Submitted:
01 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
Herpesviridae is a family of enveloped double-stranded DNA viruses that cause a wide range of diseases in hosts. Among the various components of herpesvirus particles, tegument proteins residing between the envelop and nucleocapsid play crucial roles in viral replication, immune evasion, and modulation of host cellular pathways. Understanding the three-dimensional structure of herpesvirus tegument proteins help to reveal the molecular mechanism underlying the crosstalk with other viral and cellular components, and thus is crucial for the investigation of their biological and pathological functions. In this review, we summarize the current knowledge of the structural features of herpesvirus tegument proteins, highlighting the structure-based functional implications, including the potential as targets for antiviral drug development.
Keywords:
Herpesviruses
; Tegument proteins
; Structure
; Viral pathogenesis
1. Introduction
Herpesviruses are complex pathogens that have co-evolved with their hosts, employing an array of proteins to modulate host cellular processes and establish lifelong latent infections1,2. Herpesviridae comprises three subfamilies. Representatives of each subfamily, which are all infectious to humans, are human simplex virus (HSV) and varicella-zoster virus (VZV) for α-Herpesvirinae; human cytomegalovirus (HCMV) for β-Herpesvirinae; and Epstein-Barr Virus and Kaposi’s Sarcoma virus (KSHV) for γ-Herpesvirinae. Herpesviruses share a common virion structure: a DNA core in a nucleocapsid, an outer membranous envelope with glycoprotein spikes, and a tegument between the nucleocapsid and envelope (Figure 1). Being a unique structure of herpesvirus, the tegument is constituted by a number of proteins with various sizes and structures, including large multifunctional proteins such as UL363 and BPLF14, and small regulatory factors like UL245. These proteins form a complex network that contributes to the overall stability and infectivity of viral particles. ~30 tegument proteins have been identified in Herpesviridae (Table 1), Most of these tegument proteins are common to all three herpesvirus subfamilies, but others are subfamily-specific. Taking HSV-1 tegument proteins as examples, UL7, UL11, UL13, UL14, UL16, UL17, UL21, UL23, UL25, UL36, UL37, UL51 have homologs in other herpesviruses. Three tegument proteins, BKRF4 (homolog of KSHV ORF45), BLRF2 (ORF52), and BNRF1 (ORF75), are unique to γ-Herpesvirinae.
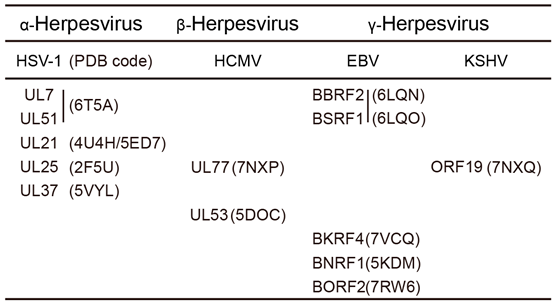
Over the past 20 years, the structures of herpesvirus glycoproteins have been extensively studied6. Recent development of cryo-electron microscopy (cryo-EM) allowed visualization of entire herpesvirus nucleocapsids of all three subfamily members at high resolution7–11. Compared to glycoproteins and nucleocapsids, the structural knowledge of tegument proteins is relatively sparse. Apparent reasons include that many tegument proteins contain rich disordered regions hindering crystallization, and most of them are too small to be unambiguously identified or discerned in cryo-EM studies. To date, only 11 tegument proteins have experimental structural annotations, which are summarized in Table 2.
Crystal structures of tegument proteins across the Herpesviridae family provide valuable insights into their intricate roles in various stages of viral life cycle. UL7/UL51 in HSV, UL103/UL71 in HCMV and BBRF2/BSRF1 in EBV form a multimeric complex that actively involved in viral replication, virion egress and secondary envelopment12–15. UL21 in HSV and BcRF1 have been reported that play a role in viral replication, B cell growth and secondary envelopment16,17. HSV UL25, HCMV UL77 and EBV BVRF1 have been illuminated in processes ranging from viral uncoating and capsid assembly to their membership in the capsid-associated tegument complex and involvement in nuclear egress.7,9,18,19. HSV UL37 interacts with capsids and other partners, such as UL36, another essential tegument protein, contributing to nucleocapsid transport and secondary envelopment20,21. pp150 associates with the capsid in viral assembly, which is essential for efficient HCMV replication22. Some tegument proteins are tightly associated with viral nucleocapsids, as part of them were discovered in single-particle reconstructed structures of herpesvirus capsids by cryogenic-election microscopy (cryo-EM)7–9. These tegument proteins are thus named capsid-associated tegument complexes (CATCs), which include UL17, UL25, UL36 in HSV, UL32/pp150 in HCMV, and BGLF1, BVRF1, and BPLF1 in EBV.
By elucidating the crystal structures of these proteins, researchers have gained insights into their molecular mechanisms and their significance in viral pathogenesis. In this article, we review the current structural data for herpesvirus tegument proteins, and discuss how the structural information aids the functional and translational research on Herpesviridae.
2. Structures of Common Herpesvirus Tegument Proteins
2.1. HSV UL7/UL51 and Homologs
The UL7-UL51 interaction was found first in HSV23 and later proved a conserved event for all three herpesvirus subfamilies, including EBV BBRF2-BSRF112 and HCMV UL103-UL7124. Loss of either UL7 or UL51 reduces the virus replication of HSV-1 and affects viral secondary envelopment25–30. Similarly, HCMV UL103 and UL71, as well as EBV BBRF2 and BSRF1, were found crucial for viral infection, secondary envelopment and virion egress13,31. In addition, HCMV UL103 was reported to interact with other HCMV proteins, such as tegument protein UL71, UL47 and capsid protein UL8632.
Recently, structural studies on tegument proteins UL7-UL51 in HSV-1 and BBRF2-BSRF1 in EBV have been characterized. UL7-UL51 forms 1:2 hetero-trimers in solution and further oligomerizes 2:4 hetero-hexamers at higher protein concentrations. Unlike this observation in solution, however, UL7 and UL51 stack in a 4:4 hetero-octamer in the crystal lattice, which is mediated by an intermolecular central β-barrel. As part of this β-barrel was encoded by the sequence derived from the vector, the 4:4 hetero-octamer observed in the crystal structure may not represent the physiological assembly of UL7-UL51 complex (Figure 2A). In the basic UL7-UL51 hetero-dimer, UL7 (containing residues 11–296) folds into a compact globular architecture, and UL51 (containing residues 41–125) includes two anti-parallel helices stabilized by hydrophobic interactions (Figure 2B). In addition, previous study reported the association between UL51 and UL14 and the importance of this association in viral envelopment33. The UL51-UL14 interaction could be abolished when residues Ile111, Leu119 and Tyr123 of UL51 were mutated.
Besides HSV-1 UL7 and UL51, crystal structures of EBV BBRF2 and BSRF1 were also reported. The crystal structure of BBRF2 was determined at high resolution as a monomer. In comparison, the solo structure of UL7 was not available, because UL7 tends to aggregate in solution23. The globular BBRF2 contains a central 6-stranded β-sheet surrounded by 10 helices as UL7, representing a new fold (named herpesvirus tegument fold 1, or HTF1; also mentioned as ‘conserved UL7 tegument assembly/release domain’, or CUSTARD fold, for HSV-1 UL7) that has not been observed for other proteins in the structural database. An uncommon feature of BBRF2 is that as a cytosolic protein, it has a hydrophilic core centered by an arginine (Arg72) residue conserved in γ-herpesvirinae. The 1.6 Å resolution allowed the discovery of a solvent channel filled with water molecules from this buried Arg27 to the surface of BBRF2 (Figure 2C), but the biological role of this special folding remains unclear. The BSRF1 portion solved in the crystal structure was a naturally degraded form (residues 41–139, noted BSRF1Δ) that contains two anti-parallel helices as observed for UL51, plus a C-terminal short helix perpendicular to the anti-parallel helices. forms dimers in solution. BBRF2 does not show significant conformational change before and after complexing with BSRF1, except for the relocation of an N-terminal loop. In the asymmetric unit of the crystal, BBRF2 and BSRF1Δ form a 6:6 hetero-dodecamer, which is composed of two triangular 3:3 hetero-hexamers stacked face-to-face (Figure 2D). Despite this somewhat complicated assembly, the basic unit of BBRF2-BSRF1Δ complex was thought to be a hetero-dimer as UL7-UL51. The association manners of basic HSV-1 UL7-UL51 hetero-dimer and EBV BBRF2-BSRF1Δ hetero-dimer are similar, both dominated by massive hydrophobic interactions. The N-terminal loop and two α-helices of UL7 and BSRF1 all participate in the contact with UL51 and BBRF2, respectively. Interestingly, several hydrophobic residues involved in the hetero-dimeric interfaces are only conserved in α- and γ-Herpesvirinea, but not β-Herpesvirinae. Structural information for UL51/UL7 homologs in β-Herpesvirinae (UL71/UL103) is unavailable. In addition, the differences in length and amino acid sequence are quite prominent at N- and C-terminus of UL51/UL7 and homologs among the Herpesviridae.
2.2. Functional Hints from the UL7-UL51 and BBRF2-BSRF1 Complex Structure
Several studies revealed membrane residence of UL51 and BSRF1, with a specific localization on the Golgi apparatus, which is conducted by the palmytoilation of a conserved cysteine at the N-terminus31,34. BSRF1 can translocate cytosplamically localized BBRF2 to the Golgi apparatus, and also binds viral glycoproteins such as gB and gH/gL. On the other hand, BBRF2 was found to interact with the components of viral nucleocapsid including MCP and BPLF135. Thus, UL7-UL51 and BBRF2-BSRF1 complexes mediate the association of nucleocapsid to the Golgi apparatus where the envelope with glycoproteins is acquired, thereby facilitating viral secondary envelopment.
For viral secondary envelopment, it is obvious that membrane remodeling events are essential to ensure the enwrapping of viral nucleocapsid. An interesting issue to note is the oligomerization tendency of UL7-UL51 and BBRF2-BSRF1 complexes. More than the crystallographic multimers mentioned in the previous section, UL7 and UL51 were found to form hetero-complexes with different stoichiometric ratios with the potential for further oligomerization23, and near-full-length BSRF1 with intact C-terminus is a dimer rather than a monomer as BSRF1Δ35. The BSRF1 homolog in HCMV, UL71, contains a basic leucine zipper (bZIP)-like domain, which accounts for the oligomerization of UL71 and is essential for viral envelopment36. In addition, UL51 and BSRF1 were suggested to possess similar folding with membrane shaping proteins like charged multivesicular body protein (CHMP) family members. Whether this similarity and the oligomerization feature implicate a membrane remodeling function of UL7-UL51 and BBRF2-BSRF1 complexes require further structural investigations using cryo-electron tomography (cryo-ET) on liposome-based systems or in situ.
2.3. HSV UL21 and Homologs
UL21 plays multiple roles in the viral replication, secondary envelopment and cell-cell spread through binding of the tegument proteins UL16, UL11, and capsid in the nucleus or cytoplasm17,37–40. The UL21 homolog in EBV is BcRF1. BcRF1 and BGLF3 make up the viral preinitiation complex (vPIC). The vPIC is responsible for initiating the synthesis of late transcripts by binding to specific promoter elements (the TATT motif) within the late gene promoters during the productive cycle of herpesviruses. The vPIC interacts with cellular RNA polymerase II, which is a crucial enzyme responsible for transcribing genes into messenger RNA (mRNA). This interaction facilitates the transcription process, enabling RNA polymerase II to start the synthesis of late viral transcripts41–43. HCMV UL87 has not been extensively investigated. In the only study on UL87, it was shown to be a late transcription factor (LFT) that activates viral early to late transcription via targeting the TATT sequence in viral promotors44.
The full-length structure of UL21 is currently unavailable, and structures of truncated versions containing either N- or C-terminal portions of UL21 were individually reported. The crystal structure of the UL21 N-terminal portion (UL21N, residues 1–216) possesses a novel sail-like fold that comprises a β-bouquet surrounded by four α-helices and two 310 helices (Figure 3A). The β-bouquet is composed of three piles of β-sheets. The largest pile contains 7 β-strands, facing the other two linearly aligned smaller piles. Evolutionary trace analysis (ETA) revealed several surface regions of UL21N which may be involved in the intramolecular contact with UL21C, or the association with its partner UL1645.
