
Submitted:
06 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
Aquaculture is a fast-emerging food-producing sector in which fishery production plays an im-perative socio-economic role, providing ample resources and tremendous potential worldwide. However, aquatic animals are exposed to the deterioration of the ecological environment and in-fection outbreaks, which represent significant issues nowadays. One of the reasons for these threats is the excessive use of antibiotics and synthetic drugs that have harmful impacts on the aquatic atmosphere. It is not surprising that functional biotic feeds such as probiotics, prebiotics, postbiotics, and synbiotics have been developed as natural alternatives to sustain a healthy microbial envi-ronment in aquaculture. These functional feed additives possess several beneficial characteristics, including gut microbiota modulation, immune response reinforcement, resistance to pathogenic organisms, improved growth performance, and enhanced feed utilization in aquatic animals. Nevertheless, their mechanisms in modulating the immune system and gut microbiota in aquatic animals are largely unclear. This review discusses current research advancements to fill research gaps and promote effective and healthy aquaculture production.
Keywords:
Probiotics
; Prebiotics
; Synbiotics
; Gut microbiota
; Fishes
; Aquaculture
1. Introduction
Aquaculture is a fast-emerging food manufacturing sector worldwide, with Asia currently contributing to 90% of the total production. This sector involves the culture of tilapia, carp fishes, catfishes (non-air breathing and air-breathing), pangasius fishes, and prawns. Tilapia, in particular, is one of the main farmed fish globally and has seen a quadrupling in production over the past few decades due to its suitability for aquaculture, market demand, and stable market prices [1]. Among the major socio-economic sectors in the world, aquaculture generates numerous employment opportunities and fulfills a vital need for nutrients [2]. China is one of the largest seafood producing country in the world, alongside others like India, Vietnam, Bangladesh, Indonesia, Norway, and Egypt. In 2018, China alone produced 62.2 million tons of fish, representing more than 60% of the total fish production [3].
Aquatic animals, especially fishes, are exposed to adverse conditions due to the deterioration of the ecological environment. Disease outbreaks pose a significant challenge for aquaculture, impacting both the financial status and the economic development of communities in the Asia-Pacific region. Various types of aquatic microbial pathogens (such as Aeromonas hydrophila, Aeromonas salmonicida, Vibrio anguillarum, Vibrio vulnificus, Vibrio salmonicida, Streptococcus sp., Yersinia ruckeri), viruses (causing diseases like hematopoietic necrosis, yellow head virus, viral hemorrhagic septicemia), and parasites (e.g., Ichthyophthirius multifiliis) can cause different types of diseases in aquatic animals, including luminous vibriosis, filamentous bacterial disease, shell disease, larval mycosis, protozoan infections white spot, velvet disease, fungal infections, and dropsy [4–6].
The aquaculture production sector typically relies on traditional methods that involve the use of antibiotics (e.g., chloramphenicol, fluoroquinolones, nitrofurans, quinolones, florfenicol, sufamerazine, chorionic gonadotropin, oxytetracycline dihydrate, oxytetracycline hydrochloride) and synthetic chemicals (e.g., nitrofurans, formalin, malachite green, potassium permanganate, copper sulfate, and Neguvon) to control diseases. However, some of these chemotherapeutic applications have been widely criticized due to their negative impacts on aquaculture such as marine debris gathering, the rise of drug resistance and immunosuppressant activity. The intensive use of these traditional practices leads to the buildup of antibiotic and chemical residues, not only in aquatic animals but also in consumers, resulting in side effects such as diarrhea, vomiting, and stomach problems. Moreover, these traditional methods mainly affecting aquatic productions have been reported to be ineffective in controlling diseases in large-scale aquaculture processes [7–12].
In fish, the gastrointestinal tract (GIT) microbiota plays several vital functions. These microbial consortia increase digestive action, enhance the immune system, protect against harmful microbes, and improve intestine development [13]. In recent years, some gnotobiotic (germ-free) animal models have been developed as wonderful tools for studying host-microbe interaction. Moreover, gnotobiotic models have been used to investigate the role of gut microbiota in xenobiotic metabolism [14,15]. Using zebrafish (Danio rerio) model, researchers observed that the presence of alkaline phosphatase in the brush border intestine plays a vital function in gut epithelium division, as well as in the modulation of gene expression in bacteria, which possesses various functional properties (e.g., epithelial maturation, hormone-secreting endocrine organs and mucous secreting goblet cells in the gastrointestinal tract in D. rerio larvae [16,17]. Recently, it was reported that TLR2/MyD88 signaling played an essential role in innate immune recognition and activation during the colonization of two indigenous bacteria (Chryseobacterium ZOR0023 and Exiguobacterium ZWU0009) in zebrafish [18]. Indigenous probiotic microbes have significant functions such as developing the immune system (nonspecific and specific immunity) and inducing different types of cytokines, namely TNF-a, interleukins (IL-6, IL-10, IL-12), and IFN-c [19]. Indigenous probiotic Bacillus pumilus SE5 activates the expression of TLR2 signaling and antibacterial peptides genes in the intestine of grouper (Epinephelus coioides) The enhanced TLR2 signaling may result from the interaction of the host with the probiotic cell components [20,21]. In addition to enhancing the immune system in fish, the gut microbiota also provides important protection against pathogenic organisms [22,23].
Functional feed additives of probiotics, prebiotics, and/or synbiotics in diets have been extensively recommended to maintain a healthy GIT microbial community, improving infection resistance, and consequently promote the health of cultured aquatic organisms [24–26]. These biotic-based ingredients, consisting in live microorganisms, inert substrates, and the combination of both, possess a wide range of multiple functionalities They represent alternative nature-based solutions for improving aquatic animal health and production [23,27,28]. This review provides insights into the current developments in the utilization of probiotics, prebiotics, postbiotics, and synbiotics in aquaculture applications. It also presents a new way to develop the healthy and modern aquaculture industry.
2. Probiotics
2.1. Definition, and characteristic features
The Food and Agriculture Organization (FAO) of the United Nations and the World Health Organization (WHO) defined probiotics as “Live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” [29]. Recently, the term probiotics is associated with microbial feed additives, when controlled in enough amounts, conferring health and beneficial impact on the host of aquatic animals [28].
Probiotics act as a defense system for the host against harmful microbes or foreign substances [30–33]. They also produce beneficial bioactive molecules such as enzymes, proteins, lipids, organic acids, and others. Some of these bioactive molecules improve adherence to probiotics and reduce therefore the activity of pathogens in the gut region through the surface competition mechanism [34]. Probiotics play a significant responsibility in strengthening the immune system of the host [35]. While earlier studies have noted the utilization of probiotics in pigs, poultry, cattle, and humans, their application in aquaculture is a relatively new idea [36,37]. Probiotics might be managed in two ways in aquaculture. They can be supplemented with feed to modulate the gut microbes, or they can be managed by direct addition into the water, thereby inhibiting the growth of pathogens. These modes of administration are very critical in the utilization of probiotics in finfish and shrimp aquaculture ([38,39]. Probiotics could be alive, dead, or microbial cell components, and provide benefits to the host when administered via the feed or to the rearing water. This is achieved at least in part via improving the microbial balance of the host or ambient environment [39]. Figure 1 summarizes the different entryways of probiotics and their benefits in the aquaculture system.
Probiotics appear a new element agent for the development of aquaculture systems by exerting several favorable effects on growth activity, immune system, digestion, water quality, inhibition of pathogens, and regulation of gut microbes of aquatic animals. The utilization of probiotics in aquaculture is a modern trend, although its effectiveness in the aquatic ecosystem has not been considered comprehensively. Probiotics are ubiquitous, commonly present in aquatic animals, and play an important protective role throughout the digestive system [40,41]. Mainly represented by Lactobacilli, these beneficial microorganisms are vital to prevent illnesses and improve the aquatic animal GIT functions by excreting secondary metabolites such as lactic acid and other bioactive compounds [42,43]. These biomolecules synthesized by probiotics protect against the inhibitory molecules from predators [44]. They also can be extracted from probiotics in terrestrial plants and marine life forms, and then utilized to enhance disease resistance, develop the immune system, reduce environmental stress, and increase feed quality levels [45,46]. Advanced studies in this field have reported microbial by-product biomolecules such as enzymes, lipids, proteins, and immune toxins [47]. Nowadays, there exist some probiotic products commercially available, which are already used in aquaculture as feed additives [48]. These microbial by-products are beneficial and are mainly helpful in enhancing the health status of aquatic animals.
The characterization and identification of potential probiotics are based on various criteria such as acid and bile tolerance, no hemolytic activity, inhibition of pathogens, no pathogenicity, survivable in storage and field, cultivable on a big scale, competent to adhere to the epithelial liner of the gut, free of plasmid-encoded antibiotic resistance genes, safe for use as a feed additive, acting as a growth promoter, anti-inflammatory, antimutagenic, immunostimulatory, and beneficial effects on host animals. Every new strain used for probiotic expansion should contain all the aforesaid features [27,49,50].
2.2. Possible modes of action of probiotics in aquaculture
Significant effects of probiotic, e.g. Bacillus sp. as feed supplements, include the improvement of growth performance, digestive enzyme activity, infection resistance, and immune response in aquatic animals [51,52].
2.2.1. Probiotics act as a growth enhancer in aquaculture
One of the mechanisms, which regulates the metabolism of amino and fatty acids, is the capacity of various probiotic strains in producing vitamin B12, as revealed by a study on carp gut [53,54]. In addition, this is helpful to enhance fish growth and eradicate vitamin B12 deficiency in fish [55]. Also, essential macronutrients are usually supplied through feeds. Various micronutrients such as amino acids, vitamins, and fatty acids are very important for physiological functions as nutrients in aquatic animals [56–58]. For instance, diverse fish species such as carp (Cyprinus carpio), rainbow trout (Oncorhynchus mykiss), channel catfish (Ictalurus punctatus), and tilapia (Oreochromis niloticus) are identified to synthesize vitamin B12 [59–61]. The growth and survival rates of juvenile black tiger shrimp (Penaeus monodon) were enhanced when these were fed for 100 days with a combination of Lactobacillus sp., previously isolated from the GIT of chicken [62]. In fact, probiotics improve the digestive function of aquatic animals by producing or inducing the secretion of different kinds of extracellular enzymes such as proteases, amylases, and lipases. They also provide various growth nutrients such as vitamins, fatty acids, and amino acids [63]. The function of probiotics results in abridged feed cost, which accounts for 60-70% of the contribution cost of fish production [64,65]. Both the maximum growth performances and most excellent feed conversion ratio were detected when O. niloticus was fed with probiotic Micrococcus luteus [66,67]. Bacillus subtilis improved feed digestibility, enhanced weight gain, feed conversion, and significantly increased the survival rate of Bullfrog (Lithobates catesbeianus) fed with different doses (2.5, 5.0, and 10.0 g/kg) [68,69]. Bacillus species aid in the digestion of aquatic animals by supplying exoenzymes (proteases, lipases, and amylases) that enhance digestive enzymatic [70]. It was observed that Streptococcus faecium and Lactic acid bacteria (LAB) improved the growth and feed competence of Israeli carp and juvenile carp, respectively [71].
2.2.2. Biocontrol of bacterial diseases in aquaculture
In the past decades, numerous studies have stated that probiotics synthesize different types of inhibitory substances responsible for the antagonistic activity against pathogens. Two probiotic strains of LAB (Lactococcus lactis MM1 and Enterococcus faecium MM4) isolated from the intestine of orange-spotted grouper (E. coioides) can secrete several inhibitory substances such as hydrogen peroxide and bacteriocin-like substances. These can be utilized to induce antimicrobial activity against different pathogens such as Staphylococcus aureus, V. harveyi, and V. metschnikovi that affect grouper (E. coioides) [72,73]. Probiotic B. pumilus H2 had strong inhibitory activity against Vibrio sp through its main mechanism of amicoumacin production to involve disrupting the cell membrane and cell lysis, thus showing anti-Vibrio activity [74,75]. Probiotic Bacillus velezensis cell-free supernatant contained different types of bioactive molecules against A. salmonicida infection [76]. The lipopeptides N3 synthesized by the probiotic Bacillus amyloliquefaciens M1 have strong antibacterial activity in the whole-cell membrane, which can exhibit significant effect of ion-conducting channels in the whole-cell membrane as well as membrane-active properties [77,78]. The probiotic species Clostridium butyricum culture supernatant included different types of inhibitory substances, lowered the pH in the intestine, and thus decreased the growth of pathogens in fish intestinal epithelial cells [79]. Probiotic E. faecium was supplemented to the diets of Olive flounder and can enhance the antibacterial activity [80].
2.2.3. Biocontrol of viral diseases in aquaculture
Microorganism strains with potential probiotic effects in aquaculture such as Pseudomonas spp., Vibrios spp., and Aeromonas spp. induce antiviral effects against hematopoietic necrosis virus (IHNV) infection [81,82]. Similarly, the potential probiotic strain Pseudoalteromonas undina VKM-124 was used to improve Yellow Jack (Carangoides bartholomaei) larval survival and enhance the antiviral effect against Neuro Necrosis Virus (SJNNV) infection [83,84].
2.2.4. Immunostimulant agents in aquaculture
Immunity development and modulation are amongst the various health benefits of probiotics in aquaculture. The majority of the earlier studies have dealt with the health-boosting capability of probiotics in aquatic organisms. Currently, probiotics are very much focused on the immunological development properties of the piscine system [19]. Different types of probiotics improve various immunological properties, and notably several fishes use the efficiency of probiotics to vitalize teleost immunity in both in situ and ex-situ conditions [85]. Although hopeful findings were reported in previous studies, most of immunostimulants did not progress to large-scale function for fish. Since various immunostimulants in aquaculture produce similar effects, researchers have demonstrated the utilization of probiotics to enhance disease resistance and the immune system of carp fish species [86,87]. Several carp fishes showed an increase in the production of total serum protein, nitric oxide, lysozyme, albumin, phagocytic activity by blood leucocytes, and the expression of IL-1b, superoxide anion, and myeloperoxidase content, respiratory burst activity, and globulin levels, complement C3, TNF-α, and lysozyme-C [86,88]. Current study reports indicate that probiotics (either single or mixed type) could enhance the immunological development of fish [89]. Those reports have emphasized the immunomodulating properties of beneficial living cell organisms and the factors to facilitate the optimal induction of defense responses in the fish community. The probiotic strain B. pumilus SE5 isolated from the intestine of fast-growing grouper E. coioides [90,91] and subsequent studies demonstrated that both the viable and heat-inactivated B. pumilus SE5 could shape the intestinal immunity and microbiota [92], and improve the growth performance and systemic immunity in E. coioides [93]. The dietary supplementation of the cell wall (CW), peptidoglycan (PG), and lipoteichoic acid (LTA) of probiotic B. pumilus SE5 and their effect on intestinal immune-related genes expression and microbiota have been evaluated in 60 days of feeding trial. PG and LTA of probiotic B. pumilus SE5 were more effective than CW in shaping the intestinal immunity and microbiota in E. coioides [20], even if the mechanisms are largely unclear and need further study.
2.2.5. Interference of quorum sensing in aquaculture
Quorum sensing (QS) is a communication system among bacterial cells, which is very useful in controlling different kinds of biological macromolecule expressions like the virulence agents in a cell thickness-dependent comparative performance [94]. In this process, control of bacterial cells in gene expressions by generative, loosening and conceiving tiny marker molecules are called auto-inducers [95]. Disruption of the QS process of pathogenic organisms has been a probable anti-infective strategy, and different types of methods have been used to investigate the analysis of QS. These include the inhibition of signal molecule biosynthesis, application of QS antagonists, chemical inactivation of QS signals by oxidized halogen antimicrobials, signal molecule biodegradation by bacterial lactonases and by bacterial and eukaryotic acylase, and application of QS agonists in aquaculture [96,97]. N-Acyl homoserine lactones (AHLs) are the most important family of QS auto-inducers utilized in Gram-negative bacteria, and their biodegradation showed as a potential method to interrupt QS [98]. Bacillus sp. QSI-1 is a quorum quencher on virulence agent production and biofilm arrangement of the fish pathogen (A. hydrophila). The fish fed with QSI-1 was experimentally performed to have a relative percentage survival of 80.8%. The results indicate those AHLs degrading bacteria should be contemplated as an alternative to antibiotics in aquaculture for the biocontrol of bacterial fish infections [98,99]. In a biofilm system, bacteria are resistant to high temperatures, phagocytic cells, surfactants, antibiotics, and antibodies, and could alter their vital transmissions by quorum sensing signaling [100]. Probiotic Bacillus strains can effectively secrete quorum quenching enzymes and could reduce the pathogenic activity of A. hydrophila YJ-1 and control gut microbiota [101,102]. However, the form of action for this strain is still limited. Dietary supplementation of probiotics with quorum quenching activity such as Bacillus cereus QSI-1 has been shown to increase the intestinal barrier function and enhance the immune system of crucian carp against A. hydrophila infection. The quorum quenching bacteria increased the expression of tight junctions (TJ) proteins, ZO-1 and Occludin, which control the permeability and absorption of the intestinal mucosal barrier of crucian carp [103].
2.2.6. Stress improvement in aquaculture system
Stress in a fish’s life cycle disrupts the entire production. The culture species may be weakened and averse to taking food. This is called food irrational fear. In this condition, probiotics in culture farms can decrease stress levels as well as help to enhance the innate immune system against pathogens and environmental stressors [6,104]. Probiotic treatment is very helpful in increasing the production of fish within the given time and it also reduces the stress level in normal aquaculture practice.
Studies have concluded that the use of some probiotic strains increased chronic stress resistance in zebrafish (D. rerio) [105,106]. An experimental nutritional probiotic Lactobacillus delbrueckii ssp. Delbrueckii supplementation of sea bass led to a decrease in cortisol levels from 25 to 59 days, which in fish tissue is a stress indicator since it is directly engaged with the host's reaction to stress [107]. One more approach evaluated that fishes treated with probiotics exhibited increased flexibility in stress tests when compared with the control group [65]. Antioxidative properties of probiotic Lactobacillus fermentum induce protective action in the intestinal microbial ecosystem and help to overcome exo- and endogenous oxidative stress [108]. The probiotic strain Bacillus coagulans SCC-19 alleviates the non-specific immune damage induced by cadmium in common carp while relieving oxidative stress induced by cadmium in fish [109].
2.2.7. Reducing heavy metals in aquaculture
Heavy metals (e.g., lead, cadmium, silver, chromium, mercury, cobalt, zinc, iron, and copper) are present in the soil, water, and atmosphere [110–112]. These metals can cause toxic effects on all organisms and pose a huge risk to food quality, crops, and environmental quality. Heavy metals are mainly connected with anthropogenic action in the ecosystem [113]. Aqueous release from metal industries (steel, mining, and electroplating) contains elevated levels of heavy metals that locate their system into water bodies which are also utilized for aquaculture action [114,115]. These heavy metals get accumulated in fish tissue, and thereby is a matter of great concern in human consuming via the food chain breathing. Their elimination is very helpful in reducing the toxic effect of the aquatic environment and outflow is subsequently imperative [116]. Among all recommended methods for eliminating heavy metal is the process of utilizing microbes, which is cost-effective [117]. Bacterial metabolic action activates the heavy metals via the production of organic and inorganic acids, complex formation with organic ligands, or oxidation reactions [118]. Microbes and microbial by-products can accumulate separate minute particles in the form of dissolved metals [119]. Generally, heavy metals activate the sporulation development of Bacillus species and thus decrease the heavy metal absorption [114,120]. In addition, probiotic microbes from aqua farming sediments can be utilized as a dietary supplement and helps to remove heavy metals, metal-resistant, and antibiotic-resistant categories from the intestine of aquatic organisms, particularly fish, to control the progress of heavy metals accumulation [121].
