
Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
In Bangkok, the capital city of Thailand, air pollution is a significant problem, and efforts are needed to define organisms that may be used as bioindicators of air pollution. Epiphytic subaerial algae, which grow on the surface of trees, may have a potential as bioindicators of urban air quality. Algae were collected from randomly selected tree trunks in five parks in Bangkok, and the air pollutants CO, NO2, O3, SO2, PM2.5, and PM10 were measured. Analysis of the subaerial algal communities was performed by metagenomics. Diversity indexes were determined for the algal taxa detected, which were separated into groups corresponding to different collection sites by cluster analysis. Relationships between taxa and air pollutants were analyzed by PCA and the Pearson correlation coefficient (r). The results showed a high diversity of epiphytic subaerial al-gae. We recorded 101 taxa belonging to the Cyanophyta (70 taxa), Chlorophyta (21 taxa), Char-ophyta (5 taxa), Bacillariophyta (3 taxa), and Eustigmatophyta (2 taxa). The most abundant taxon was Chroococcidiopsis sp. 1, for which up to 13,254 individuals/cm2 were recorded. The Shan-non–Weaver index ranged between 1.37 and 2.51, the Margalef index between 3.84 and 4.75, and the Pielou index between 0.30 and 0.54. The similarity index was between 8.00% and 64.82% ac-cording to the cluster analysis results for three groups. The PCA indicated that all air pollutants affected the diversity and abundance of the epiphytic subaerial algae. Cyanothece sp. 2 is consid-ered a potential bioindicator of air pollution. It was negatively related to O3 and positively related to NO2 and CO
Keywords:
bioindicator
; air pollution
; epiphytic subaerial algae
; metagenomics
; Bangkok
1. Introduction
Epiphytic subaerial algae are a widespread subgroup of photosynthetic organisms living on aerial surfaces [1,2,3,4]. They are either single-celled or form colonies or filaments that, when sufficiently abundant to produce mats or crusts, are visible to the naked eye. Epiphytic subaerial algae differ in many aspects from algae that live in water or in conditions of high humidity. Many subaerial algae can tolerate drought and low atmospheric humidity, although they are metabolically active only when there is liquid water or a high atmospheric moisture [2,4,5,6]. Most subaerial algae are green algae (phyla Chlorophyta & Charophyta) and Cyanobacteria with a high tolerance to ultraviolet (UV) radiation and a reduced dependence on water [7,8,9]. Epiphytic subaerial algae grow on the surface of trunks, leaves, flowers, and fruits of vascular plants all over the world, but they are usually most abundant in humid tropical environments, from plains to highlands [1,2,3,10].
Epiphytic subaerial algae can grow on the surface of trees in many settings, from forests to city boulevards, although their diversity and abundance are influenced by the different climatic conditions of different regions [1,2,4] and specific conditions of local habitats. Species growing in urban environments can tolerate air pollution and can be used as biological indicators of air pollution in different areas and weather conditions [11,12,13]. Some studies have focused on the diversity and abundance of these organisms in urban areas with high levels of air pollutants due to emissions from automobiles, industrial plants, and households [14,15,16]. [16] and [17] argued that some of these organisms may be used as bioindicators of specific air pollutants such as sulfur dioxide (SO2), nitrogen oxides (NOx), carbon monoxide (CO), and fine particulate matter (PM2.5).
Bangkok is the capital city of Thailand, and it is governed by the Bangkok Metropolitan Administration. The city is located in the tropical region, with a population of more than 10 million people [18,19,20]. Today, it suffers from severe air pollution [20,21]. The pollution derives from emissions by cars, factories, and various types of waste [22], such as hydrocarbons, particulate matter (PM10), carbon monoxide (CO), sulfur dioxide (SO2), nitrogen oxides (NOx), benzene, and black smoke from diesel-powered vehicles [23,24,25]. Epiphytic subaerial algae that grow in green spaces such as parks and urban forests are exposed to high levels of such pollutants. It is therefore plausible that some species might be used as biological indicators of current climate conditions and pollution levels. Identifying species and assessing their distribution and possible correlation with amount and/or type of air pollutants may therefore guide researchers in recognizing tolerant species that can withstand city pollution. This information can then be used for further studies focusing on several aspects of the biology of these organisms.
The objectives of this study were to determine the diversity and abundance of epiphytic subaerial algae at several sites within the Metropolitan Region of Bangkok and identify species/taxa that might be used as indicators of air pollution in this area.
2. Materials and Methods
2.1. Study Sites
Extensive collections of epiphytic subaerial algae were made in May 2022 (the wet season, which in Bangkok spans from May to October) from five parks selected as sampling sites in the Bangkok Metropolitan Region (hereafter reported simply as Bangkok). They were located in the city’s northern area (Vachirabenjatas Park, or Rot Fai Park), eastern area (Suan Luang Rama IX), central area (Lumpini Park), western area (Thawi Wanarom Park), and southern area (Thonburirom Park). Distances among these sites ranged between 10 and 30 km (Figure 1 and Table 1).
2.2. Sample Collection
At each site, samples of epiphytic subaerial algae were collected from five trees with a circumference of more than 130 cm, at approximately 1.5 m height from the ground, by the method of line transect sampling by walk in the line of 5 trees in 4 corners and a center [26]. A sterile scraper or knife was used to scrape off a surface of 1 cm2, and each sample was placed in a paper bag containing silica gel. Information concerning orientation of the sampled surface, species of tree, and characteristics of the surrounding environment was annotated for each sample. The geographical coordinates and altitude were recorded based on GPS data (Table 1). The paper bags containing the samples were sealed and transferred to the laboratory at the Department of Botany, Kasetsart University.
2.3. Measurement of Air Pollutants
The general atmospheric air quality standards for six types of air pollutants were used in this study. For ozone (O3), nitrogen dioxide (NO2), carbon monoxide (CO), and sulfur dioxide (SO2), the measurements were recorded in units of parts per billion (ppb) or parts per million (ppm). For particulate matter less than 10 microns in size (PM10) or less than 2.5 microns in size (PM2.5) the measurements were recorded in units of micrograms per cubic meter (µg/m3). The measurements used were averages of the pollutant measurements taken in every hour over 24 hours in the algal collection dates. The measurements were taken in each site by the Air Quality and Noise Management Divisions of the Pollution Control Department and the Bureau of the Environment of the Bangkok Metropolitan Administration; the data are available from http://air4thai.pcd.go.th/webV2/.
2.4. Algal Identification
The species/taxa of epiphytic subaerial algae were identified using a metagenomic approach. The samples scraped from 5 trees at each site were mixed together and DNA was extracted using the OnePCR Kit (GeneDireX, USA) following the manufacturer’s recommendations. For the amplification of the 23S rDNA marker, the primers P23SrV_f1 (GGACAGAAAGACCCT) and P23SrV_r1 (TCAGCCTGTTATCCC) were used, following the amplification protocol of [27]. These primers amplify the chloroplast-encoded 23S rDNA marker from prokaryotic and eukaryotic algae [27,28]. PCR amplification was performed using the following cycling conditions: initial denaturation at 94˚C for 2 min, followed by 35 cycles of 94˚C for 20 s, 55˚C for 30 s, and 72˚C for 30 s, with a final extension of 72˚C for 10 min [27]. PCR products with the expected size and yield were selected after a check on 2% agarose gel electrophoresis. The same amount of PCR products from each sample was pooled, end-repaired, A-tailed, and further ligated with Illumina adapters. Libraries were sequenced on a paired-end Illumina platform to generate 450-bp paired-end raw reads. The library was checked with QubitTM 4.0 Fluorometer (Thermo Fisher Scientific, USA) following [29] and the real-time polymerase chain reaction (rtPCR) for quantification and a bioanalyzer (2100 Bioanalyzer Instrument, USA) for size distribution detection. Quantified libraries were pooled and sequenced on MGISEQ-2000 Sequencing Illumina platforms, according to effective library concentrations and the amount of data required.
2.4.1. Merging of Paired-End Reads and Quality Control
Paired-end reads were assigned to samples based on their unique barcodes and were truncated by cutting off the barcodes and primer sequences. Then, the paired-end reads were merged using FLASH (Version 1.2.11, http://ccb.jhu.edu/software/FLASH/) [30], a very fast and accurate analysis tool designed to merge paired-end reads when at least some of the reads overlap with the reads generated from the opposite end of the same DNA fragment; the splicing sequences are called raw tags. Quality filtering of the raw tags was performed using fastp (Version 0.20.0) software to obtain high-quality clean tags. Finally, the clean tags were compared with the reference database (SILVA database: https://www.arbsilva.de/ for 23S; UNITE database: https://unite.ut.ee/ for rDNA) using VSEARCH (Version 2.15.0) to detect the chimera sequences, and then the chimera sequences were removed to obtain the effective tags [31].
2.4.2. Amplicon Sequence Variants (ASVs) Denoising and Species Annotation
First, for the effective tags obtained as described above, denoising was performed with the DADA2 or deblurring module in QIIME2 software (Version QIIME2-202006) to obtain the initial ASVs (default: DADA2), and then ASVs with an abundance of less than 5 were filtered out [32]. Species annotation was performed using the QIIME2 software. For 23S, the annotation database was the SILVA database from LPSN and NCBI taxonomy [33], while for markers, primer, and protocols from [27], it was the UNITE database. To study the phylogenetic relationship of each ASV and the differences in dominant species in different samples, multiple sequencing alignment was performed using the QIIME2 software. Finally, the absolute abundance (individuals, meaning an individual cell, colony or filament of subaerial algae) of ASVs was normalized using a standard sequence number corresponding to the sample with the fewest sequences.
2.5. Algal Community Structure
The diversity index (Shannon–Weaver index (H')), richness index (Margalef index), equitability (evenness) index (Pielou index (J')), and similarity index or the percentage of overlap algae, were calculated from abundance data of epiphytic subaerial algae. They were used to compare the diversity and abundance of epiphytic subaerial algae at the various sites. Cluster analysis using the Bray–Curtis method was used to determine the similarities index for the various sites of collection of epiphytic subaerial algae by PC-ORDTM 6 program [34] and IBM SPSS Statistics version 21.0 (IBM Corporation, Armonk, New York, USA) [35].
2.6. Relationship Between Algal Diversity and Abundance and the Air Pollutants
The possible relationships between diversity and abundance of the epiphytic subaerial algae and the air pollutants were analyzed using a Principal Component Analysis (PCA) with a Bray‒Curtis distance in the PC-ORDTM 6 program [34]. The statistical values of the Pearson’s correlation coefficient (r) obtained with IBM SPSS Statistics version 21.0 (IBM Corporation, Armonk, New York, USA) also analyzed the relationships between the diversity and abundance of the epiphytic subaerial algae and the air pollutants [35,36].
3. Results
3.1. Diversity and Abundance of Algae
The overall number of taxa of epiphytic subaerial algae recorded in the urban area of Bangkok was 101. Overall, 101 taxa of algae belonging to the phyla Cyanophyta (70 taxa), Chlorophyta (21 taxa), Charophyta (5 taxa), Bacillariophyta (3 taxa) and Eustigmatophyta (2 taxa) were identified in the urban area of Bangkok from the algal samples (Figure 2; Table 2).
Concerning each site individually, 72 species were recovered in the western site (Thawi Wanarom Park), 55 in the eastern site (Suan Luang RAMA IX), 53 in the northern site (Vachirabenjatas Park), 49 in the southern site (Thonburirom Park) and 44 in the central site (Lumpini Park). Moreover, when comparing the abundance of the five phyla at each site, Cyanophyta had the highest diversity of taxa, followed by Chlorophyta. Charophyta, Bacillariophyta, and Eustigmatophyta were found in smaller numbers (Figure 3).
The overall abundance of epiphytic subaerial algae in Bangkok, expressed as number of individuals, was 17,031 individuals/cm2. Cyanophyta had the highest abundance, followed by Charophyta, Chlorophyta, Eustigmatophyta, and Bacillariophyta (Figure 4 and Table A1).
Concerning the most abundant taxa, Chroococcidiopsis sp. 1 had the highest abundance, followed by Microcoleus sp. 1, Klebsormidium flaccidum, Oscillatoria sp., Scytonema mirabile, Tolypothrix sp. 1, Cyanothece sp., Trebouxia corticola and Gloeocapsa sp. (Figure 5 and Table A1).
For the epiphytic subaerial algae collected at the five sampling sites in Bangkok, in terms of numbers of individuals Cyanophyta were the most abundant group at four sites. At the fifth (the northern site), Charophyta had a slightly higher number of individuals than Cyanophyta. At the eastern, western, and southern sites, Chlorophyta was the most abundant phylum after the Cyanophyta. At the central site, Chlorophyta and Charophyta were recorded, but in small abundances (see Figure 6).
Figure 7 shows the relative abundance of different subaerial epiphytes at the five study sites within Bangkok. At the northern site and southern site, the most abundant taxa were the same, i.e., Klebsormidium flaccidum, Microcoleus sp. 1, and Chroococcidiopsis sp. 1. At the eastern site, Chroococcidiopsis sp. 1, Trebouxia corticola and Cyanothece sp. 1 had the highest abundance. At the central site, the most abundant taxa were Tolypothrix sp. 1, Cyanothece sp. 1 and Microcoleus sp.1; at the western site, Oscillatoria sp., Scytonema mirabile, and Microcoleus sp. 1. From these results it can be seen that in general Microcoleus sp. 1 and Chroococcidiopsis sp. 1 were the taxa with the highest abundance at the 5 sites sampled in Bangkok.