HSV-1 UL21C (residues 275–535) exhibits a helical-rich feature. Two helical wings, each containing five helices, flank a central long α-helix to form a dragonfly-like architecture. (Figure 3B). Three acidic patches are found on the surface of HSV-1 UL21C. One acidic patch is constituted by conserved residues and was thus considered an important region for common functions of UL21. The other two acidic patches, distributed on two helical wings, are not conserved and thought to account for virus-specific functions. Structural comparison analysis of conserved residues revealed the importance of N395 in virulence. Apart from the HSV-1 version, the structure of UL21C in pseudorabies virus (PRV), another α-herpesvirus, was also predicted. Three naturally-occurred mutations in PRV UL21, namely H37R, E355D, and V375A, were found to limit the spread and virulence of PRV46,47. Structural analysis of corresponding residues has revealed the ways these mutations affect PRV in a virus-specific manner, either by destabilizing the folding of UL21, or by disrupting its binding with other partners37,48,49.
UL21 plays a pivotal functional role in the replication and pathogenesis in HSV lies in its interaction with UL1650 and its binding to capsid51,52. And UL21 exhibits potential RNA-binding activity through the basic patch region53. The structural model proposes that UL21 consists of two specific domains connected by a flexible linker, each contributing to various functions in the virus life cycle. Notably, UL21N has much acidic regions, while UL21C possess basic character, which may facilitate the association between UL21N and UL21C via electrostatic interactions. The basic patch surface of UL21C is crucial for binding to capsid and nuclear localization. However, the precise interplay between UL21N and UL16 in binding to UL21C remains elusive. The resolved 3-dimensional structures of both UL21N and UL21C lay the foundation for targeted explorations into the diverse roles of UL21 in α-herpesvirus replication and pathogenesis. Prospective resolution of the full-length protein is expected to yield even more valuable insights.
2.4. HSV UL25 and Homologs
UL25 is renowned for its vital function in viral genome uncoating and capsid egress, yet it does not engage in the cleavage of newly replicated viral DNA or DNA encapsidation18,54,55. HSV-1 packages its DNA genome into capsids during virus assembly. Three capsid-associated DNA-packaging proteins, UL25, UL17 and UL16 are involved in this process. UL25 is added to the capsids later after the formation of the procapsids that composed of UL17 and UL16. UL25 may also play a role in stabilizing UL17 within capsids56,57. HCMV UL77, a capsid-associated structure protein in HCMV, is a homolog of HSV-1 UL25. Beside the conserved function in DNA packaging and stabilizing the capsid, it has a role in interacting with terminase subunits (UL56, UL89) during the virus infection cycle19. Another HSV-1 UL25 homolog, ORF19 in KSHV, also belongs to the capsid-associated tegument proteins and exhibits a penton-binding globular region in the KSHV virion structure58. In addition, OFR19, along with ORF32, constitutes the capsid vertex-specific component (CVSC), playing a crucial role in the assembly of viral capsids59.
In 2006, Bowman et al reported the crystal structure of a truncate HSV-1 UL25 construct that contains the C-terminal potion (residues 134–580). The structure of UL25134–580 is in a globular shape composed of dominantly α-helices (Figure 4A). Electrostatic surface analysis reveals distinct regions on the protein surface, with one surface featuring a cluster of negatively charged residues and the other displaying a multitude of positively charged residues (Figure 4B). These regions with opposing charges may be crucial for mediating homo-oligomerization of UL25, and/or interactions with other proteins including the viral capsid. Moreover, the positively-charged patch may be involve in binding viral DNA, and thus facilitating DNA encapsidation. However, due to insufficient mutagenesis-based biochemical analysis, the importance of the above regions or certain residues is still not clear for HSV-1 UL2555.
The UL25134–580-equivalent structures of HCMV UL77 and KSHV ORF19 have been subsequently reported. Crystal structure of HCMV UL77 and KSHV ORF19 reveal a flexible globular fold consisting rich α-helices and a few β-sheets (Figure 4C,D). However, KSHV ORF19 exhibits a pentameric conformation that mediated via hydrophobic interactions, hydrogen bonds and a salt bridge between adjacent protomers (Figure 4E). Notably, the extended loop of short β-strand between the adjacent protomers involves in stabilizing the pentamer. It may also extend to the ORF25 and ORF65 groove to comprise the penton vertex. While this pentameric arrangement likely caps the portal channel, it is distinct from the penton associated CVSC in KSHV. Compared with the homolog HSV UL25, the KSHV ORF19 pentamer exhibits abundant positively charged regions. This distribution of charged residues are consistent with electrostatic interactions that contribute to DNA passage through the portal vertex60. The conserved crystal structure of KSHV ORF19 and its homologs within the herpesvirus subfamilies (HSV UL25 and HCMV UL77) implies a conserved function of the pentameric assembly in capping the portal, facilitating the packaging of viral DNA genomes into capsids. Furthermore, the fitting of the pentameric ORF19 into the KSHV portal vertex suggests a related physiological role. Mutagenesis targeting its interfaces disrupt pORF19 pentamerization, severely impacting KSHV capsid assembly and progeny production. These findings provide insights into the role of ORF19 in capsid assembly and identify a potential novel drug target for herpesvirus-related diseases61.
BVRF1, along with BGLF1 and BPLF1, constitutes the EBV capsid-associated tegument complexes (CATC). The CATC of all herpesviruses links the nucleocapsid and tegument proteins. Within the EBV CATC, BVRF1 protein is composed of two distinct regions: an N-terminal Domain and a Helix Domain. The N-terminal domain has two conformers: one spans amino acids 10–36 (upper conformer), and the other spans amino acids 20–36 (lower conformer). BVRF1 also contains a Helix domain, with one spanning amino acids 37–93 (upper conformer) and the other spanning amino acids 37–87 (lower conformer). In the CATC model, these BVRF1 conformers are arranged in a helix bundle configuration, which is situated on the BGLF1 molecule. Additionally, the head domains of BVRF1 are observed as globular densities within the complex, with the left-side one being more prominent than the right-side one. It’s noteworthy that the BVRF1 head domain interacts with two neighboring penton major capsid protein (MCP) molecules within the CATC, primarily through the region of residues 291–314. This interaction contributes to the overall stability and organization of the Capsid-Associated Triplex Complex9,62. Although the specific roles of EBV BVRF1 remain unclear due to the lack of a complete structural basis, it is conceivable that BVRF1 may engage in viral packaging63, nucleocapsid uncoating64, and nuclear export65 like UL25.
2.5. HSV UL37 and Homologs
UL37 has been implicated in several key processes during the viral life cycle. UL37 is known to facilitate nucleocapsid transport20 and secondary envelopment by establishing connections between the capsid and the membrane-associated complex gK-pUL2021or by interacting with UL3666. Furthermore, UL37 has been found to interfere with the host’s antiviral responses via deamidating RIG-I67 and cGAS68. While the functional roles of UL37 in HSV-1 are well-studied, research on its homologs in other herpesviruses such as HCMV UL47, EBV BOLF1, and KSHV ORF63 remains limited. Loss of BOLF1 did not alter DNA synthesis or protein expression of EBV, but reduced viral infectivity69. KSHV ORF63 may inhibit innate immunity response by modulating inflammasome activation70.
To date, structural information is available only for the N-terminal part of UL37 (UL37N). UL37N is predominantly α-helical, and exhibits a two-bundle hairpin-like architecture. Structural comparison between HSV-1 UL37N (residues 1–575, H37N) and PrV UL37N (residues 1–479, P37N) reveal differences in specific regions (Figure 5A,B). One striking difference lies in the loops on the surface of the molecule in domains dII and dIII of H37N, while P37N features a short loop and a short helix forming a small pocket. Additionally, H37N exhibits a two-stranded sheet at the top of domain dIII, whereas P37N possesses two long outward loops linked to a helix, giving it a pointy top (Figure 5C,D)71. These divergent structural elements are likely to mediate virus-specific functions via engaging different host and/or viral binding partners, and this point needs to be verified by structural information of UL37N in other herpesviruses (e.g. EBV BOLF1 and KSHV ORF63). Furthermore, structural studies on the rest part of UL37 other than the N-terminal region is required to uncover new insights into their functions.
2.6. HSV Capsid-Associated Tegument Proteins and Homologs
Capsid-Associated Tegument Complex (CATC) is composed of specific tegument proteins that are involved in various stages of the viral life cycle, including replication, assembly, interaction with partner, and immune evasion12,72–74. This complex is responsible for linking the nucleocapsid, which contains the viral genome, to the tegument proteins and the viral membrane. In HSV-1, the CATC comprises of UL17, UL25, and UL36. UL17 and UL25 are tegument proteins involved in capsid assembly and nuclear egress18,75–77, while UL36 is a multifunctional tegument protein that participates in virion morphogenesis, egress, and nuclear targeting of the viral DNA78. Homologs of these proteins are also indispensable components in the CTACs of HCMV (UL93, UL77 and UL48) and EBV (BGLF1, BVRF1 and BPLF1). In addition, HCMV CATC contains a specific pp150 tegument protein, which has been implicated in the stabilization of the capsid and may play a role in viral assembly and maturation8,22.
Recent cryo-EM studies have provided high-resolution insights into the structure and function of CATC in different herpesviruses. In HSV-1, CATC are found present in the portal and penton vertices of the capsid. Each CATC has a UL17 core supporting a helix bundle made of UL25 and UL36. The UL17 monomer has distinct barrel and helix regions forming lobes, connected by a helix. An extended helix from UL17 guides the UL25-UL36 bundle, anchored by a hump and a hexon. The UL25 units in CATC have similar structures, binding to UL17 and triplex Tc. The UL36 dimer joins UL25 to form a four-helix coil resting on UL17 extended helix, creating the five-helix bundle. This arrangement is stabilized by hydrophobic interactions, with a break at Gln58-Arg59. This arrangement is important for nuclear capsid egress and axonal capsid transport7.
UL32/pp150, a unique tegument protein in HCMV, exists three conformers a, b and c. They attach to each triplex structure on the capsid and extend towards nearby proteins (MCPs), creating a net-like layer. An atomic model of the N-terminal part of pp150 (pp150nt) shows that it consists mainly of helical structures. These helices are arranged in upper and lower bundles connected by a central long helix. The rest of pp150 is flexible and not well-defined in the model, indicating variability in its shape. The initial 275 residues of pp150 (pp150nt) are sufficient for binding to the capsid. pp150 interacts with pentons and hexons, playing a role in stabilizing the capsid structure. The interaction between pp150nt and the capsid relies on a specific interaction between the cysteine tetrad and SCP protein. This interaction strengthens the DNA-containing capsid, making pp150 a crucial component in HCMV’s structure8.
In EBV, CATC is observed to interact with the capsid vertex connected to portal. The CATC comprises about 65% of full-length BGLF1 (331 out of 507 residues), along with two copies of the BVRF1 N-terminal region, and two copies of the BPLF1 C-terminal region. Within the CATC, BVRF1 molecules consist of distinct N-terminal and helix domains. forming a helix bundle with BGLF1. The head domains of BVRF1 are observed on one side of the bundle. The head domain of BVRF1 engages with two neighboring capsid proteins. BGLF1 is partitioned into front and back regions, facilitating the interaction with triplex Ta9.
Understanding the interactions and functions of these tegument proteins within the CATC is vital for unraveling the process of herpesvirus assembly, maturation, and egress. Targeting these interactions could potentially lead to the development of novel antiviral strategies to combat herpesvirus infections.
