
Submitted:
23 January 2024
Posted:
24 January 2024
You are already at the latest version
Abstract
A significant percentage of children suffer from neurodevelopmental aberrations, which have long-term effects on both individuals and society. Since the developmental timelines of different brain regions vary, the type, severity and timing of harmful exposures are crucial for understanding the specific pathological processes and consequences involved. During development, the cerebellum is one of the first brain structures to begin to differentiate, but one of the last to achieve maturity. This relatively long period underscores its vulnerability to detrimental environmental exposures throughout gestation. Moreover, as the postnatal functionality of the cerebellum is multifaceted, enveloping sensorimotor, cognitive, and emotional domains, prenatal disruptions in cerebellar development can result in a variety of neurological and mental health disorders. Here, we review major intrauterine insults that affect cerebellar development in both humans and rodents, ranging from abuse of toxic chemical agents such as alcohol and nicotine, to stress and sleep, malnutrition as well as infections. Understanding these pathological mechanisms in the context of the different stages of cerebellar development in humans and rodents may help us to identify critical and vulnerable periods and thereby to prevent the risk of associated prenatal and early postnatal damage that can lead to lifelong neurological and cognitive disabilities.
Keywords:
Cerebellum
; development
; critical periods
; alcohol
; smoking
; intrauterine insults
; sleep
; infection
1. Introduction
Neurodevelopmental disorders, characterized by cognitive, neurological, and psychiatric complications, affect one in six children in industrialized nations, causing enduring consequences with extensive societal and economic ramifications [1]. Detrimental prenatal environments can modify brain development, potentially leading to neurodevelopmental disorders [2,3]. The intricate influence of environmental variables on human fetal neurodevelopmental conditions is yet to be precisely determined. The developmental timelines of different brain regions vary, indicating that the type, severity, and timing of harmful exposures are crucial in determining resulting aberrations [4]. The cerebellum is a brain region that develops early and grows rapidly during gestation and continues to mature until late postnatal life [5]. Long-range connections from the cerebellum to the thalamus and cerebrum start to develop prenatally and continue to develop postnatally. This underlines the cerebellum’s vulnerability to detrimental environmental exposures during different early developmental phases [6].
2. Cerebellar function: a new perspective
Recent shifts in neuroscience perspectives have reframed the cerebellum, traditionally overshadowed by the corticocentric viewpoint, as a crucial area of study, acknowledging its role in both motor and non-motor functions [7,8,9]. Located within the skull’s fossa posterior, the cerebellum contains nearly two-thirds of the human central nervous system neurons, although occupying only one-ninth of cerebral volume. The cerebellar to cerebral neuron ratio remains relatively constant across mammals, highlighting a structural and functional interconnectivity, routed through the thalamic, pontine and inferior olivary nuclei [10]. The cerebellum’s intricate link with motor functionality has been acknowledged since Aristotle’s initial delineations in the fourth century BC [11]. Current understandings affirm the cerebellum’s pivotal role in modulating motor control, precision, and learning, correlating cerebellar damages with substantial motor disorders [12]. Schmahmann’s elucidation of the “Cerebellar Cognitive Affective Syndrome” marked a paradigmatic shift, extending cerebellar functionality to encompass cognitive and affective domains [13]. Contemporary research corroborates the cerebellum’s integration in attention, behavior, cognition, and language, but also associates its aberrations with developmental anomalies like dyslexia, autism, and attention deficit hyperactive disorder (ADHD) [8,9,14]. Its enduring neurogenetic development makes the cerebellum particularly susceptible to external disruptions in early life, highlighting the significance of studying the associated biomedical mechanisms and consequences.
3. Cerebellar anatomy
Regarding the gross anatomy, the cerebellar hemispheres, united by the vermis, can be divided into individual lobules I-X (reviewed in [15]), which are split into the anterior (lobule I-V), posterior (VI-X), and flocculonodular lobe by fissures (Figure 1).
The anterior lobe together with lobules VIIIA and B have been suggested to be the sensorimotor cerebellum, and the posterior lobe together with the lateral lobules Crus 1 and 2 of lobule VII the cognitive cerebellum [16,17,18,19]. The cerebellum receives information via the three-layered and folded cerebellar cortex that surrounds the cerebellar nuclei (CN) (Figure 1 and Figure 2).
The sole output neurons of the cerebellar cortex are the so-called inhibitory Purkinje cells (PCs). The expression patterns of different genes, proteins and enzymes, such as Aldolase-C [23], the small heat shock protein (HSP)25 [24], GRID2 [25],phospholipase C, and beta4 [26], appear to reveal a reproducible pattern of parasagittal stripes of PCs, also known as ZebrinII (ZII) zones, around which the rest of the cerebellar cortex is organized [27]. This anatomical and molecular differentiation is preserved across species and interestingly, the foundation for such a subdivision into individual cerebellar stripes and zones lies already in the early cerebellar embryology, including that of humans [28]. PCs are located in the middle layer (PCL) and their baseline simple spike firing frequency strongly depends on the intrinsic expression profile highlighted above [29,30] with the modulation thereof being tuned by various inputs. Most of these inputs reside in the outermost layer of the cerebellar cortex, the so-called molecular layer (ML). The interneurons in this layer, the MLI’s, have historically been divided into stellate and basket neurons, which inhibit PCs predominantly at their dendrites and soma, respectively. Instead, PCs receive their excitatory inputs from the climbing fibers (CFs) and parallel fibers (PFs). CFs arise in the inferior olive and PFs are the axons of granule cells (GCs), which are located in the GC layer (GL) and which in turn receive input from mossy fibers (MFs). The GL also comprises the inhibitory Golgi cells and in the nodule, uvular, and flocculus, the excitatory unipolar brush cells (UBCs). All neurons in the GL increase the diversity of the MF information, which then ultimately is relayed via the PF to the PCs. The PC axons in turn project onto the glutamatergic CN neurons that control the premotor and non-motor nuclei in the brainstem and diencephalon, as well as the GABAergic local interneurons and projection neurons that provide feedback to the inferior olive [31,32]. Even though the lamination and neuronal subtypes show similarities between humans and rodents, the ratio between the neuronal subtypes differs [33]. The fact that multiple genes and factors lead to this distinctive development of the cerebellum again underlines the vulnerability of the cerebellum when it comes to detrimental prenatal environments in humans and rodents.
4. Scope
The development of the cerebellum has been a focus of many outstanding lines of research and over the years multiple reviews have been published describing the development of the cerebellum and its cyto-architecture (for example [5,9,22,34,35]). The cerebellar anlage appears in humans from embryonic day (E) 29 and in rodents from E8.5 onwards (Figure 2 and Figure 3) [36,37,38]. Throughout prenatal development, the cerebellum can be affected by both genetic and intrauterine insults [6], often leading to major neurological and cognitive deficits. In line with the key stages of cerebellar development, we are focusing our review on intrauterine insults that are known to affect a myriad of human mothers and fetal cerebellar growth; these include maternal substance abuse such as smoking and alcohol intake, stress, sleep deprivation, undernutrition and chorioamnionitis (CA). Unlike human research [6], research in rodents permits identification of specific time points in cerebellar development that appear most critical for insults following manipulation. Indeed, abundant animal research has revealed the multifaceted negative effects of a hostile intrauterine environment on cerebellar development. However, human studies in this area are limited, focusing often on postnatal cerebellar manifestations (e.g., neuroimaging measurements), while disregarding the impact of prenatal anomalies on different subareas of cerebellar development and growth. Brain development in rodent pups generally resembles the development of human in that the orders of their different gestational periods correspond relatively well [39], but extrapolating rodent studies to the human situation remains difficult as the absolute and relative durations of the different stages vary substantially (Figure 2 and Figure 3; [22,40,41]). As a result, the critical periods of cerebellar development that are most vulnerable to particular agents vary among the different mammalian species. These differences must be considered when understanding the potential corresponding atypical developments in human and designing strategies for preventing or rescuing the related disorders.
5. Embryology of the cerebellum
Each species exhibits particularities regarding gestation. Term parturition in women occurs around 280 days after the onset of their last menstrual period [42]. In mouse and rat, gestation is shorter, lasting between 20 and 22 days [43]. Regarding the comparison between brain development of rodents and human, one must consider different aspects, since they do not run proportionally in parallel [22,44,45]. The developmental journey of the human cerebellum commences 30 days after conception and that of the rodent around E8.5 [36,37,38]. Forty to forty-five days after conception in humans, the human cerebellar ventricular zone (VZ) splits into a VZ and a subventricular zone [41]. Cerebellar maturation can be distinguished by a multifaceted, symbiotic cascade, encompassing gene expressions, electrical network interactions, and environmental factors, which are inherently interlinked with the evolution of the hindbrain (hb) and midbrain (mb). For example, instances of cerebellar hypoplasia in humans often coincide with hypoplastic conditions of the pons, attributed at least in part to secondary effects, such as axonal degeneration [46,47]. Notably, genes regulating cerebellar development exhibit expression in various brain regions, next to that in other organs such as the skin [48]. The transcription factors Otx2 and Gbx2, which are expressed anterior and posterior from the so-called isthmic organizer region between the mb and hb, are important for the general development of most of the rostral and caudal regions of the central nervous system [49]. Originating from the caudal-most primary neural tube vesicles, the rhombencephalon (hb) bifurcates into the metencephalon and myelencephalon. The foundational elements of the cerebellum emanate from the dorsal portions of the metencephalon (Alar plate) and the neural folds, termed rhombic lips (RLs) [50]. Segmented along the rostral-caudal axis into seven rhombomeres, the cerebellum’s emergence is facilitated by transcription factors and signalling molecules, leading to the formation of specialized epithelium, RL1, through bilateral expansion of the Alar plate [51] in the presence of Gbx2 and absence of Otx2 and Hoxa2 in rodents [49,52,53]. Wnt family members, fibroblast growth factors (especially Fgf8 and Fgf17), En1-2, Lmx-1, and sonic-hedgehog (shh) play a major role in defining the isthmic organizer region and the anterior-posterior as well as the dorso-ventral patterning of cerebellar development [54,55,56]. Mutations in any of these genes lead to severe implications for cerebellar development or even death in rodents [57,58,59,60]. In rodents, around E9, cerebellar histogenesis starts and the cerebellar anlage develops from two symmetric bulges. The bulges grow and give rise to the unitary cerebellar plate comprising the vermis and hemispheres. The cerebellar medial regions expand, an orthogonal rotation happens, and the cerebellar wing-like anlagen transform into a homogeneous cylindric vermis at E15.5 (Figure 2). Just above the fourth ventricle the two germinative compartments of the RLs are formed, adjacent to the roof plate and the VZ placed in the inner side. The RLs form the origin for all glutamatergic cerebellar neurons including GCs and CN neurons, amongst others, whereas all GABAergic neurons as well as glia cells, oligodendrocytes and astrocytes originate from the VZ (Figure 2 and Figure 3; [22,35,41]). Birthdating studies showed that the projection neurons are produced first, at the onset of cerebellar neurogenesis.
5.1. Glutamatergic neuron development
In rodents, the RLs are defined by the expression of the mouse homolog of Drosophila atonal (ATOH1) transcription factor [61] forming the origin of all glutamatergic cerebellar neurons [62]. Atoh1 expression begins at E9.5 in mice and from E10.5 to E12.5 progenitors leaving the rostral RLs give rise to the large CN neurons, migrating to the surface of the cerebellar anlage where they aggregate in the nuclear transitory zone. From there they move inward from the PC plate to form the four CN on both sides [63]. Progenitors migrating between E14 and E21 give rise to UBCs [64] and then GC progenitors exit the upper RLs moving tangentially along the cerebellar surface, by E16 eventually covering the entire cerebellar anlage [65]. GCs start proliferating in response to shh signalling from PCs [66]. The GC lineage arises already around E8.75. At least three transverse GC progenitor zones are identified by gene expression and birth-dating. Postmitotic GCs migrate from the external GL (eGL) to the inner GL guided by the Bergmann glia fibers, which are oriented in the same plane. Thereby the eGL topography is projected into the nascent inner GL [65]. After birth the GL is an 8 layered structure with another layer of proliferating granule precursor cells [65,67]. The proliferation window of murine GCs progenitors closes only at the end of the second postnatal week, when the last postmitotic GCs from the deepest portion of the eGL migrate inwardly to the nascent adult GL, marking the end of the eGL and ceasing Atoh-1 expression [61,68]. Interestingly, recent studies showed that the cell-fate specification among the cerebellar VZ and RL is not absolute. A so-called posterior transitory zone expresses genes to develop bipotent progenitors for cerebellar glutamatergic neurons and the choroid plexus (Figure 2 and Figure 3; [69]).
5.2. GABAergic neuron development
The VZ is defined by the pancreas transcription factor 1-a (Ptf1a), giving rise to GABAergic neurons. A Ptf1a - neurogenin 1/2 (Neurog1/2)- early B-cell factor 2 (EBF2) regulatory network is implicated in PC subtype specification [70]. In rodents, PCs are born between E10 and E13 and undergo terminal mitosis. Dividing VZ precursors emigrate into the cerebellar prospective white matter, via the cerebellar plate and form an array of clusters (E14-E18), which are suggested to aggregate into topographical organization centres (TOCs). These TOCs are not only specific for ingrowing afferent precerebellar MF and CF inputs as well as interneurons, but also for subsets of glia cells and migrating GCs [71]. As the PC clusters disperse into parasagittal stripes, all components disperse with them, forming the adult cerebellar parasagittal architecture. MFs disconnect and form local connections with GCs within the zone. At least five molecularly distinct PC subgroups have been identified throughout development with distinctive levels of Foxp1 and Foxp2, respectively. Foxp1+/Foxp2+ PCs strongly express reelin receptors and lack Ebf2. Reelin controls PC migration [72,73,74,75]. Early born PCs are likely to become ZII+ during adulthood, while late-born PCs adopt the ZII− phenotype, which is in line with the high ZII expression in the phylogenetically older vestibulocerebellum [76]. The postnatal development of PCs can be divided into different stages. The first distinction can be made between intrinsic maturation by PCs themselves, and guided maturation by stimulation of other cell types, such as GCs [77]. In rodents, intrinsic growing starts with a fast somatic growth from P0-P9 followed by a rapid dendritic growth from P9-P18 [78,79] with more processes growing outwards from the soma. During the second postnatal week, the processes become more complex by growing rapidly and increasing the number of branches. This is the start of the dendritic tree, which will be completed around postnatal week 4 in rodents [80]. Different PC subtypes display distinct susceptibilities to environmental insults or genetic mutations [81]. CFs are also involved in the dendritic arborisation during these stages, presumably by stimulating PCs. This may explain why higher-order mammals, including humans, show both a higher level of persistent multiple CF innervation and a higher complexity of the dendritic trees of their PCs [82]. GABAergic interneurons in the CN are born between E10.5–E11.5, and Golgi cells at approximately E13.5–postnatally (peak around E14–E16) [35,83,84]. Late-born GABAergic interneurons, including stellate and basket cells, derive from secondary precursors in the prospective white matter at later stages (from E13 to P5 with a peak around birth) [85]. Thus, cerebellar neuronal subtypes depend on when and where they are generated from neural progenitors. This leads to the idea that cerebellar progenitors with their own spatial and temporal identities produce specific neuronal subtypes. Additionally, the cerebellum accommodates astrocytes, glia, and oligodendrocytes, the origins of which remain subjects of ongoing discussions [86].