2.3. Major probiotic genera as biocontrol agents in aquaculture
The major probiotic genera used in aquaculture are Lactobacillus and Bacillus [122]. Most in case, Bacillus, Lactobacillus, Lactococcus, Leuconostoc, Pediococcus and Weissella have been isolated from fish and shellfish gut [123–127]. Supplementation in aquaculture feed is achieved using single-strain probiotics or association of various bacteria as multi-strain probiotics (MSP), which are reported to have more beneficial effects to host, owing to synergistic effects among various strains [128]. Table 1 lists some examples of probiotic-based functional feed additives for aquatic animals.
3. Prebiotics
Prebiotics are "non-digestible sugars, which helpfully influence the host by specifically enhancing the development of health-encouraging strains in the gut" [158,159]. Prebiotics improve the synbiotic association amid the gut microbiota of the host [160], and are also known as immunosaccharides. There are various types of prebiotic compounds, including mannan oligosaccharides (MOS), fructooligosaccharides (FOS), and arabinooligosaccharides (AOS), which play a significant role in improving the natural immune system [161]. MOS is most frequently used in animal diets. This prebiotic improves growth activity, feed utilization, survival rate, development of immune reaction, and antagonistic activity against aquatic pathogens [162–164]. Oligosaccharide-type components have been connected with the development of immunity [165,166], and were used extensively in diverse fish species such as Psetta maxima [12], Larimichthys crocea [167], Paralichthys olivaceus [168], Rutilus rutilus [169], Piaractus mesopotamicus [170] and Acipenser Persicus [171]. Previous study reports have examined the function of prebiotics in cultured finfish and shellfish, explaining that these compounds have significant effects on gut microbial composition, immune system, and infection resistance against pathogenic organisms in fishes [172,173]. Previous studies have also verified the health beneficial effects of prebiotics on growth, and physiological status [174]. Prebiotics can improve the capability and feasibility of aquaculture production. The most frequently used prebiotics, including xylooligosaccharide (XOS), FOS, transgalactooligosaccharide (TGOS), glucooligosaccharide (GOS), soybean oligosaccharide (SBOS), polydextrose, inulin, and Lactosucrose, enhance the aquaculture production [175]. Natural sources of prebiotics in vertebrates include onion, garlic, tomato, honey chicory, leek, and so on [176].
3.1. Action in the gastrointestinal tract of aquatic animals
Prebiotics exert possible effects on host biological response, protecting fish species against harmful microbes and thus decreasing their mortality. However, the evaluation of the intestinal microbiota of important commercial fishes like hybrid striped bass, channel catfish, salmonids and tilapia is necessary to infer if there are any particular bacterial species to be enhanced by the utilization of prebiotics. By increasing the production of volatile fatty acids (VFA) in the GIT, the host's advantage is the inhibition of potentially pathogenic organisms [177,178]. The synthesis of VFA in the aquatic organisms’ GIT indicates the presence of microbial communities [179]. The herbivorous fish were the first species (Kyphosus cornelii and K. sydneyanus) shown to contain VFA synthesized by the intestinal bacterial community [180]. Another fish species tilapia (Oreochromis mossambicus) was established to have VFA produced by intestinal bacterial communities [181]. Prebiotics have numerous favorable effects in aquatic animals by enhancing disease resistance and improving nutrient accessibility [182]. Recently, our group evaluated the effects of FOS on growth performance and predominant autochthonous intestinal microbiota of shrimp (L. vannamei) fed diets with fish meal partially replaced by soybean meal. The results showed that a dietary supplement of 2-4 g/kg FOS could improve the growth performance, and survival rate, and exert a beneficial effect on the intestinal microbiota of shrimp. A dose adding 2-4 g/kg FOS in shrimp diets with fish meal partially replaced by soybean meal was recommended [183,184].
3.2. Regulation in the immune system of aquatic animals
In the past decades, prebiotics has been used to regulate intestinal microbiota, modulate immunity, control pathogens, and increase the survival ability of aquatic animals. Fishes are a diverse group of organisms that include sharks, rays, and bony fishes [195]. Similar to all vertebrates, fishes rely fully on their natural immunity against pathogens due to the restrictions in their adaptive immune function [187]. There are various cellular and soluble components primarily concerned with the immune responses including phagocytes, leukocytes, and auxiliary cells, which are organized in tissues and organs with leukocytes being the most functional. The impacts of prebiotics on immunity are indirect and involve the modification of the gut microbes, thereby enhancing the immune system. Thus, these beneficial components assist in supply change effectiveness, enhance fish growth, and induce inhibitory activity against pathogens through the viable prohibition of linkage sites, synthesis of natural organic acids (e.g., formic acid, lactic acid, acetic acid), hydrogen peroxide, and numerous other compounds like bacteriocins, siderophores, lysozyme, and antibiotics. This also causes a change in physiological and immunological responses in fish’s spleen, kidney and thymus, which are major lymphoid organs [48,188]. The prebiotic components act as growth promoter for commensal microbes to inhibit the adhesion and assault of harmful microorganisms in the epithelial cells. A beneficial effect of monosaccharide components arises from enhancing immune function and acting as a protection system for lymphoid organs as well.
3.2.1. Phagocytosis
Phagocytosis is the process by which immune cells like macrophages and neutrophils, engulf and digest foreign cells or particles, such as bacteria, viruses, or cellular debris [189]. FOS (0.5%) is used to enhance phagocytosis, respiratory burst, and phenoloxidase activity of sea cucumber coelomocytes and infection resistance against V. splendidus infection [190]. The phagocytic capability of inhabitant and obtained trout macrophages are related to the circumstances (i.e., in suspension versus attached and spread) of the cells at the time of particle treatment. Substrate binding and cells spreading may play a very important function in controlling the overall phagocytic capability of macrophages. Since the host’s resistance against infectious agents depends upon the phagocytic ability of the cells, the finding that obtained trout macrophages can surround larger numbers of activity latex particles than inhabitant cells provides a better understanding of immune regulatory mechanisms in fish [191]. Dietary supplementation of FOS significantly improved lysozyme activity compared to control diet group. However, there were no significant effects of the phagocytic percentage of the phagocytic index. In addition, a combination of FOS and MOS (5.0 g/kg) showed a significant difference in phagocytic activity of Japanese flounder [171].
3.2.2. Macrophages activation
Macrophages play a very imperative function in the nonspecific and specific connection of the immune function to synthesize the highest level of immune reaction and eliminate harmful microbes. Macrophages are stimulated to produce diverse inflammatory cytokines like tumor necrosis factor (TNF), IL-1, IL-12, etc. [80]. The alterations in the physiology of macrophages as a result of environmental signals can benefit them with improved antimicrobial activity. Nevertheless, ecosystem signals do not always cause changes that improve macrophage immune activity. Both nonspecific and specific immune responses can result in macrophages that are more vulnerable to harmful infections and less prepared to generate cytokines that enhance immune system response [192].
3.2.3. Respiratory burst activity
A respiratory burst is the fast release of reactive oxygen substances namely superoxide anions, hydrogen peroxide, and hydroxyl radicals. These reactive oxygen compounds are generally used to defend the capability of the host organism to counter harmful microbes. They are synthesized by activated phagocytes that are responsible for destroying microbes [193]. Respiratory burst analyses were performed in naturally resistant cells and blood neutrophils using NBT (nitro blue tetrazolium) or MPO (myeloperoxidase) methods. Inulin (5 g kg-1) was utilized as a dietary nutrient supplement for Nile tilapia and improved lysozyme and hematocrit NBT action. It also significantly enhanced the natural immune system and increased the survival rate against A. hydrophila infection [194,195].
The marine invertebrates contain enzymes such as tyrosinases, laccases, and catecholases which can be modified to complement the system of prophenoloxidase. This enhancement improves antagonistic activity through processes like phagocytosis and respiratory burst via opsonization. In a study conducted on red swamp crayfish, supplementation of a prebiotic nutrient diet with 8 and 10 g kg-1 FOS over a 30-day trial period significantly enhanced phenoloxidase reaction, stimulated immune-related genes (lysozyme, crustin 1, SOD), and increased the survival rate and antibacterial activity against A. hydrophila infection [196].
3.2.4. Synthesis of antibodies
B lymphocytes could produce special antibodies for recognizing specific microbial antigens and these antibodies could neutralize the antigens by surface binding and attaching to the target cells. Antibodies could also perform phagocytosis activity through the activated complement system, and antibody-dependent cellular cytotoxicity (ADCC). The hematocrit or total hemocyte was analyzed to count the total number of blood cells including WBC, RBC, and platelets. The hematocrit analytical methods are used for macro immune system analysis in fish [197]. These methods are used for the enhancement of immune cells (neutrophils, basophils, eosinophils, lymphocytes, and monocyte) in fish blood. The dietary supplementation of MOS and β-glucans were used to enhance the immune system of carp fry [198,199].
3.3. Major prebiotics with biocontrol capabilities in aquaculture
3.3.1. β-glucans
There is enough evidence available regarding their positive effects on immune responses, disease resistance, and growth performance upon oral delivery to a variety of farmed animals such as salmonids [200], sea bream [201], and shellfish [202]. Supplementation of β-glucans as prebiotic enhances the growth activity and higher resistance action against pathogens in P. vannamei [203]. Prebiotic administration of β-glucans in diet is used to increase disease resistance; its efficiency depends on their origin and structure [204]. The Glucan substance extracted from the cell wall of yeast (S. cerevisiae) has the ability to enhance the non-specific immune system and disease resistance in the Atlantic salmon [205].
3.3.2. Oligosaccharides
The oligosaccharide components are crucial for the modulation of immune responses in fish species. The positive results of monosaccharide products have encouraged the development of various immunomodulating, environmentally friendly nutrient diet supplements for fish species [206]. The diet supplementation of MOS from 1 to 1.5g kg-1 was capable to get better the growth activity and the feed efficiency of common carp fingerlings, as well as their antibacterial ability against A. hydrophila infection [207]. The nutrient feed additives (FOS) in beluga (Huso huso) juveniles had numerous beneficial effects such as gut microbiota modulation, immune response, and digestive enzyme action and growth performance [208]. They also presented a 7-week study report in common carp with dietary supplementation of FOS at (0%, 0.5%, and 1%) different levels which proved to have significant effects on intestinal microbiota modulation and physiological response [209]. The dietary supplementation of MOS at 0.4% improved the growth performance and non-specific immune response of Asian catfish (Clarias batrachus) juvenile’s [210]. The prebiotics FOS, when used as feed additives in juvenile large yellow croakers was found to improve growth action and digestive enzyme action [12,211].
Not all the prebiotic substances have immunostimulant properties, only a few references are available regarding the effects of isomalto-oligosaccharides (IMO), which consist of a combination of isomaltotriose, isomaltose, panose, and isomaltotetraose, on aquatic animals. No clear statement was recorded regarding immune responses [212].
3.3.3. Chitosan
Chitosan is a linear polysaccharide component of β-(1–4) linked D-glucosamine and is synthesized through alkaline deacetylation. It is a major component of arthropods' exoskeleton like shrimps, crabs, insects, and lobsters. In aquaculture, chitosan induces immunostimulation effects in various species namely rainbow trout [213], olive flounder (Paralichthys olivaceus) [214], and salmonids [215]. The administration of chitosan in the nutrient feed of C. carpio koi for 75 days resulted in significant effects such as enhanced immune response, improved lipid metabolism, enhanced growth performance, and modulated intestine microbiota, thereby protecting the fish from pathogen invasion [216].
3.3.4. Inulin
The prebiotic component inulin, a soluble plant fiber, is used in the fish diet and plays a crucial role in enhancing the immune system in both mammals and fish. In aquaculture, tinulin finds significant use by activating the beneficial bacteria, inhibiting the pathogens, and boosting immune system activity [217]. Inulin has the potential to mitigate inflammation induced by a high-carbohydrate diet, thereby enhancing pathogen resistance in fish. Additionally, supplementing with inulin led to changes in gut microbiota composition and their metabolites. These alterations likely contribute to alleviating the metabolic syndromes induced by a high-carbohydrate diet in fish. [218].
4. Postbiotics
4.1. Concept, definition and major components of postbiotics
The use of live microorganisms as probiotics may have potential concerns associated with the gene resistance acquisition and translocation, and depends on their viability [221]. Likewise, it has been recognized that non-viable microorganisms as well as their components and metabolites can provide positive effects on health leading to the apparition of postbiotic concept [222]. Postbiotics are defined by consensus panels as a preparation of inactivated microorganisms and/or their components (cell fragments, cell walls, metabolites) that have beneficial health effects on host [223]. This definition does not include purified metabolites in the absence of cells and cells components. One defined postbiotics as dead microbes and/or cell structures or metabolites that are produced by bacterial lysis or secreted during fermentation process [224].
Postbiotics include inactivated probiotics called paraprobiotics, metabolites like short chain fatty acids (SCFAs), vitamins, and phenolic acids, secreted proteins and peptides, functional proteins and enzymes, cell wall components like LTAs and peptidoglycan (PG)-derived muropeptides, secreted and extracellular polysaccharides (EPS), cell lysates, cellular components (glycans, enzymes), microbial fraction, and surface molecules such as pili [225,226].
Figure 3 outlines main postbiotics components.
4.2. Action modes and applications of postbiotics in aquaculture
The action mechanisms of postbiotics are still unclear but it has been generally assumed that they are similar to those of live probiotics [227]. Three main mechanisms are involved in postbiotics action modes.
4.2.1. Immunomodulation by microbial compounds
Postbiotics act on the immune system through two signaling pathways, namely nuclear factor -kB (NF-kB) and mitogen-activated protein kinases (MAPK), which are involved in the immune and inflammatory responses. Postbiotics stimulate the innate and adaptive immune systems via external Toll-like receptors (TLRs) that recognize associated pathogens and bind to specific patterns such as LTAs and PGs. They also interact with intracellular nucleotide-like receptors (NLRs) and nucleotide-binding and oligomerization domain (NOD)-like receptors, which can bind to molecules like lipopolysaccharides (LPS), PG, and flagellin, thereby activating innate immune signaling pathways [224,226]. The role of PG recognition proteins in innate immune responses against pathogens has been demonstrated in fish [228,229]. PG-derived muropeptides from bacterial cell walls have been shown to boost the immune system of fish [230] and shrimp [231]. For instance, muropeptides isolated from Bifidobacterium thermophilum have been proven to enhance shrimp immunity by increasing phagocytic activity or activating immune genes [231,232].
Additionally, postbiotics can enhance epithelial barriers protection via cell surface molecules such as pili and secreted protein P40 [233]. For example, the role of Lactobacillus pentosus surface protein on immune response has been demonstrated in shrimp (L. vannamei) infected with Vibrio parahaemolyticus [234].
4.2.2. Antagonizing pathogens by antimicrobial activities
Postbiotics exhibit antimicrobial activities against various pathogens due to the presence of metabolites like peptides and organic acids [235]. Bacteriocin JFP2 isolated from B. amyloliquefaciens exhibits antimicrobial activity against fish pathogen A. hydrophila [236]. The dietary addition of postbiotic containing LAB (strain Lactobacillus) has been reported to protect rainbow trout (O. mykiss) against the bacterial fish pathogen L. garvieae after 30 days feeding [237].
4.2.3. Inhibition of oxidation by antioxidant enzyme systems and metabolites
Various postbiotics obtained from LABs have been shown to exhibit antioxidant activity mainly attributed to phenolic compounds [238]. L. plantarum postbiotics have been documented to enhance antioxidant activity in animals [239]. In aquaculture applications, the overall antioxidant status of shrimp fed with diets supplemented with C. butyricum postbiotics was improved regarding the increase of alkaline phosphatase, acid phosphatase, total nitric oxide synthase, lysozyme, peroxidase, superoxide dismutase activities, total antioxidant capacity, and phenoloxidase content in the serum [240].
In aquaculture, postbiotics have been used as growth promoters instead of antibiotics, as immune system stimulation and as disease control [197,233,241]. Recently, the potential application of postbiotics in aquaculture water quality to modulate bacterioplankton communities and to influence nutrient cycling and bacterial pathogen abundance has been reported [242]. Figure 4 illustrates potential applications of probiotics in aquaculture. Table 3 shows some recent potential applications of postbiotics in aquaculture.
5. Synbiotics
Synbiotics refer to dietary additives that blend probiotics and prebiotics in a synergistic combination, thereby enhancing their beneficial effects. When either dietary additives or supplements are used, the resulting positive effects typically follow one of three patterns: ingredient effects, synergism, or potentiation. Supplementation outcomes occur when the combined effects of both additives used together approximate the sum of the effects of the individual supplements. In the case of synergism, the amalgamated result of the two products is significantly greater than the sum of the effects of each factor administered alone. The term potentiation is used differently; some pharmacologists interchange it with synergism to describe a result that is better than that of a supplement alone, while others use it to describe an outcome that is only present when both substances are used simultaneously [248,249].
5.1. Possible modes of action of synbiotics in aquaculture
5.1.1. Synbiotics enhance digestive enzyme and growth performance
Dietary administration of synbiotics is helpful in enhancing the digestive enzyme action of fish, allowing the host to degrade more nutrients. This dietary method increases digestive action and likely enhances the weight gain rate and/or feed efficiency [250]. Nutrient diet supplementation with a mixture of probiotics and monosaccharides enhances feed efficiency and overall health in carp. However, limited data is available in aquaculture regarding the function of nutrient diet supplementation of synbiotics in carp [23]. Nutrient diet administration of synbiotics enhances the lymphocyte and white blood cells in carp [251]. Synbiotics (IMBO), a combination of probiotics (E. faecium) and prebiotics (FOS), have been used to enhance the growth performance, survival rate, and digestive enzyme function of common carp fingerlings [252]. Dietary supplementation of FOS, MOS, and B. clausii can improve growth performance and health benefits of the Japanese flounder more than the control diet [168]. Dietary supplementation of FOS and 1.35×107 CFU g-1 B. subtilis (single or mixed) increases the specific growth rate (SGR) and feed efficiency ratio (FER) compared to the groups without B. subtilis additives in juvenile large yellow croaker (Larimichthys crocea) [211]. Figure 5 illustrates the possible modes of action of synbiotics in aquaculture.
5.1.2. Synbiotics improve immune response and disease resistance
An amalgamation of probiotics and prebiotics feed supplements is mainly helpful to enhance the survival of beneficial organisms, as the presence of prebiotics protects well-organized fermentation. Finally, this rewards the host with a suitable approach [253]. The nutritional additives of probiotics and prebiotics (MOS, FOS, and inulin) enhance the fish immune system via GIT [23,254,255]. A synbiotic composed of Pediococcus acidilactici and galactooligosaccharides improved immune parameters and antagonistic activity against S. iniae when administered to rainbow trout fingerlings for 8 weeks [256]. The combination of probiotic Bacillus sp. and 0.2% of prebiotic isomaltooligosaccharide has been used to improve immune functions in shrimp (Penaeus japonicas) against V. alginolyticus infection [257]. In addition, the blended use of Bacillus and molasses improved the microbial population, enhanced the development of the probiotic community, and inhibitory activity against pathogens in pacific white shrimp [258]. The effectiveness of synbiotic treatment in conditions of defense against infectious factors could be evaluated by a confrontation examination due to its regulatory power over harmful microbes and its capability to resist infections [259]. The functional feed additives of synbiotics in aquatic animals are summarised in Table 4.
6. Limitations of the use of biotic agents in aquaculture
The use of biotic agents in aquaculture instead of antibiotics has recently gained significant interests [265]. Probiotics have been shown to be effective in promoting growth, increasing immunity, and improving resistance to infections in aquatic animals [266]. The major limitation of their use comes from the problem of possible gene resistance acquisition and translocation, as well as the question of their viability and/or ability to colonize the fish gut [221]. The use of multi-strain probiotics increases the possibility of strain survival rates and therefore improves the beneficial effects on growth, immunity and infection resistance of aquatic animals [128]. Postbiotics present an advantage over probiotics because they do not have viability problems and are less susceptible to environmental conditions [221,267]. Additionally, they generally have a complex composition made up of several compounds that play multiple roles and provide numerous beneficial effects on aquatic animals. However, their use to manage infectious disease is still in its early stages [235].