3. Structures of Subfamily-Specific Tegument Proteins
3.1. HCMV UL50 and UL53
Previous studies have showed the potential for developing antiviral drugs by targeting interactions within the HCMV Nuclear Egress Complex (NEC). Disrupting of these interactions are lethal to virus. In HCMV, the NEC is composed of UL50 and UL53, with a well-defined binding interface that can be targeted for drug discovery79,80. UL50 and UL53 play a crucial role in assisting the virus in nuclear egress during replication by forming the NEC and disrupting the nuclear lamina structure that surrounds the nucleus. Additionally, UL50 and UL53 recruit other kinase proteins, like protein kinase UL97, to facilitate this process81,82. Recently, three crystal structures were reported for UL53 (84–292), UL53 (72–292) and UL53 (72–292)-UL50 (1–169) complex. Structurally, UL53 can be partitioned into four conserved regions (CR1-CR4) (Figure 6A). each likely to confer distinct functional attributes. CR1 is implicated in nucleoplasmic subunit binding and contributes to zinc finger formation. In contrast, the precise biochemical roles of CR2–CR4, which remain relatively unexplored, may be inferred from analogous proteins, suggesting involvement in DNA packaging or protein interactions83. The construct of UL53 (84–292) lacks the anticipated region responsible for binding with UL50, whereas UL53 (72–292) encompasses the crucial residues known to facilitate UL50 binding, such as E75,L79 and M82 that around the zinc finger79. UL53 (84–292) and UL53 (72–292) have mostly same structures except the N-terminus loop. Importantly, UL53 (84–292) and UL53 (72–292) exhibit predominantly congruent structural characteristics. Both UL53 (72–292) and UL50 exhibit M50-like folding (Bergerat fold)84, characterized by a β bouquet enveloped by α-helices (Figure 6B). The UL53 (84–292) assemblage forms a heterodimer within the asymmetric unit (Figure 6C).
Regarding the Nuclear Egress Complex (NEC), UL50 establishes a clamping interaction with the helices of UL53, as evidenced by the contacts delineated in the UL53-UL50 complex structure (Figure 6D). UL53 does not exhibit substantial conformational changes before and after complexation with UL50, apart from the N-terminal helical. The NEC interaction interface is mediated by hydrophobic interactions84. The structure of the complex reveals a mechanism of hetero-dimerization, and the identified contacts may serve as a basis for designing targeted antiviral drugs. These findings also carry deep comprehension of nuclear egress85.
3.2. EBV BNRF1
Being a common target of CD4+ T cells, the major tegument BNRF1 assumes an important immune target in EBV86. BNRF1 is required for the selective viral gene expression during latency via interacting with death-domain associated protein (DAXX), a critical chaperone for histone H3.3 that plays a central role in the organization of chromatin structures87–89. Previously study has been reported that the C-terminal domains of BNRF1 is similar to the cellular purine biosynthesis enzyme FGARAT90. BNRF1 has DID domain, PurM-like domain and GATase domain as StFGARAT protein (Figure 7A). It engages with human antiviral proteins such as SP100 and DAXX through its N-terminal domains91. The complex structure of BNRF1 DAXX-interaction domain (DID) DAXX histone-binding domain (HBD) and histones H3.3-H4 via its N-terminal domain is determined lately. The structure of BNRF1 interacts with DAXX in a stable quaternary complex with histones H3.3-H4 dimer. BNRF1 embraces a β-sheet core fold enveloped by α-helices overlaying DAXX-H3.3-H4 complex. The interaction interface is facilitated by multiple loops embedded within the core fold (Figure 7B). The BNRF1 DID exhibits an overall fold akin to the PurM-like domain of the StFGARAT protein, except for the extended loop connecting the β-sheet90. This local variance is essential for binding to DAXX HBD—H3.3-H4. The BNRF1-DAXX interface is contributed to localize to PML-nuclear bodies that are involved in host-antiviral resistance and transcriptional repression. It is also indispensable in activating the transcription of latent cycle genes required for promoting B-cell proliferation. These findings illuminate the intricate strategy employed by tegument BNRF1 in harnessing the cellular antiviral histone chaperone DAXX, facilitating viral latency and cellular immortalization.
3.3. EBV BORF2
The large subunit of the EBV Ribonucleotide reductase (RNR), BORF2, interacts with cellular APOBEC3 enzymes (nuclear DNA cytosine deaminase), revealing an important role in inhibiting antiviral innate immunity92,93. BORF2 protects viral genomes from deamination during the lytic phase of DNA replication by suppressing the DNA deaminase activity of APOBEC3 enzymes. Recently, a complex structure of BORF2-AB3 was determined by cryo-EM. BORF2 exhibits distinctive attributes, including a short helix insertion (SHI) and a long loop insertion (LLI), both of which contribute to its unique functional characteristics (Figure 8A,B). The structure of BORF2 closely aligns with the RNRs of both E.coli and humans (Figure 8C). Several residues are highly conserved between BORF2 and class I RNRs, suggesting their involvement in ribonucleotide reduction. The residues within the SHI play a direct role in binding to A3B and facilitating the stability of complex interactions. Additionally, LLI, which is anchored to the RNR core, interacts with SHI, further contributing to A3B interaction. It is noteworthy that while certain class I RNRs encompass a conserved ATP cone domain in their N-terminal region, which regulates ATP activity94, this domain is absent in BORF2. This region in BORF2 contributes to mediate the novel dimer formation potentially facilitating contacts with BaRF1, another RNR subunit. The structural insights from two distinct complexes provide functional versatility. Specifically, the BORF2-BaRF1 complex may contribute to enhancing nuclear dNTP concentrations, while the BORF2-A3B complex likely plays a role in shielding viral genomic DNA during replication. In addition, the dimerization activity of BORF2 is essential to facilitates A3B re-localization from the nuclear to cytoplasmic95. This mechanism of pathogen-host interaction offers valuable prospects for drug development strategies that disrupt these interactions and provide the structure information to design DNA deaminase inhibitors.
3.4. EBV BKRF4
The Epstein-Barr virus tegument protein BKRF4, through its N-terminal acidic domain (BKRF4-N, residues 1–113), is shown to function as a histone chaperone. A specific segment within BKRF4-N (residues 15–113) is identified as the histone-binding domain (HBD). This HBD is responsible for binding to histones H2A-H2B, H3-H4, and cellular chromatin. BKRF4 also inhibits host’s DNA damage response (DDR) signaling by interacting with chromatin96,97. Recently, the structures of BKRF4-N in a co-chaperone complex with the histone chaperone ASF1b and H3.3-H4 dimer96 (Figure 9A) or in complex with H2A-H2B dimer were determined97 (Figure 9B). The co-chaperone complex likely plays a role in the manipulating the host chromatin by the virus. This unique binding mode enables BKRF4-HBD to interact with partially assembled nucleosomes, facilitating the disassembly of nucleosomes.
4. Conclusion
The structures of herpesvirus tegument proteins have revealed important functional insights. These structures have provided details on protein-protein interactions, viral-host interactions, and enzymatic activities, which contribute to viral replication, immune evasion, and pathogenesis. Further investigation of herpesvirus tegument protein structures which are not yet resolved will enhance our understanding of their roles in viral pathogenesis. Advances in cryo-EM and other cutting-edge techniques will contribute to determining the structures of more challenging tegument proteins. The structural insights gained from these structural studies will pave way for the development of targeted antiviral strategies and provide new opportunities for combatting herpesvirus infections.
Acknowledgments
This review article is dedicated to the late Prof. Jianguo Wu, our beloved mentor and a distinguished virologist. The structural study of BBRF2-BSRF1 complex mentioned in this article was presented by Dr. Hui-Ping He during Ph.D. defense as part of her thesis in 2020. As the head of the examination committee for Dr. He’s Ph.D defense, Prof. Wu has raised many constructive comments which greatly improved the study. This work was supported by grants from National Natural Science Foundation of China to H.-P.H. (32100971) and S.G. (82173098), Natural Science Foundation of Guangdong Province (2019TX05Y598).
Conflicts of Interest
The Authors declare no competing interests.
Materials & Correspondence:
Correspondence and requests for materials should be addressed to S.G. (gaosong@sysucc.org.cn) and H-P.H(hehp@sysucc.org.cn).
References
- Murata, T. & Tsurumi, T. Switching of EBV cycles between latent and lytic states. Reviews in Medical Virology (2014). https://doi.org/10.1002/rmv.1780. [CrossRef] [PubMed]
- Balfour, H. H., Dunmire, S. K. & Hogquist, K. A. Infectious mononucleosis. Clinical and Translational Immunology (2015). [CrossRef] [PubMed]
- Klupp, B. G., Fuchs, W., Granzow, H., Nixdorf, R. & Mettenleiter, T. C. Pseudorabies Virus UL36 Tegument Protein Physically Interacts with the UL37 Protein. J. Virol. 2002, 76, 3065–3071.
- Whitehurst, C. B. , Vaziri, C., Shackelford, J. & Pagano, J. S. Epstein-Barr Virus BPLF1 Deubiquitinates PCNA and Attenuates Polymerase Recruitment to DNA Damage Sites. J. Virol. 2012, 86, 8097–8106. [Google Scholar] [PubMed]
- Xu, H. , Su, C., Pearson, A., Mody, C. H. & Zheng, C. Herpes Simplex Virus 1 UL24 Abrogates the DNA Sensing Signal Pathway by Inhibiting NF-κB Activation. J. Virol. 91, (2017).
- Connolly, S. A. , Jardetzky, T. S. & Longnecker, R. The structural basis of herpesvirus entry. Nat. Rev. Microbiol. 2021, 19, 110–121. [Google Scholar]
- Dai, X. & Hong Zhou, Z. Structure of the herpes simplex virus 1 capsid with associated tegument protein complexes. Science 2018, 80, 360. [Google Scholar]
- Yu, X. , Jih, J., Jiang, J. & Zhou, Z. H. Atomic structure of the human cytomegalovirus capsid with its securing tegument layer of pp150. Science 2017, 80, 356. [Google Scholar]
- Li, Z. et al. CryoEM structure of the tegumented capsid of Epstein-Barr virus. Cell Res. 2020, 30, 873–884. [CrossRef] [PubMed]
- Xinghong Dai, Danyang Gong, Hanyoung Lim, Jonathan Jih, Ting-Ting Wu, R. & Zhou, S. and Z. H. Structure and mutagenesis reveal essential capsid protein interactions for KSHV replication. Science (80-. ). 2018, 553, 521–525. [Google Scholar]
- Liu, Y.-T. , Jih, J., Dai, X., Bi, G.-Q. & Zhou, Z. H. Cryo-EM structures of herpes simplex virus type 1 portal vertex and packaged genome. Nature 2019, 570, 257–261. [Google Scholar]
- He, H. P. et al. Structure of Epstein-Barr virus tegument protein complex BBRF2-BSRF1 reveals its potential role in viral envelopment. Nat. Commun. 2020, 11, 1–14. [CrossRef]
- Al Masud, H. M. A. et al. Epstein-barr virus BBRF2 is required for maximum infectivity. Microorganisms 2019, 7. [CrossRef] [PubMed]
- Ahlqvist, J. & Mocarski, E. Cytomegalovirus UL103 Controls Virion and Dense Body Egress. J. Virol. 2011, 85, 5125–5135. [Google Scholar]
- Roller, R. J. & Fetters, R. The Herpes Simplex Virus 1 UL51 Protein Interacts with the UL7 Protein and Plays a Role in Its Recruitment into the Virion. J. Virol. 2015, 89, 3112–3122. [Google Scholar] [PubMed]
- Gruffat, H. , Kadjouf, F., Mariamé, B. & Manet, E. The Epstein-Barr Virus BcRF1 Gene Product Is a TBP-Like Protein with an Essential Role in Late Gene Expression. J. Virol. 2012, 86, 6023–6032. [Google Scholar] [PubMed]
- Sarfo, A. et al. The UL21 Tegument Protein of Herpes Simplex Virus 1 Is Differentially Required for the Syncytial Phenotype. J. Virol. 91, (2017). [CrossRef]
- Preston, V. G. , Murray, J., Preston, C. M., McDougall, I. M. & Stow, N. D. The UL25 Gene Product of Herpes Simplex Virus Type 1 Is Involved in Uncoating of the Viral Genome. J. Virol. 2008, 82, 6654–6666. [Google Scholar] [PubMed]
- Köppen-Rung, P. , Dittmer, A. & Bogner, E. Intracellular Distribution of Capsid-Associated pUL77 of Human Cytomegalovirus and Interactions with Packaging Proteins and pUL93. J. Virol. 2016, 90, 5876–5885. [Google Scholar]
- Pasdeloup, D. , McElwee, M., Beilstein, F., Labetoulle, M. & Rixon, F. J. Herpesvirus Tegument Protein pUL37 Interacts with Dystonin/BPAG1 To Promote Capsid Transport on Microtubules during Egress. J. Virol. 2013, 87, 2857–2867. [Google Scholar] [PubMed]
- Jambunathan, N. et al. Herpes Simplex Virus 1 Protein UL37 Interacts with Viral Glycoprotein gK and Membrane Protein UL20 and Functions in Cytoplasmic Virion Envelopment. J. Virol. 2014, 88, 5927–5935. [CrossRef]
- Liu, Y. T. , Strugatsky, D., Liu, W. & Zhou, Z. H. Structure of human cytomegalovirus virion reveals host tRNA binding to capsid-associated tegument protein pp150. Nat. Commun. 2021, 12, 1–9. [Google Scholar]
- Butt, B. G. et al. Structure of herpes simplex virus pUL7:pUL51, a conserved complex required for efficient herpesvirus assembly. bioRxiv 810663 (2019).