5.3. Embryology of the precerebellar system
CF neurons are derived from the dorsal neuroepithelium (dP), the caudal hb (RL 6-8) at E9.5 to E11.5 in rodents. Olivocerebellar projections are being formed at E17.5 [87] and already at the late embryonic stage the olivocerebellar bundle shows an organized topographic projection pattern. Neurons in a particular subnucleus of the inferior olive project via their CFs to a specific part of the cerebellum. The bundles with CFs run contralaterally through the inferior cerebellar peduncle. Axons that leave the peduncle rostrally project to the vermis, whereas CFs innervating the other cerebellar areas pass through the more caudal parts of the inferior cerebellar peduncle [88]. During early development CF axons form a delicate plexus with abundant branching, while in the second postnatal week axonal branches disappear and those so-called nest terminals grow into an entire CF terminal in the following weeks forming the distinct one-to-one synaptic connection in rodents (Figure 2), (for comparison in human, see recent [82]). MFs are derived from the dP1 domain of the caudal hb (RL 6-8) and are generated at slightly later stages (E10.5-E16.5) compared to the CFs [89]. Axonal fibers of GCs that receive MF input make initially contact with the soma of PCs, but around P5-P15 they start to turn into the typical parallel fibers that establish synaptic contacts with the dendrites of PCs [80]. PCs and GCs generate their zonal circuit map early during development [90].
6. Extrinsic deterrents influencing cerebellar development
Multiple genetic and external conditions can significantly hinder both pre- and postnatal development of the cerebellum, resulting in impaired maturation and functionality. In this review we focus on four common intrauterine insults in humans (Figure 4) that affect fetal cerebellar development. These common disruptors during prenatal stages include 1. teratogenic exposures during pregnancy to substances like alcohol and nicotine; 2. increases in cortisol level (stress); 3. Intrauterine growth restriction (IUGR) resulting from e.g., malnutrition; and 4. chorioamnionitis (CA) [2]. Thus, lifestyle choices and environmental exposures of mothers can impact the cerebellar development of fetuses, potentially causing lifelong consequences for the structure and function of the cerebellum [2,6].
6.1. Maternal substance use
The negative effects of alcohol and other external prenatal cerebellotoxic substances, such as nicotine, cocaine, heroin, specific anti-epileptics (phenytoin), specific anti-depressants (lithium salts) and many more drugs have been shown to be harmful to the maturation of the fetal brain, including decreased volumes, increased apoptosis, and behavioural differences [91,92,93,94,95,96]. In a study examining alcohol consumption habits of pregnant adults aged 18–49 in the U.S. between 2018-2020 (data from the Behavioral Risk Factor Surveillance System (BRFSS)) revealed that 13.5% reported current drinking (at least one drink in the past 30 days) and 5.2% reported binge drinking (four or more drinks on a single occasion in the past 30 days) [97]. About a decade ago, a substantial part (29%) of the women engaged in drinking were also involved in smoking [98]. The reported drinking habits during early pregnancy are likely underestimated, since many women don’t modify their alcohol consumption until pregnancy confirmation, often excluding the initial post-conception period in reflections on pregnancy behavior. In this section, we focus on the maternal use of alcohol and nicotine during pregnancy in humans, and the prenatal and early postnatal exposure of these substances in rodents.
Human studies on the correlation between maternal alcohol consumption and fetal cerebellar growth have yielded inconsistent results [99,100,101,102,103,104,105,106]. Excessive alcohol exposure during pregnancy results in children with fetal alcohol spectrum disorder (FASD), which usually comes with symptoms such as balance disturbances and retardation of motor skills [107]. Cerebellar degeneration might contribute to some of the cognitive disabilities that can be observed in children with FASD [108]. For example, children with anterior vermal dysmorphology show negative verbal learning and memory performance later in life [109]. Likewise, several studies have shown decreased cerebellar size and significantly stunted cerebellar growth in the offspring of women who indulged in heavy drinking during pregnancy compared to those who abstained [99,105]. However, one other study investigating the effects of alcohol on cerebellar development did not show significant changes in cerebellar size compared to controls [109].
In rats, alcohol administration for only 1 day during the first postnatal week results in reduced brain weight at P10, with the cerebellum being diminished the most [91]. More specifically, alcohol administration between P2-P5 leads to reduced volumes of the GL and ML and a significant loss of PCs and GCs in all lobules of the vermis, except for the PCs of lobules VI and VII [91,110,111,112,113]. Due to a ‘temporal window of vulnerability’ [114], alcohol administration to rodents in their first postnatal week affects the cerebellum, whereas alcohol administration in their second postnatal week leads to non-significant effects [111]. The results of Hamre and West (1993) suggest that this window of vulnerability ends after P7, with the biggest loss of PCs after ethanol treatment at P4-5 [115]. Question is to what extent differential gene expression plays a role in this temporal window of vulnerability. The timeframe-dependent gene expression, as discussed in the first section of this review, differs between the first and second postnatal week. Math1 and Cyclin D2, both markers for immature neurons in the cerebellum and expressed in GC precursors, are found at relatively high expression levels during the first postnatal week compared to the second. After alcohol administration, the expression level of Math1 increases at both timeframes similarly, whereas the expression of Cyclin D2 significantly increases at P10 compared to P4 [111]. Chronic maternal alcohol exposure during pregnancy in a rodent FASD model results in increased oxidative stress, measured as Reactive Oxygen Species (ROS) levels, Monoamina Oxidase (MAO) and lipid peroxidation. Additionaly, chronic alcohol exposure results in decreased GluR1, PSD-95, and integrin-linked kinase levels in the cerebellum of 6-weeks-old offspring [116]. The latter in turn may result in changes in synaptic plasticity and dendritic morphology in neurons of the cerebellum [116]. Capase-3 levels also increase in the cerebellum following prenatal alcohol exposure, which may indicate increased apoptosis. However, neither PCs were found to be decreased nor the cerebellar to body-weight ratio to be reduced after alcohol adminisattion during the third week of pregnancy but ethanol treatment before pregnancy, during pregnancy and during lactation resulted in changes. One study shows morphological differences in the eGL, GCs and Bergmann glia in offspring [117]. Additionally, alcohol exposure during both E12-19 and P2-9 resulted in decreased numbers of inhibitory interneurons in lobule II, lower numbers of PCs in lobules II, IV-V and IX, and decreases volumes of lobules II, IV-V, VI-VII , IX and X of the vermis [118]. The latter results again support the idea that periods of vulnerability exists when it comes to alcohol exposure and cerebellar development.
Taken together, the literature is not consistent about the impact of maternal alcohol exposure on cerebellar maturation in human and rodents. However, rodent studies revealed that there is a clear negative impact of alcohol usage on cerebellar development but that the effects depend distinct periods of vulnerability. The developing cerebellum seems to be most vulnerable to alcohol administration during the first week of the rodents’ postnatal life, which corresponds with last trimester of pregnancy in humans. Within the cerebellum, PCs located in lobules I-V and IX-X of the vermis are most affected by alcohol abuse, whereas lobules VI and VII of the vermis seem to be less sensitive to alcohol abuse during the first postnatal week. Whether such periods of vulnerability also exist in humans needs to be determined.
6.2. Impact of smoking and nicotine exposure on cerebellar maturation
6.2.1. Maternal smoking
Research on maternal smoking has revealed its notable adverse influence on fetal cerebellar development [95,96]. Several human studies conducted have consistently shown associations between maternal smoking and underdevelopment in fetal cerebellar structure, often revealing smaller transcerebellar diameter and smaller head parameters in the offspring of smoking mothers compared to those of non-smoking mothers [95,96]. Indeed, smoking during pregnancy is associated with altered maturation, such as reduced brain volumes of the cerebellum and prefrontal cortex at birth in human studies [119]. In addition, defective maturation and migration of PCs were noted in postmortem histology specimens of fetuses [95]. The high incidence of developmental abnormalities in these cases suggests a strong link between tobacco exposure and malformations in the cerebellum during pregnancy. There is an urgent need for more data on the impact of nicotine on human fetal brain development, especially in light of the rising popularity of new nicotine products like e-cigarettes during pregnancy [120,121,122].
In rodents, smoking is known to affect cerebellar development during different developmental stages. Many studies have investigated the impact of smoking or nicotine exposure on the maturing brain. Nicotine, which is the psychoactive drug in tobacco, is able to cross the placenta and can even be found for longer periods of time and in higher concentrations in the fetus compared to those in the mother [123]. When exposed to the maturing embryo, nicotine has an inhibitory effect on the development of stem cells [124]. Prenatally, nicotine exposure during almost the whole pregnancy results in a significant increase in the density of dying PCs and a reduced density of surviving PCs in adolescent and adult rats [125,126]. Moreover, glial fibrillary acidic protein (GFAP) immunoreactivity expression is significantly increased in the GL and white matter of the cerebellum in adolescent and adult rats [125,126], as well as increased levels of neuroinflammation and oxidative stress have been measured [127]. The vulnerability to maternal nicotine exposure seems to be time sensitive, with more severe histomorphological differences of the PCs after 3 weeks of daily nicotine exposure compared to 2 weeks of exposure in offspring examined at P10 [128]. Postnatally, nicotine exposure in the first postnatal week increases apoptosis in the inner GL [129], and nicotine exposure in the first and second postnatal week reduces PCs in the cerebellar vermis in rats [110].
6.2.2. E-cigarettes
Maternal exposure to e-cigarettes, containing propylene glycol and vegetable glycerol, during the first three weeks of pregnancy in mice results in increased pro-inflammatory cytokine IL-6 levels in the cerebellum of adolescent offspring [130], indicating an increase in neuroinflammation. Surprisingly, mother dams that are exposed to e-cigarettes that do not only contain propylene glycol and vegetable glycerol but also nicotine, do not show increased levels of the tested cytokines in the cerebellum [130]. How nicotine might have a protective effect in this regard remains to be elucidated.
6.3. Stress and sleep
6.3.1. Impact of stress
Changes in the cortisol levels of mammals can affect the neuronal development of the unborn offspring significantly. Indeed, in humans maternal stress during pregnancy can influence fetal cerebellar development through a process known as ‘fetal programming’ [131]. This concept posits that variations in the intrauterine environment during critical fetal developmental phases can induce enduring changes in both the structure and function of the fetus, including the brain [132]. Such alterations arise when the fetus adapts or prepares for the expected postnatal environment based on these prenatal signals. Increased maternal anxiety can intensify the release of glucocorticoids and diminish the integrity of the placental barrier, facilitating greater glucocorticoid transmission to the fetus. Additionally, heightened anxiety can reduce uterine blood flow, and negatively impacting brain development. Such prenatal environmental changes may lead to cognitive, motor, and behavioral challenges in children [132]. Moreover, disruptions in neuroendocrine pathways like the hypothalamic-pituitary axis have been observed in offspring of mothers with heightened anxiety, suggesting potential long-term impacts on brain structure and function [133]. In a study by Buss et al. (2010) high pregnancy anxiety at 19 weeks gestation was associated with gray matter volume reductions in several brain regions, including the cerebellum, in offspring aged 6 to 9 years, as revealed by structural MRI scans [134]. Furthermore, prenatal exposure to maternal psychological stress is linked with increased sleep problems in toddlers and is associated with decreased fetal cerebellar-insular connectivity, although the specific mediating effects of fetal brain regions remain unidentified [134].
Similarly, in rats, stress can induce many developmental abnormalities. For example, maternal stress at E7 and E14 during the rats second week of pregnancy resulted in decreased nuclear sizes of PCs and GCs, increased PC proliferation, increased density of PCs, reduced synapse-to-GC ratio, reduced GC-to-PC ratio, as well as decreased synaptophysin expression in the GCs during adolescence [135,136]. Likewise, maternal stress in rats during their third week of pregnancy results in long-lasting morphological differences in PCs located in the vermis of their pups. The surface of the dendritic trees of their PCs increases during adolescence, but decreases during adulthood, accompanied by an increase in anxiety-related behaviour [137,138]. Instead, prenatal stress induced by glucocorticoid administration at the end of the pregnancy results in reduced numbers of dendritic branches of the PCs in both adolescent and adult rats, as well as in increased levels of mGluR1 in adults [139]. Importantly, the negative impact of stress extends beyond pregnancy. For example, stress induced by daily separation of the pups from the mothers for 1 hour a day at P2-14 leads to a decrease in glucocorticoid receptor expression in the CN, while it impairs associative cerebellum-dependent learning later in life [140]. Along the same vein, stress induced by transportation of rodents during the 2nd postnatal week leads to changes in the excitability of CN neurons [141].
Overall, the evidence for the negative effects of stress during pregnancy or during early postnatal life on development of the cerebellum is robust, but the specific impact depends on the precise period of stress induction and presumably also on the intensity thereof. For example, whereas a relatively short period of 3 hours of maternal deprivation a day during the first two postnatal weeks increases neurogenesis and cell density in the GL of rats [142], 24 hours of maternal deprivation at the end of the second postnatal week increases cell death in the GL [143]. These findings may be explained by differences in neuronal vulnerability to stress during the first weeks of postnatal life [144,145]. Indeed, during the first 2 weeks after birth the stress-response may be relatively mild, which comes with decreased corticosterone levels and reduced levels of the adrenocorticotropic hormone (ACTH) and corticosterone release after mild stressful events [144,145]. This ‘stress-hyporesponsive period’ [145] is suggested to have a protective function for the maturing brain against elevated glucocorticoid levels [145]. High levels of glucocorticoids during these timeframes are known to affect maturation and are for example related to increased apoptosis of cerebellar GCs in the eGL and inner GL [146]. However, during high stressful events, the sensitivity to stress is increased, indicating that this stress-hyporesponsive period is probably only a mechanism to protect the brain against mild stressors, i.e., not life-threatening situations [147]. It will be interesting to gain a better understanding to what extent the beneficial impact of the stress-hyporesponsive period during early life exactly affects the cerebellum and other brain regions, and for how long during postnatal life these beneficial effects last[142].
6.3.1. Impact of sleep deprivation
Both during late gestational periods and early postnatal periods, new-born humans and rodent pups sleep almost 80% of their time and it has been suggested that active sleep periods play a major role in the functional development of the cerebellum. For example, given that the cerebellum receives a copy of motor commands as well as subsequent signals about sensory feedback during sleep periods with muscle twitches, the cerebellum may be entrained during sleep to develop predictive coding of movements [148,149]. During such active sleep synapses may be strengthened or weakened for the sensorimotor system to develop. As a consequence, without sleep related twitching the cerebellum may not develop its distinct ability to process the motor commands and sensory feedback signals within the expected time period [150,151,152,153]. Therefore, it is not surprising that Tfap2b, which is a gene that acts during early embryonic stages, controls not only sleep in mice, but also affects functioning of GABAergic neurons in the cerebellum [154]. In this regard too, it will be interesting to find out which sleep-control genes and how sleep restriction affects development of the cerebellum and/or that of other brain regions [155]. Considering that sleep deprivation is a major stressor of pregnant mothers, a study testing the effects of sleep deprivation is overdue, both with regard to the development of the cerebellum and that of other brain regions [156,157].
6.4. Intrauterine growth restriction
IUGR affects around 10% of human pregnancies and is associated with long-term motor and cognitive problems [158,159]. The cause for IUGR can be maternal factors such as undernutrition and maternal smoking but also placental and cord abnormalities, as well as fetal factors such as congenital heart disease. Obviously from the description, since the causes for IUGR are heterogeneous, there is also overlap with the consequences described for the other insults in this review [160]. IUGR leads to a heterogeneous set of fetal clinical pathologies. Recent studies suggest that certain motor deficits in patients can result from abnormal cerebellar development due to IUGR [161,162].
