Submitted:
23 October 2023
Posted:
24 October 2023
You are already at the latest version
Abstract
Research conducted on individuals with depression reveals that major depressive disorders (MDDs) coincide with diminished levels of the inhibitory neurotransmitter γ-aminobutyric acid (GABA) in the brain, as well as modifications in the subunit composition of the primary receptors (GABAA receptors) responsible for mediating GABAergic inhibition. Furthermore, there is substantial evidence supporting the significant role of GABA in regulating stress within the brain, which is a pivotal vulnerability factor in mood disorders. Although there is substantial evidence indicating that GABA may be affecting GABA receptors in peripheral tissues, there is comparatively less evidence supporting its direct action within the brain. Emerging evidence highlights that GABA may exert beneficial effects on the brain and psyche through the gut-brain axis. GABA is readily available and approved as a food supplement in many countries. While GABA enjoys wide consumer acceptance in Eastern Asian markets, with many consumers reporting favorable effects on stress regulation, mood, and sleep, rigorous independent research is still largely lacking. Basic research, coupled with initial clinical findings, makes GABA an intriguing neuro-nutritional compound deserving of clinical studies in individuals with depression and other psychological problems.
Keywords:
nutrition
; mood disorder
; neurotransmitter
; nutreaceuticals
; microbiome
; gut-brain axis
; enteric nervous system
; insomnia
; stress
1. Introduction
Depressive disorders are among the most common mental disorders, it is estimated that about 5% of adults worldwide suffer from depression [1]. During a depressive episode, a person experiences depressed mood (feeling sad, irritable, empty) or a loss of pleasure or interest in activities, for most of the day, nearly every day, for at least two weeks. Several other symptoms are also present, which may include poor concentration, feelings of excessive guilt or low self-worth, hopelessness about the future, thoughts about dying or suicide, disrupted sleep, changes in appetite or weight, and feeling especially tired or low in energy [2]. In addition to these symptoms, depression is also characterized by cognitive impairment. For instance, deficits in the episodic memory are associated with higher depression scores and pathological changes in depression [3,4]. Depression-related cognitive dysfunctions are generally associated with a higher rate of relapse [5], seen as a major factor contributing to functional impairments [6] and frequently persist after remission of a major depressive episode [7].
Globally, depression is a leading cause of disability and is, thus, a major contributor to the worldwide overall disease burden. The economic burden of major depressive disorder was estimated to amount to 326.2 billion $US in the United States alone [8]. Depression affects women more frequently than man. Depression can lead to suicide. A psychiatric diagnosis is present in about 98% of individuals committing suicide with mood disorders being the leading cause with 30.2% [9]. Depression is oftentimes categorized mild, moderate and severe forms, for all of which effective treatments exist. However, many depressed individuals receive no mental health care, especially in low- and middle-income countries where 75% of patients receive no treatment [10]. Most contemporary treatment guidelines recommend evidence-based psychological therapy as first-line treatment for ‘milder’ presentations of depression and a combination of psychological and pharmacological therapy for moderate-to-severe depression [11,12,13]. Acknowledging the indispensable role of psychotherapy in depression, even in the so-called treatment refractory depression, this review lays its focus on treatment with psychotropic agents [14]. Antidepressants are first-line pharmacotherapy in most guidelines for the management of severe depression [11,12,13]. For patients who do not respond to the initial antidepressant prescribed, guidelines recommend alternative pharmacotherapeutic strategies (e.g., switching or augmenting with additional antidepressants and/or antipsychotic medications). Hence, despite increasing emphasis on psychological interventions supported by patients expressing a preference for these, in reality antidepressant therapy is the mainstay of depression management, while psychotropic polypharmacy is the norm—especially in severe and complex mood disorders [15,16]. Despite substantial advances in treatment and management strategies for major depression, less than 50% of patients respond to first-line antidepressant treatment or psychotherapy [17]. If there is a lack of response to, or tolerability of initial treatment, alternatives can be tried. In other instances, acute responses may be obtained but subsequent relapses occur even on treatment [17,18]. “Treatment Resistant Depression (TRD)” is a term that has been used for several decades [19]. In the last couple of years, a new but related term has arisen: “Difficult-to-Treat Depression (DTD)” [20]. In such cases neurostimulation techniques, especially electroconvulsive therapy (ECT), show a considerable success rate [21]. However, ECT carries the additional risk of anesthesia, its availability is still limited in many countries, and, despite firm evidence for its effectiveness, it is not universally accepted by the public [21,22,23,24]. Given the high individual suffering and societal burden, there is a strong rationale to identify novel preventive measures and treatment approaches to treat depression.
In this review, we aim to examine the neurobiological evidence supporting the role of the neurotransmitter gamma-aminobutyric acid (GABA) in depression, stress, sleep, and cognitive function. We also aim to summarize the evidence regarding the potential beneficial effects of oral GABA on stress and mood regulation, with the goal of exploring its potential as a neuro-nutraceutical for depression. Additionally, we will focus on GABA’s production by commensal and probiotic microorganisms and its interaction with the enteric nervous system, emphasizing its actions through the gut-brain axis.
2. GABAergic Hypothesis in Depression
Growing scientific findings suggest a correlation between major depressive disorder (MDDs) and a variety of GABAergic deficiencies. The GABAergic hypothesis of MDD proposes that changes in GABAergic neurotransmission are fundamentally significant elements in the development of depression’s underlying causes [25]. Major depression’s pathophysiology is still largely elusive, despite accumulating data highlighting alterations in various central nervous neurotransmitter systems. GABA is a non-proteinogenic amino acid and a firmly established inhibitory neurotransmitter abundant in the central nervous system of animals [26]. In recent years, GABA has become a hot topic in medical and pharmaceutical studies that highlight its multiple beneficial physiological effects, including neuroprotection, stress-relieve [27], promotion of sleep, neuronal cell injury prevention [28,29] as well as antioxidant properties [30], blood pressure regulation [31,32] and protection from cancer [33]. GABA is produced from the excitatory neurotransmitter glutamate (Glu) in the brain [34]. GABAergic neurons are present throughout all levels of the neuraxis, representing between 20 and 40% of all neurons depending on the brain region and are known to balance and fine-tune excitatory neurotransmission of various neuronal systems [25]. GABA is the predominant inhibitory neurotransmitter in the mature brain [35]. GABAergic neurons account for one-third of the total synapses in the CNS, crucially impacting on neural network dynamics [36]. GABAergic neurons play pivotal roles in key processes altered in psychiatric disorders, such as neural plasticity, sensory processing, stress reactivity, memory, and attention [36,37]. In contrast to Glu, which triggers the depolarization of postsynaptic neurons inducing an excitatory postsynaptic potential, GABA induces the hyperpolarization of postsynaptic neurons resulting in an inhibitory postsynaptic potential [34]. GABA exerts its effects by activation of two entirely different classes of receptors: the ionotropic GABAA receptors (GABAARs) and the metabotropic GABABRs. GABAARs are known as key control elements of anxiety state based on the potent anxiolytic activity of benzodiazepines [38,39]. Alterations of the GABAergic system are involved in numerous psychiatric disorders, including major depressive disorder, schizophrenia, bipolar affective disorder, and autism spectrum disorders [40,41,42,43].
Clinical studies with depressed individuals using magnetic resonance spectroscopy (MRS) and positron emission tomography (PET) have identified alterations in Glu and GABA concentrations and activity, suggesting that dysfunction in excitatory and/or inhibitory neurotransmitter signaling mechanisms may play a critical role in depression [44,45]. Using proton MRS (1H-MRS), an in vivo imaging technique for total tissue detection of neurochemicals, including N-acetylaspartate, GABA, Glu, glutamine (Gln), and a combination of Glu/Gln with a minor contribution from GABA (known as Glx), aberrant amino acid neurotransmitter levels measured by 1H-MRS have been found in patients with major depression [46,47,48,49]. Compared to healthy subjects, these differences became especially apparent in the dorsolateral prefrontal cortex (PFC) [50]. In a meta-analysis of 17 1H-MRS studies investigating patients with major depression, reductions of Glx in the PFC were associated with number of failed antidepressant treatments, a measure of chronicity and a proxy for severity of depressive illness course [49]. This finding is complemented by the detection of a selective loss of calbindin-positive GABAergic interneurons in the dorsal PFC inpatients with depression [51]. Moreover, the GABAergic deficit hypothesis is supported by the detection of reduced plasma GABA levels [52,53] as well as a GABA reduction in cerebrospinal fluid and resected cortical tissue samples from depressed individuals [54,55]. Interestingly, GABA deficits seem to be most pronounced in treatment-resistant cases [56]. While altered Glu levels show now synchronicity with current mood, GABA levels seem to correlate with mood and show similar levels between remitted patients with major depression and subjects without a history of depression [57]. A pivotal role of glutamatergic/GABAergic neurotransmission in the pathophysiology of and treatment response in major depression is supported by strong evidence for the antidepressant efficacy of ketamine, a N-methyl-D-aspartate (NMDA) receptor antagonist [58,59,60]. The most effective method to manage treatment-resistant/difficult-to-treat depression is electroconvulsive therapy (ECT), although ECT’s precise mechanism of action is unclear. Evidence suggests that ECT’s strong antidepressant effect might be mediated by an increase in GABAergic activity [61]. GABA-agonists such as pregabalin showed efficacy against major depression and depressive symptoms as an adjunct treatment [62,63]. Other depression treatment modalities such as cognitive behavioral therapy and selective serotonin reuptake inhibitors also seem to restore the GABAergic deficit in major depression [64,65]. Many studies have reported how the inhibition of GABA signals allows for the continuous release of corticotropin-releasing factor (CRF) by paraventricular nucleus (PVN) neurons, resulting in cortisol overexpression and HPA axis hyperactivity, thus suggesting that such GABAergic activity is a key neurological factor in the maintenance of non-negative, anti-depressive emotional states [66].
3. GABA and Cognitive Function in Depression
Low GABA levels are also considered to play a crucial role in the cognitive symptoms of depression. The inhibitory system of GABAergic interneurons allocated throughout the hippocampus tightly controls and synchronizes the hippocampal activity [67]. This modulation of hippocampal activity is a key mechanism to control neuronal plasticity and the ability to learn [68,69]. Specifically, somatostatin (SST) expressing GABAergic interneurons and alpha 5 GABAA receptors, which are highly present in the hippocampus [70], play a crucial role in the cognitive function [71]. Reduced SST expression, for instance, is associated with impaired cognition in normal aging [72] and Alzheimer disease [73]. It is believed that the low expression of SST reduces the excitatory signal-to-noise ratio and, thus, the synchronization of the cellular and neural activity in the hippocampus; resulting in cognitive dysfunctions [74,75]. Augmenting SST+ cell post-synaptic α5-GABA-A receptor activity, on the other hand, ameliorates stress- and age-related cognitive dysfunction by balancing the hippocampal inhibitory/exhibitory activity [76,77]. Interestingly, SST expressing neurons seem to be typically affected in MDD-related GABAergic phenotypes [78,79]. Therefore, low GABA levels are considered one of most promising endophenotypes for therapeutic targets in depression [79].
4. GABA and the Microbiota-Gut-Brain Axis
The microbiome-gut-brain axis (MGBA) describes the bidirectional communication between the gastrointestinal tract including its resident microbiota and the brain; linking emotional and cognitive centers of the brain with peripheral intestinal functions. The human gut microbiome counts of trillions of symbiotic bacteria that play a key role in regulating the host brain and behavior [80]. The precise mechanisms underlying this bidirectional regulation between the gut microbiome and the brain are still open to discussion, but the vagus nerve, the endocrine and immune system as well as the synthesis and metabolism of metabolites and neurotransmitters in the gut are critically involved (reviewed in [81]). Moreover, merging evidence suggests that the gut microbiome plays a causal role in the etiology and psychopathology of MDD [82,83]. Two independent studies claimed that rodents who received fecal microbiome transplantation (FMT) from depressed patients had a higher level of inflammation and acted in a more depressive-like manner than rodents who received FMT from healthy volunteers [84,85]. Further FMT studies in rats also underscore the causal role of the gut microbiota regarding cognition. FMT from old to young rats impaired cognition and reduced the brain derived neurotrophic factor (BDNF) expression [86] whereas FMT from young rats to old rats ameliorated cognition and affected the hippocampal metabolome [87]. Numerous taxonomic alterations in bacterial composition have been identified in both individuals diagnosed with MDD and in animal models designed to simulate depressive states [88]. These alterations include species-specific microbial shifts, such as increased levels of Eggerthella, Paraprevotella, Flavonifractor, and Holdemania, and decreased levels of Christensenellaceae_R-7_group, Coprococcus, Fusicatenibacter, and Lachnospiraceae_ND3007_group in humans. In rodent models, there is a reproducible reduction in Acetatifactor. Additionally, a disrupted balance of the microbiome and functional changes are consistently observed across species. This disruption is characterized by an increase in pro-inflammatory bacteria, like Desulfovibrio and Escherichia/Shigella, and a decrease in anti-inflammatory bacteria that produce butyrate, such as Bifidobacterium and Faecalibacterium. At the species taxonomic level, the abundance of Bacteroides fragilis, Eggerthella lenta, and Ruminococcus gnavus, was found to be elevated only in patients suffering from depression. In depressed mice, there was an increased presence of Mucispirillum schaedleri and Helicobacter rodentium [88].