3.2. Types of Air Pollutants
The air pollutants affecting the general atmospheric air quality at the time of sampling in Bangkok are presented in Figure 8. The amounts of ozone (O3) were at 16.34 to 23.84 ppb. Amounts of nitrogen dioxide (NO2) ranged between 6.67 to 24.18 ppb; carbon monoxide between (CO) 29 and 233 ppb (a large amount), and sulfur dioxide (SO2) between 1.34 and 2.41 ppb. Particulate matter less than 10 microns in size (PM10) was present at high levels, from 26.8 to 53.85 µg/m3. Finally, particulate matter less than 2.5 microns (PM2.5) was present at 10 to 15 µg/m3.
3.3. Index Values for Algal Communities
The diversity index (Shannon–Weaver index (H')), richness index (Margalef index), equitability, or evenness, index (Pielou index (J')), and similarity index were determined for the algal communities found at the sampling sites in Bangkok (Table 3 and Table 4). The diversity index ranged between 1.37 and 2.51, with the highest value for the southern site and the lowest value for the northern site. The western site had the highest richness index value (6.19), whereas the lowest was found for the central site (3.84). The equitability index values ranged between 0.30 and 0.54. Finally, the similarity index values ranged from 8.00% to 64.82%, with the highest similarity recorded between the central and southern sites, and the lowest similarity between the northern and eastern sites.
A cluster analysis showed that the composition of species and abundance of epiphytic subaerial algae separated the subaerial epiphytic communities into three groups of sampling sites (Figures 9), with 75% of the information remaining. For the Group 1, represented by the northern site, Klebsormidium flaccidum, Microcoleus sp. 1, and Chroococcidiopsis sp. 1 were the dominant taxa. In Group 2 (eastern site), Chroococcidiopsis sp. 1, Trebouxia corticola, and Cyanothece sp. were the dominant taxa. In Group 3 (which clustered the central, western, and southern sites), Microcoleus sp.1, and Tolypothrix sp. 1 were the dominant taxa.
3.4. Relationship between Diversity of Species and Air Pollutants
The PCA showed the influence of air pollutants on the presence of diversity and abundance of algal species (41.623% and 28.075% of variance explained for the first and second axis, respectively) (Figure 10). Only PM2.5 was a major contributor to the first PCA axis (r = −0.468, r = 0.187), which was directly positively correlated for the western site but negatively correlated for the southern and central sites. CO, NO2, and PM10 were the major contributors to the second PCA axis (r = 0.423, r = 0.856; r = 0.646, r = 0.673; and r = 0.527, r = 0.621, respectively), which was directly positively correlated for the northern site. SO2 also was a major contributor to the second PCA axis (r = 0.455, r = −0.634), which was directly positively correlated for the eastern site. Finally, O3 also was a major contributor to the second PCA axis (r = −0.514, r = −0.611), which was negatively correlated for the northern site.
The relationship between the diversity of epiphytic subaerial algae found in Bangkok and air pollutants showed abundance of subaerial species and many air pollutants (see Figure 10). The abundance of subaerial species was broadly classified into 3 groups. The first group, based on PM2.5, was the only major factor on the first PCA axis, meaning that areas with high PM2.5 pollution had high abundance and composition of most subaerial algal communities. The second group, CO, NO2 and PM10, contributed significantly to the second PCA axis and had the same abundance and composition of most subaerial algal communities as PM2.5, but a different species group. Whereas O3 was a similar major contributor to the second PCA axis as CO, NO2 and PM10, but in the opposite direction; that is, when the area had high O3, the abundance and composition of subaerial algal communities was low. Finally, the third group, following SO2, contributed significantly to the second PCA axis in most epiphytic subaerial algae, as did CO, N2, and PM10, but had an impact on several species’ groups.
The possible relationship between abundance of each subaerial algal taxon and chemical pollutants was tested using the Pearson’s correlation coefficient (r). Pearson’s correlation coefficient (r) showed that all air pollutants had a significant correlation (* = statistically significant difference (p ≤ .05), 0.01, ** = high statistically significant difference (p ≤ .01)) with the abundance of several epiphytic subaerial algae (Table A2). Cyanothece sp. 2 had a correlation many air pollutants: it was negatively correlated with O3 (−0.941**) and positively with NO2 (0.988**) and CO (0.914*) (Table A2). Leptolyngbya sp. 1, Nostoc carneum, Phormidium tinctorium, Synechococcus sp., Symbiochloris irregularis, Stichococcus sp., Chlamydomonas zebra, and Nannochloropsis oculata were positively correlated with CO (0.986**) and PM10 (0.964**). Significant correlations with these pollutants were also detected for Klebsormidium flaccidum (0.986**) (0.963**), Tolypothrix sp. 2 (0.925*) (0.999**), Nostoc sp. 1 (0.975*) (0.943*), Microcoleus sp. 1 (0.939*) (0.916*), Oscillatoria nigro-viridis (0.949*) (0.893*), Cylindrospermum sp. (0.949*) (0.994**), Diadesmis sp. (0.986**) (0.940*), and Parachlorella kessleri (0.957*) (0.980**).
Chroococcidiopsis thermalis and Watanabea reniformis were positively correlated with SO2 (0.880* and 0.957*, respectively). Scytonema hofmannii (0.964**) was positively correlated with CO. Calothrix sp. 4 (0.917*) was positively correlated with O3, whereas Chloromonas perforata (−0.962**) was negatively correlated with this pollutant. Symbiochloris reticulata (0.944*) was positively correlated with PM10. Finally, Acutodesmus sp. (−0.896*) was negatively correlated with PM2.5.
4. Discussion
The high diversity of epiphytic subaerial algae found in this study was not unexpected, due to the fact that Bangkok is located in a tropical region in Southeast Asia, with a humid climate characterized by high levels of rainfall, and therefore particularly suitable for growth of subaerial algae [13,16,38,39].
A noteworthy aspect of our study is the lack of sequences of Trentepohliales in our metagenomic sequencing results. The Trentepohliales is an order of subaerial green algae widespread in areas with humid climates, particularly in tropical regions [3,40,41]. These algae have been reported from urban habitats in the tropics [5,42,43], where they typically form bright red or orange patches on the colonized surfaces. At the time of sampling, growths with typical trentepohlialean habit were not visible on the tree trunks selected for this study. However, previous studies based on high-throughput sequencing carried out in the tropics recovered large numbers of sequences of Trentepohliales [10,44], revealing a great phylogenetic diversity of these algae in natural environments. Therefore, we expected to record some trentepohlialean sequences in our results. Further studies will be necessary to clarify if this reflects a real rarity of these chlorophytes in the urban area of Bangkok or is related to some technical problems for extraction of DNA from thalli of Trentepohliales. This result is even more striking considering that the sampling for this study was carried out in Bangkok’s wet season, which should be a favourable period for the growth of Trentepohliales. The high diversity of epiphytic subaerial algae recorded in this study was certainly related to this aspect. Obviously, the wet season’s high humidity and rainfall provide optimal conditions for the vegetative growth of subaerial algae [2]. Differences in microhabitat conditions related to microvariations in humidity may play a crucial role in determining the diversity and composition of tropical bark-growing algal assemblages [39].
Cyanobacteria (Cyanophyta) had the highest diversity and abundance among the epiphytic subaerial algae recorded in our study, followed by chlorophytan green algae and diatoms (Bacillariophyta). The success of cyanobacteria in subaerial environments is due to their adaptations to desiccation, favoured by the production of mucilaginous sheaths [2,45]. The tropics are well known to host a large diversity of Cyanobateria, which often grow in large amounts on many surfaces [46], especially artificial [3,47]. Chlorophyte green algae have a wide range of thallus and cellular organization compared to other groups of microalgae, and have developed several adaptations for survival and growth in subaerial habitats [3]. Diatoms are another algal group widely distributed in this type of environment [46,48,49]. Because they are limited by the availability of dissolved silica, their numbers are higher during periods of higher humidity and in habitats in which silica is more readily available [46,50,51,52], in a study similar to ours conducted in tropical South America and Hawaii, recorded environmental sequences inferred to be diatoms.
Chroococcidiopsis sp. 1, Microcoleus sp. 1, and Klebsormidium flaccidum were the most abundant subaerial algae found in Bangkok. Unicellular cyanobacteria belonging to the genus Chroococcidiopsis are extremely resistant to desiccation, UV irradiation, salt toxicity and high temperatures [53,54]. They require high light intensities for growth and are protected from high-energy UV light by pigments such as carotenoids and scytonemin [53,54]. Microcoleus is a well-known filamentous cyanobacterium that is resistant to desiccation thanks to several desiccation tolerance mechanisms, including accumulation of trehalose [55], stabilization of the photosynthetic apparatus [56] and accumulation of UV-protecting pigments [57]. A recent study reported the expression of genes involved in the oxidative and osmotic stress response, the desaturation of membrane lipids, and the production of EPS at the onset of desiccation [58]. Klebsormidium flaccidum was one of the most common subaerial algae recorded in this study. It is generally reported as one of the most widespread subaerial algae in the world, with cosmopolitan distribution according to morphology-based identifications [59,60,61]. The taxonomic circumscription of this species has been reassessed in recent years based on molecular data. In general, species of Klebsorbidium are known as fast-evolving organisms, which may rapidly develop physiological adaptations enabling them to colonize new habitats, including some with extreme conditions [62,63].
The wet season in Bangkok has an AQI of 28, or green, which indicates that a “good quality of air” is present and that residents “can participate in outdoor activities and travel as usual.” The AQI is a universal index with five levels from 0 to 201 or more [64,65]. It is widely used in many countries, such as the United States, Australia, Singapore, Malaysia, and Thailand, and measures levels of O3, NO2, CO, SO2, PM10, and PM2.5 [64,65,66].
The diversity index in the wet season in Bangkok was 1.37 to 2.51 for taxa of epiphytic subaerial algae. The richness index ranged between 3.84 and 4.75, indicating that in the sites studied there were generally high abundances of epiphytic subaerial algae [67]. The equitability index had values ranging from 0.30 to 0.54, showing that there was a similar representation of taxa in terms of abundances. [68] stated that if the equitability index approaches 1, this indicates that the study site/area hosts taxa with generally similar abundances. The similarity index had values ranging from 8.00% to 64.82%; these values were consistent with the results of the cluster analysis.
Cluster analysis showed that the subaerial algal communities could be subdivided in three separate groups distributed in different sampling sites. Group 1 was represented by the northern site, Group 2 by the eastern site, and Group 3 by the central, western, and southern sites. Each group had different abundances of taxa and ecological index values (i.e., diversity index, richness index, and equitability index), in agreement with the similarity index results. In all groups cyanobacteria were the dominant group, whereas differences in abundance of the other algal groups were detected. In Group 1 Charophyta were well represented, and this group had the lowest value for the ecological index values. In Group 2 (eastern site) Chlorophyta were well represented; this group had a low-value ecological index. Group 3, which comprised the central, western, and southern sites, consisted largely of Cyanobacteria and had the highest ecological index values. The abundance of cyanobacteria in urban areas is well documented, with particular abundance in tropical regions [3,45].
A significant relationship between air pollutants and some epiphytic subaerial algae was detected, in particular PM10. Cyanothece sp. 2 corresponds to Cyanothece sp. ATCC 51142 in the genomic database from NCBI (National Center for Biotechnology Information of USA) [69,70,71,72]. In this study it showed a negative correlation with O3 and a positive correlation with NO2 and CO. Species of Cyanothece are widely distributed in various environments worldwide, usually at a pH lower than 7 [73]. Typically, this genus is associated with water in benthic marine environments, rice fields, acidic marshes, peaty bogs, intertidal zones, moors, and clear lakes, but it is sometimes found in mountain soils [69,74], and records of it are also available from rocks and trees. The taxa Leptolyngbya sp. 1, Nostoc carneum, Phormidium tinctorium, Synechococcus sp., Symbiochloris irregularis, Stichococcus sp., Chlamydomonas zebra, Nannochloropsis oculata, and Klebsormidium flaccidum were also positively correlated with CO and PM10 and were found only at some sites. These correlations indicate their potential value as bioindicators of these pollutants, which should be further explored in future studies.
5. Conclusions
This study unraveled the presence of 101 taxa of epiphytic subaerial algae on tree barks in Bangkok, consisting mainly of Cyanophyta. In terms of abundance, the three most abundant taxa were Chroococcidiopsis sp. 1, Microcoleus sp. 1 and Klebsormidium flaccidum. All environmental pollutants had correlations with the abundance of epiphytic subaerial algae. Cyanothece sp. 2 was the taxon that showed the strongest correlation and has therefore the best potential to be used as bioindicator, being negatively correlated to O3 and positively correlated to NO2 and CO.
Author Contributions
Conceptualization, S.S. and N.S.; Methodology, S.S., N.S., and F.R.; Software, S.S.; Validation, S.S., and N.S.; Formal Analysis, S.S.; Investigation, S.S.; Resources, S.S.; Data Curation, S.S.; Writing—Original Draft Preparation, S.S.; Writing—Review and Editing, S.S., N.S., and F.R.; Visualization, S.S. and N.S.; Supervision, N.S.; Project Administration, N.S.; Funding Acquisition, S.S. and N.S.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author, Nuttha Sanevas. E-mail: fscintsv@ku.ac.th.
