
Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
Staphylococcus aureus are commensal bacteria present on skin and mucosae in both humans and animals, but they can also be found in food, water and in various environments. They are also considered a major pathogen having a great morbidity leading to a wide range of infections. They are included in the list of most virulent and antimicrobic resistant bacterial pathogens (ESKAPE, Escherichia coli, Staphylococcus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, Enterococcus faecalis and faecium)) by the Center for Disease Control and Prevention (CDC), and S. aureus is part of the worldwide threat represented by the presence of antimicrobial resistance (AMR). The outcomes of an epidemiological investigation on 26,430 S. aureus isolates from a global public database (NPDIB; NCBI Pathogen Detection Isolate Browser) support the evidence of significant differences in the distribution of isolates and ARGs (Antimicrobial Resistance Genes) clusters among geographical areas of origin and among sources of the isolates. Moreover, the ARGs cluster patterns suggest an association between the isolates from human and animal populations. This outcome and the large diffusion of the pathogens among human and animal populations support the importance of gaining information on the epidemiology of these infections with a One Health approach.
Keywords:
S. aureus
; One Health
; antimicrobial resistance
; molecular epidemiology
; geographical distribution.
1. Introduction
Staphylococcus aureus are commensal bacteria present on skin and mucosae in both humans and animals, but they can also be found in food, water and in various environments [1,2]. Unfortunately, they are also major pathogens having a great morbidity leading to a wide range of infections including bacteremia, infective endocarditis, complicated skin and soft tissue infections, pleuropulmonary infections, urinary tract infections, toxic shock syndrome and prosthetic devices infections [3]. Most of the cases are observed in healthcare and community settings [4] and it has been estimated that the global mortality due to S. aureus infections, was of about 1 million deaths in 2019 [5].
S. aureus is also present in almost every animal species from wild animals [6] to livestock and pets[7,8], and it can lead to different kinds of infections that can be a health and economic burden, especially in farm animals: mastitis in ruminants [9], septicemia, osteoarticular infections and pododermatitis in poultry[10,11,12,13], exudative epidermitis in piglets [14], cutaneous abscesses [15] and mastitis in rabbits [16].
S. aureus is also considered one of the principal pathogens responsible for foodborne diseases [17]. Indeed, 241,000 illnesses per year are estimated in the United States [18], while in Europe the official diagnosed cases in 2021 were 640 with more than 50 hospitalizations [19]; these latter small numbers probably underestimate the frequency of the infections due to their mild symptoms, not requiring medical attention, and thus leading to unreported foodborne infections in many Countries.
S. aureus are included in the list of most virulent and antimicrobic resistant bacterial pathogens (ESKAPE) by the Center for Disease Control and Prevention (CDC), and S. aureus is part of the worldwide threat represented by AMR. This latter problem, and the large diffusion of these pathogens among human and animal populations support the importance of gaining information on the epidemiology of these infections with a One Health approach. This approach is fundamental to develop efficient control strategies that consider isolates from both humans and animals as well as the possible risks related to bacteria and ARGs spreading via the environment. The access to publicly available databases, collecting isolates from numerous locations and sources, may help to investigate the distribution of AMR genes among isolates, as shown in a previous study [20]. This paper is focused on reporting the results of the epidemiological analysis on the distribution of S. aureus AMR genes based on the isolation characteristics: geography, source and clinical characteristics.
2. Results
2.1. Data description
We considered the worldwide public database NCBI Pathogen Detection Isolate Browser (NPDIB). At the date of 30.04.2022 the public database included 35,026 S. aureus isolates. The isolates were classified in three groups: human associated (HUA), non-human associated (NHA), and unknown (UNK). The HUA category contains samples taken in healthcare settings, while the NHA category includes isolates coming from animals, food and environment. Isolates with little or no information about the origin of isolation have been labelled as unknown origin, thus forming the UNK category. Isolates without information on their geographical origin were not furthermore considered in the analysis. After data verification, we included 26,430 isolates in the epidemiological analysis. Most of the isolates submitted were from North America (USA, Canada and Mexico) and Europe with 35% and 30.8% of the isolates, respectively. Asia accounted for about 20% of the isolates, whereas Oceania, South America, and Africa had the lowest percentages, with frequencies of 3-7% (Figure 1).
Table 1 reports the distribution of the isolates among the different geographical regions by the source characteristics: 63% of them were classified as HUA, 7.8% were NHA, while the remaining isolates had an unknown origin. The HUA group was the most frequent in each geographical area; a significant statistical difference among NHA, HUA and UNK isolates using χ2 test (α=0.05) was observed in each area, except Africa.
2.2. European isolates
Given the substantial number of isolates originating from Europe and the United States, we comprehensively analyzed this dataset. In Europe (Figure 2), the UK had 2212 (27%), followed by Germany 1547 (19%), Denmark 928 (11.4%), Netherlands 754 (9.3%), Switzerland 599 (7.4%), and Italy 515 (6.3%). Other European states had lower isolate frequencies. The proportion of UK isolates was significantly larger than the other Countries, but when the ratio cases/population is considered the proportion of cases is higher for Denmark (154 records/million people), Switzerland (68 records/million people) and Netherlands (42 records/million people), while in UK 33 records/million people and in Germany 19 records/million people were registered.
The trend observed in the worldwide distribution among NHA, HUA and UNK isolates, was observed for the European data as well (Table 2), where a significant statistical difference between the three groups is present in every area considered.
2.3. USA isolates
Within the different States of USA, Massachusetts had the highest frequency of isolates (1530, 17%), followed by California with 15% of the US isolates (1388 isolates), New York state with 13% (1160 isolates), and Iowa with 11% (1007 isolates). The remaining States have frequencies lower than 10% (Figure 3). For the statistical analysis all the US States with a frequency < 2% were included in the category “Other States”.
Table 3 reports the distribution and the statistical differences observed among NHA, HUA and UNK isolates among the States. The results of the statistical analysis
Table 3 reports the distribution and the statistical differences observed among NHA, HUA and UNK isolates among the States. The statistical analysis results showed that a statistically significant difference among the three groups of isolates was observed for each State, except for New York, Iowa, Pennsylvania and Missouri.
A similar scenario to Europe also emerged in the USA when population is considered. Indeed, 314 records/million people were submitted from Iowa, a typical agricultural State, while Massachusetts, which supplied the highest number of isolates, had a proportion of 218 records/million people, New York 59 records/million people and California 35 records/million people.
2.4. Resistance gene distribution
Among the 67 ARGs reported in the database only those with a total prevalence >2% were considered for the statistical analyses. Furthermore, regulatory genes such as blaI, blaR1 for blaZ and mecI and mecR1 for mecA were excluded from this study. The most frequent ARGs identified were those conferring resistance to the tetracycline antimicrobial family with more than 77,000 positive identifications and those responsible for resistance against penams and fosfonic acid, with 51,459 and 45,659 identifications respectively; ARGs related to resistance to aminoglycosides and fluoroquinolones had both more than 36,000 positive identifications and genes related to resistance to macrolides had 15,784 positive identifications. Our prior study examined single ARG prevalence in depth [20].
2.5. Cluster analyses
In order to identify a possible pattern in the distribution of the ARGs, a cluster analysis has been performed on the dataset, not only to recognize a particular asset in the AMR of the various isolates, but also to identify potential associations between the different patterns and the source or the region of origin of the isolates. A detailed description of the patterns obtained after cluster analysis has been reported in our previous work [20]. Briefly, Clusters 2 and 3 were identified as the clusters with the lowest presence of ARGs while Clusters 4, 5 and 6 have high rates of ARGs related to nine different antibiotic classes. Clusters 1 and 7 showed a mild resistance pattern with a progressively increase of ARGs frequency from Cluster 7 to Cluster 1. In Figure 4 is visualized the distribution of the 7 clusters identified in the subset of isolates considered in this paper.
2.5.1. Association between gene-cluster and geographical area of submission
The statistical analysis was performed to identify possible associations between the origin of the isolates and the cluster membership (Table 4, Table 5 and Table 6). The statistical analysis showed large and significant variations among clusters in relation to the geographical origin (Table 4). Indeed, North America had the highest significant prevalence of Clusters 4 and 5 (64.9% and 73.5%, respectively); Cluster 1 is prevalent in Europe with 2034 isolates, representing more than half of all isolates in this clusters (53.7%); Clusters 2 with 1153 isolates (40.3%), Cluster 7 with 2228 isolates (41.4%) and Cluster 3 with 1138 isolates (33.3%) are also more frequently reported in Europe, even if with lower prevalence. Cluster 6 showed the highest prevalence in Other Asian Countries with 1019 isolates (56.3%).
2.5.2. European and USA isolates
Since most of the isolates were reported from Europe and USA, and the numbers between these two areas were comparable, we also analyzed the different distribution of clusters between these two areas. The results of the χ2 analysis (Figure 5a) and of the residues (Figure 5b) confirmed significant differences in the clusters frequencies between European and USA isolates. These differences are particularly sigificant for Clusters 1, 5, 6 and 7.