In humans, instances of decreased cerebellar volume, or cerebellar hypoplasia, are frequently observed in fetuses experiencing preterm birth (i.e., babies born <37 weeks of gestation) and/or IUGR (also called fetal growth restriction: FGR) [163]. Alterations to typical cerebellar growth can occur because of influences that can be relatively direct or indirect [164]. The neuropathology underlying IUGR is intricate and unique compared to preterm infants without IUGR and term infants exposed to acute hypoxia. Research, spanning human imaging, post-mortem examinations, and animal models, often paints a picture of IUGR brains having diminished volume, compromised gray and white matter structures, and cellular anomalies. Specifically, gray matter regions exhibit fewer cells with a chaotic cortical configuration, whereas white matter appears immature with signs of inflammation and astrogliosis [163]. The structural connectivity, especially along motor and cortico-striatal-thalamic tracts, has been suggested to be altered in IUGR brains, correlating with adverse neurodevelopmental outcomes in affected children [165,166]. The risks of neurodevelopmental impairments in IUGR are modulated by the severity of growth restriction, its onset timing, the presence of relative “brain sparing” and gestational age at birth.
Rodent studies have shown that bilateral uterine vessel ligation to restrict blood flow to the fetus can be used as a model of IUGR in that it leads to a general decrease in body weight, including that of the brain. Artery ligation in mice from E12.5 days onwards leads to cerebellar changes in myelination especially when it is combined with hyperoxia [167]. Unfortunately, no specifics about the region of the cerebellum analysed or the neuron types affected by demyelination were given. Applying artery ligation at E18 in rats leads towards the end of the first postnatal week to a 30% increase in the width of the eGL, while there is no difference in the width of the proliferative zone or the proliferating marker Ki67 in GCs. The increase in eGL following artery ligation may be partly because Bergman glia cells and their fiber density become disorganized and decreased [168,169]. Indeed, since the Bergman glia fibers normally guide the migration of the GCs from the eGL to the GL during early development, any structural disturbance in the Bergman fibers may affect GC transfer [168,169]. Additionally, the expression of genes that are necessary for a healthy migration to the GL may be affected following uterine vessel ligation, which could further worsen the deficits in cerebellar development. GC defects in turn may affect normal PC development, as suggested by guinea pig studies [170]. Malnutrition of the rat fetus during the last 5 embryonic days leads to reduced levels of glutamic acid decarboxylase (GAD) only at P2, examined with High-performance liquid chromatography (HPLC) of the whole cerebellum [171]. Compared to controls, HPLC also revealed an increase in the amino acids, alanine and taurine. Thus, rodent data suggest that in babies with IUGR the cerebellum is likely to be affected, since GCs cannot sufficiently migrate from eGL to GL, which in turn may impact development and myelination of the PCs. However, one has to consider the limitations and caveats of the uterine vessel ligation animal model, since human placental insufficiency usually develops more gradually across the different gestational periods with the consequence that cerebellar development can be affected differentially with more or less potential for compensatory mechanisms to be engaged dependent on the specific cause.
6.5. Infections
Chorioamnionitis, or CA, is an infection of the chorion and amnion of the mother that can lead to a fetal inflammatory response, with adverse consequences for the developing fetal brain [172]. When brain inflammation is prolonged and/or becomes severe, it can exacerbate damage through further influx of cytokines, chemokines, and other inflammatory mediators released from glial cells. In humans, there is a strong causal link between CA, preterm brain injury, and the pathogenesis of severe postnatal neurological deficits, such as cerebral palsy [173]. When measured in premature infants, there is a significant association between exposure to CA and neurodevelopmental impairments from 18 to 30 months of corrected age [174,175,176], decreased cognitive performance at 5 years [173], and autism spectrum disorder [177]. The most frequent route that causes CA development in humans is the ascending microbial invasion from the lower genital tract. In a recent study conducted by Jain and colleagues [178], moderate to severe acute histological CA was found to elevate the risk of structural brain anomalies also in the cerebellum as seen on MRI, both directly and by prompting premature birth.
In mice, one model to induce CA is ureaplasma-induced perinatal inflammation at E13.5, which induces a significant decrease in calbindin-positive neurons as well as a moderate decrease in myelin basic protein (MBP) in neocortex myelinization [179]. Another mouse model for CA makes use of inflammation-induced encephalopathy of prematurity driven by systemic administration of pro-inflammatory IL-1β. This model has been shown to interfere with the physiological roles of microglia in the brain, in particular that of the cerebellum; indeed, inducing inflammatory activation with this approach results in perturbed oligodendrocyte development and myelination in whole cerebellum lysates used for Western blotting[180]. IL-1β administration during the first postnatal week, a timing equivalent to the last trimester for brain development in humans, leads to specific reductions in gray and white matter volumes of the mouse cerebellar lobules, most specifically lobules I and II, and the nucleus interpositus from the second postnatal week onwards. These volume changes, which can be detected with MRI as of the second week, are preceded by reduced proliferation of OLIG2+ cells as well as reduced levels of MBP and myelin-associated glycoprotein (MAG). Moreover, the density of IBA1+ cerebellar microglia is increased both during the first postnatal week and P45, with evidence for increased microglial proliferation during the first two weeks postnatal. CA also induces a significant enrichment of pro-inflammatory markers in microglia from cerebellum and cerebrum, with the cerebellar microglia displaying a unique type I interferon signalling dysregulation. In summary, perinatal inflammation driven by systemic IL-1β leads to cerebellar volume deficits, especially in lobules I and II but also other lobules, which likely reflect oligodendrocyte pathology downstream of microglial activation [180].
7. Vulnerable and critical periods in cerebellar development affected by intrauterine insults
Intrauterine and postnatal insults can lead to a multitude of cerebellar deficiencies (Figure 4), which in turn can disrupt the maturation of neocortical and subcortical structures causing lifetime neurological motor and cognitive disabilities. Here we provide a perspective on the potential critical periods for healthy cerebellar development during gestation, highlighting opportunities to prevent risks of perinatal brain injury considering rodent and human research.
In human and rodent studies on developmental disorders caused by insults, the cerebellum is still a relatively neglected brain region and thus the effects of insults on cerebellar development are underestimated. It has for example been shown that stress during the second and third gestational period leads to changes in maternal care later in life as well as epigenetic variations [181], suggesting affecting also cerebellar functionality, but what this explicitly means for cerebellar development still needs to be determined.
When the impact of insults on cerebellar development is analysed, the focus lies often on PCs and GCs, since they form the sole output neurons of the cerebellar cortex and most of all neurons in the brain, respectively. This does not mean that the other neurons are less important when it comes to the functionality of the cerebellum. Depending on the time point, duration, and severity of the insult different subareas, layers, and neuron types are affected (Figure 3 and Figure 4). The critical periods are extended over the whole gestational period until 3-4 weeks postnatally in rodents and 3 years in human. Most likely GABAergic interneuron proliferation is affected slightly later during gestation compared to excitatory CN neurons but there are hardly any studies looking at the effects on both CN neurons and interneurons. CN neurons will be important to study with respect to insults since excitatory CNs develop first within the cerebellum, they are the output neurons of the cerebellum and they are the relay structure of the whole cerebellar cortex, being of major importance for cerebellar-thalamo-cortical communication [182,183,184,185,186].
When interpreting results of our review, it is important to consider three key factors. First, it must be mentioned that all information provided in this review is dependent on study design chosen by the investigators. Therefore, it is inevitable that there are differences in cerebellar modifications, such as cell type, and timeframes when comparing the studies within and between different intrauterine and postnatal insults. This means that information provided in this review must been seen as an overview; if different cell types are not mentioned to be affected by a specific intrauterine insult, this does not mean that they are unaffected. Additionally, in the cerebellum a single neuron type could cause a chain reaction of malfunctioning cells due to the aforementioned developmental dependencies.
Second, and as previously mentioned, there are differences in the development of the cerebellum as a whole and cerebellar subareas comparing humans and rodents. The folia complexity of human cerebellum is greater with enlarged hemispheres relative to the medial cerebellar vermis [22]. In human, foliation and growth of the cerebellum takes place during gestation, whereas in rodents the largest growth is postnatal. Besides the surface area is greater and the neuronal subtype ratios differ significantly between individual cerebellar areas [33,187]. Next to that, the developmental timeline of individual cell types is different with slight regional differences between areas (Figure 3 [35,41,188]).
Third, the fact that there are regional differences in the vulnerability of the cerebellum to intrauterine and postnatal insults shows that all studies discussed in this review should be interpreted with care and that where necessary the lobules and ideally microzones should be reinvestigated and seen as individual functional regions. In most studies, it is not obvious, whether researchers selected cerebellar areas to be analysed a priori to the data gathering. One cannot see the cerebellum as one structure when it comes to vulnerability and functional development (see e.g., [30]).
Below, we separately discuss the major mechanistic principles for the three prenatal trimesters in humans and discuss separately the first two postnatal weeks in rodents, where the development of the rodent cerebellum resembles the human cerebellum during the third gestational period.
7.1. First trimester (until week 13 human, until E12/13 rodents)
Even though cerebellar development of mammals starts during the first trimester of pregnancy, hardly any study in human or rodent focuses on the effects of intrauterine insults limited to this trimester. In humans PCs are born within this period (beginning of 7th gestational week, rodents between E10-E13.5) until the PC plate is formed (13th gestational week). Also, the eGL starts to develop (10-11th gestational week humans, E12.5-16.5 rodents), next to the large projection neurons and the GABAergic neurons of the CN. If insults affect the fetus during the first gestational period, these insults may be more severe compared to later during gestation, especially if they affect crucial genes or neurogenesis through mutagenesis or if the insults happen at very important key days, e.g., E9 in rodents. In rodents with genetic variations that affect the development of the cerebellar anlage, the mutation often leads to severe developmental problems, e.g.,[63,68]. If insults are not deadly, developmental compensations, as is often seen in rodent research, might also occur, e.g.,[189]. Stress has been shown to affect the nuclear size of PCs and GCs as well as the dendritic structure of CN neurons, if it occurs during the second (1st trimester) or third gestational week (2nd trimester) in rodents, respectively. Maternal alcohol and nicotine usage starting in the first week and ending in the last week of pregnancy mainly results in increased oxidative stress and neuroinflammation. Some intrauterine insults may be less harmful for the cerebellum during the very first weeks because of developmental compensations. Furthermore, some of the here discussed insults and consequences started during the first gestational period in humans, but effects cannot be isolated and assigned to this specific gestational period.
7.2. Second trimester (week 13-26 human, E13-birth rodents)
During the second trimester of pregnancy most cerebellar neuron types migrate, as well as UBCs and GABAergic interneurons develop and start migrating. IUGR and CA induced at E12.5 and E13.5 in rodents (like end of first or beginning second trimester in humans), respectively, both lead to adverse effects on myelination and the previous was also tested on synaptogenesis. CA furthermore has been shown to lead to a decrease in calbindin-positive neurons, which are PCs in the cerebellar cortex. If intrauterine insults lead to diminished PC, GC proliferation and migration, or Bergmann glia morphology, and/or neurogenesis one always has to keep in mind that the effects on one neuron type will most likely also affect the migration of the other cerebellar neurons [77,190]. Thereby insults lasting longer, such as often happens with CA and IUGR, lead to adverse effects that might be rather general with yet important implications. Maternal stress during only the third week of gestation affects the morphology of the PC dendrites. Alcohol administration during certain periods of the second trimester in rodents seems to be less harmful for PCs compared to other periods, although alcohol administration during both the first and second trimester does lead to changes in the cerebellum. Nicotine during the first and second trimester affects PC survival.
To summarize, in rodents during the last weeks before being born, mainly Bergmann glia, PC, and GC development is affected by defects seen in the migration, morphology, myelinization, and synaptogenesis. To our current knowledge, no clear regional differences in the vulnerability of the cerebellum have been described when harmful insults occur exclusively during the second trimester.
7.3. Third trimester (week 27 - 40 human)
The development of the cerebellum during the third trimester in humans and the comparable developmental weeks in rodents is different. Whereas the development of the cerebellum of rodents during the first two weeks after birth (P0-P12) resembles the cerebellum of a last trimester embryo in humans, the development of rodent cerebellum of course relies less on maternal risk factors since pups are already born. Also, the growth of the cerebellum occurs mainly after birth in rodents and less so in humans. Nevertheless, substance use such as alcohol and smoking, but also stress, IUGR as well as CA, especially during the last gestational period affect cerebellar development significantly. A recently uploaded preprint has shown that CA leads to PCs loss in non-human primates and a disrupted maturation of GCs, with the PC loss being accompanied by decreased shh signalling from PCs to GC [191]. CA furthermore accelerated pre-oligodendrocyte maturation into myelinating oligodendrocytes, which is not in line with rodent research but in line with increased expression of MBP in the cerebellum of CA-exposed fetuses. These findings are also consistent with reported histopathological findings in individuals with autism and suggest a potential mechanism through which perinatal inflammation contributes to neurodevelopmental disorders in human.
7.4. Two weeks postnatal rodents
In rodents, the first and second week postnatally (last weeks of human gestation) might be the most vulnerable time period for the cerebellum with respect to substance use but also the period where protection mechanisms against certain insults have evolved that protect the embryo from being too vulnerable. Nicotine usage during these first two weeks affects both the GCs and PCs. Stress leads to changes in GC maturation, showing an increase in the GL. Stress during the first postnatal week of rodents leads to life-lasting increases in corticosterone levels but no changes in GC apoptosis, however, stress during the second week postnatal does lead to changes in GC apoptosis. Regarding alcohol, it seems to be the other way around: where administration during the first week after birth leads to both PC and GC loss and increased pro-inflammatory cytokine levels, administration during the second week does not. Also, the effects of CA on cerebellar development have been assessed. Microglia proliferation increases with infections, also leading to cerebellar volume decrease and CN functionality being affected.
To conclude, in rodents the first weeks postnatally are very vulnerable and lead to severe effects, however dependent on the insults there are certain critical periods where the cerebellum is more or less vulnerable. There are time periods in the rodents’ life where the cerebellum is less vulnerable to both stress and alcohol. Whether there is also such a period in human cerebellar development is unknown since alcohol and stress research often neglects the cerebellum as a brain area with developmental problems and in humans both stress and alcohol abuse are often not limited to one week if they occur during pregnancy.
7.5. Regional differences in how intrauterine insults effect cerebellar development
Regarding regional differences, PCs in lobules VI and VII were less vulnerable when it comes to alcohol exposure in the first week postnatally in rodents. This either means that these lobules are in general less affected by alcohol, or it means that these lobules develop slightly differently compared to the other lobules. Data suggest that different lobules and neuronal subtypes are affected differently depending on the insult happening, which may at least partially be explained by a difference in the developmental timeline between areas [22,184,188]. Whether the insult enters the cerebellum via the blood vessels, or the blood-brain barrier, will also lead to regional differences in vulnerability, solely due to the location with respect to the blood-brain barrier and blood vessels (Figure 1).
8. Conclusion
To conclude, the cerebellum is a brain region that develops early and grows rapidly during gestation and continues to mature until late postnatal life in humans and rodents. The cerebellum is still often neglected in developmental research studying intrauterine insults. If cerebellar development has been examined with respect to intrauterine insults, the cerebellum has often been considered as one homogenously developing brain region, which it is not. In humans, the third trimester (gestational period) is the critical period where the cerebellum is most vulnerable for intrauterine insults to affect its development. The critical periods for vulnerability in a rodent’s life are the first two to three postnatal weeks, dependent on the severity of the insult and the cerebellar area affected.