Acknowledgments
This work was financially supported by the Office of the Ministry of Higher Education, Science, Research and Innovation and the Thailand Science Research and Innovation through the Kasetsart University Reinventing University Program 2021.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

Table A1.
The abundances of species (individuals/cm2) of epiphytic subaerial algae found during the wet season in Bangkok at five sites.
Table A1.
The abundances of species (individuals/cm2) of epiphytic subaerial algae found during the wet season in Bangkok at five sites.
| Northern Site | Eastern Site | Central Site | Western Site | Southern Site | |
|---|---|---|---|---|---|
| Oscillatoria sp. | 163 | 9 | 22 | 7911 | 102 |
| Scytonema mirabile | 1106 | 265 | 255 | 5232 | 656 |
| Microcoleus sp. 1 | 7941 | 1454 | 2922 | 1144 | 1004 |
| Chroococcidiopsis sp. 1 | 1302 | 12391 | 602 | 962 | 1013 |
| Gloeocapsopsis sp. | 7 | 28 | 8 | 900 | 19 |
| Stanieria sp. | 16 | 7 | 18 | 793 | 14 |
| Tolypothrix sp. 1 | 61 | 140 | 6186 | 242 | 301 |
| Microchaete diplosiphon | 0 | 0 | 0 | 199 | 3 |
| Scytonema sp. 1 | 0 | 164 | 22 | 160 | 444 |
| Tolypothrix sp. 2 | 506 | 244 | 140 | 152 | 165 |
| Calothrix sp. 3 | 36 | 0 | 0 | 131 | 1 |
| Pleurocapsa minor | 90 | 22 | 20 | 122 | 32 |
| Cyanothece sp. 1 | 4 | 2057 | 3543 | 92 | 291 |
| Hyella patelloides | 0 | 0 | 0 | 78 | 2 |
| Calothrix sp. 2 | 64 | 6 | 18 | 40 | 156 |
| Nostoc piscinale | 0 | 1 | 1 | 39 | 0 |
| Chondrocystis sp. | 2 | 0 | 0 | 37 | 0 |
| Chroococcidiopsis thermalis | 15 | 98 | 10 | 33 | 52 |
| Brasilonema sp. | 0 | 2 | 0 | 17 | 2 |
| Calothrix sp. 1 | 0 | 396 | 6 | 17 | 6 |
| Cyanothece sp. 3 | 6 | 3 | 0 | 13 | 5 |
| Gloeocapsa sp. | 164 | 1118 | 282 | 13 | 95 |
| Microcoleus sp. 3 | 1 | 3 | 0 | 12 | 0 |
| Lusitaniella coriacea | 0 | 0 | 0 | 12 | 198 |
| Cyanothece sp. 4 | 0 | 3 | 0 | 11 | 0 |
| Nostoc linckia | 1 | 76 | 1 | 11 | 0 |
| Calothrix brevissima | 0 | 2 | 3 | 10 | 0 |
| Cyanothece sp. 5 | 2 | 0 | 0 | 10 | 0 |
| Nostoc sp. 1 | 61 | 1 | 1 | 8 | 0 |
| Scytonema hofmannii | 33 | 0 | 11 | 8 | 12 |
| Oscillatoria nigro-viridis | 28 | 0 | 0 | 8 | 0 |
| Nostoc sp. 2 | 0 | 0 | 0 | 6 | 0 |
| Cylindrospermum stagnale | 0 | 551 | 13 | 6 | 0 |
| Scytonema crispum | 0 | 0 | 7 | 5 | 17 |
| Gloeobacter kilaueensis | 0 | 0 | 0 | 5 | 0 |
| Coleofasciculus chthonoplastes | 1 | 0 | 0 | 5 | 2 |
| Oculatella neakameniensis | 0 | 0 | 0 | 4 | 0 |
| Leptolyngbya sp. 2 | 0 | 0 | 0 | 4 | 1 |
| Leptolyngbya sp. 3 | 0 | 0 | 0 | 4 | 0 |
| Scytonema sp. 2 | 0 | 7 | 0 | 3 | 0 |
| Lyngbya aestuarii | 0 | 0 | 0 | 3 | 0 |
| Calothrix sp. 4 | 0 | 3 | 5 | 3 | 0 |
| Synechococcus lividus | 0 | 0 | 0 | 2 | 4 |
| Oscillatoria acuminata | 0 | 0 | 0 | 2 | 0 |
| Trichormus azollae | 0 | 0 | 0 | 1 | 0 |
| Nodosilinea sp. | 0 | 0 | 0 | 1 | 0 |
| Thermosynechococcus sp. | 0 | 0 | 0 | 1 | 0 |
| Crinalium epipsammum | 0 | 0 | 0 | 1 | 0 |
| Sphaerospermopsis kisseleviana | 0 | 0 | 0 | 1 | 0 |
| Fischerella muscicola | 0 | 19 | 1 | 1 | 0 |
| Pleurocapsa sp. | 1 | 0 | 0 | 1 | 0 |
| Dolichospermum compactum | 2 | 0 | 0 | 0 | 72 |
| Stanieria cyanosphaera | 2 | 0 | 0 | 0 | 23 |
| Cyanothece sp. 2 | 40 | 4 | 0 | 0 | 22 |
| Nostoc punctiforme | 1 | 211 | 2 | 0 | 15 |
| Cylindrospermum muscicola | 0 | 0 | 0 | 0 | 1 |
| Calothrix sp. 6 | 0 | 0 | 0 | 0 | 1 |
| Anabaena sp. 1 | 0 | 0 | 25 | 0 | 1 |
| Calothrix sp. 5 | 0 | 0 | 5 | 0 | 0 |
| Jaaginema litorale | 9 | 0 | 4 | 0 | 0 |
| Cylindrospermum sp. | 75 | 13 | 0 | 0 | 0 |
| Cyanothece sp. 6 | 0 | 6 | 0 | 0 | 0 |
| Camptylonemopsis sp. | 0 | 3 | 0 | 0 | 0 |
| Gloeocapsopsis crepidinum | 0 | 2 | 0 | 0 | 0 |
| Chroococcidiopsis sp. 2 | 0 | 2 | 0 | 0 | 0 |
| Cyanobium gracile | 0 | 1 | 0 | 0 | 0 |
| Leptolyngbya sp. 1 | 5 | 0 | 0 | 0 | 0 |
| Nostoc carneum | 3 | 0 | 0 | 0 | 0 |
| Phormidium tinctorium | 2 | 0 | 0 | 0 | 0 |
| Synechococcus sp. | 2 | 0 | 0 | 0 | 0 |
| Diadesmis sp. | 49 | 3 | 8 | 5 | 5 |
| Nitzschia sp. | 0 | 0 | 0 | 3 | 0 |
| Cylindrotheca closterium | 0 | 0 | 0 | 1 | 0 |
| Watanabea reniformis | 21 | 97 | 17 | 16 | 53 |
| Ignatius tetrasporus | 37 | 20 | 5 | 20 | 27 |
| Nannochloris normandinae | 17 | 135 | 36 | 47 | 13 |
| Scenedesmus | 4 | 104 | 24 | 236 | 13 |
| Trebouxia corticola | 5 | 2090 | 38 | 6 | 5 |
| Trebouxia australis | 0 | 9 | 0 | 14 | 2 |
| Chloromonas perforata | 1 | 0 | 0 | 0 | 2 |
| Pyramimonas disomata | 3 | 13 | 0 | 232 | 2 |
| Xylochloris irregularis | 0 | 1 | 0 | 6 | 1 |
| Lobosphaera incisa | 3 | 13 | 0 | 18 | 1 |
| Edaphochlorella mirabilis | 0 | 4 | 2 | 2 | 1 |
| Dilabifilum sp. | 0 | 0 | 0 | 46 | 0 |
| Heterochlorella | 0 | 0 | 4 | 5 | 0 |
| Friedmannia sp. | 0 | 0 | 0 | 1 | 0 |
| Symbiochloris reticulata | 12 | 7 | 0 | 1 | 0 |
| Acutodesmus sp. | 0 | 2 | 5 | 0 | 0 |
| Parachlorella kessleri | 46 | 6 | 4 | 0 | 0 |
| Trebouxia decolorans | 0 | 1 | 0 | 0 | 0 |
| Symbiochloris irregularis | 20 | 0 | 0 | 0 | 0 |
| Stichococcus sp. | 4 | 0 | 0 | 0 | 0 |
| Chlamydomonas zebra | 2 | 0 | 0 | 0 | 0 |
| Monodopsis sp. 2 | 22 | 16 | 14 | 0 | 28 |
| Nannochloropsis oculata | 29 | 0 | 0 | 0 | 0 |
| Chlorokybus atmophyticus | 0 | 0 | 0 | 1 | 21 |
| Klebsormidium sp. 2 | 12 | 3 | 35 | 11 | 6 |
| Klebsormidium sp. 1 | 0 | 19 | 13 | 83 | 1 |
| Klebsormidium flaccidum | 12763 | 10 | 91 | 22 | 0 |
| Klebsormidium sp. 3 | 0 | 20 | 23 | 0 | 0 |
Table A2.
Pearson’s correlation coefficient (r) of CO, NO2, O3, PM2.5, PM10, and SO2 found during the wet season in Bangkok (p ≤ .05, 0.01, n = 30 and * = statistically significant difference (p ≤ .05), 0.01, ** = high statistically significant difference (p ≤ .01)).
Table A2.
Pearson’s correlation coefficient (r) of CO, NO2, O3, PM2.5, PM10, and SO2 found during the wet season in Bangkok (p ≤ .05, 0.01, n = 30 and * = statistically significant difference (p ≤ .05), 0.01, ** = high statistically significant difference (p ≤ .01)).