When the clusters distribution was analysed within European Countries, great differences were also observed (Table 5). Indeed, Cluster 1 was mainly recovered in UK, while Germany supplied about one third of the isolates of Cluster 7, as well as Switzerland for Cluster 4. More generally, each Country appears to be characterized by one or two clusters that have a prevalence largely higher than all the others.

Table 5.
Geographical distribution of clusters in Europe and statistical differences among Countries.
Table 5.
Geographical distribution of clusters in Europe and statistical differences among Countries.
| State | Cluster 1 N (%) |
Cluster 2 N (%) |
Cluster 3 N (%) |
Cluster 4 N (%) |
Cluster 5 N (%) |
Cluster 6 N (%) |
Cluster 7 N (%) |
|---|---|---|---|---|---|---|---|
| United Kingdom | 1243e 1 (61.1) |
377b (32.6) |
150d (13.2) |
273e (29.3) |
11b (3.3) |
53b (16.2) |
105f (4.7) |
| Other Europe2 | 408d (20.1) |
252a (21.9) |
266c (23.3) |
157c, e (16.8) |
89a (26.9) |
107d (32.8) |
310d (13.9) |
| Germany | 227 b (11.2) |
164b (14.2) |
210b, c (18.5) |
90b (9.6) |
90a (27.2) |
57b (17.4) |
709b (31.8) |
| Denmark | 90a (4.4) |
153a (13.3) |
289a (25.4) |
12a (1.3) |
61a (18.4) |
3a (0.9) |
320a (14.4) |
| Netherlands | 7c (0.3) |
82b (7.1) |
102b, c (9) |
0d (0.0) |
1b (0.3) |
0a (0.0) |
562c (25.2) |
| Switzerland | 11c (0.5) |
56b (4.9) |
56b,d (4.9) |
364f (39) |
41a (12.4) |
23b, d (7) |
48e (2.2) |
| Italy | 48a (2.4) |
69a, b (6) |
65b, c (5.7 ) |
37b, c (4) |
38a (11.5) |
84c (25.7) |
174a (7.8) |
| Total | 2034 (100) |
1153 (100) |
1138 (100) |
933 (100) |
331 (100) |
327 (100) |
2228 (100) |
1 Values with different superscripts among rows statistically differ at χ2 test or Fisher’s exact test (α=0.05). 2 Other Europe includes: Austria, Belgium, Belarus, Croatia, Finland, Greece, Latvia, Lithuania, Luxembourg, Poland, Portugal, Czech Republic, Romania, Serbia, Slovenia and Hungary.
The same analysis applied to USA (Table 6) gave similar results with the distribution of clusters largely associated to a specific State. Indeed, Cluster 4 was mainly associated to Massachusetts isolates, Cluster 5 to New York isolates, Cluster 6 to California and Cluster 7 from Iowa isolates.
Table 6.
Geographical distribution of clusters in USA and statistical differences among States.
| State | Cluster 1 N (%) |
Cluster 2 N (%) |
Cluster 3 N (%) |
Cluster 4 N (%) |
Cluster 5 N (%) |
Cluster 6 N (%) |
Cluster 7 N (%) |
|---|---|---|---|---|---|---|---|
| Other States1 | 95b, d, f, h 2 (20.9) |
290a (35.2) |
253d (39.9) |
496c (14.4) |
387b (13.8) |
13a, b, c (22.7) |
365b (46.1) |
| Massachusetts | 15i (3.3) |
7g (0.8) |
18a (2.8) |
1191b (34.4) |
268b (9.6) |
1c (1.8) |
30a, b, c (3.8) |
| California | 87a, b, c, d, e, f, g (19.1) |
170a, b (20.6) |
10a (1.6) |
587a (17) |
492a (17.5) |
20a (35.1) |
22a (2.8) |
| New York | 101 c, g (22.2) |
122a, b, c (14.8) |
97b (15.3) |
114e (3.3) |
661c (23.6) |
13a, b (22.7) |
52c (6.6) |
| Iowa | 31h (6.8) |
90b, c, d, e, f (10.9) |
78b (12.3) |
402a (11.6) |
227b (8.1) |
1b, c (1.8) |
178b (22.5) |
| Pennsylvania | 54a, c, e, g (11.9) |
37c, d, e, f (4.5) |
36b (5.7) |
231a (6.7) |
221a (7.9) |
5a, b, c (8.8) |
18a, c (2.3) |
| Maryland | 35e, f, g (7.7) |
18e, f (2.2) |
21b (3.3) |
85c (2.5) |
251c (9) |
3a, b, c (5.3) |
16a, c (2) |
| Ohio | 11b, d, f, h, i (2.4) |
32a, b, c, d, e, f (3.9) |
19b (3) |
143a (4.1) |
122a (4.4) |
0a, b, c (0.0) |
36d (4.5) |
| Missouri | 9d, h, i (2) |
46a, b (5.6) |
86c (13.6) |
20e (0.6) |
120a (4.3) |
1a, b, c (1.8) |
70b (8.8) |
| Michigan | 17a, b, c, d, e, f, g, h (3.7) |
12d, f (1.5) |
16b (2.5) |
185d (5.4) |
49b (1.8) |
0a, b, c (0.0) |
5a, c (0.6) |
| Total | 455 (100) |
824 (100) |
634 (100) |
3454 (100) |
2798 (100) |
57 (100) |
792 (100) |
1 Other States includes: Alabama, Alaska, Arizona, Arkansas, Colorado, Connecticut, Delaware, Florida, Georgia, Hawaii, Idaho, Illinois, Indiana, Kansas, Kentucky, Louisiana, Maine, Minnesota, Mississippi, Montana, Nebraska, Nevada, New Hampshire, New Jersey, New Mexico, North Carolina, North Dakota, Oklahoma, Oregon, Rhode Island, South Carolina, South Dakota, Tennessee, Texas, Utah, Vermont, Virginia, Washington, West Virginia, Wisconsin, Wyoming.2Values with different superscripts among rows statistically differ at χ2 test or Fisher’s exact test (α=0.05).
2.6. Isolates from humans (clinical sources)
We investigated in detail also human (clinical) isolates which are the ones with the more precise characterization in the database. They are more frequently classified in Clusters 4, 5 and 7 (Table 7), and the statistical analysis of the frequencies among geographical areas supports the difference observed in the general database. Most isolates in North America were classified in Clusters 4 and 5, while Cluster 1 was the most frequent in Europe, supplying nearly 50% of the isolates classified in this cluster. Cluster 2 isolates came mainly from North America, Europe and Asian Countries, except China, where Cluster 6 was observed with the highest frequency.
When the statistical analysis was performed within European Countries, the results showed as 75% of the isolates in Cluster 1 were from UK. Clusters 3 and 7 were more frequently associated to Germany, while Italy supplied about one third of Cluster 6 isolates (Table 8). The same analysis performed on USA isolates (
Table 1Table 9), showed that Clusters 4 and 5 represented more than 70% of the total HUA isolates, Massachusetts was the area where Cluster 4 isolates were more frequently isolated, while New York State was the major source of Cluster 5 isolates.
Table 8.
Distribution and Statistical differences of HUA isolates among clusters in European Countries.
Table 8.
Distribution and Statistical differences of HUA isolates among clusters in European Countries.
| State | Cluster 1 N (%) |
Cluster 2 N (%) |
Cluster 3 N (%) |
Cluster 4 N (%) |
Cluster 5 N (%) |
Cluster 6 N (%) |
Cluster 7 N (%) |
|---|---|---|---|---|---|---|---|
| United Kingdom | 880e 1 (75.5) |
244b (40.6) |
39f (8.1) |
217g (65.7) |
7c (4.5) |
25a, c (26.3) |
28c (4.6) |
| Other Europe2 | 122d (10.5) |
122b, c (20.3) |
97a, c, d, e (20.2) |
69f, g (20.8) |
21a (13.4) |
14a, b, c (14.7) |
148a (24.2) |
| Germany | 74b (6.4) |
106b, c (17.6) |
132d, e (27.4) |
15d, e (4.5) |
47b (29.9) |
14a, b, c (14.7) |
198b (32.4) |
| Denmark | 53a (4.5) |
9a (1.5) |
37a, b, c, d, e (7.7) |
2a, b, c, d, e (0.6) |
14a (8.9) |
2a, b, c (2.1) |
36a, b (5.9) |
| Netherlands | 7c (0.6) |
72c (12) |
97b (20.2) |
0b, e (0.0) |
0c (0.0) |
0c (0.0) |
106b (17.3) |
| Switzerland | 5b, c (0.4) |
6a (1) |
31a, b, c, d, e (6.4) |
8a, c, d, f, g (2.4) |
41d (26.1) |
7b, d (7.4) |
17a (2.8) |
| Italy | 24b (2.1) |
42a, b, c (7) |
48c, e (10) |
20c, f (6) |
27b (17.2) |
33d (34.8) |
78a, b (12.8) |
| Total | 1165 (100) |
601 (100) |
481 (100) |
331 (100) |
157 (100) |
95 (100) |
611 (100) |
1 Values with different superscripts among rows statistically differ at χ2 test or Fisher’s exact test (α=0.05). 2 Other Europe includes: Austria, Belgium, Belarus, Croatia, Finland, Greece, Latvia, Lithuania, Luxembourg, Poland, Portugal, Czech Republic, Romania, Serbia, Slovenia and Hungary.
