
Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
Protein tyrosine phosphatases (PTPs) of the polymerase and histidinol phosphatases (PHP) superfamily with characteristic phosphatase activity dependent on divalent metal ions are found in many gram-positive bacteria. Although members of this family are co-purified with metal ions, they still require the exogenous supply of metal ions for full activation. However, the specific roles these metal ions play during catalysis are yet to be well understood. Here, we report the metal ion requirement for phosphatase activities of S. aureus Cap8C and L. rhamnosus Wzb. AlphaFold-predicted structures of the two PTPs suggest that they are members of the PHP family. Like other PHP phosphatases, the two enzymes have catalytic preference for Mn2+, Co2+, and Ni2+ ions. Cap8C and Wzb show an unusual thermophilic property with optimum activities over 75oC. Pre-incubation experiments show that the two enzymes are not inherently thermostable suggesting that certain catalytic step(s) in the reaction mechanism has a high activation energy. Consistent with this model, the activity-temperature profiles of the two enzymes are dependent on the divalent metal ion activating the enzyme.
Keywords:
protein tyrosine phosphatases
; polymerase and histidinol phosphatases
; Cap8C
; Wzb
; metal ion activation
; thermophilic enzymes
1. Introduction
Protein tyrosine phosphorylation and dephosphorylation exist in both prokaryotes and eukaryotes playing critical roles in pigment production, biofilm formation, biosynthesis of capsular/extracellular polysaccharides, control of cell growth, cell differentiation, along with other biological functions [1,2]. In these processes, protein tyrosine phosphorylation and dephosphorylation are characterized by the opposing actions of protein tyrosine kinases and protein tyrosine phosphatases, respectively [2].
Protein tyrosine phosphatases (PTPs EC 3.1.3.48) catalyse the removal of phosphate groups from phosphorylated tyrosine residues on proteins [3]. They are classified as classical PTPs, dual specificity PTPs (DSPs), low molecular weight phosphatase (LMPTP), and pTyr-specific phosphatases [4]. Recently, numerous manganese dependent prokaryotic O-phosphatases have also been characterized and classified based on amino acid sequence into phosphoprotein phosphatases (PPP), Mg2+/Mn2+-dependent protein phosphatases (PPM) and polymerase and histidinol phosphatases (PHP) superfamilies [5]. Protein tyrosine phosphatases of the PHP superfamily are commonly found in gram-positive bacteria with catalytic mechanism dependent on metals [6]. Except for PHP family members, protein tyrosine phosphatases generally share a signature catalytic motif C–X5–R–(S/T) [6]. Sequence alignment showed that C–X5–R–(S/T) motif is absent in PTPs such as Staphylococcus aureus Cap8C and Lactobacillus rhamnosus Wzb of the PHP superfamily [6,7].
Although S. aureus Cap8C is yet to be purified and characterised, comparative sequence analyses show that the phosphatase contained similar highly conserved motifs of the PHP domain as found in Streptococcus pneumoniae CpsB (the first characterised member of the PHP family) and L. rhamnosus Wzb [7]. Phosphatases of the PHP superfamily are metalloenzymes which are usually co-purified with three metal ions bound to the active site but still require exogenous supply of metal ions (particularly divalent metal ions) for full activation [8]. Recently, Kim et al. modeled the active site bound metal ions of YwqE from Bacillus subtilis and CpsB from S. pneumoniae as iron (Fe), iron (Fe), and magnesium (Mg) ions [9]. This contradicts Hagelueken et al. model for S. pneumoniae CpsB which proposed that the metals at the active site are all manganese ions [8]. Although the catalytic mechanism of PHP family members is not fully characterized, it appears to rely chiefly on signature sequence motif 4 which consists of five histidine residues and a single aspartic acid residue [10]. Mutations of conserved histidine and aspartic acid residues in the signature motif 4 greatly impair phosphatase activity and formation of cell capsule as observed for S. pneumonia CpsB [10].
Histidine residues are proposed to be involved in divalent metal ion binding at the catalytic site, but very few site-directed mutagenesis studies support this claim [7]. Divalent metal ions such as manganese, cobalt and copper have been widely reported to activate members of the PHP superfamily while others like magnesium, calcium and nickel are reported to have little impact on phosphatase activity [6,7]. While the histidine residues of the four conserved motifs of PHP family are proposed to be responsible for divalent metal ion binding, the aspartate residue coordinates the catalytic site [7]. Site directed mutagenesis studies have revealed that the inactivation of the aspartate or histidine residues of motif IV resulted in the loss of activity of S. pneumoniae Cps2B [11]. The finding implied that metal ion binding was necessary for PHP phosphoesterase activity and histidine residues were crucial for the binding. Like typical PHP family member, L. rhamnosus Wzb have their full activation dependent on metal ions [7]. The role these metal ions play during catalysis are yet to be well understood, some may be structural, for substrate binding, stabilization of enzyme-substrate complex or may be involved in different aspects of catalysis.
Even though L. rhamnosus Wzb originates from a mesophilic organism, the enzyme possesses potential thermophilic properties in the presence of metal ions and maybe a potential thermozyme with industrial benefits [7]. The presence of metal ions have been demonstrated to assist some enzymes maintain stability at temperatures that are otherwise lethal in the absence of such metal ions [12]. In view of this, it is important to study how different metal ions modify the phosphatase activities and thermophilic potentials of both S. aureus Cap8C and L. rhamnosus Wzb to be able to work out the possible roles of these metal ions.
The unusual thermophilic property of L. rhamnosus Wzb had earlier been suggested by Lapointe et al. [7], where the phosphatase activity of the enzyme was observed to peak at 75°C following a 30-minute pre-incubation at the same temperature. Interestingly, the residual activity of Wzb at 50°C was reported to have declined to 10% after 30 minutes pre-incubation [7]. These properties revealed that Wzb from a mesophilic source might have interesting thermophilic potentials that can be further investigated.
In this study, we investigated the metal ion requirements of L. rhamnosus Wzb and S. aureus Cap8C to gain insight into the possible roles of these metal ions. Interestingly, the unusual high temperature dependence of some members of the PHP superfamily in the presence of metal ions has also been suggested [7]. Hence, the temperature dependence of Cap8C and Wzb and how they are affected by metal ion were also investigated.
Some structural information of the active site of PTPs of the PHP family reveals that they are co-purified with metal ions bound to catalytic site but still require unusual complex mixture of metal ions for their full activation [7,8,11]. Lapointe et al. reported that several divalent metal ions activated L. rhamnosus Wzb [7]. Even though these metal ions seem important for enzyme activation, the specific roles they play in catalysis are yet to be fully understood although some structural roles have been suggested [8,9].
Members of the polymerase and histidinol phosphatase family of proteins have active site characterized by a trinuclear metal center and distorted Tim barrel protein fold [13]. The active site architecture of some members of the PHP family such as CpsB from S. pneumonia, Bacillus subtilis YwqE and Lactococcus lactis HPP have been determined [8,9,13]. The catalytic structure of PTP of the PHP family would be described with reference to Cps4B from S. pneumonia. The structure consists of two three-stranded parallel β-sheets with a ring of α-helices on the outside of the structure [8]. Three metal ions are found bounded adjacent to one another between the β-sheets with three water molecules serving as ligands, the fold can be described as a distorted TIM barrel (Figure 6) [8,14]. Enzymes in the PHP family are co-purified with metal ions bound to the active site; Cps4B is co-purified with three metal ions bound to the active site (Figure 6a) [8]. Kim et al. reported that metal analyses of recombinant YwqE and CpsB by inductively coupled plasma atomic emission indicated that they contain mostly iron (Fe) and magnesium (Mg), but not other metal ions such as nickel (Ni2+), copper (Cu2+), zinc (Zn2+), manganese (Mn2+), and cobalt (Co2+) [9]. Kim et al. also modeled M1, M2 and M3 of YwqE and CpsB as iron (Fe), and magnesium (Mg) ions [9], which is different from what was reported by Hagelueken et al. for CpsB were M1, M2 and M3 were modeled as manganese (Mn2+) ion [8].
2. Results
2.1. AlphaFold2-Predicted Structures of Cap8C and Wzb show the Polymerase and Histidinol Phosphatase (PHP) Domain
The structures of Cap8C and Wzb as predicted by AlphaFold2 [15,16] was downloaded from the AlphaFold Protein Structure Database (https://alphafold.ebi.ac.uk/) and) and visualized using PyMOL ((https://pymol.org/2/). Both structures reflect the typical Polymerase and Histidinol Phosphatase (PHP) domain. The PHP domain is characterized by four conserved sequence motifs that contain invariant histidine and aspartate residues that function in coordinating metal ions [17]. The motif has a signature distorted (α/β)7 barrel fold (Figure 1). This unique fold is similar to the TIM-barrel fold found in metallohydrolases. The PHP-like distorted (α/β)7 barrel is also found in the protein YcdX from E. coli which has been shown to display phosphatase activity to pNPP [18]. Interestingly, the (α/β)7 barrel has a trinuclear Zn2+ metal-binding site on the C-terminal side of the barrel.
2.2. Metal ion Activation of Cap8C and Wzb Activities
Analysis of AlphaFold-predicted structures of Cap8C and Wzb along with evidence from biochemical characterization of related PTPs [7] suggest that the two proteins belong to the PHP phosphatase family. The PHP PTPs typically prefer Mn2+ as cofactor and studies of Wzb showed that it is activated by a wide range of divalent metal ions [7]. To confirm which metal ions activated Cap8C, we tested the effect of selected divalent metal cofactors (Co2+, Ni2+, Mn2+, Mg2+ and Ca2+) on the phosphatase activities of Cap8C and Wzb. Staphylococcus aureus Cap8C and Lactobacillus rhamnosus Wzb proteins were established to have phosphatase activity using pNPP as substrate. The protein content of samples of the purified enzyme solutions of both Cap8C and Wzb were determined as 0.106 mg/ml (3.61 µmol/ml) and 0.099 mg/ml (3.43 µmol/ml), respectively. From these solutions, 2.5 µM solutions were prepared which were then used for subsequent experiments.
As shown in Figure 2, Cap8C and Wzb showed very low activities (barely detectable) when the reaction buffer is not supplemented with any metal ions. Three of the divalent cations studied (Co2+, Ni2+, Mn2+) showed significant activation of the activities of both enzymes, while Mg2+ and Ca2+ did not have any noticeable effects on the activities of Cap8C and Wzb. This confirms that the two PTPs behaved like the PHP group of protein tyrosine phosphatases. Co2+ (1.0 mM) had a significant activating effect on Cap8C and Wzb activities, yielding 17- and 29-fold increase in activity, respectively. In the presence of Ni2+ (1.0 mM), the activity of Cap8C increased 12-fold, while Wzb had a 4-fold increase in activity. Wzb showed a 24-fold increase in activity in the presence of 1.0 mM Mn2+, while Cap8C showed a 7-fold increase in activity.
In summary, among the divalent cations that we studied, Co2+, Ni2+, and Mn2+ had significant activating effect on Cap8C and Wzb in the order Co2⁺(17-fold) > Ni2⁺(12-fold) > Mn2⁺(7-fold) and Co2⁺(29-fold) > Mn2⁺(24-fold) > Ni2⁺(4-fold), respectively.
2.3. Concentration-Dependent Activation of Cap8c and Wzb by Co2+, Ni2+ and Mn2+ ions
Having established that Cap8C and Wzb are activated to different extent by Co2+, Ni2+ and Mn2+ ions, we determined the effect of the concentration of each metal ion on the activities of the two enzymes. We added increasing concentrations (0.25, 0.5, 1.0 and 2.0 mM) of the three divalent cofactors to the reaction buffer and measured the activity of the two enzymes.
Cap8C: As shown in Figure 3, all three metal ions caused a concentration-dependent and significant (p < 0.05) increase in phosphatase activity of Cap8C. All the four concentrations (0.25, 0.5, 1.0 and 2.0 mM) of Co2⁺ studied showed increased and sustained activating effect on Cap8C catalyzed hydrolysis of pNPP. The activating effect peaked in the presence of 1.0 mM Co2⁺. Ni2⁺ had a progressive activating effect on Cap8C with a steady increase in activity as the metal ion concentration was increased from 0.25 mM to 2.0 mM. At all the concentrations of Mn2⁺ studied, high activating effects on Cap8C catalyzed hydrolysis of pNPP were observed. This activating effect peaked at 1.0 mM (9-fold increase), and there was a slight decrease in activity as the metal ion concentration was increased from 1.0 mM to 2.0 mM. Overall, 1.0 mM Co2⁺ had the highest activating effect on Cap8C activity with approximately 17-fold increase (Figure 3).