| CO | NO2 | O3 | PM2.5 | PM10 | SO2 | |
|---|---|---|---|---|---|---|
| Oscillatoria sp. | ‒0.236 | –0.509 | 0.400 | 0.498 | –0.358 | –0.547 |
| Scytonema mirabile | ‒0.097 | –0.378 | 0.274 | 0.549 | –0.239 | –0.580 |
| Microcoleus sp. 1 | 0.939* | 0.730 | –0.510 | –0.307 | 0.916* | –0.354 |
| Chroococcidiopsis sp. 1 | ‒0.327 | –0.276 | 0.294 | –0.026 | 0.046 | 0.784 |
| Gloeocapsopsis sp. | ‒0.264 | –0.532 | 0.420 | 0.500 | –0.377 | –0.524 |
| Stanieria sp. | ‒0.247 | –0.521 | 0.413 | 0.487 | –0.370 | –0.551 |
| Tolypothrix sp. 1 | ‒0.287 | –0.334 | 0.475 | –0.857 | –0.368 | –0.369 |
| Microchaete diplosiphon | ‒0.254 | –0.521 | 0.409 | 0.499 | –0.375 | –0.538 |
| Scytonema sp. 1 | ‒0.386 | 0.034 | –0.317 | 0.678 | –0.460 | 0.562 |
| Tolypothrix sp. 2 | 0.925* | 0.773 | –0.610 | –0.022 | 0.999** | –0.048 |
| Calothrix sp. 3 | 0.017 | –0.312 | 0.244 | 0.498 | –0.114 | –0.633 |
| Pleurocapsa minor | 0.399 | 0.061 | –0.081 | 0.505 | 0.258 | –0.648 |
| Cyanothece sp. 1 | ‒0.499 | –0.495 | 0.638 | –0.866 | –0.371 | 0.082 |
| Hyella patelloides | –0.255 | –0.520 | 0.406 | 0.504 | –0.377 | –0.536 |
| Calothrix sp. 2 | 0.222 | 0.583 | –0.781 | 0.572 | 0.000 | 0.209 |
| Nostoc piscinale | –0.277 | –0.555 | 0.450 | 0.469 | –0.387 | –0.536 |
| Chondrocystis sp. | –0.194 | –0.484 | 0.386 | 0.486 | –0.317 | –0.573 |
| Chroococcidiopsis thermalis | –0.470 | –0.263 | 0.145 | 0.329 | –0.175 | 0.880* |
| Brasilonema sp. | –0.320 | –0.553 | 0.419 | 0.557 | –0.413 | –0.432 |
| Calothrix sp. 1 | –0.388 | –0.340 | 0.353 | –0.041 | –0.017 | 0.734 |
| Cyanothece sp. 3 | 0.079 | –0.110 | –0.036 | 0.785 | –0.051 | –0.384 |
| Gloeocapsa sp. | –0.332 | –0.279 | 0.347 | –0.262 | 0.042 | 0.713 |
| Microcoleus sp. 3 | –0.262 | –0.555 | 0.459 | 0.502 | –0.305 | –0.427 |
| Lusitaniella coriacea | –0.122 | 0.324 | –0.561 | 0.527 | –0.288 | 0.414 |
| Cyanothece sp. 4 | –0.364 | –0.631 | 0.524 | 0.491 | –0.380 | –0.347 |
| Nostoc linckia | –0.402 | –0.390 | 0.398 | 0.004 | –0.035 | 0.682 |
| Calothrix brevissima | –0.445 | –0.750 | 0.691 | 0.205 | –0.522 | –0.564 |
| Cyanothece sp. 5 | –0.056 | –0.374 | 0.297 | 0.495 | –0.183 | –0.615 |
| Nostoc sp. 1 | 0.975* | 0.752 | –0.591 | –0.001 | 0.943* | –0.342 |
| Scytonema hofmannii | 0.964** | 0.841 | –0.709 | –0.059 | 0.805 | –0.444 |
| Oscillatoria nigro-viridis | 0.949* | 0.689 | –0.547 | 0.079 | 0.893* | –0.427 |
| Nostoc sp. 2 | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Cylindrospermum stagnale | –0.376 | –0.327 | 0.349 | –0.070 | –0.003 | 0.734 |
| Scytonema crispum | –0.329 | 0.044 | –0.259 | 0.296 | –0.577 | 0.111 |
| Gloeobacter kilaueensis | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Coleofasciculus chthonoplastes | –0.097 | –0.226 | 0.044 | 0.737 | –0.300 | –0.447 |
| Oculatella neakameniensis | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Leptolyngbya sp. 2 | –0.287 | –0.451 | 0.280 | 0.634 | –0.450 | –0.446 |
| Leptolyngbya sp. 3 | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Scytonema sp. 2 | –0.486 | –0.563 | 0.530 | 0.178 | –0.167 | 0.491 |
| Lyngbya aestuarii | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Calothrix sp. 4 | –0.585 | –0.790 | 0.917* | –0.705 | –0.503 | 0.281 |
| Synechococcus lividus | –0.226 | 0.103 | –0.384 | 0.728 | –0.441 | 0.185 |
| Oscillatoria acuminata | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Trichormus azollae | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Nodosilinea sp. | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Thermosynechococcus sp. | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Crinalium epipsammum | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Sphaerospermopsis kisseleviana | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Fischerella muscicola | –0.402 | –0.367 | 0.389 | –0.077 | –0.031 | 0.711 |
| Pleurocapsa sp. | 0.600 | 0.231 | –0.184 | 0.356 | 0.485 | –0.662 |
| Dolichospermum compactum | –0.079 | 0.375 | –0.600 | 0.493 | –0.238 | 0.436 |
| Stanieria cyanosphaera | –0.010 | 0.438 | –0.654 | 0.495 | –0.172 | 0.423 |
| Cyanothece sp. 2 | 0.914* | 0.988** | –0.941** | 0.210 | 0.842 | 0.045 |
| Nostoc punctiforme | –0.377 | –0.293 | 0.299 | –0.031 | –0.009 | 0.783 |
| Cylindrospermum muscicola | –0.106 | 0.350 | –0.578 | 0.491 | –0.263 | 0.440 |
| Calothrix sp. 6 | –0.106 | 0.350 | –0.578 | 0.491 | –0.263 | 0.440 |
| Anabaena sp. 1 | –0.272 | –0.310 | 0.453 | –0.861 | –0.353 | –0.358 |
| Calothrix sp. 5 | –0.265 | –0.321 | 0.472 | –0.873 | –0.339 | –0.372 |
| Jaaginema litorale | 0.875 | 0.668 | –0.430 | –0.456 | 0.818 | –0.442 |
| Cylindrospermum sp. | 0.949* | 0.774 | –0.599 | –0.066 | 0.994** | –0.139 |
| Cyanothece sp. 6 | –0.364 | –0.312 | 0.331 | –0.055 | 0.008 | 0.743 |
| Camptylonemopsis sp. | –0.364 | –0.312 | 0.331 | –0.055 | 0.008 | 0.743 |
| Gloeocapsopsis crepidinum | –0.364 | –0.312 | 0.331 | –0.055 | 0.008 | 0.743 |
| Chroococcidiopsis sp. 2 | –0.364 | –0.312 | 0.331 | –0.055 | 0.008 | 0.743 |
| Cyanobium gracile | –0.364 | –0.312 | 0.331 | –0.055 | 0.008 | 0.743 |
| Leptolyngbya sp. 1 | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Nostoc carneum | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Phormidium tinctorium | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Synechococcus sp. | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Diadesmis sp. | 0.986** | 0.802 | –0.627 | –0.107 | 0.940* | –0.332 |
| Nitzschia sp. | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Cylindrotheca closterium | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Watanabea reniformis | –0.371 | –0.112 | 0.035 | 0.164 | –0.063 | 0.957* |
| Ignatius tetrasporus | 0.774 | 0.801 | –0.835 | 0.589 | 0.761 | 0.142 |
| Nannochloris normandinae | –0.494 | –0.538 | 0.563 | –0.090 | –0.138 | 0.564 |
| Scenedesmus | –0.461 | –0.713 | 0.611 | 0.437 | –0.429 | –0.252 |
| Trebouxia corticola | –0.370 | –0.319 | 0.340 | –0.069 | 0.003 | 0.739 |
| Trebouxia australis | –0.500 | –0.676 | 0.547 | 0.528 | –0.404 | –0.010 |
| Chloromonas perforata | 0.547 | 0.857 | –0.962** | 0.424 | 0.386 | 0.236 |
| Pyramimonas disomata | –0.263 | –0.537 | 0.428 | 0.500 | –0.364 | –0.511 |
| Xylochloris irregularis | –0.396 | –0.572 | 0.406 | 0.632 | –0.460 | –0.285 |
| Lobosphaera incisa | –0.369 | –0.605 | 0.524 | 0.469 | –0.230 | –0.021 |
| Edaphochlorella mirabilis | –0.758 | –0.735 | 0.700 | –0.073 | –0.462 | 0.542 |
| Dilabifilum sp. | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Heterochlorella | –0.406 | –0.701 | 0.693 | –0.086 | –0.566 | –0.750 |
| Friedmannia sp. | –0.252 | –0.524 | 0.416 | 0.491 | –0.370 | –0.542 |
| Symbiochloris reticulata | 0.762 | 0.584 | –0.415 | –0.038 | 0.944* | 0.106 |
| Acutodesmus sp. | –0.452 | –0.481 | 0.641 | –0.896* | –0.333 | 0.015 |
| Parachlorella kessleri | 0.957* | 0.771 | –0.578 | –0.153 | 0.980** | –0.211 |
| Trebouxia decolorans | –0.364 | –0.312 | 0.331 | –0.055 | 0.008 | 0.743 |
| Symbiochloris irregularis | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Stichococcus sp. | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Chlamydomonas zebra | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Monodopsis sp. 2 | 0.382 | 0.763 | –0.775 | –0.034 | 0.384 | 0.560 |
| Nannochloropsis oculata | 0.986** | 0.807 | –0.641 | –0.055 | 0.964** | –0.268 |
| Chlorokybus atmophyticus | –0.119 | 0.329 | –0.564 | 0.520 | –0.284 | 0.419 |
| Klebsormidium sp. 2 | –0.085 | –0.230 | 0.392 | –0.802 | –0.232 | –0.618 |
| Klebsormidium sp. 1 | –0.405 | –0.689 | 0.597 | 0.381 | –0.459 | –0.461 |
| Klebsormidium flaccidum | 0.986** | 0.806 | –0.638 | –0.060 | 0.963** | –0.272 |
| Klebsormidium sp. 3 | –0.506 | –0.516 | 0.661 | –0.798 | –0.288 | 0.243 |
References
- John, D.M. Phylum Chlorophyta (green algae). In The freshwater algal flora of the British Isles; John, D.M., Whitton, B.A., Brook, A.J., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 287–612. [Google Scholar]
- López-Bautista, J.M.; Rindi, F.; Casamatta, D. The systematics of subaerial algae. In Algae and cyanobacteria in extreme environments; Seckbach, J., Ed.; Springer: Dordrecht, Netherlands, 2007; pp. 599–617. [Google Scholar]
- Rindi, F. Diversity, distribution and ecology of green algae and cyanobacteria in urban habitats. In Algae and Cyanobacteria in Extreme Environments; Seckbach, J., Ed.; Springer: Dordrecht, Netherlands, 2007; pp. 619–638. [Google Scholar]
- Graham, L.; Graham, J.; Wilcox, L. Algae, 2nd ed.; Benjamin Cummings (Pearson): San Francisco, USA, 2009. [Google Scholar]
- Ong, B.M.; Lim, M.; Wee, Y.C. Effects of desiccation and illumination of photosynthesis and pigmentation of an edaphic population of Trentepohlia odorata (Chlorophyta). J. Phycol. 1992, 28, 768–772. [Google Scholar] [CrossRef]
- Lüttge, U.; Büdel, B. Resurrection kinetics of photosynthesis in desiccation-tolerant terrestrial green algae (Chlorophyta) on tree bark. Plant Biol. 2010, 12, 437–44. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U.; Friedl, T.; Schumann, R.; Hoyer, K.; Lembcke, S. Mycosporine-like amino acids and phylogenies in green algae: Prasiola and its relatives from the Trebouxiophyceae (Chlorophyta). J. Phycol. 2005, 41, 557–566. [Google Scholar] [CrossRef]
- Karsten, U.; Karsten, U.; Lembcke, S.; Schumann, R. The effects of ultraviolet radiation on photosynthetic performance, growth, and sunscreen compounds in aeroterrestrial biofilm algae isolated from building facades. Planta 2007, 225, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U.; Schumann, R.; Mostaert, A.S. Aeroterrestrial algae growing on man-made surfaces: what are the secrets of their ecological success? In Algae and Cyanobacteria in Extreme Environments; Seckbach, J., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 583–597. [Google Scholar]
- Zhu, H.; Li, S.; Hu, Z.; Liu, G. Molecular characterization of eukaryotic algal communities in the tropical phyllosphere based on real-time sequencing of the 18S rDNA gene. BMC Plant Biol. 2018, 18, 365. [Google Scholar] [CrossRef] [PubMed]
- Haapala, H.; Goltsova, N.; Seppälä, H.S.; Kouki, J.; Lamppu, J.; Popovichev, B. Ecological condition of forests around the eastern part of the Gulf of Finland. Environ. Pollut. 1996, 91, 253–265. [Google Scholar] [CrossRef]
- Marmor, L.; Degtjarenko, P. Trentepohlia umbrina on Scots pine as bioindicator of alkaline dust pollution. Ecol. Indic. 2014, 45, 717–720. [Google Scholar] [CrossRef]
- Razli, S.A.; Azlam, A.; Ismail, A.; Murnira, O.; Ahmad, M.A.A.; Hafizah, B.N.; Kadaruddin, A.; Talib, L.M. Epiphytic microalgae as biological indicators for carbon monoxide concentrations in different areas of Peninsular Malaysia. Environ. Forensics 2022, 23, 314–323. [Google Scholar] [CrossRef]
- Roy-Ocotla, G.; Carrera, J. Aeroalgae: response to some aerobiological questions. Grana 1993, 32, 48–56. [Google Scholar] [CrossRef]
- Ra, H.S.Y.; Crang, F.E. Changes in thallus and algal cell components of two lichen species in response to low-level air pollution at Pacific Northeast Forests. Microsc. Microanal. 2002, 8, 1078–1079. [Google Scholar] [CrossRef]
- Ismail, A.; Wahab, N.A.; Latif, M.T.; Said, M.; Ismail, A.; Zulkifli, A.R.; Alwi, I.; Daud, D.; Sulaiman, F.N. Atmospheric air pollution and roughness of bark as possible factors in increasing density of epiphytic terrestrial algae. Asian J. Applied Sci. 2016, 4, 256–261. [Google Scholar]
- Hemond, H.F.; Fechner-Levy, E.J. Chemical Fate and Transport in The Environment; Academic Press: London, UK, 2000. [Google Scholar]
- Zhang, B.N.; Oanh, N.T. Photochemical smog pollution in the Bangkok Metropolitan Region of Thailand in relation to O3 precursor concentrations and meteorological conditions. Atmospheric Environment 2002, 36, 4211–4222. [Google Scholar] [CrossRef]
- Organization for Economic Cooperation and Development [OECD]. OECD Green Growth Studies: Green Growth in Bangkok, Thailand; OECD: Paris, France, 2015. [Google Scholar]
- Dhakal, S.; Shrestha, A. Bangkok, Thailand. In Cities on A Finite Planet: Towards Transformative Responses to Climate Change; Bartlett, S., Satterthwaite, D., Eds.; Routledge: London, UK, 2016; pp. 63–81. [Google Scholar]
- Klongvessa, P.; Chotpantarat, S. Statistical analysis of rainfall variations in the Bangkok urban area, Thailand. Arab. J. Geosci. 2015, 8, 4207–4219. [Google Scholar] [CrossRef]
- Bangkok Metropolitan Administration and Chulalongkorn University. Executive Summary 20-Year Development Plan for Bangkok Metropolis; Strategy and Evaluation Department Bangkok Metropolitan Administration and Faculty of Political Sciences, Chulalongkorn University: Bangkok, Thailand, 2011. [Google Scholar]
- Schwela, D.; Zali, O. Urban traffic pollution; E & FN Spon: London, UK, 1999. [Google Scholar]
- Muttamara, S.; Leong, S.T. Monitoring and assessment of exhaust emission in Bangkok Street air. Environ. Monit. Assess. 2000, 60, 163–180. [Google Scholar] [CrossRef]
- Afroz, R. Review of air pollution and health impacts in Malaysia. Environ. Res. 2003, 92, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Jorge, N.; Díaz-Gamboa, R.E. Chapter: 4. line transect sampling. In Introduction to Ecological Sampling; Manly, B., Navarro, J., Eds.; CRC Press: Boca Raton, Florida, 2015; pp. 47–62. [Google Scholar]
- Sherwood, A.R.; Presting, G.G. Universal primers amplify a 23S rDNA plastid marker in eukaryotic algae and Cyanobacteria. J. Phycol. 2007, 43, 605–608. [Google Scholar] [CrossRef]
- Sherwood, A.R.; Chan, Y.L.; Presting, G.G. Application of universally amplifying plastid primers to environmental sampling of a stream periphyton community. Mol. Ecol. Resour. 2008, 8, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Haendiges, J.; Timme, R.; Ramachandran, P.; Balkey, M. DNA Quantification Using the Qubit Fluorometer. Available online: https://www.protocols.io/view/dna-quantification-using-the-qubit-fluorometer-81wgbp3x3vpk/v1 5 (accessed on 1 July 2022).
