Submitted:
19 October 2023
Posted:
19 October 2023
You are already at the latest version
Abstract
We evaluated the transcriptional response of interferon (IFN)-related genes in rock bream iridovirus (RBIV)-infected rock bream under high-, low-, or no-mortality conditions induced by different stocking water temperatures. Under the high susceptibility condition (group A, water temperature 26 °C, 100% mortality), only the Mx gene was expressed early, with prolonged expression, and with heavy viral loads of approximately 106–107 major capsid protein gene copies/μL from 4 to 10 days post infection (dpi). However, IRF1, IRF3, IRF8, STAT1, ISG15, PKR, Viperin, GVIN1, IFI44, and ISG56 were activated at later time points (8 dpi) and then quickly decreased (10 dpi). For the low susceptibility condition, the water temperature was set at 23 °C for 7 days (group B) and then reduced to 17 °C. Group B exhibited a 28% mortality rate, in which persistent and effective antiviral responses were observed for long periods of time. In particular, at 20 and 22 dpi, when virus replication was peaked at approximately 107/μL, the expressions of most of the IFN-related genes (IRF1, IRF3, IRF8, Mx, STAT1, ISG15, PKR, Viperin, GVIN1, IFI44, and ISG56) were significantly higher in group B than in the control group. Moreover, prolonged and higher levels of IRF3 (at least 30 dpi), IRF8 (at least 30 dpi), ISG15 (at least 30 dpi), PKR (at least 28 dpi), Viperin (at least 30 dpi), and IFI44 (at least 30 dpi) were also observed in the recovery stage of infection. Under the no-susceptibility condition at 17 °C (0% mortality), significantly elevated levels of IRF3, Mx, ISG15, and PKR were observed mostly until 20 dpi. The findings indicate that RBIV infection can induce an efficient IFN-mediated antiviral immune response in low- and no-susceptibility conditions. The findings could be valuable for effective control of viral pathogens in fish.
Keywords:
rock bream
; rock bream iridovirus
; water temperature
; virus replication
; inteferons
1. Introduction
Rock bream iridovirus (RBIV) belongs to the genus Megalocytivirus [1]. In Korea, an outbreak of RBIV in rock bream (Oplegnathus fasciatus) was first reported in the summer of 1998 in southern coastal areas with high water temperatures (23–27 °C) [2]. Since the first outbreak, high RBIV-related mortality has occurred annually in rock bream. This remains an important health problem in the rock bream aquaculture industry.
Water temperature is closely related to host immune responses and the susceptibility of rock bream to RBIV infection [3,4,5,6,7]. RBIV infection of rock bream can result in pronounced pathogenicity [3]. A shift in water temperature from 23 °C to 17 °C, which does not cause mortality, allows rock bream to survive RBIV infection, with the survivors acquiring protective immunity [3]. However, once RBIV replicates at high susceptibility water temperatures over 23 °C, it is difficult to inactivate the virus in the fish. The surviving fish need at least 50 days from the peak virus level or death of other fish in the population to avoid re-activation of the virus, thereby completely and slowly eliminating the virus from their bodies [7]. This phenomenon shows that water temperature greatly affects the health status of RBIV-infected rock bream. Although water temperature shifting induces effective protection, the mechanism of protection remains unknown. Evaluation of antiviral immune responses needed to inhibit virus replication could identify important factor(s) for survival.
In general, the first line of defense against virus infection is based on type I interferon (IFN) expression [8], which leads to the transcription of many downstream genes. The expression of IFNs is regulated by transcription factors called IFN-regulatory factors (IRFs) [9]. IRFs were initially identified as factors that regulate the transcription of IFNs and IFN-stimulated genes (ISGs) [10]. Mx, ISG15, and protein kinase R (PKR) are stimulated by type I IFN [11,12,13]. IFN-inducible protein 44 (IFI44) is rapidly induced during viral infections and encodes an intracellular protein with anti-proliferative activity [14]. IFN-γ-inducible lysosomal thiol reductase 30 (GILT or IFI30) is constitutively expressed in most antigen presenting cells and is induced by IFN-γ in other cell types via signal transducer and activator of transcription (STAT) [15]. The IFN-inducible antiviral proteins inhibit viral replication during viral protein synthesis, virion assembly, and mRNA synthesis [16]. Furthermore, the IFN-related immune response in fish shows broad antiviral activity against many fish viruses [17,18,19,20,21,22,23].
The immune defense response in rock bream to RBIV infection currently remains unclear. Our recent studies described changes in the expressions of a number of genes in immune pathways after RBIV infection. These genes include toll-like receptor 9 (TLR9)-mediated myeloid differentiation primary response gene 88 (MyD88)-dependent genes, pro-inflammatory cytokines, apoptosis pathway genes, antigen presenting (MHC class I), and T cell response (cytotoxic CD8+ cell) genes [24,25,26]. We have also reported IFN-associated immune gene responses in rock bream with or without the administration of CpG oligonucleotide (ODN) 1668, polyinosinic:polycytidylic acid [poly (I:C)], and DNA vaccine [27,28,29]. Furthermore, type I IFN-related molecules have been identified and characterized in rock bream [30,31,32,33]. However, the role of the IFN, IRFs or ISGs pathways of rock bream during RBIV replication is still unclear.