Prebiotics, as inert biotic agents, are relatively safe and cost-effective alternative to probiotics. Several studies on their immunostimulant properties and growth promotion in fish and shellfish have shown some evidence for their interest in aquaculture [268]. Nevertheless, studies on the optimal dose should be carried out, as inadequate dose may lead to detrimental effects on aquatic animals [182,208]. Synbiotics improve the colonization of microorganisms in the intestines and are generally more effective than probiotics or prebiotics alone [267]. For example, Nile tilapia (O. niloticus) fed with synbiotic showed the highest increase in the specific growth rate compared to the group fed with probiotics or prebiotics alone [251,269]. Extensive studies are still needed to specify the role of prebiotics, probiotics, postbiotics, and synbiotics in growth performance, intestinal health, and immune aspects with a focus on the mechanisms underlying the synbiotic diet in aquatic animals against various pathogens. The mode of administration and dose of the biotic agents are also important and have certainly an impact on their effectiveness [270].
7. Concluding remarks and future perspectives
Aquaculture is one of the fastest-growing food manufacturing sectors in the world. Disease outbreaks pose significant problems for aquaculture, impacting the financial status and economic development of people in the Asia-Pacific region as a whole. Globally, most researchers have analyzed and concluded that functional feeds (probiotics, prebiotics, and synbiotics) or immunostimulants are potential alternatives for the development of the modern aquaculture industry. These methods are particularly useful in enhancing aquaculture production as they offer numerous health benefits, such as the modulation of gut microbiota and immune systems, enhancement of disease resistance and survival rates, improvement of growth performance, and efficient feed utilization.
While several study reports are available regarding probiotics, prebiotics, and synbiotics, these are not sufficient to drive the development of the aquaculture industry. Extensive studies are needed at different levels, corresponding to the role of prebiotics, probiotics, and synbiotics in growth performance, intestinal health, and immune aspects. Additionally, there is a need for a deeper understanding of the mechanisms underlying synbiotic diets in aquatic animals against various pathogens.
This review concludes that functional feed additives, such as probiotics, prebiotics, and synbiotics, are utilized to enhance the immune system, disease resistance, gut microbiota survival rates, growth performance, and feed utilization in numerous commercially available fish species. However, data from zebrafish modeling studies are very limited. In the future, not only probiotics, prebiotics, and synbiotics but also various other beneficial substances, such as functional amino acids, fatty acids, enzymes, organic acids, and herbs, can be added or substituted to enhance the innate immune system, disease resistance, and growth and survival rates in the zebrafish model. These additions can also improve feed quality levels in aquaculture. Furthermore, postbiotics, which are components or metabolites from dead probiotic microorganisms, show promise as functional feed components."
Author Contributions
Conceptualization, S.V. and H.L.R.; writing—original draft preparation, S.V.; writing—review and editing, H.L.R., H.N.R, and H-TN.; supervision, H.L.R. and Y-ZS. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Acknowledgments
We wish to acknowledge the national natural science foundation of China (Grant No. 32072990), Xiamen Marine and Fisheries Development Fund (Grant No. 19CZP018HJ04), Industry-University Cooperation Project of Fujian Province (Grant No. 2018N5011) for supporting this research work.
Conflict of interest
The authors have declared no conflict of interest.
References
- Moyo, N.A.; Rapatsa, M.M. A Review of the Factors Affecting Tilapia Aquaculture Production in Southern Africa. Aquaculture 2021, 535, 736386. [Google Scholar] [CrossRef]
- Gephart, J.A.; Golden, C.D.; Asche, F.; Belton, B.; Brugere, C.; Froehlich, H.E.; Fry, J.P.; Halpern, B.S.; Hicks, C.C.; Jones, R.C. Scenarios for Global Aquaculture and Its Role in Human Nutrition. Rev. Fish. Sci. Aquac. 2020, 29, 122–138. [Google Scholar] [CrossRef]
- FAO/WHO The State of World Fisheries and Aquaculture 2020: Sustainability in Action; Food and Agriculture Organization of the United Nations, 2020; ISBN 92-5-132692-4.
- Baticados, M.C.L.; Paclibare, J.O. The Use of Chemotherapeutic Agents in Aquaculture in the Philippines. In Proceedings of the Diseases in Asian Aquaculture I. Proceedings of the First Symposium on Diseases in Asian Aquaculture, 26-29 November 1990, Bali, Indonesia; Asian Fisheries Society, Fish Health Section, 1992; pp. 531–546.
- Sadat Hoseini Madani, N.; Adorian, T.J.; Ghafari Farsani, H.; Hoseinifar, S.H. The Effects of Dietary Probiotic Bacilli (Bacillus Subtilis and Bacillus Licheniformis) on Growth Performance, Feed Efficiency, Body Composition and Immune Parameters of Whiteleg Shrimp (Litopenaeus Vannamei) Postlarvae. Aquac. Res. 2018, 49, 1926–1933. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2020, 29, 198–217. [Google Scholar] [CrossRef]
- Alfred, O.; Shaahu, A.; Orban, D.A.; Egwenomhe, M. An Overview on Understanding the Basic Concept of Fish Diseases in Aquaculture. IRE J. 2020, 4, 83–91. [Google Scholar]
- Essawi, T.; Srour, M. Screening of Some Palestinian Medicinal Plants for Antibacterial Activity. J. Ethnopharmacol. 2000, 70, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Zou, H.K.; Miandare, H.K.; Van Doan, H.; Romano, N.; Dadar, M. Enrichment of Common Carp (Cyprinus Carpio) Diet with Medlar (Mespilus Germanica) Leaf Extract: Effects on Skin Mucosal Immunity and Growth Performance. Fish Shellfish Immunol. 2017, 67, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Musthafa, M.S.; Asgari, S.M.; Kurian, A.; Elumalai, P.; Ali, A.R.J.; Paray, B.A.; Al-Sadoon, M.K. Protective Efficacy of Mucuna Pruriens (L.) Seed Meal Enriched Diet on Growth Performance, Innate Immunity, and Disease Resistance in Oreochromis Mossambicus against Aeromonas Hydrophila. Fish Shellfish Immunol. 2018, 75, 374–380. [Google Scholar] [CrossRef]
- Safety, S. FDA Needs to Improve Oversight of Imported Seafood and Better Leverage Limited Resources. U. S. Gov. Account. Off. 2011. [Google Scholar]
- Yilmaz, S.; Yilmaz, E.; Dawood, M.A.; Ringø, E.; Ahmadifar, E.; Abdel-Latif, H.M. Probiotics, Prebiotics, and Synbiotics Used to Control Vibriosis in Fish: A Review. Aquaculture 2022, 547, 737514. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in Fish Gastrointestinal Microbiota Research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef]
- Luna, G.M.; Quero, G.M.; Kokou, F.; Kormas, K. Time to Integrate Biotechnological Approaches into Fish Gut Microbiome Research. Curr. Opin. Biotechnol. 2022, 73, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Jia, P.-P.; Junaid, M.; Wen, P.-P.; Yang, Y.-F.; Li, W.-G.; Yang, X.-G.; Pei, D.-S. Role of Germ-Free Animal Models in Understanding Interactions of Gut Microbiota to Host and Environmental Health: A Special Reference to Zebrafish. Environ. Pollut. 2021, 279, 116925. [Google Scholar] [CrossRef]
- Bates, J.M.; Mittge, E.; Kuhlman, J.; Baden, K.N.; Cheesman, S.E.; Guillemin, K. Distinct Signals from the Microbiota Promote Different Aspects of Zebrafish Gut Differentiation. Dev. Biol. 2006, 297, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Rawls, J.F. Microbial Influences on Gut Development and Gut-Brain Communication. Development 2021, 148, dev194936. [Google Scholar] [CrossRef]
- Koch, B.E.; Yang, S.; Lamers, G.; Stougaard, J.; Spaink, H.P. Intestinal Microbiome Adjusts the Innate Immune Setpoint during Colonization through Negative Regulation of MyD88. Nat. Commun. 2018, 9, 4099. [Google Scholar] [CrossRef]
- Tran, N.T.; Yang, W.; Nguyen, X.T.; Zhang, M.; Ma, H.; Zheng, H.; Zhang, Y.; Chan, K.-G.; Li, S. Application of Heat-Killed Probiotics in Aquaculture. Aquaculture 2022, 548, 737700. [Google Scholar] [CrossRef]
- Yang, H.-L.; Sun, Y.-Z.; Hu, X.; Ye, J.; Lu, K.-L.; Hu, L.-H.; Zhang, J.-J. Bacillus Pumilus SE5 Originated PG and LTA Tuned the Intestinal TLRs/MyD88 Signaling and Microbiota in Grouper (Epinephelus Coioides). Fish Shellfish Immunol. 2019, 88, 266–271. [Google Scholar] [CrossRef]
- Van Doan, H. Bacillus Spp. in Aquaculture-Mechanisms and Applications: An Update View. Probiotic Bact. Postbiotic Metab. Role Anim. Hum. Health 2021, 1–59. [Google Scholar] [CrossRef]
- Kim, Y.-R.; Kim, E.-Y.; Choi, S.; Hossain, M.T.; Oh, R.-K.; Heo, W.-S.; Lee, J.-M.; Cho, Y.-C.; Kong, I.-S. Effect of a Probiotic Strain, Enterococcus Faecium, on the Immune Responses of Olive Flounder (Paralichthys Olivaceus). J. Microbiol. Biotechnol. 2012, 22, 526–529. [Google Scholar] [CrossRef]
- Mugwanya, M.; Dawood, M.A.; Kimera, F.; Sewilam, H. Updating the Role of Probiotics, Prebiotics, and Synbiotics for Tilapia Aquaculture as Leading Candidates for Food Sustainability: A Review. Probiotics Antimicrob. Proteins 2021, 1–28. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Shakouri, M.; Yousefi, S.; Van Doan, H.; Shafiei, S.; Yousefi, M.; Mazandarani, M.; Mozanzadeh, M.T.; Tulino, M.G.; Faggio, C. Humoral and Skin Mucosal Immune Parameters, Intestinal Immune Related Genes Expression and Antioxidant Defense in Rainbow Trout (Oncorhynchus Mykiss) Fed Olive (Olea Europea L.) Waste. Fish Shellfish Immunol. 2020, 100, 171–178. [Google Scholar] [CrossRef] [PubMed]
- da Silva Liebl, A.R.; Cáo, M.A.; dos Santos Nascimento, M.; Castro, P.D. da S.; Duncan, W.L.P.; Pantoja-Lima, J.; Aride, P.H.R.; Bussons, M.R.F.M.; Furuya, W.M.; Faggio, C. Dietary Lysine Requirements of Colossoma Macropomum (Cuvier, 1818) Based on Growth Performance, Hepatic and Intestinal Morphohistology and Hematology. Vet. Res. Commun. 2022, 1–17. [CrossRef]
- Mahboub, H.H.; Faggio, C.; Hendam, B.M.; Algharib, S.A.; Alkafafy, M.; Hashem, M.A.; Mahmoud, Y.K.; Khamis, T.; Abdel-Ghany, H.M.; Masoud, S.R. Immune-Antioxidant Trait, Aeromonas Veronii Resistance, Growth, Intestinal Architecture, and Splenic Cytokines Expression of Cyprinus Carpio Fed Prunus Armeniaca Kernel-Enriched Diets. Fish Shellfish Immunol. 2022, 124, 182–191. [Google Scholar] [CrossRef]
- Al-Shawi, S.G.; Dang, D.S.; Yousif, A.Y.; Al-Younis, Z.K.; Najm, T.A.; Matarneh, S.K. The Potential Use of Probiotics to Improve Animal Health, Efficiency, and Meat Quality: A Review. Agriculture 2020, 10, 452. [Google Scholar] [CrossRef]
- Adel, M.; Dawood, M.A. Probiotics Application: Implications for Sustainable Aquaculture. Probiotic Bact. Postbiotic Metab. Role Anim. Hum. Health 2021, 191–219. [Google Scholar]
- FAO/WHO, E.C. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; World Health Organization Córdoba, 2001.
- Guardiola, F.A.; Porcino, C.; Cerezuela, R.; Cuesta, A.; Faggio, C.; Esteban, M.A. Impact of Date Palm Fruits Extracts and Probiotic Enriched Diet on Antioxidant Status, Innate Immune Response and Immune-Related Gene Expression of European Seabass (Dicentrarchus Labrax). Fish Shellfish Immunol. 2016, 52, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Van Doan, H.; Hoseinifar, S.H.; Ringø, E.; Ángeles Esteban, M.; Dadar, M.; Dawood, M.A.; Faggio, C. Host-Associated Probiotics: A Key Factor in Sustainable Aquaculture. Rev. Fish. Sci. Aquac. 2020, 28, 16–42. [Google Scholar] [CrossRef]
- Morshedi, V.; Bojarski, B.; Hamedi, S.; Torahi, H.; Hashemi, G.; Faggio, C. Effects of Dietary Bovine Lactoferrin on Growth Performance and Immuno-Physiological Responses of Asian Sea Bass (Lates Calcarifer) Fingerlings. Probiotics Antimicrob. Proteins 2021, 13, 1790–1797. [Google Scholar] [CrossRef]
- Mirbakhsh, M.; Ghaednia, B.; Zorriehzahra, M.J.; Esmaeili, F.; Faggio, C. Dietary Mixed and Sprayed Probiotic Improves Growth Performance and Digestive Enzymes of Juvenile Whiteleg Shrimp (Litopenaeus Vannamei, Boone, 1931). J. Appl. Aquac. 2022, 1–14. [Google Scholar] [CrossRef]
- Misra, S.; Pandey, P.; Mishra, H.N. Novel Approaches for Co-Encapsulation of Probiotic Bacteria with Bioactive Compounds, Their Health Benefits and Functional Food Product Development: A Review. Trends Food Sci. Technol. 2021, 109, 340–351. [Google Scholar] [CrossRef]
- El-Kady, A.A.; Magouz, F.I.; Mahmoud, S.A.; Abdel-Rahim, M.M. The Effects of Some Commercial Probiotics as Water Additive on Water Quality, Fish Performance, Blood Biochemical Parameters, Expression of Growth and Immune-Related Genes, and Histology of Nile Tilapia (Oreochromis Niloticus). Aquaculture 2022, 546, 737249. [Google Scholar] [CrossRef]
- Vijayaram, S.; Kannan, S. Probiotics: The Marvelous Factor and Health Benefits. Biomed. Biotechnol. Res. J. BBRJ 2018, 2, 1. [Google Scholar] [CrossRef]
- Cıl, G.I.; Bulut, G.; Budak, D.; Camkerten, G.; Camkerten, I. Probiotics and Functional Feed. In Probiotics, the Natural Microbiota in Living Organisms; CRC Press, 2021; pp. 315–342 ISBN 1-351-02754-9.
- Foysal, M.J.; Alam, M.; Kawser, A.R.; Hasan, F.; Rahman, M.M.; Tay, C.-Y.; Prodhan, M.S.H.; Gupta, S.K. Meta-Omics Technologies Reveals Beneficiary Effects of Lactobacillus Plantarum as Dietary Supplements on Gut Microbiota, Immune Response and Disease Resistance of Nile Tilapia (Oreochromis Niloticus). Aquaculture 2020, 520, 734974. [Google Scholar] [CrossRef]
- Zaineldin, A.I.; Hegazi, S.; Koshio, S.; Ishikawa, M.; Dawood, M.A.; Dossou, S.; Yukun, Z.; Mzengereza, K. Singular Effects of Bacillus Subtilis C-3102 or Saccharomyces Cerevisiae Type 1 on the Growth, Gut Morphology, Immunity, and Stress Resistance of Red Sea Bream (Pagrus Major). 2021. [CrossRef]
- Kong, Y.; Gao, C.; Du, X.; Zhao, J.; Li, M.; Shan, X.; Wang, G. Effects of Single or Conjoint Administration of Lactic Acid Bacteria as Potential Probiotics on Growth, Immune Response and Disease Resistance of Snakehead Fish (Channa Argus). Fish Shellfish Immunol. 2020, 102, 412–421. [Google Scholar] [CrossRef]
- Kord, M.I.; Maulu, S.; Srour, T.M.; Omar, E.A.; Farag, A.A.; Nour, A.A.M.; Hasimuna, O.J.; Abdel-Tawwab, M.; Khalil, H.S. Impacts of Water Additives on Water Quality, Production Efficiency, Intestinal Morphology, Gut Microbiota, and Immunological Responses of Nile Tilapia Fingerlings under a Zero-Water-Exchange System. Aquaculture 2022, 547, 737503. [Google Scholar] [CrossRef]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C. da; Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J. Oxidative Stress Tolerance and Antioxidant Capacity of Lactic Acid Bacteria as Probiotic: A Systematic Review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.K.; Dicks, L.M.; Popov, I.V.; Karaseva, A.; Ermakov, A.M.; Suvorov, A.; Tagg, J.R.; Weeks, R.; Chikindas, M.L. Probiotics at War against Viruses: What Is Missing from the Picture? Front. Microbiol. 2020, 11, 1877. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, K.; Samayanpaulraj, V.; Narayanadoss, V.; Uthandakalaipandian, R. Isolation of Lactic Acid Bacteria from Intestine of Freshwater Fishes and Elucidation of Probiotic Potential for Aquaculture Application. Probiotics Antimicrob. Proteins 2021, 13, 1598–1610. [Google Scholar] [CrossRef] [PubMed]
- Khalid, F.; Khalid, A.; Fu, Y.; Hu, Q.; Zheng, Y.; Khan, S.; Wang, Z. Potential of Bacillus Velezensis as a Probiotic in Animal Feed: A Review. J. Microbiol. 2021, 59, 627–633. [Google Scholar] [CrossRef]
- Vallesi, A.; Pucciarelli, S.; Buonanno, F.; Fontana, A.; Mangiagalli, M. Bioactive Molecules from Protists: Perspectives in Biotechnology. Eur. J. Protistol. 2020, 75, 125720. [Google Scholar] [CrossRef]
- Butt, U.D.; Lin, N.; Akhter, N.; Siddiqui, T.; Li, S.; Wu, B. Overview of the Latest Developments in the Role of Probiotics, Prebiotics and Synbiotics in Shrimp Aquaculture. Fish Shellfish Immunol. 2021, 114, 263–281. [Google Scholar] [CrossRef]
- Labba, I.-C.M.; Andlid, T.; Lindgren, Å.; Sandberg, A.-S.; Sjöberg, F. Isolation, Identification, and Selection of Strains as Candidate Probiotics and Starters for Fermentation of Swedish Legumes. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef]
- Rajyalakshmi, K.; Babu, M.K.; Shabana, S.; Satya, A.K. Identification and Screening of Probiotics as a Biocontrol Agent against Pathogenic Vibriosis in Shrimp Aquaculture. Ann. Romanian Soc. Cell Biol. 2021, 25, 12292–12305. [Google Scholar]
- Mingmongkolchai, S.; Panbangred, W. Bacillus Probiotics: An Alternative to Antibiotics for Livestock Production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Ringø, E. Probiotics in Shellfish Aquaculture. Aquac. Fish. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, E.; Lu, M.; Xie, J. Effects of Probiotic Supplementation on Gut Microbiota as Well as Metabolite Profiles within Nile Tilapia, Oreochromis Niloticus. Aquaculture 2020, 527, 735428. [Google Scholar] [CrossRef]
- Yukgehnaish, K.; Kumar, P.; Sivachandran, P.; Marimuthu, K.; Arshad, A.; Paray, B.A.; Arockiaraj, J. Gut Microbiota Metagenomics in Aquaculture: Factors Influencing Gut Microbiome and Its Physiological Role in Fish. Rev. Aquac. 2020, 12, 1903–1927. [Google Scholar] [CrossRef]
- Wu, G. Nutrition and Metabolism: Foundations for Animal Growth, Development, Reproduction, and Health. Recent Adv. Anim. Nutr. Metab. 2022, 1–24. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial Effects on Host Energy Metabolism of Short-Chain Fatty Acids and Vitamins Produced by Commensal and Probiotic Bacteria. Microb. Cell Factories 2017, 16, 1–10. [Google Scholar] [CrossRef]
- Morais, T.; Inácio, A.; Coutinho, T.; Ministro, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed Potential in the Animal Feed: A Review. J. Mar. Sci. Eng. 2020, 8, 559. [Google Scholar] [CrossRef]
- Hardy, R.W.; Kaushik, S.J.; Mai, K.; Bai, S.C. Fish Nutrition—History and Perspectives. In Fish Nutrition; Elsevier, 2022; pp. 1–16.