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Gevers, D.; Earl, A.M. , Feldgarden, M.; Ward, D.V., Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Shao, D.; Zhou, J.; Gu, J.; Qin, J.; Chen, W.; Wei, W. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma. Chin. J. Cancer Res. 2020, 32, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Berney, C.; Ciuprina, A.; Bender, S.; Brodie, J.; Edgcomb, V.; Kim, E.; Rajan, J.; Parfrey, L.W.; Adl, S.; Audic, S.; et al. UniEuk: Time to speak a common language in protistology! J. Eukaryot. Microbiol. 2017, 64, 407–411. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Mefford, M.J. PC-ORD Multivariate Analysis of Ecological Data. Version 6; MjM Software, Gleneden Beach, Oregon. 2011.
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Plymouth Marine Laboratory: Plymouth, UK, 1994. [Google Scholar]
- McCune, B. Community Structure and Analysis Biology 570/670; Department of Botany and Plant Pathology, Oregon State University: Corvallis, Oregon, USA, 2007. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: https://www.algaebase.org (accessed on 10 September 2022).
- Neustupa, J.; Škaloud, P. Diversity of subaerial algae and cyanobacteria on tree bark in tropical mountain habitats. Biologia 2008, 63, 806–812. [Google Scholar] [CrossRef]
- Neustupa, J.; Škaloud, P. Diversity of subaerial algae and cyanobacteria growing on bark and wood in the lowland tropical forests of Singapore. Plant Ecol. Evol. 2010, 143, 51–62. [Google Scholar] [CrossRef]
- Škaloud, P.; Rindi, F.; Boedeker, C.; Leliaert, F. Chlorophyta: Ulvophyceae. In Süßwasserflora von Mitteleuropa – Freshwater Flora of Central Europe 13; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum: Berlin, Germany, 2018; pp. 1–289. [Google Scholar]
- Klimešová, M.; Rindi, F.; Skaloud, P. DNA cloning demonstrates high genetic heterogeneity in populations of the subaerial green alga Trentepohlia (Trentepohliales, Chlorophyta). J. Phycol. 2019, 55, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.K.M.N. Subaerial algae of Bangladesh. Bangladesh J. Bot. 1972, 1, 13–64. [Google Scholar]
- Ho, K.K.; Tan, K.H.; Wee, Y.C. Growth conditions of Trentepohlia odorata (Chlorophyta, Ulotrichales). Phycologia 1983, 22, 303–308. [Google Scholar] [CrossRef]
- Johnston, E.; Conklin, K.; Fredrick, P.; Sherwood, A. Pyrosequencing and culturing of Hawaiian corticolous biofilms demonstrate high diversity and confirm phylogenetic placement of the green alga Spongiochrysis hawaiiensis in Cladophorales (Ulvophyceae). Phycologia 2018, 57, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Nienow, J.A. Ecology of subaerial algae. Nova Hedwigia 1996, 112, 537–552. [Google Scholar]
- Nienow, J.A. Subaerial communities. In Encyclopedia of Environmental Microbiology; Bitton, G., Ed.; Wiley: New York, USA, 2002; pp. 3055–3065. [Google Scholar]
- Gorbushina, A.A. Life on the rocks. Environ. Microbiol. 2007, 9, 1613–1631. [Google Scholar] [CrossRef]
- Sharma, N.K.; Rai, A.K.; Singh, S.; Brown, J.R.M. Airborne algae: Their present status and relevance. J. Phycol. 2007, 43, 615–627. [Google Scholar] [CrossRef]
- Neustupa, J.; Štifterová, A. Distribution patterns of subaerial corticolous microalgae in two European regions. Plant Ecol. Evol. 2013, 146, 279–289. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Beyens, L.; Vincke, S.; Gremmen, N.J.M. Moss inhibiting diatom communities from Heard Island, sub-Antarctic. Polar Biol. 2004, 27, 532–543. [Google Scholar] [CrossRef]
- Taylor, J.C.; Levanets, A.A.; Blanco, S.; Ector, L. Microcostatus schoemanii sp. nov., M. cholnokyi sp. nov. and M. angloensis sp. nov.: three new terrestrial diatoms (Bacillariophyceae) from South Africa. Phycological Res. 2010, 58, 177–187. [Google Scholar] [CrossRef]
- Lam, D.W. Biodiversity and systematics of subaerial algae from the neotropics and Hawaii. Doctoral Degree, University of Alabama, Tuscaloosa, Alabama, 2010. [Google Scholar]
- Billi, D.; Wilmotte, A.; McKay, C.P. Desert strains of Chroococcidiopsis: a platform to investigate genetic diversity in extreme environments and explore survival potential beyond Earth. In EPSC Abstracts; European Planetary Science Congress: Rome, Italy, 2010. [Google Scholar]
- Bothe, H. The cyanobacterium Chroococcidiopsis and its potential for life on Mars. J. Astrobiol. Space Sci. Rev. 2019, 2, 398–412. [Google Scholar]
- Hershkovitz, N.; Oren, A.; Cohen, Y. Accumulation of trehalose and sucrose in cyanobacteria exposed to matric water stress. Appl. Environ. Microbiol. 1991, 57, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Harel, Y.; Ohad, I.; Kaplan, A. Activation of photosynthesis and resistance to photoinhibition in cyanobacteria within biological desert crust. Plant Physiol. 2004, 136, 3070–3079. [Google Scholar] [CrossRef]
- Quesada, A.; Vincent, W. F. Strategies of adaptation by Antarctic cyanobacteria to ultraviolet radiation. Eur. J. Phycol. 1997, 32, 335–342. [Google Scholar] [CrossRef]
- Rajeev, L.; da Rocha, U.N.; Klitgord, N.; Luning, E.G.; Fortney, J.; Axen, S.D.; Shih, P.M.; Bouskill, N.J.; Bowen, B.P.; et al. Dynamic cyanobacterial response to hydration and dehydration in a desert biological soil crust. ISME 2013, 7, 2178–2191. [Google Scholar] [CrossRef]
- Lokhorst, G.M. Comparative taxonomic studies on the genus Klebsormidium (Charophyceae) in Europe. In Cryptogamic Studies; Gustav Fischer Verlag: Stuttgart, Germany, 1996; Volume 5, pp. 1–132. [Google Scholar]
- Gaysina, L.A.; Purina, E.S.; Safiullina, L.M.; Bakieva, G.R. Resistance of Klebsormidium flaccidum (Kützing) Silva, Mattox & Blackwell (Streptophyta) to heavy metals. NBU J. Plant Sci. 2009, 3, 39–42. [Google Scholar]
- Gaysina, L.A.; Bohunická, M.; Hazuková, V.; Johansen, J.R. Biodiversity of terrestrial cyanobacteria of the South Ural Region. Cryptogam. Algol. 2018, 39, 167–198. [Google Scholar] [CrossRef]
- Škaloud, P.; Rindi, F. Ecological differentiation of cryptic species within an asexual protist morphospecies: a case study of filamentous green alga Klebsormidium (Streptophyta). J. Eukaryot. Microbiol. 2013, 60, 350–362. [Google Scholar] [CrossRef]
- Škaloud, P.; Lukešova, A.; Malavasi, V.; Ryšánek, D.; Hrčková, K.; Rindi, F. Molecular evidence for the polyphyletic origin of low pH adaptation in the genus Klebsormidium (Klebsormidiophyceae, Streptophyta). Pl. Ecol. Evol. 2014, 147, 333–345. [Google Scholar] [CrossRef]
- Kowalska, M.; Ośródka, L.; Klejnowski, K.; Zejda, J.; Krajny, E.; Wojtylak, M. Air quality index and its significance in environmental health risk communication. Arch. Environ. Prot. 2009, 35, 13–21. [Google Scholar]
- Environment Bureau Bangkok. AQI Information. Available online: https://bangkokairquality.com/bma/aqi?lang=en (accessed on 4 October 2022).
- Hossain, I.; Rahman, S.; Sattar, S.; Haque, M.; Mullick, A.; Siraj, S.; Sultana, N.; Uz-zaman, M.A.; Samima, I.; Haidar, A.; Khan, M. Environmental overview of air quality index (AQI) in Bangladesh: characteristics and challenges in present era. Int. J. Res. Eng. Technol. 2021, 4, 110–115. [Google Scholar]
- Magurran, A.E. 2004. Measuring Biological Diversity; Blackwell: Oxford, USA.
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification and Use as Bioindicators, 2nd ed.; Wiley–Blackwell: Hoboken, New Jersey, USA, 2010. [Google Scholar]
- Reddy, K.J.; Haskell, J.B.; Sherman, D.M.; Sherman, L.A. Unicellular, aerobic nitrogen-fixing cyanobacteria of the genus Cyanothece. J. Bacteriol. 1993, 175, 1284–1292. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, C.T. Origins of replication in Cyanothece 51142. Proc. Natl. Acad. Sci. 2008, 105, 125. [Google Scholar] [CrossRef]
- Welsh, E.A.; Liberton, M.; Stockel, J.; Loh, T.; Elvitigala, T.; Wang, C.; Wollam, A.; Fulton, R.S.; Clifton, S.W.; Jacobs, J.M.; et al. The genome of Cyanothece 51142, a unicellular diazotrophic cyanobacterium important in the marine nitrogen cycle. Proc. Natl. Acad. Sci. 2008, 105, 15094–15099. [Google Scholar] [CrossRef] [PubMed]
- Mareš, J.; Johansen, J.R.; Hauer, T.; Zima, J.Jr.; Ventura, S.; Cuzman, O.; Tiribilli, B.; Kaštovský, J. Taxonomic resolution of the genus Cyanothece (Chroococcales, Cyanobacteria), with a treatment on Gloeothece and three new genera, Crocosphaera, Rippkaea, and Zehria. J. Phycol. 2019, 55, 578–610. [Google Scholar] [CrossRef] [PubMed]
- Komarek, J.; Cepak, V. Cytomorphological characters supporting the taxonomic validity of Cyanothece (Cyanoprokaryota). Plant Syst. Evol. 1998, 210, 25–39. [Google Scholar] [CrossRef]
- Porta, D.; Rippka, R.; Hernandez-Marine, M. Unusual ultrastructural features in three strains of Cyanothece (cyanobacteria). Arch. Microbiol. 2000, 173, 154–163. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map showing the five sampling sites within the urban area of Bangkok.
Figure 2.
The numbers of taxa of the five phyla of epiphytic subaerial algae found at the sampling sites in Bangkok.
Figure 2.
The numbers of taxa of the five phyla of epiphytic subaerial algae found at the sampling sites in Bangkok.
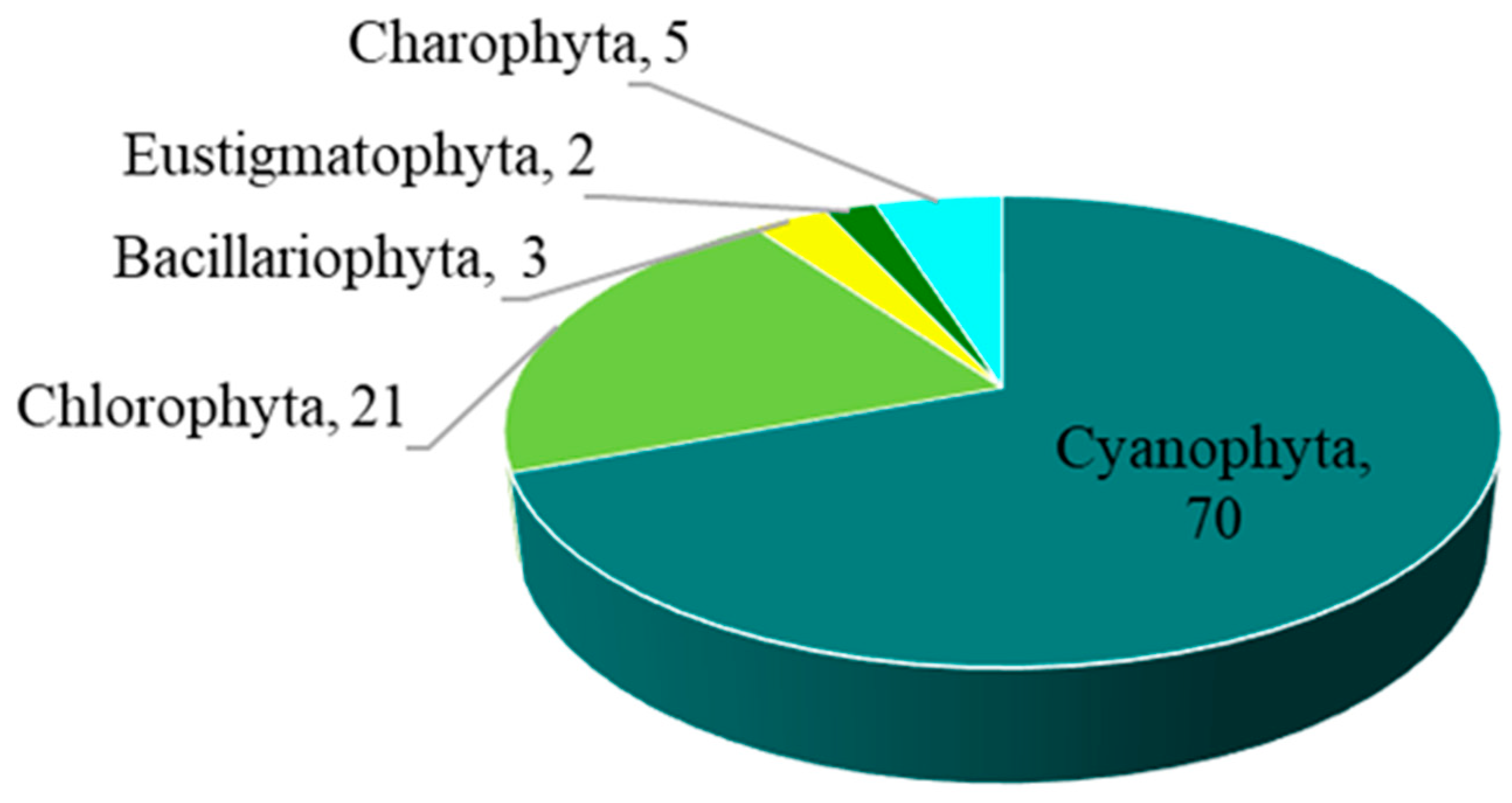
Figure 3.