- Womack, A. & Shenk, T. Human Cytomegalovirus Tegument Protein pUL71 Is Required for Efficient Virion Egress. MBio 1, (2010).
- Tanaka, M., Sata, T. & Kawaguchi, Y. The product of the Herpes simplex virus 1 UL7 gene interacts with a mitochondrial protein, adenine nucleotide translocator 2. Virol. J. 2008, 5.
- Xu, X. et al. The mutated tegument protein UL7 attenuates the virulence of herpes simplex virus 1 by reducing the modulation of α-4 gene transcription. Virol. J. 2016, 13.
- Nozawa, N. et al. Herpes Simplex Virus Type 1 UL51 Protein Is Involved in Maturation and Egress of Virus Particles. J. Virol. 2005, 79, 6947–6956. [CrossRef]
- Albecka, A. et al. Dual Function of the pUL7-pUL51 Tegument Protein Complex in Herpes Simplex Virus 1 Infection. J. Virol. 2017, 91.
- Fuchs, W. et al. The UL7 Gene of Pseudorabies Virus Encodes a Nonessential Structural Protein Which Is Involved in Virion Formation and Egress. J. Virol. 2005, 79, 11291–11299. [CrossRef] [PubMed]
- Roller, R. J. , Haugo, A. C., Yang, K. & Baines, J. D. The Herpes Simplex Virus 1 UL51 Gene Product Has Cell Type-Specific Functions in Cell-to-Cell Spread. J. Virol. 2014, 88, 4058–4068. [Google Scholar] [PubMed]
- Yanagi, Y. et al. Initial characterization of the Epstein–Barr virus BSRF1 gene product. Viruses 2019, 11.
- Ortiz, D. A. , Glassbrook, J. E. & Pellett, P. E. Protein-Protein Interactions Suggest Novel Activities of Human Cytomegalovirus Tegument Protein pUL103. J. Virol. 2016, 90, 7798–7810. [Google Scholar] [PubMed]
- Oda, S. , Arii, J., Koyanagi, N., Kato, A. & Kawaguchi, Y. The Interaction between Herpes Simplex Virus 1 Tegument Proteins UL51 and UL14 and Its Role in Virion Morphogenesis. J. Virol. 2016, 90, 8754–8767. [Google Scholar]
- Nozawa, N. et al. Subcellular Localization of Herpes Simplex Virus Type 1 UL51 Protein and Role of Palmitoylation in Golgi Apparatus Targeting. J. Virol. 2003, 77, 3204–3216. [CrossRef]
- He, H. P. et al. Structure of Epstein-Barr virus tegument protein complex BBRF2-BSRF1 reveals its potential role in viral envelopment. Nat. Commun. 2020, 11, 5405. [CrossRef]
- Meissner, C. S. , Suffner, S., Schauflinger, M., von Einem, J. & Bogner, E. A Leucine Zipper Motif of a Tegument Protein Triggers Final Envelopment of Human Cytomegalovirus. J. Virol. 2012, 86, 3370–3382. [Google Scholar]
- Klupp, B. G., Böttcher, S., Granzow, H., Kopp, M. & Mettenleiter, T. C. Complex Formation between the UL16 and UL21 Tegument Proteins of Pseudorabies Virus. J. Virol. (2005). [CrossRef]
- Harper, A. L. et al. Interaction Domains of the UL16 and UL21 Tegument Proteins of Herpes Simplex Virus. J. Virol. 2010, 84, 2963–2971. [CrossRef]
- Le Sage, V. et al. The Herpes Simplex Virus 2 UL21 Protein Is Essential for Virus Propagation. J. Virol. 2013, 87, 5904–5915. [CrossRef] [PubMed]
- Han, J. , Chadha, P., Starkey, J. L. & Wills, J. W. Function of glycoprotein E of herpes simplex virus requires coordinated assembly of three tegument proteins on its cytoplasmic tail. Proc. Natl. Acad. Sci. 2012, 109, 19798–19803. [Google Scholar] [PubMed]
- Aubry, V. et al. Epstein-Barr Virus Late Gene Transcription Depends on the Assembly of a Virus-Specific Preinitiation Complex. J. Virol. 2014, 88, 12825–12838. [CrossRef]
- Li, J. , Walsh, A., Lam, T. T., Delecluse, H.-J. & El-Guindy, A. A single phosphoacceptor residue in BGLF3 is essential for transcription of Epstein-Barr virus late genes. PLOS Pathog. 2019, 15, e1007980. [Google Scholar]
- Gruffat, H. , Kadjouf, F., Mariamé, B. & Manet, E. The Epstein-Barr Virus BcRF1 Gene Product Is a TBP-Like Protein with an Essential Role in Late Gene Expression. J. Virol. 2012, 86, 6023–6032. [Google Scholar] [PubMed]
- Li, M. et al. Cytomegalovirus late transcription factor target sequence diversity orchestrates viral early to late transcription. PLoS Pathogens 2021, 17.
- Metrick, C. M. , Chadha, P. & Heldwein, E. E. The Unusual Fold of Herpes Simplex Virus 1 UL21, a Multifunctional Tegument Protein. J. Virol. 2015, 89, 2979–2984. [Google Scholar] [PubMed]
- Curanović, D. , Lyman, M. G., Bou-Abboud, C., Card, J. P. & Enquist, L. W. Repair of the U L 21 Locus in Pseudorabies Virus Bartha Enhances the Kinetics of Retrograde, Transneuronal Infection In Vitro and In Vivo. J. Virol. 2009, 83, 1173–1183. [Google Scholar] [PubMed]
- Klupp, B. G. , Lomniczi, B., Visser, N., Fuchs, W. & Mettenleiter, T. C. Viral Pathogens and ImmunityMutations Affecting the UL21 Gene Contribute to Avirulence of Pseudorabies Virus Vaccine Strain Bartha. Virology 1995, 212, 466–473. [Google Scholar] [PubMed]
- de Wind, N. , Wagenaar, F., Pol, J., Kimman, T. & Berns, A. The pseudorabies virus homology of the herpes simplex virus UL21 gene product is a capsid protein which is involved in capsid maturation. J. Virol. 1992, 66, 7096–7103. [Google Scholar] [PubMed]
- Yang, L. Duck enteritis virus UL21 is a late gene encoding a protein that interacts with pUL16. BMC Vet. Res. 2020, 16, 1–11. [Google Scholar] [CrossRef]
- Klupp, B. G. , Böttcher, S., Granzow, H., Kopp, M. & Mettenleiter, T. C. Complex Formation between the UL16 and UL21 Tegument Proteins of Pseudorabies Virus. J. Virol. 2005, 79, 1510–1522. [Google Scholar]
- de Wind, N. , Wagenaar, F., Pol, J., Kimman, T. & Berns, A. The pseudorabies virus homology of the herpes simplex virus UL21 gene product is a capsid protein which is involved in capsid maturation. J. Virol. 1992, 66, 7096–7103. [Google Scholar] [PubMed]
- Takakuwa, H. et al. Herpes simplex virus encodes a virion-associated protein which promotes long cellular processes in over-expressing cells. Genes to Cells 2001, 6, 955–966. [CrossRef] [PubMed]
- Metrick, C. M. & Heldwein, E. E. Novel Structure and Unexpected RNA-Binding Ability of the C-Terminal Domain of Herpes Simplex Virus 1 Tegument Protein UL21. J. Virol. 2016, 90, 5759–5769. [Google Scholar] [PubMed]
- Klupp, B. G. , Granzow, H., Keil, G. M. & Mettenleiter, T. C. The Capsid-Associated UL25 Protein of the Alphaherpesvirus Pseudorabies Virus Is Nonessential for Cleavage and Encapsidation of Genomic DNA but Is Required for Nuclear Egress of Capsids. J. Virol. 2006, 80, 6235–6246. [Google Scholar] [PubMed]
- Bowman, B. R. et al. Structural Characterization of the UL25 DNA-Packaging Protein from Herpes Simplex Virus Type 1. J. Virol. 2006, 80, 2309–2317. [Google Scholar] [CrossRef] [PubMed]
- Thurlow, J. K. , Murphy, M., Stow, N. D. & Preston, V. G. Herpes Simplex Virus Type 1 DNA-Packaging Protein UL17 Is Required for Efficient Binding of UL25 to Capsids. J. Virol. 2006, 80, 2118–2126. [Google Scholar] [PubMed]
- Ali, M. A. , Forghani, B. & Cantin, E. M. Characterization of an essential HSV-1 protein encoded by the UL25 gene reported to be involved in virus penetration and capsid assembly. Virology 1996, 216, 278–283. [Google Scholar]
- Dai, X. , Gong, D., Wu, T.-T., Sun, R. & Zhou, Z. H. Organization of Capsid-Associated Tegument Components in Kaposi’s Sarcoma-Associated Herpesvirus. J. Virol. 2014, 88, 12694–12702. [Google Scholar] [PubMed]
- Grzesik, P. et al. Incorporation of the Kaposi’s sarcoma-associated herpesvirus capsid vertex-specific component (CVSC) into self-assembled capsids. Virus Res. 2017, 236, 9–13. [CrossRef]
- Liu, Y.-T. , Jih, J., Dai, X., Bi, G.-Q. & Zhou, Z. H. Cryo-EM structures of herpes simplex virus type 1 portal vertex and packaged genome. Nature 2019, 570, 257–261. [Google Scholar] [PubMed]
- Naniima, P. et al. Assembly of infectious Kaposi’s sarcoma-associated herpesvirus progeny requires formation of a pORF19 pentamer. PLoS Biology vol. 19 (2021).
- Liu, W. et al. Structures of capsid and capsid-associated tegument complex inside the Epstein–Barr virus. Nat. Microbiol. 2020, 5, 1285–1298. [CrossRef] [PubMed]
- Heming, J. D., Conway, J. F. & Homa, F. L. Herpesvirus Capsid Assembly and DNA Packaging. in Microorganisms vol. 7 119–142 (2017).
- Draganova, E. B. , Zhang, J., Zhou, Z. H. & Heldwein, E. E. Structural basis for capsid recruitment and coat formation during HSV-1 nuclear egress. Elife 2020, 9, 7073–7082. [Google Scholar]
- McNab, A. R. et al. The Product of the Herpes Simplex Virus Type 1 UL25 Gene Is Required for Encapsidation but Not for Cleavage of Replicated Viral DNA. J. Virol. 1998, 72, 1060–1070. [CrossRef]
- Desai, P. J. A Null Mutation in the UL36 Gene of Herpes Simplex Virus Type 1 Results in Accumulation of Unenveloped DNA-Filled Capsids in the Cytoplasm of Infected Cells. J. Virol. 2000, 74, 11608–11618. [Google Scholar] [CrossRef]
- Zhao, J. et al. A Viral Deamidase Targets the Helicase Domain of RIG-I to Block RNA-Induced Activation. Cell Host Microbe 2016, 20, 770–784. [Google Scholar] [CrossRef]
- Zhang, J. et al. Species-Specific Deamidation of cGAS by Herpes Simplex Virus UL37 Protein Facilitates Viral Replication. Cell Host Microbe 2018, 24, 234–248. [Google Scholar] [CrossRef]
- Masud, H. M. A. Al et al. The BOLF1 gene is necessary for effective Epstein–Barr viral infectivity. Virology 2019, 531, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J. P. & Monie, T. P. Computational analysis predicts the Kaposi’s sarcoma-associated herpesvirus tegument protein ORF63 to be alpha helical. Proteins Struct. Funct. Bioinforma. 2012, 80, 2063–2070. [Google Scholar]
- Pitts, J. D. , Klabis, J., Richards, A. L., Smith, G. A. & Heldwein, E. E. Crystal Structure of the Herpesvirus Inner Tegument Protein UL37 Supports Its Essential Role in Control of Viral Trafficking. J. Virol. 2014, 88, 5462–5473. [Google Scholar] [PubMed]
- Whitehurst, C. B. et al. The Epstein-Barr Virus (EBV) Deubiquitinating Enzyme BPLF1 Reduces EBV Ribonucleotide Reductase Activity. J. Virol. 2009, 83, 4345–4353. [Google Scholar] [CrossRef]
- Sugimoto, A. , Yamashita, Y., Kanda, T., Murata, T. & Tsurumi, T. Epstein-Barr virus genome packaging factors accumulate in BMRF1-cores within viral replication compartments. PLoS One 2019, 14, 1–14. [Google Scholar]
- Lui, W. Y. et al. Suppression of cGAS- and RIG-I-mediated innate immune signaling by Epstein-Barr virus deubiquitinase BPLF1. PLoS Pathogens vol. 19 (2023).