Table 9.
Distribution and statistical differences of HUA isolates among clusters in USA countries.
| State | Cluster 1 N (%) |
Cluster 2 N (%) |
Cluster 3 N (%) |
Cluster 4 N (%) |
Cluster 5 N (%) |
Cluster 6 N (%) |
Cluster 7 N (%) |
|---|---|---|---|---|---|---|---|
| Other States1 | 56b, d, f, g, h, i, j, k 2 (16.4) |
166a(27) |
145d (29.3) |
363f (12.9) |
204b, c (9.2.) |
10a, b, c (20.8) |
209b (34.7) |
| Massachusetts | 10l (2.9) |
6g (1) |
13b (2.6) |
974b (34.7) |
207c (9.3) |
1c (2.1) |
28a (4.7) |
| California | 49a, b, c, d, e, f, g, h, i, j, k (14.3) |
113a, b (18.4) |
9a, b (1.8) |
340a (12.1) |
292a (13.2) |
14a (29.1) |
12a (2) |
| New York | 96c (28.1) |
107a, b, c, e (17.4) |
85c (17.1) |
112e (4) |
616d (27.7) |
13a, b (27.1) |
39a (6.5) |
| Iowa | 31h, i, j, k (9) |
89b, c, d, e, f (14.5) |
78c (15.7) |
401a (14.3) |
227b (10.2) |
1b, c (2.1) |
176b (29.3) |
| Pennsylvania | 54a, c, e (15.7) |
36c, d, e, f (5.9) |
35c (7) |
226a, f (8) |
219a (9.9) |
5a, b, c (10.4) |
18a (3) |
| Maryland | 22a, b, c, d, e, f, g (6.4) |
13f (2.1) |
11a, c (2.2) |
56c (2) |
166d (7.5) |
3a, b, c (6.3) |
11a, c (1.8) |
| Ohio | 8b, d, f, g, h, i, j, k, l (2.3) |
28a, b, c, d, e, f (4.6) |
19c (3.8) |
136a, f (4.8) |
119a (5.4) |
0a, b, c (0.0) |
34c (5.7) |
| Missouri | 9d, g, i, k, l (2.6) |
46a, b (7.5) |
86e (17.3) |
20e (0.7) |
120a (5.4) |
1a, b, c, (2.1) |
70b (11.6) |
| Michigan | 8e, f, g, j, k, l (2.3) |
10d, f (1.6) |
16c, d (3.2) |
183d (6.5) |
49b, c (2.2) |
0a, b, c (0.0) |
4a (0.7) |
| Total | 343 (100) |
614 (100) |
497 (100) |
2811 (100) |
2219 (100) |
48 (100) |
601 (100) |
1 Other States includes: Alabama, Alaska, Arizona, Arkansas, Colorado, Connecticut, Delaware, Florida, Georgia, Hawaii, Idaho, Illinois, Indiana, Kansas, Kentucky, Louisiana, Maine, Minnesota, Mississippi, Montana, Nebraska, Nevada, New Hampshire, New Jersey, New Mexico, North Carolina, North Dakota, Oklahoma, Oregon, Rhode Island, South Carolina, South Dakota, Tennessee, Texas, Utah, Vermont, Virginia, Washington, West Virginia, Wisconsin, Wyoming. 2 Values with different superscripts among rows statistically differ at χ2 test or Fisher’s exact test (α=0.05).
2.7. Isolates from animals, food and environment
As stated before, the amount of NHA isolate was very low representing only 7.8% of the whole database. Nonetheless, the data reported in Supplementary Table 1 show that Cluster 7 is mainly associated to China and Europe, with 42.2% and 31.2% of the isolates respectively, while Cluster 2 is prevalent in Europe and North America (40.7% and 36.1% respectively). Notably the distribution of the NHA isolates in Europe is characterized by the complete absence of them in Clusters 5 and 6 (Supplementary Table 2), whilst Cluster 7 is more present in Germany (61.3%). In USA most of the isolates fall in Cluster 5 (166) with a great contribution from Maryland (42.2%), while there are no isolates in Cluster 6 (Supplementary Table 3).
3. Discussion
S. aureus is a highly adapted microorganism, with different lineages associated with specific hosts [21]; while a change of the major host is rare, spillover events can be more common and lead to infection in unusual hosts [22]. The risk of transmission should consider not only zoonotic or antropozoonotic (reverse-zoonotic) infections but also the ARGs spread among species, through pathways that still need to be investigated [23]. The presence of these risks supports the importance of epidemiological studies on the characteristics and distribution of the isolates with different genetic patterns [24]. Publicly available datasets, collecting isolates from various Countries and sources, allow to monitor the epidemiology of S. aureus, and foresee changes in the distribution of different lineages and ARGs [25].
The analysis of the database considered in this study supports the evidence of significant differences in the distribution of isolates and ARGs clusters among geographical areas of origin and sources of the isolates. Geographical differences in the genetic characteristics of isolates were already known in the case of bovine mastitis [26,27,28], but these differences were only recently investigated in the case of human isolates [24,29,30]. More than 60% of the records originated in Europe and USA, suggesting the relevance of the problem in these areas, but the proportion of records from Asia (19%) is not negligible and confirms the increasing importance of S. aureus infections and AMR problems also in this area [25,31,32,33].
As reported in other studies, the scarcity of information derived from low-middle income Countries, also evident in this study, reflecting the limits of the local healthcare systems where resources for the control and prevention of AMR are limited [34,35]. Indeed, one of the limits of this study is represented by the voluntariness of the submissions of the isolates, and the uneven frequencies of reporting information among Countries could be attributed to economic limitations, missed diagnosis or the lack of interest in sharing the data.
Most records are related to human clinical cases, and relatively few to environmental, animal or food isolates. This unbalance could be a source of bias in the analysis when the different sources are compared. However, the close values of the NHA isolates from North America, USA and China suggest that this problem has common features in these highly populated areas. This may also be due to the low prevalence of severe illnesses in humans, usually not requiring hospitalization, leading to an underestimation of the frequency of these infections.
Despite the population size and the Public Health conditions are similar within different European Countries, the frequency and relevance of the problem seem to be different, suggesting the presence of local factors that could influence the spread and the characteristics of the infections. For example, Netherlands and Denmark have significant food animal populations, mainly cows and pigs, that may play a role in the epidemiology of S. aureus infections as already shown for MRSA infections [36].
These results support the importance of a One Health approach to investigate these infections and the need for a larger number of isolates from animal, environmental and food sources to confirm the pattern emerged with the data considered in this study. The analysis of the distribution of ARGs clusters among and within continents fully supports the previous observations and suggests that the circulation of the different isolates is associated with relatively small areas, and the development of AMR may be mainly due to the therapeutical protocols applied locally, and it cannot be generalized. Indeed, the results of this study suggest that the ARGs clusters characterized by higher AMR (Clusters 4,5, and 6) [20] are recovered with significantly higher frequency in North America, while the other clusters, characterized by lower AMR patterns originate mainly from Europe. These differences are also supported by the evidence of different distributions of the clusters even when relatively homogeneous economic and political areas (USA and Europe) were compared.
In Europe the prevalent cluster among HUA isolates is Cluster 1; this cluster is mainly associated to ARGs towards fluoroquinolones, penam and tetracycline, other prevalent clusters are number 2 and 7 that involve resistance toward fosfonic acid, tetracycline and penams. It’s interesting to note that in European States the NHA isolates with ARGs are also included in Clusters 2 and 7, which are characterized by a quite high prevalence of ARGs towards rifamycin and fosfonic acid which are not allowed for veterinary use [37]. Clusters 5 and 6 are both characterized by ARGs for nucleosides [20], in Europe this antimicrobial class is not authorized for veterinary use, and this could explain why we didn’t find any NHA isolates in Clusters 5 and 6 among European isolates [37].
Similarly, in USA, Massachusetts was the major source of Cluster 4, while New York State was for Cluster 5. These differences may result from a higher transmission frequency of genetically similar isolates in the specific geographical area and/or from applying of different therapeutical protocols among the different States. Indeed, Cluster 4 is characterized by a high frequency of genes leading to fluorquinolones and glycopeptides resistance, while Cluster 5 is characterized by a high frequency of genes related to penams and nucleosides resistance [20]. It’s important to highlight that Iowa is the single State with the highest frequency of insolates in Cluster 7, supporting the hypothesis of an association with livestock. Indeed, this cluster is related mostly to ARGs directed against tetracycline and penams [20] which are largely applied for livestock treatments [38] .
Cluster 7 was also frequently observed within Asian isolates, suggesting a similar epidemiological patterns as in the other continents.
Overall, the results of this study support the evidence that the occurrence of isolates with peculiar characteristics, including higher morbidity and AMR, may be identified, and should be considered [29]. It also implies that the preventive measures to reduce the occurrence and development of AMR should be aimed at the clusters of S. aureus with the highest frequency at a local level [24].