Generally, the gut microbiome can be modulated by diet and nutritional supplements, like live bacteria (probiotics) and non-digestible food components (prebiotics) [89,90]. There is evidence that the regular intake of a probiotic supplement improves depressive symptoms [91,92]. The probiotic supplements most frequently consist of bacteria belonging to the genera Lactobacillus and Bifidobacterium [93,94], followed by genera such as Lactococcus, Enterococcus, Streptococcus and Leuconostoc [92,95]. Interestingly, all these bacterial genera are classified as lactic acid bacteria, which are mainly used for the fermentation of raw food [96]. Thus, the consumption of fermented food seems to have beneficial effects on mood [97]. The group of prebiotics, probiotics and postbiotics (metabolites produced by the microbiome) that, when ingested, confer mental health benefits through interactions with commensal gut bacteria are defined as psychobiotics [98]. Especially, GABA, a crucial ‘postbiotic’ found in the enteric nervous system, is extensively distributed in foods and beverages and contributes significantly to MGBA functions and related disorders, including depression, anxiety, inflammatory, and cardiovascular disorders [95]. Prior research has demonstrated that bacteria have the capability to synthesize gamma-aminobutyric acid (GABA), a significant inhibitory neurotransmitter in the brain, through two distinct mechanisms. On one hand, a series of enzymes can convert arginine, ornithine, and agmatine into putrescine, and subsequently into GABA. This process involves GABA serving as an intermediate in the production of succinate via the GABA shunt pathway. This mechanism represents a means for bacteria to utilize carbon and nitrogen sources when faced with limited nutrient availability [99]. On the other hand, GABA synthesis can occur through the glutamate decarboxylase (GAD) system. In this pathway, a key enzyme, which is dependent on pyridoxal-5’-phosphate and encoded by gadA or gadB, converts glutamate into GABA. This conversion results in the production of carbon dioxide (CO2) and the consumption of a proton [100,101]. In an initial survey of the Integrated Microbial Genomes/Human Microbiome Project database, it was revealed that there are 26 distinct bacterial genera that contain orthologs of the gadB gene. Notably, this includes Bacteroides, which is known to be one of the most abundant and widespread genera in the gastrointestinal tract (GIT) [101]. Strandwitz et al. subsequently validated this discovery using a collection of 1,159 gut bacterial genomes representing diverse taxonomic groups [102]. Within this dataset, they identified 45 strains of Bacteroides that possessed orthologs of the gadB gene. Notably, GABA production was experimentally confirmed in six of these strains [102]. Through a combination of 16S rRNA sequencing and functional magnetic resonance imaging (fMRI) in individuals diagnosed with MDD, the authors observed an inverse relationship between the relative abundance of Bacteroides in the fecal microbiota and patterns in the brain that are linked to depressive symptoms [102].
Pathways of the MGBA such as the vagus nerve, prevertebral sympathetic ganglia, and the hypothalamic-pituitary-adrenal axis might be important mechanisms affected by GABA (see Figure 1). For instance, GABA-producing Lactobacilli have been found to reduce anxiety behaviors and markers of depression in mice, and alter the expression of GABAA receptor subunits in key brain regions that are involved in regulating mood and anxiety such as the hippocampus and amygdala. The study also demonstrated the importance of vagal afferents in communicating GABAergic activity from the gut to the brain as these modulatory effects were prevented in vagotomized mice [103,104,105].
5. GABA’s Impact on the Enteric Nervous System
The reciprocal communication between the brain and the Enteric Nervous System (ENS) plays a crucial role in preserving homeostasis [105]. Commensal bacteria belonging to Lactobacillus and Bifidobacterium strains have the potential to elevate GABA levels within the ENS [106]. GABA and its receptors are broadly distributed throughout the ENS [107]. Significant communication occurs between the gut and the brain via the vagal nerve [108]. An experiment conducted in mice revealed that the introduction of Lactobacillus rhamnosus (JB-1) consistently influenced the mRNA expression of GABAAα2, GABAAα1, and GABAB1b receptor subunits [103]. These receptors are frequently linked to anxiety-like behavior [109]. The application of these bacteria resulted in a decrease in stress-induced corticosterone levels compared to the control group. Importantly, none of these effects were observed in mice that had undergone vagotomy [103]. In humans, the primary application of stimulating the vagus nerve through vagus nerve stimulation (VNS) has been in the treatment of refractory epilepsy [110]. VNS has obtained FDA approval for depression treatment. The realm of VNS is expanding, and the rapid advancements in noninvasive VNS are noteworthy [111]. VNS is likely to influence multiple neurotransmitter systems in the brain [110]. GABAA receptors could potentially play a role in the therapeutic effectiveness of VNS. In a study employing single photon emission computed tomography (SPECT) using the benzodiazepine receptor inverse agonist iomazenil to assess cortical GABAA receptor density (GRD), researchers examined 10 individuals with drug-resistant partial epilepsy before and one year after the implantation of a VNS device. The findings indicated a significant correlation between therapeutic responses to VNS and the restoration of GRD to normal levels [112]. VNS also appears to elevate the concentration of unbound GABA in the cerebrospinal fluid [113].
These data suggest that while GABA may not directly cross the blood-brain barrier in humans, an indirect influence through the ENS could potentially provide a viable pathway for the impact of GABA dietary supplements. Although the connection between oral GABA administration, the vagal nerve, and GABA levels in the brain has not been firmly established, considering the existing evidence, it represents a promising avenue for future research.
6. Traditional Diets and Their Impact on Mood
The processes required for the production of fermented foods are ancient [114]. Modern humans’ paleolithic ancestors had plenty of access to food items subject to natural microbial fermentation (such as honey, fruits and berries and their juices) long before a biochemical understanding of fermentation has emerged. Long before the discovery of microbes and the underlying biochemical processes, it has been realized that fermentation renders certain foods palatable, analgesic, and psychotropic or ensures their preservation [115]. Since the dawn of culture fermented foods and beverages became valuable cultural goods aiding human nutrition, traditional medicine, and certain ritual practices [116,117,118]. There is plenty of evidence suggesting that besides alcohol production the fermentation of cereals, dairy, vegetables, fish and meats were an integral element of ancestral diets [119]. Despite the introduction of chemical preservatives and refrigeration, fermented foods still account for up to one third of consumed foods globally [120]. Mounting evidence highlights that fermentation can enhance the nutritional value of a wide variety of foods [121]. As the knowledge on the human gut microbiome grows rapidly, the crucial impact that fermented food items exert on commensal microbes, with important health implications, is uncovered inch by inch.
Many authors conceive depression to be a “disease of modernity” [122], highlighting the notion that a shift away from traditional life styles is responsible for its rising prevalence [123,124]. This theory is related to the so-called evolutionary mismatch hypothesis [125], which assumes that the rising incidence of disease such as depression arises from a flagrant discrepancy between the environments, in which humans have evolved and adapted anatomically, and the modern environment. Food is among the paramount factors defining modern environments and being markedly different from pre-modern conditions [126]. The research community shows increasing interest in food as a variable influencing mood and mental health [124]. It seems obvious that, given the brain’s dependence on nutrition to provide for its structure and energy metabolism, diet is expected to be paramount for mental health. Thus, one would expect human nutrition to be a major focus of mental health research. In reality, however, there is a dearth of research, particularly high-quality research, on the role of diet for mental health. The so-called field of nutritional psychiatry has historically been neglected and is populated with mostly poorly designed studies [121]. For millennia, diverse plant and animal products were subjected to fermentation by various bacteria, yeasts and fungi to produce palatable foods [127]. As such, fermented products are an essential part of many traditional diets. When discussing the health implications of traditional diet, the Japanese and Mediterranean models receive considerable interest [128,129,130,131,132,133,134]. The traditional Japanese diet is rich in various fermentation products, such as foods containing probiotic bacteria, black rice vinegar (kurosu), soy sauce (shoyu), soybean-barley paste (miso), natto and tempeh. These foods are produced by traditional methods that harness mixed cultures of various microorganisms such as lactic acid bacteria, acetic acid bacter sake yeast, koji molds and natto bacteria [135]. Several population studies established a link between adherence to traditional dietary practices and a lower anxiety and depression risk [136,137,138,139,140,141]. Adherence to a traditional Japanese dietary custom was linked with lower depressive symptom rates [132,133]. A substantial body of evidence indicates that the Mediterranean diet is protective against depressive symptoms and major depression [142,143]. This finding is supported by population surveys as well as randomized controlled trials [144,145,146,147,148,149,150]. Mediterranean diet’s health benefits might be explained by its high content of antioxidants, fibers, monosaturated and omega-3 fatty acids, phytosterols and probiotic microorganisms [151,152]. However, the Mediterranean diet is also rich in lacto-fermented foods (that is, foods fermented by lactic acid bacteria) such as lacto-fermented pickles and dairy products such as yoghurt and cheese [153]. The health benefits of lacto-fermented foods have received considerable attention and they might be a crucial factor underlying the Mediterranean diets health promoting effects [153]. Additional foods characteristic of the Mediterranean diet that contain GABA include items such as beans, tomatoes, spinach, mushrooms, and buckwheat [154].
7. Fermented Foods Enriched with GABA
Many fermented foods are rich in GABA. Lactic acid bacteria and yeasts exploited in fermented food production show GABA-yielding properties [155]. Numerous GABA-producing microorganisms have been identified in fermented products. For example, Lactobacillus brevis J1 is a lactic acid bacterial strain isolated from fermented cow milk, it accumulates up to 9.87 g/L GABA when cultured in Man, Rogosa, Sharpe (MRS) medium [156]. Lactobacillus plantarum M-6 strain that was isolated from traditional Chinese fermented food displays good GABA-producing trait by accumulating 545.33 mg/L GABA while inoculated in MRS medium with chickpea milk, fortified with monosodium glutamate [29]. Other than Lactobacillus spp., Bacillus cereus strain KBC was also isolated from fermented soy moromi, shown to produced maximum GABA value of 532.74 mg/L in MRS broth for 7 days of fermentation [157]. The Streptococcus thermophilus APC151 strain accumulates accumulate 2.1 mg/mL GABA and is suitable for the manufacture of GABA-enriched bioactive yogurt [158].
Regarding probiotics, Lactobacillus rhamnosus strains are among the best characterized probiotic microorganisms. L. rhamnosus is a rod shaped, facultative heterofermentative and anaerobic commensal bacterium abundant in the gastrointestinal tract. It is believed to contribute to the maintenance of gut homeostasis [159]. L. rhamnosus are well known for their GABA-producing properties [160]. Strains of L. rhamnosus such as JB-1 and HN001 can regulate depressive states in both humans and mice; particularly, the JB-1 strain has been shown to increase cortical GABAergic activity in mice [103,161,162]. L. rhamnosus (JB-1) increases CNS GABA levels in mice, modulating GABAA and GABAB receptor expression by activating GABA signaling pathways via vagal afferents [102,163]. L. rhamnosus GG has been found to increase GABA concentration within fermented adzuki bean milk under optimized cultural conditions [164]. In a study investigating the beneficial properties of quinoa yogurt beverages, Lactobacillus rhamnosus SP1 and Lactobacillus plantarum T6B10 raised the beverages’ GABA levels up to 211 mg/kg [165]. A randomized controlled trial evaluated the effect of Lactobacillus rhamnosus HN001 (HN001) given in pregnancy and postpartum on symptoms of maternal depression and anxiety in the postpartum period. Women who were administered HN001 exhibited notably reduced depression and anxiety scores during the postpartum period [162].