The number of taxa of epiphytic subaerial algae recorded at the five sites in Bangkok.
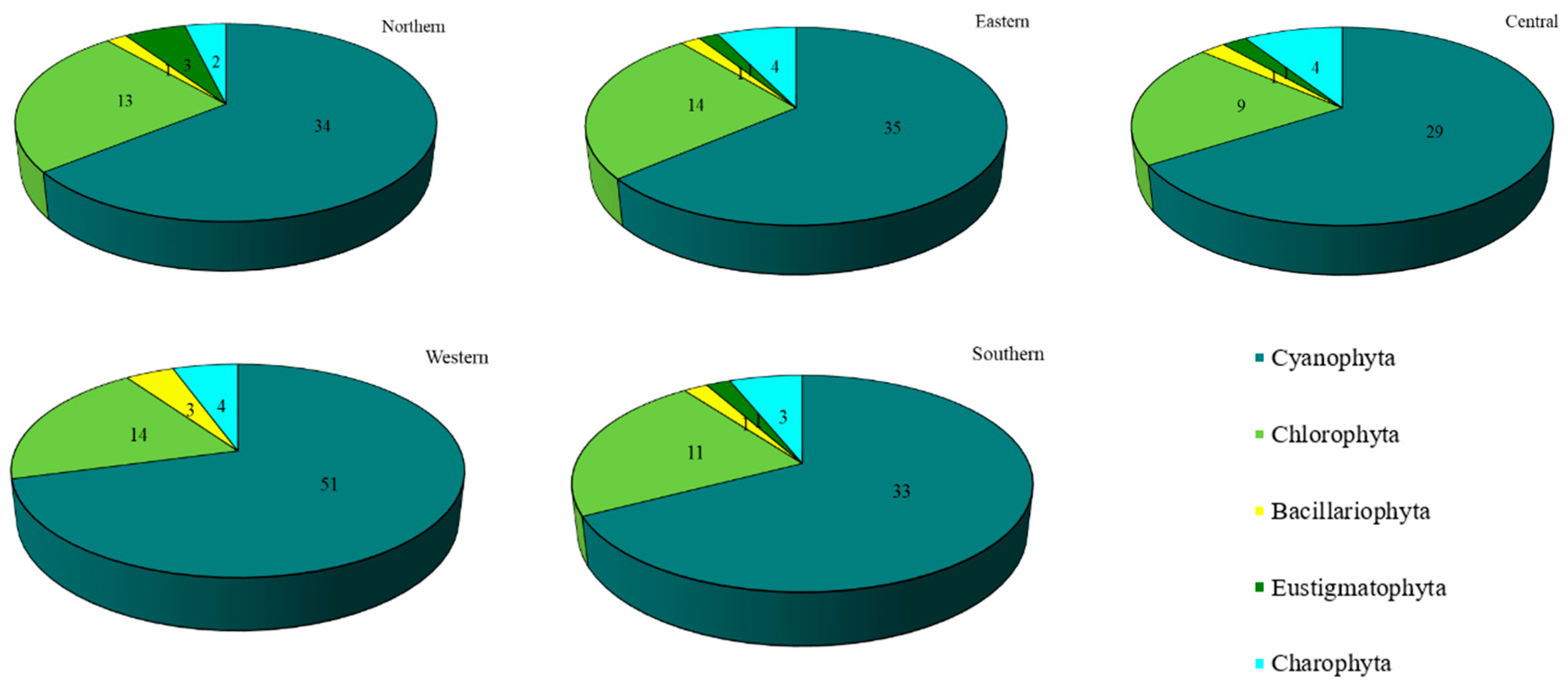
Figure 4.
The abundance of the five phyla of epiphytic subaerial algae found during the wet season in Bangkok, expressed as number of individuals.
Figure 4.
The abundance of the five phyla of epiphytic subaerial algae found during the wet season in Bangkok, expressed as number of individuals.
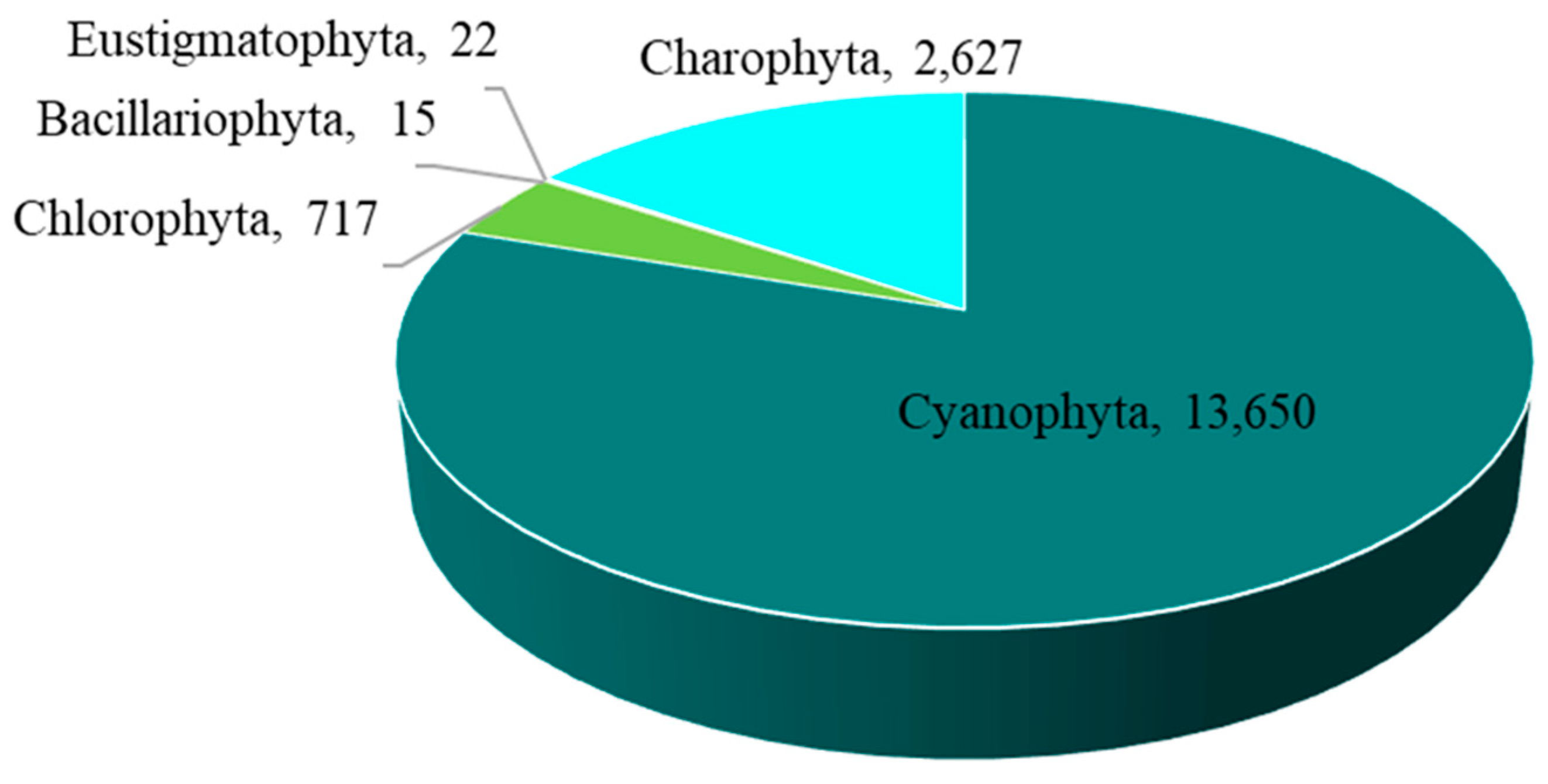
Figure 5.
The abundance of individuals of the dominant taxa of epiphytic subaerial algae (individuals/cm2) found during the wet season in Bangkok.
Figure 5.
The abundance of individuals of the dominant taxa of epiphytic subaerial algae (individuals/cm2) found during the wet season in Bangkok.
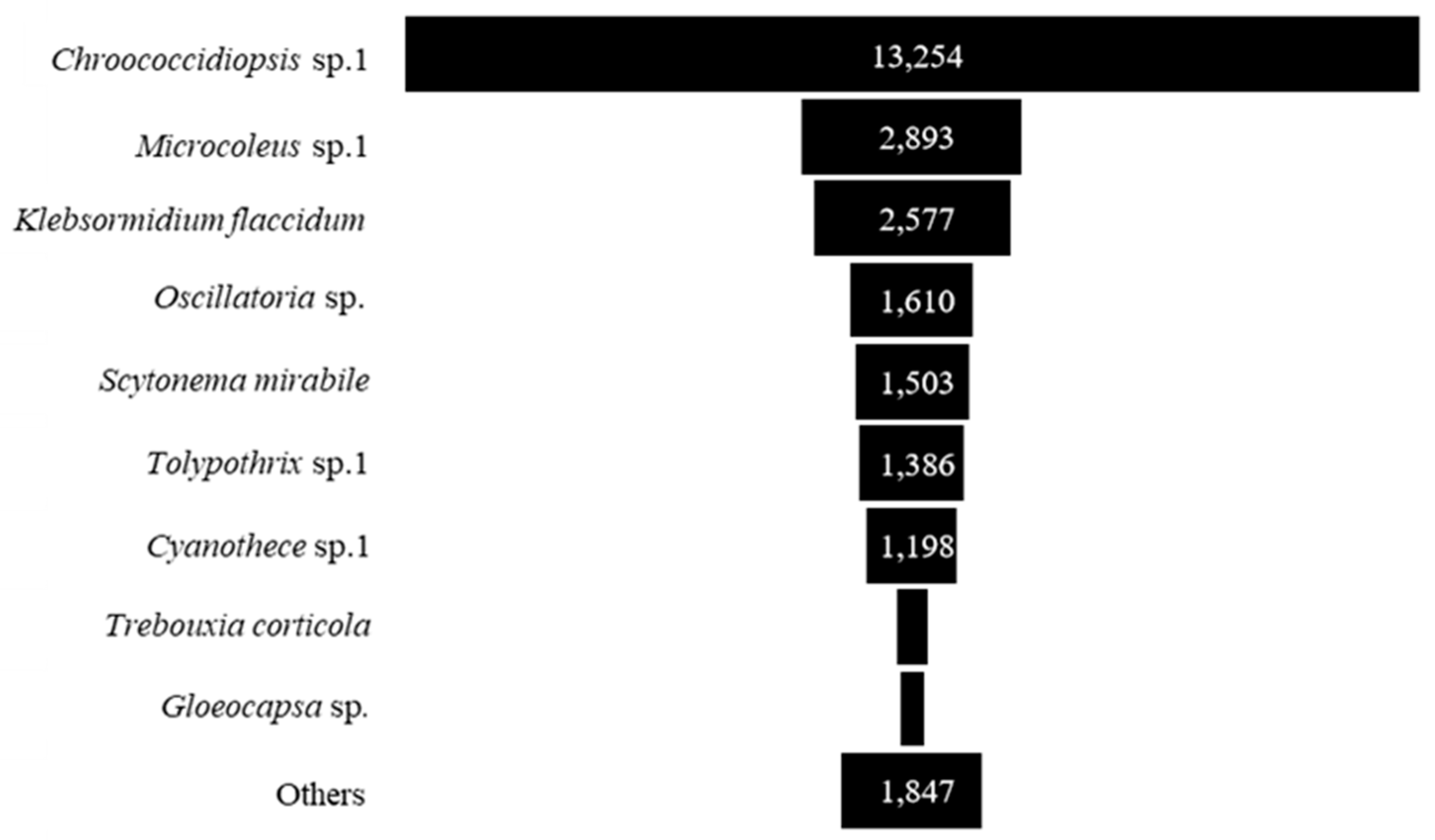
Figure 6.