- Uma, A.; Subash, P.; Abraham, T.J. Importance of Gut Microbiota in Fish–A Review. Indian J Anim Hlth 2020, 59, 181–194. [Google Scholar] [CrossRef]
- Singh, S.K.; Bhandari, M.P.; Shrestha, S.; Koirala, U.; Gurung, G.B. Supplementation of Commercial Probiotics in Feed for Growth and Survival of Rainbow Trout (Oncorhynchus Mykiss). In Proceedings of the thNational Workshop on Livestock and Fisheries Research in Nepal; 2021; Vol. 3, p. 275.
- Chen, J.; Sun, D.; Cui, H.; Rao, C.; Li, L.; Guo, S.; Yang, S.; Zhang, Y.; Cao, X. Toxic Effects of Carbon Quantum Dots on the Gut–Liver Axis and Gut Microbiota in the Common Carp Cyprinus Carpio. Environ. Sci. Nano 2022, 9, 173–188. [Google Scholar] [CrossRef]
- Phianphak, W.; Rengpipat, S.; Piyatiratitivorakul, S.; Menasveta, P. Probiotic Use of Lactobacillus Spp. for Black Tiger Shrimp, Penaeus Monodon. J Sci Res Chula Univ 1999, 24, 41–58. [Google Scholar]
- Zhang, W.; Li, C.; Guo, M. Use of Ecofriendly Alternatives for the Control of Bacterial Infection in Aquaculture of Sea Cucumber Apostichopus Japonicus. Aquaculture 2021, 545, 737185. [Google Scholar] [CrossRef]
- Tuan, T.N.; Duc, P.M.; Hatai, K. Overview of the Use of Probiotics in Aquaculture. Int. J. Res. Fish. Aquac. 2013, 3, 89–97. [Google Scholar]
- El-Saadony, M.T.; Alagawany, M.; Patra, A.K.; Kar, I.; Tiwari, R.; Dawood, M.A.; Dhama, K.; Abdel-Latif, H.M. The Functionality of Probiotics in Aquaculture: An Overview. Fish Shellfish Immunol. 2021, 117, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Yassir, R.Y.; Adel, M.E.; Azze, A. Use of Probiotic Bacteria as Growth Promoters, Antibacterial and the Effect on Physiological Parameters of Orechromis Niloticus. J Fish Dis 2002, 22, 633–642. [Google Scholar]
- Swain, S.; Hauzoukim, S.K.G.; Das, S.K.; Roy, A. Application of Probiotics in Aquaculture. 2021.
- Franca, F.M.; Danielle de Carla, D.; Teixeira, P.C.; Marcantonio, A.S.; de Stefani, M.V.; Antonucci, A.; Da Rocha, G.; RANZANI-PAIVA, M.J.T.; Ferreira, C.M. Efeito Do Probiótico Bacillus Subtilis No Crescimento, Sobrevivência e Fisiologia de Rãs-Touro (Rana Catesbeiana). Bol. Inst. Pesca 2008, 34, 403–412. [Google Scholar]
- Wang, Z.; Yang, M.; Wang, L.; Lu, K.; Song, K.; Zhang, C. Bacillus Subtilis LCBS1 Supplementation and Replacement of Fish Meal with Fermented Soybean Meal in Bullfrog (Lithobates Catesbeianus) Diets: Effects on Growth Performance, Feed Digestibility and Gut Health. Aquaculture 2021, 545, 737217. [Google Scholar] [CrossRef]
- Assan, D.; Kuebutornye, F.K.A.; Hlordzi, V.; Chen, H.; Mraz, J.; Mustapha, U.F.; Abarike, E.D. Effects of Probiotics on Digestive Enzymes of Fish (Finfish and Shellfish); Status and Prospects: A Mini Review. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2022, 257, 110653. [Google Scholar] [CrossRef]
- Bahnasawy, M.H.; El-Ghobashy, A.E.; El-Ebiary, E.-S.H.; Helal, A.M.; El-Sisy, D.M. Effect of Probiotic on Water Quality, Growth Performance and Body Composition of Nile Tilapia (Oreochromis Niloticus). Int J Fish Aquat. Stud 2020, 8, 86–91. [Google Scholar]
- Carnevali, O.; Sun, Y.-Z.; Merrifield, D.L.; Zhou, Z.; Picchietti, S. Probiotic Applications in Temperate and Warm Water Fish Species. Aquac. Nutr. Gut Health Probiotics Prebiotics 2014, 253–289. [Google Scholar] [CrossRef]
- Loh, J.Y.; Chan, H.K.; Yam, H.C.; In, L.L.A.; Lim, C.S.Y. An Overview of the Immunomodulatory Effects Exerted by Probiotics and Prebiotics in Grouper Fish. Aquac. Int. 2020, 28, 729–750. [Google Scholar] [CrossRef]
- Gao, X.-Y.; Liu, Y.; Miao, L.-L.; Li, E.-W.; Hou, T.-T.; Liu, Z.-P. Mechanism of Anti-Vibrio Activity of Marine Probiotic Strain Bacillus Pumilus H2, and Characterization of the Active Substance. AMB Express 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kuebutornye, F.K.; Abarike, E.D.; Lu, Y.; Hlordzi, V.; Sakyi, M.E.; Afriyie, G.; Wang, Z.; Li, Y.; Xie, C.X. Mechanisms and the Role of Probiotic Bacillus in Mitigating Fish Pathogens in Aquaculture. Fish Physiol. Biochem. 2020, 46, 819–841. [Google Scholar] [CrossRef]
- Emam, A.M.; Dunlap, C.A. Genomic and Phenotypic Characterization of Bacillus Velezensis AMB-Y1; a Potential Probiotic to Control Pathogens in Aquaculture. Antonie Van Leeuwenhoek 2020, 113, 2041–2052. [Google Scholar] [CrossRef]
- Xu, H.-M.; Rong, Y.-J.; Zhao, M.-X.; Song, B.; Chi, Z.-M. Antibacterial Activity of the Lipopetides Produced by Bacillus Amyloliquefaciens M1 against Multidrug-Resistant Vibrio Spp. Isolated from Diseased Marine Animals. Appl. Microbiol. Biotechnol. 2014, 98, 127–136. [Google Scholar] [CrossRef]
- Chau, K.M.; Van, T.T.H.; Quyen, D.V.; Le, H.D.; Phan, T.H.T.; Ngo, N.D.T.; Vo, T.D.T.; Dinh, T.T.; Le, H.T.; Khanh, H.H.N. Molecular Identification and Characterization of Probiotic Bacillus Species with the Ability to Control Vibrio Spp. in Wild Fish Intestines and Sponges from the Vietnam Sea. Microorganisms 2021, 9, 1927. [Google Scholar] [CrossRef]
- Yin, Z.; Liu, Q.; Liu, Y.; Gao, S.; He, Y.; Yao, C.; Huang, W.; Gong, Y.; Mai, K.; Ai, Q. Early Life Intervention Using Probiotic Clostridium Butyricum Improves Intestinal Development, Immune Response, and Gut Microbiota in Large Yellow Croaker (Larimichthys Crocea) Larvae. Front. Immunol. 2021, 12, 640767. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B.; Liu, C.; Zhou, H.; Wang, X.; Mai, K.; He, G. Effects of Dietary Raw or Enterococcus Faecium Fermented Soybean Meal on Growth, Antioxidant Status, Intestinal Microbiota, Morphology, and Inflammatory Responses in Turbot (Scophthalmus Maximus L.). Fish Shellfish Immunol. 2020, 100, 261–271. [Google Scholar] [CrossRef]
- Kamei, Y.; Yoshimizu, M.; Ezura, Y.; Kimura, T. Screening of Bacteria with Antiviral Activity from Fresh Water Salmonid Hatcheries. Microbiol. Immunol. 1988, 32, 67–73. [Google Scholar] [CrossRef]
- Hasan, K.N.; Banerjee, G. Recent Studies on Probiotics as Beneficial Mediator in Aquaculture: A Review. J. Basic Appl. Zool. 2020, 81, 1–16. [Google Scholar] [CrossRef]
- Maeda, M.; Nogami, K.; Kanematsu, M.; Hirayama, K. The Concept of Biological Control Methods in Aquaculture. Hydrobiologia 1997, 358, 285–290. [Google Scholar] [CrossRef]
- Mondal, H.; Chandrasekaran, N.; Mukherjee, A.; Thomas, J. Viral Infections in Cultured Fish and Shrimps: Current Status and Treatment Methods. Aquac. Int. 2022, 1–36. [Google Scholar] [CrossRef]
- de Andrade Belo, M.A.; Charlie-Silva, I. Teleost Fish as an Experimental Model for Vaccine Development. Vaccine Des. Methods Protoc. Vol. 2 Vaccines Vet. Dis. 2022, 175–194. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S. Recent Advances in the Role of Probiotics and Prebiotics in Carp Aquaculture: A Review. Aquaculture 2016, 454, 243–251. [Google Scholar] [CrossRef]
- Akbari, H.; Shekrabi, S.P.H.; Soltani, M.; Mehrgan, M.S. Effects of Potential Probiotic Enterococcus Casseliflavus (EC-001) on Growth Performance, Immunity, and Resistance to Aeromonas Hydrophila Infection in Common Carp (Cyprinus Carpio). Probiotics Antimicrob. Proteins 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rachmawati, R.A.; Mulyani, Y.; Rochima, E.; Grandiosa, R. The Effect of Induction of Bacteria Bacillus Subtilis in Feed on the Immune System of Carp (Cyprinus Carpio Linnaeus, 1758). World Sci. News 2021, 160, 203–216. [Google Scholar]
- Shah, S.; Chesti, A.; Rather, M.; Manzoor, S.; Malik, R.; Khan, J. Effect of Probiotic (Bacillus Subtilis) on the Immune System of Fingerlings of Grass Carp, Ctenopharyngodon Idella. Pharma Innov. J. 2021, 10, 769–772. [Google Scholar]
- Yan, Y.-Y.; Xia, H.-Q.; Yang, H.-L.; Hoseinifar, S.H.; Sun, Y.-Z. Effects of Dietary Live or Heat-inactivated Autochthonous Bacillus Pumilus SE 5 on Growth Performance, Immune Responses and Immune Gene Expression in Grouper Epinephelus Coioides. Aquac. Nutr. 2016, 22, 698–707. [Google Scholar] [CrossRef]
- Liu, Z.-Y.; Yang, H.-L.; Hu, L.-H.; Yang, W.; Ai, C.-X.; Sun, Y.-Z. Dose-Dependent Effects of Histamine on Growth, Immunity and Intestinal Health in Juvenile Grouper (Epinephelus Coioides). Front. Mar. Sci. 2021, 8, 685720. [Google Scholar] [CrossRef]
- Yang, H.-L.; Xia, H.-Q.; Ye, Y.-D.; Zou, W.-C.; Sun, Y.-Z. Probiotic Bacillus Pumilus SE5 Shapes the Intestinal Microbiota and Mucosal Immunity in Grouper Epinephelus Coioides. Dis. Aquat. Organ. 2014, 111, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-L.; Hu, X.; Ye, J.-D.; Seerengaraj, V.; Yang, W.; Ai, C.-X.; Sun, Y.-Z. Cell Wall Components of Bacillus Pumilus SE5 Improved the Growth, Digestive and Immunity of Grouper (Epinephelus Coioides). Curr. Chin. Sci. 2021, 1, 231–239. [Google Scholar] [CrossRef]
- Boo, A.; Amaro, R.L.; Stan, G.-B. Quorum Sensing in Synthetic Biology: A Review. Curr. Opin. Syst. Biol. 2021, 28, 100378. [Google Scholar] [CrossRef]
- Saeki, E.K.; Kobayashi, R.K.T.; Nakazato, G. Quorum Sensing System: Target to Control the Spread of Bacterial Infections. Microb. Pathog. 2020, 142, 104068. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T.; Boon, N.; Bossier, P.; Verstraete, W. Disruption of Bacterial Quorum Sensing: An Unexplored Strategy to Fight Infections in Aquaculture. Aquaculture 2004, 240, 69–88. [Google Scholar] [CrossRef]
- Alexpandi, R.; Abirami, G.; Satish, L.; Swasthikka, R.P.; Krishnaveni, N.; Jayakumar, R.; Pandian, S.K.; Ravi, A.V. Tocopherol and Phytol Possess Anti-Quorum Sensing Mediated Anti-Infective Behavior against Vibrio Campbellii in Aquaculture: An in Vitro and in Vivo Study. Microb. Pathog. 2021, 161, 105221. [Google Scholar] [CrossRef]
- Samrot, A.V.; Abubakar Mohamed, A.; Faradjeva, E.; Si Jie, L.; Hooi Sze, C.; Arif, A.; Chuan Sean, T.; Norbert Michael, E.; Yeok Mun, C.; Xiao Qi, N. Mechanisms and Impact of Biofilms and Targeting of Biofilms Using Bioactive Compounds—A Review. Medicina (Mex.) 2021, 57, 839. [Google Scholar] [CrossRef]
- Chu, W.; Zhou, S.; Zhu, W.; Zhuang, X. Quorum Quenching Bacteria Bacillus Sp. QSI-1 Protect Zebrafish (Danio Rerio) from Aeromonas Hydrophila Infection. Sci. Rep. 2014, 4, 1–6. [Google Scholar] [CrossRef]
- Mamun, M.A.A.; Nasren, S.; Abhiman, P.B.; Rathore, S.S.; Sowndarya, N.S.; Ramesh, K.S.; Shankar, K.M. Investigation of Production, Formation and Characterization of Biofilm Cells of Aeromonas Hydrophila for Oral Vaccination of Fish. J Exp Zool India 2019, 22, 1115–1123. [Google Scholar]
- Hoseinifar, S.H.; Sun, Y.-Z.; Wang, A.; Zhou, Z. Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Ran, C.; Wang, Y.; Zhang, Z.; Ding, Q.; Yang, Y.; Olsen, R.E.; Ringø, E.; Bindelle, J.; Zhou, Z. Use of Probiotics in Aquaculture of China—a Review of the Past Decade. Fish Shellfish Immunol. 2019, 86, 734–755. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhou, S.; Sarkodie, E.K.; Chu, W. The Effects of Bacillus Cereus QSI-1 on Intestinal Barrier Function and Mucosal Gene Transcription in Crucian Carp (Carassius Auratus Gibelio). Aquac. Rep. 2020, 17, 100356. [Google Scholar] [CrossRef]
- Mohapatra, S.; Chakraborty, T.; Kumar, V.; DeBoeck, G.; Mohanta, K.N. Aquaculture and Stress Management: A Review of Probiotic Intervention. J. Anim. Physiol. Anim. Nutr. 2013, 97, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Vianello, S.; Brazzoduro, L.; Dalla Valle, L.; Belvedere, P.; Colombo, L. Myostatin Expression during Development and Chronic Stress in Zebrafish (Danio Rerio). J. Endocrinol. 2003, 176, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Lutfi, E.; Basili, D.; Falcinelli, S.; Morillas, L.; Carnevali, O.; Capilla, E.; Navarro, I. The Probiotic Lactobacillus Rhamnosus Mimics the Dark-Driven Regulation of Appetite Markers and Melatonin Receptors’ Expression in Zebrafish (Danio Rerio) Larvae: Understanding the Role of the Gut Microbiome. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2021, 256, 110634. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, O.; de Vivo, L.; Sulpizio, R.; Gioacchini, G.; Olivotto, I.; Silvi, S.; Cresci, A. Growth Improvement by Probiotic in European Sea Bass Juveniles (Dicentrarchus Labrax, L.), with Particular Attention to IGF-1, Myostatin and Cortisol Gene Expression. Aquaculture 2006, 258, 430–438. [Google Scholar] [CrossRef]
- Castex, M.; Lemaire, P.; Wabete, N.; Chim, L. Effect of Dietary Probiotic Pediococcus Acidilactici on Antioxidant Defences and Oxidative Stress Status of Shrimp Litopenaeus Stylirostris. Aquaculture 2009, 294, 306–313. [Google Scholar] [CrossRef]
- Chang, X.; Chen, Y.; Feng, J.; Huang, M.; Zhang, J. Amelioration of Cd-Induced Bioaccumulation, Oxidative Stress and Immune Damage by Probiotic Bacillus Coagulans in Common Carp (Cyprinus Carpio L.). Aquac. Rep. 2021, 20, 100678. [Google Scholar] [CrossRef]
- Merola, C.; Bisegna, A.; Angelozzi, G.; Conte, A.; Abete, M.C.; Stella, C.; Pederiva, S.; Faggio, C.; Riganelli, N.; Perugini, M. Study of Heavy Metals Pollution and Vitellogenin Levels in Brown Trout (Salmo Trutta Trutta) Wild Fish Populations. Appl. Sci. 2021, 11, 4965. [Google Scholar] [CrossRef]
- Jyoti, D.; Sinha, R.; Faggio, C. Advances in Biological Methods for the Sequestration of Heavy Metals from Water Bodies: A Review. Environ. Toxicol. Pharmacol. 2022, 103927. [Google Scholar] [CrossRef] [PubMed]
- Shahjahan, M.; Taslima, K.; Rahman, M.S.; Al-Emran, M.; Alam, S.I.; Faggio, C. Effects of Heavy Metals on Fish Physiology–a Review. Chemosphere 2022, 134519. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Al-Yahyai, R.; Ameen, A.; Sharif, Y.; Ali, L.; Fatima, M.; Khan, K.A.; Li, S. Health and Environmental Effects of Heavy Metals. J. King Saud Univ.-Sci. 2022, 34, 101653. [Google Scholar] [CrossRef]
- Stefanescu, I.A. Bioaccumulation of Heavy Metals by Bacillus Megaterium from Phosphogypsum Waste. Sci. Study Res. Chem. Chem. Eng. Biotechnol. Food Ind. 2015, 16, 93. [Google Scholar]
- Sonone, S.S.; Jadhav, S.; Sankhla, M.S.; Kumar, R. Water Contamination by Heavy Metals and Their Toxic Effect on Aquaculture and Human Health through Food Chain. Lett Appl NanoBioScience 2020, 10, 2148–2166. [Google Scholar] [CrossRef]
- Moiseenko, T.I.; Gashkina, N.A. Distribution and Bioaccumulation of Heavy Metals (Hg, Cd and Pb) in Fish: Influence of the Aquatic Environment and Climate. Environ. Res. Lett. 2020, 15, 115013. [Google Scholar] [CrossRef]
- Rahman, Z.; Singh, V.P. Bioremediation of Toxic Heavy Metals (THMs) Contaminated Sites: Concepts, Applications and Challenges. Environ. Sci. Pollut. Res. 2020, 27, 27563–27581. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, Y.; Zhang, C.; Zhang, D.; Yao, C.; Meng, Q.; Zhao, R.; Wei, Z. Speciation, Toxicity Mechanism and Remediation Ways of Heavy Metals during Composting: A Novel Theoretical Microbial Remediation Method Is Proposed. J. Environ. Manage. 2020, 272, 111109. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.J.; Moses, V. Metal Recovery and Processing. In Biotechnology; CRC Press, 2020; pp. 623–637 ISBN 1-00-307843-5.