8. GABA-Enriched Fermented Foods as Neuro-Therapeutics
The growing global demand for functional (probiotic) dairy foods can be largely attributed to the high level of interest consumers have in food products that promote health. Yogurt has gained widespread consumer acceptance and is widely regarded as the ideal medium for delivering beneficial functional ingredients [166,167], whereas GABA has emerged as a promising bioactive ingredient in functional foods [168]. Due to its numerous physiological functions and positive effects on metabolic disorders, GABA has been the subject of extensive research [169]. One of its most significant benefits is its demonstrated hypotensive effect in animal studies and human intervention trials [170]. The biosynthesis of GABA and its optimization, while preserving sensory characteristics, are crucial factors in creating GABA-enriched food products that offer health benefits. Lactic acid bacteria (LAB) are the primary producers of GABA, making it possible to create a wide range of GABA-enriched fermented foods that are natural, safe, and eco-friendly. The increased understanding of bioactive compounds in food has opened up new opportunities for the development of naturally occurring functional foods that offer added health benefits [171]. Due to its lipophilic and charged nature at physiological pH, GABA is unable to easily cross the blood-brain barrier passively [172]. This has led to debate over whether oral administration of GABA can directly affect brain function, with some studies suggesting that the actions of oral GABA may be attributed to its effects on the enteric nervous system [95]. Earlier studies found that under normal circumstances, intravenous, intraperitoneal, or oral administration of GABA did not increase brain GABA levels [173,174]. However, recent studies in mice have identified a GABA transporter, GAT2/BGT-1, responsible for GABA transport across the blood-brain barrier, and further research is needed to explore GABA blood-brain-barrier permeability in humans [175]. Despite this, there is extensive evidence that oral supplementation of GABA can reach the brain and exert biological effects in both humans and animals, including mood improvement and activity in the central nervous system. High plasma concentrations of GABA have also been shown to increase GABA concentration in the brain [176]. For instance, in rats, one month of oral GABA supplementation enhanced novel object recognition memory and working memory [177]. Similarly, in healthy human adults at rest, the administration of 100 mg GABA in water increased the alpha:beta wave ratio measured by EEG, indicating improved relaxation [27]. Additionally, oral GABA administration during a mental stress task resulted in a smaller decrease in alpha waves, suggesting an acute stress-reducing effect in human adults [178]. In another study, GABA-enriched yeast supplementation altered the balance between cortical excitation and inhibition as shown by EEG in humans [179]. There are various possible mechanisms through which oral GABA may exert its effects. GABAA receptors can be modulated by a range of steroids, including neuroactive steroids and neurosteroids that are synthesized in the brain [180]. As such, exogenous GABA may indirectly affect GABA activity in the brain by acting on these steroid and other modulators [181]. Moreover, GABA may act as a source of energy via the GABA shunt, which can bypass the usual TCA (tricarboxylic acid) cycle and increase ATP (adenosine triphosphate) [182].
Principally, the human body is capable of producing its own GABA. However, factors such as a deficiency in estrogen, zinc, or vitamins, as well as an excess of salicylic acid and food additives, can inhibit the body’s ability to produce GABA [183]. GABA-enriched food is required since the GABA content of daily diets is rather low [184]. A GABA concentration as low as 2.01 mg in 200 mL of GABA-enriched Oolong tea showed stress decreasing effects in high stressed individuals [181]. Consumption of 100 mg of biosynthetic GABA for a week might improve sleep value in human subjects with poor sleep quality [185]. A dosage of 300 mg biosynthetic GABA from fermented rice germ helps to shorten sleep latency [186]. Abdou and colleagues have investigated the potential of orally administered GABA to promote relaxation and immunity during times of stress [27]. Two studies were conducted, the first of which evaluated the effect of GABA intake on the brain waves of 13 subjects. Each volunteer underwent 3 tests—one with only water, one with GABA, and one with L-theanine. Electroencephalograms (EEG) were taken after each test, and the results showed that after 60 minutes, GABA significantly increased alpha waves and decreased beta waves when compared to water or L-theanine. This indicates that GABA not only induces relaxation, but also reduces anxiety. The second study examined the role of GABA intake as a relaxant and anxiolytic agent in promoting immunity in stressed volunteers. Eight acrophobic subjects were divided into two groups, one receiving a placebo and the other GABA. Both groups were required to cross a suspended bridge as a stressful stimulus, and their saliva was monitored for levels of immunoglobulin A (IgA) during the crossing. The placebo group exhibited a marked decrease in IgA levels, while the GABA group showed significantly higher levels. In conclusion, GABA has demonstrated potential as a natural relaxant, with effects that can be observed within an hour of administration, and as a means of reducing anxiety. Furthermore, GABA administration may enhance immunity under conditions of stress [27]. Kanehira and colleagues studied the effects of a GABA-rich beverage on occupational fatigue [187]. The authors assigned an arithmetic task for the Uchida-Kraepelin Psychodiagnostic Test (UKT) to 30 healthy Japanese subjects, 9 of whom were diagnosed as having chronic fatigue. The subjects were administered 250 mL of a test beverage containing GABA at the dose of 0, 25, and 50 mg before assigning task for the UKT. Psychological fatigue assessed by the Visual Analogue Scale (VAS) was significantly lower in the group administrated the beverage containing 50 mg GABA than in the control group. The results of the Profile of Mood States (POMS) also indicated that psychological fatigue was significantly reduced in the 50-mg-GABA group. The salivary secretion levels of chromogranin A and cortisol—markers of physical fatigue—in both 25-mg and 50-mg-GABA groups were significantly lower than those in the control group. The 50-mg-GABA group also showed higher score on UKT by solving the arithmetic task more accurately than the control group. The results suggest that intake of GABA-containing beverages, especially those containing 50 mg of GABA, may help reduce both psychological and physical fatigue and improve task-solving ability [187].
Moreover, GABA-enriched foods are considered to improve memory and learning abilities [188]. A particularly promising functional food is fermented Laminaria japonica (FLJ), a sea tangle typically used as food resource in Pacific and Asian regions because of its high contents of dietary fiber, carbohydrates, minerals, and protein. Using a specific LAB fermentation process involving Lactobacillus brevis BJ20, the glutamic acid contained in the sea tangle is bio-converted into GABA [189]. There is convincing evidence that the consumption of FLJ has beneficial effects on cognition. An intake of 1 g/day of FLJ containing 5% GABA for eight weeks significantly increased the brain derived neurotrophic factor (BDNF), a biomarker tightly linked to hippocampal neurogenesis and memory, in elderly women compared to a placebo group [189,190,191,192]. If FLJ (5% GABA) was consumed for 6 weeks (1.5 g/day), global cognition, working memory, logical reasoning, information processing, and selective attention also improved significantly in elderly women compared to a placebo group. In mice, the supplementation of FLJ ameliorated short-term memory impairment and the hippocampal-dependent spatial learning ability to an equivalent degree as conventional therapy in scopolamine-, ethanol- and trimethyltin chloride-induced dementia [189,193]. Moreover, there is evidence that kefir, a traditional fermented milk beverage from the Caucasus mountains that is produced by adding a kefir grain to milk, increases the gut microbiota’s capacity to produce GABA in mice. On the behavioral side, the consumption of kefir improved reward learning and fear-dependent contextual memory [194]. Even though all these studies reported convincing evidence that GABA-enriched foods can improve cognition, there are also studies reporting null or even negative results. Tınok et al. could not find a positive effect of a GABA supplement (800 mg) dissolved in orange juice on spatial attention and visual working memory [195]. Also, Leonte et al. did not find a positive effect of GABA supplement (800 mg) on spatial attention but on temporal attention [196]. However, the improvement of temporal attention could not be replicated by Tınok et al. [195].
Interestingly, there are no studies investigating the beneficial effect of GABA-enriched foods on cognitive deficits in depression. Nevertheless, two studies examined the effect of a probiotic supplement on cognitive symptoms in depressed patients [197,198]. Both studies found improved verbal episodic memory after the consumption of a probiotic supplement for either four or eight weeks. In both studies, the administered supplements contained Lactobacillus and Bifidobacterium strains such as Lactobacillus plantarum and Bifidobacterium brevi, which are known to produce GABA [95]. Thus, it may well be assumed that an increased production of GABA was responsible for the improvement in hippocampal dependent verbal episodic memory. However, there is currently a lack of direct evidence to confirm this.
9. Conclusion
It’s possible that the quantity of GABA reaching the brain may be too minimal to have clinical relevance but sufficient to produce an effect in a stop-change paradigm. Nonetheless, oral GABA obtained from oral supplements, probiotics, GABA-rich fermented foods, or fortified food products may exert an effect on the brain through complex peripheral mechanisms, primarily involving the Enteric Nervous System (ENS) and the gut-brain axis. We maintain the belief that Magnetic Resonance Spectroscopy (MRS) studies offer the most auspicious approach for directly evaluating the impact of GABA supplementation on GABA levels in the human brain.
Notably, in one of the rat studies under consideration, the administration of GABA alone led to a 33% increase in brain GABA levels. However, when GABA was administered in conjunction with L-arginine, brain GABA levels surged by an impressive 383.3% [199]. It would indeed be intriguing to investigate whether this effect can be replicated in humans, potentially serving as a leverage point to enhance the efficacy of GABA in human applications. Considering the widespread distribution of GABA and its receptors in peripheral tissues, it is highly justified to conduct further research into the impact of oral GABA on peripheral tissues and the gut-brain axis as part of the investigation into the potential benefits of GABA-enriched foods.
The dietary supplement form of GABA is readily accessible to consumers. While many individuals assert that they derive advantages from using these products, it remains uncertain whether these supplements provide benefits beyond what could be attributed to a placebo effect. There is some supportive evidence suggesting a calming impact of GABA dietary supplements; however, it’s worth noting that much of this evidence comes from researchers who may have a potential conflict of interest [172]. Furthermore, it’s important to mention that in some of the studies identified, the sample sizes were relatively small, which can limit the strength of the conclusions that can be drawn [200]. Additional evidence from independent studies is required to establish the favorable psychological effects of GABA conclusively. LD50 tests conducted on rats using a 5000 mg/kg dose of natural GABA did not result in any fatalities [201]. This demonstrates a favorable safety profile, rendering it an appealing adjunct therapeutic option deserving of further comprehensive study. However, it is evident that additional safety data covering various dosage ranges in humans are clearly needed.
In summary, the various potential physiological benefits of GABA in mood regulation, stress tolerance, and cognitive performance, coupled with its favorable safety profile, make GABA an intriguing natural compound worthy of investigation in prospective clinical trials for depression.
Author Contributions
Conceptualization, T.L. and E.S..; Writing—Original Draft Preparation, T.L. and E.S..; Writing—Review & Editing, T.L., E.S., U.L. and A.B..; Visualization, T.L. and E.S..
Conflicts of Interest
The authors declare no conflict of interest.
References
- Institute of Health Metrics and Evaluation Global Health Data Exchange (GHDx).
- World Health Organization Depression.