- Madrigano, J. & Adam Moser and Darrin M. York, K. R. Labeling and localization of the herpes simplex virus capsid protein UL25 and its interaction with the two triplexes closest to the penton. J Mol Biol 2008, 23, 1–7. [Google Scholar]
- Trus, B. L. et al. Allosteric Signaling and a Nuclear Exit Strategy: Binding of UL25/UL17 Heterodimers to DNA-Filled HSV-1 Capsids. Mol. Cell 2007, 26, 479–489. [Google Scholar] [CrossRef]
- Toropova, K. , Huffman, J. B., Homa, F. L. & Conway, J. F. The Herpes Simplex Virus 1 UL17 Protein Is the Second Constituent of the Capsid Vertex-Specific Component Required for DNA Packaging and Retention. J. Virol. 2011, 85, 7513–7522. [Google Scholar] [PubMed]
- Newcomb, W. W. & Brown, J. C. Structure and Capsid Association of the Herpesvirus Large Tegument Protein UL36. J. Virol. 2010, 84, 9408–9414. [Google Scholar] [PubMed]
- Sam, M. D. , Evans, B. T., Coen, D. M. & Hogle, J. M. Biochemical, Biophysical, and Mutational Analyses of Subunit Interactions of the Human Cytomegalovirus Nuclear Egress Complex. J. Virol. 2009, 83, 2996–3006. [Google Scholar] [PubMed]
- Leigh, K. E. et al. Structure of a herpesvirus nuclear egress complex subunit reveals an interaction groove that is essential for viral replication. Proc. Natl. Acad. Sci. 2015, 112, 9010–9015. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M. , Kamil, J. P., Coughlin, M., Reim, N. I. & Coen, D. M. Human Cytomegalovirus UL50 and UL53 Recruit Viral Protein Kinase UL97, Not Protein Kinase C, for Disruption of Nuclear Lamina and Nuclear Egress in Infected Cells. J. Virol. 2014, 88, 249–262. [Google Scholar] [PubMed]
- Milbradt, J. , Auerochs, S., Sticht, H. & Marschall, M. Cytomegaloviral proteins that associate with the nuclear lamina: components of a postulated nuclear egress complex. J. Gen. Virol. 2009, 90, 579–590. [Google Scholar]
- Popa, M. et al. Dominant Negative Mutants of the Murine Cytomegalovirus M53 Gene Block Nuclear Egress and Inhibit Capsid Maturation. J. Virol. 2010, 84, 9035–9046. [Google Scholar] [CrossRef] [PubMed]
- Leigh, K. E. et al. Structure of a herpesvirus nuclear egress complex subunit reveals an interaction groove that is essential for viral replication. Proc. Natl. Acad. Sci. U. S. A. 2015, 112, 9010–9015. [Google Scholar] [CrossRef]
- Lye, M. F. et al. Unexpected features and mechanism of heterodimer formation of a herpesvirus nuclear egress complex. EMBO J. 2015, 34, 2937–2952. [Google Scholar] [CrossRef]
- Adhikary, D. , Damaschke, J., Mautner, J. & Behrends, U. The Epstein-Barr Virus Major Tegument Protein BNRF1 Is a Common Target of Cytotoxic CD4 + T Cells. J. Virol. 2020, 94, 93–95. [Google Scholar]
- Cohen, J. I. Epstein–Barr Virus Infection. N. Engl. J. Med. 2000, 343, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, P. M. Chromatin Structure of Epstein–Barr Virus Latent Episomes. in Cancers vol. 10 71–102 (2015).
- Schreiner, S. & Wodrich, H. Virion Factors That Target Daxx To Overcome Intrinsic Immunity. J. Virol. 2013, 87, 10412–10422. [Google Scholar] [PubMed]
- Huang, H. et al. Structural basis underlying viral hijacking of a histone chaperone complex. 2016, 7. [CrossRef]
- Tsai, K. , Messick, T. E. & Lieberman, P. M. Disruption of host antiviral resistances by gammaherpesvirus tegument proteins with homology to the FGARAT purine biosynthesis enzyme. Curr. Opin. Virol. 2015, 14, 30–40. [Google Scholar] [PubMed]
- Cheng, A. Z. et al. A Conserved Mechanism of APOBEC3 Relocalization by Herpesviral Ribonucleotide Reductase Large Subunits. J. Virol. 93, (2019). [CrossRef] [PubMed]
- Cheng, A. Z. et al. Epstein–Barr virus BORF2 inhibits cellular APOBEC3B to preserve viral genome integrity. Nat. Microbiol. 2018, 4, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Greene, B. L. et al. Ribonucleotide Reductases: Structure, Chemistry, and Metabolism Suggest New Therapeutic Targets. Annu. Rev. Biochem. 2020, 89, 45–75. [Google Scholar] [CrossRef] [PubMed]
- Shaban, N. M. et al. Cryo-EM structure of the EBV ribonucleotide reductase BORF2 and mechanism of APOBEC3B inhibition. Sci. Adv. 2022, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. et al. Epstein-Barr Virus Tegument Protein BKRF4 is a Histone Chaperone. J. Mol. Biol. 2022, 434, 167756. [Google Scholar] [CrossRef]
- Chen, J. et al. Epstein-Barr virus protein BKRF4 restricts nucleosome assembly to suppress host antiviral responses. Proc. Natl. Acad. Sci. U. S. A. 2022, 119, 1–10. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The architecture of Herpesviruses virion. The glycoproteins are anchored in viral envelope and pointing outwards. The viral genome is packed in the regularly shaped capsid. The tegument is the space between the viral envelope and capsid, and contains a variety of tegument proteins with various functions.
Figure 1.
The architecture of Herpesviruses virion. The glycoproteins are anchored in viral envelope and pointing outwards. The viral genome is packed in the regularly shaped capsid. The tegument is the space between the viral envelope and capsid, and contains a variety of tegument proteins with various functions.
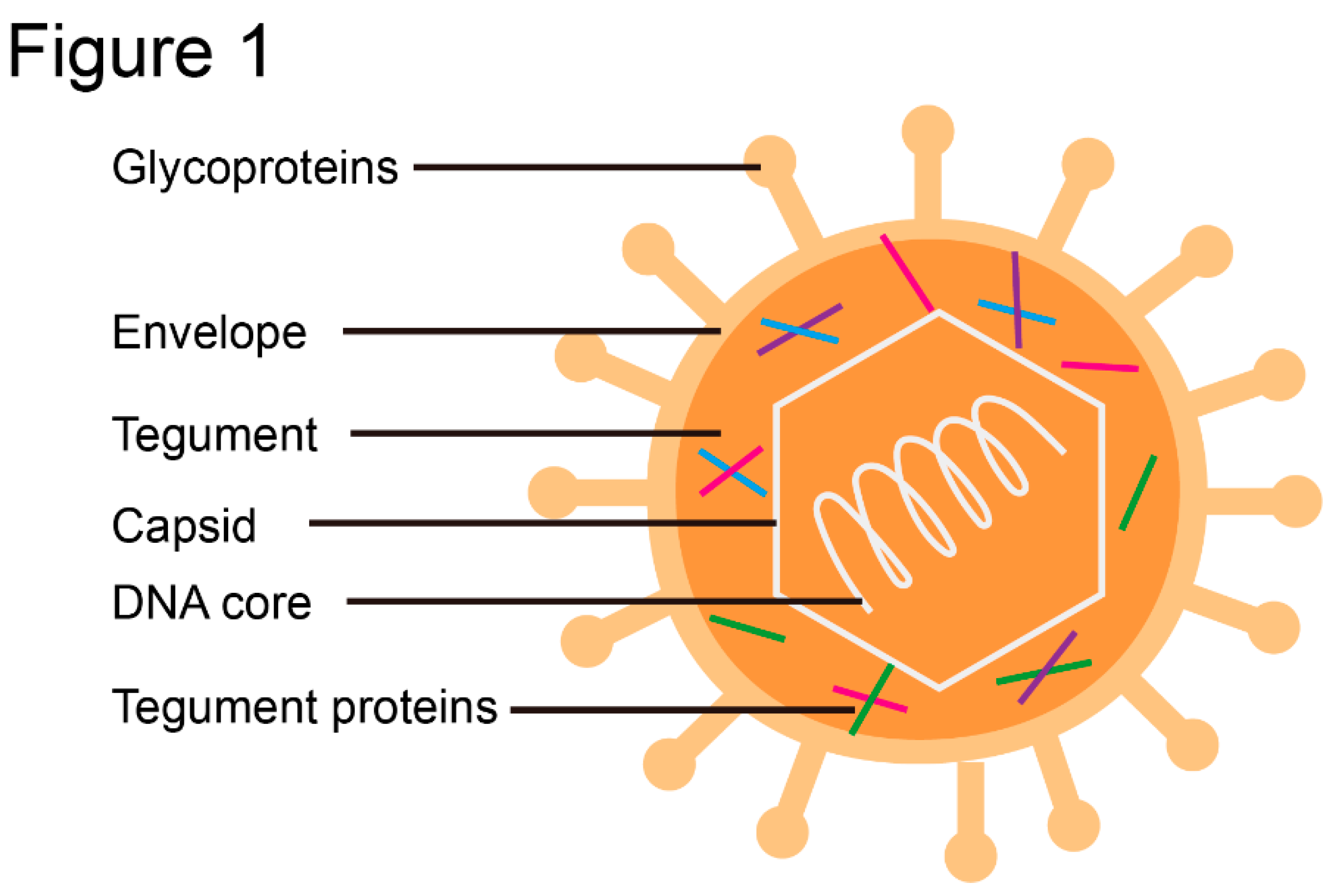
Figure 2.
Complex structures of HSV UL7-UL51Δ and EBV BBRF2-BSRF1Δ. A, Hetero-octamer of UL7-UL51 in the asymmetric unit. UL7 and pUL51 (8-142) are shown as cyan and pink. (Protein data bank code: 6T5A). B, Heterodimer of UL7-UL51 (41-125). C, Structure of BBRF2Δ. Helices and β-strands are coloured cyan and magenta, respectively. Details of the hydrophilic core of BBRF2Δ. D, Left to the 6:6 BBRF2Δ-BSRF1Δ heterododecamer in the asymmetric unit: the 3:3 heterohexamer. (PDB: 6LQO).
Figure 2.
Complex structures of HSV UL7-UL51Δ and EBV BBRF2-BSRF1Δ. A, Hetero-octamer of UL7-UL51 in the asymmetric unit. UL7 and pUL51 (8-142) are shown as cyan and pink. (Protein data bank code: 6T5A). B, Heterodimer of UL7-UL51 (41-125). C, Structure of BBRF2Δ. Helices and β-strands are coloured cyan and magenta, respectively. Details of the hydrophilic core of BBRF2Δ. D, Left to the 6:6 BBRF2Δ-BSRF1Δ heterododecamer in the asymmetric unit: the 3:3 heterohexamer. (PDB: 6LQO).