- Fakhar, A.; Gul, B.; Gurmani, A.R.; Khan, S.M.; Ali, S.; Sultan, T.; Chaudhary, H.J.; Rafique, M.; Rizwan, M. Heavy Metal Remediation and Resistance Mechanism of Aeromonas, Bacillus, and Pseudomonas: A Review. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1868–1914. [Google Scholar] [CrossRef]
- Rekadwad, B. Microbial Systematics; CRC Press, Boca Raton, FL, USA, 2021.
- Ringø, E.; Van Doan, H.; Lee, S.H.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, Lactic Acid Bacteria and Bacilli: Interesting Supplementation for Aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef]
- Valipour, A.; Nedaei, S.; Noori, A.; Khanipour, A.A.; Hoseinifar, S.H. Dietary Lactobacillus Plantarum Affected on Some Immune Parameters, Air-Exposure Stress Response, Intestinal Microbiota, Digestive Enzyme Activity and Performance of Narrow Clawed Crayfish (Astacus Leptodactylus, Eschscholtz). Aquaculture 2019, 504, 121–130. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, Z.; Zhao, F.; Liu, H.; Yu, L.; Zha, J.; Wang, G. Probiotic Potential of Bacillus Velezensis JW: Antimicrobial Activity against Fish Pathogenic Bacteria and Immune Enhancement Effects on Carassius Auratus. Fish Shellfish Immunol. 2018, 78, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-B.; Wu, Y.-C.; Chi, S.-C. Dietary Supplementation of Pediococcus Pentosaceus Enhances Innate Immunity, Physiological Health and Resistance to Vibrio Anguillarum in Orange-Spotted Grouper (Epinephelus Coioides). Fish Shellfish Immunol. 2014, 39, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Dejene, F.; Regasa Dadi, B.; Tadesse, D. In Vitro Antagonistic Effect of Lactic Acid Bacteria Isolated from Fermented Beverage and Finfish on Pathogenic and Foodborne Pathogenic Microorganism in Ethiopia. Int. J. Microbiol. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Huy, N.D.; Ngoc, L.M.T.; Loc, N.H.; Lan, T.T.; Quang, H.T.; Dung, T. Isolation of Weissella Cibaria from Pacific White Shrimp (Litopenaeus Vannamei) Gastrointestinal Tract and Evaluation of Its Pathogenic Bacterial Inhibition. Indian J. Sci. Technol. 2020, 13, 1200–1212. [Google Scholar] [CrossRef]
- Puvanasundram, P.; Chong, C.M.; Sabri, S.; Yusoff, M.S.; Karim, M. Multi-Strain Probiotics: Functions, Effectiveness and Formulations for Aquaculture Applications. Aquac. Rep. 2021, 21, 100905. [Google Scholar] [CrossRef]
- Midhun, S.J.; Neethu, S.; Arun, D.; Vysakh, A.; Divya, L.; Radhakrishnan, E.K.; Jyothis, M. Dietary Supplementation of Bacillus Licheniformis HGA8B Improves Growth Parameters, Enzymatic Profile and Gene Expression of Oreochromis Niloticus. Aquaculture 2019, 505, 289–296. [Google Scholar] [CrossRef]
- Yang, G.; Tian, X.; Dong, S.; Peng, M.; Wang, D. Effects of Dietary Bacillus Cereus G19, B. Cereus BC-01, and Paracoccus Marcusii DB11 Supplementation on the Growth, Immune Response, and Expression of Immune-Related Genes in Coelomocytes and Intestine of the Sea Cucumber (Apostichopus Japonicus Selenka). Fish Shellfish Immunol. 2015, 45, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yuan, L.; Wan, J.; Sun, Z.; Wang, Y.; Sun, H. Effects of Potential Probiotic Bacillus Cereus EN25 on Growth, Immunity and Disease Resistance of Juvenile Sea Cucumber Apostichopus Japonicus. Fish Shellfish Immunol. 2016, 49, 237–242. [Google Scholar] [CrossRef]
- Zhang, J.-J.; Yang, H.-L.; Yan, Y.-Y.; Zhang, C.-X.; Ye, J.; Sun, Y.-Z. Effects of Fish Origin Probiotics on Growth Performance, Immune Response and Intestinal Health of Shrimp (Litopenaeus Vannamei) Fed Diets with Fish Meal Partially Replaced by Soybean Meal. Aquac. Nutr. 2020, 26, 1255–1265. [Google Scholar] [CrossRef]
- Newaj-Fyzul, A.; Adesiyun, A.A.; Mutani, A.; Ramsubhag, A.; Brunt, J.; Austin, B. Bacillus Subtilis AB1 Controls Aeromonas Infection in Rainbow Trout (Oncorhynchus Mykiss, Walbaum). J. Appl. Microbiol. 2007, 103, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Sahandi, J.; Jafaryan, H.; Soltani, M.; Ebrahimi, P. The Use of Two Bifidobacterium Strains Enhanced Growth Performance and Nutrient Utilization of Rainbow Trout (Oncorhynchus Mykiss) Fry. Probiotics Antimicrob. Proteins 2019, 11, 966–972. [Google Scholar] [CrossRef]
- Ringø, E.; Seppola, M.; Berg, A.; Olsen, R.E.; Schillinger, U.; Holzapfel, W. Characterization of Carnobacterium Divergens Strain 6251 Isolated from Intestine of Arctic Charr (Salvelinus Alpinus L.). Syst. Appl. Microbiol. 2002, 25, 120–129. [Google Scholar] [CrossRef]
- Ringø, E. The Ability of Carnobacteria Isolated from Fish Intestine to Inhibit Growth of Fish Pathogenic Bacteria: A Screening Study. Aquac. Res. 2008, 39, 171–180. [Google Scholar] [CrossRef]
- Vendrell, D.; Balcazar, J.L.; de Blas, I.; Ruiz-Zarzuela, I.; Gironés, O.; Muzquiz, J.L. Protection of Rainbow Trout (Oncorhynchus Mykiss) from Lactococcosis by Probiotic Bacteria. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Al-Dohail, M.A.; Hashim, R.; Aliyu-Paiko, M. Evaluating the Use of Lactobacillus Acidophilus as a Biocontrol Agent against Common Pathogenic Bacteria and the Effects on the Haematology Parameters and Histopathology in African Catfish Clarias Gariepinus Juveniles: L. Acidophilus as a Probiotic in African Catfish. Aquac. Res. 2011, 42, 196–209. [Google Scholar] [CrossRef]
- Zheng, C.N.; Wang, W. Effects of Lactobacillus Pentosus on the Growth Performance, Digestive Enzyme and Disease Resistance of White Shrimp, Litopenaeus Vannamei (Boone, 1931). Aquac. Res. 2017, 48, 2767–2777. [Google Scholar] [CrossRef]
- Lee, S.H.; Beck, B.R.; Hwang, S.-H.; Song, S.K. Feeding Olive Flounder (Paralichthys Olivaceus) with Lactococcus Lactis BFE920 Expressing the Fusion Antigen of Vibrio OmpK and FlaB Provides Protection against Multiple Vibrio Pathogens: A Universal Vaccine Effect. Fish Shellfish Immunol. 2021, 114, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Balcázar, J.L.; Vendrell, D.; De Blas, I.; Ruiz-Zarzuela, I.; Múzquiz, J.L. Effect of Lactococcus Lactis CLFP 100 and Leuconostoc Mesenteroides CLFP 196 on Aeromonas Salmonicida Infection in Brown Trout (Salmo Trutta). Microb. Physiol. 2009, 17, 153–157. [Google Scholar] [CrossRef]
- Thao, T.T.P.; Lan, T.T.P.; Phuong, T.V.; Truong, H.T.H.; Khoo, K.S.; Manickam, S.; Hoa, T.T.; Tram, N.D.Q.; Show, P.L.; Huy, N.D. Characterization Halotolerant Lactic Acid Bacteria Pediococcus Pentosaceus HN10 and in Vivo Evaluation for Bacterial Pathogens Inhibition. Chem. Eng. Process.-Process Intensif. 2021, 168, 108576. [Google Scholar] [CrossRef]
- Tarkhani, R.; Imani, A.; Hoseinifar, S.H.; Moghanlou, K.S.; Manaffar, R. The Effects of Host-Associated Enterococcus Faecium CGMCC1. 2136 on Serum Immune Parameters, Digestive Enzymes Activity and Growth Performance of the Caspian Roach (Rutilus Rutilus Caspicus) Fingerlings. Aquaculture 2020, 519, 734741. [Google Scholar] [CrossRef]
- Safari, R.; Adel, M.; Lazado, C.C.; Caipang, C.M.A.; Dadar, M. Host-Derived Probiotics Enterococcus Casseliflavus Improves Resistance against Streptococcus Iniae Infection in Rainbow Trout (Oncorhynchus Mykiss) via Immunomodulation. Fish Shellfish Immunol. 2016, 52, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Soltanian, S.; Mirghaed, A.T.; Akhlaghi, M.; Hoseinifar, S.H.; Esmailnejad, A. The Effect of Oral Administration of Lactic Acid Bacteria Isolated from Kefir on Intestinal Microbiota, Growth Performance and Survival in Juvenile Rainbow Trout, Oncorhynchus Mykiss. Int. J. Aquat. Biol. 2020, 8, 35–49. [Google Scholar] [CrossRef]
- Sakai, M.; Yoshida, T.; Atsuta, S.; Kobayashi, M. Enhancement of Resistance to Vibriosis in Rainbow Trout, Oncorhynchus Mykiss (Walbaum), by Oral Administration of Clostridium Butyricum Bacterin. J. Fish Dis. 1995, 18, 187–190. [Google Scholar] [CrossRef]
- Meng, X.; Wu, S.; Hu, W.; Zhu, Z.; Yang, G.; Zhang, Y.; Qin, C.; Yang, L.; Nie, G. Clostridium Butyricum Improves Immune Responses and Remodels the Intestinal Microbiota of Common Carp (Cyprinus Carpio L.). Aquaculture 2021, 530, 735753. [Google Scholar] [CrossRef]
- Kahyani, F.; Pirali-Kheirabadi, E.; Shafiei, S.; Shenavar Masouleh, A. Effect of Dietary Supplementation of Potential Probiotic Weissella Confusa on Innate Immunity, Immune-related Genes Expression, Intestinal Microbiota and Growth Performance of Rainbow Trout (Oncorhynchus Mykiss). Aquac. Nutr. 2021, 27, 1411–1420. [Google Scholar] [CrossRef]
- Rengpipat, S.; Rueangruklikhit, T.; Piyatiratitivorakul, S. Evaluations of Lactic Acid Bacteria as Probiotics for Juvenile Seabass Lates Calcarifer. Aquac. Res. 2008, 39, 134–143. [Google Scholar] [CrossRef]
- Jinendiran, S.; Archana, R.; Sathishkumar, R.; Kannan, R.; Selvakumar, G.; Sivakumar, N. Dietary Administration of Probiotic Aeromonas Veronii V03 on the Modulation of Innate Immunity, Expression of Immune-Related Genes and Disease Resistance against Aeromonas Hydrophila Infection in Common Carp (Cyprinus Carpio). Probiotics Antimicrob. Proteins 2021, 13, 1709–1722. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Rhman, A.M.; Khattab, Y.A.E.; Shalaby, A.M.E. Micrococcus Luteus and Pseudomonas Species as Probiotics for Promoting the Growth Performance and Health of Nile Tilapia, Oreochromis Niloticus. Fish Shellfish Immunol. 2009, 27, 175–180. [Google Scholar] [CrossRef]
- ABdel-Tawwab, M.; Mousa, M.A.A.; Mohammed, M.A. Use of Live Baker’s Yeast, Saccharomyces Cerevisiae, in Practical Diet to Enhance the Growth Performance of Galilee Tilapia, Sarotherodon Galilaeus (L.), and Its Resistance to Environmental Copper Toxicity. J. World Aquac. Soc. 2010, 41, 214–223. [Google Scholar] [CrossRef]
- Banu, M.R.; Akter, S.; Islam, M.R.; Mondol, M.N.; Hossain, M.A. Probiotic Yeast Enhanced Growth Performance and Disease Resistance in Freshwater Catfish Gulsa Tengra, Mystus Cavasius. Aquac. Rep. 2020, 16, 100237. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Alamillo, E.; Angulo, C. Probiotic and Immunomodulatory Activity of Marine Yeast Yarrowia Lipolytica Strains and Response against Vibrio Parahaemolyticus in Fish. Probiotics Antimicrob. Proteins 2021, 13, 1292–1305. [Google Scholar] [CrossRef] [PubMed]
- Raida, M.K.; Larsen, J.L.; Nielsen, M.E.; Buchmann, K. Enhanced Resistance of Rainbow Trout, Oncorhynchus Mykiss (Walbaum), against Yersinia Ruckeri Challenge Following Oral Administration of Bacillus Subtilis and B. Licheniformis (BioPlus2B). J. Fish Dis. 2003, 26, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Sayed Hassani, M.H.; Jourdehi, A.Y.; Zelti, A.H.; Masouleh, A.S.; Lakani, F.B. Effects of Commercial Superzist Probiotic on Growth Performance and Hematological and Immune Indices in Fingerlings Acipenser Baerii. Aquac. Int. 2020, 28, 377–387. [Google Scholar] [CrossRef]
- Zhang, M.; Pan, L.; Fan, D.; He, J.; Su, C.; Gao, S.; Zhang, M. Study of Fermented Feed by Mixed Strains and Their Effects on the Survival, Growth, Digestive Enzyme Activity and Intestinal Flora of Penaeus Vannamei. Aquaculture 2021, 530, 735703. [Google Scholar] [CrossRef]
- Joseph, T.C.; Kumar, A.; Basha, K.A. Prophylactic Health Products in Aquaculture. In; ICAR-Central Institute of Fisheries Technology, 2017.
- Lordan, C.; Thapa, D.; Ross, R.P.; Cotter, P.D. Potential for Enriching Next-Generation Health-Promoting Gut Bacteria through Prebiotics and Other Dietary Components. Gut Microbes 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Peredo-Lovillo, A.; Romero-Luna, H.E.; Jiménez-Fernández, M. Health Promoting Microbial Metabolites Produced by Gut Microbiota after Prebiotics Metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, Y.; Lu, Y.; Hao, H.; Liu, J.; Huang, R. Structural Features, Interaction with the Gut Microbiota and Anti-Tumor Activity of Oligosaccharides. RSC Adv. 2020, 10, 16339–16348. [Google Scholar] [CrossRef] [PubMed]
- Elumalai, P.; Kurian, A.; Lakshmi, S.; Musthafa, M.S.; Ringo, E.; Faggio, C. Effect of Leucas Aspera Against Aeromonas Hydrophila in Nile Tilapia (Oreochromis Niloticus): Immunity and Gene Expression Evaluation. Turk. J. Fish. Aquat. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Kumar, J.; Priyadharshini, M.; Madhavi, M.; Begum, S.S.; Ali, A.J.; Musthafa, M.S.; Faggio, C. Impact of Hygrophila Auriculata Supplementary Diets on the Growth, Survival, Biochemical and Haematological Parameters in Fingerlings of Freshwater Fish Cirrhinus Mrigala (Hamilton, 1822). Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2022, 263, 111097. [Google Scholar] [CrossRef]
- Aliab, S.S.R.; Ambasankar, K.; Praveena, P.E. Effect of Dietary Mannanoligosaccharide on Intestinal Microbiota and Immune Parameters of Asian Seabass (Lates Calcarifer) Juveniles. J Fish Res 2021 5 5 16-21 J Fish Res 2021 Vol. 5 Issue 5.