- Bierman, E.J.M.; Comijs, H.C.; Jonker, C.; Beekman, A.T.F. Effects of Anxiety Versus Depression on Cognition in Later Life. The American Journal of Geriatric Psychiatry 2005, 13, 686–693. [Google Scholar] [CrossRef]
- Dere, E.; Pause, B.M.; Pietrowsky, R. Emotion and Episodic Memory in Neuropsychiatric Disorders. Behavioural Brain Research 2010, 215, 162–171. [Google Scholar] [CrossRef]
- Majer, M.; Ising, M.; Künzel, H.; Binder, E.B.; Holsboer, F.; Modell, S.; Zihl, J. Impaired Divided Attention Predicts Delayed Response and Risk to Relapse in Subjects with Depressive Disorders. Psychol Med 2004, 34, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, R.S.; Cha, D.S.; Soczynska, J.K.; Woldeyohannes, H.O.; Gallaugher, L.A.; Kudlow, P.; Alsuwaidan, M.; Baskaran, A. Cognitive Deficits and Functional Outcomes in Major Depressive Disorder: Determinants, Substrates, and Treatment Interventions. Depress Anxiety 2013, 30, 515–527. [Google Scholar] [CrossRef]
- Semkovska, M.; Quinlivan, L.; O’Grady, T.; Johnson, R.; Collins, A.; O’Connor, J.; Knittle, H.; Ahern, E.; Gload, T. Cognitive Function Following a Major Depressive Episode: A Systematic Review and Meta-Analysis. Lancet Psychiatry 2019, 6, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, P.E.; Fournier, A.-A.; Sisitsky, T.; Simes, M.; Berman, R.; Koenigsberg, S.H.; Kessler, R.C. The Economic Burden of Adults with Major Depressive Disorder in the United States (2010 and 2018). Pharmacoeconomics 2021, 39, 653–665. [Google Scholar] [CrossRef]
- Bertolote, J.M.; Fleischmann, A.; De Leo, D.; Wasserman, D. Psychiatric Diagnoses and Suicide: Revisiting the Evidence. Crisis 2004, 25, 147–155. [Google Scholar] [CrossRef]
- Evans-Lacko, S.; Aguilar-Gaxiola, S.; Al-Hamzawi, A.; Alonso, J.; Benjet, C.; Bruffaerts, R.; Chiu, W.T.; Florescu, S.; de Girolamo, G.; Gureje, O.; et al. Socio-Economic Variations in the Mental Health Treatment Gap for People with Anxiety, Mood, and Substance Use Disorders: Results from the WHO World Mental Health (WMH) Surveys. Psychol Med 2018, 48, 1560–1571. [Google Scholar] [CrossRef]
- Malhi, G.S.; Bassett, D.; Boyce, P.; Bryant, R.; Fitzgerald, P.B.; Fritz, K.; Hopwood, M.; Lyndon, B.; Mulder, R.; Murray, G.; et al. Royal Australian and New Zealand College of Psychiatrists Clinical Practice Guidelines for Mood Disorders. Australian & New Zealand Journal of Psychiatry 2015, 49, 1087–1206. [Google Scholar] [CrossRef]
- Middleton, H.; Shaw, I.; Hull, S.; Feder, G. NICE Guidelines for the Management of Depression. BMJ 2005, 330, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Härter, M.; Prien, P. The Diagnosis and Treatment of Unipolar Depression—National Disease Management Guideline. Dtsch Arztebl Int 2023. [Google Scholar] [CrossRef]
- Cuijpers, P.; Karyotaki, E.; Eckshtain, D.; Ng, M.Y.; Corteselli, K.A.; Noma, H.; Quero, S.; Weisz, J.R. Psychotherapy for Depression Across Different Age Groups: A Systematic Review and Meta-Analysis. JAMA Psychiatry 2020, 77, 694. [Google Scholar] [CrossRef]
- Malhi, G.S.; Bell, E. Make News: Treatment-Resistant Depression – an Irreversible Problem in Need of a Reversible Solution? Australian & New Zealand Journal of Psychiatry 2020, 54, 111–113. [Google Scholar] [CrossRef]
- Malhi, G.S.; Das, P.; Mannie, Z.; Irwin, L. Treatment-Resistant Depression: Problematic Illness or a Problem in Our Approach? British Journal of Psychiatry 2019, 214, 1–3. [Google Scholar] [CrossRef]
- Thase, M.E.; Friedman, E.S.; Biggs, M.M.; Wisniewski, S.R.; Trivedi, M.H.; Luther, J.F.; Fava, M.; Nierenberg, A.A.; McGrath, P.J.; Warden, D.; et al. Cognitive Therapy versus Medication in Augmentation and Switch Strategies as Second-Step Treatments: A STAR*D Report. Am J Psychiatry 2007, 164, 739–752. [Google Scholar] [CrossRef]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D.; et al. Acute and Longer-Term Outcomes in Depressed Outpatients Requiring One or Several Treatment Steps: A STAR*D Report. Am J Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Overall, J.E. Methodologic Issues in the Epidemiology of Treatment Resistant Depression. Contribution to Epidemiology. Pharmakopsychiatr Neuropsychopharmakol 1974, 7, 129–138. [Google Scholar] [CrossRef]
- Kupfer, D.J.; Charney, D.S. “Difficult-to-Treat Depression”. Biol Psychiatry 2003, 53, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Bayes, A.J.; Parker, G.B. Comparison of Guidelines for the Treatment of Unipolar Depression: A Focus on Pharmacotherapy and Neurostimulation. Acta Psychiatr Scand 2018, 137, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Bwalya, G.M.; Srinivasan, V.; Wang, M. Electroconvulsive Therapy Anesthesia Practice Patterns: Results of a UK Postal Survey. J ECT 2011, 27, 81–85. [Google Scholar] [CrossRef] [PubMed]
- González-Pando, D.; González-Menéndez, A.; Aparicio-Basauri, V.; Sanz de la Garza, C.L.; Torracchi-Carrasco, J.E.; Pérez-Álvarez, M. Ethical Implications of Electroconvulsive Therapy: A Review. Ethical Hum Psychol Psychiatry 2021, 23, 25–38. [Google Scholar] [CrossRef]
- Gergel, T.; Howard, R.; Lawrence, R.; Seneviratne, T. Time to Acknowledge Good Electroconvulsive Therapy Research. Lancet Psychiatry 2021, 8, 1032–1033. [Google Scholar] [CrossRef]
- Luscher, B.; Shen, Q.; Sahir, N. The GABAergic Deficit Hypothesis of Major Depressive Disorder. Mol Psychiatry 2011, 16, 383–406. [Google Scholar] [CrossRef]
- Spiering, M.J. The Discovery of GABA in the Brain. J Biol Chem 2018, 293, 19159–19160. [Google Scholar] [CrossRef]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and Immunity Enhancement Effects of γ-Aminobutyric Acid (GABA) Administration in Humans. BioFactors 2006, 26, 201–208. [Google Scholar] [CrossRef]
- Lee, B.-J.; Kim, J.-S.; Kang, Y.M.; Lim, J.-H.; Kim, Y.-M.; Lee, M.-S.; Jeong, M.-H.; Ahn, C.-B.; Je, J.-Y. Antioxidant Activity and γ-Aminobutyric Acid (GABA) Content in Sea Tangle Fermented by Lactobacillus Brevis BJ20 Isolated from Traditional Fermented Foods. Food Chem 2010, 122, 271–276. [Google Scholar] [CrossRef]
- Li, W.; Wei, M.; Wu, J.; Rui, X.; Dong, M. Novel Fermented Chickpea Milk with Enhanced Level of γ -Aminobutyric Acid and Neuroprotective Effect on PC12 Cells. PeerJ 2016, 4, e2292. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Xu, L.; Zeng, X.; Li, Z.; Qin, B.; He, N. New Perspective of GABA as an Inhibitor of Formation of Advanced Lipoxidation End-Products: It’s Interaction with Malondiadehyde. J Biomed Nanotechnol 2010, 6, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Furuya, Y.; Endo, Y.; Fujimoto, K. Effect of Gamma-Aminobutyric Acid-Enriched Tempeh-like Fermented Soybean (GABA-Tempeh) on the Blood Pressure of Spontaneously Hypertensive Rats. Biosci Biotechnol Biochem 2003, 67, 1806–1808. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.K.; Kim, N.Y.; Ahn, H.J.; Ji, G.E. γ-Aminobutyric Acid (GABA) Production and Angiotensin-I Converting Enzyme (ACE) Inhibitory Activity of Fermented Soybean Containing Sea Tangle by the Co-Culture of Lactobacillus Brevis with Aspergillus Oryzae. J Microbiol Biotechnol 2015, 25, 1315–1320. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, C.; Wang, C.; Hu, Y.; Qiu, L.; Xu, P. Neurotransmitter γ-Aminobutyric Acid-Mediated Inhibition of the Invasive Ability of Cholangiocarcinoma Cells. Oncol Lett 2011, 2, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.C.; Kemp, J.A. Glutamate- and GABA-Based CNS Therapeutics. Curr Opin Pharmacol 2006, 6, 7–17. [Google Scholar] [CrossRef]
- Patel, A.B.; Rothman, D.L.; Cline, G.W.; Behar, K.L. Glutamine Is the Major Precursor for GABA Synthesis in Rat Neocortex in Vivo Following Acute GABA-Transaminase Inhibition. Brain Res 2001, 919, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Möhler, H. Molecular Regulation of Cognitive Functions and Developmental Plasticity: Impact of GABA A Receptors. J Neurochem 2007, 102, 1–12. [Google Scholar] [CrossRef]
- Mody, I.; Pearce, R.A. Diversity of Inhibitory Neurotransmission through GABA(A) Receptors. Trends Neurosci 2004, 27, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Mombereau, C.; Kaupmann, K.; Froestl, W.; Sansig, G.; van der Putten, H.; Cryan, J.F. Genetic and Pharmacological Evidence of a Role for GABA(B) Receptors in the Modulation of Anxiety- and Antidepressant-like Behavior. Neuropsychopharmacology 2004, 29, 1050–1062. [Google Scholar] [CrossRef]
- Mombereau, C.; Kaupmann, K.; Gassmann, M.; Bettler, B.; van der Putten, H.; Cryan, J.F. Altered Anxiety and Depression-Related Behaviour in Mice Lacking GABAB(2) Receptor Subunits. Neuroreport 2005, 16, 307–310. [Google Scholar] [CrossRef]
- Luscher, B.; Shen, Q.; Sahir, N. The GABAergic Deficit Hypothesis of Major Depressive Disorder. Mol Psychiatry 2011, 16, 383–406. [Google Scholar] [CrossRef]
- Gonzalez-Burgos, G.; Fish, K.N.; Lewis, D.A. GABA Neuron Alterations, Cortical Circuit Dysfunction and Cognitive Deficits in Schizophrenia. Neural Plast 2011, 2011, 723184. [Google Scholar] [CrossRef]
- Brambilla, P.; Perez, J.; Barale, F.; Schettini, G.; Soares, J.C. GABAergic Dysfunction in Mood Disorders. Mol Psychiatry 2003, 8, 721–737. [Google Scholar] [CrossRef]
- Horder, J.; Petrinovic, M.M.; Mendez, M.A.; Bruns, A.; Takumi, T.; Spooren, W.; Barker, G.J.; Künnecke, B.; Murphy, D.G. Glutamate and GABA in Autism Spectrum Disorder-a Translational Magnetic Resonance Spectroscopy Study in Man and Rodent Models. Transl Psychiatry 2018, 8, 106. [Google Scholar] [CrossRef]
- Lener, M.S.; Niciu, M.J.; Ballard, E.D.; Park, M.; Park, L.T.; Nugent, A.C.; Zarate, C.A. Glutamate and Gamma-Aminobutyric Acid Systems in the Pathophysiology of Major Depression and Antidepressant Response to Ketamine. Biol Psychiatry 2017, 81, 886–897. [Google Scholar] [CrossRef]
- Knudsen, M.K.; Near, J.; Blicher, A.B.; Videbech, P.; Blicher, J.U. Magnetic Resonance (MR) Spectroscopic Measurement of γ-Aminobutyric Acid (GABA) in Major Depression before and after Electroconvulsive Therapy. Acta Neuropsychiatr 2019, 31, 17–26. [Google Scholar] [CrossRef]