4. Materials and Methods
4.1. NCBI pathogen detection isolate browser and antibacterial data (NPDIB)
With the NCBI pathogen detection isolate browser (NPDIB) more than one million isolates from 80 different bacteria are available. The strains submitted to the database, used in this epidemiological study, were analyzed with the same parameters described in a previous study [20]. Identification data from the database have been exported and organized into columns with Microsoft Excel™. Each column represented an AMR gene, and the value of the cell was associated with a dichotomic variable: 1 if the ARG was present, and 0 if it was not. The information in the other columns (e.g., source of isolation, geographical area, etc.) were kept as in the original database.
4.2. Statistical analysis
The statistical analysis has been performed on SPSS 28.0.1.1 (IBM Corp., USA, 2022). We used a χ2 test with Bonferroni adjustment in order to analyse the frequency distribution. When cell numerosity was below 6 a Fisher’s exact test was applied instead of χ2 test. With the aim to classify isolates based on the different combination of AMR genes, cluster analysis was performed with the following parameters: squared Euclidean distance, Ward's agglomeration method and truncation at 20% of total distance [39]. Cluster analysis is a multivariate technique which allows to group isolates on the base of their characteristics (e.g. AMR genes).
5. Conclusion
The availability of public databases collecting genetic information on pathogens allows to investigate the epidemiology of the different pathogens and the ARGs patterns. Data analysis on S. aureus isolates from NPDIB databases confirmed that both isolate distribution and ARGs patterns are peculiar of different geographical areas. This can be related to the diagnostic capability, to different local therapeutical approaches, the different availability of antimicrobial molecules, and the epidemiology of these bacteria in animals, and particularly livestock. From a One Health point of view this type of analysis is crucial, because allows to identify both similarities and differences among human, animal and environmental isolates, and the possible interaction between the groups.
The quantity and quality of information collected in the database should be implemented in order to intensify the surveillance of S. aureus resistance, not only in human medicine purview, but also in veterinary medicine and in the field of food and environmental contamination. A larger number of isolates from different sources and a periodical analysis of prevalence and ARGs patterns are pivotal in developing effective control programs.
Supplementary Materials
Supplementary Table 1: distribution and statistical difference of NHA isolates among clusters in different geographical regions. Supplementary Table 1: distribution and statistical difference of NHA isolates among European Countries. Supplementary Table 2: distribution and statistical difference of NHA isolates among clusters in USA Countries.
Author Contributions
Conceptualization, G.M. and V.M.S.; methodology, A.Z. and A.S.; formal analysis, A.Z.; resources, F.Z.; data curation, G.L. and A.S.; writing—original draft preparation, A.Z., V.M.S. and F.Z.; writing—review and editing, G.M., P.A.M. and A.S.; funding acquisition, A.Z. and L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by FEASR—Programma di Sviluppo Rurale 2014–2020 Misura 16.1 project MOOH, PRIN 2017MZ5KWM_001 project “SAFE MILK: OMICS SCIENCE FOR MILK SAFETY AND QUALITY” and University of Milan PSR2021 line 6 One Health Action Hub.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: https://www.ncbi.nlm.nih.gov/pathogens/isolates/#taxgroup_name:%22Staphylococcus%20aureus%22 (accessed on 30 April 2022).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gajdács, M. The Continuing Threat of Methicillin-Resistant Staphylococcus Aureus. Antibiotics 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, L.R.; Haack, S.K.; Johnson, H.E.; Brennan, A.K.; Isaacs, N.M.; Spencer, C. Staphylococcus Aureus and Methicillin-Resistant s. Aureus (MRSA) at Ambient Freshwater Beaches. J Water Health 2015, 13. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin Microbiol Rev 2015, 28. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.L.; Melles, D.C.; Vos, M.C.; Van Leeuwen, W.; Van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The Role of Nasal Carriage in Staphylococcus Aureus Infections. Lancet Infectious Diseases 2005, 5. [Google Scholar] [CrossRef] [PubMed]
- Ikuta, K.S.; Swetschinski, L.R.; Robles Aguilar, G.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Davis Weaver, N.; Wool, E.E.; Han, C.; Gershberg Hayoon, A.; et al. Global Mortality Associated with 33 Bacterial Pathogens in 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. The Lancet 2022, 400. [Google Scholar] [CrossRef] [PubMed]
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus Aureus Epidemiology in Wildlife: A Systematic Review. Antibiotics 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ronholm, J. Staphylococcus Aureus in Agriculture: Lessons in Evolution from a Multispecies Pathogen. Clin Microbiol Rev 2021, 34. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moein, K.A.; Samir, A. Isolation of Enterotoxigenic Staphylococcus Aureus from Pet Dogs and Cats: A Public Health Implication. Vector-Borne and Zoonotic Diseases 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Liao, G.; Wu, Z.; Lv, J.; Chen, W. Prevalence and Characterization of Staphylococcus Aureus Isolates from Subclinical Bovine Mastitis in Southern Xinjiang, China. J Dairy Sci 2020, 103. [Google Scholar] [CrossRef] [PubMed]
- Szafraniec, G.M.; Szeleszczuk, P.; Dolka, B. Review on Skeletal Disorders Caused by Staphylococcus Spp. in Poultry. Veterinary Quarterly 2022, 42. [Google Scholar]
- Xu, Y.; Rivas, J.M.; Brown, E.L.; Liang, X.; Höök, M. Virulence Potential of the Staphylococcal Adhesin CNA in Experimental Arthritis Is Determined by Its Affinity for Collagen. Journal of Infectious Diseases 2004, 189. [Google Scholar] [CrossRef] [PubMed]
- Ali, Y.; Islam, M.A.; Muzahid, N.H.; Sikder, M.O.F.; Hossain, M.A.; Marzan, L.W. Characterization, Prevalence and Antibiogram Study of Staphylococcus Aureus in Poultry. Asian Pac J Trop Biomed 2017, 7, 253–256. [Google Scholar] [CrossRef]
- Youssef, F.M.; Soliman, A.A.; Ibrahim, G.A.; Saleh, H.A. Advanced Bacteriological Studies on Bumblefoot Infections in Broiler Chicken with Some Clinicopathological Alteration. Veterinary Science and research 2019, 1. [Google Scholar]
- Van Duijkeren, E.; Jansen, M.D.; Flemming, S.C.; De Neeling, H.; Wagenaar, J.A.; Schoormans, A.H.W.; Van Nes, A.; Fluit, A.C. Methicillin-Resistant Staphylococcus Aureus in Pigs with Exudative Epidermitis. Emerg Infect Dis 2007, 13. [Google Scholar] [CrossRef] [PubMed]