- Asaduzzaman, M.D.; Iehata, S.; Akter, S.; Kader, M.A.; Ghosh, S.K.; Khan, M.N.A.; Abol-Munafi, A.B. Effects of Host Gut-Derived Probiotic Bacteria on Gut Morphology, Microbiota Composition and Volatile Short Chain Fatty Acids Production of Malaysian Mahseer Tor Tambroides. Aquac. Rep. 2018, 9, 53–61. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, J.; Wu, S. The Growth Performance and Non-Specific Immunity of Juvenile Grass Carp (Ctenopharyngodon Idella) Affected by Dietary Alginate Oligosaccharide. 3 Biotech 2021, 11, 46. [Google Scholar] [CrossRef]
- Liu, Y.; Miao, Y.; Xu, N.; Ding, T.; Cui, K.; Chen, Q.; Zhang, J.; Fang, W.; Mai, K.; Ai, Q. Effects of Dietary Astragalus Polysaccharides (APS) on Survival, Growth Performance, Activities of Digestive Enzyme, Antioxidant Responses and Intestinal Development of Large Yellow Croaker (Larimichthys Crocea) Larvae. Aquaculture 2020, 517, 734752. [Google Scholar] [CrossRef]
- Hamidoghli, A.; Won, S.; Lee, S.; Lee, S.; Farris, N.W.; Bai, S.C. Nutrition and Feeding of Olive Flounder Paralichthys Olivaceus: A Review. Rev. Fish. Sci. Aquac. 2020, 28, 340–357. [Google Scholar] [CrossRef]
- Kazuń, B.; Małaczewska, J.; Kazuń, K.; Kamiński, R.; Adamek-Urbańska, D.; Żylińska-Urban, J. Dietary Administration of β-1, 3/1, 6-Glucan and Lactobacillus Plantarum Improves Innate Immune Response and Increases the Number of Intestine Immune Cells in Roach (Rutilus Rutilus). BMC Vet. Res. 2020, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Deon, M.P.P.; Bicudo, Á.J. de A.; Sado, R.Y. Performance, Hematology, and Immunology of Pacu in Response to Dietary Supplementation with Fructooligosaccharides. Pesqui. Agropecuária Bras. 2021, 56. [Google Scholar] [CrossRef]
- Yousefi, S.; Shokri, M.M.; Noveirian, H.A.; Hoseinifar, S.H. Effects of Dietary Yeast Cell Wall on Biochemical Indices, Serum and Skin Mucus Immune Responses, Oxidative Status and Resistance against Aeromonas Hydrophila in Juvenile Persian Sturgeon (Acipenser Persicus). Fish Shellfish Immunol. 2020, 106, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Serradell, A.; Torrecillas, S.; Makol, A.; Valdenegro, V.; Fernández-Montero, A.; Acosta, F.; Izquierdo, M.S.; Montero, D. Prebiotics and Phytogenics Functional Additives in Low Fish Meal and Fish Oil Based Diets for European Sea Bass (Dicentrarchus Labrax): Effects on Stress and Immune Responses. Fish Shellfish Immunol. 2020, 100, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Priya, P.S.; Ashwitha, A.; Thamizharasan, K.; Harishkumar, M.; Dinesh, S.; Nithya, T.G.; Kamaraj, M. Synergistic Effect of Durian Fruit Rind Polysaccharide Gel Encapsulated Prebiotic and Probiotic Dietary Supplements on Growth Performance, Immune-Related Gene Expression, and Disease Resistance in Zebrafish (Danio Rerio). Heliyon 2021, 7, e06669. [Google Scholar] [CrossRef]
- Mohammadian, T.; Ghanei-Motlagh, R.; Molayemraftar, T.; Mesbah, M.; Zarea, M.; Mohtashamipour, H.; Nejad, A.J. Modulation of Growth Performance, Gut Microflora, Non-Specific Immunity and Gene Expression of Proinflammatory Cytokines in Shabout (Tor Grypus) upon Dietary Prebiotic Supplementation. Fish Shellfish Immunol. 2021, 112, 38–45. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Faggio, C.; Chitmanat, C.; Mai, N.T.; Jaturasitha, S.; Ringø, E. Effects of Corncob Derived Xylooligosaccharide on Innate Immune Response, Disease Resistance, and Growth Performance in Nile Tilapia (Oreochromis Niloticus) Fingerlings. Aquaculture 2018, 495, 786–793. [Google Scholar] [CrossRef]
- Włodarczyk, M.; Śliżewska, K. Efficiency of Resistant Starch and Dextrins as Prebiotics: A Review of the Existing Evidence and Clinical Trials. Nutrients 2021, 13, 3808. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; González, M.; Murado, M.A. Effects of Lactic Acid Bacteria Cultures on Pathogenic Microbiota from Fish. Aquaculture 2005, 245, 149–161. [Google Scholar] [CrossRef]
- Poolsawat, L.; Li, X.; Xu, X.; Rahman, M.M.; Boonpeng, N.; Leng, X. Dietary Xylooligosaccharide Improved Growth, Nutrient Utilization, Gut Microbiota and Disease Resistance of Tilapia (Oreochromis Niloticus x O. Aureus). Anim. Feed Sci. Technol. 2021, 275, 114872. [Google Scholar] [CrossRef]
- Maas, R.M.; Deng, Y.; Dersjant-Li, Y.; Petit, J.; Verdegem, M.C.; Schrama, J.W.; Kokou, F. Exogenous Enzymes and Probiotics Alter Digestion Kinetics, Volatile Fatty Acid Content and Microbial Interactions in the Gut of Nile Tilapia. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Wang, M.; Zhou, J.; Selma-Royo, M.; Simal-Gandara, J.; Collado, M.C.; Barba, F.J. Potential Benefits of High-Added-Value Compounds from Aquaculture and Fish Side Streams on Human Gut Microbiota. Trends Food Sci. Technol. 2021, 112, 484–494. [Google Scholar] [CrossRef]
- Maas, R.M.; Verdegem, M.C.; Wiegertjes, G.F.; Schrama, J.W. Carbohydrate Utilisation by Tilapia: A Meta-analytical Approach. Rev. Aquac. 2020, 12, 1851–1866. [Google Scholar] [CrossRef]
- Wee, W.; Hamid, N.K.A.; Mat, K.; Khalif, R.I.A.R.; Rusli, N.D.; Rahman, M.M.; Kabir, M.A.; Wei, L.S. The Effects of Mixed Prebiotics in Aquaculture: A Review. Aquac. Fish. 2022. [Google Scholar] [CrossRef]
- Hu, X.; Yang, H.-L.; Yan, Y.-Y.; Zhang, C.-X.; Ye, J.; Lu, K.-L.; Hu, L.-H.; Zhang, J.-J.; Ruan, L.; Sun, Y.-Z. Effects of Fructooligosaccharide on Growth, Immunity and Intestinal Microbiota of Shrimp (Litopenaeus Vannamei) Fed Diets with Fish Meal Partially Replaced by Soybean Meal. Aquac. Nutr. 2019, 25, 194–204. [Google Scholar] [CrossRef]
- Mustafa, A.; Buentello, A.; Gatlin III, D.; Lightner, D.; Hume, M.; Lawrence, A. Effects of Fructooligosaccharides (FOS) on Growth, Survival, Gut Microflora, Stress, and Immune Response in Pacific White Shrimp, Litopenaeus Vannamei, Cultured in a Recirculating System. J. Immunoassay Immunochem. 2020, 41, 45–59. [Google Scholar] [CrossRef]
- Van Doan, H.; Soltani, M.; Ringø, E. In Vitro Antagonistic Effect and in Vivo Protective Efficacy of Gram-Positive Probiotics versus Gram-Negative Bacterial Pathogens in Finfish and Shellfish. Aquaculture 2021, 540, 736581. [Google Scholar] [CrossRef]
- Yamamoto, F.Y.; Older, C.E.; Hume, M.E.; Hoffmann, A.R.; Gatlin III, D.M. Effects of Butyrate, Propionate, and Their Combination in Vitro, and the Impacts of Their Supplementation in High-Plant-Protein Diets to the Production Performance, Innate Immune Responses, and Intestinal Microbiota of Red Drum (Sciaenops Ocellatus). Aquaculture 2021, 545, 737225. [Google Scholar] [CrossRef]
- Zhu, W.; Su, J. Immune Functions of Phagocytic Blood Cells in Teleost. Rev. Aquac. 2022, 14, 630–646. [Google Scholar] [CrossRef]
- Tran, N.T.; Li, S. Potential Role of Prebiotics and Probiotics in Conferring Health Benefits in Economically Important Crabs. Fish Shellfish Immunol. Rep. 2022, 3, 100041. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Querol, E.; Rosales, C. Phagocytosis: Our Current Understanding of a Universal Biological Process. Front. Immunol. 2020, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Mai, K.; Xu, W.; Zhang, W.; Ai, Q.; Zhang, Y.; Wang, X.; Liufu, Z. Influence of Dietary Probiotic Bacillus TC22 and Prebiotic Fructooligosaccharide on Growth, Immune Responses and Disease Resistance against Vibrio Splendidus Infection in Sea Cucumber Apostichopus Japonicus. J. Ocean Univ. China 2011, 10, 293–300. [Google Scholar] [CrossRef]
- Park, Y.; Zhang, Q.; Wiegertjes, G.F.; Fernandes, J.M.; Kiron, V. Adherent Intestinal Cells from Atlantic Salmon Show Phagocytic Ability and Express Macrophage-Specific Genes. Front. Cell Dev. Biol. 2020, 8, 580848. [Google Scholar] [CrossRef] [PubMed]
- Picchietti, S.; Miccoli, A.; Fausto, A.M. Gut Immunity in European Sea Bass (Dicentrarchus Labrax): A Review. Fish Shellfish Immunol. 2021, 108, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, E.; Rojas, D.A.; Morales, S.; Miralles, V.; Solari, A. Dual and Opposite Roles of Reactive Oxygen Species (ROS) in Chagas Disease: Beneficial on the Pathogen and Harmful on the Host. Oxid. Med. Cell. Longev. 2020, 2020, 1–17. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Ringo, E. Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics; John Wiley & Sons, 2014; ISBN 0-470-67271-4.
- Ghafarifarsani, H.; Rashidian, G.; Bagheri, T.; Hoseinifar, S.H.; Van Doan, H. Study on Growth Enhancement and the Protective Effects of Dietary Prebiotic Inulin on Immunity Responses of Rainbow Trout (Oncorhynchus Mykiss) Fry Infected with Aeromonas Hydrophila. Ann. Anim. Sci. 2021, 21, 543–559. [Google Scholar] [CrossRef]
- Dong, C.; Wang, J. Immunostimulatory Effects of Dietary Fructooligosaccharides on Red Swamp Crayfish, Procambarus Clarkii (Girard). Aquac. Res. 2013, 44, 1416–1424. [Google Scholar] [CrossRef]
- Wu, X.; Teame, T.; Hao, Q.; Ding, Q.; Liu, H.; Ran, C.; Yang, Y.; Zhang, Y.; Zhou, Z.; Duan, M. Use of a Paraprobiotic and Postbiotic Feed Supplement (HWFTM) Improves the Growth Performance, Composition and Function of Gut Microbiota in Hybrid Sturgeon (Acipenser Baerii x Acipenser Schrenckii). Fish Shellfish Immunol. 2020, 104, 36–45. [Google Scholar] [CrossRef]
- Geraylou, Z.; Souffreau, C.; Rurangwa, E.; De Meester, L.; Courtin, C.M.; Delcour, J.A.; Buyse, J.; Ollevier, F. Effects of Dietary Arabinoxylan-Oligosaccharides (AXOS) and Endogenous Probiotics on the Growth Performance, Non-Specific Immunity and Gut Microbiota of Juvenile Siberian Sturgeon (Acipenserábaerii). Fish Shellfish Immunol. 2013, 35, 766–775. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Ghaedi, G.; Sharifinia, M. Effects of Diets Containing Β-glucan on Survival, Growth Performance, Haematological, Immunity and Biochemical Parameters of Rainbow Trout (Oncorhynchus Mykiss) Fingerlings. Aquac. Res. 2022, 53, 1842–1850. [Google Scholar] [CrossRef]
- Marcos-López, M.; Rodger, H.D. Amoebic Gill Disease and Host Response in Atlantic Salmon (Salmo Salar L.): A Review. Parasite Immunol. 2020, 42, e12766. [Google Scholar] [CrossRef] [PubMed]
- Reis, B.; Gonçalves, A.T.; Santos, P.; Sardinha, M.; Conceição, L.E.; Serradeiro, R.; Pérez-Sánchez, J.; Calduch-Giner, J.; Schmid-Staiger, U.; Frick, K. Immune Status and Hepatic Antioxidant Capacity of Gilthead Seabream Sparus Aurata Juveniles Fed Yeast and Microalga Derived β-Glucans. Mar. Drugs 2021, 19, 653. [Google Scholar] [CrossRef]
- Pogue, R.; Murphy, E.J.; Fehrenbach, G.W.; Rezoagli, E.; Rowan, N.J. Exploiting Immunomodulatory Properties of β-Glucans Derived from Natural Products for Improving Health and Sustainability in Aquaculture-Farmed Organisms: Concise Review of Existing Knowledge, Innovation and Future Opportunities. Curr. Opin. Environ. Sci. Health 2021, 21, 100248. [Google Scholar] [CrossRef]
- Mameloco, E.J.; Traifalgar, R.F. Supplementation of Combined Mannan Oligosaccharide and β-Glucan Immunostimulants Improves Immunological Responses and Enhances Resistance of Pacific Whiteleg Shrimp, Penaeus Vannamei, against Vibrio Parahaemolyticus Infection. Int. Aquat. Res. 2020, 12, 291. [Google Scholar] [CrossRef]
- Cornet, V.; Khuyen, T.D.; Mandiki, S.N.; Betoulle, S.; Bossier, P.; Reyes-López, F.E.; Tort, L.; Kestemont, P. GAS1: A New β-Glucan Immunostimulant Candidate to Increase Rainbow Trout (Oncorhynchus Mykiss) Resistance to Bacterial Infections with Aeromonas Salmonicida Achromogenes. Front. Immunol. 2021, 12, 693613. [Google Scholar] [CrossRef]
- Hansen, J.Ø.; Lagos, L.; Lei, P.; Reveco-Urzua, F.E.; Morales-Lange, B.; Hansen, L.D.; Schiavone, M.; Mydland, L.T.; Arntzen, M.Ø.; Mercado, L. Down-Stream Processing of Baker’s Yeast (Saccharomyces Cerevisiae)–Effect on Nutrient Digestibility and Immune Response in Atlantic Salmon (Salmo Salar). Aquaculture 2021, 530, 735707. [Google Scholar] [CrossRef]
- El-Nobi, G.; Hassanin, M.; Khalil, A.A.; Mohammed, A.Y.; Amer, S.A.; Montaser, M.M.; El-Sharnouby, M.E. Synbiotic Effects of Saccharomyces Cerevisiae, Mannan Oligosaccharides, and β-Glucan on Innate Immunity, Antioxidant Status, and Disease Resistance of Nile Tilapia, Oreochromis Niloticus. Antibiotics 2021, 10, 567. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, Gh.; Ouraji, H.; Khalesi, M.K.; Sudagar, M.; Barari, A.; Zarei Dangesaraki, M.; Jani Khalili, K.H. Effects of a Prebiotic, Immunogen®, on Feed Utilization, Body Composition, Immunity and Resistance to Aeromonas Hydrophila Infection in the Common Carp Cyprinus Carpio (Linnaeus) Fingerlings. J. Anim. Physiol. Anim. Nutr. 2012, 96, 591–599. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Mirvaghefi, A.; Mojazi Amiri, B.; Rostami, H.K.; Merrifield, D.L. The Effects of Oligofructose on Growth Performance, Survival and Autochthonous Intestinal Microbiota of Beluga (Huso Huso) Juveniles. Aquac. Nutr. 2011, 17, 498–504. [Google Scholar] [CrossRef]
- Ziółkowska, E.; Bogucka, J.; Dankowiakowska, A.; Rawski, M.; Mazurkiewicz, J.; Stanek, M. Effects of a Trans-Galactooligosaccharide on Biochemical Blood Parameters and Intestine Morphometric Parameters of Common Carp (Cyprinus Carpio L.). Animals 2020, 10, 723. [Google Scholar] [CrossRef] [PubMed]
- Akter, M.N.; Zahan, K.; Zafar, M.A.; Khatun, N.; Rana, M.S.; Mursalin, M.I. Effects of Dietary Mannan Oligosaccharide on Growth Performance, Feed Utilization, Body Composition and Haematological Parameters in Asian Catfish (Clarias Batrachus) Juveniles. Turk. J. Fish. Aquat. Sci. 2021, 21, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Ai, Q.; Xu, H.; Mai, K.; Xu, W.; Wang, J.; Zhang, W. Effects of Dietary Supplementation of Bacillus Subtilis and Fructooligosaccharide on Growth Performance, Survival, Non-Specific Immune Response and Disease Resistance of Juvenile Large Yellow Croaker, Larimichthys Crocea. Aquaculture 2011, 317, 155–161. [Google Scholar] [CrossRef]
- Hu, Y.; Winter, V.; Gänzle, M. In Vitro Digestibility of Commercial and Experimental Isomalto-Oligosaccharides. Food Res. Int. 2020, 134, 109250. [Google Scholar] [CrossRef] [PubMed]
- Oushani, A.K.; Soltani, M.; Sheikhzadeh, N.; Mehrgan, M.S.; Islami, H.R. Effects of Dietary Chitosan and Nano-Chitosan Loaded Clinoptilolite on Growth and Immune Responses of Rainbow Trout (Oncorhynchus Mykiss). Fish Shellfish Immunol. 2020, 98, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Rashidpour, A.; Almajano, M.P.; Metón, I. Chitosan-Based Drug Delivery System: Applications in Fish Biotechnology. Polymers 2020, 12, 1177. [Google Scholar] [CrossRef] [PubMed]
- Andresen, A.M.S.; Gjøen, T. Chitosan Nanoparticle Formulation Attenuates Poly (I: C) Induced Innate Immune Responses against Inactivated Virus Vaccine in Atlantic Salmon (Salmo Salar). Comp. Biochem. Physiol. Part D Genomics Proteomics 2021, 40, 100915. [Google Scholar] [CrossRef]
- Kamilya, D.; Khan, M.I.R. Chitin and Chitosan as Promising Immunostimulant for Aquaculture. In Handbook of Chitin and Chitosan; Elsevier, 2020; pp. 761–771.