- Sanacora, G.; Treccani, G.; Popoli, M. Towards a Glutamate Hypothesis of Depression: An Emerging Frontier of Neuropsychopharmacology for Mood Disorders. Neuropharmacology 2012, 62, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Lener, M.S.; Iosifescu, D. V In Pursuit of Neuroimaging Biomarkers to Guide Treatment Selection in Major Depressive Disorder: A Review of the Literature. Ann N Y Acad Sci 2015, 1344, 50–65. [Google Scholar] [CrossRef]
- Yüksel, C.; Öngür, D. Magnetic Resonance Spectroscopy Studies of Glutamate-Related Abnormalities in Mood Disorders. Biol Psychiatry 2010, 68, 785–794. [Google Scholar] [CrossRef]
- Arnone, D.; Mumuni, A.N.; Jauhar, S.; Condon, B.; Cavanagh, J. Indirect Evidence of Selective Glial Involvement in Glutamate-Based Mechanisms of Mood Regulation in Depression: Meta-Analysis of Absolute Prefrontal Neuro-Metabolic Concentrations. Eur Neuropsychopharmacol 2015, 25, 1109–1117. [Google Scholar] [CrossRef]
- Yildiz-Yesiloglu, A.; Ankerst, D.P. Review of 1H Magnetic Resonance Spectroscopy Findings in Major Depressive Disorder: A Meta-Analysis. Psychiatry Res 2006, 147, 1–25. [Google Scholar] [CrossRef]
- Rajkowska, G.; O’Dwyer, G.; Teleki, Z.; Stockmeier, C.A.; Miguel-Hidalgo, J.J. GABAergic Neurons Immunoreactive for Calcium Binding Proteins Are Reduced in the Prefrontal Cortex in Major Depression. Neuropsychopharmacology 2007, 32, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Petty, F.; Schlesser, M.A. Plasma GABA in Affective Illness. J Affect Disord 1981, 3, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Petty, F.; Sherman, A.D. Plasma GABA Levels in Psychiatric Illness. J Affect Disord 1984, 6, 131–138. [Google Scholar] [CrossRef]
- Gerner, R.H.; Hare, T.A. CSF GABA in Normal Subjects and Patients with Depression, Schizophrenia, Mania, and Anorexia Nervosa. Am J Psychiatry 1981, 138, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Honig, A.; Bartlett, J.R.; Bouras, N.; Bridges, P.K. Amino Acid Levels in Depression: A Preliminary Investigation. J Psychiatr Res 1988, 22, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Price, R.B.; Shungu, D.C.; Mao, X.; Nestadt, P.; Kelly, C.; Collins, K.A.; Murrough, J.W.; Charney, D.S.; Mathew, S.J. Amino Acid Neurotransmitters Assessed by Proton Magnetic Resonance Spectroscopy: Relationship to Treatment Resistance in Major Depressive Disorder. Biol Psychiatry 2009, 65, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Schür, R.R.; Draisma, L.W.R.; Wijnen, J.P.; Boks, M.P.; Koevoets, M.G.J.C.; Joëls, M.; Klomp, D.W.; Kahn, R.S.; Vinkers, C.H. Brain GABA Levels across Psychiatric Disorders: A Systematic Literature Review and Meta-Analysis of (1) H-MRS Studies. Hum Brain Mapp 2016, 37, 3337–3352. [Google Scholar] [CrossRef] [PubMed]
- Kohrs, R.; Durieux, M.E. Ketamine. Anesth Analg 1998, 87, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Fond, G.; Loundou, A.; Rabu, C.; Macgregor, A.; Lançon, C.; Brittner, M.; Micoulaud-Franchi, J.-A.; Richieri, R.; Courtet, P.; Abbar, M.; et al. Ketamine Administration in Depressive Disorders: A Systematic Review and Meta-Analysis. Psychopharmacology (Berl) 2014, 231, 3663–3676. [Google Scholar] [CrossRef]
- Newport, D.J.; Carpenter, L.L.; McDonald, W.M.; Potash, J.B.; Tohen, M.; Nemeroff, C.B. Ketamine and Other NMDA Antagonists: Early Clinical Trials and Possible Mechanisms in Depression. American Journal of Psychiatry 2015, 172, 950–966. [Google Scholar] [CrossRef]
- Esel, E.; Kose, K.; Hacimusalar, Y.; Ozsoy, S.; Kula, M.; Candan, Z.; Turan, T. The Effects of Electroconvulsive Therapy on GABAergic Function in Major Depressive Patients. J ECT 2008, 24, 224–228. [Google Scholar] [CrossRef]
- Vitali, M.; Tedeschini, E.; Mistretta, M.; Fehling, K.; Aceti, F.; Ceccanti, M.; Fava, M. Adjunctive Pregabalin in Partial Responders with Major Depressive Disorder and Residual Anxiety. J Clin Psychopharmacol 2013, 33, 95–98. [Google Scholar] [CrossRef]
- Stein, D.J.; Baldwin, D.S.; Baldinetti, F.; Mandel, F. Efficacy of Pregabalin in Depressive Symptoms Associated with Generalized Anxiety Disorder: A Pooled Analysis of 6 Studies. Eur Neuropsychopharmacol 2008, 18, 422–430. [Google Scholar] [CrossRef]
- Sanacora, G.; Fenton, L.R.; Fasula, M.K.; Rothman, D.L.; Levin, Y.; Krystal, J.H.; Mason, G.F. Cortical Gamma-Aminobutyric Acid Concentrations in Depressed Patients Receiving Cognitive Behavioral Therapy. Biol Psychiatry 2006, 59, 284–286. [Google Scholar] [CrossRef]
- Sanacora, G.; Mason, G.F.; Rothman, D.L.; Hyder, F.; Ciarcia, J.J.; Ostroff, R.B.; Berman, R.M.; Krystal, J.H. Increased Cortical GABA Concentrations in Depressed Patients Receiving ECT. Am J Psychiatry 2003, 160, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Peng, T.; Gaur, U.; Silva, M.; Little, P.; Chen, Z.; Qiu, W.; Zhang, Y.; Zheng, W. Role of Corticotropin Releasing Factor in the Neuroimmune Mechanisms of Depression: Examination of Current Pharmaceutical and Herbal Therapies. Front Cell Neurosci 2019, 13, 290. [Google Scholar] [CrossRef]
- Cobb, S.R.; Buhl, E.H.; Halasy, K.; Paulsen, O.; Somogyi, P. Synchronization of Neuronal Activity in Hippocampus by Individual GABAergic Interneurons. Nature 1995, 378, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Q.; Barberis, A.; Higley, M.J. Preserving the Balance: Diverse Forms of Long-Term GABAergic Synaptic Plasticity. Nat Rev Neurosci 2019, 20, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Wilcke, T.; Fuchs, E.; Funke, K.; Vlachos, A.; Müller-Dahlhaus, F.; Puts, N.A.J.; Harris, R.E.; Edden, R.A.E. GABA—from Inhibition to Cognition: Emerging Concepts. The Neuroscientist 2018, 24, 501–515. [Google Scholar] [CrossRef]
- Möhler, H.; Rudolph, U. Disinhibition, an Emerging Pharmacology of Learning and Memory. F1000Res 2017, 6. [Google Scholar] [CrossRef]
- Collinson, N.; Kuenzi, F.M.; Jarolimek, W.; Maubach, K.A.; Cothliff, R.; Sur, C.; Smith, A.; Otu, F.M.; Howell, O.; Atack, J.R.; et al. Enhanced Learning and Memory and Altered GABAergic Synaptic Transmission in Mice Lacking the Alpha 5 Subunit of the GABAA Receptor. J Neurosci 2002, 22, 5572–5580. [Google Scholar] [CrossRef]
- Martel, G.; Dutar, P.; Epelbaum, J.; Viollet, C. Somatostatinergic Systems: An Update on Brain Functions in Normal and Pathological Aging. Front Endocrinol (Lausanne) 2012, 3. [Google Scholar] [CrossRef]
- Kumar, U. Expression of Somatostatin Receptor Subtypes (SSTR1-5) in Alzheimer’s Disease Brain: An Immunohistochemical Analysis. Neuroscience 2005, 134, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Fee, C.; Banasr, M.; Sibille, E. Somatostatin-Positive Gamma-Aminobutyric Acid Interneuron Deficits in Depression: Cortical Microcircuit and Therapeutic Perspectives. Biol Psychiatry 2017, 82, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Gentet, L.J.; Kremer, Y.; Taniguchi, H.; Huang, Z.J.; Staiger, J.F.; Petersen, C.C.H. Unique Functional Properties of Somatostatin-Expressing GABAergic Neurons in Mouse Barrel Cortex. Nat Neurosci 2012, 15, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Piantadosi, S.C.; French, B.J.; Poe, M.M.; Timić, T.; Marković, B.D.; Pabba, M.; Seney, M.L.; Oh, H.; Orser, B.A.; Savić, M.M.; et al. Sex-Dependent Anti-Stress Effect of an A5 Subunit Containing GABAA Receptor Positive Allosteric Modulator. Front Pharmacol 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Koh, M.T.; Rosenzweig-Lipson, S.; Gallagher, M. Selective GABA(A) A5 Positive Allosteric Modulators Improve Cognitive Function in Aged Rats with Memory Impairment. Neuropharmacology 2013, 64, 145–152. [Google Scholar] [CrossRef]
- Prévot, T.; Sibille, E. Altered GABA-Mediated Information Processing and Cognitive Dysfunctions in Depression and Other Brain Disorders. Mol Psychiatry 2021, 26, 151–167. [Google Scholar] [CrossRef]
- Hasler, G.; Northoff, G. Discovering Imaging Endophenotypes for Major Depression. Mol Psychiatry 2011, 16, 604–619. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A. V.; et al. The Microbiota-Gut-Brain Axis. Physiol Rev 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Pereira, J.S.; Rea, K.; Nolan, Y.M.; O’Leary, O.F.; Dinan, T.G.; Cryan, J.F. Depression’s Unholy Trinity: Dysregulated Stress, Immunity, and the Microbiome. Annu Rev Psychol 2020, 71, 49–78. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Hart, R.A.; Charlesworth, R.P.G.; Sharpley, C.F. Gut Microbiome and Depression: What We Know and What We Need to Know. Rev Neurosci 2018, 29, 629–643. [Google Scholar] [CrossRef]
- Kelly, J.R.; Borre, Y.; O’ Brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G.; et al. Transferring the Blues: Depression-Associated Gut Microbiota Induces Neurobehavioural Changes in the Rat. J Psychiatr Res 2016, 82, 109–118. [Google Scholar] [CrossRef]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut Microbiome Remodeling Induces Depressive-like Behaviors through a Pathway Mediated by the Host’s Metabolism. Mol Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ning, L.; Yin, Y.; Wang, R.; Zhang, Z.; Hao, L.; Wang, B.; Zhao, X.; Yang, X.; Yin, L.; et al. Age-Related Shifts in Gut Microbiota Contribute to Cognitive Decline in Aged Rats. Aging 2020, 12, 7801–7817. [Google Scholar] [CrossRef]
- Boehme, M.; Guzzetta, K.E.; Bastiaanssen, T.F.S.; van de Wouw, M.; Moloney, G.M.; Gual-Grau, A.; Spichak, S.; Olavarría-Ramírez, L.; Fitzgerald, P.; Morillas, E.; et al. Microbiota from Young Mice Counteracts Selective Age-Associated Behavioral Deficits. Nat Aging 2021, 1, 666–676. [Google Scholar] [CrossRef]
- Liu, L.; Wang, H.; Zhang, H.; Chen, X.; Zhang, Y.; Wu, J.; Zhao, L.; Wang, D.; Pu, J.; Ji, P.; et al. Toward a Deeper Understanding of Gut Microbiome in Depression: The Promise of Clinical Applicability. Advanced Science 2022, 9. [Google Scholar] [CrossRef]
- Firth, J.; Gangwisch, J.E.; Borsini, A.; Wootton, R.E.; Mayer, E.A. Food and Mood: How Do Diet and Nutrition Affect Mental Wellbeing? BMJ 2020, m2382. [Google Scholar] [CrossRef] [PubMed]
