Reproduced with permission from BirdLife International and Handbook of the Birds of the World 2021. Aegypius monachus. The IUCN Red List of Threatened Species. Version 2022-2. [6].
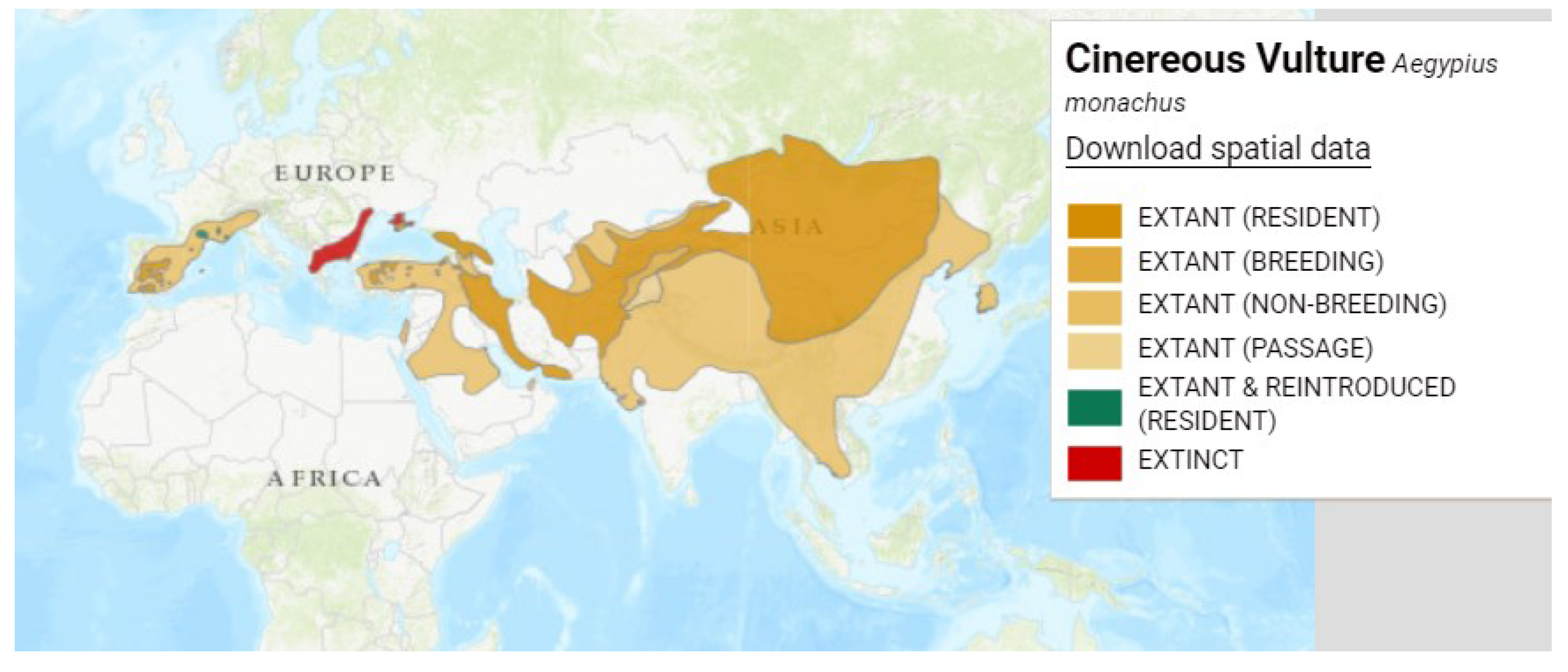
Table 1.
Causes of free-living vulture morbidity and mortality (adapted from the review by Ives et al. [15]).
Table 1.
Causes of free-living vulture morbidity and mortality (adapted from the review by Ives et al. [15]).
| Species | IUCN global statusa | Toxic | Trauma | Infectious | Idiopathic | Metabolic | Inflammatory | Totalb |
|---|---|---|---|---|---|---|---|---|
| Aegypius monachus | NT | 9 (485) |
1 (9) |
6 (46) |
1 (1) |
2 (16) |
1 (1) |
14 (542) |
|
Gypaetus barbatus |
NT |
8 (61) |
7 (80) |
1 (3) |
2 (2) |
0 |
1 (1) |
9 (143) |
|
Gyps fulvus |
LC |
18 (615) |
12 (978) |
5 (73) |
1 (5) |
1 (51) |
0 |
32 (1722) |
| Neophron percnopterus |
EN |
15 (500) |
13 (176) |
3 (38) |
0 |
0 |
0 |
25 (714) |
a LC – Least concern; NT – Near threatened; EN – Endangered; CR – Critically endangered. b Number of studies (number of affected individual vultures reported).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