- de Campos, C.M.; Zanuzzo, F.S.; Gimbo, R.Y.; Favero, G.C.; Soares, M.P.; Pilarski, F.; Urbinati, E.C. Dietary Inulin Modulated the Cortisol Response and Increased the Protection against Pathogens in Juvenile Pacu (Piaractus Mesopotamicus). Aquac. Res. 2022, 53, 860–869. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, N.; Yu, X.-B.; Qiao, F.; Chen, L.-Q.; Du, Z.-Y.; Zhang, M.-L. Inulin Alleviates Adverse Metabolic Syndrome and Regulates Intestinal Microbiota Composition in Nile Tilapia (Oreochromis Niloticus) Fed with High-Carbohydrate Diet. Br. J. Nutr. 2021, 126, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Xia, B.; Zhang, B.; Li, L.; Zou, Y.; Shen, Z.; Xiang, Y.; Han, Y.; Chen, W. Mannan Oligosaccharide (MOS) on Growth Performance, Immunity, Inflammatory and Antioxidant Responses of the Common Carp (Cyprinus Carpio) under Ammonia Stress. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Zhou, L.; Li, H.; Qin, J.G.; Wang, X.; Chen, L.; Xu, C.; Li, E. Dietary Prebiotic Inulin Benefits on Growth Performance, Antioxidant Capacity, Immune Response and Intestinal Microbiota in Pacific White Shrimp (Litopenaeus Vannamei) at Low Salinity. Aquaculture 2020, 518, 734847. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Sabharwal, V.; Kaushik, P.; Joshi, A.; Aayushi, A.; Suri, M. Postbiotics: From Emerging Concept to Application. Front Sustain Food Syst 2022, 6, 887642. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021. [Google Scholar] [CrossRef]
- Mantziari, A.; Salminen, S.; Szajewska, H. Postbiotics against Pathogens Commonly Involved in Pediatric Infectious Diseases. Microorganisms 2020, 8, 1510. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-Parabiotics: The New Horizons in Microbial Biotherapy and Functional Foods. Microb. Cell Factories 2020, 19, 1–22. [Google Scholar] [CrossRef]
- Jastrząb, R.; Graczyk, D.; Siedlecki, P. Molecular and Cellular Mechanisms Influenced by Postbiotics. Int. J. Mol. Sci. 2021, 22, 13475. [Google Scholar] [CrossRef]
- Vallejo-Cordoba, B.; Castro-López, C.; García, H.S.; González-Córdova, A.F.; Hernández-Mendoza, A. Postbiotics and Paraprobiotics: A Review of Current Evidence and Emerging Trends. Adv. Food Nutr. Res. 2020, 94, 1–34. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, S.A.; Roher, N.; Boltaña, S.; Goetz, F.W. Peptidoglycan, Not Endotoxin, Is the Key Mediator of Cytokine Gene Expression Induced in Rainbow Trout Macrophages by Crude LPS. Mol. Immunol. 2010, 47, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cui, K.; Xu, D.; Wu, M.; Mai, K.; Ai, Q. Molecular Identification of Peptidoglycan Recognition Protein 5 and Its Functional Characterization in Innate Immunity of Large Yellow Croaker, Larimichthys Crocea. Dev. Comp. Immunol. 2021, 124, 104130. [Google Scholar] [CrossRef]
- Casadei, E.; Bird, S.; Wadsworth, S.; Vecino, J.L.G.; Secombes, C.J. The Longevity of the Antimicrobial Response in Rainbow Trout (Oncorhynchus Mykiss) Fed a Peptidoglycan (PG) Supplemented Diet. Fish Shellfish Immunol. 2015, 44, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhang, Y.; Wei, S.; Huang, J. Effects of Different Enzymatic Hydrolysis Methods on the Bioactivity of Peptidoglycan in Litopenaeus Vannamei. Chin. J. Oceanol. Limnol. 2013, 31, 374–383. [Google Scholar] [CrossRef]
- Itami, T.; Asano, M.; Tokushige, K.; Kubono, K.; Nakagawa, A.; Takeno, N.; Nishimura, H.; Maeda, M.; Kondo, M.; Takahashi, Y. Enhancement of Disease Resistance of Kuruma Shrimp, Penaeus Japonicus, after Oral Administration of Peptidoglycan Derived from Bifidobacterium Thermophilum. Aquaculture 1998, 164, 277–288. [Google Scholar] [CrossRef]
- Teame, T.; Wang, A.; Xie, M.; Zhang, Z.; Yang, Y.; Ding, Q.; Gao, C.; Olsen, R.E.; Ran, C.; Zhou, Z. Paraprobiotics and Postbiotics of Probiotic Lactobacilli, Their Positive Effects on the Host and Action Mechanisms: A Review. Front. Nutr. 2020, 7, 570344. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wang, B.; Jiang, K.; Wang, M.; Zhou, S.; Liu, M.; Wang, L. Exploring the Influence of the Surface Proteins on Probiotic Effects Performed by Lactobacillus Pentosus HC-2 Using Transcriptome Analysis in Litopenaeus Vannamei Midgut. Fish Shellfish Immunol. 2019, 87, 853–870. [Google Scholar] [CrossRef] [PubMed]
- Sudhakaran, G.; Guru, A.; Haridevamuthu, B.; Murugan, R.; Arshad, A.; Arockiaraj, J. Molecular Properties of Postbiotics and Their Role in Controlling Aquaculture Diseases. Aquac. Res. 2022, 53, 3257–3273. [Google Scholar] [CrossRef]
- Kim, D.-H.; Subramanian, D.; Park, S.-H.; Jang, Y.-H.; Heo, M.-S. Assessment and Potential Application of the Probiotic Strain, Bacillus Amyloliquefaciens JFP2, Isolated from Fermented Seafood-Jeotgal in Flounder Paralichthys Olivaceus Juveniles. Isr. J. Aquac.-Bamidgeh 2017, 69. [Google Scholar] [CrossRef]
- Mora-Sánchez, B.; Balcázar, J.L.; Pérez-Sánchez, T. Effect of a Novel Postbiotic Containing Lactic Acid Bacteria on the Intestinal Microbiota and Disease Resistance of Rainbow Trout (Oncorhynchus Mykiss). Biotechnol. Lett. 2020, 42, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- İncili, G.K.; Karatepe, P.; Akgöl, M.; Güngören, A.; Koluman, A.; İlhak, O.İ.; Kanmaz, H.; Kaya, B.; Hayaloğlu, A.A. Characterization of Lactic Acid Bacteria Postbiotics, Evaluation in-Vitro Antibacterial Effect, Microbial and Chemical Quality on Chicken Drumsticks. Food Microbiol. 2022, 104, 104001. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Foo, H.L.; Loh, T.C.; Lim, E.T.C.; Abdul Mutalib, N.E. Comparative Studies of Inhibitory and Antioxidant Activities, and Organic Acids Compositions of Postbiotics Produced by Probiotic Lactiplantibacillus Plantarum Strains Isolated from Malaysian Foods. Front. Vet. Sci. 2021, 7, 602280. [Google Scholar] [CrossRef]
- Luo, K.; Tian, X.; Wang, B.; Wei, C.; Wang, L.; Zhang, S.; Liu, Y.; Li, T.; Dong, S. Evaluation of Paraprobiotic Applicability of Clostridium Butyricum CBG01 in Improving the Growth Performance, Immune Responses and Disease Resistance in Pacific White Shrimp, Penaeus Vannamei. Aquaculture 2021, 544, 737041. [Google Scholar] [CrossRef]
- del Valle, J.C.; Bonadero, M.C.; Gimenez, A.V.F. Saccharomyces Cerevisiae as Probiotic, Prebiotic, Synbiotic, Postbiotics and Parabiotics in Aquaculture: An Overview. Aquaculture 2023, 739342. [Google Scholar] [CrossRef]
- Sousa, J.M.G.; Louvado, A.; Coelho, F.; Oliveira, V.; Oliveira, H.; Cleary, D.F.R.; Gomes, N.C.M. In Vitro Study of the Modulatory Effects of Heat-Killed Bacterial Biomass on Aquaculture Bacterioplankton Communities. Sci. Rep. 2022, 12, 19699. [Google Scholar] [CrossRef]
- Feng, J.; Cai, Z.; Chen, Y.; Zhu, H.; Chang, X.; Wang, X.; Liu, Z.; Zhang, J.; Nie, G. Effects of an Exopolysaccharide from Lactococcus Lactis Z-2 on Innate Immune Response, Antioxidant Activity, and Disease Resistance against Aeromonas Hydrophila in Cyprinus Carpio L. Fish Shellfish Immunol. 2020, 98, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Gao, Q.; Min, M.; Zhang, C.; Peng, S.; Shi, Z. Ability of Lactobacillus Plantarum Lipoteichoic Acid to Inhibit Vibrio Anguillarum-Induced Inflammation and Apoptosis in Silvery Pomfret (Pampus Argenteus) Intestinal Epithelial Cells. Fish Shellfish Immunol. 2016, 54, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Hao, Q.; Zhang, Q.; Yu, Z.; Liu, S.; Yang, Y.; Ran, C.; Zhang, Z.; Zhou, Z. A Compound of Paraprobiotic and Postbiotic Derived from Autochthonous Microorganisms Improved Growth Performance, Epidermal Mucus, Liver and Gut Health and Gut Microbiota of Common Carp (Cyprinus Carpio). Aquaculture 2023, 570, 739378. [Google Scholar] [CrossRef]
- Rimoldi, S.; Gini, E.; Koch, J.F.A.; Iannini, F.; Brambilla, F.; Terova, G. Effects of Hydrolyzed Fish Protein and Autolyzed Yeast as Substitutes of Fishmeal in the Gilthead Sea Bream (Sparus Aurata) Diet, on Fish Intestinal Microbiome. BMC Vet. Res. 2020, 16, 1–13. [Google Scholar] [CrossRef]
- Van Nguyen, N.; Onoda, S.; Van Khanh, T.; Hai, P.D.; Trung, N.T.; Hoang, L.; Koshio, S. Evaluation of Dietary Heat-Killed Lactobacillus Plantarum Strain L-137 Supplementation on Growth Performance, Immunity and Stress Resistance of Nile Tilapia (Oreochromis Niloticus). Aquaculture 2019, 498, 371–379. [Google Scholar] [CrossRef]
- Dawood, M.A.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of Transcriptomic Profile in Aquatic Animals: Probiotics, Prebiotics and Synbiotics Scenarios. Fish Shellfish Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-W.; Chang, C.-C.; Cheng, W. Synbiotic Combination of Prebiotic, Cacao Pod Husk Pectin and Probiotic, Lactobacillus Plantarum, Improve the Immunocompetence and Growth of Litopenaeus Vannamei. Fish Shellfish Immunol. 2021, 118, 333–342. [Google Scholar] [CrossRef]
- Ghafarifarsani, H.; Hoseinifar, S.H.; Talebi, M.; Yousefi, M.; Van Doan, H.; Rufchaei, R.; Paolucci, M. Combined and Singular Effects of Ethanolic Extract of Persian Shallot (Allium Hirtifolium Boiss) and Synbiotic Biomin® IMBO on Growth Performance, Serum-and Mucus-Immune Parameters and Antioxidant Defense in Zebrafish (Danio Rerio). Animals 2021, 11, 2995. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, G.; Hafezieh, M.; Karimi, A.A.; Azra, M.N.; Van Doan, H.; Tapingkae, W.; Abdelrahman, H.A.; Dawood, M.A. The Synergistic Effects of Plant Polysaccharide and Pediococcus Acidilactici as a Synbiotic Additive on Growth, Antioxidant Status, Immune Response, and Resistance of Nile Tilapia (Oreochromis Niloticus) against Aeromonas Hydrophila. Fish Shellfish Immunol. 2022, 120, 304–313. [Google Scholar] [CrossRef]
- Dehaghani, P.G.; Baboli, M.J.; Moghadam, A.T.; Ziaei-Nejad, S.; Pourfarhadi, M. Effect of Synbiotic Dietary Supplementation on Survival, Growth Performance, and Digestive Enzyme Activities of Common Carp (Cyprinus Carpio) Fingerlings. Czech J. Anim. Sci. 2015, 60, 224–232. [Google Scholar] [CrossRef]
- Kapoor, D.; Sharma, P.; Sharma, M.M.M.; Kumari, A.; Kumar, R. Microbes in Pharmaceutical Industry. Microb. Divers. Interv. Scope 2020, 259–299. [Google Scholar] [CrossRef]
- Elabd, H.; Faggio, C.; Mahboub, H.H.; Emam, M.A.; Kamel, S.; El Kammar, R.; Abdelnaeim, N.S.; Shaheen, A.; Tresnakova, N.; Matter, A. Mucuna Pruriens Seeds Extract Boosts Growth, Immunity, Testicular Histology, and Expression of Immune-Related Genes of Mono-Sex Nile Tilapia (Oreochromis Niloticus). Fish Shellfish Immunol. 2022, 127, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Lumsangkul, C.; Linh, N.V.; Chaiwan, F.; Abdel-Tawwab, M.; Dawood, M.A.; Faggio, C.; Jaturasitha, S.; Van Doan, H. Dietary Treatment of Nile Tilapia (Oreochromis Niloticus) with Aquatic Fern (Azolla Caroliniana) Improves Growth Performance, Immunological Response, and Disease Resistance against Streptococcus Agalactiae Cultured in Bio-Floc System. Aquac. Rep. 2022, 24, 101114. [Google Scholar] [CrossRef]
- Jamal, M.T.; Sumon, A.A.; Pugazhendi, A.; Al Harbi, M.; Hussain, A.; Haque, F. Use of Probiotics in Commercially Important Finfish Aquaculture. Int. J. Probiotics Prebiotics 2020, 15. [Google Scholar] [CrossRef]
- Prabawati, E.; Hu, S.-Y.; Chiu, S.-T.; Balantyne, R.; Risjani, Y.; Liu, C.-H. A Synbiotic Containing Prebiotic Prepared from a By-Product of King Oyster Mushroom, Pleurotus Eryngii and Probiotic, Lactobacillus Plantarum Incorporated in Diet to Improve the Growth Performance and Health Status of White Shrimp, Litopenaeus Vannamei. Fish Shellfish Immunol. 2022, 120, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, A.; Das, R.R.; Sivakumar, M.R.; Saravanan, A.; Saranya, C.; Sudheer, N.S.; Vasagam, K.K.; Mahalakshmi, P.; Kannappan, S.; Gopikrishna, G. Bio-Augmentation of Heterotrophic Bacteria in Biofloc System Improves Growth, Survival, and Immunity of Indian White Shrimp Penaeus Indicus. Fish Shellfish Immunol. 2020, 98, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Rajeev, R.; Adithya, K.K.; Kiran, G.S.; Selvin, J. Healthy Microbiome: A Key to Successful and Sustainable Shrimp Aquaculture. Rev. Aquac. 2021, 13, 238–258. [Google Scholar] [CrossRef]
- Safari, O.; Sarkheil, M.; Shahsavani, D.; Paolucci, M. Effects of Single or Combined Administration of Dietary Synbiotic and Sodium Propionate on Humoral Immunity and Oxidative Defense, Digestive Enzymes and Growth Performances of African Cichlid (Labidochromis Lividus) Challenged with Aeromonas Hydrophila. Fishes 2021, 6, 63. [Google Scholar] [CrossRef]
- Ye, J.-D.; Wang, K.; Li, F.-D.; Sun, Y.-Z. Single or Combined Effects of Fructo- and Mannan Oligosaccharide Supplements and Bacillus Clausii on the Growth, Feed Utilization, Body Composition, Digestive Enzyme Activity, Innate Immune Response and Lipid Metabolism of the Japanese Flounder Paralichth: Dietary Benefits of FOS, MOS and B. Clausii in the Japanese Flounder. Aquac. Nutr. 2011, 17, e902–e911. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Hoseini, S.M.; Bagheri, D. Effects of Galactooligosaccharide and Pediococcus Acidilactici on Antioxidant Defence and Disease Resistance of Rainbow Trout, Oncorhynchus Mykiss. Ann. Anim. Sci. 2017, 17, 217–227. [Google Scholar] [CrossRef]
- Wan, J.-J.; Pan, J.; Shen, M.-F.; Xue, H.; Sun, M.; Zhang, M.-Q.; Zhu, X.-H.; Ma, X. Changes in the Growth Performance, Antioxidant Enzymes and Stress Resistance Caused by Dietary Administration of Synbiotic (Fructooligosaccharide and Probiotics) in Juvenile Chinese Mitten Crab, Eriocheir Sinensis. Aquac. Int. 2022, 1–15. [Google Scholar] [CrossRef]
- Maniat, M.; Salati, A.P.; Zanguee, N.; Mousavi, S.M.; Hoseinifar, S.H. Effects of Dietary Pediococcus Acidilactici and Isomaltooligosaccharide on Growth Performance, Immunity, and Antioxidant Defense in Juvenile Common Carp. Aquac. Nutr. 2023, 2023, e1808640. [Google Scholar] [CrossRef]
- Rohani, M.F.; Islam, S.M.; Hossain, M.K.; Ferdous, Z.; Siddik, M.A.; Nuruzzaman, M.; Padeniya, U.; Brown, C.; Shahjahan, M. Probiotics, Prebiotics and Synbiotics Improved the Functionality of Aquafeed: Upgrading Growth, Reproduction, Immunity and Disease Resistance in Fish. Fish Shellfish Immunol. 2022, 120, 569–589. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Abdel-Daim, M.M.; Van Doan, H. Probiotic Application for Sustainable Aquaculture. Rev. Aquac. 2019, 11, 907–924. [Google Scholar] [CrossRef]
- Goh, J.X.H.; Tan, L.T.-H.; Law, J.W.-F.; Ser, H.-L.; Khaw, K.-Y.; Letchumanan, V.; Lee, L.-H.; Goh, B.-H. Harnessing the Potentialities of Probiotics, Prebiotics, Synbiotics, Paraprobiotics, and Postbiotics for Shrimp Farming. Rev. Aquac. 2022, 14, 1478–1557. [Google Scholar] [CrossRef]
- Carbone, D.; Faggio, C. Importance of Prebiotics in Aquaculture as Immunostimulants. Effects on Immune System of Sparus Aurata and Dicentrarchus Labrax. Fish Shellfish Immunol. 2016, 54, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, F.M.; Hussein, S.H.; Awad, S.M.; El-Makhzangy, A.A. Growth Promoter, Immune Response, and Histopathological Change of Prebiotic, Probiotic and Synbiotic Bacteria on Nile Tilapia. Saudi J. Biol. Sci. 2023, 30, 103539. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, L.; Esteban, M.Á. Administration of Probiotics in the Water in Finfish Aquaculture Systems: A Review. Fishes 2018, 3, 33. [Google Scholar] [CrossRef]
Figure 1.
Illustration of the use and impact of probiotics in aquaculture systems.
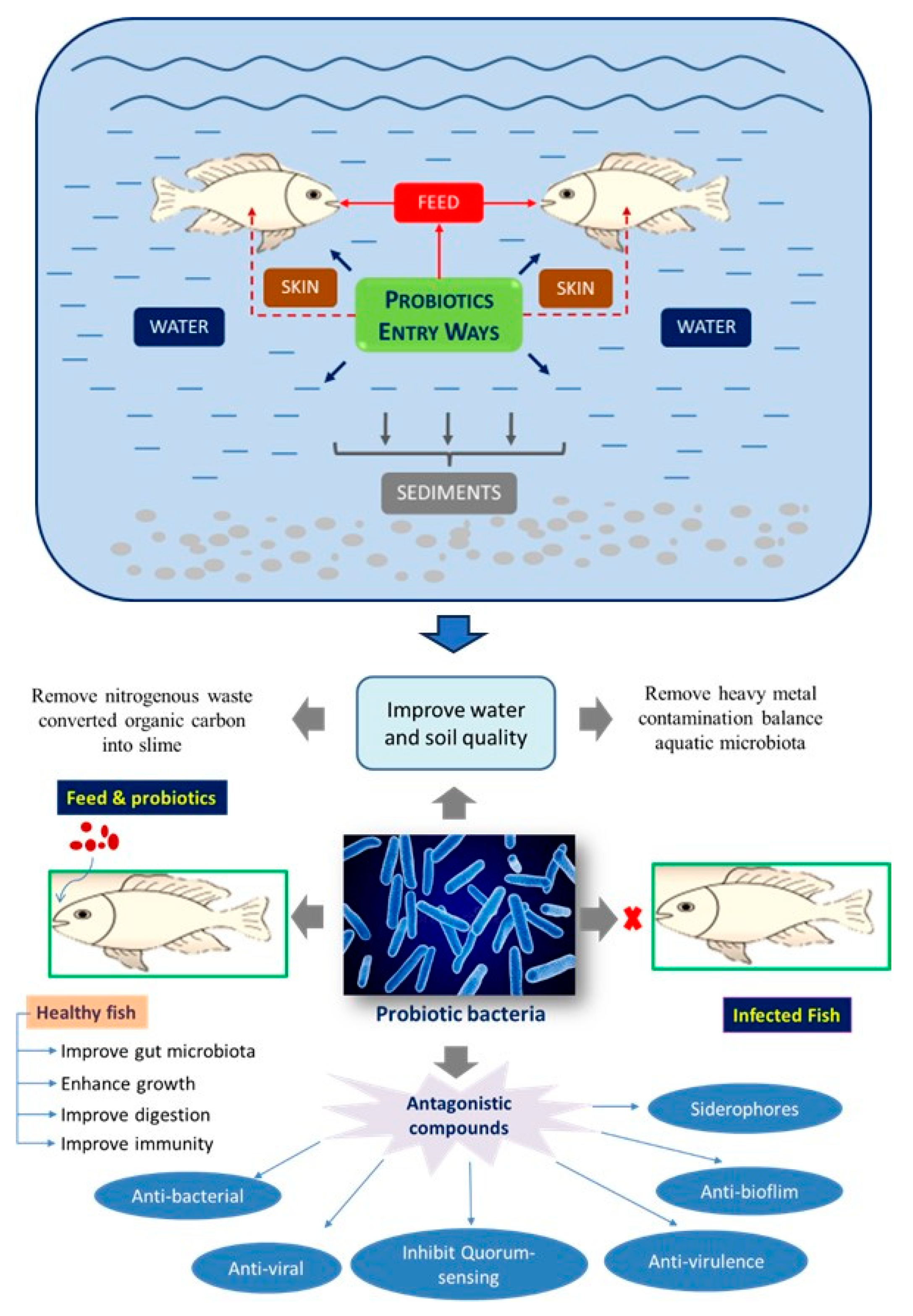
Figure 2.
Main chemical components of prebiotics from natural sources and their action modes in improving host health.
Figure 2.
Main chemical components of prebiotics from natural sources and their action modes in improving host health.
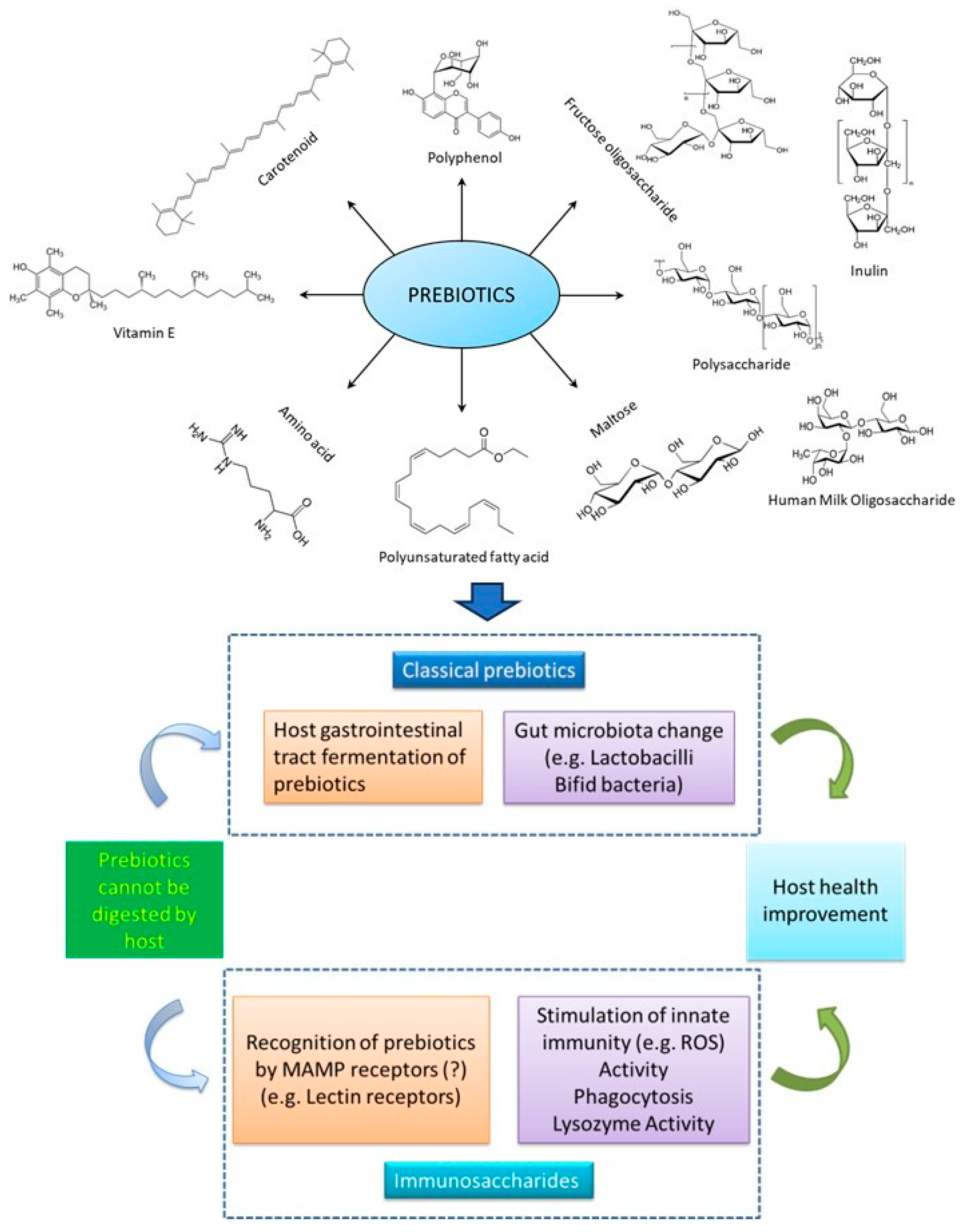
Figure 3.