- Liwinski, T.; Elinav, E. Harnessing the Microbiota for Therapeutic Purposes. American Journal of Transplantation 2020, 20, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Schaub, A.-C.; Schneider, E.; Vazquez-Castellanos, J.F.; Schweinfurth, N.; Kettelhack, C.; Doll, J.P.K.; Yamanbaeva, G.; Mählmann, L.; Brand, S.; Beglinger, C.; et al. Clinical, Gut Microbial and Neural Effects of a Probiotic Add-on Therapy in Depressed Patients: A Randomized Controlled Trial. Transl Psychiatry 2022, 12, 227. [Google Scholar] [CrossRef]
- Ng, Q.X.; Peters, C.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.-S. A Meta-Analysis of the Use of Probiotics to Alleviate Depressive Symptoms. J Affect Disord 2018, 228, 13–19. [Google Scholar] [CrossRef]
- Nikolova, V.L.; Cleare, A.J.; Young, A.H.; Stone, J.M. Acceptability, Tolerability, and Estimates of Putative Treatment Effects of Probiotics as Adjunctive Treatment in Patients With Depression. JAMA Psychiatry 2023, 80, 842. [Google Scholar] [CrossRef]
- Schaub, A.-C.; Schneider, E.; Vazquez-Castellanos, J.F.; Schweinfurth, N.; Kettelhack, C.; Doll, J.P.K.; Yamanbaeva, G.; Mählmann, L.; Brand, S.; Beglinger, C.; et al. Clinical, Gut Microbial and Neural Effects of a Probiotic Add-on Therapy in Depressed Patients: A Randomized Controlled Trial. Transl Psychiatry 2022, 12, 227. [Google Scholar] [CrossRef] [PubMed]
- Diez-Gutiérrez, L.; San Vicente, L.; Barrón, L.J.R.; del Carmen Villarán, M.; Chávarri, M. Gamma-Aminobutyric Acid and Probiotics: Multiple Health Benefits and Their Future in the Global Functional Food and Nutraceuticals Market. J Funct Foods 2020, 64, 103669. [Google Scholar] [CrossRef]
- Vodnar, D.; Paucean, A.; Dulf, F.; Socaciu, C. HPLC Characterization of Lactic Acid Formation and FTIR Fingerprint of Probiotic Bacteria during Fermentation Processes. Not Bot Horti Agrobot Cluj Napoca 2010, 38, 109–113. [Google Scholar]
- Aslam, H.; Green, J.; Jacka, F.N.; Collier, F.; Berk, M.; Pasco, J.; Dawson, S.L. Fermented Foods, the Gut and Mental Health: A Mechanistic Overview with Implications for Depression and Anxiety. Nutr Neurosci 2020, 23, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria–Gut–Brain Signals. Trends Neurosci 2016, 39, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Shaibe, E.; Metzer, E.; Halpern, Y.S. Metabolic Pathway for the Utilization of L-Arginine, L-Ornithine, Agmatine, and Putrescine as Nitrogen Sources in Escherichia Coli K-12. J Bacteriol 1985, 163, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.K.; Kassam, T.; Singh, B.; Elliott, J.F. Escherichia Coli Has Two Homologous Glutamate Decarboxylase Genes That Map to Distinct Loci. J Bacteriol 1992, 174, 5820–5826. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Johnson, C.; Luk, B.; Uribe, G.; Fu, Y.; Oezguen, N.; Matsunami, R.K.; Lugo, M.; Major, A.; Mori-Akiyama, Y.; et al. <scp>GABA</Scp> -producing Bifidobacterium Dentium Modulates Visceral Sensitivity in the Intestine. Neurogastroenterology & Motility 2017, 29. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat Microbiol 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus Strain Regulates Emotional Behavior and Central GABA Receptor Expression in a Mouse via the Vagus Nerve. Proc Natl Acad Sci U S A 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-Altering Microorganisms: The Impact of the Gut Microbiota on Brain and Behaviour. Nat Rev Neurosci 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann Gastroenterol 2015, 28, 203–209. [Google Scholar] [PubMed]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric Acid Production by Culturable Bacteria from the Human Intestine. J Appl Microbiol 2012, 113, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Auteri, M.; Zizzo, M.G.; Serio, R. GABA and GABA Receptors in the Gastrointestinal Tract: From Motility to Inflammation. Pharmacol Res 2015, 93, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front Neurosci 2018, 12. [Google Scholar] [CrossRef]
- Bravo, J.A.; Julio-Pieper, M.; Forsythe, P.; Kunze, W.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Communication between Gastrointestinal Bacteria and the Nervous System. Curr Opin Pharmacol 2012, 12, 667–672. [Google Scholar] [CrossRef]
- Carron, R.; Roncon, P.; Lagarde, S.; Dibué, M.; Zanello, M.; Bartolomei, F. Latest Views on the Mechanisms of Action of Surgically Implanted Cervical Vagal Nerve Stimulation in Epilepsy. Neuromodulation: Technology at the Neural Interface 2023, 26, 498–506. [Google Scholar] [CrossRef]
- Austelle, C.W.; O’Leary, G.H.; Thompson, S.; Gruber, E.; Kahn, A.; Manett, A.J.; Short, B.; Badran, B.W. A Comprehensive Review of Vagus Nerve Stimulation for Depression. Neuromodulation: Technology at the Neural Interface 2022, 25, 309–315. [Google Scholar] [CrossRef]
- Marrosu, F.; Serra, A.; Maleci, A.; Puligheddu, M.; Biggio, G.; Piga, M. Correlation between GABAA Receptor Density and Vagus Nerve Stimulation in Individuals with Drug-Resistant Partial Epilepsy. Epilepsy Res 2003, 55, 59–70. [Google Scholar] [CrossRef]
- Ben-Menachem, E.; Hamberger, A.; Hedner, T.; Hammond, E.J.; Uthman, B.M.; Slater, J.; Treig, T.; Stefan, H.; Ramsay, R.E.; Wernicke, J.F.; et al. Effects of Vagus Nerve Stimulation on Amino Acids and Other Metabolites in the CSF of Patients with Partial Seizures. Epilepsy Res 1995, 20, 221–227. [Google Scholar] [CrossRef]
- Junker, B. Fermentation. In Kirk-Othmer Encyclopedia of Chemical Technology; Wiley, 2004.
- Steinkraus, K.H. Fermentations in World Food Processing. Compr Rev Food Sci Food Saf 2002, 1, 23–32. [Google Scholar] [CrossRef]
- Henderson, J.S.; Joyce, R.A.; Hall, G.R.; Hurst, W.J.; McGovern, P.E. Chemical and Archaeological Evidence for the Earliest Cacao Beverages. Proc Natl Acad Sci U S A 2007, 104, 18937–18940. [Google Scholar] [CrossRef]
- McGovern Anticancer Activity of Botanical Compounds in Ancient Fermented Beverages (Review). Int J Oncol 2010, 37. [CrossRef]
- McGovern, P.E.; Zhang, J.; Tang, J.; Zhang, Z.; Hall, G.R.; Moreau, R.A.; Nuñez, A.; Butrym, E.D.; Richards, M.P.; Wang, C.; et al. Fermented Beverages of Pre- and Proto-Historic China. Proceedings of the National Academy of Sciences 2004, 101, 17593–17598. [Google Scholar] [CrossRef]
- Caplice, E. Food Fermentations: Role of Microorganisms in Food Production and Preservation. Int J Food Microbiol 1999, 50, 131–149. [Google Scholar] [CrossRef]
- C. Borresen, E.; J. Henderson, A.; Kumar, A.; L. Weir, T.; P. Ryan, E. Fermented Foods: Patented Approaches and Formulations for Nutritional Supplementation and Health Promotion. Recent Patents on Food, Nutrition & Agriculturee 2012, 4, 134–140. [Google Scholar] [CrossRef]
- Selhub, E.M.; Logan, A.C.; Bested, A.C. Fermented Foods, Microbiota, and Mental Health: Ancient Practice Meets Nutritional Psychiatry. J Physiol Anthropol 2014, 33, 2. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, B.H. Depression as a Disease of Modernity: Explanations for Increasing Prevalence. J Affect Disord 2012, 140, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Logan, A.C.; Selhub, E.M. Vis Medicatrix Naturae: Does Nature “Minister to the Mind”? Biopsychosoc Med 2012, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Villegas, A.; Martínez-González, M.A. Diet, a New Target to Prevent Depression? BMC Med 2013, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Li, N.P.; van Vugt, M.; Colarelli, S.M. The Evolutionary Mismatch Hypothesis: Implications for Psychological Science. Curr Dir Psychol Sci 2018, 27, 38–44. [Google Scholar] [CrossRef]
- Chang, Y.; Durante, K.M. Why Consumers Have Everything but Happiness: An Evolutionary Mismatch Perspective. Curr Opin Psychol 2022, 46, 101347. [Google Scholar] [CrossRef] [PubMed]
- Hesseltine, C.W.; Wang, H.L. Traditional Fermented Foods. Biotechnol Bioeng 1967, 9, 275–288. [Google Scholar] [CrossRef]
- Takashima, N.; Katayama, A.; Dokai Mochimasu, K.; Hishii, S.; Suzuki, H.; Miyatake, N. A Pilot Study of the Relationship between Diet and Mental Health in Community Dwelling Japanese Women. Medicina (B Aires) 2019, 55, 513. [Google Scholar] [CrossRef]
- Ventriglio, A.; Sancassiani, F.; Contu, M.P.; Latorre, M.; Di Slavatore, M.; Fornaro, M.; Bhugra, D. Mediterranean Diet and Its Benefits on Health and Mental Health: A Literature Review. Clinical Practice & Epidemiology in Mental Health 2020, 16, 156–164. [Google Scholar] [CrossRef]
- Koga, M.; Toyomaki, A.; Miyazaki, A.; Nakai, Y.; Yamaguchi, A.; Kubo, C.; Suzuki, J.; Ohkubo, I.; Shimizu, M.; Musashi, M.; et al. Mediators of the Effects of Rice Intake on Health in Individuals Consuming a Traditional Japanese Diet Centered on Rice. PLoS One 2017, 12, e0185816. [Google Scholar] [CrossRef]
- Nanri, A.; Kimura, Y.; Matsushita, Y.; Ohta, M.; Sato, M.; Mishima, N.; Sasaki, S.; Mizoue, T. Dietary Patterns and Depressive Symptoms among Japanese Men and Women. Eur J Clin Nutr 2010, 64, 832–839. [Google Scholar] [CrossRef]
- Nanri, A.; Mizoue, T.; Poudel-Tandukar, K.; Noda, M.; Kato, M.; Kurotani, K.; Goto, A.; Oba, S.; Inoue, M.; Tsugane, S. Dietary Patterns and Suicide in Japanese Adults: The Japan Public Health Center-Based Prospective Study. British Journal of Psychiatry 2013, 203, 422–427. [Google Scholar] [CrossRef]
- Suzuki, T.; Miyaki, K.; Tsutsumi, A.; Hashimoto, H.; Kawakami, N.; Takahashi, M.; Shimazu, A.; Inoue, A.; Kurioka, S.; Kakehashi, M.; et al. Japanese Dietary Pattern Consistently Relates to Low Depressive Symptoms and It Is Modified by Job Strain and Worksite Supports. J Affect Disord 2013, 150, 490–498. [Google Scholar] [CrossRef]
- Sanada, M.; Imai, T.; Sezaki, A.; Miyamoto, K.; Kawase, F.; Shirai, Y.; Abe, C.; Suzuki, N.; Inden, A.; Kato, T.; et al. Changes in the Association between the Traditional Japanese Diet Score and Suicide Rates over 26 Years: A Global Comparative Study. J Affect Disord 2021, 294, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Murooka, Y.; Yamshita, M. Traditional Healthful Fermented Products of Japan. J Ind Microbiol Biotechnol 2008, 35, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Jacka, F.N.; Mykletun, A.; Berk, M.; Bjelland, I.; Tell, G.S. The Association Between Habitual Diet Quality and the Common Mental Disorders in Community-Dwelling Adults. Psychosom Med 2011, 73, 483–490. [Google Scholar] [CrossRef]