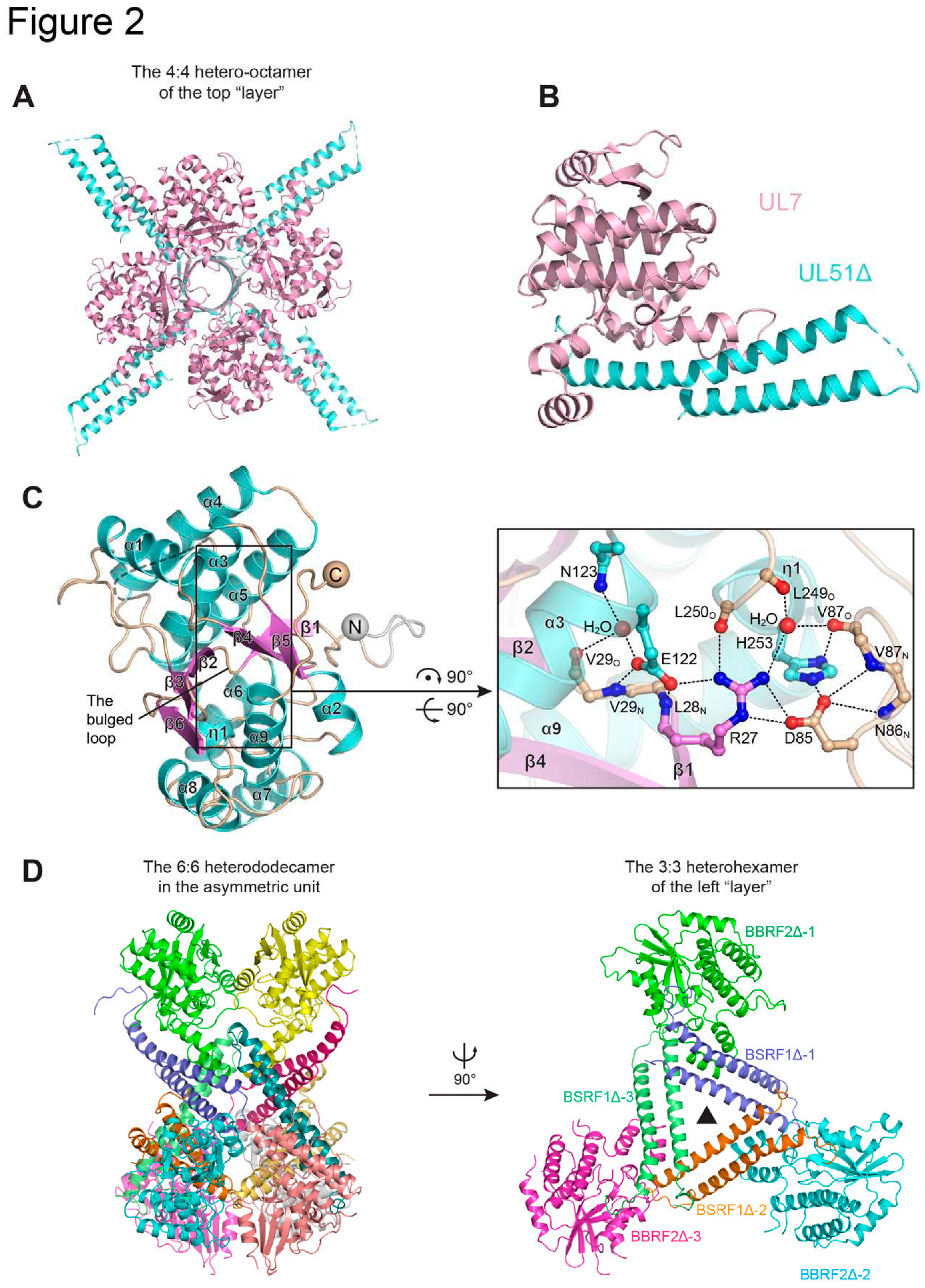
Figure 3.
Structures of HSV UL21. A, Structure of the N-terminal portion of UL21N (4U4H). B, Structure of the C-terminal portion of UL21C (5ED7).
Figure 3.
Structures of HSV UL21. A, Structure of the N-terminal portion of UL21N (4U4H). B, Structure of the C-terminal portion of UL21C (5ED7).
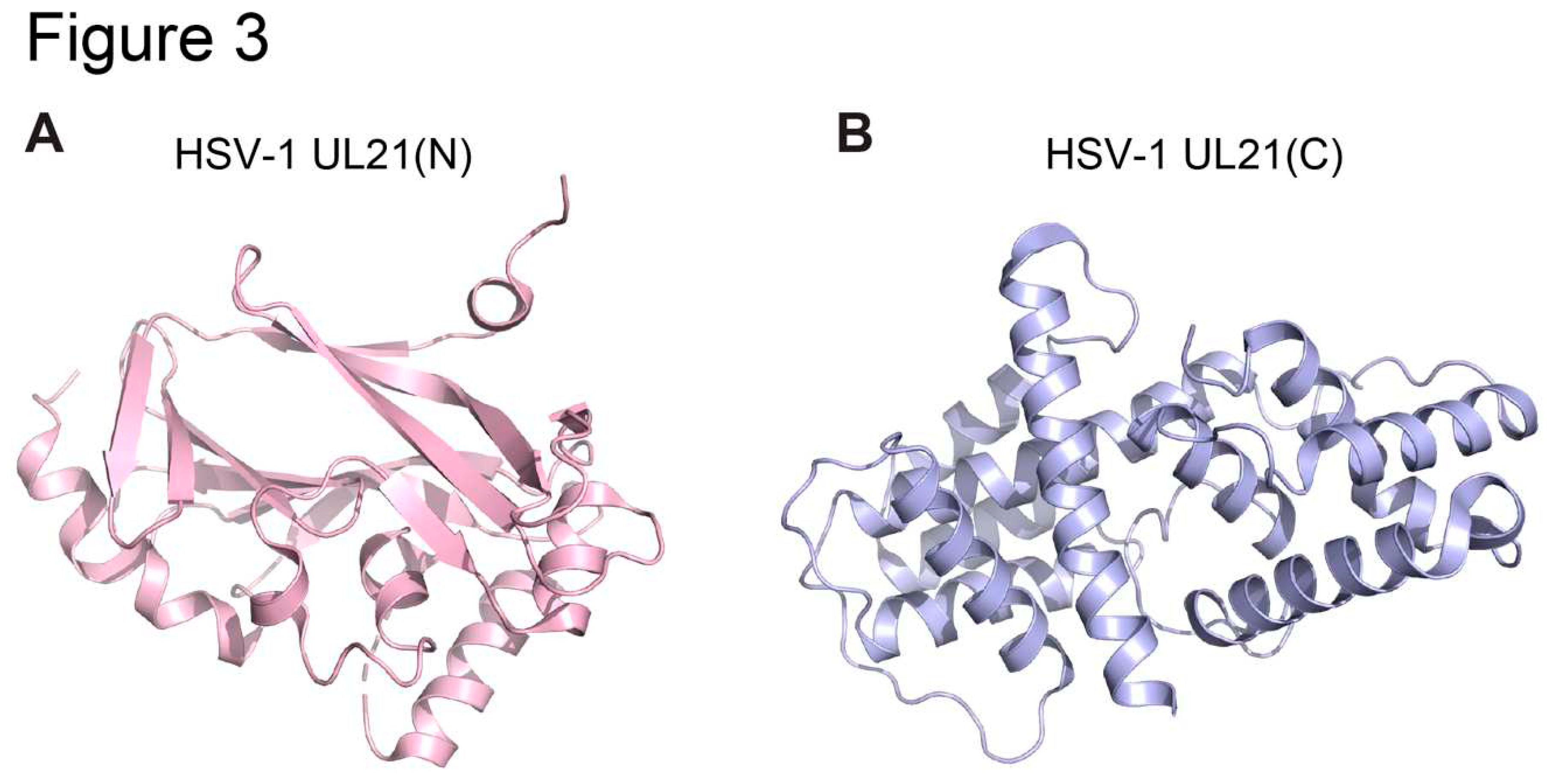
Figure 4.
Structure of HSV UL25 and homologs. A, Structure of HSV-1 UL25 (2F5U). B, Surface electrostatic of HSV-1 UL25. C, Structure of HCMV UL77 (7NXP). D, Structure of KSHV ORF19 (7NXQ). E, The pentamer structure of KSHV ORF19 in solution.
Figure 4.
Structure of HSV UL25 and homologs. A, Structure of HSV-1 UL25 (2F5U). B, Surface electrostatic of HSV-1 UL25. C, Structure of HCMV UL77 (7NXP). D, Structure of KSHV ORF19 (7NXQ). E, The pentamer structure of KSHV ORF19 in solution.
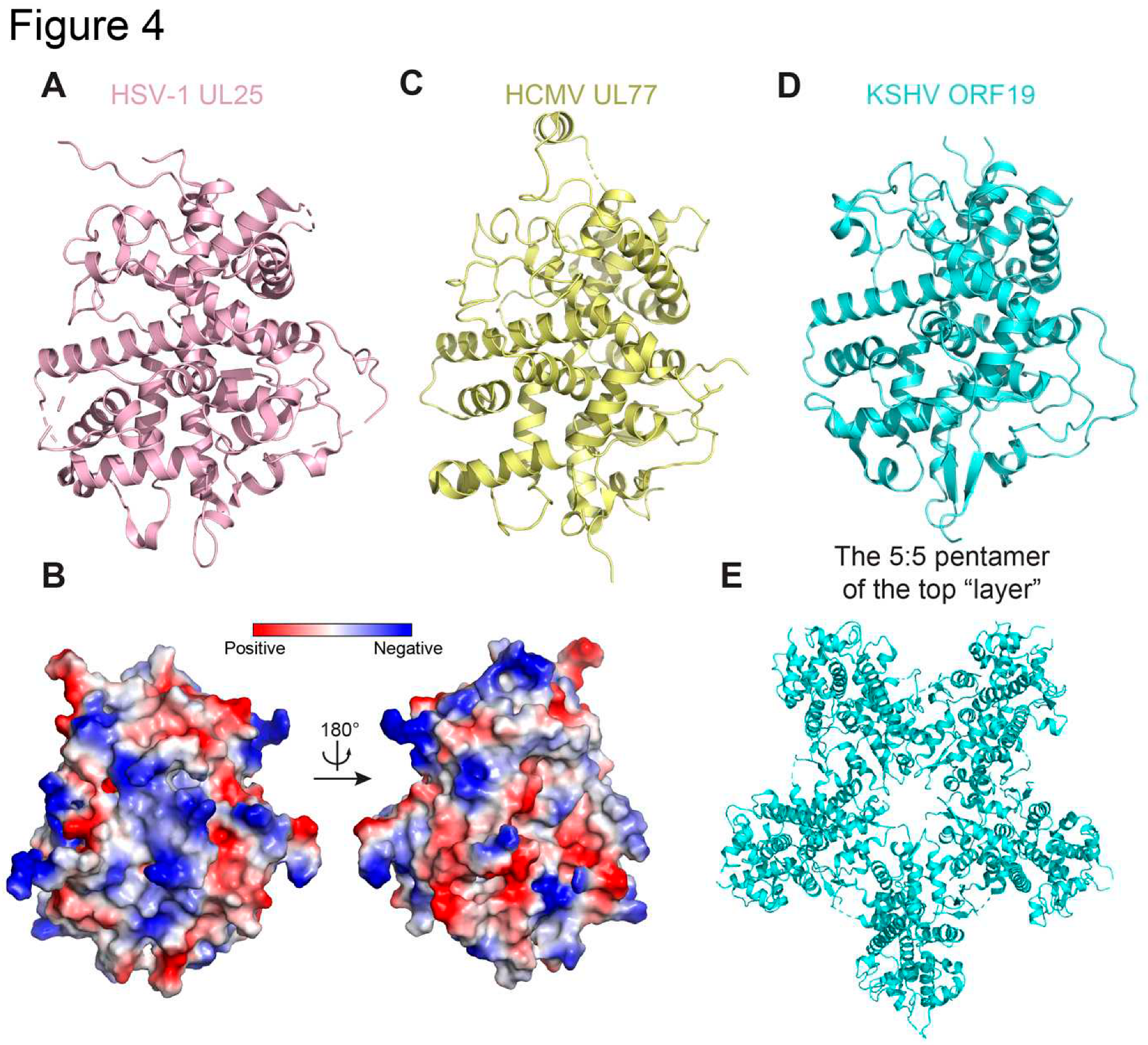
Figure 5.