Wzb: A similar pattern of concentration-based activation of Cap8C was observed for Wzb (Figure 3). Co2⁺ had highest activating effect on Wzb catalyzed hydrolysis of pNPP. This activating effect started steadily as concentration increased from 0.25–0.5 mM followed by a rapid increase as Co2⁺ concentration approached 1.0 mM where the activity peaked, yielding a 27-fold increase in activity. There was no significant change in activity as Co2+ concentration was increased from 1.0 mM to 2.0 mM.
Ni2+ showed limited activating effect on Wzb (Figure 3) with no effect seen in the presence of 0.25 and 0.5 mM of the divalent cation. Increase in Ni2+ concentration to 1.0 mM and 2.0 mM led to corresponding increase in its activating effect of Wzb with a 7-fold increase.
Mn2⁺ showed a concentration-dependent steady activation of Wzb from 0.25 mM to 1.0 mM, where it peaked with a 26-fold increase in activity. However, increasing the concentration of Mn2+ to 2.0 mM led to a sharp drop-in activity, giving a 10-fold enhancement over the control compared to 26-fold seen with 1.0 mM concentration (Figure 3).
2.4. Temperature Dependence of Cap8C and Wzb Activities
LaPointe et al. showed that the optimum temperature of Wzb was 75 °C [7]. In addition, Phyre2 analysis of the structure of Cap8c showed some structural similarity to some thermophilic proteins. Hence, we determined the effect of temperature (30–90 °C) on the activities of Cap8C and Wzb.
Cap8C: Figure 4A shows that Cap8C exhibited high activity when activated by its cofactors at 50–90 °C. In the presence of 1.0 mM Co2+, the activity of Cap8C increased with temperature from 93.66 mmol/min at 30 °C to 2931.61 mmol/min at 90 °C. With Co2+ ion, the activity of Cap8C peaked at 90 °C with an activity of 2931.61 mmol/min which is 31-fold higher than that of 30 °C. Similarly, the activity of Cap8C increased with temperature in the presence of 1.0 mM Ni2+ from 31.93 mmol/min at 30 °C to 543.33 mmol/min at 90 °C (Figure 4). However, at 70 °C, this activity peaked (609.84 mmol/min) yielding a 19-fold increase in activity when compared with the activity observed at 30 °C. In the presence of 1.0 mM Mn2+, Cap8C activity increased with temperature from 49.31 mmol/min at 30 °C to 503.95 mmol/min at 90 °C. Cap8C activity peaked at 90 °C in the presence of Mn2+ with a 10-fold increase (Figure 4).
Wzb: The effect of temperature (30–90 °C) on Wzb catalyzed hydrolysis of pNPP in the presence of the divalent cations (Co2+, Ni2+ and Mn2+) is shown in Figure 4B. The result revealed that in the presence of 1.0 mM Co2+, the activity of Wzb increased rapidly with increase in temperature peaking at 90 °C. With Co2+, Wzb activity at 90 °C (4696.49 mmol/min) was 6-fold higher than that observed at 30 °C. When Ni2+ was used as the cofactor, the activity of Wzb also increased with temperature peaking at 90 °C (1132.95 mmol/min) representing a 4-fold increase. This increase in activity is significantly less than what was seen when Co2+ was used as the cofactor. In the presence of 1.0 mM Mn2+, Wzb activity rapidly increased with temperature and peaked at 70 °C with an activity of 3092.65 mmol/min (~ 3-fold increase). But as temperature approached 90 °C, this activity declined rapidly from 3092.65 to 904.67 mmol/min (~ 3-fold decrease).
2.5. Effect of Metal ion Cofactors on Thermostability of Cap8C and Wzb
2.5.1. Effect of Mn2+ on Temperature-Dependent Activation Thermostability of Cap8C and Wzb
The effect of Mn2+ on the stability profile of Cap8C (Figure 5A) and Wzb (Figure 5B) at high temperatures (50–90 °C) were evaluated and compared at time 0–30 minutes. Pre-incubation of Cap8C at 50 °C in the presence of Mn2+ resulted in a steady increase in activity as pre-incubation time increased. Activity peaked at 50 °C (58% increase over the starting time) and was reduced to 54% and 27%, at 20 and 30 minutes, respectively. In contrast, recovered activities following pre-incubation at 60, 70, 80 and 90 °C were characterized by initial increase after 2 minutes followed by sharp decline from 5 to 30 minutes. The severity of the loss of activity correlates directly with the pre-incubation temperature.
Pre-incubation of Wzb at different temperatures in the presence of Mn2+ showed a trend (Figure 5B) that is nearly identical to the Cap8C pattern shown in Figure 5A. At 50 °C, Wzb’s activity was at initial pre-incubation time (0–5 minutes) observed to increase slightly (2651.171 to 2721.42 mmol/min), but as pre-incubation time increased to 30 minutes activity decreased steadily leading to 12% loss of enzyme stability. A similar pattern of increased Wzb activity at initial pre-incubation time 2 and 5 minutes and decrease in activity was observed at 60 and 70 °C as pre-incubation time increased to 30 minutes. The activity of Wzb at 60 and 70 °C peaked at 5 minutes pre-incubation, followed by steady loss of activities over the remaining time points assayed. Similarly, recovered activities following pre-incubation at 80 and 90 °C were characterized by initial increase after 2 minutes followed by sharp decline from 5 to 30 minutes. In all cases, the extent of the loss of activity correlated directly with the pre-incubation temperature.
2.5.2. Effect of Co2+ on Temperature-Dependent Activation Thermostability of Cap8C and Wzb
The effect of Co2+ on the stability profile of Cap8C and Wzb at high temperatures (50–90 °C) were evaluated and compared at time 0–30 minutes (Figure 6). Pre-incubating Cap8C with 1.0 mM Co2+ at 50 and 60 °C for 0–30 minutes had very limited effect on the recovered activity of the enzyme producing a slight increase in activity from 1125.49 mmol/min (0 minutes) to 1337.29 mmol/min (30 minutes) and from 1123.90 mmol/min (0 minutes) to 1339.95 mmol/min (30 minutes), respectively (Figure 6). Incubating the enzyme at 70 and 80 C had a similar effect of increasing the activity with a more linear trend over the 30 minutes studied. The recovered activities were higher at all time points were higher for the 80 °C incubation than the 70 °C. At 70 °C, there was a steady increase in Cap8C activity from pre-incubation time 0 (1118.05 mmol/min) to 30 minutes (1344.74 mmol/min) in the presence of 1.0 mM Co2+. The activity of Cap8C peaked at 70 °C after pre-incubating for 20 minutes (1803.46 mmol/min) when compared with activity at the start of pre-incubation (0 minute). This was followed by a rapid decline as pre-incubation time approached 30 minutes (1344.74 mmol/min with 120% thermostability) when compared with activity at 20 minutes pre-incubation at 70 °C. Interestingly, at 80°C and 90 °C, in the presence of 1.0 mM Co2+, Cap8C activity increased progressively with increase in pre-incubation time, but a slight decline was observed at 90 °C after pre-incubation for 20 and 30 minutes. At 80 °C, Cap8C activity peaked after 30 minutes pre-incubation yielding 2120.618 mmol/min when compared with activity at 0 minute pre-incubation. Pre-incubation at 90 °C leads to a sharp and gradual increase in activity over the first 15 minutes. At 90 °C, maximal Cap8C activity was observed at 15 minutes (3321.68 mmol/min with 300% thermostability) compared with activity at 0 minute pre-incubation in the presence of 1.0 mM Co2+. Further incubation did not lead to increase in activity. This trend suggests that Co2+ activation of the enzyme is temperature dependent and may indicate a structural transition mechanism that switches on the activity of the enzyme.
Pre-incubation of Wzb at different temperatures in the presence of Co2+ showed a trend different from what was observed with Cap8C (Figure 6B). Incubation at 50 °C resulted in increase in recovered activity peaking at 15 minutes (5467.84 mmol/min) after which there was a gradual decrease in activity to 4482.29 mmol/min. Similar effect was seen when the enzyme was incubated at 60 °C. Although a higher increase in residual activity was observed with a peak at 5 minutes followed by a sharp linear decline resulting in about the same activity seen for the 50 C incubation for 30 minutes. The pattern seen for pre-incubation at 70 °C and 80 °C are similar: an initial increase in activity at 2 minutes followed by a linear but moderate drop-in activity. In contrast, pre-incubation at 90 °C caused a sharp and sustained decline in activity after 2 minutes resulting in loss of over 90% activity after 10 minutes.
2.5.3. Effect of Ni2+ on Temperature-Dependent Activation Thermostability of Cap8C and Wzb:
The effect of Ni2+ on the stability profile of Cap8C (Figure 7A) and Wzb (Figure 7B) at high temperatures (50–90 °C) were evaluated and compared at time 0–30 minutes. Following pre-incubation at 50 and 60 °C, in the presence of 1.0 mM Ni2+ and for 30 minutes, the activity of Cap8C increased from 1098.89 mmol/min to 2669.26 mmol/min and 1097.83 mmol/min to 3293.48 mmol/min, respectively. In contrast to what was observed at 50 and 60 °C, pre-incubation at 70 °C and 80 °C led to an increase in activity after 2 minutes but followed by a steady decrease in activity up to the 30 minutes time point that we investigated. A similar pattern was seen with the 90 °C pre-incubation with the difference been that the loss of activity was seen after 2 minutes and continue steadily leaving about 4% activity after 30 minutes. This trend suggests a temperature-dependent activation of the enzyme up to a point after which the protein is denatured.
Remarkably, the effect of Ni2+ on temperature-dependent activation and thermostability seen with Wzb is generally similar to the pattern seen with Cap8C (Figure 7B). In the presence of Ni2+ at 50 °C, Wzb activity increased with increase in pre-incubation time from 858.36 mmol/min at the start, peaked at 5 minutes to 1062.44 mmol/min and decreased slowly to 908.38 mmol/min at 30 minute. In contrast, at 60 °C, Wzb activity increased initially after 2 minutes pre-incubation to 1195.48 mmol/min. This was followed by a rapid decline as pre-incubation time increased to 30 minutes (677.96 mmol/min). At 70 °C, a slight increase in Wzb activity (which was also the peak) was observed at 2 minutes pre-incubation. This was followed by a rapid decrease as pre-incubation time increased to 30 minutes. In a trend similar to what was seen with Cap8C (Figure 7A), at 80 and 90 °C, Wzb activity decreased rapidly as pre-incubation time was increased from 0 to 30 minutes, with 52 and 38% residual activities, respectively.
3. Discussion
3.1. Divalent Metal Ions as Cofactors of Cap8C and Wzb Phosphatase Activities
AlphaFold2-predicted structure of Cap8C and Wzb puts the two protein tyrosine phosphatases (PTPs) in the polymerase and histidinol phosphatase (PHP) family (Figure 1). PTPs are generally not considered to have catalytic activities that are dependent on metal ions except for those in the PHP superfamily [4,6]. However, some PTPs have been reported to have activities modulated by the presence of some metal ions [19,20,21]. Although phosphatases of the PHP superfamily are co-purified with metal ions attached to the active site, their activities in vitro in the absence of the exogenous supply of some metal ions remain barely detectable [8,9,13], indicating non-structural roles for the metal ions. Hence, one of the objectives of this work was to identify the divalent metal ions that serve as activating cofactors for Cap8C and Wzb enzymes. The predicted catalytic site of PHP which consists of four motifs with conserved histidine residues have been proposed to be involved in metal dependent catalysis of phosphoester bond hydrolysis [7,14], through metal ion binding and coordination for enzymatic activity (14). Also, the conserved aspartate residue of the fourth motif has been predicted to be important for catalysis by participating in electron transfer [11].