Author Contributions
Conceptualization, J.A.W.W., J.D., C.B.C.; Writing – Original Draft Preparation, J.A.W.W., J.D., C.B.C.; Writing – Review & Editing, J.A.W.W., J.D., C.I.D.Z., C.B.C.; Supervision, J.D., C.B.C.; Funding acquisition, J.D., C.I.D.Z., C.B.C.
Funding
This study was enabled by funding from the Netherlands Organization for Scientific Research (NWO STEM - VBT 2021 19224 and NWO 863.14.005, C.B.C; NWO-ALW 824.02.001, C.I.D.Z.), the Dutch Organization for Medical Sciences (ZonMW 91120067, C.I.D.Z.), Medical Neuro-Delta (MD 01092019-31082023, C.I.D.Z.), INTENSE LSH-NWO (TTW/00798883, C.I.D.Z.), ERC-adv (GA-294775, C.I.D.Z.) and ERC-POC (nrs. 737619 and 768914, C.I.D.Z.), the NIN Vriendenfonds for Albinism (C.I.D.Z.), the Dutch NWO Gravitation Program, and the Dutch Brain Interface Initiative (DBI2, C.I.D.Z.).
Institutional Review Board Statement
“Not applicable”
Acknowledgments
We would like to thank F.A.H. Ooms, the Cerebellar Coordination and Cognition group, the UMC Utrecht Brain Center, Division Woman and Baby, Department of Neonatology for their contribution to useful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dietrich, K.N.; Eskenazi, B.; Schantz, S.; Yolton, K.; Rauh, V.A.; Johnson, C.B.; Alkon, A.; Canfield, R.L.; Pessah, I.N.; Berman, R.F. Principles and Practices of Neurodevelopmental Assessment in Children: Lessons Learned from the Centers for Children’s Environmental Health and Disease Prevention Research. Environ Health Perspect 2005, 113, 1437–1446. [Google Scholar] [CrossRef]
- Koning, I. V; Dudink, J.; Groenenberg, I.A.L.; Willemsen, S.P.; Reiss, I.K.M.; Steegers-Theunissen, R.P.M. Prenatal Cerebellar Growth Trajectories and the Impact of Periconceptional Maternal and Fetal Factors. Hum Reprod 2017, 32, 1230–1237. [Google Scholar] [CrossRef]
- Mwaniki, M.K.; Atieno, M.; Lawn, J.E.; Newton, C.R. Long-Term Neurodevelopmental Outcomes after Intrauterine and Neonatal Insults: A Systematic Review. The Lancet 2012, 379, 445–452. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.J.; Meaney, M.J. Fetal Origins of Mental Health: The Developmental Origins of Health and Disease Hypothesis. American Journal of Psychiatry 2017, 174, 319–328. [Google Scholar] [CrossRef]
- van Essen, M.J.; Nayler, S.; Becker, E.B.E.; Jacob, J. Deconstructing Cerebellar Development Cell by Cell. PLoS Genet 2020, 16, e1008630. [Google Scholar] [CrossRef] [PubMed]
- Koning, I. V; Tielemans, M.J.; Hoebeek, F.E.; Ecury-Goossen, G.M.; Reiss, I.K.M.; Steegers-Theunissen, R.P.M.; Dudink, J. Impacts on Prenatal Development of the Human Cerebellum: A Systematic Review. J Matern Fetal Neonatal Med 2017, 30, 2461–2468. [Google Scholar] [CrossRef]
- Badura, A.; Verpeut, J.L.; Metzger, J.W.; Pereira, T.D.; Pisano, T.J.; Deverett, B.; Bakshinskaya, D.E.; Wang, S.S.-H. Normal Cognitive and Social Development Require Posterior Cerebellar Activity. Elife 2018, 7. [Google Scholar] [CrossRef]
- Bruchhage, M.M.K.; Bucci, M.-P.; Becker, E.B.E. Cerebellar Involvement in Autism and ADHD. Handb Clin Neurol 2018, 155, 61–72. [Google Scholar] [CrossRef]
- Sathyanesan, A.; Zhou, J.; Scafidi, J.; Heck, D.H.; Sillitoe, R. V; Gallo, V. Emerging Connections between Cerebellar Development, Behaviour and Complex Brain Disorders. Nat Rev Neurosci 2019, 20, 298–313. [Google Scholar] [CrossRef]
- Herculano-Houzel, S. The Human Brain in Numbers: A Linearly Scaled-up Primate Brain. Front Hum Neurosci 2009, 3. [Google Scholar] [CrossRef] [PubMed]
- Ross, W.D. The Works of Aristotle. Translated into English Metaphysica . Oxford: Clarendon Press; 1928.
- Manto, M.; Bower, J.M.; Conforto, A.B.; Delgado-García, J.M.; da Guarda, S.N.F.; Gerwig, M.; Habas, C.; Hagura, N.; Ivry, R.B.; Mariën, P.; et al. Consensus Paper: Roles of the Cerebellum in Motor Control—The Diversity of Ideas on Cerebellar Involvement in Movement. The Cerebellum 2012, 11, 457–487. [Google Scholar] [CrossRef]
- Schmahmann, J.D. Dysmetria of Thought: Clinical Consequences of Cerebellar Dysfunction on Cognition and Affect. Trends Cogn Sci 1998, 2, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.-H.; Kloth, A.D.; Badura, A. The Cerebellum, Sensitive Periods, and Autism. Neuron 2014, 83, 518–532. [Google Scholar] [CrossRef] [PubMed]
- Apps, R.; Hawkes, R. Cerebellar Cortical Organization: A One-Map Hypothesis. Nat Rev Neurosci 2009, 10, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Grodd, W.; Hülsmann, E.; Lotze, M.; Wildgruber, D.; Erb, M. Sensorimotor Mapping of the Human Cerebellum: FMRI Evidence of Somatotopic Organization. Hum Brain Mapp 2001, 13, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.M.; Strick, P.L. Cerebellar Loops with Motor Cortex and Prefrontal Cortex of a Nonhuman Primate. J Neurosci 2003, 23, 8432–8444. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. The Cerebellum and Cognition. Neurosci Lett 2019, 688, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Kansal, K.; Yang, Z.; Fishman, A.M.; Sair, H.I.; Ying, S.H.; Jedynak, B.M.; Prince, J.L.; Onyike, C.U. Structural Cerebellar Correlates of Cognitive and Motor Dysfunctions in Cerebellar Degeneration. Brain 2017, 140, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, E.D.; Cui, K.; Hernandez-Miranda, L.R. Regulation of Early Cerebellar Development. FEBS J 2023, 290, 2786–2804. [Google Scholar] [CrossRef]
- Martinez, S.; Andreu, A.; Mecklenburg, N.; Echevarria, D. Cellular and Molecular Basis of Cerebellar Development. Front Neuroanat 2013, 7. [Google Scholar] [CrossRef]
- Haldipur, P.; Millen, K.J.; Aldinger, K.A. Human Cerebellar Development and Transcriptomics: Implications for Neurodevelopmental Disorders. Annu Rev Neurosci 2022, 45, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Brochu, G.; Maler, L.; Hawkes, R. Zebrin II: A Polypeptide Antigen Expressed Selectively by Purkinje Cells Reveals Compartments in Rat and Fish Cerebellum. J Comp Neurol 1990, 291, 538–552. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.L.; Krueger-Naug, A.M.; Currie, R.W.; Hawkes, R. Constitutive Expression of the 25-KDa Heat Shock Protein Hsp25 Reveals Novel Parasagittal Bands of Purkinje Cells in the Adult Mouse Cerebellar Cortex. J Comp Neurol 2000, 416, 383–397. [Google Scholar] [CrossRef]
- Hashizume, M.; Miyazaki, T.; Sakimura, K.; Watanabe, M.; Kitamura, K.; Kano, M. Disruption of Cerebellar Microzonal Organization in GluD2 (GluRδ2) Knockout Mouse. Front Neural Circuits 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Sarna, J.R.; Marzban, H.; Watanabe, M.; Hawkes, R. Complementary Stripes of Phospholipase Cbeta3 and Cbeta4 Expression by Purkinje Cell Subsets in the Mouse Cerebellum. J Comp Neurol 2006, 496, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.; Hawkes, R. Zones and Stripes. In Essentials of Cerebellum and Cerebellar Disorders A Primer For Graduate Students Second Edition; 2023; pp. 99–106.
- Ament, S.A.; Cortes-Gutierrez, M.; Herb, B.R.; Mocci, E.; Colantuoni, C.; McCarthy, M.M. A Single-Cell Genomic Atlas for Maturation of the Human Cerebellum during Early Childhood. Sci Transl Med 2023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Lin, Z.; Voges, K.; Ju, C.; Gao, Z.; Bosman, L.W.; Ruigrok, T.J.; Hoebeek, F.E.; De Zeeuw, C.I.; Schonewille, M. Cerebellar Modules Operate at Different Frequencies. Elife 2014, 3, e02536. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, C.I. Bidirectional Learning in Upbound and Downbound Microzones of the Cerebellum. Nat Rev Neurosci 2021, 22, 92–110. [Google Scholar] [CrossRef]
- Wang, X.; Novello, M.; Gao, Z.; Ruigrok, T.J.H.; De Zeeuw, C.I. Input and Output Organization of the Mesodiencephalic Junction for Cerebro-cerebellar Communication. J Neurosci Res 2022, 100, 620–637. [Google Scholar] [CrossRef]
- De Zeeuw, C.I.; Hoebeek, F.E.; Bosman, L.W.J.; Schonewille, M.; Witter, L.; Koekkoek, S.K. Spatiotemporal Firing Patterns in the Cerebellum. Nat Rev Neurosci 2011, 12, 327–344. [Google Scholar] [CrossRef]
- Lange, W. Cell Number and Cell Density in the Cerebellar Cortex of Man and Some Other Mammals. Cell Tissue Res 1975, 157. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y. Signaling Pathways in Cerebellar Granule Cells Development. Am J Stem Cells 2019, 8, 1–6. [Google Scholar]
- Leto, K.; Arancillo, M.; Becker, E.B.E.; Buffo, A.; Chiang, C.; Ding, B.; Dobyns, W.B.; Dusart, I.; Haldipur, P.; Hatten, M.E.; et al. Consensus Paper: Cerebellar Development. Cerebellum 2015, 1–40. [Google Scholar] [CrossRef]
- Carletti, B.; Rossi, F. Neurogenesis in the Cerebellum. Neuroscientist 2008, 14, 91–100. [Google Scholar] [CrossRef]
- Hausmann, B.; Mangold, U.; Sievers, J.; Berry, M. Derivation of Cerebellar Golgi Neurons from the External Granular Layer: Evidence from Explantation of External Granule Cells in Vivo. J Comp Neurol 1985, 232, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Haldipur, P.; Bharti, U.; Alberti, C.; Sarkar, C.; Gulati, G.; Iyengar, S.; Gressens, P.; Mani, S. Preterm Delivery Disrupts the Developmental Program of the Cerebellum. PLoS One 2011, 6, e23449. [Google Scholar] [CrossRef]
- Workman, A.D.; Charvet, C.J.; Clancy, B.; Darlington, R.B.; Finlay, B.L. Modeling Transformations of Neurodevelopmental Sequences across Mammalian Species. The Journal of Neuroscience 2013, 33, 7368–7383. [Google Scholar] [CrossRef] [PubMed]
- Haldipur, P.; Dang, D.; Millen, K.J. Embryology. Handb Clin Neurol 2018, 154, 29–44. [Google Scholar] [CrossRef]
- Haldipur, P.; Aldinger, K.A.; Bernardo, S.; Deng, M.; Timms, A.E.; Overman, L.M.; Winter, C.; Lisgo, S.N.; Razavi, F.; Silvestri, E.; et al. Spatiotemporal Expansion of Primary Progenitor Zones in the Developing Human Cerebellum. Science 2019, 366, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Jukic, A.M.; Baird, D.D.; Weinberg, C.R.; McConnaughey, D.R.; Wilcox, A.J. Length of Human Pregnancy and Contributors to Its Natural Variation. Hum Reprod 2013, 28, 2848–2855. [Google Scholar] [CrossRef]
- Jackson, C.M.; Mukherjee, S.; Wilburn, A.N.; Cates, C.; Lewkowich, I.P.; Deshmukh, H.; Zacharias, W.J.; Chougnet, C.A. Pulmonary Consequences of Prenatal Inflammatory Exposures: Clinical Perspective and Review of Basic Immunological Mechanisms. Front Immunol 2020, 11, 1285. [Google Scholar] [CrossRef]
- Cappelletti, M.; Presicce, P.; Kallapur, S.G. Immunobiology of Acute Chorioamnionitis. Front Immunol 2020, 11, 649. [Google Scholar] [CrossRef]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain Development in Rodents and Humans: Identifying Benchmarks of Maturation and Vulnerability to Injury across Species. Prog Neurobiol 2013, 106–107, 1–16. [Google Scholar] [CrossRef]
- Limperopoulos, C.; Soul, J.S.; Haidar, H.; Huppi, P.S.; Bassan, H.; Warfield, S.K.; Robertson, R.L.; Moore, M.; Akins, P.; Volpe, J.J.; et al. Impaired Trophic Interactions Between the Cerebellum and the Cerebrum Among Preterm Infants. Pediatrics 2005, 116, 844–850. [Google Scholar] [CrossRef]
- Accogli, A.; Addour-Boudrahem, N.; Srour, M. Diagnostic Approach to Cerebellar Hypoplasia. The Cerebellum 2021, 20, 631–658. [Google Scholar] [CrossRef] [PubMed]
- Aldinger, K.A.; Thomson, Z.; Phelps, I.G.; Haldipur, P.; Deng, M.; Timms, A.E.; Hirano, M.; Santpere, G.; Roco, C.; Rosenberg, A.B.; et al. Spatial and Cell Type Transcriptional Landscape of Human Cerebellar Development. Nat Neurosci 2021, 24, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Joyner, A.L. Otx2 and Gbx2 Are Required for Refinement and Not Induction of Mid-Hindbrain Gene Expression. Development 2001, 128, 4979–4991. [Google Scholar] [CrossRef] [PubMed]
- Beckinghausen, J.; Sillitoe, R. V. Insights into Cerebellar Development and Connectivity. Neurosci Lett 2019, 688, 2–13. [Google Scholar] [CrossRef]
- Wingate, R.J.T. The Rhombic Lip and Early Cerebellar Development. Curr Opin Neurobiol 2001, 11, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Millet, S.; Campbell, K.; Epstein, D.J.; Losos, K.; Harris, E.; Joyner, A.L. A Role for Gbx2 in Repression of Otx2 and Positioning the Mid/Hindbrain Organizer. Nature 1999, 401, 161–164. [Google Scholar] [CrossRef]
- Broccoli, V.; Boncinelli, E.; Wurst, W. The Caudal Limit of Otx2 Expression Positions the Isthmic Organizer. Nature 1999, 401, 164–168. [Google Scholar] [CrossRef]
- Chizhikov, V. V.; Lindgren, A.G.; Mishima, Y.; Roberts, R.W.; Aldinger, K.A.; Miesegaes, G.R.; Currle, D.S.; Monuki, E.S.; Millen, K.J. Lmx1a Regulates Fates and Location of Cells Originating from the Cerebellar Rhombic Lip and Telencephalic Cortical Hem. Proceedings of the National Academy of Sciences 2010, 107, 10725–10730. [Google Scholar] [CrossRef]
- Corrales, J.D.; Rocco, G.L.; Blaess, S.; Guo, Q.; Joyner, A.L. Spatial Pattern of Sonic Hedgehog Signaling through Gli Genes during Cerebellum Development. Development 2004, 131, 5581–5590. [Google Scholar] [CrossRef]
- Belzunce, I.; Belmonte-Mateos, C.; Pujades, C. The Interplay of Atoh1 Genes in the Lower Rhombic Lip during Hindbrain Morphogenesis. PLoS One 2020, 15, e0228225. [Google Scholar] [CrossRef]
- French, C.R.; Seshadri, S.; Destefano, A.L.; Fornage, M.; Arnold, C.R.; Gage, P.J.; Skarie, J.M.; Dobyns, W.B.; Millen, K.J.; Liu, T.; et al. Mutation of FOXC1 and PITX2 Induces Cerebral Small-Vessel Disease. J Clin Invest 2014, 124, 4877–4881. [Google Scholar] [CrossRef]
- Aldinger, K.A.; Lehmann, O.J.; Hudgins, L.; Chizhikov, V. V; Bassuk, A.G.; Ades, L.C.; Krantz, I.D.; Dobyns, W.B.; Millen, K.J. FOXC1 Is Required for Normal Cerebellar Development and Is a Major Contributor to Chromosome 6p25.3 Dandy-Walker Malformation. Nat Genet 2009, 41, 1037–1042. [Google Scholar] [CrossRef]
- Aldinger, K.A.; Timms, A.E.; Thomson, Z.; Mirzaa, G.M.; Bennett, J.T.; Rosenberg, A.B.; Roco, C.M.; Hirano, M.; Abidi, F.; Haldipur, P.; et al. Redefining the Etiologic Landscape of Cerebellar Malformations. Am J Hum Genet 2019, 105, 606–615. [Google Scholar] [CrossRef]
- Thomas, K.R.; Musci, T.S.; Neumann, P.E.; Capecchi, M.R. Swaying Is a Mutant Allele of the Proto-Oncogene Wnt-1. Cell 1991, 67, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, C.; Ishibashi, M.; Shimizu, C.; Nakanishi, S.; Kageyama, R. A Mammalian Helix-Loop-Helix Factor Structurally Related to the Product of Drosophila Proneural Gene Atonal Is a Positive Transcriptional Regulator Expressed in the Developing Nervous System. Journal of Biological Chemistry 1995, 270, 8730–8738. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, M.; Nakamura, S.; Mori, K.; Kawauchi, T.; Terao, M.; Nishimura, Y. V.; Fukuda, A.; Fuse, T.; Matsuo, N.; Sone, M.; et al. Ptf1a, a BHLH Transcriptional Gene, Defines GABAergic Neuronal Fates in Cerebellum. Neuron 2005, 47, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Machold, R.; Fishell, G. Math1 Is Expressed in Temporally Discrete Pools of Cerebellar Rhombic-Lip Neural Progenitors. Neuron 2005, 48, 17–24. [Google Scholar] [CrossRef]
- Sekerková, G.; Ilijic, E.; Mugnaini, E. Time of Origin of Unipolar Brush Cells in the Rat Cerebellum as Observed by Prenatal Bromodeoxyuridine Labeling. Neuroscience 2004, 127, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Consalez, G.G.; Goldowitz, D.; Casoni, F.; Hawkes, R. Origins, Development, and Compartmentation of the Granule Cells of the Cerebellum. Front Neural Circuits 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Chizhikov, V.; Millen, K.J. Development and Malformations of the Cerebellum in Mice. Mol Genet Metab 2003, 80, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Komuro, H.; Yacubova, E.; Yacubova, E.; Rakic, P. Mode and Tempo of Tangential Cell Migration in the Cerebellar External Granular Layer. The Journal of Neuroscience 2001, 21, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Helms, A.W.; Gowan, K.; Abney, A.; Savage, T.; Johnson, J.E. Overexpression of MATH1 Disrupts the Coordination of Neural Differentiation in Cerebellum Development. Molecular and Cellular Neuroscience 2001, 17, 671–682. [Google Scholar] [CrossRef]
- Wizeman, J.W.; Guo, Q.; Wilion, E.M.; Li, J.Y. Specification of Diverse Cell Types during Early Neurogenesis of the Mouse Cerebellum. Elife 2019, 8. [Google Scholar] [CrossRef]
- Zordan, P.; Croci, L.; Hawkes, R.; Consalez, G.G. Comparative Analysis of Proneural Gene Expression in the Embryonic Cerebellum. Developmental Dynamics 2008, 237, 1726–1735. [Google Scholar] [CrossRef] [PubMed]
- Miterko, L.N.; Sillitoe, R. V.; Hawkes, R. Zones and Stripes: Development of Cerebellar Topography. In Handbook of the Cerebellum and Cerebellar Disorders: Second Edition: Volume 3; 2021; Vol. 3.