Structure UL37 in α-herpesvirinae. A, Schematic representation showing the domain of full-length UL37 in HSV-1 and PrV. B, Overlay of the two structures of UL37 N-terminal domain in HSV-1 (PDB:5VYL) and in PrV ( 4K70). C, D Structural differences in dII domain (C) and dIII domain (D) between HSV-1 (PDB:5VYL) and in PrV. Structurally consistent regions are shown in transparent cartoons to highlight the difference.
Figure 5.
Structure UL37 in α-herpesvirinae. A, Schematic representation showing the domain of full-length UL37 in HSV-1 and PrV. B, Overlay of the two structures of UL37 N-terminal domain in HSV-1 (PDB:5VYL) and in PrV ( 4K70). C, D Structural differences in dII domain (C) and dIII domain (D) between HSV-1 (PDB:5VYL) and in PrV. Structurally consistent regions are shown in transparent cartoons to highlight the difference.
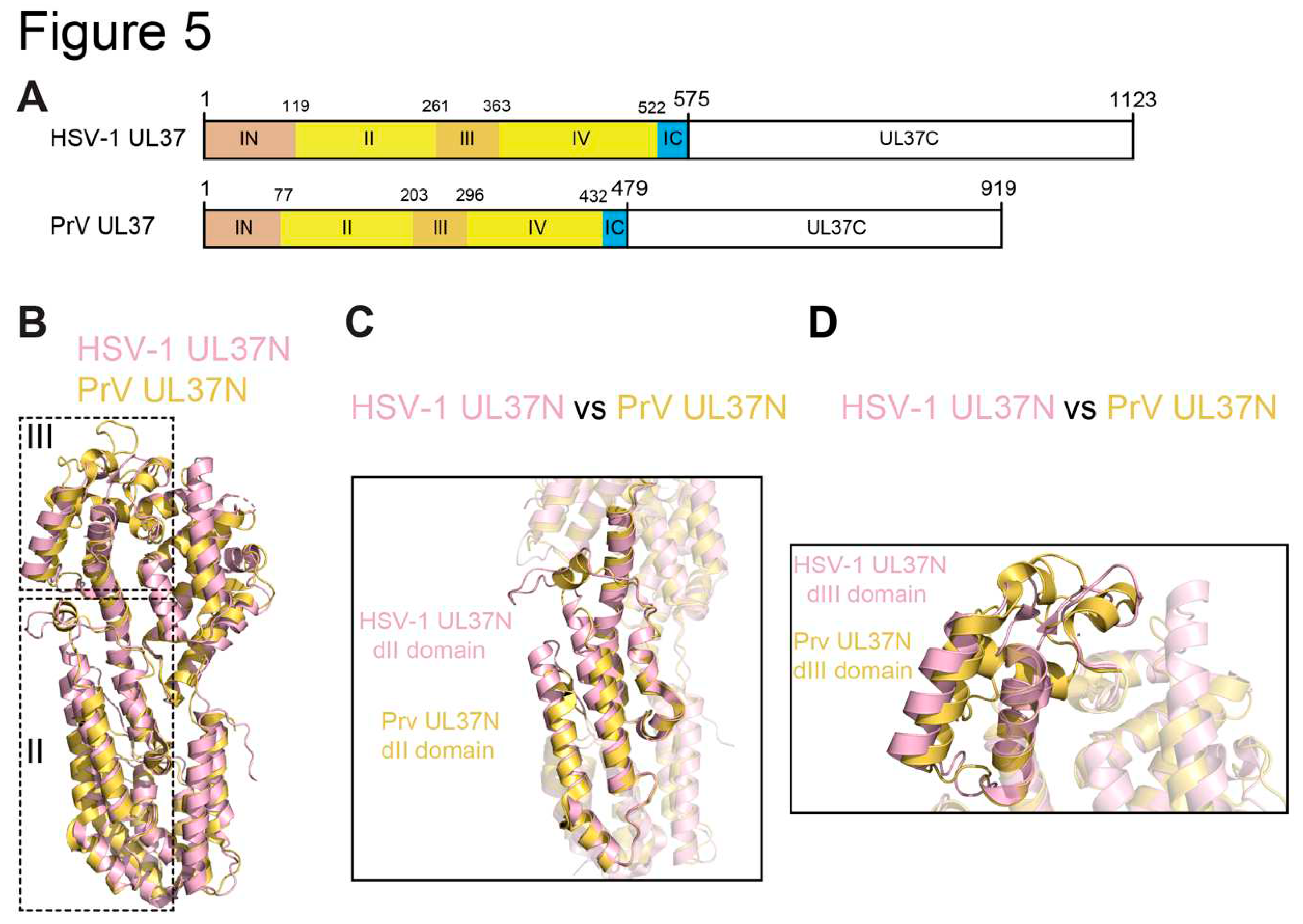
Figure 6.
Structures of UL53 and UL50. A, Schematic representation showing the domain of full-length HCMV UL53 and UL50. B, The structure of HCMV UL53 (residues 72-292,5DOE), Helices, β-strands and Zinc atom are specified, and coloured pink, yellow and wheat, respectively. C, The homodimer structure of HCMV UL53 (residues 84-292,5DOC). D. Complex structure of HCMV UL53-UL50 (5DOB). UL53 is shown in light blue and UL50 in wheat.
Figure 6.
Structures of UL53 and UL50. A, Schematic representation showing the domain of full-length HCMV UL53 and UL50. B, The structure of HCMV UL53 (residues 72-292,5DOE), Helices, β-strands and Zinc atom are specified, and coloured pink, yellow and wheat, respectively. C, The homodimer structure of HCMV UL53 (residues 84-292,5DOC). D. Complex structure of HCMV UL53-UL50 (5DOB). UL53 is shown in light blue and UL50 in wheat.
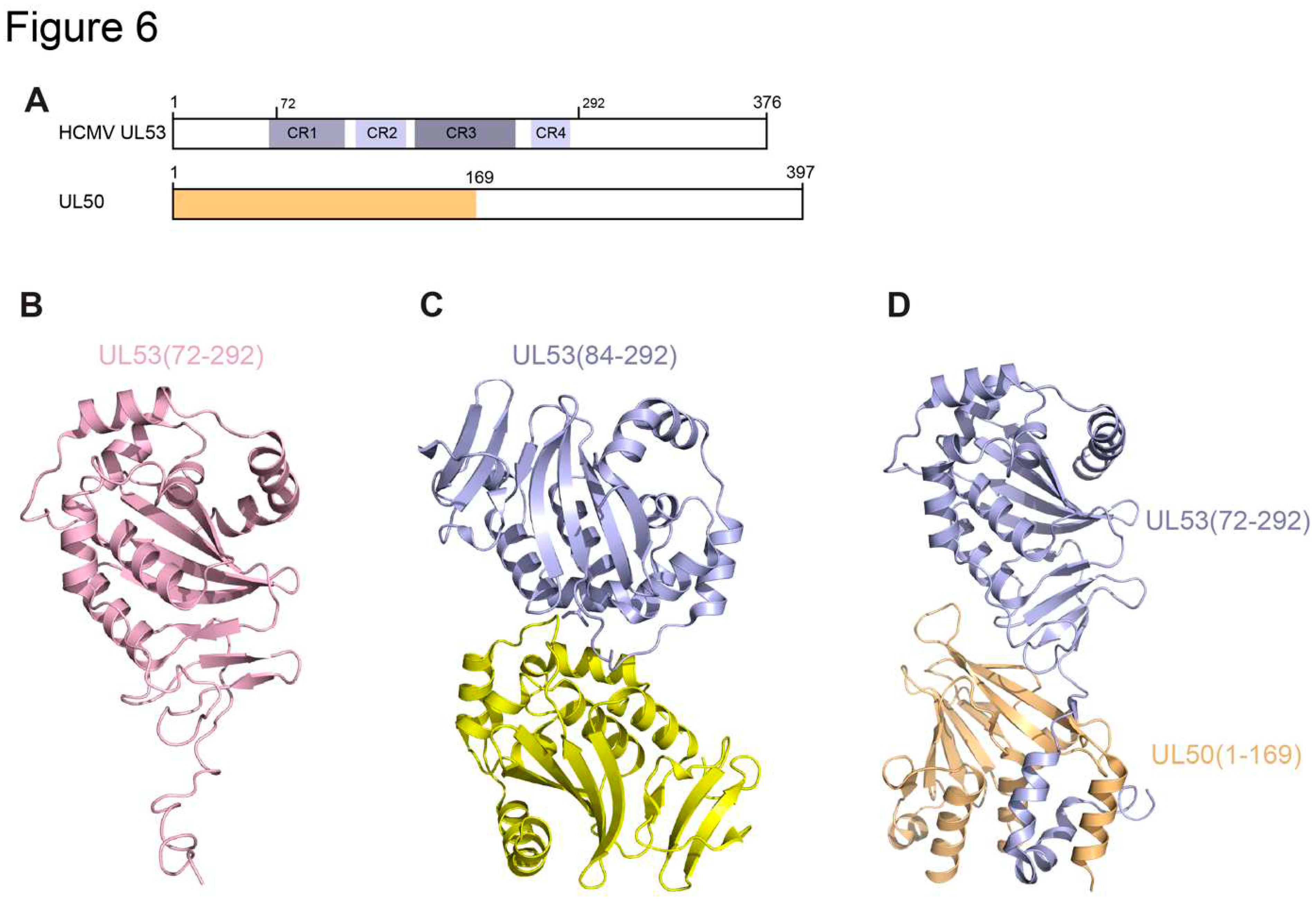
Figure 7.
Structures of BNRF1-DAXX-H3.3-H4 complex. A, Schematic diagram of full-length BNRF1 and homologous hFGART. Each domain are assigned with divergent colors. And BNRF1 DID domain is shown in slate as in the complex. B. Complex structure of EBV BNRF1(DID domain)- DAXX HBD-H3.3-H4 (5KDM), and coloured as in Figure 7B.
Figure 7.
Structures of BNRF1-DAXX-H3.3-H4 complex. A, Schematic diagram of full-length BNRF1 and homologous hFGART. Each domain are assigned with divergent colors. And BNRF1 DID domain is shown in slate as in the complex. B. Complex structure of EBV BNRF1(DID domain)- DAXX HBD-H3.3-H4 (5KDM), and coloured as in Figure 7B.