- Corpa, J.M.; Hermans, K.; Haesebrouck, F. Main Pathologies Associated with Staphylococcus Aureus Infections in Rabbits: A Review. World Rabbit Science 2010, 17. [Google Scholar] [CrossRef]
- Viana, D.; Selva, L.; Callanan, J.J.; Guerrero, I.; Ferrian, S.; Corpa, J.M. Strains of Staphylococcus Aureus and Pathology Associated with Chronic Suppurative Mastitis in Rabbits. Veterinary Journal 2011, 190. [Google Scholar] [CrossRef] [PubMed]
- Kadariya, J.; Smith, T.C.; Thapaliya, D. Staphylococcus Aureus and Staphylococcal Food-Borne Disease: An Ongoing Challenge in Public Health. Biomed Res Int 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States-Major Pathogens. Emerg Infect Dis 2011, 17. [Google Scholar] [CrossRef] [PubMed]
- The European Union One Health 2021 Zoonoses Report. EFSA Journal 2022, 20. [CrossRef]
- Zaghen, F.; Sora, V.M.; Meroni, G.; Laterza, G.; Martino, P.A.; Soggiu, A.; Bonizzi, L.; Zecconi, A. Epidemiology of Antimicrobial Resistance Genes in Staphyloccocus Aureus Isolates from a Public Database in a One Health Perspective—Sample Characteristics and Isolates’ Sources. Antibiotics 2023, 12, 1225. [Google Scholar] [CrossRef]
- Sung, J.M.L.; Lloyd, D.H.; Lindsay, J.A. Staphylococcus Aureus Host Specificity: Comparative Genomics of Human versus Animal Isolates by Multi-Strain Microarray. Microbiology (N Y) 2008, 154. [Google Scholar] [CrossRef] [PubMed]
- Shepheard, M.A.; Fleming, V.M.; Connor, T.R.; Corander, J.; Feil, E.J.; Fraser, C.; Hanage, W.P. Historical Zoonoses and Other Changes in Host Tropism of Staphylococcus Aureus, Identified by Phylogenetic Analysis of a Population Dataset. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Bergeron, G.; Bourassa, M.W.; Dickson, J.; Gomes, F.; Howe, A.; Kahn, L.H.; Morley, P.S.; Scott, H.M.; Simjee, S.; et al. Complexities in Understanding Antimicrobial Resistance across Domesticated Animal, Human, and Environmental Systems. Ann N Y Acad Sci 2019, 1441. [Google Scholar] [CrossRef] [PubMed]
- Pennone, V.; Cobo-Díaz, J.F.; Prieto-Maradona, M.; Álvarez-Ordóñez, A. Integration of Genomics in Surveillance and Risk Assessment for Outbreak Investigation. EFSA Journal 2022, 20. [Google Scholar] [CrossRef] [PubMed]
- Meroni, G.; Sora, V.M.; Martino, P.A.; Sbernini, A.; Laterza, G.; Zaghen, F.; Soggiu, A.; Zecconi, A. Epidemiology of Antimicrobial Resistance Genes in Streptococcus Agalactiae Sequences from a Public Database in a One Health Perspective. Antibiotics 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Piccinini, R.; Borromeo, V.; Zecconi, A. Relationship between S. Aureus Gene Pattern and Dairy Herd Mastitis Prevalence. Vet Microbiol 2010, 145. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Wegener, H.C.; Jensen, N.E.; Jonsson, O.; Myllys, V.; Thorberg, B.M.; Waage, S.; Rosdahl, V.T. A Study of Phage- and Ribotype Patterns of Staphylococcus Aureus Isolated from Bovine Mastitis in the Nordic Countries. Acta Vet Scand 1997, 38. [Google Scholar] [CrossRef]
- Larsen, H.D.; Aarestrup, F.M.; Jensen, N.E. Geographical Variation in the Presence of Genes Encoding Superantigenic Exotoxins and β-Hemolysin among Staphylococcus Aureus Isolated from Bovine Mastitis in Europe and USA. Vet Microbiol 2002, 85. [Google Scholar] [CrossRef] [PubMed]
- Coppens, J.; Xavier, B.B.; Vlaeminck, J.; Larsen, J.; Lammens, C.; Van Puyvelde, S.; Goossens, H.; Larsen, A.R.; Malhotra-Kumar, S. Genomic Analysis of Methicillin-Resistant Staphylococcus Aureus Clonal Complex 239 Isolated from Danish Patients with and without an International Travel History. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- De Backer, S.; Xavier, B.B.; Vanjari, L.; Coppens, J.; Lammens, C.; Vemu, L.; Carevic, B.; Hryniewicz, W.; Jorens, P.; Kumar-Singh, S.; et al. Remarkable Geographical Variations between India and Europe in Carriage of the Staphylococcal Surface Protein-Encoding SasX/SesI and in the Population Structure of Methicillin-Resistant Staphylococcus Aureus Belonging to Clonal Complex 8. Clinical Microbiology and Infection 2019, 25. [Google Scholar] [CrossRef] [PubMed]
- Fetsch, A.; Etter, D.; Johler, S. Livestock-Associated Meticillin-Resistant Staphylococcus Aureus—Current Situation and Impact From a One Health Perspective. Curr Clin Microbiol Rep 2021, 8. [Google Scholar] [CrossRef]
- Wu, Q.; Sabokroo, N.; Wang, Y.; Hashemian, M.; Karamollahi, S.; Kouhsari, E. Systematic Review and Meta-Analysis of the Epidemiology of Vancomycin-Resistance Staphylococcus Aureus Isolates. Antimicrob Resist Infect Control 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Tong, X.; Liu, S.; Wang, D.; Wang, L.; Fan, H. Prevalence of Methicillin-Resistant Staphylococcus Aureus in Healthy Chinese Population: A System Review and Meta-Analysis. PLoS One 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Sulis, G.; Sayood, S.; Gandra, S. Antimicrobial Resistance in Low- and Middle-Income Countries: Current Status and Future Directions. Expert Rev Anti Infect Ther 2022, 20. [Google Scholar] [CrossRef] [PubMed]
- Lawal, O.U.; Ayobami, O.; Abouelfetouh, A.; Mourabit, N.; Kaba, M.; Egyir, B.; Abdulgader, S.M.; Shittu, A.O. A 6-Year Update on the Diversity of Methicillin-Resistant Staphylococcus Aureus Clones in Africa: A Systematic Review. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Stefani, S.; Chung, D.R.; Lindsay, J.A.; Friedrich, A.W.; Kearns, A.M.; Westh, H.; MacKenzie, F.M. Meticillin-Resistant Staphylococcus Aureus (MRSA): Global Epidemiology and Harmonisation of Typing Methods. Int J Antimicrob Agents 2012, 39. [Google Scholar] [CrossRef] [PubMed]
- EU Veterinary Medicinal Product Database [Internet]. European Medicines Agency; 2019 [Cited 2023 Sep 5]. Available from: Http://Data.Europa.Eu/88u/Dataset/Eu-Veterinary-Medicinal-Product-Database.
- Tiseo, K.; Huber, L.; Gilbert, M.; Robinson, T.P.; Van Boeckel, T.P. Global Trends in Antimicrobial Use in Food Animals from 2017 to 2030. Antibiotics 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Everitt, B.S.; Landau, S.; Leese, M.; Stahl, D. Cluster Analysis 5th Ed.; 5th ed.; John Wiley & Sons 2011, 2011; ISBN 978-0-470-74991-3.
Figure 1.
Distribution of S. aureus isolates by the geographical regions. 1 Other Asia includes: Saudi Arabia, Bangladesh, Cambodia, United Arab Emirates, Jordan, Hong Kong, Kazakhstan, Kuwait, Lebanon, Nepal, Oman, Pakistan, Russia, Singapore, Syria, Sri Lanka, South Korea, Thailand, Taiwan, Turkey and Vietnam.
Figure 1.
Distribution of S. aureus isolates by the geographical regions. 1 Other Asia includes: Saudi Arabia, Bangladesh, Cambodia, United Arab Emirates, Jordan, Hong Kong, Kazakhstan, Kuwait, Lebanon, Nepal, Oman, Pakistan, Russia, Singapore, Syria, Sri Lanka, South Korea, Thailand, Taiwan, Turkey and Vietnam.
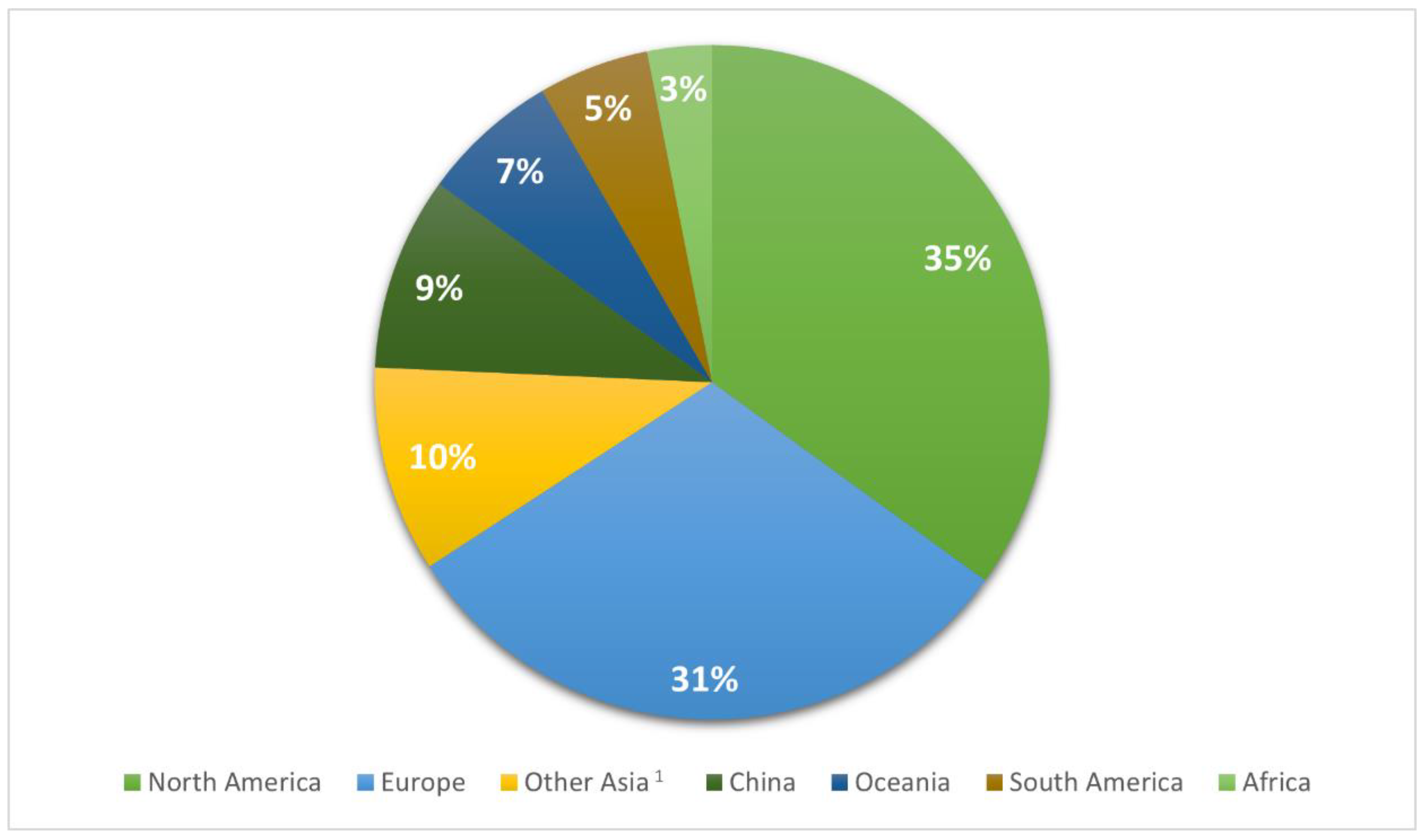
Figure 2.