- Sheldon, M.; Rice, D.S.; D’Arcangelo, G.; Yoneshima, H.; Nakajima, K.; Mikoshiba, K.; Howell, B.W.; Cooper, J.A.; Goldowitz, D.; Curran, T. Scrambler and Yotari Disrupt the Disabled Gene and Produce a Reeler -like Phenotype in Mice. Nature 1997, 389, 730–733. [Google Scholar] [CrossRef]
- Howell, B.W.; Hawkes, R.; Soriano, P.; Cooper, J.A. Neuronal Position in the Developing Brain Is Regulated by Mouse Disabled-1. Nature 1997, 389, 733–737. [Google Scholar] [CrossRef]
- D’Arcangelo, G.; G. Miao, G.; Chen, S.-C.; Scares, H.D.; Morgan, J.I.; Curran, T. A Protein Related to Extracellular Matrix Proteins Deleted in the Mouse Mutant Reeler. Nature 1995, 374, 719–723. [Google Scholar] [CrossRef]
- Chung, S.-H.; Marzban, H.; Croci, L.; Consalez, G.G.; Hawkes, R. Purkinje Cell Subtype Specification in the Cerebellar Cortex: Early B-Cell Factor 2 Acts to Repress the Zebrin II-Positive Purkinje Cell Phenotype. Neuroscience 2008, 153, 721–732. [Google Scholar] [CrossRef]
- Zhang, J.; Tran-Anh, K.; Hirata, T.; Sugihara, I. Striped Distribution Pattern of Purkinje Cells of Different Birthdates in the Mouse Cerebellar Cortex Studied with the Neurog2-CreER Transgenic Line. Neuroscience 2021, 462, 122–140. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, M.E.; Lackey, E.P.; Perez, R.; Ișleyen, F.S.; Brown, A.M.; Donofrio, S.G.; Lin, T.; Zoghbi, H.Y.; Sillitoe, R. V Maturation of Purkinje Cell Firing Properties Relies on Neurogenesis of Excitatory Neurons. Elife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- McKay, B.E.; Turner, R.W. Physiological and Morphological Development of the Rat Cerebellar Purkinje Cell. J Physiol 2005, 567, 829–850. [Google Scholar] [CrossRef]
- Beekhof, G.C.; Schonewille, M. Lobule-Related Action Potential Shape- and History-Dependent Current Integration in Purkinje Cells of Adult and Developing Mice. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Kapfhammer, J.P. Cellular and Molecular Control of Dendritic Growth and Development of Cerebellar Purkinje Cells. Prog Histochem Cytochem 2004, 39, 131–182. [Google Scholar] [CrossRef]
- Sarna, J.R.; Hawkes, R. Patterned Purkinje Cell Death in the Cerebellum. Prog Neurobiol 2003, 70, 473–507. [Google Scholar] [CrossRef]
- Busch, S.E.; Hansel, C. Climbing Fiber Multi-Innervation of Mouse Purkinje Dendrites with Arborization Common to Human. Science (1979) 2023, 381, 420–427. [Google Scholar] [CrossRef]
- Sultan, F.; Czubayko, U.; Thier, P. Morphological Classification of the Rat Lateral Cerebellar Nuclear Neurons by Principal Component Analysis. Journal of Comparative Neurology 2003, 455, 139–155. [Google Scholar] [CrossRef]
- Leto, K.; Carletti, B.; Williams, I.M.; Magrassi, L.; Rossi, F. Different Types of Cerebellar GABAergic Interneurons Originate from a Common Pool of Multipotent Progenitor Cells. The Journal of Neuroscience 2006, 26, 11682–11694. [Google Scholar] [CrossRef]
- Leto, K.; Bartolini, A.; Yanagawa, Y.; Obata, K.; Magrassi, L.; Schilling, K.; Rossi, F. Laminar Fate and Phenotype Specification of Cerebellar GABAergic Interneurons. The Journal of Neuroscience 2009, 29, 7079–7091. [Google Scholar] [CrossRef]
- Grimaldi, P.; Parras, C.; Guillemot, F.; Rossi, F.; Wassef, M. Origins and Control of the Differentiation of Inhibitory Interneurons and Glia in the Cerebellum. Dev Biol 2009, 328, 422–433. [Google Scholar] [CrossRef]
- Chedotal, A.; Sotelo, C. Early Development of Olivocerebellar Projections in the Fetal Rat Using CGRP Immunocytochemistry. European Journal of Neuroscience 1992, 4, 1159–1179. [Google Scholar] [CrossRef]
- Sugihara, I. Microzonal Projection and Climbing Fiber Remodeling in Single Olivocerebellar Axons of Newborn Rats at Postnatal Days 4–7. Journal of Comparative Neurology 2005, 487, 93–106. [Google Scholar] [CrossRef]
- Rodriguez, C.I.; Dymecki, S.M. Origin of the Precerebellar System. Neuron 2000, 27, 475–486. [Google Scholar] [CrossRef]
- Kim, T.; Park, H.; Tanaka-Yamamoto, K.; Yamamoto, Y. Developmental Timing-Dependent Organization of Synaptic Connections between Mossy Fibers and Granule Cells in the Cerebellum. Commun Biol 2023, 6, 446. [Google Scholar] [CrossRef] [PubMed]
- Goodlett, C.R.; Marcussen, L.; West, J.R. A Single Day of Alcohol Exposure During the Brain Growth Spurt Induces Brain Weight Restriction and Cerebellar Purkinje Cell Loss. Alcohol 1989, 7, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sim, M.E.; Lyoo, I.K.; Streeter, C.C.; Covell, J.; Sarid-Segal, O.; Ciraulo, D.A.; Kim, M.J.; Kaufman, M.J.; Yurgelun-Todd, D.A.; Renshaw, P.F. Cerebellar Gray Matter Volume Correlates with Duration of Cocaine Use in Cocaine-Dependent Subjects. Neuropsychopharmacology 2007, 32, 2229–2237. [Google Scholar] [CrossRef] [PubMed]
- Manto, M.; Perrotta, G. Toxic-Induced Cerebellar Syndrome: From the Fetal Period to the Elderly. In; 2018; pp. 333–352.
- Dow-Edwards, D.L.; Benveniste, H.; Behnke, M.; Bandstra, E.S.; Singer, L.T.; Hurd, Y.L.; Stanford, L.R. Neuroimaging of Prenatal Drug Exposure. Neurotoxicol Teratol 2006, 28, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Falk, L.; Nordberg, A.; Seiger, Å.; Kjældgaard, A.; Hellström-Lindahl, E. Smoking during Early Pregnancy Affects the Expression Pattern of Both Nicotinic and Muscarinic Acetylcholine Receptors in Human First Trimester Brainstem and Cerebellum. Neuroscience 2005, 132, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Lavezzi, A.M.; Corna, M.F.; Repetti, M.L.; Matturri, L. Cerebellar Purkinje Cell Vulnerability to Prenatal Nicotine Exposure in Sudden Unexplained Perinatal Death. Folia Neuropathol 2013, 4, 290–301. [Google Scholar] [CrossRef]
- Gosdin, L.K.; Deputy, N.P.; Kim, S.Y.; Dang, E.P.; Denny, C.H. Alcohol Consumption and Binge Drinking During Pregnancy Among Adults Aged 18–49 Years — United States, 2018–2020. MMWR Morb Mortal Wkly Rep 2022, 71, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Dukes, K.; Tripp, T.; Willinger, M.; Odendaal, H.; Elliott, A.J.; Kinney, H.C.; Robinson, F.; Petersen, J.M.; Raffo, C.; Hereld, D.; et al. Drinking and Smoking Patterns during Pregnancy: Development of Group-Based Trajectories in the Safe Passage Study. Alcohol 2017, 62, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Handmaker, N.S.; Rayburn, W.F.; Meng, C.; Bell, J.B.; Rayburn, B.B.; Rappaport, V.J. Impact of Alcohol Exposure After Pregnancy Recognition on Ultrasonographic Fetal Growth Measures. Alcohol Clin Exp Res 2006, 30, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Astley, S.J.; Aylward, E.H.; Olson, H.C.; Kerns, K.; Brooks, A.; Coggins, T.E.; Davies, J.; Dorn, S.; Gendler, B.; Jirikowic, T.; et al. Magnetic Resonance Imaging Outcomes From a Comprehensive Magnetic Resonance Study of Children With Fetal Alcohol Spectrum Disorders. Alcohol Clin Exp Res 2009, 33, 1671–1689. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.L.; Crocker, N.; Mattson, S.N.; Riley, E.P. Neuroimaging and Fetal Alcohol Spectrum Disorders. Dev Disabil Res Rev 2009, 15, 209–217. [Google Scholar] [CrossRef]
- West, J. Acute and Long-Term Changes in the Cerebellum Following Developmental Exposure to Ethanol. Alcohol Alcohol Suppl. 1993, 2, 199–202. [Google Scholar]
- Sowell, E.R.; Jernigan, T.L.; Mattson, S.N.; Riley, E.P.; Sobel, D.F.; Jones, K.L. Abnormal Development of the Cerebellar Vermis in Children Prenatally Exposed to Alcohol: Size Reduction in Lobules I-V. Alcohol Clin Exp Res 1996, 20, 31–34. [Google Scholar] [CrossRef]
- Mattson, S.N.; Riley, E.P.; Jernigan, T.L.; Garcia, A.; Kaneko, W.M.; Ehlers, C.L.; Jones, K.L. A Decrease in the Size of the Basal Ganglia in Children with Fetal Alcohol Syndrome. Neurotoxicol Teratol 1994, 16, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Kfir, M.; Yevtushok, L.; Onishchenko, S.; Wertelecki, W.; Bakhireva, L.; Chambers, C.D.; Jones, K.L.; Hull, A.D. Can Prenatal Ultrasound Detect the Effects of In-utero Alcohol Exposure? A Pilot Study. Ultrasound in Obstetrics & Gynecology 2009, 33, 683–689. [Google Scholar] [CrossRef]
- Autti-Rämö, I.; Autti, T.; Korkman, M.; Kettunen, S.; Salonen, O.; Valanne, L. MRI Findings in Children with School Problems Who Had Been Exposed Prenatally to Alcohol. Dev Med Child Neurol 2002, 44, 98. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.P.; McGee, C.L. Fetal Alcohol Spectrum Disorders: An Overview with Emphasis on Changes in Brain and Behavior. Exp Biol Med 2005, 230, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Jaatinen, P.; Rintala, J. Mechanisms of Ethanol-Induced Degeneration in the Developing, Mature, and Aging Cerebellum. Cerebellum 2008, 7, 332–347. [Google Scholar] [CrossRef] [PubMed]
- O’Hare, E.D.; Kan, E.; Yoshii, J.; Mattson, S.N.; Riley, E.P.; Thompson, P.M.; Toga, A.W.; Sowell, E.R. Mapping Cerebellar Vermal Morphology and Cognitive Correlates in Prenatal Alcohol Exposure. Neuroreport 2005, 16, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-J.A.; Parnell, S.E.; West, J.R.; Chen, W.-J.A.; Parnell, S.E.; Neonatal, J.R.W. Neonatal Alcohol and Nicotine Exposure Limits Brain Growth and Depletes Cerebellar Purkinje Cells; 1998; Vol. 15.