Previous reports have shown that Co2+, Ni2+, Mn2+and Mg2+ stimulate phosphatase activity to different extents, while Ca2+ has little or no activating effect on phosphatases [5]. The effect of these selected metal ions on the phosphatase activities of Cap8C and Wzb were investigated in this study, with the aim of establishing their dependence on these cations. Our results show that in the absence of Co2+, Ni2+, and Mn2+, the activities of Cap8C and Wzb on pNPP hydrolysis were barely detectable at pH7.5 (Figure 2). This shows both enzymes are metalloenzymes that require the presence of metal ions for full activation just as reported for members of the PHP superfamily that show low activities in the absence of metal ions [8,11]. Mijakovic et al. reported a metal dependent PTP of the PHP superfamily YwqE from B. subtilis, with barely noticeable activity in the absence of 1.0 mM Mn2+, 1.0 mM Cu2+ and 1.0 mM Zn2+ [22]. A baseline level of phosphatase activity was also observed for S. pneumoniae CpsB in the absence of 1.0 mM Co2+, 1.0 mM Mn2+ and 1.0 mM Mg2+ [11].
Our results show that Co2+ best activated both Cap8C and Wzb with 17-fold and 29-fold increase in activity, respectively. This study is the first to report Co2+ activation of Cap8C activity and shows that the PTP behaves in a similar pattern in the presence of Co2+ as other members of the PHP family. S. aureus Cap8C had been predicted to be a homologue of S. pneumoniae CpsB.
The PHP family members of PTPs have been generally described to have optimal activity at basic pH and are also sensitive to Mn2+ [9,23]. Streptococcus pneumoniae CpsB, the first PTP representative of the PHP superfamily to be described, has phosphatase activity dependent on Mn2+ (11). Like S. pneumoniae CpsB, B. subtilis YwqE (another member of the PHP superfamily) exhibited maximal phosphatase activity in the presence of Mn2+ (22). Similarly, Standish et al. reported that DNA polymerase PolC exonuclease activity was significantly higher in the presence of Mn2+ than Mg2+ [23]. In our study, both Cap8C and Wzb were significantly activated by Mn2+, 7-fold and 24-fold, respectively. The characteristic activation of Cap8C and Wzb by Mn2+ reveal a significant difference from what was described for S. pneumoniae CpsB (manganese-dependent PTP) [11]. This suggests that although Cap8C has been proposed to be a homologue of S. pneumoniae CpsB, its sensitivity to Mn2+ differs and its phosphatase activity does not seem to be dependent on the cation. The responsiveness of Cap8C to Mn2+ in addition to earlier claim of sequence homology with the novel member of PHP superfamily, S. pneumoniae CpsB, shows that Cap8C belongs to the PHP family.
The conserved histidine residues on the catalytic motif of phosphatases of the PHP superfamily have been suggested to be involved in metal binding, especially of Ni2+ [24,25]. Although studies are yet to confirm that Ni2+ is essential for activating phosphatase of PHP superfamily, it has however been shown to activate them to a lesser extent compared with the activating effects of Co2+, Cu2+ and Mn2+ [5,8]. In this study, we show that Ni2+ significantly increased the activities of Cap8C and Wzb (Figure 2). Ni2+ was observed to have a higher activating effect on Cap8C (12-fold) than on Wzb (4-fold). LaPointe showed that the absence of Ni2+ in reaction mixtures containing other cations resulted in a minimal reduction in the phosphatase activity of Wzb [7]. Also, the activity of S. pneumoniae CpsB, the pioneer phosphatase of the PHP superfamily in the presence of Ni2+ was observed to have increased by approximately 5-fold [8]. Shi et al. reported that the phosphatases from the phosphoprotein phosphatase (PPP) superfamily activities were significantly activated by Ni2+ and they include, S. typhimurium PrpA, S. typhimurium PrpB, B. subtilis PrpE and S. coelicolor SppA [26]. However, the specific role of Ni2+ in these systems are yet to be established.
Mg2+ and Ca2+ ions are generally not known to be essential for the activity of PTP members of the PHP superfamily but can stimulate to some extent the activity of some phosphatases [5]. On the basis of their ionization potential, Mg2+ and Ca2+ are not considered to possess strong Lewis acidity when compared to other divalent metal ions such as Co2+, Ni2+ and Mn2+. This might contribute to their poor activating effect during the catalysis of hydrolysis reactions [27]. Our results show that both Mg2+ and Ca2+ had no significant effect on the phosphatase activity of Cap8C and Wzb, as both enzymes only yielded baseline activities in the presence of the cation (Figure 2). These results agree with what has been reported for other PHP PTPs. Mijakovic et al. showed that Mg2+ and Ca2+ have no effect on the phosphatase activity of B. subtilis YwqE [22]. Morona et al. also reported a baseline level of phosphatase activity for S. pneumoniae CpsB in the presence of Ca2+ [11].
3.2. Activation of Cap8C and Wzb by Divalent Cations is Concentration-Dependent
With the activating effects of Co2+, Ni2+ and Mn2+ on Cap8C and Wzb already established in the previous section, the effect of varying concentrations of these ions on the phosphatase activity of Cap8C and Wzb were investigated. The findings generally showed that the metal ions had concentration dependent activating effect on the phosphatase activities of both enzymes (Figures 3).
Protein tyrosine phosphatases of the PHP superfamily have been reported to be sensitive to Mn2+ [9], and the findings from this study show that Cap8C and Wzb are no exception. The findings in this study reveal that all concentrations of (0.25, 0.50, 1.00 and 2.00 mM) Mn2+ studied had high and sustained activating effect on Cap8C. But even at its peak (1.0 mM), the activating effect of Mn2+ was observed to be lower than what was obtained in the presence of Co2+. This suggests that although Cap8C is sensitive to Mn2+, the enzyme’s activity was maximal in the presence of Co2+. On the other hand, for all the concentrations used in this study, Mn2+ had a progressive activating effect on Wzb that peaked at 1.0 mM (Figure 3). The findings in this study reveal that Cap8C and Wzb pose slightly different sensitivity to Mn2+, as the cation was observed to favour the phosphatase activity of Wzb more. This finding agrees with the reports by LaPointe et al., where 0.1 mM Mn2+ was reported to only increase the activity of L. rhamnosus Wzb by only 2-folds [7]. Hageleuken et al. reported that 0.5 mM Mn2+ resulted in a 13-folds increase in the phosphatase activity of S. pneumoniae Cps4B [8]; this is however higher than what was obtained in our study for both enzymes.
Our findings show that the activating effect of Co2+ on Cap8C and Wzb was optimal at 1.0 mM for both enzymes. This trend of activation for Cap8C and Wzb suggests that Co2+ might be performing the same role during catalysis by both enzymes as the patterns of activation were similar for both enzymes. Also, the role of Co2+ in the catalysis by both enzymes may not be structural, as many metal ions that have been implicated in structural functions during catalysis are reported to be deeply buried within the active site of enzymes [28]. The metal ions that carry out structural functions act by helping the enzyme adopt proper conformation which promotes interactions and consequently enhances enzymatic catalysis [28]. Crystal structures that have been determined for some members of the PHP family have not shown Co2+ to be part of the metal ion cluster at the active site of the purified enzymes [8,9]. Kim et al. modeled the metal ion cluster at the active site of YwqE from Bacillus subtilis and CpsB from S. pneumoniae as two iron (Fe) and magnesium (Mg) ions [8]. It follows that a structural role for Co2+ in Cap8C is a possibility that is worthy of further exploration.
Generally, Ni2+ has been reported to be a poor activator of phosphatases especially of the polymerase and histidinol phosphatase superfamily [5]. For Cap8C, unlike other phosphatase members of the PHP superfamily, the activating effect was observed to be high. On the other hand, Ni2+ had the least activating effect on Wzb as expected for PTP members of the PHP superfamily. Although the activating effect of Ni2+ was maximal at 2.0 mM, Co2+ was a better activator of Cap8C than Ni2+. The pattern of Co2+ and Ni2+ activation suggests that the cations might be influencing enzyme-substrate binding by facilitating the release of proton from bound water, yielding nucleophilic hydroxyl ion [8,28]. These cations may also boost Cap8C activity by interacting with negatively charged amino acid residues such as aspartic and glutamic acid at the active site, thereby stabilizing negative charges of trigonal-bipyramidal intermediate, thus translating into increased activity [8,9]. In agreement with our findings in this study, 0.5 mM Ni2+ was observed to increase the activity of wild-type Cps4B from S. pneumoniae by 5-fold [8]. LaPointe et al. reported that the absence of Ni2+ in a reaction mixture that contained other metal ions resulted in slight decrease in L. rhamnosus Wzb activity [7].
The summary of the investigation carried out on the effect of varying concentration of the metal ions (Co2+, Ni2+ and Mn2+) on the phosphatase activity of Cap8C and Wzb reveal that both enzymes had higher levels of dependence on Co2+ compared to the other two cations. These characteristics imply that though both Cap8C and Wzb belong to the PHP superfamily, they are significantly different from the phosphatase described for S. pneumoniae, CpsB.
3.3. Temperature Dependence of Cap8C and Wzb Activities
Rise in temperature is expected to exponentially increase the rate of enzyme catalysed reaction at least until a point is reached where observable decline in activity begins due to loss of the protein’s native structure [29]. For an enzyme catalysed reaction, increase in temperature below the optimal temperature usually results in corresponding increase in activity. This is partly because the inactivation effect of temperature is not pronounced at suboptimal temperatures [29]. However, at supraoptimal temperatures, activity decreases because of increased thermal-induced enzyme inactivation [29]. Thermophilic and hyperthermophilic enzymes are intrinsically active and stable at high temperatures and they offer major biotechnological advantages over mesophilic or psychrophilic enzymes. They are easily purified by heat treatment, are more resistant to chemical denaturation, have higher reaction rates, and they tolerate higher substrate concentrations [30,31].
The effect of temperature (30–90 °C) on the phosphatase activities of Cap8C and Wzb was investigated in the presence of Co2+, Ni2+ and Mn2+. This was done to understand the catalytic behaviour and stability of these enzymes in the presence of the divalent ions at various temperatures.
Results of this study showed that in the presence of Co2+, Ni2+ and Mn2+, Cap8C showed thermostable tendency as activities were observed to increase with temperature beyond what is observed for enzymes of mesophilic origin. In the presence of Co2+, Cap8C had optimal activity at 90 °C which is significantly higher than the optimal growth temperature (37 °C) of S. aureus. This indicates that in the presence of the cation, increase in Cap8C activity was maintained with increase in temperature. This suggests that as stabilizing bonds of Cap8C break and reform rapidly, Co2+ was able to promote and maintain conformational changes in the protein needed for catalysis to occur at a faster pace [32]. The results also showed that in the presence of Co2+, Cap8C was more resistant to heat–induced inactivation that might have taken place at high temperatures.
Although with lower levels of activities, in the presence of Ni2+ and Mn2+, Cap8C activity increased steadily with increase in temperature with optimal activity at 70 and 90 °C, respectively (Figure 4a), as against 37 °C optimal growth temperature of S. aureus. The result implies that the binding of Ni2+ and Mn2+ to Cap8C helped to some extent in stabilizing the conformational changes necessary for catalysis to take place at high temperature. In summary, our findings showed that in the presence of the metal ions, Cap8C displayed thermostable potentials in the order Co2+>Ni2+>Mn2+ and the thermostability is higher than what was reported for other PTPs from the same species [33]. Earlier, PTPa and PTPb from S. aureus were characterised and both enzymes had optimal activity at 40 °C [33]. A protein tyrosine phosphatase from mesophilic fungi, Metarhizium anisopliae, has been reported to show thermostable properties in the absence of metal ions, with optimum activity between 70 to 75 °C [34].
Like Cap8C, Wzb had thermostable properties in the presence of the metal ions, although to different extent as its activity remarkably increased with temperature. With Co2+, Wzb had progressive increase in activity with increase in temperature which peaked at 90 °C; this is significantly higher than the optimal growth temperature (6 to 41 °C) of L. rhamnosus. This implies that Co2+ was able to initiate and stabilize conformational changes of Wzb that was required for catalysis to happen faster. Also, in the presence of Co2+, more energy would be required to denature Wzb. In a like manner, Ni2+ might also have shielded Wzb from the deleterious effect of high temperature by ensuring that the continuous breaking and reforming of Wzb stabilizing bonds were not disrupted as temperature increased [32,35].