Distribution of S. aureus isolates in Europe. 1 Other Europe includes: Austria, Belgium, Belarus, Croatia, Finland, Greece, Latvia, Lithuania, Luxembourg, Poland, Portugal, Czech Republic, Romania, Serbia, Slovenia and Hungary.
Figure 2.
Distribution of S. aureus isolates in Europe. 1 Other Europe includes: Austria, Belgium, Belarus, Croatia, Finland, Greece, Latvia, Lithuania, Luxembourg, Poland, Portugal, Czech Republic, Romania, Serbia, Slovenia and Hungary.
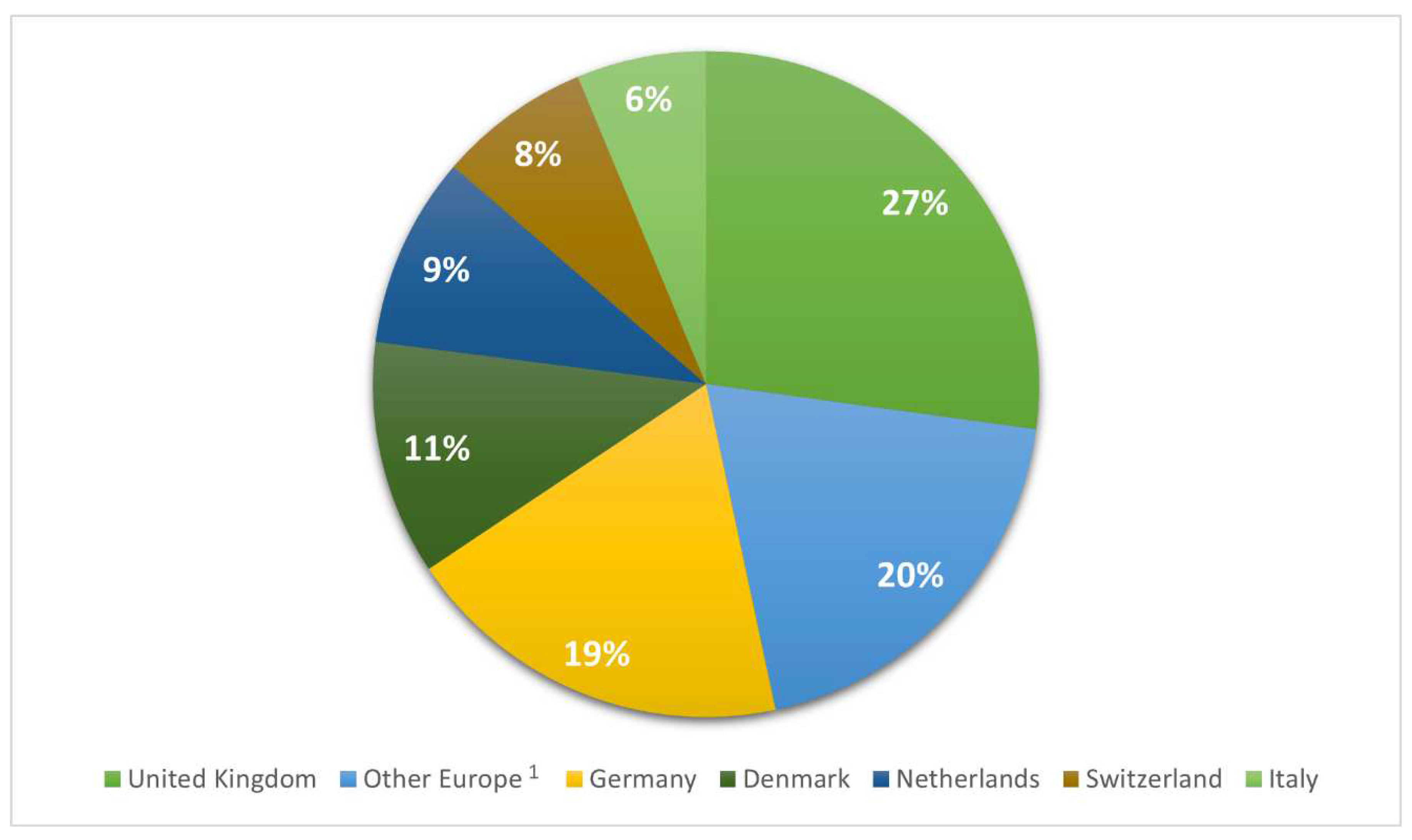
Figure 3.
Distribution of S. aureus isolates in USA. 1 Other States includes: Alabama, Alaska, Arizona, Arkansas, Colorado, Connecticut, Delaware, Florida, Georgia, Hawaii, Idaho, Illinois, Indiana, Kansas, Kentucky, Louisiana, Maine, Minnesota, Mississippi, Montana, Nebraska, Nevada, New Hampshire, New Jersey, New Mexico, North Carolina, North Dakota, Oklahoma, Oregon, Rhode Island, South Carolina, South Dakota, Tennessee, Texas, Utah, Vermont, Virginia, Washington, West Virginia, Wisconsin, Wyoming.
Figure 3.
Distribution of S. aureus isolates in USA. 1 Other States includes: Alabama, Alaska, Arizona, Arkansas, Colorado, Connecticut, Delaware, Florida, Georgia, Hawaii, Idaho, Illinois, Indiana, Kansas, Kentucky, Louisiana, Maine, Minnesota, Mississippi, Montana, Nebraska, Nevada, New Hampshire, New Jersey, New Mexico, North Carolina, North Dakota, Oklahoma, Oregon, Rhode Island, South Carolina, South Dakota, Tennessee, Texas, Utah, Vermont, Virginia, Washington, West Virginia, Wisconsin, Wyoming.
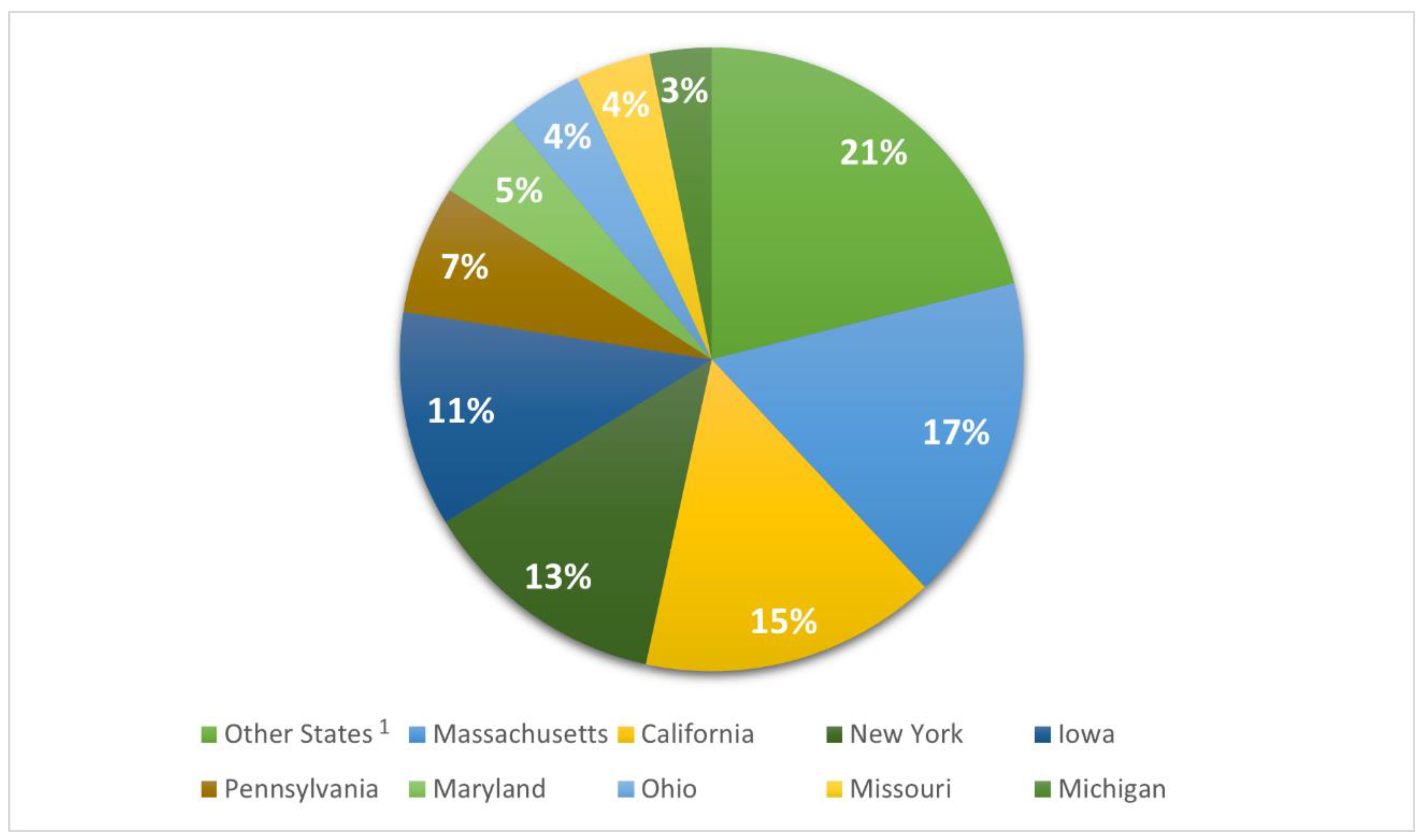
Figure 4.