- Karaçay, B.; Li, S.; Bonthius, D.J. Maturation-Dependent Alcohol Resistance in the Developing Mouse: Cerebellar Neuronal Loss and Gene Expression during Alcohol-Vulnerable and -Resistant Periods. Alcohol Clin Exp Res 2008, 32, 1439–1450. [Google Scholar] [CrossRef]
- Lee, Y.; Rowe, J.; Eskue, K.; West, J.R.; Maier, S.E. Alcohol Exposure on Postnatal Day 5 Induces Purkinje Cell Loss and Evidence of Purkinje Cell Degradation in Lobule I of Rat Cerebellum. Alcohol 2008, 42, 295–302. [Google Scholar] [CrossRef]
- Topper, L.A.; Baculis, B.C.; Valenzuela, C.F. Exposure of Neonatal Rats to Alcohol Has Differential Effects on Neuroinflammation and Neuronal Survival in the Cerebellum and Hippocampus. J Neuroinflammation 2015, 12. [Google Scholar] [CrossRef]
- Goodlett, C.R.; Eilers, A.T. Alcohol-Induced Purkinje Cell Loss with a Single Binge Exposure in Neonatal Rats: A Stereological Study of Temporal Windows of Vulnerability. Alcohol Clin Exp Res 1997, 21, 738–744. [Google Scholar] [CrossRef]
- Hamre, K.M.; West, J.R. The Effects of the Timing of Ethanol Exposure during the Brain Growth Spurt on the Number of Cerebellar Purkinje and Granule Cell Nuclear Profiles; 1993; Vol. 17.
- Bhattacharya, D.; Majrashi, M.; Ramesh, S.; Govindarajulu, M.; Bloemer, J.; Fujihashi, A.; Crump, B.R.; Hightower, H.; Bhattacharya, S.; Moore, T.; et al. Assessment of the Cerebellar Neurotoxic Effects of Nicotine in Prenatal Alcohol Exposure in Rats. Life Sci 2018, 194, 177–184. [Google Scholar] [CrossRef]
- González-Burgos, I.; Alejandre-Gómez, M. Cerebellar Granule Cell and Bergmann Glial Cell Maturation in the Rat Is Disrupted by Pre- and Post-natal Exposure to Moderate Levels of Ethanol. International Journal of Developmental Neuroscience 2005, 23, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Nirgudkar, P.; Taylor, D.H.; Yanagawa, Y.; Valenzuela, C.F. Ethanol Exposure during Development Reduces GABAergic/Glycinergic Neuron Numbers and Lobule Volumes in the Mouse Cerebellar Vermis. Neurosci Lett 2016, 632, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Ekblad, M.; Korkeila, J.; Parkkola, R.; Lapinleimu, H.; Haataja, L.; Lehtonen, L. Maternal Smoking during Pregnancy and Regional Brain Volumes in Preterm Infants. Journal of Pediatrics 2010, 156. [Google Scholar] [CrossRef] [PubMed]
- Ekblad, M.O. Association of Smoking During Pregnancy With Compromised Brain Development in Offspring. JAMA Netw Open 2022, 5, e2224714. [Google Scholar] [CrossRef]
- Bhandari, N.R.; Day, K.D.; Payakachat, N.; Franks, A.M.; McCain, K.R.; Ragland, D. Use and Risk Perception of Electronic Nicotine Delivery Systems and Tobacco in Pregnancy. Women’s Health Issues 2018, 28, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Stroud, L.R.; Papandonatos, G.D.; Borba, K.; Kehoe, T.; Scott-Sheldon, L.A.J. Flavored Electronic Cigarette Use, Preferences, and Perceptions in Pregnant Mothers: A Correspondence Analysis Approach. Addictive Behaviors 2019, 91, 21–29. [Google Scholar] [CrossRef]
- Banerjee, S.; Deacon, A.; Suter, M.A.; Aagaard, K.M. Understanding the Placental Biology of Tobacco Smoke, Nicotine, and Marijuana (THC) Exposures During Pregnancy. Clin Obstet Gynecol 2022, 65, 347–359. [Google Scholar] [CrossRef]
- Kim, C.W.; Lee, S.M.; Ko, E.B.; Go, R.E.; Jeung, E.B.; Kim, M.S.; Choi, K.C. Inhibitory Effects of Cigarette Smoke Extracts on Neural Differentiation of Mouse Embryonic Stem Cells. Reproductive Toxicology 2020, 95, 75–85. [Google Scholar] [CrossRef]
- Abdel-Rahman, A.; Dechkovskaia, A.M.; Sutton, J.M.; Chen, W.C.; Guan, X.; Khan, W.A.; Abou-Donia, M.B. Maternal Exposure of Rats to Nicotine via Infusion during Gestation Produces Neurobehavioral Deficits and Elevated Expression of Glial Fibrillary Acidic Protein in the Cerebellum and CA1 Subfield in the Offspring at Puberty. Toxicology 2005, 209, 245–261. [Google Scholar] [CrossRef]
- Abou-Donia, M.B.; Khan, W.A.; Dechkovskaia, A.M.; Goldstein, L.B.; Bullman, S.L.; Abdel-Rahman, A. In Utero Exposure to Nicotine and Chlorpyrifos Alone, and in Combination Produces Persistent Sensorimotor Deficits and Purkinje Neuron Loss in the Cerebellum of Adult Offspring Rats. Arch Toxicol 2006, 80, 620–631. [Google Scholar] [CrossRef]
- Chan, Y.L.; Saad, S.; Pollock, C.; Oliver, B.; Al-Odat, I.; Zaky, A.A.; Jones, N.; Chen, H. Impact of Maternal Cigarette Smoke Exposure on Brain Inflammation and Oxidative Stress in Male Mice Offspring. Sci Rep 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Al-Amri, I.S.; Kadim, I.T.; Alkindi, A.; Khalaf, S.K.; Hamaed, A.; Al-Yaqoobi, S.S.; Al-Harrasi, A.S.; Al-Hashmi, S.A.; Al-Maasbi, A.A.; Al-Harthy, R.S.; et al. Effects of Prenatal Tobacco Smoke on Cerebellum Histomorphological Changes at Critical Developmental Stages in CD-1 Mice. International Journal of Morphology 2021, 39, 318–326. [Google Scholar] [CrossRef]
- Huang, L.Z.; Abbott, L.C.; Winzer-Serhan, U.H. Effects of Chronic Neonatal Nicotine Exposure on Nicotinic Acetylcholine Receptor Binding, Cell Death and Morphology in Hippocampus and Cerebellum. Neuroscience 2007, 146, 1854–1868. [Google Scholar] [CrossRef] [PubMed]
- Church, J.S.; Chace-Donahue, F.; Blum, J.L.; Ratner, J.R.; Zelikoff, J.T.; Schwartzer, J.J. Neuroinflammatory and Behavioral Outcomes Measured in Adult Offspring of Mice Exposed Prenatally to E-Cigarette Aerosols. Environ Health Perspect 2020, 128. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of Stress throughout the Lifespan on the Brain, Behaviour and Cognition. Nat Rev Neurosci 2009, 10, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Adamson, B.; Letourneau, N.; Lebel, C. Prenatal Maternal Anxiety and Children’s Brain Structure and Function: A Systematic Review of Neuroimaging Studies. J Affect Disord 2018, 241, 117–126. [Google Scholar] [CrossRef]
- Dunkel Schetter, C.; Tanner, L. Anxiety, Depression and Stress in Pregnancy. Curr Opin Psychiatry 2012, 25, 141–148. [Google Scholar] [CrossRef]
- Buss, C.; Davis, E.P.; Muftuler, L.T.; Head, K.; Sandman, C.A. High Pregnancy Anxiety during Mid-Gestation Is Associated with Decreased Gray Matter Density in 6–9-Year-Old Children. Psychoneuroendocrinology 2010, 35, 141–153. [Google Scholar] [CrossRef]
- Ulupinar, E.; Yucel, F. Prenatal Stress Reduces Interneuronal Connectivity in the Rat Cerebellar Granular Layer. In Proceedings of the Neurotoxicology and Teratology; May 2005; Vol. 27; pp. 475–484. [Google Scholar]
- Ulupinar, E.; Yucel, F.; Ortug, G. The Effects of Prenatal Stress on the Purkinje Cell Neurogenesis. Neurotoxicol Teratol 2006, 28, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Pascual, R.; Ebner, D.; Araneda, R.; Urqueta, M.J.; Bustamante, C. Maternal Stress Induces Long-Lasting Purkinje Cell Developmental Impairments in Mouse Offspring. Eur J Pediatr 2010, 169, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Pascual, R.; Valencia, M.; Bustamante, C. Purkinje Cell Dendritic Atrophy Induced by Prenatal Stress Is Mitigated by Early Environmental Enrichment. Neuropediatrics 2015, 46, 37–43. [Google Scholar] [CrossRef]
- Pascual, R.; Santander, O.; Cuevas, I.; Valencia, M. Prenatal Glucocorticoid Administration Persistently Increased the Immunohistochemical Expression of Type-1 Metabotropic Glutamate Receptor and Purkinje Cell Dendritic Growth in the Cerebellar Cortex of the Rat. Rom J Morphol Embryol 2017, 58, 67–72. [Google Scholar]
- Wilber, A.A.; Lin, G.L.; Wellman, C.L. Neonatal Corticosterone Administration Impairs Adult Eyeblink Conditioning and Decreases Glucocorticoid Receptor Expression in the Cerebellar Interpositus Nucleus. Neuroscience 2011, 177, 56–65. [Google Scholar] [CrossRef]
- Schneider, E.R.; Civillico, E.F.; Wang, S.S.-H. Calcium-Based Dendritic Excitability and Its Regulation in the Deep Cerebellar Nuclei. J Neurophysiol 2013, 109, 2282–2292. [Google Scholar] [CrossRef]
- Roque, A.; Lajud, N.; Valdez, J.J.; Torner, L. Early-Life Stress Increases Granule Cell Density in the Cerebellum of Male Rats. Brain Res 2019, 1723. [Google Scholar] [CrossRef]
- Zhang, L.-X.; Levine, S.; Dent, G.; Zhan, Y.; Xing, G.; Okimoto, D.; Gordon, M.K.; Post, R.M.; Smith, M.A. Maternal Deprivation Increases Cell Death in the Infant Rat Brain; 2002; Vol. 133.
- Schmidt, M.; Enthoven, L.; Van Der Mark, M.; Levine, S.; De Kloet, E.R.; Oitzl, M.S. The Postnatal Development of the Hypothalamic-Pituitary-Adrenal Axis in the Mouse. International Journal of Developmental Neuroscience 2003, 21, 125–132. [Google Scholar] [CrossRef]
- Sapolsky’, R.M.; Meaney’, M.J. Maturation of the Adrenocortical Stress Response: Neuroendocrine Control Mechanisms and the Stress Hyporesponsive Period; 1986; Vol. 11.
- Noguchi, K.K.; Walls, K.C.; Wozniak, D.F.; Olney, J.W.; Roth, K.A.; Farber, N.B. Acute Neonatal Glucocorticoid Exposure Produces Selective and Rapid Cerebellar Neural Progenitor Cell Apoptotic Death. Cell Death Differ 2008, 15, 1582–1592. [Google Scholar] [CrossRef]
- Schmidt, M. V. Stress-Hyporesponsive Period. In Stress: Physiology, Biochemistry, and Pathology Handbook of Stress Series, Volume 3; Elsevier, 2019; pp. 49–56 ISBN 9780128131466.