The abundance of the phyla of epiphytic subaerial algae found during the wet season at the five sites in Bangkok.
Figure 6.
The abundance of the phyla of epiphytic subaerial algae found during the wet season at the five sites in Bangkok.
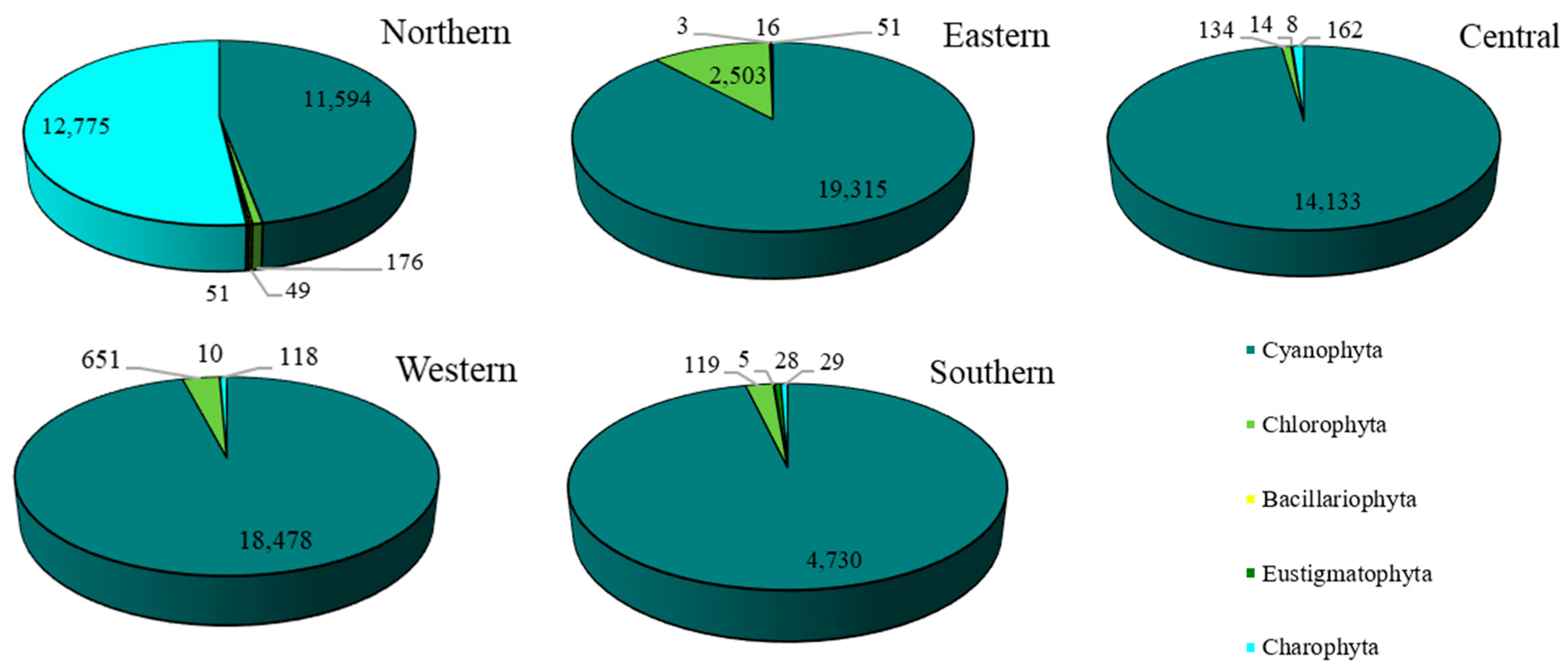
Figure 7.
Species abundance of epiphytic subaerial algae expressed in terms of numbers of individuals at the five study sites in Bangkok.
Figure 7.
Species abundance of epiphytic subaerial algae expressed in terms of numbers of individuals at the five study sites in Bangkok.
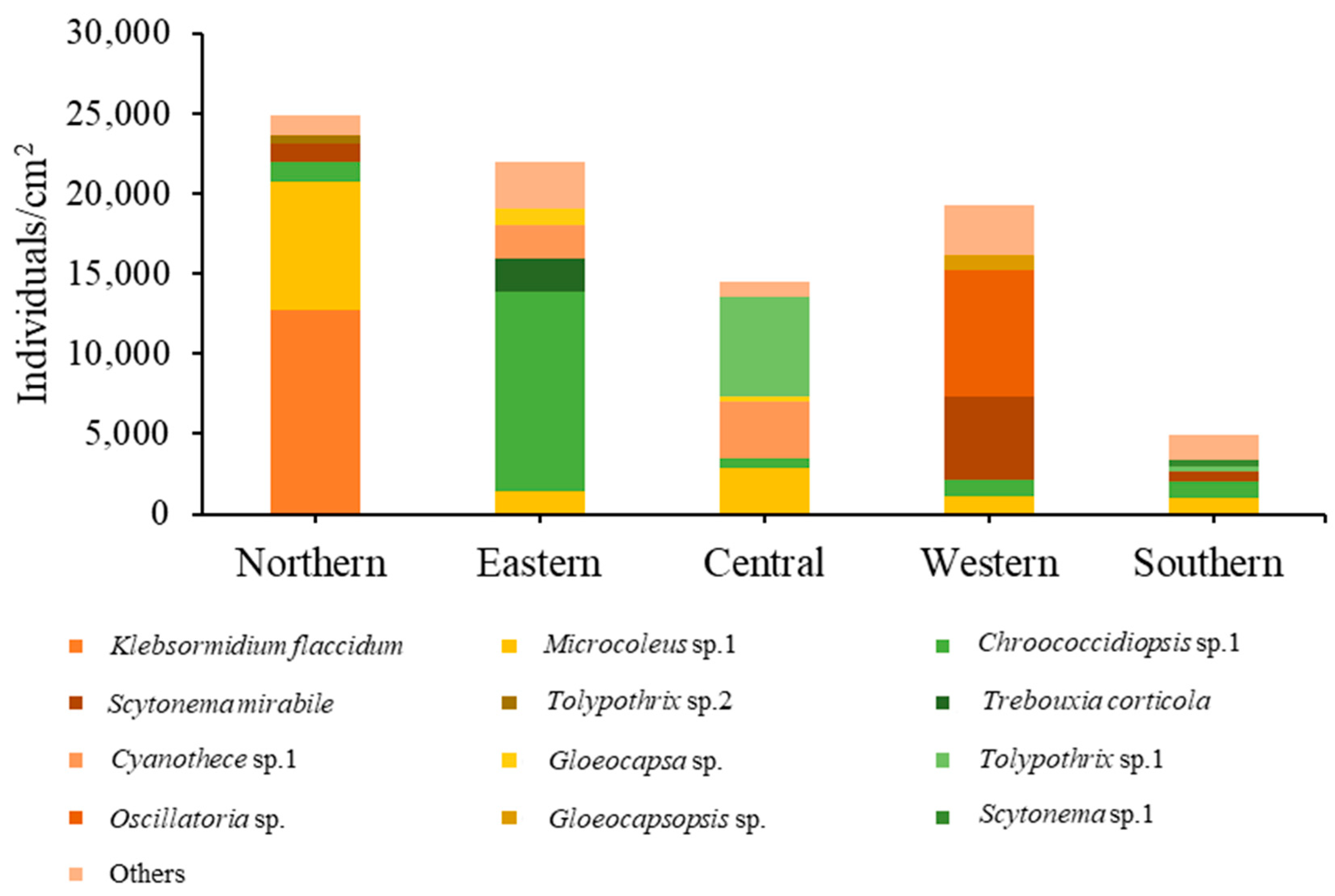
Figure 8.
Air pollutants (O3, NO2, CO, SO2, PM10, and PM2.5) found during the wet season in Bangkok (the error bars meaning to standard error, these were the pollutant measurements taken in every hour over 24 hours (24 times) in algal collection date).
Figure 8.
Air pollutants (O3, NO2, CO, SO2, PM10, and PM2.5) found during the wet season in Bangkok (the error bars meaning to standard error, these were the pollutant measurements taken in every hour over 24 hours (24 times) in algal collection date).
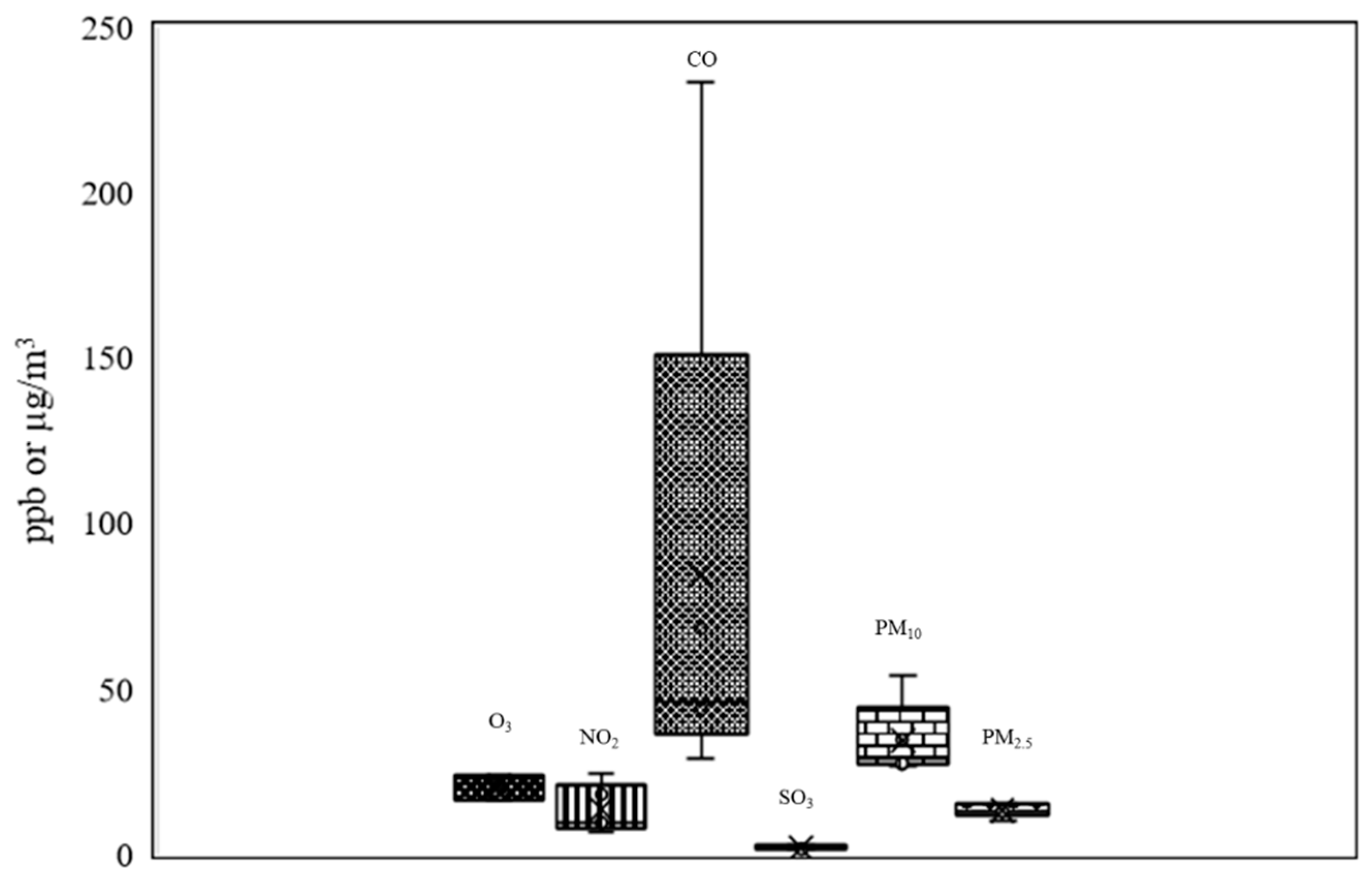
Figure 9.
Cluster analysis results of the epiphytic subaerial algal communities found at the five sampling sites in Bangkok.
Figure 9.
Cluster analysis results of the epiphytic subaerial algal communities found at the five sampling sites in Bangkok.