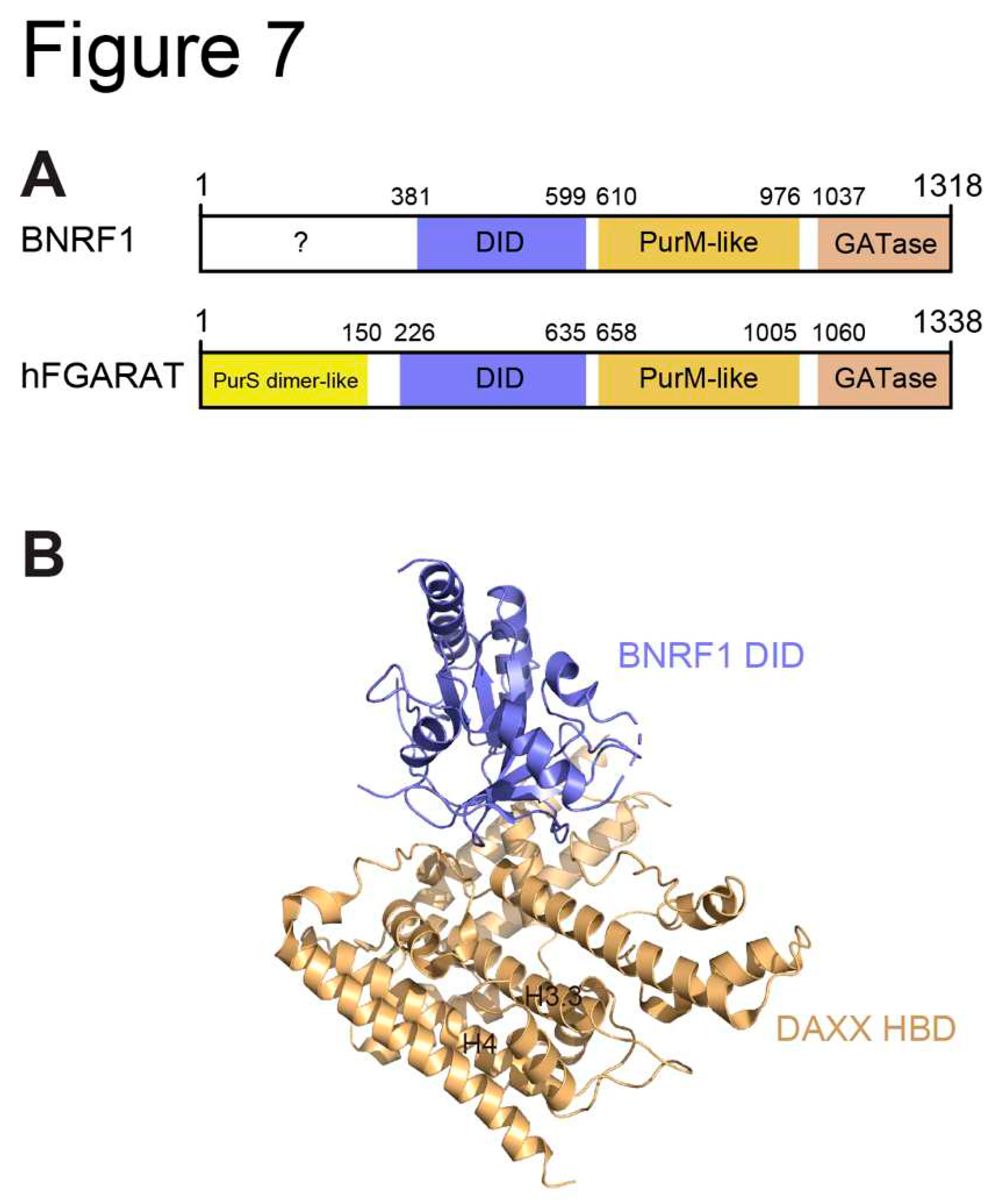
Figure 8.
Structures of BORF2. A, Schematic of BORF2 showing the A3B binding regions (yellow) and the unique SHI and LLI domain (hotpink). B, Structure of BORF2-A3Bctd complex (7RW6) in dimer, BORF2 colored slate, A3Bctd colored wheat and RNR core colored yellow. C, RNR subunit structural conservation in BORF2 (lightblue), E. coli RNR subunit (6W4X, cyan) and human RNR subunit (6AUI, pink) share a conserved RNR core and overall fold.
Figure 8.
Structures of BORF2. A, Schematic of BORF2 showing the A3B binding regions (yellow) and the unique SHI and LLI domain (hotpink). B, Structure of BORF2-A3Bctd complex (7RW6) in dimer, BORF2 colored slate, A3Bctd colored wheat and RNR core colored yellow. C, RNR subunit structural conservation in BORF2 (lightblue), E. coli RNR subunit (6W4X, cyan) and human RNR subunit (6AUI, pink) share a conserved RNR core and overall fold.
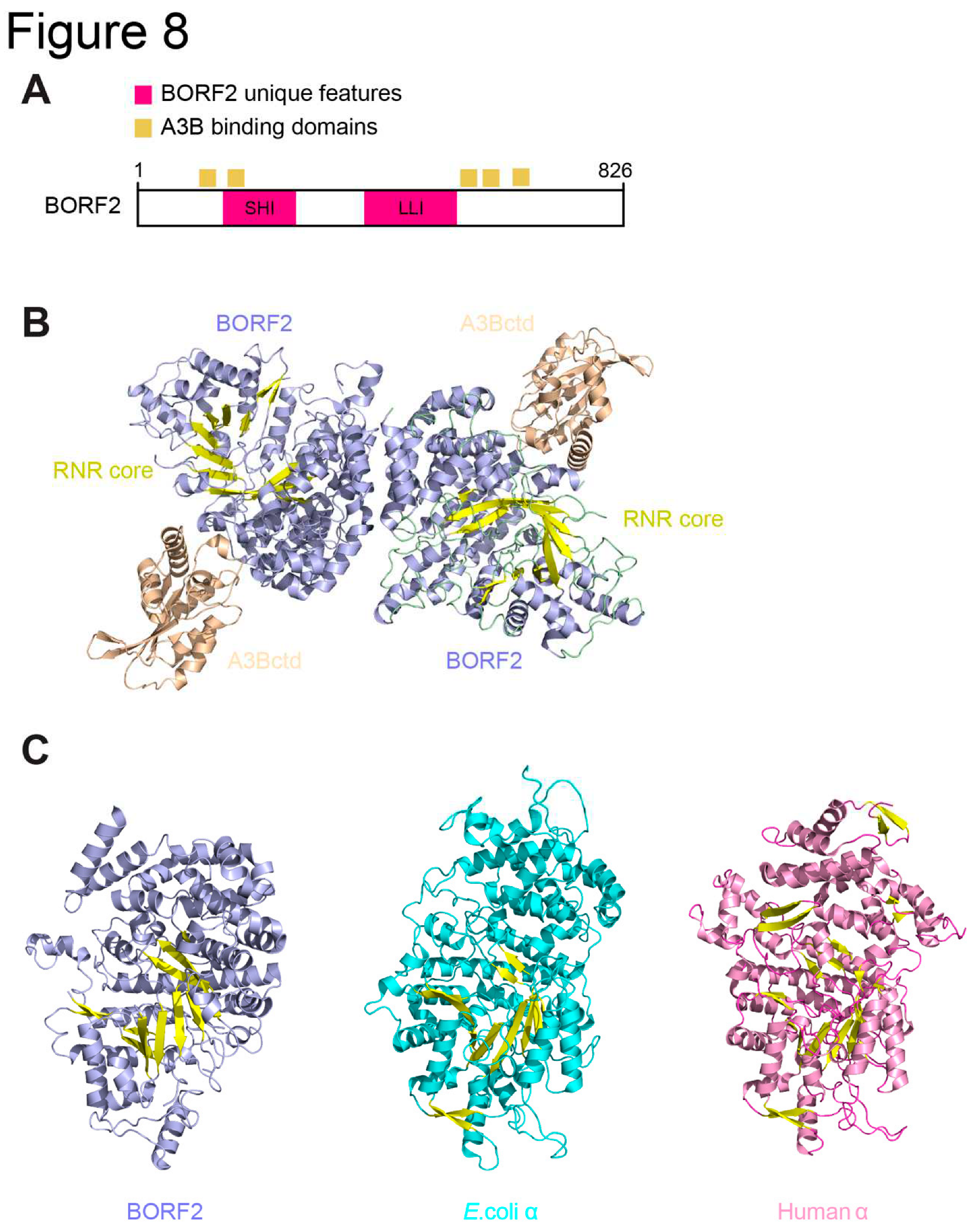
Figure 9.
Structures of BKRF4. A, The structures of BKRF4-N in a co-chaperone complex with the histone chaperone ASF1b and H3.3-H4 (7VCQ). B, Structure of BKRF4-N and H2A-H2B dimer (7VCL). BKRF4-N colored cyan, ASFIb colored gray, H3.3 in pink, H2A in purple, H4 and H2B in wheat.
Figure 9.
Structures of BKRF4. A, The structures of BKRF4-N in a co-chaperone complex with the histone chaperone ASF1b and H3.3-H4 (7VCQ). B, Structure of BKRF4-N and H2A-H2B dimer (7VCL). BKRF4-N colored cyan, ASFIb colored gray, H3.3 in pink, H2A in purple, H4 and H2B in wheat.
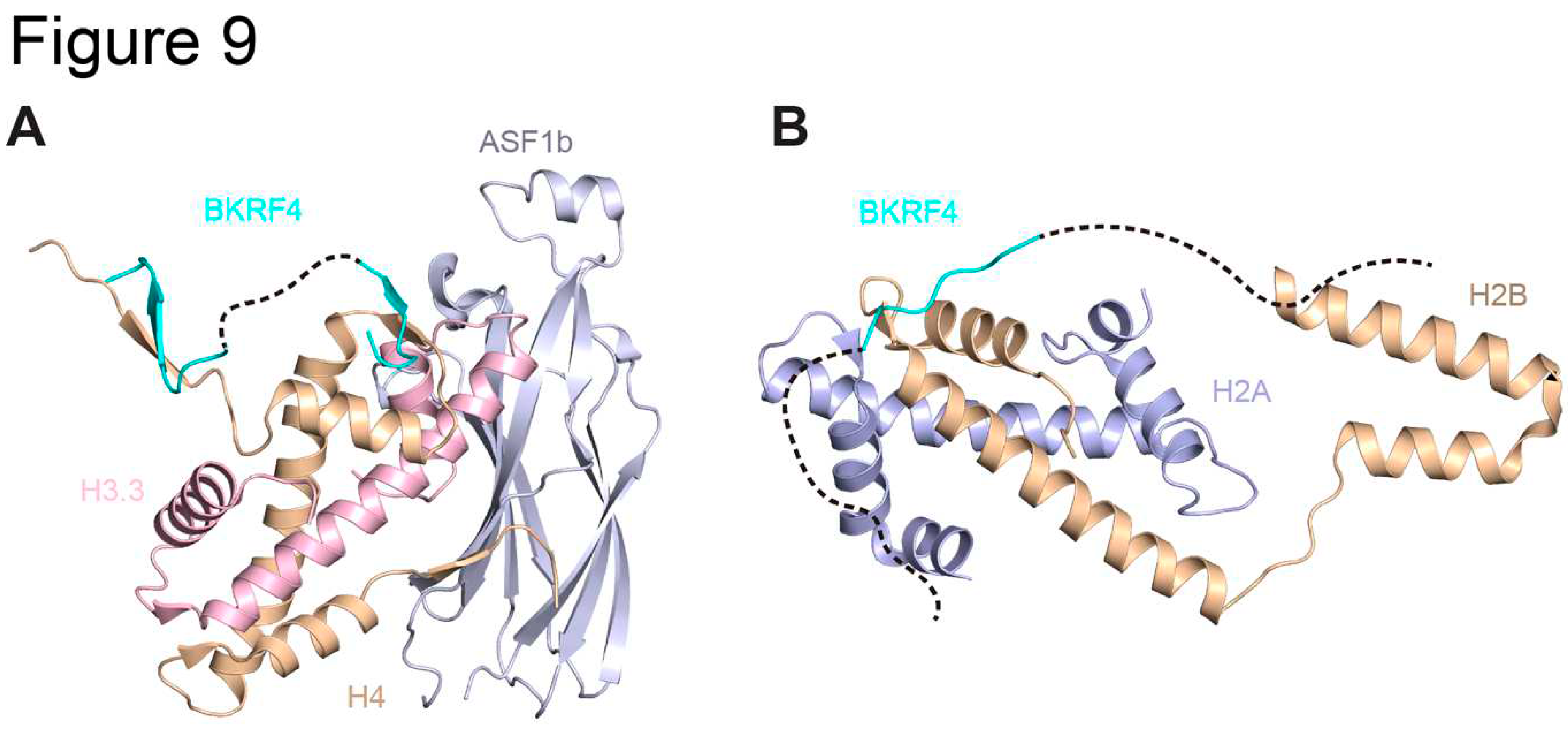

Table 1.
Conservation of Herpesvirus tegument proteins.
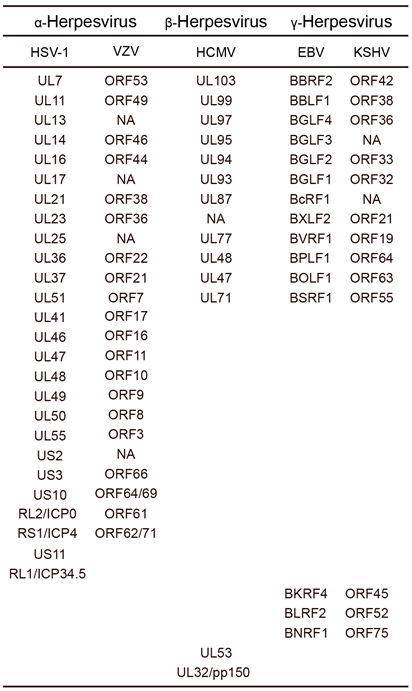
Table 2.
Structure of Herpesvirus tegument proteins.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