Postbiotic main components and molecules.
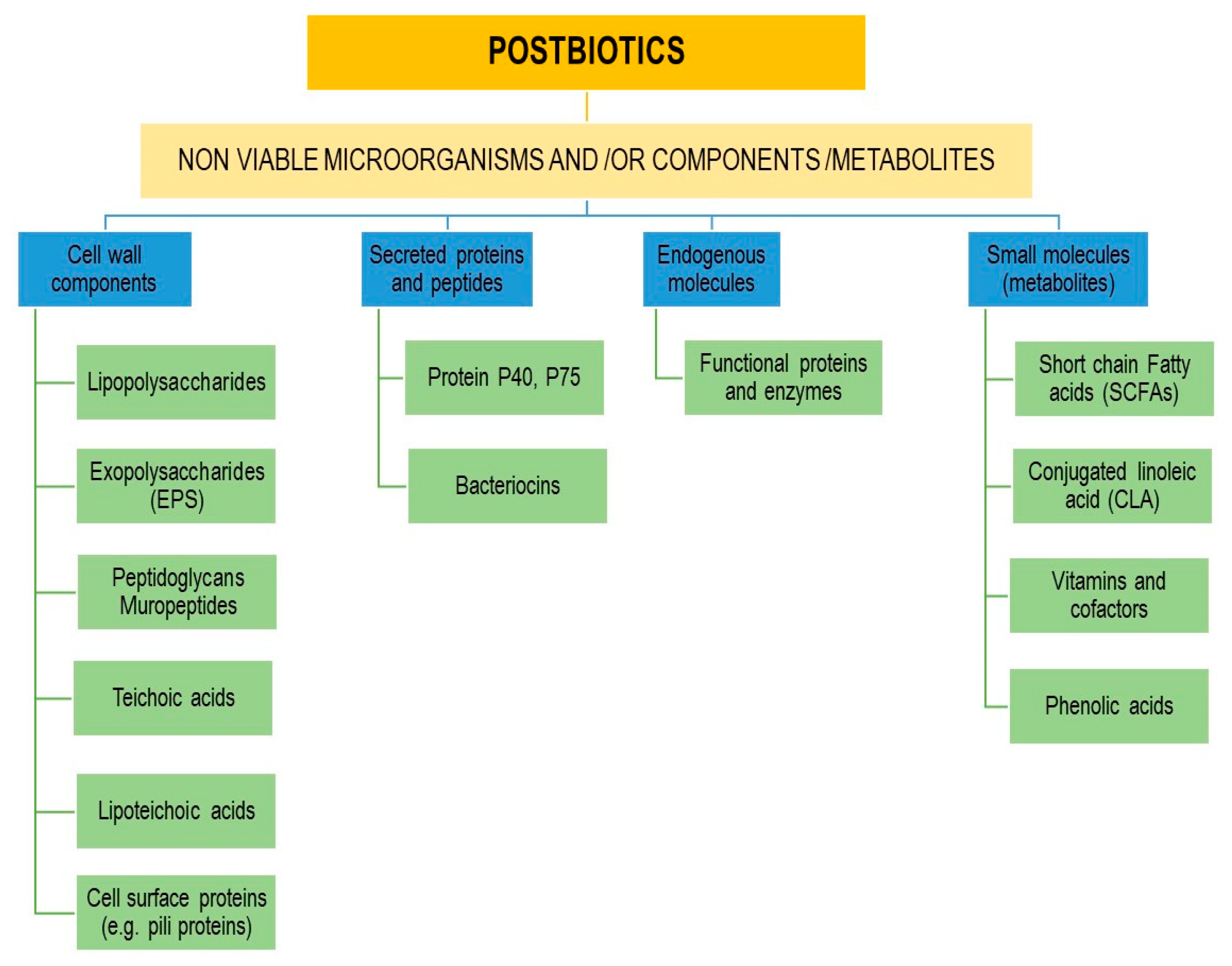
Figure 4.
Postbiotics in aquaculture.
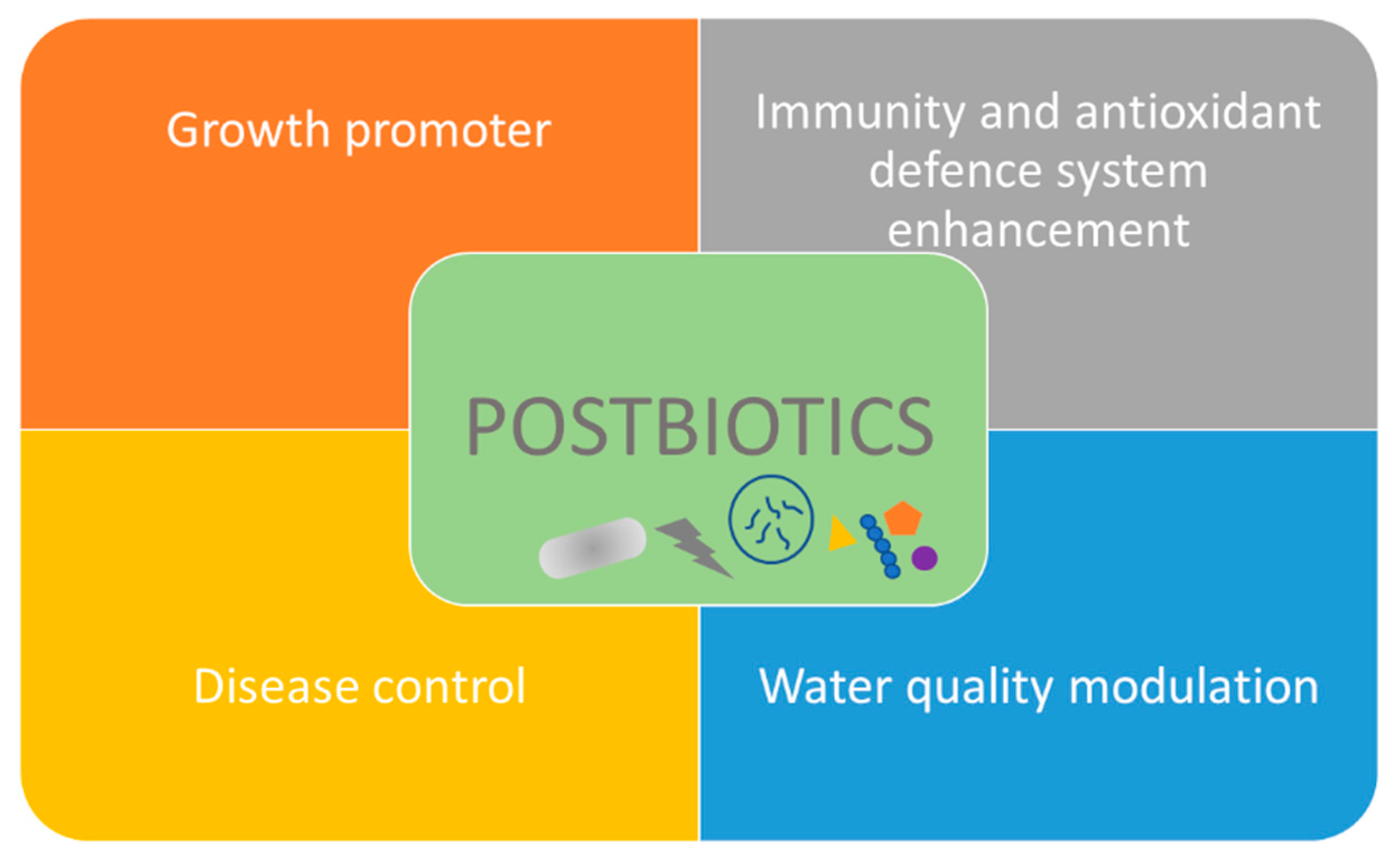
Figure 5.
Illustration of modes of action of synbiotics in aquaculture.
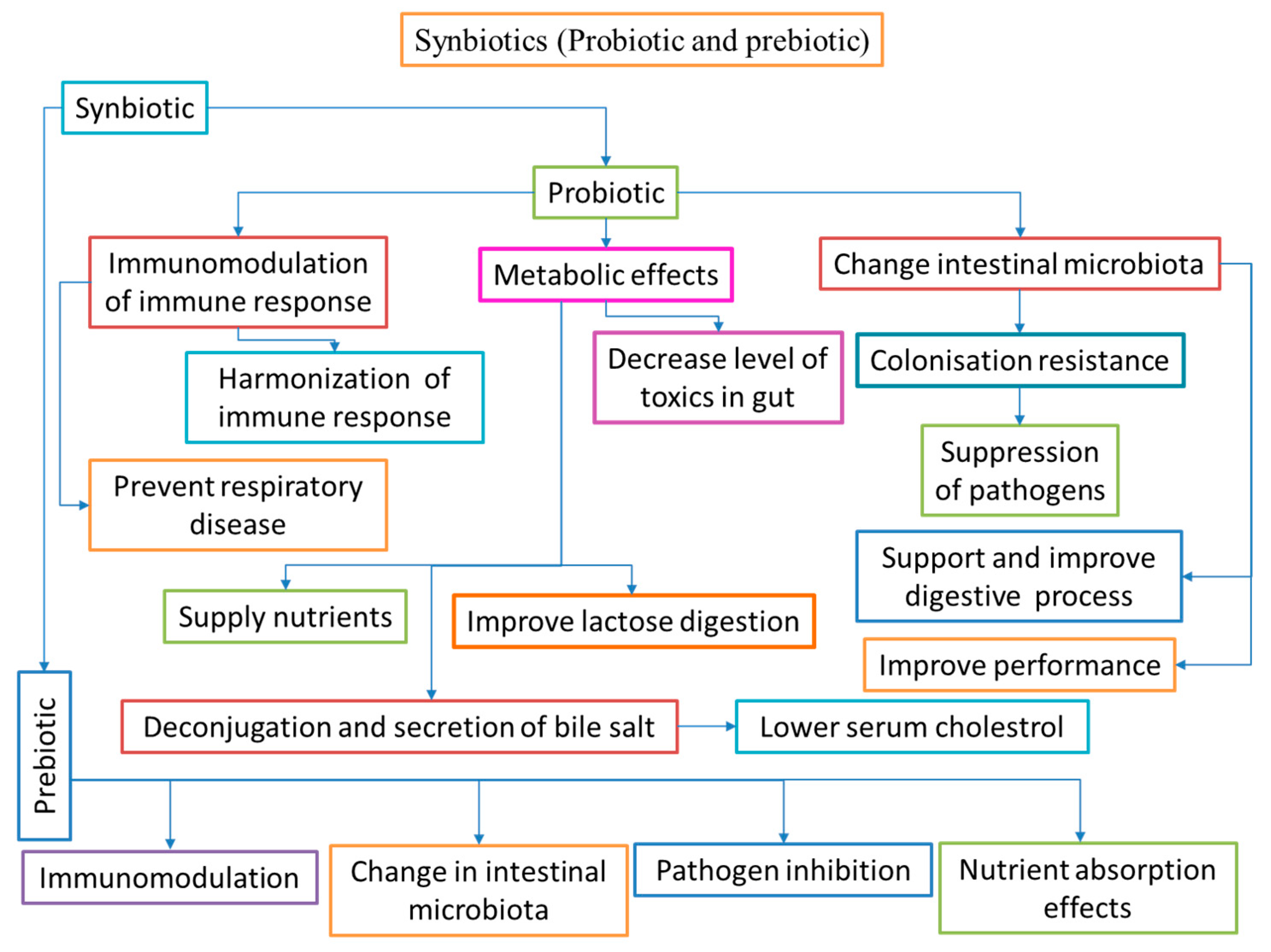

Table 1.
Functional feed additives of major probiotics in aquatic animals.
| Probiotics organisms | Functions | Aquatic organisms | References |
|---|---|---|---|
| Bacillus | |||
| B. licheniformis HGA8B | ↑growth performances and feed conversion ratio Up-regulation of immune genes |
O. niloticus | [129] |
|
B. cereus G19 B. cereus BC-01 |
↑ growth and immunity | Apostichopus japonicus | [130] |
| B. cereus EN25 | Immunity and resistance against Vibrio splendidus | A. japonicus | [131] |
| B. pumilus SE5 | ↑ growth and immunity | L. vannamei | [132] |
| B. subtilis AB1 | Bactericidal activity against Aeromonas infection | O. mykiss | [133] |
| Bifidobacterium | |||
| Bifidobacterium animalis PTCC-1631 | ↑growth performance, digestion and nutrient utilisation | O. mykiss | [134] |
| B. lactis PTCC-1736 | ↑growth, nutrient digestability and carcass composition | O. mykiss | [134] |
| Carnobacterium | |||
|
C. divergens C. maltaromaticum |
Antagonistic effects against V. anguillarum, V. viscosus and A. salmonicida | - | [135,136] |
| Lactobacillus | |||
| L. plantarum CLFP | ↓mortalities against harmful strain L. garvieae | O. mykiss | [137] |
| L. acidophilus | Against Staphylococcus xylosus, Aeromonas hydrophila gr.2 and Streptococcus agalactiae infection | Clarias gariepinus | [138] |
| L. pentosus | ↑growth performances and feed conversion ratio ↑survival against Vibrio species |
L. vannamei | [139] |
| Lactococcus | |||
| Lactococcus lactis BFE920 | Activation of non-specific immune system Bactericidal activity against S. iniae |
Paralichthys olivaceus | [140] |
| Leuconostoc | |||
| Lc. Mesenteroides CLFP 196 | ↑survival against A. salmonicida infection | Salmo trutta | [141] |
| Pediococcus | |||
| P. pentosaceus HN10 | ↑ feed utilization, digestive enzyme activity and antivibrio activity | L. vannamei | [142] |
| Enterococcus | |||
| E. casseliflavus CGMCC1.2136 | ↑growth performances, immunity and digestive enzyme activity | Rutilus rutilus caspicus | [143] |
| E. casseliflavus | ↑growth performances and disease resistance against S. iniae | O. mykiss | [144] |
| E. durans | ↑growth performances and survival rate | O. mykiss | [145] |
| Clostridium | |||
|
C. butyricum |
↑antibacterial activity against Vibriosis infection | O. mykiss | [146] |
| C. butyricum | ↑ immunity, regulate gut microbiota, antagonistic effects against Aeromonas sp., Vibrio sp., Pseudomonas sp. | C. carpio | [147] |
| Weissella | |||
| W. confusa | ↑growth performances | O. mykiss | [148] |
| W. confusa | ↑growth performances, antibacterial activity against A. hydrophila | Lates calcarifer | [149] |
| Other strains | |||
| A. veronii BA-1 | ↑immune system and antibacterial activity | C. carpio | [150] |
| Micrococcus luteus | ↑growth performances and feed conversion ratio | O. niloticus | [151] |
| Pseudoalteromonas undina VKM-124 | ↑survival and antiviral activity | Carangoides bartholomaei | [83] |
| Yeast | |||
| S.cerevisiae | ↑growth performances and resistance into waterborne Cu toxicity | Sarotherodon galilaeus | [152] |
| S. cerevisiae | ↑ immunity and ↓ mortality against P. fluorescens | Mystus cavasius | [153] |
| Yarrowia lipolytica | ↑ immune response, antioxidant status, and disease resistance against V. parahaemolyticus infection | Lutjanus peru | [154] |
| Multi-strain | |||
| B. subtilis and Bacillus licheniformis (BioPlus2B) | ↑ resistance against Y. ruckeri | O. mykiss | [155] |
|
Lactobacillus delbrueckii Lactobacillus rhamnosus L. plantarum B. bifidum |
↑growth performances and immunity | Acipenser baerii | [156] |
| Lactobacillus plantarum (STBL1), Saccharomyces cerevisiae (STBS1), and Bacillus safensis (SQVG18) | ↑growth, antioxidant capacity, digestion, and gut microflora | P. vannamei | [157] |
↓Decrease or reduce, ↑Increase or improve
Table 2.
Functional feed additives of prebiotics in aquatic animals.
| Prebiotics | Functions | Aquatic species | References |
|---|---|---|---|
| FOS | ↑growth, survival and gut microbiota section | L. vannamei | [184] |
| β-glucans | ↑growth, survival and immune system | Sparus aurata | [201] |
| MOS | ↑growth, immune system, antioxidant capacity and intestinal health | Cyprinus carpio | [219] |
| Chitosan | ↑growth, feed utilization, lipid metabolism, gut microbiota composition and immune system | Cyprinus carpio koi | [216] |
| Inulin | ↑growth, antioxidant capacity, immunity, and gut microbiota at low salinity | L. vannamei | [220] |
↓Decrease or reduce, ↑Increase or improve.
Table 3.
Some recent potential applications of postbiotics in aquaculture.
| Postbiotics | Microorganism producer | Aquatic species | Applications | References |
|---|---|---|---|---|
| Exopolysaccharides | Lactococcus lactis Z-2 | Common carp (C. carpio) |
Immunity enhancement Resistance against A. hydrophila |
[243] |
| Cell surface proteins | L. pentosus | Shrimp (Litopenaeus vannamei) |
Immune response improvement | [234] |
| Cell wall components (PGs and LTA) | B. pumilus SE5 | Grouper (E. coioides) |
Growth performance improvement Innate and adaptive immunity amelioration |
[93] |
| Lipoteichoic acids | L. plantarum LTA | Silvery pomfret (Pampus argenteus) |
Against V. anguillarum-caused vibriosis | [244] |
| Non live microorganisms | ||||
| S. cerevisiae, B. velezensis and Cetobacterium somerae | Common carp (C. carpio) |
Gut microbiota improvement Enhance non specific immunity Antioxidant status improvement |
[245] | |
| Dried autolyzed yeast | Gilthead sea bream (Sparus aurata) | Intestinal microbiota improvement | [246] | |
| Rhodotorula minuta and Cetobacterium somerae | Hybrid sturgeon (Acipenser baerii x Acipenser schrencki) |
Growth performance improvement Non-specific immunity improvement |
[197] | |
| Heat killed L. plantarum L-137 | Nile tilapia (O. niloticus) |
Growth performance stress resistance and immunity enhancement | [247] |
Table 4.
Functional feed additives of synbiotics in aquatic animals.
| Synbiotics | Functions | Aquatic organisms | References |
|---|---|---|---|
| P.acidilactici + GOS | ↑growth, survival and digestive enzyme function | Labidochromis lividus | [260] |
| B. clausii + FOS, MOS | ↑growth, survival and digestive enzyme function | Paralichthys olivaceus | [261] |
| P.acidilactici + GOS | ↑immunity and antagonistic activity against S. iniae infections | Oncorhynchus mykiss | [262] |
| B. subtilis + L. acidophilus + S. cerevisiae + FOS | ↑growth, and feed efficiency ratio | Eriocheir sinensis | [263] |
| P.acidilactici + IMO | ↑growth, immune response, and antioxidant capacity | C. carpio | [264] |
↓Decrease or reduce, ↑Increase or improve.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