The findings that Cap8C and Wzb are PTPs found in mesophilic organisms with thermophilic properties in the presence of Co2+, Ni2+ and Mn2+ is not unprecedented. LaPointe et al. (7) reported the thermostable properties of L. rhamnosus Wzb in the presence of metal ions with activity peaking at 75 °C; however, the thermostability was studied in the presence of Cu2+, Co2+, Fe3+, Mn2+ and Mg2+ combination. John et al. have also characterised a thermostable protein tyrosine phosphatase from Trypanosoma evansi with optimal temperature at 70 °C [36]. This thermostable PTP as described for T. evansi was suggested to play important role in the adaptation of the organism to extreme temperatures in vivo. In addition to the claim that thermostable protein tyrosine phosphatase can be found in mesophiles is the purification and characterization of thermostable PTP from Metarhizium anisopliae which showed optimal activity at 70 °C [37]. Zhang and colleagues suggested that the high percentage of polar amino acid and proline content of Metarhizium anisopliae PTP might be responsible for its thermostability as these features play significant role in protein stability [37].
3.4. Effect of Metal Ion Cofactors on Thermostability of Cap8C and Wzb
The factors that confer thermostability on enzymes have been carefully studied and the general conclusion is that no single universal mechanism can promote stability at high temperatures [31]. However, some structural features have been proposed to contribute to enzyme thermostability. These features include more interactions (like hydrophobic, electrostatic, hydrogen bonds, disulfide bonds and metal binding) than those found in less stable enzymes. Others include superior conformational structure such as higher packing efficiency, more rigidity, reduced entropy of unfolding, conformational strain release and stability of α-helix [30,31].
Having established that Cap8C and Wzb showed thermostable properties in the presence of the cofactors, to further understand this, the stability profiles of these enzymes at different temperatures (50, 60, 70, 80 and 90 °C) were investigated at different pre-incubation times (0, 2, 5, 10, 15, 20 and 30 minutes) in the presence of metal ion cofactors. The objective of this investigation was to further probe into the predicted thermostable properties of both enzymes.
Since most technological processes require elevated bioprocessing temperatures, interest has been drawn into the development of “thermoactive and thermostable” enzymes that would achieve higher catalytic rates and provide significant processing advantages [30]. Thermophilic enzymes have been reported to be stable and perform optimally within the temperature range of 60 to 80 °C and typically do not perform well below 40 °C [30].
The thermophilic behaviour of Cap8C in the presence of Co2+ may imply that the cation acts to lock the active site in a stable conformation thereby ensuring increase in activity as temperature increases. This is an interesting finding that might require further investigations into the stability of Cap8C at high temperatures in the presence of the cation for longer duration to fully establish its thermostability. The stability of Cap8C at 90 °C in the presence of Co2+ as cofactor was observed to be higher than that of PTP from Thermus thermophiles, an extreme thermophile as reported [29]. Vieille et al. reported that the presence of Co2+ remarkably stabilized Bacillus licheniformis xylose isomerase, as both Ea of thermal denaturation and inactivation temperature were significantly higher than those of the apoenzyme [35].
The Wzb stability profile study done in the presence of Co2+ (Figure 6) revealed that Wzb was less stable in the presence of the cofactor when compared with Cap8C. Findings of this investigation indicated that pre-incubation of Wzb for 2–30 minutes at high temperature (90 °C) resulted in loss of activity which might be due to loss of the enzyme’s native conformation. It could be that lesser thermal energy would be required for Wzb to lose tightly bound Co2+, ultimately leading to unfolding thereby exposing the enzyme to chemical modifications (such as cysteine oxidation, deamidation and peptide bond hydrolysis) that eventually result in irreversible inactivation [30,32].
The study of the stability profile of Cap8C in the presence of Ni2+ revealed that the enzyme was stable within the temperature range of 50 to 70 °C (Figure 7). Loss of Wzb activity was visible at 80 and 90 °C with increased pre-incubation time. Findings gathered from this study revealed that at 50, 60 and 70 °C, Ni2+ was able to maintain a shielding effect on Cap8C against the deleterious effect of the high temperatures likely due to ensuring that the active site conformation for activity was maintained.
Like Cap8C, Wzb was observed to be less stable at high temperatures in the presence of Ni2+ with increase in pre-incubation time (Figure 7). The poor stabilizing effect of Ni2+ on Wzb as observed in this section was expected, as it had already been established that the cofactor is a poor activator of the enzyme. One possible explanation is that the cation was unable to lock the active site of Wzb in a stable conformation for a long period of time as temperature increased [32].
Protein tyrosine phosphatases of the PHP superfamily have been reported to be significantly activated by Mn2+, with some members such as Streptococcus pneumoniae CpsB having activity dependent on the cofactor [11]. This study has shown that Mn2+ significantly activated Cap8C but to a lesser extent when compared with other metal ions like Co2+ and Ni2+. Based on the results from this investigation, it was observed that Cap8C was less stable at high temperatures in the presence of Mn2+ (Figure 5). Cap8C was observed in this study to be most stable at 50 °C. Similar pattern of initial activation followed by rapid loss of Cap8C activity was recorded at 60, 70, 80 and 90 °C as pre-incubation time approached 30 minutes. The initial increase in Cap8C activity that was observed at 60, 70, 80 and 90 °C supports earlier findings that Mn2+ activates the enzyme at high temperatures. However, after prolonged pre-incubation, Mn2+ was unable to maintain the protein structure at high temperatures, thereby leading to the disruption of the stabilizing interactions and in turn loss of Cap8C activity.
The differences in the stabilization efficacy of Co2+, Ni2+ and Mn2+ observed might be due to their ability to adopt different geometries in the same site in the absence of substrate. In comparison with Co2+ and Ni2+, Mn2+ had lesser affinity for Cap8C as increasing pre-incubation time at 60 to 90 °C might lead to steady dissociation of the enzyme-metal complex. This steady dissociation of the holoenzyme would result in the corresponding increase in exposure of Cap8C to the inactivating effects of high temperatures. Mn2+ was reported to significantly shield and stabilize Bacillus licheniformis xylose isomerase (BLXI) at high temperature slightly more than Co2+ [35]. Mutations of metal binding sites on xylose isomerase (XI) have been reported to result in destabilization of the enzyme [35].
Figures 6 shows that Co2+ best activated Wzb, while Mn2+ was only slightly less activating. Results from the investigation of the effect of Mn2+ on the stability profile of Wzb showed that the cofactor was less efficient than Co2+ and Ni2+ in stabilizing Wzb at high temperatures (Figure 6 and Figure 7). In the presence of Mn2+, Wzb was less stable as increasing temperature and pre-incubation time had corresponding inactivating effect on Wzb. Initial pre-incubation of Wzb at 50, 60, 70 and 80 °C led to the activation of the enzyme. These were immediately followed by decline in activity. The initial increase in Wzb activity showed that for short time exposure, Mn2+ was able to effect activation and prevent denaturation at these temperatures. At 90 °C, Wzb was observed to be completely inactivated as activity after 30 minutes pre-incubation was barely detectable (Figure 5). This suggests that increasing pre-incubation time at high temperatures may lead to quick dissociation of the Wzb- Mn2+ complex, resulting in the release of the bound cofactor, exposing Wzb to the deleterious effects of high temperatures. Contrary to our findings, LaPointe et al. reported that Wzb activity increased with temperature (up to 75 °C) after 30 minutes pre-incubation in the presence of a combination of metal ion Cu2+, Co2+, Fe3+, Mn2+ and Mg2+ [7]. The disparity in the findings here and that of LaPointe et al. [7] implies that for Wzb to be stable at high temperatures, it may require the concerted action of multiple activating metal ions. The inactivation pattern of Wzb after 30 minutes pre-incubation in this study was observed to be similar to what was reported by Wang et al. for a PTP purified from Thermus thermophiles (an extreme thermophilic bacteria) [29].
An apparent theme that has been described for thermostable enzymes is the sequence feature with high percentage of small amino acid residues (Gly, Ala, Ser, and Val). The packing of small residues lowers the entropy cost of packing at high temperature and promotes intimate interactions between helices. Additionally, relative scarcity of cysteine and destabilizing polar residues (36Asp, Asn, Glu, Gln, and Arg) has also been suggested to improve thermostability [38]. Interestingly, both Cap8C and Wzb do not seem to possess most of these sequence features due to their mesophilic origin. This further suggests that their thermostable tendencies may be dependent on the presence of the metal ions. The stability profiles of Cap8C and Wzb in the presence of the 1.0 mM metal ions can be summarized in the order Co2+>Ni2+>Mn2+.
Bacteria can be classified based on their optimal growth temperature into four groups; psychrophiles (-5 to 15°C), mesophiles (15 to 45°C), thermophiles (45 to 80°C), and hyperthermophiles (≥80°C) [39]. Generally, enzymes that are synthesized by thermophiles and hyperthermophiles are regarded as thermozymes, they are usually thermostable, or resistant to irreversible inactivation at high temperature and are thermophilic (optimally active at temperatures between 60 and 125°C) [31]. Psychrozymes originate from psychrophiles, while mesophiles produce enzymes known as mesozymes [40]. S. aureus and L. rhamnosus are typical examples of mesophiles with optimum growth temperature (OGT) of (7–48°C) and (6–41°C) respectively [41]. Lactobacillus rhamnosus Wzb is important for extracellular polysaccharide production and polymerization [42], and Staphylococcus aureus Cap8C is part of a group of enzymes required for the biosynthesis of Capsular polysaccharide [43]. There are reports of mesozymes with thermophilic potentials. For example, two thermophilic restriction endonucleases PtaI and PpaAII from mesophilic cyanobacteria strains have been reported with optimum activity at 65–80°C, which is far above the lethal temperature of the parent organism (40°C) [44]. Lapointe et al. showed that L. rhamnosus Wzb had optimal activity at 75°C which is almost 30 °C higher than its OGT [7]. These interesting albeit unusual patterns are very good and require further investigation.
Merone et al. reported a thermostable PHP phopshotriesterase from Sulfolobus solfataricus that is activated by Mg2+, Co2+ and Ni2+ ions [45]. Further studies on this and other systems could help provide insight into the structural and mechanistic basis for the thermophilic properties of these enzymes. Baros et al. also showed that mutations in the degenerate metal-binding site of E. coli DNA polymerase III PHP domain decrease the overall stability of the polymerase and reduce its activity [46]. This suggest that the observed thermophilic properties of the PHP PTPs might be linked to the unique distorted (α/β)7 barrel of the PHP domain.
4. Materials and Methods
4.1. Reagents
All reagents including salts of divalent metal ions and the reaction substrate (p-nitrophenylphosphate) were purchased from Sigma-Aldrich, UK.
4.2. Cloning, Expression and Purification of Wzb and Cap8C
Modified versions of the molecular cloning protocols of Sambrook and Russell [47] were used for the cloning of plasmid vectors. Codon-optimized sequences of Lactobacillus rhamnosus Wzb (GenBank: AAW22448) and Staphylococcus aureus Cap8c (GenBank: AAB49432), were obtained from GeneArt (Invitrogen). The coding sequences were cloned into the pET-28a(+) vector (Novagen) between the NdeI and XhoI sites for use in protein overexpression. The resulting pET-28a(+) vector carries a tag that contains an N-terminal hexahistidine tag (MGSSHHHHHHSSGLVPRGSHM) to facilitate purification via nickel affinity chromatography. Both plasmid vectors were sequenced (Eurofins Genomics) to verify the sequence of the protein tyrosine phosphatase open reading frame.
Expression and purification of Wzb and Cap8C are as described in Sulyman et al. [48]. Briefly, BL21(DE3)pLysS strain of E. coli were transformed with the pET-28a(+) vectors of Wzb and Cap8C and protein expression was induced with 0.5 mM IPTG. The histidine-tagged Cap8C and Wzb were purified using nickel affinity chromatography, and selected fractions were dialysed in Protein Dilution Buffer (PDB; 25 mM Tris–HCl (pH 7.5), 1 mM DTT, 1 M NaCl and 50% v/v glycerol), and stored at -20 ◦C.