- Blumberg, M.; Dooley, J.; Sokoloff, G. The Developing Brain Revealed during Sleep. Curr Opin Physiol 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Canto, C.B.; Bauer, S.; Hoogland, T.M.; Hoedemaker, H.H.; Geelen, C.; Loyola, S.; Miaja, P.; Zeeuw, C.I. De Sleep-State Dependent Cerebellar Processing in Adult Mice. bioRxiv 2023, 2023.10.30.564769, doi:10.1101/2023.10.30.564769. [CrossRef]
- Li, W.; Ma, L.; Yang, G.; Gan, W.-B. REM Sleep Selectively Prunes and Maintains New Synapses in Development and Learning. Nat Neurosci 2017, 20, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Dooley, J.C.; Sokoloff, G.; Blumberg, M.S. Movements during Sleep Reveal the Developmental Emergence of a Cerebellar-Dependent Internal Model in Motor Thalamus. Curr Biol 2021, 31, 5501–5511. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Sokoloff, G.; Blumberg, M.S. Corollary Discharge in Precerebellar Nuclei of Sleeping Infant Rats. Elife 2018, 7. [Google Scholar] [CrossRef]
- Crapse, T.B.; Sommer, M.A. Corollary Discharge across the Animal Kingdom. Nat Rev Neurosci 2008, 9, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Korovaichuk, A.; Astiz, M.; Schroeder, H.; Islam, R.; Barrenetxea, J.; Fischer, A.; Oster, H.; Bringmann, H. Functional Divergence of Mammalian TFAP2a and TFAP2b Transcription Factors for Bidirectional Sleep Control. Genetics 2020, 216, 735–752. [Google Scholar] [CrossRef]
- Pardo, G.V.E.; Goularte, J.F.; Hoefel, A.L.; de Castro, A.L.; Kucharski, L.C.; da Rosa Araujo, A.S.; Lucion, A.B. Effects of Sleep Restriction during Pregnancy on the Mother and Fetuses in Rats. Physiol Behav 2016, 155, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Canto, C.B.; Onuki, Y.; Bruinsma, B.; van der Werf, Y.D.; De Zeeuw, C.I. The Sleeping Cerebellum. Trends Neurosci 2017, 40, 309–323. [Google Scholar] [CrossRef] [PubMed]
- De Zeeuw, C.I.; Canto, C.B. Sleep Deprivation Directly Following Eyeblink-Conditioning Impairs Memory Consolidation. Neurobiol Learn Mem 2020, 170, 107165. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, B.; Edmonds, C.J. School Age Neurological and Cognitive Outcomes of Fetal Growth Retardation or Small for Gestational Age Birth Weight. Front Endocrinol (Lausanne) 2019, 10, 186. [Google Scholar] [CrossRef]
- Hartkopf, J.; Schleger, F.; Keune, J.; Wiechers, C.; Pauluschke-Froehlich, J.; Weiss, M.; Conzelmann, A.; Brucker, S.; Preissl, H.; Kiefer-Schmidt, I. Impact of Intrauterine Growth Restriction on Cognitive and Motor Development at 2 Years of Age. Front Physiol 2018, 9, 1278. [Google Scholar] [CrossRef]
- Devaskar, S.U.; Chu, A. Intrauterine Growth Restriction: Hungry for an Answer. Physiology (Bethesda) 2016, 31, 131–146. [Google Scholar] [CrossRef]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The Consequences of Fetal Growth Restriction on Brain Structure and Neurodevelopmental Outcome. J Physiol 2016, 594, 807–823. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, W.; Liu, J. Neurodevelopment in Children with Intrauterine Growth Restriction: Adverse Effects and Interventions. J Matern Fetal Neonatal Med 2016, 29, 660–668. [Google Scholar] [CrossRef]
- Olivier, P.; Baud, O.; Bouslama, M.; Evrard, P.; Gressens, P.; Verney, C. Moderate Growth Restriction: Deleterious and Protective Effects on White Matter Damage. Neurobiol Dis 2007, 26, 253–263. [Google Scholar] [CrossRef]
- Volpe, J.J. Brain Injury in Premature Infants: A Complex Amalgam of Destructive and Developmental Disturbances. Lancet Neurol 2009, 8, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Eixarch, E.; Muñoz-Moreno, E.; Bargallo, N.; Batalle, D.; Gratacos, E. Motor and Cortico-Striatal-Thalamic Connectivity Alterations in Intrauterine Growth Restriction. Am J Obstet Gynecol 2016, 214, 725–e1. [Google Scholar] [CrossRef] [PubMed]
- Fischi-Gomez, E.; Muñoz-Moreno, E.; Vasung, L.; Griffa, A.; Borradori-Tolsa, C.; Monnier, M.; Lazeyras, F.; Thiran, J.-P.; Hüppi, P.S. Brain Network Characterization of High-Risk Preterm-Born School-Age Children. Neuroimage Clin 2016, 11, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.L.; Bashir, M.; Santiago, C.; Farrow, K.; Fung, C.; Brown, A.S.; Dettman, R.W.; Dizon, M.L.V. Intrauterine Growth Restriction and Hyperoxia as a Cause of White Matter Injury. Dev Neurosci 2018, 40, 344–357. [Google Scholar] [CrossRef] [PubMed]
- McDougall, A.R.A.; Wiradjaja, V.; Azhan, A.; Li, A.; Hale, N.; Wlodek, M.E.; Hooper, S.B.; Wallace, M.J.; Tolcos, M. Intrauterine Growth Restriction Alters the Postnatal Development of the Rat Cerebellum. Dev Neurosci 2017, 39, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Iskusnykh, I.Y.; Fattakhov, N.; Buddington, R.K.; Chizhikov, V. V Intrauterine Growth Restriction Compromises Cerebellar Development by Affecting Radial Migration of Granule Cells via the JamC/Pard3a Molecular Pathway. Exp Neurol 2021, 336, 113537. [Google Scholar] [CrossRef] [PubMed]
- Tolcos, M.; McDougall, A.; Shields, A.; Chung, Y.; O’Dowd, R.; Turnley, A.; Wallace, M.; Rees, S. Intrauterine Growth Restriction Affects Cerebellar Granule Cells in the Developing Guinea Pig Brain. Dev Neurosci 2018, 40, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Chanez, C.; Rabin, O.; Heroux, M.; Giguere, J.F. Cerebral Amino Acid Changes in an Animal Model of Intrauterine Growth Retardation. Metab Brain Dis 1993, 8, 61–72. [Google Scholar] [CrossRef]
- Tita, A.T.N.; Andrews, W.W. Diagnosis and Management of Clinical Chorioamnionitis. Clin Perinatol 2010, 37, 339–354. [Google Scholar] [CrossRef]
- Ylijoki, M.; Lehtonen, L.; Lind, A.; Ekholm, E.; Lapinleimu, H.; Kujari, H.; Haataja, L. Chorioamnionitis and Five-Year Neurodevelopmental Outcome in Preterm Infants. Neonatology 2016, 110, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Hendson, L.; Russell, L.; Robertson, C.M.T.; Liang, Y.; Chen, Y.; Abdalla, A.; Lacaze-Masmonteil, T. Neonatal and Neurodevelopmental Outcomes of Very Low Birth Weight Infants with Histologic Chorioamnionitis. J Pediatr 2011, 158, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Nasef, N.; Shabaan, A.E.; Schurr, P.; Iaboni, D.; Choudhury, J.; Church, P.; Dunn, M.S. Effect of Clinical and Histological Chorioamnionitis on the Outcome of Preterm Infants. Am J Perinatol 2013, 30, 59–68. [Google Scholar] [CrossRef]
- Pappas, A.; Kendrick, D.E.; Shankaran, S.; Stoll, B.J.; Bell, E.F.; Laptook, A.R.; Walsh, M.C.; Das, A.; Hale, E.C.; Newman, N.S.; et al. Chorioamnionitis and Early Childhood Outcomes among Extremely Low-Gestational-Age Neonates. JAMA Pediatr 2014, 168, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Limperopoulos, C.; Bassan, H.; Sullivan, N.R.; Soul, J.S.; Robertson, R.L.J.; Moore, M.; Ringer, S.A.; Volpe, J.J.; du Plessis, A.J. Positive Screening for Autism in Ex-Preterm Infants: Prevalence and Risk Factors. Pediatrics 2008, 121, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.G.; Willis, K.A.; Jobe, A.; Ambalavanan, N. Chorioamnionitis and Neonatal Outcomes. Pediatr Res 2022, 91, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Normann, E.; Lacaze-Masmonteil, T.; Eaton, F.; Schwendimann, L.; Gressens, P.; Thébaud, B. A Novel Mouse Model of Ureaplasma-Induced Perinatal Inflammation: Effects on Lung and Brain Injury. Pediatr Res 2009, 65, 430–436. [Google Scholar] [CrossRef]
- Klein, L.; Van Steenwinckel, J.; Fleiss, B.; Scheuer, T.; Bührer, C.; Faivre, V.; Lemoine, S.; Blugeon, C.; Schwendimann, L.; Csaba, Z.; et al. A Unique Cerebellar Pattern of Microglia Activation in a Mouse Model of Encephalopathy of Prematurity. Glia 2022, 70, 1699–1719. [Google Scholar] [CrossRef]
- Gudsnuk, K.M.A.; Champagne, F.A. Epigenetic Effects of Early Developmental Experiences. Clin Perinatol 2011, 38, 703–717. [Google Scholar] [CrossRef]
- Broersen, R.; Canto, C.B.; De Zeeuw, C.I. Cerebellar Nuclei: Associative Motor Learning in Zebrafish. Current Biology 2023, 33, R867–R870. [Google Scholar] [CrossRef] [PubMed]
- Canto, C.B.; Broersen, R.; De Zeeuw, C.I. Intrinsic Excitement in Cerebellar Nuclei Neurons during Learning. Proceedings of the National Academy of Sciences 2018, 115, 9824–9826. [Google Scholar] [CrossRef] [PubMed]
- Beekhof, G.C.; Gornati, S. V; Canto, C.B.; Libster, A.M.; Schonewille, M.; De Zeeuw, C.I.; Hoebeek, F.E. Activity of Cerebellar Nuclei Neurons Correlates with ZebrinII Identity of Their Purkinje Cell Afferents. Cells 2021, 10. [Google Scholar] [CrossRef]
- Witter, L.; Canto, C.B.; Hoogland, T.M.; de Gruijl, J.R.; De Zeeuw, C.I. Strength and Timing of Motor Responses Mediated by Rebound Firing in the Cerebellar Nuclei after Purkinje Cell Activation. Front Neural Circuits 2013, 7, 133. [Google Scholar] [CrossRef] [PubMed]
- Hoogland, T.M.; De Gruijl, J.R.; Witter, L.; Canto, C.B.; De Zeeuw, C.I. Role of Synchronous Activation of Cerebellar Purkinje Cell Ensembles in Multi-Joint Movement Control. Curr Biol 2015, 25, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Lange, W. Regional Differences in the Distribution of Golgi Cells in the Cerebellar Cortex of Man and Some Other Mammals. Cell Tissue Res 1974, 153, 219–226. [Google Scholar] [CrossRef]
- Beekhof, G.C.; Osório, C.; White, J.J.; van Zoomeren, S.; van der Stok, H.; Xiong, B.; Nettersheim, I.H.; Mak, W.A.; Runge, M.; Fiocchi, F.R.; et al. Differential Spatiotemporal Development of Purkinje Cell Populations and Cerebellum-Dependent Sensorimotor Behaviors. Elife 2021, 10. [Google Scholar] [CrossRef]
- Wojcinski, A.; Morabito, M.; Lawton, A.K.; Stephen, D.N.; Joyner, A.L. Genetic Deletion of Genes in the Cerebellar Rhombic Lip Lineage Can Stimulate Compensation through Adaptive Reprogramming of Ventricular Zone-Derived Progenitors. Neural Dev 2019, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- van der Heijden, M.E.; Sillitoe, R. V. Interactions Between Purkinje Cells and Granule Cells Coordinate the Development of Functional Cerebellar Circuits. Neuroscience 2021, 462, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.; Tong, X.; Tan, A.; Paiva, V.N. De; Presicce, P.; Kannan, P.S.; Williams, K.; Damianos, A.; Newsam, M.T.; Benny, M.; et al. Single-Nucleus RNA-Seq Reveals Disrupted Cell Maturation by Chorioamnionitis in the Preterm Cerebellum of Nonhuman Primates. bioRxiv 2022, 2022.07.12.499720. [CrossRef]
Figure 1.
Gross anatomy of the cerebellum. An unfolded view of the cerebellum showing medial (dark blue), intermediate (lighter blue) and lateral zones (light blue), which are interconnected with different parts of the cerebellar nuclei (indicated with the same color-coding on the left). The areas of the cortex served by the cerebellar arteries are indicated on the left. The image was adapted with courtesy and permission from I. Koning.
Figure 1.
Gross anatomy of the cerebellum. An unfolded view of the cerebellum showing medial (dark blue), intermediate (lighter blue) and lateral zones (light blue), which are interconnected with different parts of the cerebellar nuclei (indicated with the same color-coding on the left). The areas of the cortex served by the cerebellar arteries are indicated on the left. The image was adapted with courtesy and permission from I. Koning.
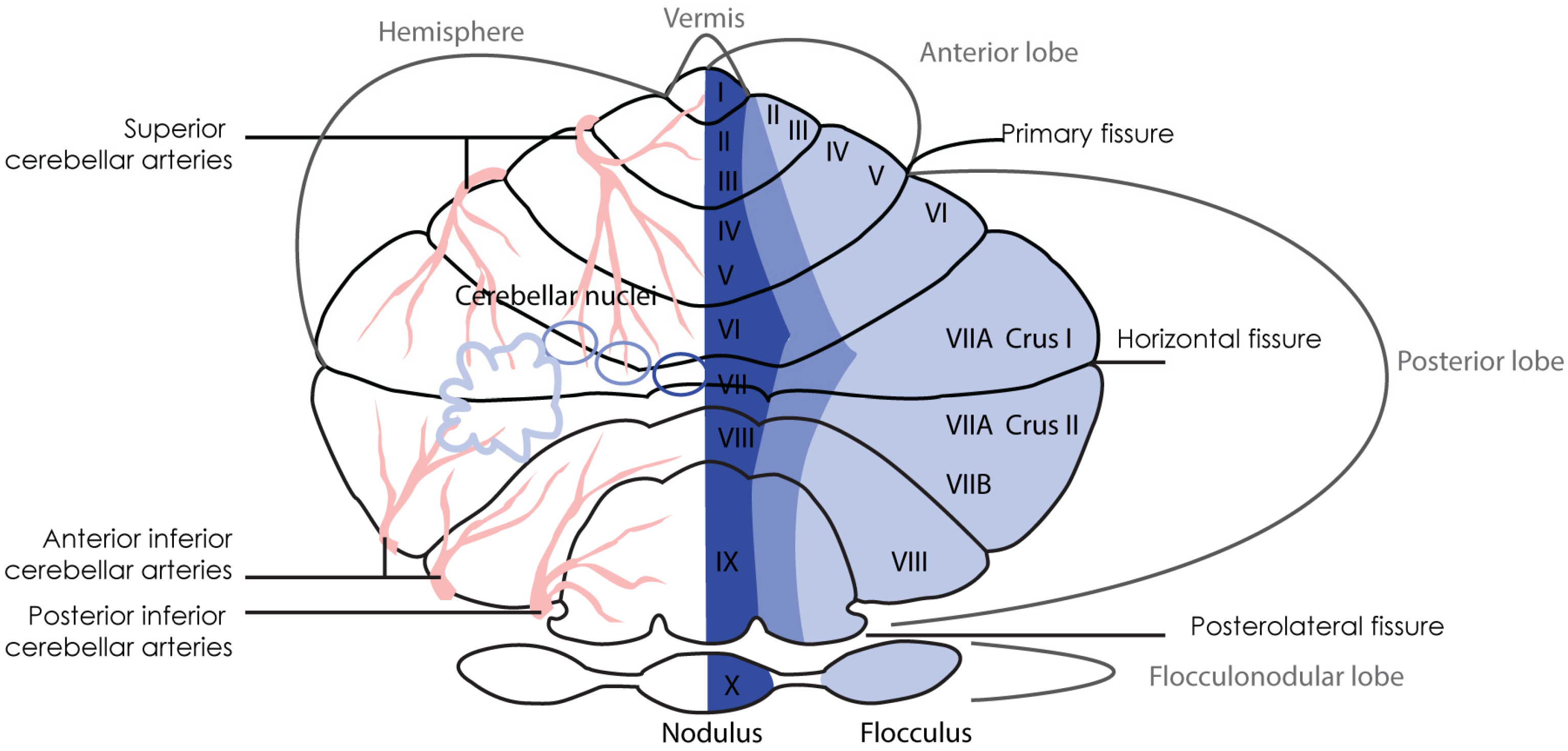
Figure 2.
Timeline of cerebellar development of humans (dark blue, top) and rodents (light blue, bottom). Top: Sagittal sections of the outline of the human cerebellum at 6 time points during development. Top and bottom: The duration and timing of different parts of cerebellar development are indicated with a color-coded bar with a distinct bar length. The text explains what is happening during that period (adapted from [6]). Bottom: Rodent cerebellar development during different time points. The first two outlines show the differentiation of the hind- and midbrain and the relative location of the cerebellar anlage [20]. Embryonic day (E)15.5, E18.5, postnatal ~(P)4 and P21 show the outlines of cerebellar midsagittal cuts (adapted from [21]). Arrowheads indicate the migration direction of individual neuron types. At ~P4 and P21 also a zoom-in of the cerebellar cortex is presented next to the sagittal section to show the location of individual cell types in the three-layered structure, the inputs of the two precerebellar systems (Climbing fibers (CF, blue) and mossy fibers (mf, green)) and the output to the cerebellar nuclei (CN, purple). Color-coding is as follows: Excitatory neurons derive from the rhombic lips (RL, red): Large cerebellar nuclei neurons (CN neurons, purple), granule cells (GCs, orange) in the granule layer (GL), and unipolar brush cells (UBCs). Inhibitory GABAergic interneurons (IN, green) derive from the ventricular zone (VZ, light green): Purkinje cells (PCs, pink) in the Purkinje cell layer (PCL), stellate and basket cells (located in the molecular layer (ML)), Golgi cells, and inhibitory CN neurons (green). Excitatory neurons are also indicated with (+) and inhibitory neurons with (-). There are two progenitor zones, the VZ and the RLs at the start of cerebellar development. Later the external granule layer (eGL, orange) is a third progenitor zone. Early during development GCs are located in the eGL which then travel along Bergmann glia to the internal GL. The development of the rodent starts E8.5 and takes around 30-35 days and the cerebellar anlage in human appears from E29, and takes around 2-3 years. In humans, the RLs are spatiotemporal expanded during gestation and promotes growth and maintenance of the posterior lobe. In mouse RLs presence is transient [22]. Cerebellar size expansion and increased lobular complexity occurs from gw 20 in human and from birth (P0) in mice.
Figure 2.