Cluster distribution.
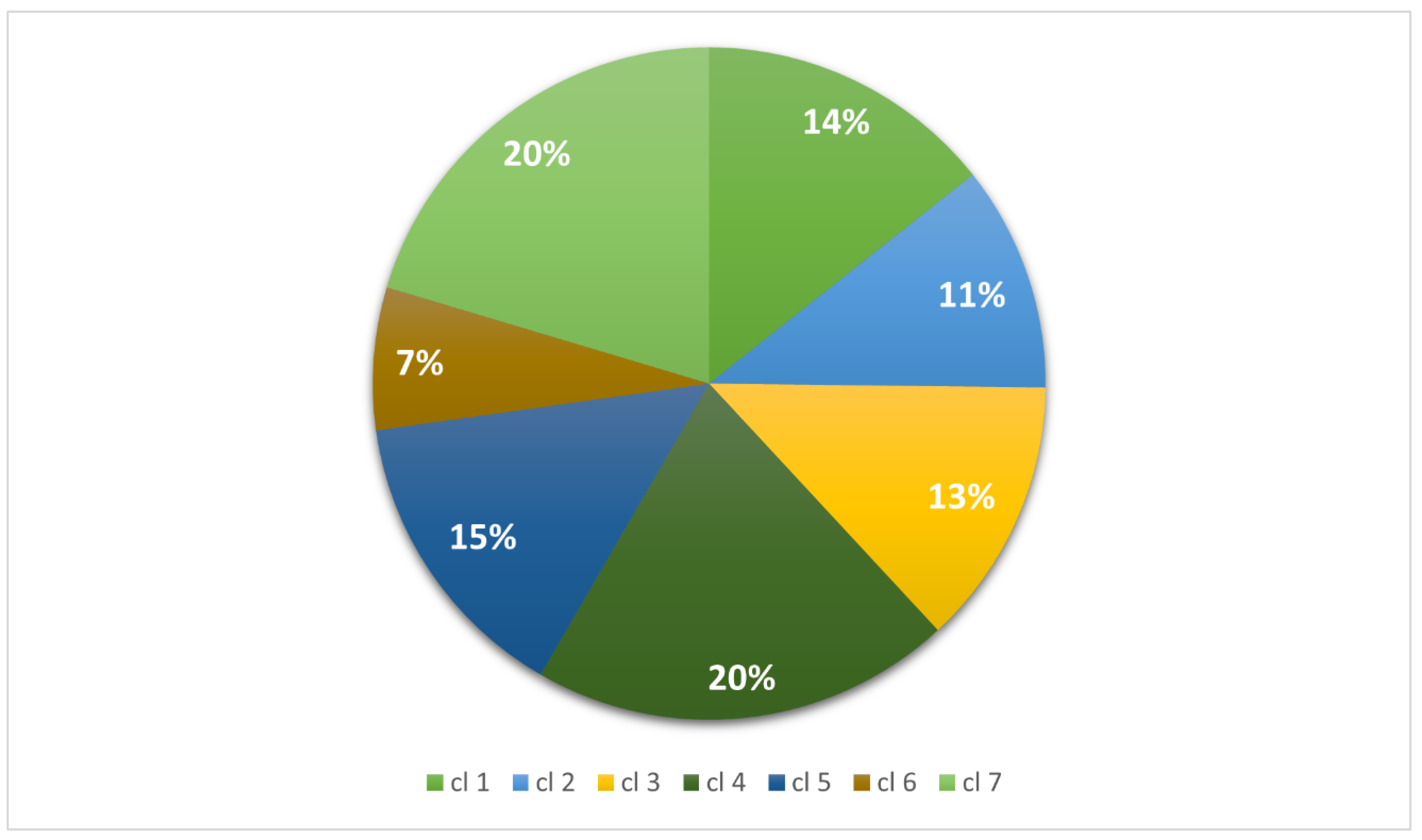
Figure 5.
Comparison between the distribution of clusters (a) and residues (difference between expected and observed frequency) (b) among European and USA S. aureus isolates.
Figure 5.
Comparison between the distribution of clusters (a) and residues (difference between expected and observed frequency) (b) among European and USA S. aureus isolates.
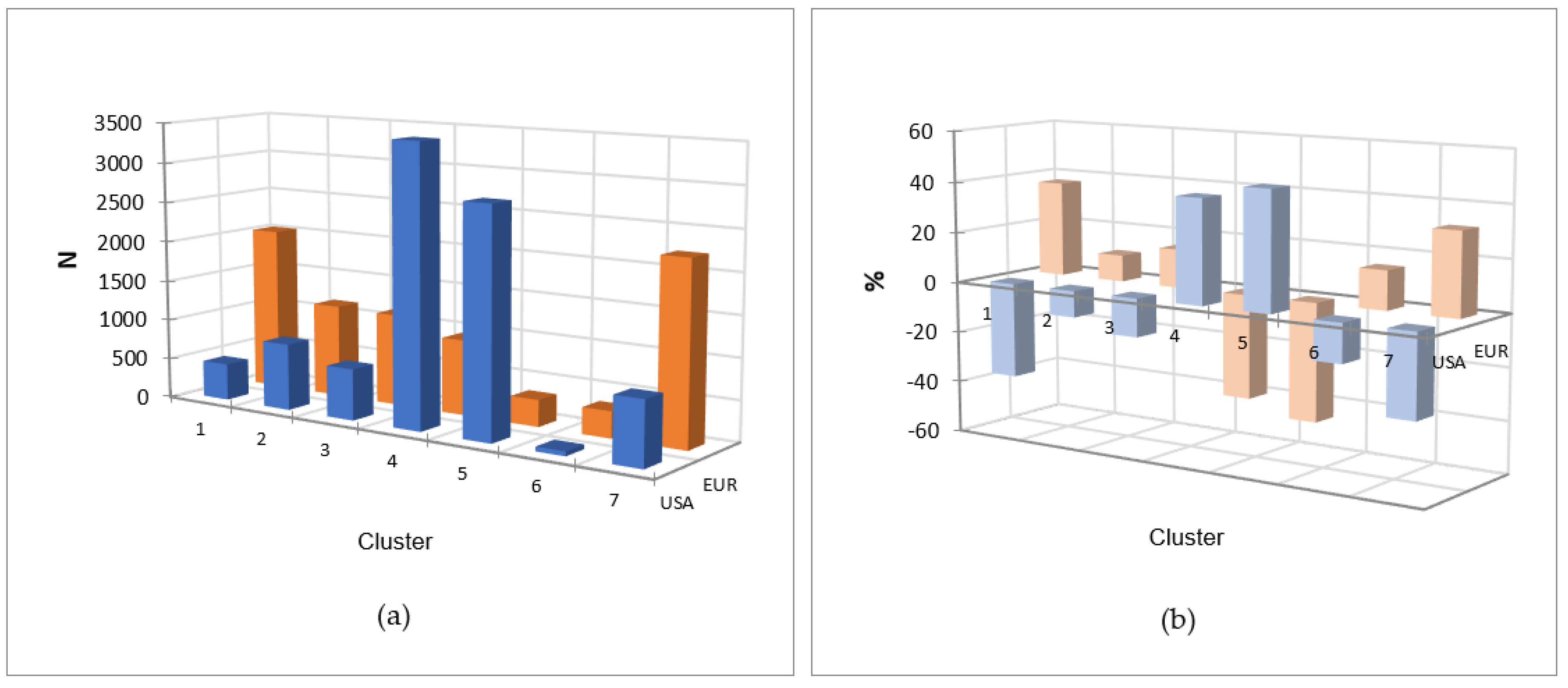
Table 1.
Geographical distribution and statistical differences of non-human associated, human associated and unknown isolates.
Table 1.