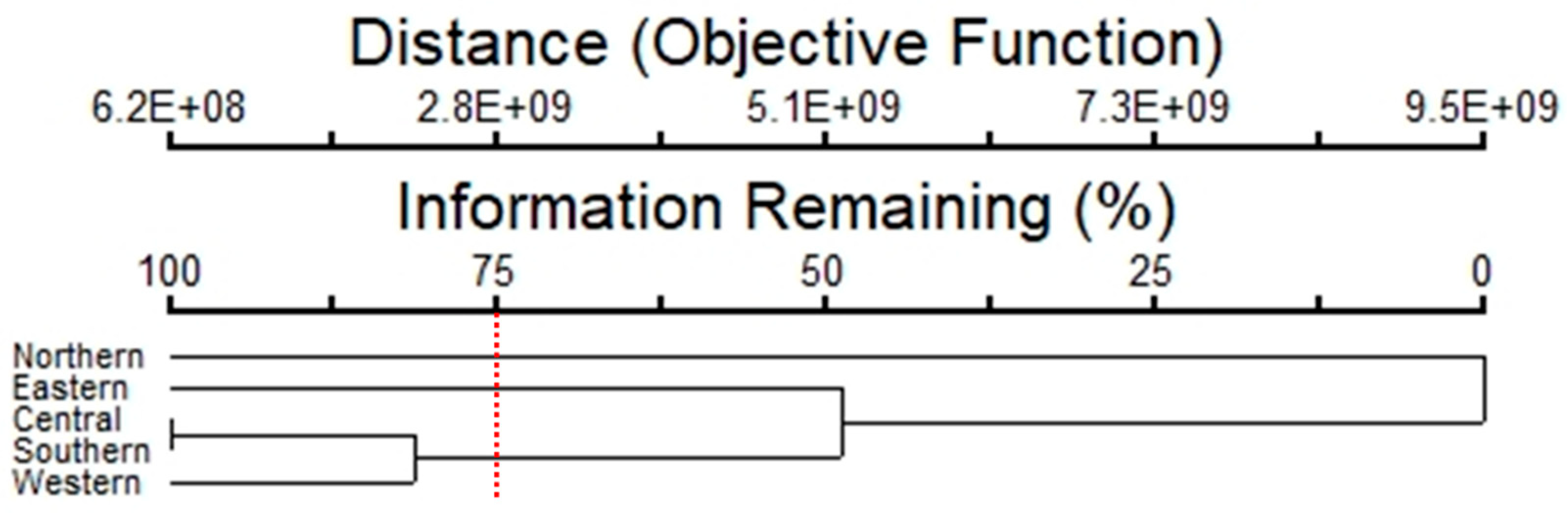
Figure 10.
PCA performed on data of air pollutants, diversity, and abundance of epiphytic subaerial algae found during the wet season in Bangkok. Axis 1 represents 41.623% and Axis 2 represents 28.075% of the observed variance.
Figure 10.
PCA performed on data of air pollutants, diversity, and abundance of epiphytic subaerial algae found during the wet season in Bangkok. Axis 1 represents 41.623% and Axis 2 represents 28.075% of the observed variance.
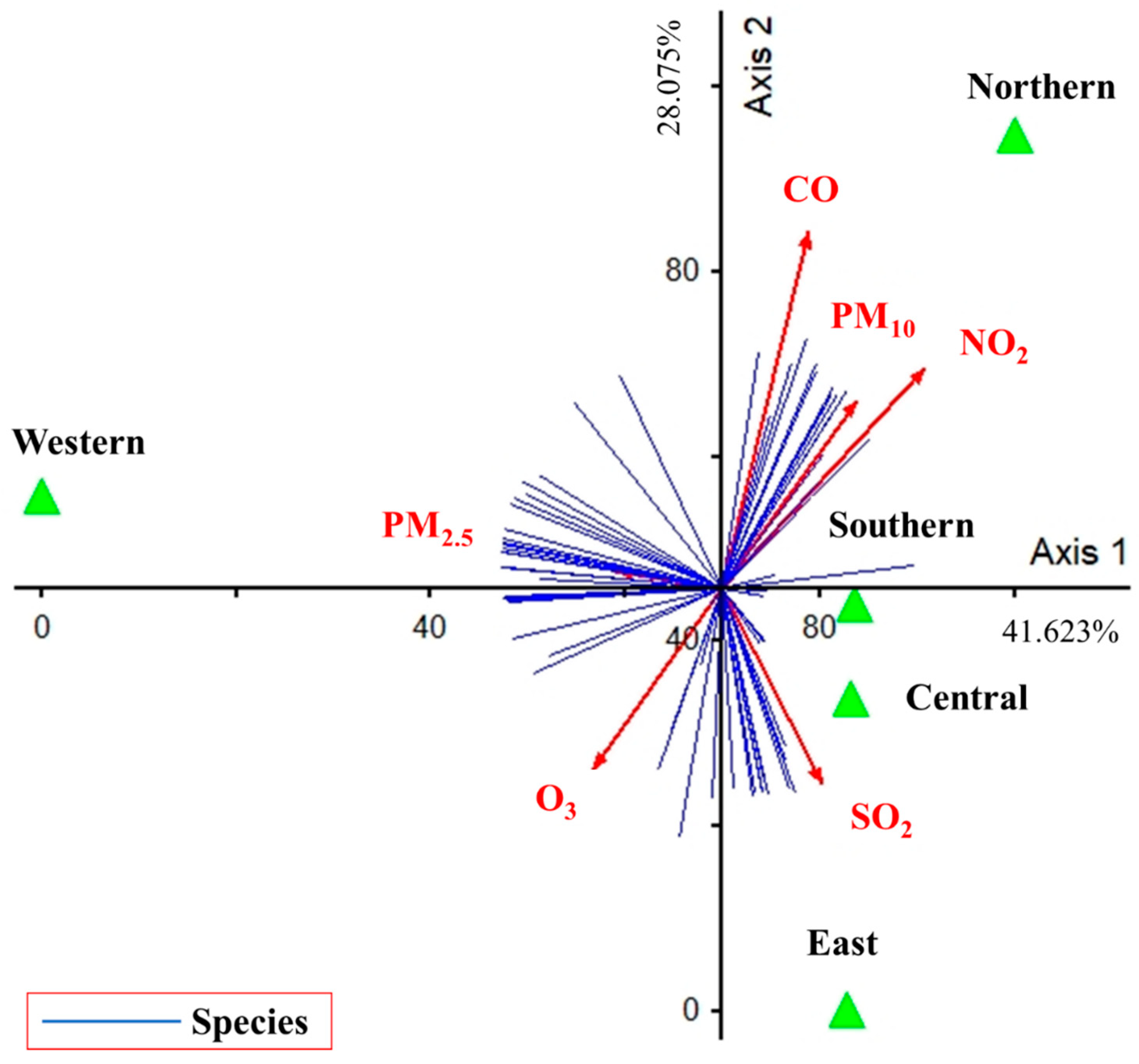
Table 1.
Details of the five sampling sites within the urban area of Bangkok.
| Sampling Sites | Location | Approximate GPS Coordinates | Altitude (m) | Date of Collection |
|---|---|---|---|---|
| Northern Urban Site (Vachirabenjatas Park) |
Bangkok | 13° 48.605' N, 100° 33.265' E | 2-7 | May 14, 2022 |
| Eastern Urban Site (Suan Luang Rama IX) |
Bangkok | 13° 41.288' N 100°, 39.488' E | 2-7 | May 15, 2022 |
| Central Urban Site (Lumpini Park) |
Bangkok | 13° 43.884' N 100°, 32.486' E | 4-6 | May 14, 2022 |
| Western Urban Site (Thawi Wanarom Park) |
Bangkok | 13° 44.645' N 100°, 21.139' E | 3-6 | May 13, 2022 |
| Southern Urban Site (Thonburirom Park) |
Bangkok | 13° 39.121' N 100°, 29.484' E | 4-5 | May 13, 2022 |
Table 2.
List of epiphytic subaerial algae recorded from sites within Bangkok. Nomenclature and taxonomy follow AlgaeBase [37].
Table 2.
List of epiphytic subaerial algae recorded from sites within Bangkok. Nomenclature and taxonomy follow AlgaeBase [37].
| Division | Order | Family | (n) Taxa |
|---|---|---|---|
| Cyanophyta | Unclassified | Unclassified | (1) Lusitaniella coriacea |
| Synechococcales | Leptolyngbyaceae | (5) Leptolyngbya sp. 1, Leptolyngbya sp. 2, Oculatella neakameniensis, Leptolyngbya sp. 3, Nodosilinea sp. | |
| Pseudanabaenaceae | (1) Jaaginema littorale | ||
| Synechococcaceae | (4) Synechococcus lividus, Synechococcus sp. Cyanobium gracile, Thermosynechococcus sp. |
||
| Pleurocapsales | Dermocarpellaceae | (2) Stanieria sp.*, Stanieria cyanosphaera | |
| Hyellaceae | (3) Pleurocapsa minor*, Hyella patelloides, Pleurocapsa sp. | ||
| Oscillatoriales | Coleofasciculaceae | (1) Coleofasciculus chthonoplastes | |
| Cyanothecaceae | (6) Cyanothece sp. 1*, Cyanothece sp. 2, Cyanothece sp. 3, Cyanothece sp. 4, Cyanothece sp. 5, Cyanothece sp. 6 | ||
| Gomontiellaceae | (1) Crinalium epipsammum | ||
| Microcoleaceae | (2) Microcoleus sp. 1*, Microcoleus sp. 3 | ||
| Oscillatoriaceae | (5) Oscillatoria nigro-viridis, Lyngbya aestuarii, Phormidium tinctorium, Oscillatoria acuminata, Oscillatoria sp.* | ||
| Nostocales | Aphanizomenonaceae | (2) Dolichospermum compactum, Sphaerospermopsis kisseleviana | |
| Hapalosiphonaceae | (1) Fischerella muscicola | ||
| Nostocaceae | (12) Cylindrospermum stagnale, Nostoc punctiforme, Nostoc linckia, Cylindrospermum sp., Nostoc sp. 1, Nostoc piscinale, Anabaena sp. 1, Nostoc sp. 2, Camptylonemopsis sp., Nostoc carneum, Trichormus azollae, Cylindrospermum muscicola | ||
| Rivulariaceae | (8) Calothrix sp. 1, Calothrix sp. 2*, Microchaete diplosiphon, Calothrix sp. 3, Calothrix brevissima, Calothrix sp. 4, Calothrix sp. 5, Calothrix sp. 6 | ||
| Scytonemataceae | (6) Scytonema mirabile*, Scytonema sp. 1, Scytonema hofmannii, Scytonema crispum, Brasilonema sp., Scytonema sp. 2 | ||
| Tolypothrichaceae | (2) Tolypothrix sp. 1*, Tolypothrix sp. 2* | ||
| Gloeobacterales | Gloeobacteraceae | (1) Gloeobacter kilaueensis | |
| Chroococcidiopsidales | Chroococcidiopsidaceae | (3) Chroococcidiopsis sp. 1*, Chroococcidiopsis thermalis*, Chroococcidiopsis sp. 2 | |
| Chroococcales | Chroococcaceae | (4) Gloeocapsa sp.*, Gloeocapsopsis sp.*, Chondrocystis sp., Gloeocapsopsis crepidinum |
|
| Bacillariophyta | Bacillariales | Bacillariaceae | (2) Nitzschia sp., Cylindrotheca closterium |
| Diadesmidaceae | (1) Diadesmis sp.* | ||
| Chlorophyta | Chaetophorales | Chaetophoraceae | (1) Dilabifilum sp. |
| Chlamydomonadales | Chlamydomonadaceae | (2) Chloromonas perforata, Chlamydomonas zebra | |
| Sphaeropleales | Scenedesmaceae | (2) Scenedesmus sp.*, Acutodesmus sp. | |
| Microthamniales | Unclassified | (1) Trebouxia corticola* | |
| Chlorellales | Chlorellaceae | (2) Nannochloris normandinae*, Lobosphaera incisa | |
| Unclassified | (1) Parachlorella kessleri | ||
| Prasiolales | Prasiolaceae | (1) Edaphochlorella mirabilis | |
| Microthamniales | Unclassified | (1) Friedmannia sp. | |
| Unclassified | Unclassified | (3) Heterochlorella, Watanabea reniformis*, Xylochloris irregularis | |
| Microthamniales | Unclassified | (5) Stichococcus sp., Symbiochloris irregularis, Symbiochloris reticulata, Trebouxia australis, Trebouxia decolorans | |
| Ignatiales | Unclassified | (1) Ignatius tetrasporus* | |
| Pyramimonadales | Unclassified | (1) Pyramimonas disomata | |
| Eustigmatophyta | Eustigmatales | Monodopsidaceae | (2) Monodopsis sp. 2, Nannochloropsis oculata |
| Charophyta | Chlorokybales | Chlorokybaceae | (1) Chlorokybus atmophyticus |
| Klebsormidiales | Klebsormidiaceae | (4) Klebsormidium flaccidum, Klebsormidium sp. 1, Klebsormidium sp. 2*, Klebsormidium sp. 3 |
* The 19 taxa of epiphytic subaerial algae collected within Bangkok were found in more than one park, belonging to the divisions Cyanophyta (13 species), Chlorophyta (5 species) and Bacillariophyta (1 species).
Table 3.
The diversity indices, richness indices, and equitability indices for the algal communities found in the five sampling sites in Bangkok.
Table 3.
The diversity indices, richness indices, and equitability indices for the algal communities found in the five sampling sites in Bangkok.
| Diversity Index (H') | Richness Index | Equitability Index (J') | |
|---|---|---|---|
| Northern Site | 1.37 | 4.43 | 0.3 |
| Eastern Site | 1.71 | 4.65 | 0.37 |
| Central Site | 1.58 | 3.84 | 0.34 |
| Western Site | 1.95 | 6.19 | 0.42 |
| Southern Site | 2.51 | 4.75 | 0.54 |
Table 4.
The similarity indices for the algal communities found at the five sampling sites in Bangkok.
Table 4.
The similarity indices for the algal communities found at the five sampling sites in Bangkok.
| Northern Site | Eastern Site | Central Site | Western Site | Southern Site | |
|---|---|---|---|---|---|
| Northern Site | ‒ | 8 | 22.99 | 15.19 | 27.22 |
| Eastern Site | ‒ | ‒ | 31.92 | 24.29 | 41.04 |
| Central Site | ‒ | ‒ | ‒ | 41 | 64.82 |
| Western Site | ‒ | ‒ | ‒ | ‒ | 54.26 |
| Southern Site | ‒ | ‒ | ‒ | ‒ | ‒ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