4.3. Assay of Protein Tyrosine Phosphatase Activity
Phosphatase reactions were initiated by the addition of 5 mM pNPP to a reaction mixture containing 0.25 µM Cap8C/Wzb and 92 mM Tris HCl pH 7.5 in the presence or absence of metal ions. Each metal ion (Co2+, Mn2+, Ni2+, Mg2+ and Ca2+) was added at a final concentration of 1.0 mM. The assay was carried out at 75 oC for 15 minutes. The reaction was stopped by the addition of 100 µL Stop Buffer (containing 25 mM Tris-HCl, pH 7.5, 25 mM EDTA, and 0.1% SDS); the absorbance was read at 405 nm (characteristic for the reaction product p-nitrophenol) against blank. The blank contained everything as the test samples except that Cap8C/Wzb was substituted with PDB (Protein Dilution Buffer). Absorbance readings were taken on a Clariostar Micro plate Reader. All measurements of reaction rate were performed in triplicates, and activities are expressed as change in absorbance at 405 nm per minute.
In experiments where the effects of temperature were investigated, reactions were incubated at 30, 40, 50, 60, 70 80 and 90 oC for 15 minutes, after which the reaction was stopped by the addition of 100 µl stop buffer.
In pre-incubation experiments to test the effect of metal ions on thermostability of the enzymes, reaction mixture containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and metal ion (Co2+, Ni2+ or Mn2+) were pre incubated at different temperature (50, 60, 70, 80 and 90 oC) for time 0, 2, 5, 10, 15, 20 and 30 min. The reaction mixtures were cooled on ice then reaction was initiated by the addition of 5 mM pNPP to the reaction mixture and assay was done at 75 oC for 15 min.
4.4. Data and Statistical Analysis
Data analysis was done using Microsoft excel and Duncan multiple range test. Each value is a mean of three determinations ± S.E.M. Difference between means were considered different for p < 0.05.
5. Conclusions
The purification and characterization of Cap8C and Wzb are of particular importance since basic analysis of enzyme complex system is an important step into gaining understanding of their biological functions. Although several enzyme catalytic systems have been characterized in the past decades with most newly found enzymes falling neatly into well-known patterns, the system described here seems to represent a novel paradigm. The unusual preference for high temperature for optimal activities of the two enzymes presents a curious structural and mechanistic challenge. The reason being that the two enzymes are from mesophilic organisms rather than thermophiles. It is common to find enzymes from thermophilic organisms to have preference for high temperature for their catalytic activities. Such enzymes often possess structural domains or motifs that are resistant to heat denaturation. An example is Taq polymerase from Thermus aquaticus, an enzyme routinely used in polymerase chain reactions (PCR), T. aquaticus is found in hot springs and Taq polymerase is remarkably heat-stable both structurally and functionally [49]. Further studies would be required to understand the structural and mechanistic origin of the preference for high temperature by Cap8C and Wzb. It may be that these two enzymes represent a new family of heat-loving enzymes with no immediate obvious known structural features that account for such thermophilic property.
Findings from this study reveal the distinct dependence of Cap8C and Wzb on metal ions and unique catalytic mechanisms which differ from their eukaryotic counterparts. These findings are significant as PTPs have been described as notoriously undruggable, meaning that it is often difficult to find specific inhibitors that are not general, and wide-acting protein inactivators [37,50]. The metal ion dependence properties of Cap8C and Wzb and their mode of activation described here may provide an important starting point for the development of more efficient antibacterial drugs. This study is relevant as it serves as a foundation on which studies on mechanism of S. aureus and L. rhamnosus extracellular polysaccharides and capsular polysaccharides biosynthesis and pathogenesis can be based.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, F.O. and S.M.; methodology, A.A., A.B., and F.O; software, F.O.; validation, A.A. and A.I.; formal analysis, A.A. and A.I.; investigation, A.A. and E.J; resources, F.O. and S.M.; data curation, A.A.; A.B., and F.O.; writing—original draft preparation, A.A., A.B., A.I. and F.O.; writing—review and editing, A.B., S.M., and F.O.; visualization, A.A. and E.J; supervision, A.I., S.M., and F.O..; project administration, S.M. and F.O.; funding acquisition, S.M. and F.O.. All authors have read and agreed to the published version of the manuscript.”.
Funding
This researched was supported by institutional Quality-Related research (QR) and Global Challenges Research Fund (GCRF) funding from Liverpool John Moores University, Liverpool U.K. to A.A. and F.J.O.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
Declare conflicts of interest or state “The authors declare no conflict of interest.” Authors must identify and declare any personal circumstances or interest that may be perceived as inappropriately influencing the representation or interpretation of reported research results. Any role of the funders in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results must be declared in this section. If there is no role, please state “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Cheng, H.-C.; Qi, R. Z.; Paudel, H.; Zhu, H.-J. Regulation and Function of Protein Kinases and Phosphatases. Enzyme Res. 2011, 2011. [Google Scholar] [CrossRef]
- Hale, A. J.; Ter Steege, E.; den Hertog, J. Recent Advances in Understanding the Role of Protein-Tyrosine Phosphatases in Development and Disease. Dev. Biol. 2017, 428, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Goebel-Goody, S. M.; Baum, M.; Paspalas, C. D.; Fernandez, S. M.; Carty, N. C.; Kurup, P.; Lombroso, P. J. Therapeutic Implications for Striatal-Enriched Protein Tyrosine Phosphatase (STEP) in Neuropsychiatric Disorders. Pharmacol. Rev. 2012, 64, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Tonks, N. K. Protein Tyrosine Phosphatases—from Housekeeping Enzymes to Master Regulators of Signal Transduction. FEBS J. 2013, 280, 346–378. [Google Scholar] [CrossRef]
- Shi, L. Manganese-Dependent Protein O-Phosphatases in Prokaryotes and Their Biological Functions. Front. Biosci.-Landmark 2004, 9, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Standish, A. J.; Morona, R. The Role of Bacterial Protein Tyrosine Phosphatases in the Regulation of the Biosynthesis of Secreted Polysaccharides. Antioxid. Redox Signal. 2014, 20, 2274–2289. [Google Scholar] [CrossRef]
- LaPointe, G.; Atlan, D.; Gilbert, C. Characterization and Site-Directed Mutagenesis of Wzb, an O-Phosphatase from Lactobacillus Rhamnosus. BMC Biochem. 2008, 9 (1), 10. [CrossRef]
- Hagelueken, G.; Huang, H.; Mainprize, I. L.; Whitfield, C.; Naismith, J. H. Crystal Structures of Wzb of Escherichia Coli and CpsB of Streptococcus Pneumoniae, Representatives of Two Families of Tyrosine Phosphatases That Regulate Capsule Assembly. J. Mol. Biol. 2009, 392, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. S.; Lee, S. J.; Yoon, H. J.; An, D. R.; Kim, S.-J.; Suh, S. W. Crystal Structures of YwqE from Bacillus Subtilis and CpsB from Streptococcus Pneumoniae, Unique Metal-Dependent Tyrosine Phosphatases. J. Struct. Biol. 2011, 175, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Geno, K. A.; Hauser, J. R.; Gupta, K.; Yother, J. Streptococcus Pneumoniae Phosphotyrosine Phosphatase CpsB and Alterations in Capsule Production Resulting from Changes in Oxygen Availability. J. Bacteriol. 2014, 196, 1992–2003. [Google Scholar] [CrossRef]
- Morona, J. K.; Morona, R.; Miller, D. C.; Paton, J. C. Streptococcus Pneumoniae Capsule Biosynthesis Protein CpsB Is a Novel Manganese-Dependent Phosphotyrosine-Protein Phosphatase. J. Bacteriol. 2002, 184, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Kasumi, T.; Mori, S.; Kaneko, S.; Koyama, Y. Molecular Cloning and Characterization of D-Xylose Isomerase from a Novel Actinobacteria, Thermobifida Fusca MBL 10003. J. Appl. Glycosci. 2011, 59, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Ghodge, S. V.; Fedorov, A. A.; Fedorov, E. V.; Hillerich, B.; Seidel, R.; Almo, S. C.; Raushel, F. M. Structural and Mechanistic Characterization of l -Histidinol Phosphate Phosphatase from the Polymerase and Histidinol Phosphatase Family of Proteins. Biochemistry 2013, 52, 1101–1112. [Google Scholar] [CrossRef]
- Aravind, L.; Koonin, E. V. Phosphoesterase Domains Associated with DNA Polymerases of Diverse Origins. Nucleic Acids Res. 1998, 26, 3746–3752. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Omi, R.; Goto, M.; Miyahara, I.; Manzoku, M.; Ebihara, A.; Hirotsu, K. Crystal Structure of Monofunctional Histidinol Phosphate Phosphatase from Thermus Thermophilus HB8. Biochemistry 2007, 46, 12618–12627. [Google Scholar] [CrossRef] [PubMed]
- Teplyakov, A.; Obmolova, G.; Khil, P. P.; Howard, A. J.; Camerini-Otero, R. D.; Gilliland, G. L. Crystal Structure of the Escherichia Coli YcdX Protein Reveals a Trinuclear Zinc Active Site. 2003.
- Morona, J. K.; Morona, R.; Miller, D. C.; Paton, J. C. Mutational Analysis of the Carboxy-Terminal (YGX) 4 Repeat Domain of CpsD, an Autophosphorylating Tyrosine Kinase Required for Capsule Biosynthesis in Streptococcus Pneumoniae. J. Bacteriol. 2003, 185, 3009–3019. [Google Scholar] [CrossRef] [PubMed]
- Grangeasse, C.; Cozzone, A. J.; Deutscher, J.; Mijakovic, I. Tyrosine Phosphorylation: An Emerging Regulatory Device of Bacterial Physiology. Trends Biochem. Sci. 2007, 32, 86–94. [Google Scholar] [CrossRef]
- Bellomo, E.; Singh, K. B.; Massarotti, A.; Hogstrand, C.; Maret, W. The Metal Face of Protein Tyrosine Phosphatase 1B. Coord. Chem. Rev. 2016, 327, 70–83. [Google Scholar] [CrossRef]
- Mijakovic, I.; Petranovic, D.; Macek, B.; Cepo, T.; Mann, M.; Davies, J.; Jensen, P. R.; Vujaklija, D. Bacterial Single-Stranded DNA-Binding Proteins Are Phosphorylated on Tyrosine. Nucleic Acids Res. 2006, 34, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Standish, A. J.; Salim, A. A.; Capon, R. J.; Morona, R. Dual Inhibition of DNA Polymerase PolC and Protein Tyrosine Phosphatase CpsB Uncovers a Novel Antibiotic Target. Biochem. Biophys. Res. Commun. 2013, 430, 167–172. [Google Scholar] [CrossRef]
- McCleverty, J. A. Comprehensive Coordination Chemistry II; Elsevier Ltd., 2003.
- Chen, Y. The Structure and Function Study of Three Metalloenzymes That Utilize Three Histidines as Metal Ligands. 2013.
- Shi, L.; Kehres, D. G.; Maguire, M. E. The PPP-Family Protein Phosphatases PrpA and PrpB of Salmonella Enterica Serovar Typhimurium Possess Distinct Biochemical Properties. J. Bacteriol. 2001, 183, 7053–7057. [Google Scholar] [CrossRef] [PubMed]
- Hernick, M.; Fierke, C. Mechanisms of Metal-Dependent Hydrolases in Metabolism. Compr. Nat. Prod. II Chem. Biol. Neth. Elseiver 2010, 547–581. [Google Scholar]
- Prejanò, M.; Alberto, M. E.; Russo, N.; Toscano, M.; Marino, T. The Effects of the Metal Ion Substitution into the Active Site of Metalloenzymes: A Theoretical Insight on Some Selected Cases. Catalysts 2020, 10 (9), 1038.