Timeline of cerebellar development of humans (dark blue, top) and rodents (light blue, bottom). Top: Sagittal sections of the outline of the human cerebellum at 6 time points during development. Top and bottom: The duration and timing of different parts of cerebellar development are indicated with a color-coded bar with a distinct bar length. The text explains what is happening during that period (adapted from [6]). Bottom: Rodent cerebellar development during different time points. The first two outlines show the differentiation of the hind- and midbrain and the relative location of the cerebellar anlage [20]. Embryonic day (E)15.5, E18.5, postnatal ~(P)4 and P21 show the outlines of cerebellar midsagittal cuts (adapted from [21]). Arrowheads indicate the migration direction of individual neuron types. At ~P4 and P21 also a zoom-in of the cerebellar cortex is presented next to the sagittal section to show the location of individual cell types in the three-layered structure, the inputs of the two precerebellar systems (Climbing fibers (CF, blue) and mossy fibers (mf, green)) and the output to the cerebellar nuclei (CN, purple). Color-coding is as follows: Excitatory neurons derive from the rhombic lips (RL, red): Large cerebellar nuclei neurons (CN neurons, purple), granule cells (GCs, orange) in the granule layer (GL), and unipolar brush cells (UBCs). Inhibitory GABAergic interneurons (IN, green) derive from the ventricular zone (VZ, light green): Purkinje cells (PCs, pink) in the Purkinje cell layer (PCL), stellate and basket cells (located in the molecular layer (ML)), Golgi cells, and inhibitory CN neurons (green). Excitatory neurons are also indicated with (+) and inhibitory neurons with (-). There are two progenitor zones, the VZ and the RLs at the start of cerebellar development. Later the external granule layer (eGL, orange) is a third progenitor zone. Early during development GCs are located in the eGL which then travel along Bergmann glia to the internal GL. The development of the rodent starts E8.5 and takes around 30-35 days and the cerebellar anlage in human appears from E29, and takes around 2-3 years. In humans, the RLs are spatiotemporal expanded during gestation and promotes growth and maintenance of the posterior lobe. In mouse RLs presence is transient [22]. Cerebellar size expansion and increased lobular complexity occurs from gw 20 in human and from birth (P0) in mice.
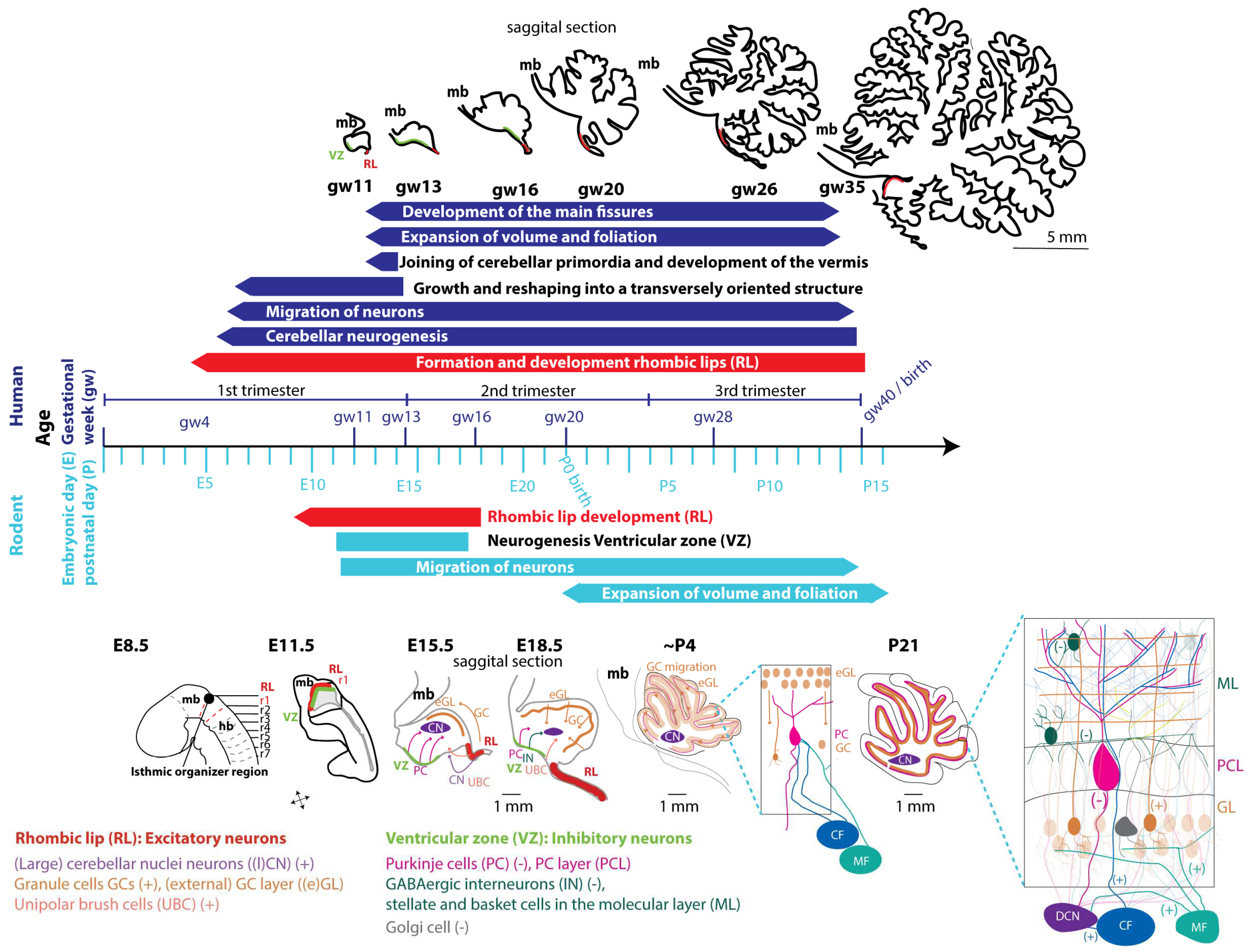
Figure 3.
Developmental timeline of the different cerebellar neuron types. The timeline of the development of individual cerebellar cell types in humans (top) and rodents (bottom). The duration and timing of development is indicated with a color-coded bar with a distinct bar length. The text explains what is happening during that period. Color-coding as follows: Excitatory neurons derive from the rhombic lips (RL, red): Large cerebellar nuclei neurons (CN neurons, purple), granule cells (GCs, orange, (+)), unipolar brush cells (UBCs, dark orange)). Inhibitory GABAergic neurons (green) derive from the ventricular zone (VZ, light green): Stellate and basket cells, Golgi cells, Lugaro cells and inhibitory CN neurons (green), Purkinje cells (PC, pink, (-)). Abbreviations: Embryonic day (E), postnatal (P), gestational week (gw).
Figure 3.
Developmental timeline of the different cerebellar neuron types. The timeline of the development of individual cerebellar cell types in humans (top) and rodents (bottom). The duration and timing of development is indicated with a color-coded bar with a distinct bar length. The text explains what is happening during that period. Color-coding as follows: Excitatory neurons derive from the rhombic lips (RL, red): Large cerebellar nuclei neurons (CN neurons, purple), granule cells (GCs, orange, (+)), unipolar brush cells (UBCs, dark orange)). Inhibitory GABAergic neurons (green) derive from the ventricular zone (VZ, light green): Stellate and basket cells, Golgi cells, Lugaro cells and inhibitory CN neurons (green), Purkinje cells (PC, pink, (-)). Abbreviations: Embryonic day (E), postnatal (P), gestational week (gw).
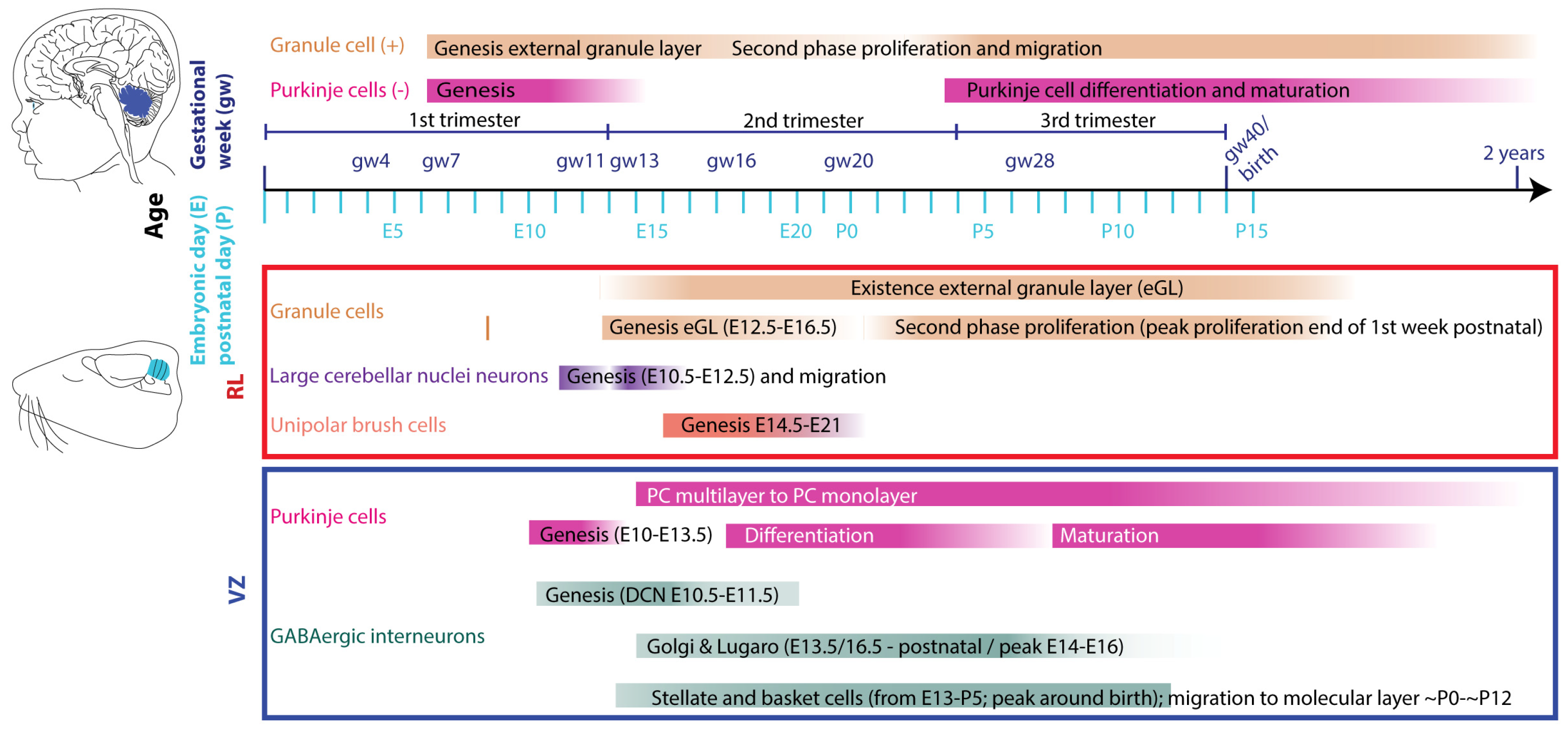
Figure 4.
Developmental timeline indicating when and how 4 common intrauterine insults have been shown to affect cerebellar development until birth in humans and until the second postnatal week in rodents. Top: Effect on cerebellar development for each prenatal intrauterine insult in humans. Bottom: Effect on cerebellar development for the prenatal intrauterine insults in rodents. P0-P14 in rodents corresponds to the last gestational period in humans when it comes to cerebellar development. The length of the bars indicates the duration and when exactly intrauterine insults have been tested and the text explains the described effects on cerebellar development. Color-coding is as follows: Cerebellar nuclei (CN) in purple, granule cells (GCs) orange, Purkinje cells (PCs) in pink and if literature mentions general cerebellar deficits it is coloured in blue (dark blue humans, light blue rodents).
Figure 4.
Developmental timeline indicating when and how 4 common intrauterine insults have been shown to affect cerebellar development until birth in humans and until the second postnatal week in rodents. Top: Effect on cerebellar development for each prenatal intrauterine insult in humans. Bottom: Effect on cerebellar development for the prenatal intrauterine insults in rodents. P0-P14 in rodents corresponds to the last gestational period in humans when it comes to cerebellar development. The length of the bars indicates the duration and when exactly intrauterine insults have been tested and the text explains the described effects on cerebellar development. Color-coding is as follows: Cerebellar nuclei (CN) in purple, granule cells (GCs) orange, Purkinje cells (PCs) in pink and if literature mentions general cerebellar deficits it is coloured in blue (dark blue humans, light blue rodents).
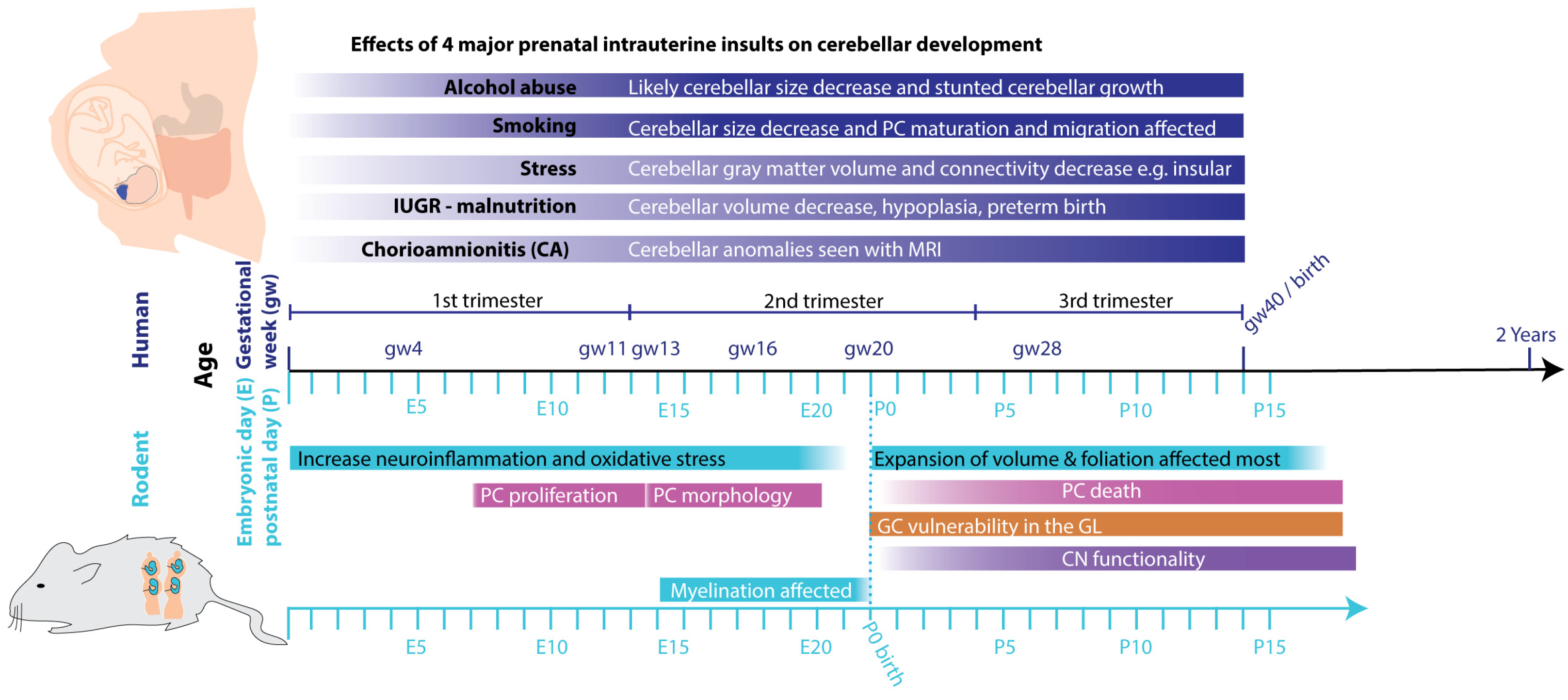
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