Geographical distribution and statistical differences of non-human associated, human associated and unknown isolates.
| Geographical Region | NHA1 | HUA | UNK | Total | |
|---|---|---|---|---|---|
| North America | N % |
595a 2 (6.4) |
7940b (77.7) |
1470c (15.9) |
9255 (100) |
| Europe | N % |
490a (6) |
3441b (42.3) |
4213c (51.7) |
8144 (100) |
| Other Asia3 | N % |
135a (5.2) |
1832b (70.3) |
639c (24.5) |
2606 (100) |
| China | N % |
685a (27.8) |
802b (32.6) |
974c (39.6) |
2461 (100) |
| Oceania | N % |
85a (4.9) |
1472b (84.6) |
183c (10.5) |
1740 (100) |
| South America | N % |
13a (0.9) |
1369b (98.2) |
12c (0.9) |
1394 (100) |
| Africa | N % |
56a (6.7) |
545a (65.7) |
229a (27.6) |
830 (100) |
1 NHA= non-human associated, HUA= human associated, UNK= unknown origin. 2 Values with different superscripts among lines statistically differ at χ2 test (α=0.05). 3 Other Asia includes: Saudi Arabia, Bangladesh, Cambodia, United Arab Emirates, Jordan, Hong Kong, Kazakhstan, Kuwait, Lebanon, Nepal, Oman, Pakistan, Russia, Singapore, Syria, Sri Lanka, South Korea, Thailand, Taiwan, Turkey and Vietnam.
Table 2.
European distribution of NHA, HUA and UNK isolates.
| State | NHA1 | HUA | UNK | Total | |
|---|---|---|---|---|---|
| United Kingdom | N % |
73a 2 (3.3) |
1440b (65.1) |
699a (31.6) |
2212 (100) |
| Other Europe3 | N % |
129a (8.1) |
593b (37.3) |
867c (54.6) |
1589 (100) |
| Germany | N % |
135a (8.7) |
586b (37.9) |
826c (53.4) |
1547 (100) |
| Denmark | N % |
3a (0.3) |
153b (16.5) |
722c (83.2) |
928 (100) |
| Netherlands | N % |
8a (1.1) |
282b (37.4) |
464c (61.5) |
754 (100) |
| Switzerland | N % |
81a (13.5) |
115b (19.2) |
403c (67.3) |
599 (100) |
| Italy | N % |
61a (11.8) |
272b (52.8) |
182c (35.3) |
515 (100) |
1 NHA= non-human associated, HUA= human associated, UNK= unknown origin. 2 Values with different superscripts among lines statistically differ at χ2 test or Fisher’s exact test (α=0.05). 3 Other Europe includes: Austria, Belgium, Belarus, Croatia, Finland, Greece, Latvia, Lithuania, Luxembourg, Poland, Portugal, Czech Republic, Romania, Serbia, Slovenia and Hungary.
Table 3.
USA distribution of NHA, HUA and UNK isolates.
| State | NHA1 | HUA | UNK | Total | |
|---|---|---|---|---|---|
| Other States2 | N % |
243a 3 (12.8) |
1153b (60.7) |
503c (26.5) |
1899 (100) |
| Massachusetts | N % |
0a (0.0) |
1239b (81) |
291c (19) |
1530 (100) |
| California | N % |
0a (0.0) |
829b (59.7) |
559c (40.3) |
1388 (100) |
| New York | N % |
74a (6.4) |
1068a (92.1) |
18b (1.6) |
1160 (100) |
| Iowa | N % |
0a (0.0) |
1003b (99.6) |
4a (0.4) |
1007 (100) |
| Pennsylvania | N % |
2a (0.3) |
593b (98.5) |
7a (1.2) |
602 (100) |
| Maryland | N % |
121a (28.2) |
282b (65.7) |
26c (6.1) |
429 (100) |
| Ohio | N % |
0a (0.0) |
344b (94.8) |
19c (5.2) |
363 (100) |
| Missouri | N % |
0a (0.0) |
352b (100) |
0a (0.0) |
352 (100) |
| Michigan | N % |
3a (1.1) |
270b (95.1) |
11a (3.9) |
284 (100) |
1 NHA= non-human associated, HUA= human associated, UNK= unknown origin. 2 Other States includes: Alabama, Alaska, Arizona, Arkansas, Colorado, Connecticut, Delaware, Florida, Georgia, Hawaii, Idaho, Illinois, Indiana, Kansas, Kentucky, Louisiana, Maine, Minnesota, Mississippi, Montana, Nebraska, Nevada, New Hampshire, New Jersey, New Mexico, North Carolina, North Dakota, Oklahoma, Oregon, Rhode Island, South Carolina, South Dakota, Tennessee, Texas, Utah, Vermont, Virginia, Washington, West Virginia, Wisconsin, Wyoming. 3 Values with different superscripts among lines statistically differ at χ2 test or Fisher’s exact test (α=0.05).
Table 4.
Geographical distribution of clusters and statistical difference among geographical regions within each cluster.
Table 4.
Geographical distribution of clusters and statistical difference among geographical regions within each cluster.
| Geographical Region | Cluster 1 N (%) |
Cluster 2 N (%) |
Cluster 3 N (%) |
Cluster 4 N (%) |
Cluster 5 N (%) |
Cluster 6 N (%) |
Cluster 7 N (%) |
|---|---|---|---|---|---|---|---|
| North America | 456e 1 (12.1) |
977c (34.1) |
651c (19.1) |
3471f (64.9) |
2802f (73.5) |
59f (3.3) |
839e (15.6) |
| Europe | 2034b (53.7) |
1153a (40.3) |
1138d (33.3) |
933a (17.5) |
331d (8.7) |
327d (18.1) |
2228a (41.4) |
| Other Asia2 | 229a (6) |
193b (6.7) |
225c (6.6) |
236b, d (4.4) |
255a (6.7) |
1019c (56.2) |
449d (8.3) |
| China | 622b (16.4) |
159b (5.5) |
468b (13.7) |
359e (6.7) |
28b (0.7) |
38b (2.1) |
787b (14.6) |
| Oceania | 335c (8.8) |
122b (4.3) |
401a (11.7) |
131c, d (2.4) |
43e (1.1) |
33b (1.8) |
675c (12.5) |
| South America | 38a (1) |
127b, c (4.4) |
344c (10.1) |
136a, b, c, d (2.5) |
289e (7.6) |
256e (14.2) |
204d (3.8) |
| Africa | 75a (2) |
136a (4.7) |
188a, b (5.5) |
84a, b, c, d (1.6) |
66a (1.7) |
77a (4.3) |
204a (3.8) |
| Total | 3789 (100) |
2867 (100) |
3415 (100) |
5350 (100) |
3814 (100) |
1809 (100) |
5386 (100) |
1 Values with different superscripts among rows statistically differ at χ2 test or Fisher’s exact test (α=0.05). 2 Other Asia includes: Saudi Arabia, Bangladesh, Cambodia, United Arab Emirates, Jordan, Hong Kong, Kazakhstan, Kuwait, Lebanon, Nepal, Oman, Pakistan, Russia, Singapore, Syria, Sri Lanka, South Korea, Thailand, Taiwan, Turkey and Vietnam.
Table 7.
Distribution and Statistical differences of HUA isolates among clusters in geographical regions.
Table 7.
Distribution and Statistical differences of HUA isolates among clusters in geographical regions.
| Geographical Region | Cluster 1 N (%) |
Cluster 2N (%) | Cluster 3N (%) | Cluster 4N (%) | Cluster 5N (%) | Cluster 6N (%) | Cluster 7N (%) |
|---|---|---|---|---|---|---|---|
| North America | 344a 1 (14.7) |
631a (39.6) |
508d (25.5) |
2824d (76) |
2222e (76.7) |
49b (3.3) |
612d (23.3) |
| Europe | 1165f (49.6) |
601d (37.7) |
481b (24.3) |
331a (8.9) |
157c (5.4) |
95d (6.4) |
611c (23.3) |
| Other Asia2 | 161c (6.8) |
43c (2.7) |
37c (1.9) |
187a, c (5) |
141a (4.9) |
989e (66.7) |
274c (10.4) |
| China | 328d (13.9) |
56a, b (3.5) |
89b (4.5) |
109c (2.9) |
5b (0.2) |
4b,c (0.3) |
211a (8) |
| Oceania | 288e (12.2) |
83b (5.2) |
386a (19.5) |
87b (2.4) |
42c (1.4) |
28c, d (1.9) |
558b (21.2) |
| South America | 37b (1.6) |
121a (7.6) |
340a (17.1) |
133a, c (3.6) |
287d (9.9) |
250f (16.9) |
201c (7.7) |
| Africa | 29a, b, c, d (1.2) |
59a (3.7) |
142a (7.2) |
45a, b, c (1.2) |
43a (1.5) |
67a (4.5) |
160a (6.1) |
| Total | 2352 (100) |
1594 (100) |
1983 (100) |
3716 (100) |
2897 (100) |
1482 (100) |
2627 (100) |
1 Values with different superscripts among rows statistically differ at χ2 test or Fisher’s exact test (α=0.05). 2 Other Asia include: Saudi Arabia, Bangladesh, Cambodia, United Arab Emirates, Jordan, Hong Kong, Kazakhstan, Kuwait, Lebanon, Nepal, Oman, Pakistan, Russia, Singapore, Syria, Sri Lanka, South Korea, Thailand, Taiwan, Turkey and Vietnam.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