- Wang, Y.; Meng, F.; Zhang, Y. Expression, Purification and Characterization of Recombinant Protein Tyrosine Phosphatase from Thermus Thermophilus HB27. Acta Biochim Biophys Sin 2009, 41, 689–698. [Google Scholar] [CrossRef]
- Vieille, C.; Zeikus, G. J. Hyperthermophilic Enzymes: Sources, Uses, and Molecular Mechanisms for Thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef]
- Li, W. F.; Zhou, X. X.; Lu, P. Structural Features of Thermozymes. Biotechnol. Adv. 2005, 23, 271–281. [Google Scholar] [CrossRef]
- Fields, P. A.; Dong, Y.; Meng, X.; Somero, G. N. Adaptations of Protein Structure and Function to Temperature: There Is More than One Way to ‘Skin a Cat. ’ J. Exp. Biol. 2015, 218, 1801–1811. [Google Scholar] [CrossRef]
- Soulat, D.; Vaganay, E.; Duclos, B.; Genestier, A.-L.; Etienne, J.; Cozzone, A. J. Staphylococcus Aureus Contains Two Low-Molecular-Mass Phosphotyrosine Protein Phosphatases. J. Bacteriol. 2002, 184, 5194–5199. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Z.; Peng, G.; Yin, Y.; Zhao, H.; Cao, Y.; Xia, Y. Purification and Characterization of a Novel Thermostable Extracellular Protein Tyrosine Phosphatase from Metarhizium Anisopliae Strain CQMa102. Biosci. Biotechnol. Biochem. 2006, 70, 1961–1968. [Google Scholar] [CrossRef]
- Vieille, C.; Epting, K. L.; Kelly, R. M.; Zeikus, J. G. Bivalent Cations and Amino-acid Composition Contribute to the Thermostability of Bacillus Licheniformis Xylose Isomerase. Eur. J. Biochem. 2001, 268, 6291–6301. [Google Scholar] [CrossRef]
- John, D.; Nzelibe, H.; Sallau, A.; Albaba, S. Purification and Characterization of Protein Tyrosine Phosphatase from Trypanosoma Evansi.
- Zhang, Z.-Y. Drugging the Undruggable: Therapeutic Potential of Targeting Protein Tyrosine Phosphatases. Acc. Chem. Res. 2017, 50, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Meruelo, A. D.; Han, S. K.; Kim, S.; Bowie, J. U. Structural Differences between Thermophilic and Mesophilic Membrane Proteins. Protein Sci. 2012, 21, 1746–1753. [Google Scholar] [CrossRef]
- Taylor, T. J.; Vaisman, I. I. Discrimination of Thermophilic and Mesophilic Proteins. BMC Struct. Biol. 2010, 10 (Suppl 1), S5. [CrossRef]
- Vieille, C.; Burdette, D. S.; Zeikus, J. G. Thermozymes. Biotechnol. Annu. Rev. 1996, 2, 1–83. [Google Scholar]
- Valík, Ľ.; Medveďová, A.; Čižniar, M.; Liptáková, D. Evaluation of Temperature Effect on Growth Rate of Lactobacillus Rhamnosus GG in Milk Using Secondary Models. Chem. Pap. 2013, 67. [Google Scholar] [CrossRef]
- Bertsch, A.; Roy, D.; LaPointe, G. Enhanced Exopolysaccharide Production by Lactobacillus Rhamnosus in Co-Culture with Saccharomyces Cerevisiae. Appl. Sci. 2019, 9 (19), 4026.
- Gruszczyk, J.; Fleurie, A.; Olivares-Illana, V.; Béchet, E.; Zanella-Cleon, I.; Morera, S.; Meyer, P.; Pompidor, G.; Kahn, R.; Grangeasse, C. Structure Analysis of the Staphylococcus Aureus UDP-N-Acetyl-Mannosamine Dehydrogenase Cap5O Involved in Capsular Polysaccharide Biosynthesis. J. Biol. Chem. 2011, 286, 17112–17121. [Google Scholar] [CrossRef]
- Piechula, S.; Waleron, K.; Światek, W.; Biedrzycka, I.; Podhajska, A. J. Mesophilic Cyanobacteria Producing Thermophilic Restriction Endonucleases. FEMS Microbiol. Lett. 2001, 198, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Merone, L.; Mandrich, L.; Rossi, M.; Manco, G. A Thermostable Phosphotriesterase from the Archaeon Sulfolobus Solfataricus: Cloning, Overexpression and Properties. Extremophiles 2005, 9, 297–305. [Google Scholar] [CrossRef]
- Barros, T.; Guenther, J.; Kelch, B.; Anaya, J.; Prabhakar, A.; O’Donnell, M.; Kuriyan, J.; Lamers, M. H. A Structural Role for the PHP Domain in E. Coli DNA Polymerase III. BMC Struct. Biol. 2013, 13 (1), 8. [CrossRef]
- Sambrook, J.; Russell, D. W. The Condensed Protocols from Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press, 2006.
- Sulyman, A. O.; Fulcher, J.; Crossley, S.; Fatokun, A. A.; Olorunniji, F. J. Shikonin and Juglone Inhibit Mycobacterium Tuberculosis Low-Molecular-Weight Protein Tyrosine Phosphatase a (Mt-PTPa). BioTech 2023, 12 (3), 59.
- Liu, C.; LiCata, V. J. The Stability of Taq DNA Polymerase Results from a Reduced Entropic Folding Penalty; Identification of Other Thermophilic Proteins with Similar Folding Thermodynamics. Proteins Struct. Funct. Bioinforma. 2014, 82, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Heneberg, P. Use of Protein Tyrosine Phosphatase Inhibitors as Promising Targeted Therapeutic Drugs. Curr. Med. Chem. 2009, 16, 706–733. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
AlphaFold-predicted structures of Staphylococcus aureus Cap8C (A) and Lactobacillus rhamnosus Wzb (B). The PDB files (Wzb: AF-A0A806JCW3-F1-; Cap8C: AF-P72369-F1) were downloaded from the AlphaFold Protein Structure Database (https://alphafold.ebi.ac.uk/) and visualised with PyMOL (https://pymol.org/2/). The characteristic PHP motifs are shown for both structures: Motif I (red), Motif II (blue), Motif III (magenta), and Motif IV (yellow).
Figure 1.
AlphaFold-predicted structures of Staphylococcus aureus Cap8C (A) and Lactobacillus rhamnosus Wzb (B). The PDB files (Wzb: AF-A0A806JCW3-F1-; Cap8C: AF-P72369-F1) were downloaded from the AlphaFold Protein Structure Database (https://alphafold.ebi.ac.uk/) and visualised with PyMOL (https://pymol.org/2/). The characteristic PHP motifs are shown for both structures: Motif I (red), Motif II (blue), Motif III (magenta), and Motif IV (yellow).
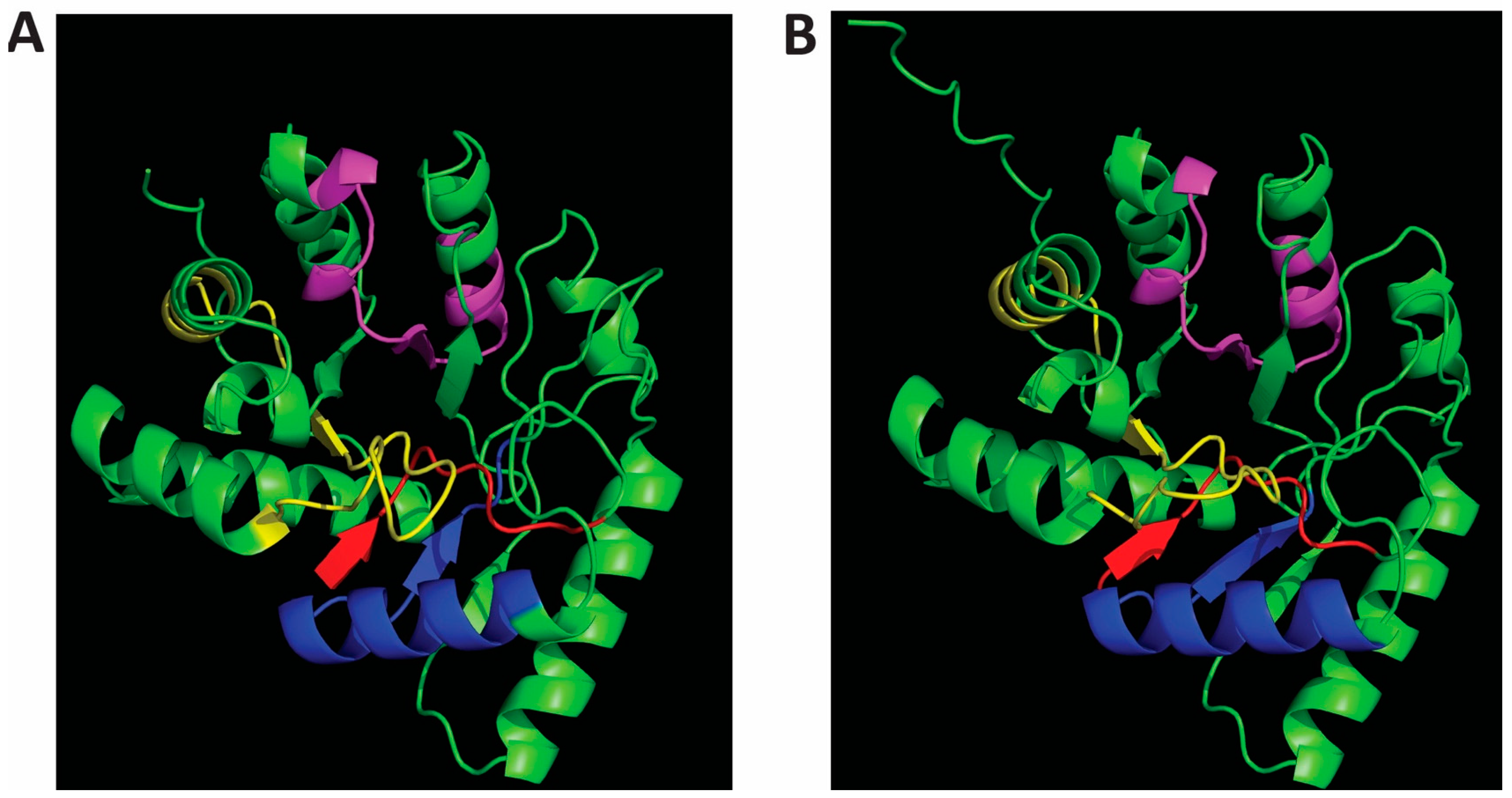
Figure 2.
Effects of divalent metal ions on Cap8c and Wzb activities. Each reaction was initiated by the addition of 5 mM pNPP to a reaction mixture containing 0.25 µM Cap8C/Wzb and 92 mM Tris HCl pH 7.5 in the presence of metal ion. Each metal ion (Co2+, Mn2+, Ni2+, Mg2+ and Ca2+) was added at a final concentration of 1.0 mM. The assay was carried out at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer (containing 25 mM Tris-HCl, pH 7.5, 25 mM EDTA, and 0.1% SDS); the absorbance was read at 405 nm against blank. The blank contained everything as the test samples except that Cap8C/Wzb was substituted with PDB (Protein Dilution Buffer). Each value is a mean of three determinations ± S.E.M. Bars with different alphabets for each enzyme are significantly different (p < 0.05).
Figure 2.
Effects of divalent metal ions on Cap8c and Wzb activities. Each reaction was initiated by the addition of 5 mM pNPP to a reaction mixture containing 0.25 µM Cap8C/Wzb and 92 mM Tris HCl pH 7.5 in the presence of metal ion. Each metal ion (Co2+, Mn2+, Ni2+, Mg2+ and Ca2+) was added at a final concentration of 1.0 mM. The assay was carried out at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer (containing 25 mM Tris-HCl, pH 7.5, 25 mM EDTA, and 0.1% SDS); the absorbance was read at 405 nm against blank. The blank contained everything as the test samples except that Cap8C/Wzb was substituted with PDB (Protein Dilution Buffer). Each value is a mean of three determinations ± S.E.M. Bars with different alphabets for each enzyme are significantly different (p < 0.05).