- Jacka, F.N.; Pasco, J.A.; Mykletun, A.; Williams, L.J.; Hodge, A.M.; O’Reilly, S.L.; Nicholson, G.C.; Kotowicz, M.A.; Berk, M. Association of Western and Traditional Diets With Depression and Anxiety in Women. American Journal of Psychiatry 2010, 167, 305–311. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Delgado-Rodríguez, M.; Alonso, A.; Schlatter, J.; Lahortiga, F.; Majem, L.S.; Martínez-González, M.A. Association of the Mediterranean Dietary Pattern With the Incidence of Depression. Arch Gen Psychiatry 2009, 66, 1090. [Google Scholar] [CrossRef]
- Akbaraly, T.N.; Brunner, E.J.; Ferrie, J.E.; Marmot, M.G.; Kivimaki, M.; Singh-Manoux, A. Dietary Pattern and Depressive Symptoms in Middle Age. British Journal of Psychiatry 2009, 195, 408–413. [Google Scholar] [CrossRef]
- Skarupski, K.A.; Tangney, C.C.; Li, H.; Evans, D.A.; Morris, M.C. Mediterranean Diet and Depressive Symptoms among Older Adults over Time. J Nutr Health Aging 2013, 17, 441–445. [Google Scholar] [CrossRef]
- Rienks, J.; Dobson, A.J.; Mishra, G.D. Mediterranean Dietary Pattern and Prevalence and Incidence of Depressive Symptoms in Mid-Aged Women: Results from a Large Community-Based Prospective Study. Eur J Clin Nutr 2013, 67, 75–82. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Henríquez, P.; Bes-Rastrollo, M.; Doreste, J. Mediterranean Diet and Depression. Public Health Nutr 2006, 9, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Lang, U.E.; Beglinger, C.; Schweinfurth, N.; Walter, M.; Borgwardt, S. Nutritional Aspects of Depression. Cellular Physiology and Biochemistry 2015, 37, 1029–1043. [Google Scholar] [CrossRef]
- Yin, W.; Löf, M.; Chen, R.; Hultman, C.M.; Fang, F.; Sandin, S. Mediterranean Diet and Depression: A Population-Based Cohort Study. International Journal of Behavioral Nutrition and Physical Activity 2021, 18, 153. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Villegas, A.; Martínez-González, M.A.; Estruch, R.; Salas-Salvadó, J.; Corella, D.; Covas, M.I.; Arós, F.; Romaguera, D.; Gómez-Gracia, E.; Lapetra, J.; et al. Mediterranean Dietary Pattern and Depression: The PREDIMED Randomized Trial. BMC Med 2013, 11, 208. [Google Scholar] [CrossRef] [PubMed]
- Bayes, J.; Schloss, J.; Sibbritt, D. A Randomised Controlled Trial Assessing the Effect of a Mediterranean Diet on the Symptoms of Depression in Young Men (the ‘AMMEND’ Study): A Study Protocol. British Journal of Nutrition 2021, 126, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Villegas, A.; Cabrera-Suárez, B.; Molero, P.; González-Pinto, A.; Chiclana-Actis, C.; Cabrera, C.; Lahortiga-Ramos, F.; Florido-Rodríguez, M.; Vega-Pérez, P.; Vega-Pérez, R.; et al. Preventing the Recurrence of Depression with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil. The PREDI-DEP Trial: Study Protocol. BMC Psychiatry 2019, 19, 63. [Google Scholar] [CrossRef]
- Parletta, N.; Zarnowiecki, D.; Cho, J.; Wilson, A.; Bogomolova, S.; Villani, A.; Itsiopoulos, C.; Niyonsenga, T.; Blunden, S.; Meyer, B.; et al. A Mediterranean-Style Dietary Intervention Supplemented with Fish Oil Improves Diet Quality and Mental Health in People with Depression: A Randomized Controlled Trial (HELFIMED). Nutr Neurosci 2019, 22, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Opie, R.S.; O’Neil, A.; Jacka, F.N.; Pizzinga, J.; Itsiopoulos, C. A Modified Mediterranean Dietary Intervention for Adults with Major Depression: Dietary Protocol and Feasibility Data from the SMILES Trial. Nutr Neurosci 2018, 21, 487–501. [Google Scholar] [CrossRef]
- Wade, A.T.; Davis, C.R.; Dyer, K.A.; Hodgson, J.M.; Woodman, R.J.; Keage, H.A.D.; Murphy, K.J. A Mediterranean Diet Supplemented with Dairy Foods Improves Mood and Processing Speed in an Australian Sample: Results from the MedDairy Randomized Controlled Trial. Nutr Neurosci 2020, 23, 646–658. [Google Scholar] [CrossRef]
- Morris, L.; Bhatnagar, D. The Mediterranean Diet. Curr Opin Lipidol 2016, 27, 89–91. [Google Scholar] [CrossRef]
- Davis, C.; Bryan, J.; Hodgson, J.; Murphy, K. Definition of the Mediterranean Diet; a Literature Review. Nutrients 2015, 7, 9139–9153. [Google Scholar] [CrossRef]
- Naureen, Z.; Bonetti, G.; Medori, M.C.; Aquilanti, B.; Velluti, V.; Matera, G.; Iaconelli, A.; Bertelli, M. Foods of the Mediterranean Diet: Lacto-Fermented Food, the Food Pyramid and Food Combinations. J Prev Med Hyg 2022, 63, E28–E35. [Google Scholar] [CrossRef]
- Briguglio, M.; Dell’Osso, B.; Panzica, G.; Malgaroli, A.; Banfi, G.; Zanaboni Dina, C.; Galentino, R.; Porta, M. Dietary Neurotransmitters: A Narrative Review on Current Knowledge. Nutrients 2018, 10, 591. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J Funct Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, P.; Pan, D.; Zeng, X.; Guo, Y.; Zhao, G. Effect of Adzuki Bean Sprout Fermented Milk Enriched in γ-Aminobutyric Acid on Mild Depression in a Mouse Model. J Dairy Sci 2021, 104, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Wan-Mohtar, W.A.A.Q.I.; Sohedein, M.N.A.; Ibrahim, M.F.; Ab Kadir, S.; Suan, O.P.; Weng Loen, A.W.; Sassi, S.; Ilham, Z. Isolation, Identification, and Optimization of γ-Aminobutyric Acid (GABA)-Producing Bacillus Cereus Strain KBC from a Commercial Soy Sauce Moromi in Submerged-Liquid Fermentation. Processes 2020, 8, 652. [Google Scholar] [CrossRef]
- Linares, D.M.; O’Callaghan, T.F.; O’Connor, P.M.; Ross, R.P.; Stanton, C. Streptococcus Thermophilus APC151 Strain Is Suitable for the Manufacture of Naturally GABA-Enriched Bioactive Yogurt. Front Microbiol 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Chamignon, C.; Mhedbi-Hajri, N.; Chain, F.; Derrien, M.; Escribano-Vázquez, U.; Garault, P.; Cotillard, A.; Pham, H.P.; Chervaux, C.; et al. The Potential Probiotic Lactobacillus Rhamnosus CNCM I-3690 Strain Protects the Intestinal Barrier by Stimulating Both Mucus Production and Cytoprotective Response. Sci Rep 2019, 9, 5398. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.-W.; Cheng, M.-Y.; Yang, X.; Lu, Y.-Y.; Yin, H.-D.; Zeng, Y.; Wang, R.-Y.; Jiang, Y.-L.; Yang, W.-T.; Wang, J.-Z.; et al. Probiotic Lactobacillus Rhamnosus GG Promotes Mouse Gut Microbiota Diversity and T Cell Differentiation. Front Microbiol 2020, 11, 607735. [Google Scholar] [CrossRef]
- Janik, R.; Thomason, L.A.M.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic Resonance Spectroscopy Reveals Oral Lactobacillus Promotion of Increases in Brain GABA, N-Acetyl Aspartate and Glutamate. Neuroimage 2016, 125, 988–995. [Google Scholar] [CrossRef]
- Slykerman, R.F.; Hood, F.; Wickens, K.; Thompson, J.M.D.; Barthow, C.; Murphy, R.; Kang, J.; Rowden, J.; Stone, P.; Crane, J.; et al. Effect of Lactobacillus Rhamnosus HN001 in Pregnancy on Postpartum Symptoms of Depression and Anxiety: A Randomised Double-Blind Placebo-Controlled Trial. EBioMedicine 2017, 24, 159–165. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA Production and Structure of GadB/GadC Genes in Lactobacillus and Bifidobacterium Strains from Human Microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef]
- Zhou, B.; Jin, G.; Pang, X.; Mo, Q.; Bao, J.; Liu, T.; Wu, J.; Xie, R.; Liu, X.; Liu, J.; et al. Lactobacillus Rhamnosus GG Colonization in Early Life Regulates Gut-Brain Axis and Relieves Anxiety-like Behavior in Adulthood. Pharmacol Res 2022, 177, 106090. [Google Scholar] [CrossRef]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C.G. Use of Selected Lactic Acid Bacteria and Quinoa Flour for Manufacturing Novel Yogurt-Like Beverages. Foods 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Hashemi Gahruie, H.; Eskandari, M.H.; Mesbahi, G.; Hanifpour, M.A. Scientific and Technical Aspects of Yogurt Fortification: A Review. Food Science and Human Wellness 2015, 4, 1–8. [Google Scholar] [CrossRef]
- McKinley, M.C. The Nutrition and Health Benefits of Yoghurt. Int J Dairy Technol 2005, 58, 1–12. [Google Scholar] [CrossRef]
- Lee, X.Y.; Tan, J.S.; Cheng, L.H. Gamma Aminobutyric Acid (GABA) Enrichment in Plant-Based Food – A Mini Review. Food Reviews International 2022, 1–22. [Google Scholar] [CrossRef]
- Nikmaram, N.; Dar, B.; Roohinejad, S.; Koubaa, M.; Barba, F.J.; Greiner, R.; Johnson, S.K. Recent Advances in γ -Aminobutyric Acid (GABA) Properties in Pulses: An Overview. J Sci Food Agric 2017, 97, 2681–2689. [Google Scholar] [CrossRef]
- Inoue, K.; Shirai, T.; Ochiai, H.; Kasao, M.; Hayakawa, K.; Kimura, M.; Sansawa, H. Blood-Pressure-Lowering Effect of a Novel Fermented Milk Containing γ-Aminobutyric Acid (GABA) in Mild Hypertensives. Eur J Clin Nutr 2003, 57, 490–495. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J Funct Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as Food Supplements: The Effects of GABA on Brain and Behavior. Front Psychol 2015, 6. [Google Scholar] [CrossRef]
- TSUKADA, Y.; NAGATA, Y.; HIRANO, S. Active Transport of Gamma-Aminobutyric Acid in Brain Cortex Slices, with Special Reference to Phosphorus-32 Turnover of Phospholipids in Cytoplasmic Particulates. Nature 1960, 186, 474–475. [Google Scholar] [CrossRef]
- Gelder, N.M.; Elliott, K.A.C. DISPOSITION OF γ-AMINOBUTYRIC ACID ADMINISTERED TO MAMMALS. J Neurochem 1958, 3, 139–143. [Google Scholar] [CrossRef]
- Takanaga, H.; Ohtsuki, S.; Hosoya, Ki; Terasaki, T. GAT2/BGT-1 as a System Responsible for the Transport of Gamma-Aminobutyric Acid at the Mouse Blood-Brain Barrier. J Cereb Blood Flow Metab 2001, 21, 1232–1239. [Google Scholar] [CrossRef]
- Hossain, S.J.; Hamamoto, K.; Aoshima, H.; Hara, Y. Effects of Tea Components on the Response of GABA A Receptors Expressed in Xenopus Oocytes. J Agric Food Chem 2002, 50, 3954–3960. [Google Scholar] [CrossRef]