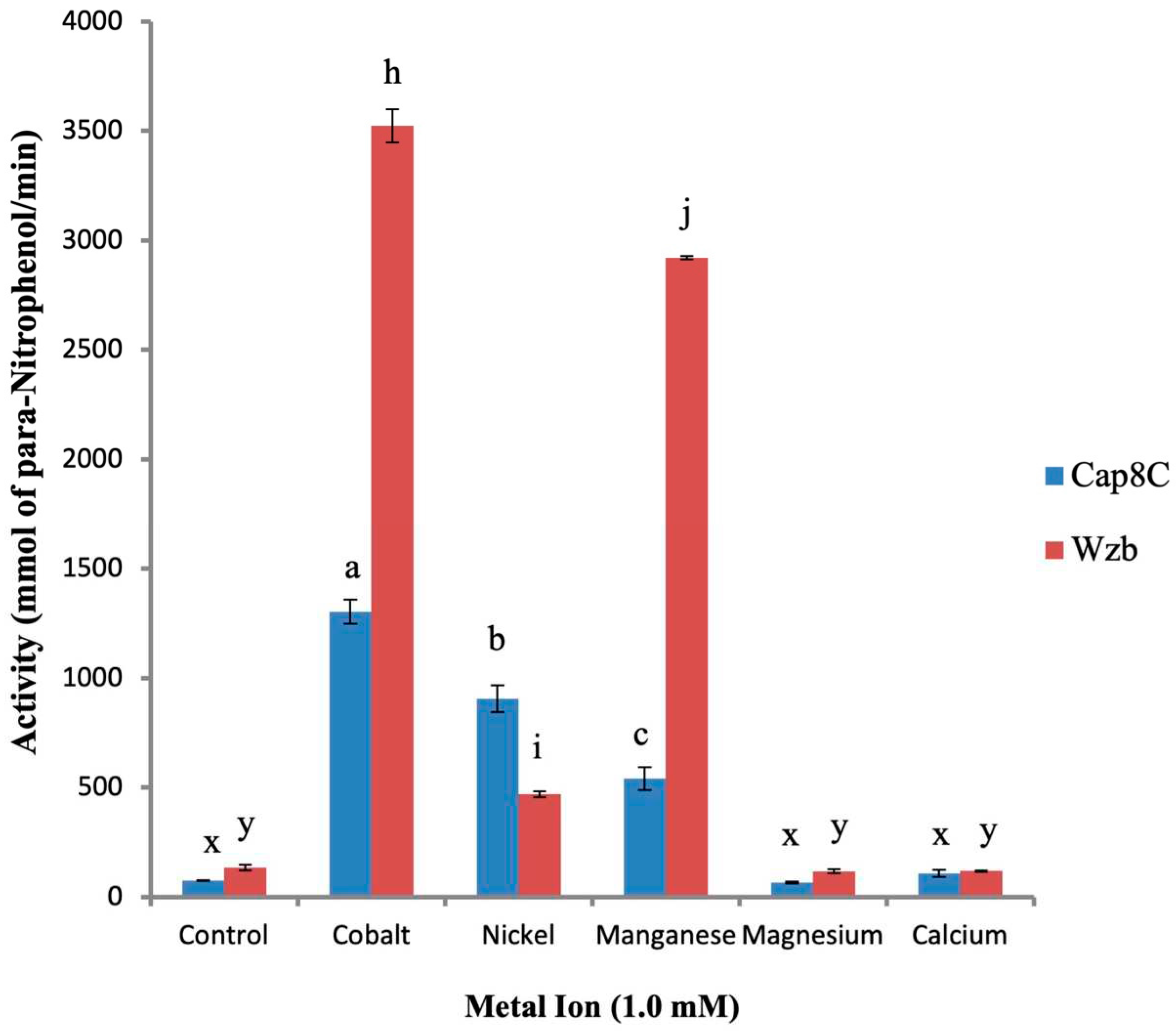
Figure 3.
Effect of increasing concentrations of Co2+, Mn2+, and Ni2+ on the activities of Cap8C (A) and Wzb (B). Each metal ion (Co2+, Ni2+ and Mn2+) was added at a final concentration of 0.25, 0.5, 1.0 and 2.0 mM. The assay was carried out at 75 oC for 15 minutes and treated as described in Figure 2. Values shown at each point represent means ± SD of three replicate determinations.
Figure 3.
Effect of increasing concentrations of Co2+, Mn2+, and Ni2+ on the activities of Cap8C (A) and Wzb (B). Each metal ion (Co2+, Ni2+ and Mn2+) was added at a final concentration of 0.25, 0.5, 1.0 and 2.0 mM. The assay was carried out at 75 oC for 15 minutes and treated as described in Figure 2. Values shown at each point represent means ± SD of three replicate determinations.
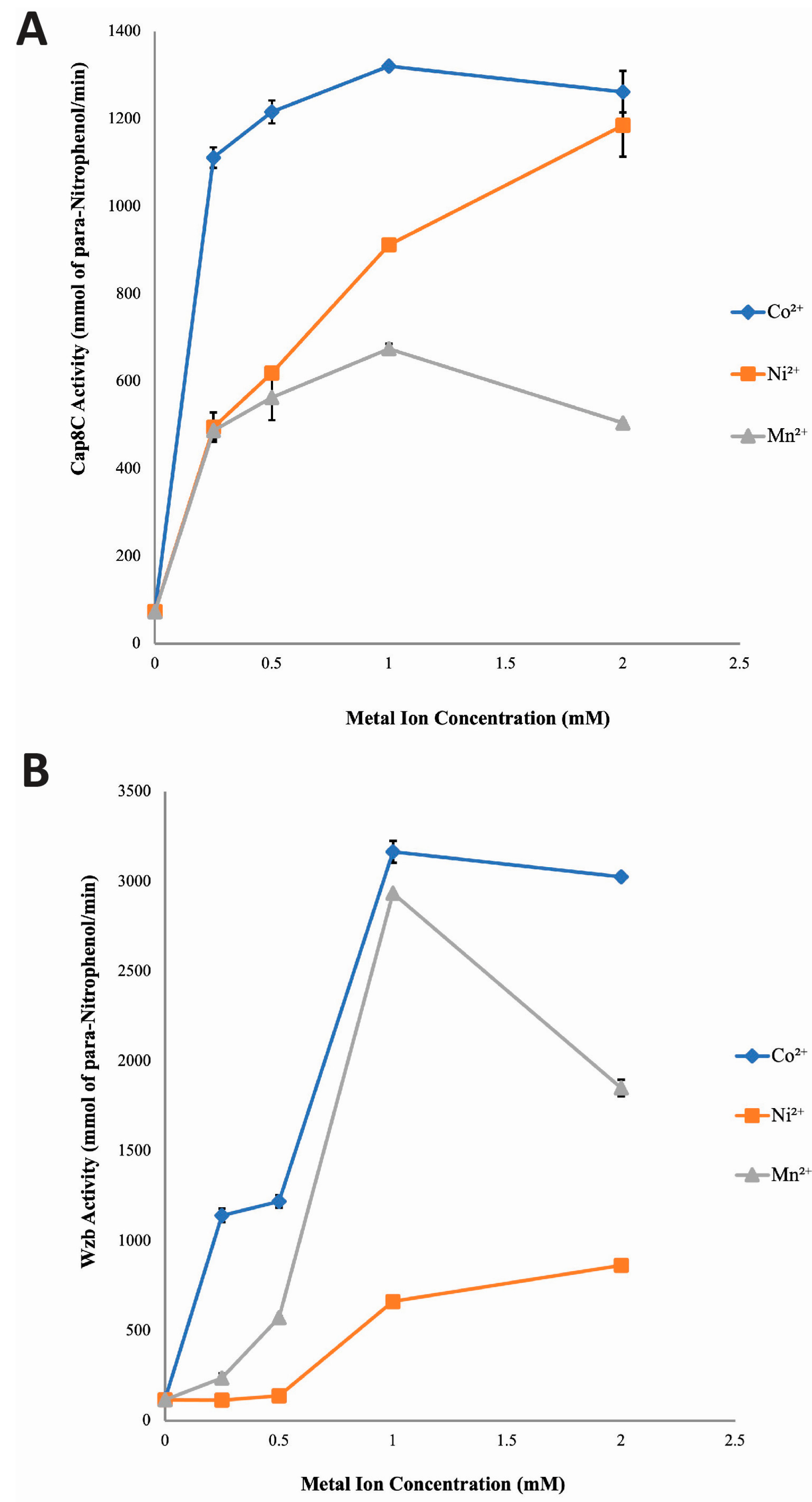
Figure 4.
Temperature dependence of the catalytic activities of Cap8C (A) and Wzb (B) in the presence of Co2+, Ni2+ and Mn2+ as catalytic cofactors. Each metal ion (Co2+, Ni2+ and Mn2+) was added at a final concentration of 1.0 mM, and assays were carried out at different temperatures (30, 40, 50, 60, 70 80 and 90 oC) for 15 minutes. All other conditions are as described in Figure 2. Values shown at each point represent means ± SD of three replicate determinations.
Figure 4.
Temperature dependence of the catalytic activities of Cap8C (A) and Wzb (B) in the presence of Co2+, Ni2+ and Mn2+ as catalytic cofactors. Each metal ion (Co2+, Ni2+ and Mn2+) was added at a final concentration of 1.0 mM, and assays were carried out at different temperatures (30, 40, 50, 60, 70 80 and 90 oC) for 15 minutes. All other conditions are as described in Figure 2. Values shown at each point represent means ± SD of three replicate determinations.
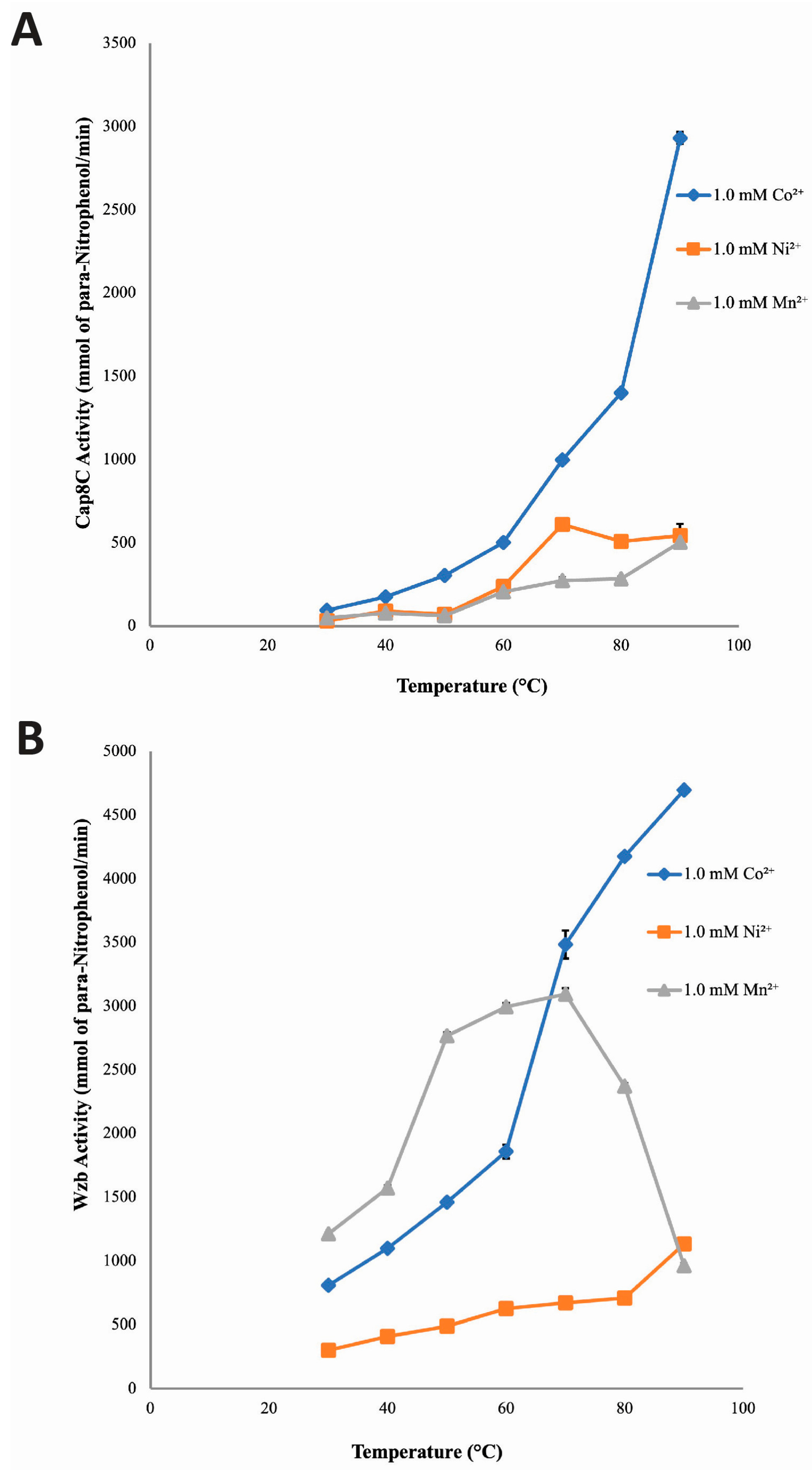
Figure 5.
Thermostability studies of Cap8C (A) and Wzb (B) in the presence of 1.0 mM Mn2+ as catalytic cofactors. Reaction mixtures containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and 1.0 mM Mn2+ were pre incubated at different temperature s(50, 60, 70, 80 and 90 °C) for 0, 2, 5, 10, 15, 20 and 30 minutes. The mixtures were cooled on ice for 5 minutes, after which reactions were initiated by the addition of 5 mM pNPP to the reaction mixture and incubated at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer; the absorbance was read at 405 nm against blank. Values shown at each point represent means ± SD of three replicate determinations.
Figure 5.
Thermostability studies of Cap8C (A) and Wzb (B) in the presence of 1.0 mM Mn2+ as catalytic cofactors. Reaction mixtures containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and 1.0 mM Mn2+ were pre incubated at different temperature s(50, 60, 70, 80 and 90 °C) for 0, 2, 5, 10, 15, 20 and 30 minutes. The mixtures were cooled on ice for 5 minutes, after which reactions were initiated by the addition of 5 mM pNPP to the reaction mixture and incubated at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer; the absorbance was read at 405 nm against blank. Values shown at each point represent means ± SD of three replicate determinations.
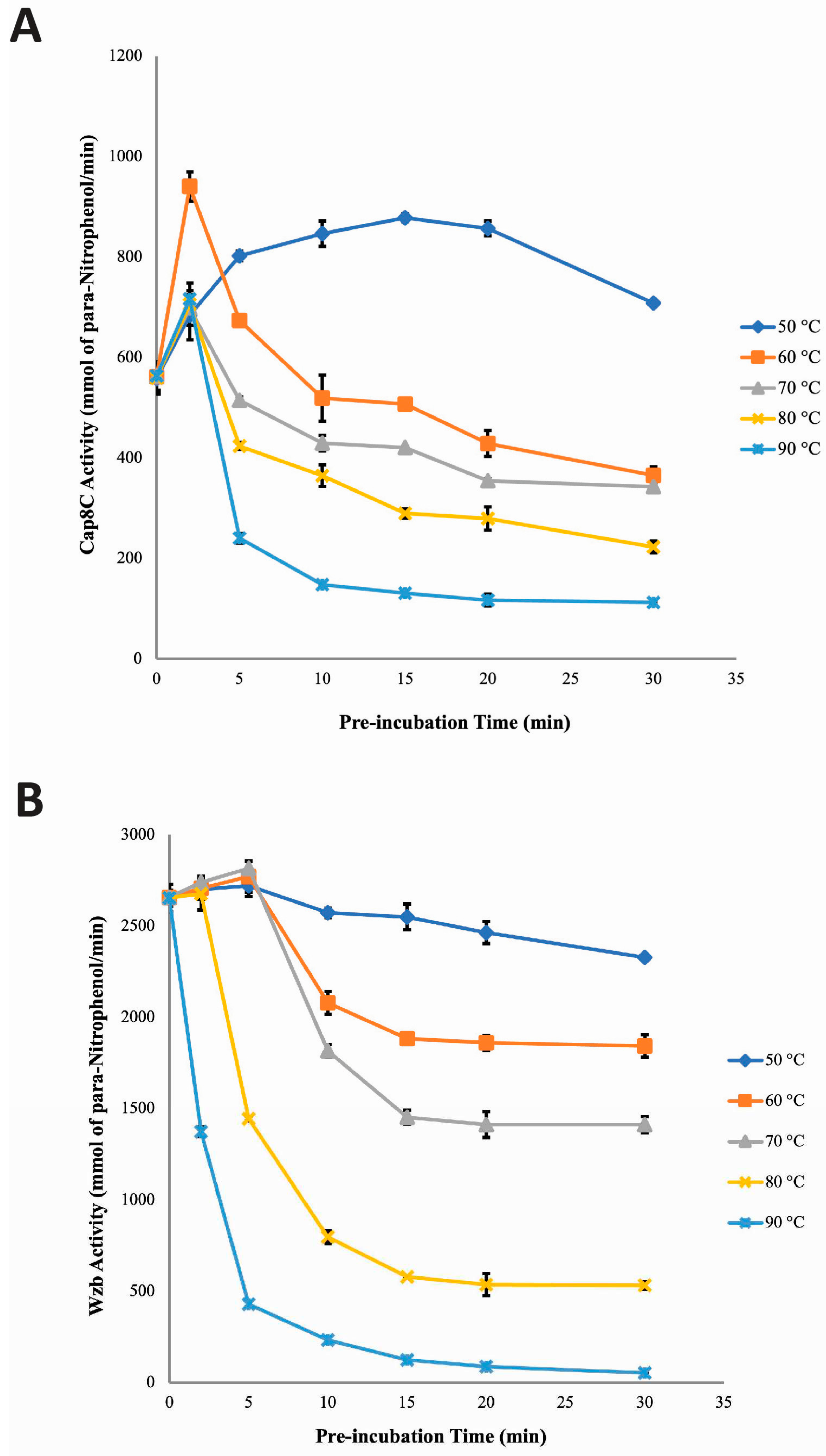
Figure 6.
Thermostability studies of Cap8C (A) and Wzb (B) in the presence of 1.0 mM Co2+ as catalytic cofactors. Reaction mixtures containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and 1.0 mM Co2+ were pre incubated at different temperatures (50, 60, 70, 80 and 90 oC) for 0, 2, 5, 10, 15, 20 and 30 minutes. The mixtures were cooled on ice for 5 minutes, after which reactions were initiated by the addition of 5 mM pNPP to the reaction mixture and incubated at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer; the absorbance was read at 405 nm against blank. Values shown at each point represent means ± SD of three replicate determinations.
Figure 6.
Thermostability studies of Cap8C (A) and Wzb (B) in the presence of 1.0 mM Co2+ as catalytic cofactors. Reaction mixtures containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and 1.0 mM Co2+ were pre incubated at different temperatures (50, 60, 70, 80 and 90 oC) for 0, 2, 5, 10, 15, 20 and 30 minutes. The mixtures were cooled on ice for 5 minutes, after which reactions were initiated by the addition of 5 mM pNPP to the reaction mixture and incubated at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer; the absorbance was read at 405 nm against blank. Values shown at each point represent means ± SD of three replicate determinations.
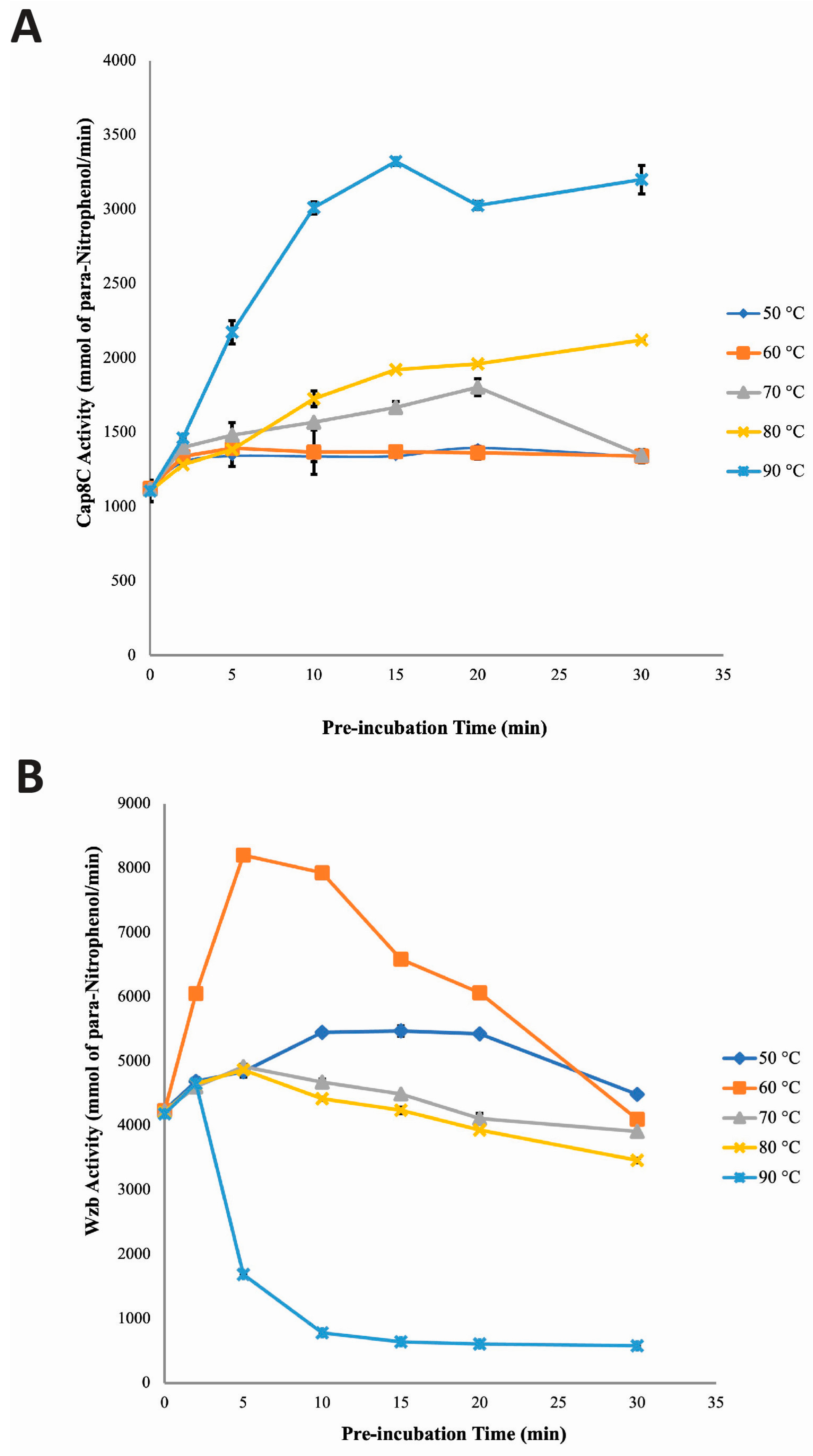
Figure 7.
Thermostability studies of Cap8C (A) and Wzb (B) in the presence of 1.0 mM Ni2+ as catalytic cofactors. Reaction mixtures containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and 1.0 mM Ni2+ were pre incubated at different temperatures (50, 60, 70, 80 and 90 oC) for 0, 2, 5, 10, 15, 20 and 30 minutes. The mixtures were cooled on ice for 5 minutes, after which reactions were initiated by the addition of 5 mM pNPP to the reaction mixture and incubated at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer; the absorbance was read at 405 nm against blank. Values shown at each point represent means ± SD of three replicate determinations.
Figure 7.
Thermostability studies of Cap8C (A) and Wzb (B) in the presence of 1.0 mM Ni2+ as catalytic cofactors. Reaction mixtures containing 0.25 µM Cap8C/Wzb, 92 mM Tris HCl pH 7.5 and 1.0 mM Ni2+ were pre incubated at different temperatures (50, 60, 70, 80 and 90 oC) for 0, 2, 5, 10, 15, 20 and 30 minutes. The mixtures were cooled on ice for 5 minutes, after which reactions were initiated by the addition of 5 mM pNPP to the reaction mixture and incubated at 75 oC for 15 min. The reaction was stopped by the addition of 100 µL stop buffer; the absorbance was read at 405 nm against blank. Values shown at each point represent means ± SD of three replicate determinations.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