- THANAPREEDAWAT, P.; KOBAYASHI, H.; INUI, N.; SAKAMOTO, K.; KIM, M.; YOTO, A.; YOKOGOSHI, H. GABA Affects Novel Object Recognition Memory and Working Memory in Rats. J Nutr Sci Vitaminol (Tokyo) 2013, 59, 152–157. [Google Scholar] [CrossRef]
- Yoto, A.; Murao, S.; Motoki, M.; Yokoyama, Y.; Horie, N.; Takeshima, K.; Masuda, K.; Kim, M.; Yokogoshi, H. Oral Intake of γ-Aminobutyric Acid Affects Mood and Activities of Central Nervous System during Stressed Condition Induced by Mental Tasks. Amino Acids 2012, 43, 1331–1337. [Google Scholar] [CrossRef]
- Smith, A.K.; Wade, A.R.; Penkman, K.E.; Baker, D.H. Dietary Modulation of Cortical Excitation and Inhibition. Journal of Psychopharmacology 2017, 31, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.J.; Cooper, M.A.; Simmons, R.D.J.; Weir, C.J.; Belelli, D. Neurosteroids: Endogenous Allosteric Modulators of GABAA Receptors. Psychoneuroendocrinology 2009, 34, S48–S58. [Google Scholar] [CrossRef] [PubMed]
- Hinton, T.; Jelinek, H.F.; Viengkhou, V.; Johnston, G.A.; Matthews, S. Effect of GABA-Fortified Oolong Tea on Reducing Stress in a University Student Cohort. Front Nutr 2019, 6. [Google Scholar] [CrossRef]
- Waagepetersen, H.S.; Sonnewald, U.; Schousboe, A. The GABA Paradox. J Neurochem 2002, 73, 1335–1342. [Google Scholar] [CrossRef] [PubMed]
- Aoshima, H.; Tenpaku, Y. Modulation of GABA Receptors Expressed in Xenopus Oocytes by 13- <scp>l</Scp> -Hydoxylinoleic Acid and Food Additives. Biosci Biotechnol Biochem 1997, 61, 2051–2057. [Google Scholar] [CrossRef]
- Oh, S.-H.; Moon, Y.-J.; Oh, C.-H. γ -Aminobutyric Acid (GABA) Content of Selected Uncooked Foods. Prev Nutr Food Sci 2003, 8, 75–78. [Google Scholar] [CrossRef]
- Yamatsu, A.; Yamashita, Y.; Pandharipande, T.; Maru, I.; Kim, M. Effect of Oral γ-Aminobutyric Acid (GABA) Administration on Sleep and Its Absorption in Humans. Food Sci Biotechnol 2016, 25, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.-I.; Shin, Y.Y.; Chung, S.-E.; Shin, W.C. Safety and Efficacy of Gamma-Aminobutyric Acid from Fermented Rice Germ in Patients with Insomnia Symptoms: A Randomized, Double-Blind Trial. Journal of Clinical Neurology 2018, 14, 291. [Google Scholar] [CrossRef] [PubMed]
- KANEHIRA, T.; NAKAMURA, Y.; NAKAMURA, K.; HORIE, K.; HORIE, N.; FURUGORI, K.; SAUCHI, Y.; YOKOGOSHI, H. Relieving Occupational Fatigue by Consumption of a Beverage Containing Γ-Amino Butyric Acid. J Nutr Sci Vitaminol (Tokyo) 2011, 57, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of Gaba (γ - Aminobutyric Acid) by Microorganisms: A Review. Brazilian Journal of Microbiology 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.N.S.; Ryu, J.; Kim, Y.; Jeon, B.H. GABA-Enriched Fermented Laminaria Japonica Improves Cognitive Impairment and Neuroplasticity in Scopolamine- and Ethanol-Induced Dementia Model Mice. Nutr Res Pract 2018, 12, 199. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-Derived Neurotrophic Factor (BDNF) Is Required for the Enhancement of Hippocampal Neurogenesis Following Environmental Enrichment. European Journal of Neuroscience 2006, 24, 1850–1856. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF Val66met Polymorphism Affects Activity-Dependent Secretion of BDNF and Human Memory and Hippocampal Function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front Cell Neurosci 2019, 13. [Google Scholar] [CrossRef]
- Park, H.-J.; Lee, M.-S.; Shim, H.S.; Lee, G.-R.; Chung, S.Y.; Kang, Y.M.; Lee, B.-J.; Seo, Y.B.; Kim, K.S.; Shim, I. Fermented Saccharina Japonica (Phaeophyta) Improves Neuritogenic Activity and TMT-Induced Cognitive Deficits in Rats. ALGAE 2016, 31, 73–84. [Google Scholar] [CrossRef]
- van de Wouw, M.; Walsh, A.M.; Crispie, F.; van Leuven, L.; Lyte, J.M.; Boehme, M.; Clarke, G.; Dinan, T.G.; Cotter, P.D.; Cryan, J.F. Distinct Actions of the Fermented Beverage Kefir on Host Behaviour, Immunity and Microbiome Gut-Brain Modules in the Mouse. Microbiome 2020, 8, 67. [Google Scholar] [CrossRef]
- Tınok, A.A.; Karabay, A.; Jong, J.D.; Balta, G.; Akyürek, E.G. Effects of Gamma-Aminobutyric Acid on Working Memory and Attention: A Randomized, Double-Blinded, Placebo-Controlled, Crossover Trial. J Psychopharmacol 2023, 37, 554–565. [Google Scholar] [CrossRef]
- Leonte, A.; Colzato, L.S.; Steenbergen, L.; Hommel, B.; Akyürek, E.G. Supplementation of Gamma-Aminobutyric Acid (GABA) Affects Temporal, but Not Spatial Visual Attention. Brain Cogn 2018, 120, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Małus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus Plantarum 299v Decreases Kynurenine Concentration and Improves Cognitive Functions in Patients with Major Depression: A Double-Blind, Randomized, Placebo Controlled Study. Psychoneuroendocrinology 2019, 100, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Doll, J.P.K.; Schweinfurth, N.; Kettelhack, C.; Schaub, A.-C.; Yamanbaeva, G.; Varghese, N.; Mählmann, L.; Brand, S.; Eckert, A.; et al. Effect of Short-Term, High-Dose Probiotic Supplementation on Cognition, Related Brain Functions and BDNF in Patients with Depression: A Secondary Analysis of a Randomized Controlled Trial. Journal of Psychiatry and Neuroscience 2023, 48, E23–E33. [Google Scholar] [CrossRef]
- Shyamaladevi, N.; Jayakumar, A.R.; Sujatha, R.; Paul, V.; Subramanian, E.H. Evidence That Nitric Oxide Production Increases γ-Amino Butyric Acid Permeability of Blood-Brain Barrier. Brain Res Bull 2002, 57, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Takishima, T.; Kometani, T.; Yokogoshi, H. Psychological Stress-Reducing Effect of Chocolate Enriched with γ-Aminobutyric Acid (GABA) in Humans: Assessment of Stress Using Heart Rate Variability and Salivary Chromogranin A. Int J Food Sci Nutr 2009, 60, 106–113. [Google Scholar] [CrossRef]
- Gamma-Aminobutyric Acid (GABA), Monograph. Altern Med Rev 2007, 12, 274–279.
Figure 1.
The diagram illustrates the intricate relationship between the depressive gut-brain axis and the impact of GABA on this axis. Prolonged exposure to chronic stress activates the hypothalamic-pituitary-adrenal (HPA) axis, resulting in the production of cortisol. This hormonal response disrupts the equilibrium of the gut microbiota and subsequently leads to increased permeability of the intestinal lining, commonly referred to as “leaky gut.” Consequently, potentially harmful substances, notably lipopolysaccharides (LPS), gain access to the brain. Moreover, cortisol not only triggers an inflammatory reaction but also activates the endocannabinoid system. Concurrently, the disturbed gut microbiota generates various bioactive compounds, including neuropeptides, hormones, and short-chain fatty acids (SCFAs). The effects of these substances, in conjunction with those of inflammatory mediators, are predominantly mediated through the vagus nerve. The stress-induced activation of the HPA axis contributes to dysbiosis and heightened intestinal permeability, exacerbating the “leaky gut” condition. Consequently, proinflammatory cytokines further stimulate the HPA axis by influencing vagal nuclei (NTS/DMN) while also modulating the tryptophan-kynurenine pathway and altering neurotransmitter metabolism. It is noteworthy that GABA is not only consumed but also synthesized directly by specific genera of gut bacteria, particularly Bifidobacterium and Lactobacillus. Therefore, the administration of GABA, such as through fortified foods or probiotic bacteria capable of producing GABA, holds the potential to restore the equilibrium of the gut microbiome. This restoration may facilitate the normalization of gut-brain communication via the enteric nervous system and vagus nerve, ultimately alleviating stress-related effects in the brain and improving depressive symptoms. Abbreviations: GABA: Gamma-Aminobutyric Acid; CRH: Corticotropin-Releasing Hormone; ACTH: Adrenocorticotropic Hormone; SCFAs: Short-Chain Fatty Acids.
Figure 1.
The diagram illustrates the intricate relationship between the depressive gut-brain axis and the impact of GABA on this axis. Prolonged exposure to chronic stress activates the hypothalamic-pituitary-adrenal (HPA) axis, resulting in the production of cortisol. This hormonal response disrupts the equilibrium of the gut microbiota and subsequently leads to increased permeability of the intestinal lining, commonly referred to as “leaky gut.” Consequently, potentially harmful substances, notably lipopolysaccharides (LPS), gain access to the brain. Moreover, cortisol not only triggers an inflammatory reaction but also activates the endocannabinoid system. Concurrently, the disturbed gut microbiota generates various bioactive compounds, including neuropeptides, hormones, and short-chain fatty acids (SCFAs). The effects of these substances, in conjunction with those of inflammatory mediators, are predominantly mediated through the vagus nerve. The stress-induced activation of the HPA axis contributes to dysbiosis and heightened intestinal permeability, exacerbating the “leaky gut” condition. Consequently, proinflammatory cytokines further stimulate the HPA axis by influencing vagal nuclei (NTS/DMN) while also modulating the tryptophan-kynurenine pathway and altering neurotransmitter metabolism. It is noteworthy that GABA is not only consumed but also synthesized directly by specific genera of gut bacteria, particularly Bifidobacterium and Lactobacillus. Therefore, the administration of GABA, such as through fortified foods or probiotic bacteria capable of producing GABA, holds the potential to restore the equilibrium of the gut microbiome. This restoration may facilitate the normalization of gut-brain communication via the enteric nervous system and vagus nerve, ultimately alleviating stress-related effects in the brain and improving depressive symptoms. Abbreviations: GABA: Gamma-Aminobutyric Acid; CRH: Corticotropin-Releasing Hormone; ACTH: Adrenocorticotropic Hormone; SCFAs: Short-Chain Fatty Acids.
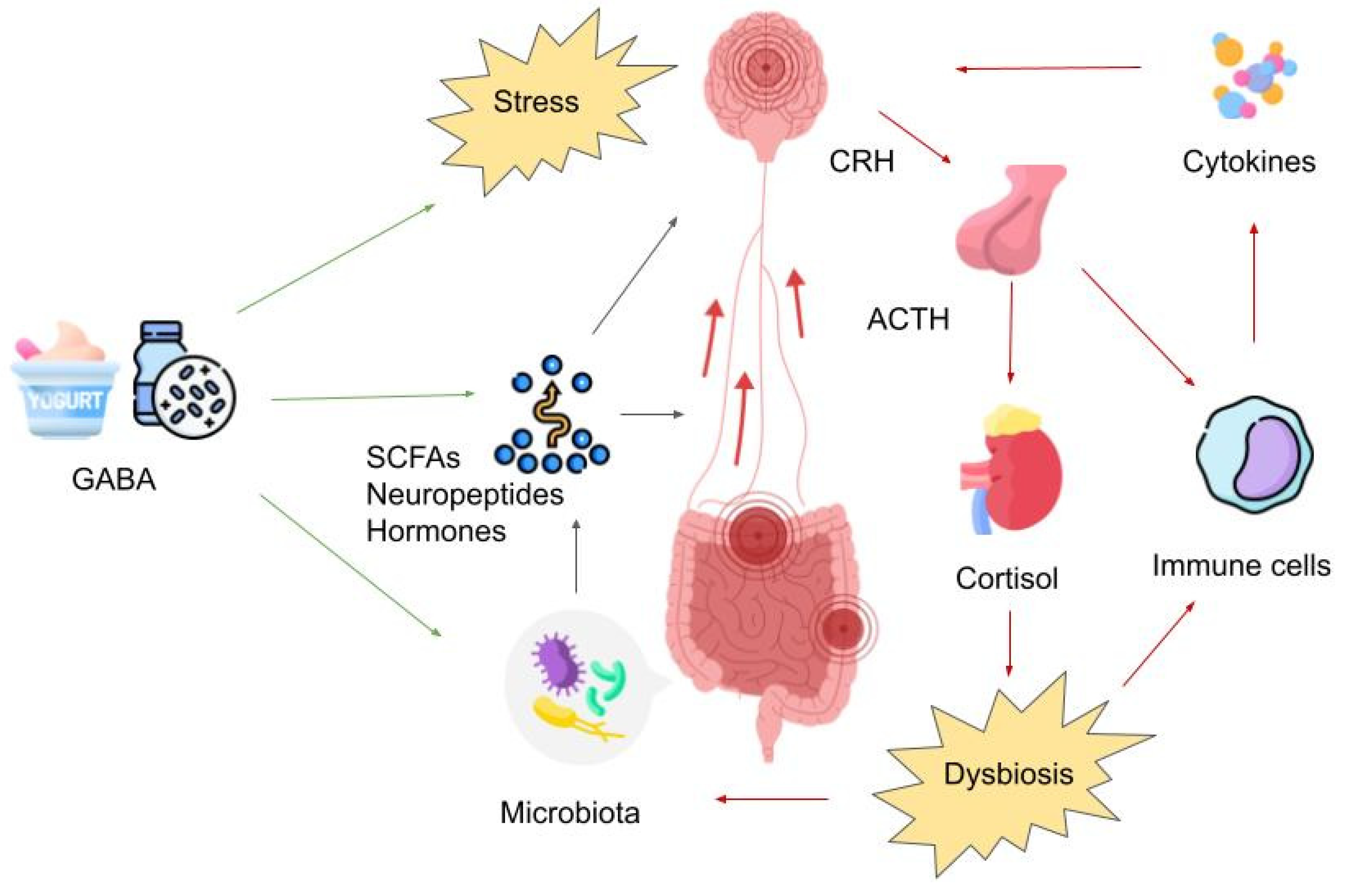
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