This study was conducted as a continuation of our previous research [24,25] to investigate the basis of RBIV mortality under different water temperatures in rock bream, and to identify an RBIV immune defense mechanism. The effects of RBIV replication on IFN-related immune gene responses were investigated using rock bream sampled from high-mortality (virus infection at 26 °C), low-mortality (virus infection at 23 °C, shifted to 17 °C) and no-mortality (virus infection at 17 °C) groups. The findings emphasize the importance of genes related to IFN responses in rock bream immunity for protection against RBIV. These genes include IRF1, IRF3, IRF4, IRF8, STAT1, Mx, ISG15, PKR, Viperin, IFN-induced very large GTPase 1-like (GVIN1), IFN-induced protein 44-like (IFI44), IFN-inducible protein 56 (ISG56), and IFI30.
2. Materials and Methods
2.1. Virus Preparation
The virus aliquots originated from rock bream infected with RBIV in September 2010 [25]. The major capsid protein (MCP) gene copies of RBIV in the supernatant preparations were quantified by quantitative real-time polymerase chain reaction (qRT-PCR). The virus titer was calculated as 1.1 × 107/100 μL MCP gene copies. Although some studies have demonstrated the use of cell lines to culture Megalocytivirus, RBIV does not replicate well using in vitro cell culture conditions. In our experiment, we tried to culture RBIV using grunt fin (GF) cells but the titer was very low. The highest titer we obtained was 103 to 104 median tissue culture infectious dose (TCID50)/mL/100 mg spleen, which is approximately 108 to 109 MCP gene copies/100 mg spleen. Hence, the TCID50 method was not used in this study. Future studies that develop cell lines allowing high virus growth to compare TCID50 and qRT-PCR data will be essential.
2.2. Experimental Infection
2.2.1. High-Mortality Condition (water temperature fixed at 26 °C; group A)
RBIV-free rock bream were obtained from a local farm. The experimental design was identical to that described previously [24]. To evaluate the IFN-related immune gene expression under RBIV infection at 26 °C, each fish in each group of 50 fish (11.5 ± 0.7 cm, 29.5 ± 5.7 g) was intraperitoneally (i.p.) injected with 100 μL of either RBIV (1.1 × 107 MCP gene copies) or phosphate buffered saline (PBS). The fish were maintained in aquaria containing 250 L of ultraviolet light (UV)-treated seawater. Six fish were randomly chosen from each group at 1, 4, 8, and 10 days post infection (dpi) (Table 1). The head kidney and spleen were collected and then stored at –80 °C.
To determine the mortality rate, 20 fish (11.5 ± 0.7 cm, 29.5 ± 5.7 g) were i.p. injected with RBIV (1.1 × 107 MCP gene copies; 100 μL/fish) or PBS (100 μL/fish)as a control. The fish were maintained in aquaria containing 30 L of UV-treated seawater at 26 °C.
2.2.2. Low-Mortality Condition (water temperature Shifted from 23 °C to 17 °C; group B)
We had previously described that RBIV causes 100% mortality in rock bream at water temperatures of 20–26 °C and 0% mortality at a temperature of 17 °C [24]. In the present study, for group B, the water temperature was shifted from 23 °C to 17 °C to reduce mortality and to analyze the immune factors involved in fish survival. Fish (11.7 ± 1.7 cm, 34.1 ± 2.1 g) were i.p. injected with RBIV and PBS in the same manner as group A and were maintained in aquaria containing 250 L of UV-treated seawater. Sixty fish in the virus and PBS groups were maintained at 23 °C for 7 days prior to the shift of the water temperature to 17 °C at a rate of 2 °C per day. To determine immune gene expression and viral copy number, five fish of each groups were randomly chosen at 2, 4, 7, 10, 15, 20, 22, 25, 28, and 30 dpi. The head kidney and spleen were collected aseptically from each fish and stored at –80 °C until use.
2.2.3. No-Mortality Condition (water temperature fixed at 17 °C; group C)
The experimental design was explained previously [24]. Fifty fish (12.0 ± 1.2 cm, 35.0 ± 2.8 g) were i.p. injected with RBIV or PBS in the same manner as described for groups A and B, and were maintained in aquaria containing 250 L of UV-treated seawater. Five fish were randomly chosen from each group at 2, 4, 7, 10, 15, 20, 25, 30, and 40 dpi (Table 1). The head kidney and spleen were collected and then stored at –80 °C.
2.3. Quantitative expression of immune genes and determination of viral copy numbers
All quantitative real-time polymerase chain reaction (qRT-PCR) assays were performed using the Exicycler 96 Real-Time Quantitative Thermal Block (Bioneer, Korea) using a commercial master mix (AccuPre®2X Greenstar qPCR Master mix; Bioneer) and the previously described cycling condition [25].
For immune gene expression, total RNA was extracted from head kidney samples using RNAiso Plus reagent (TaKaRa Bio, Japan), followed by treatment with RNase-free recombinant DNase I (TaKaRa Bio) using the ReverTra Ace qPCR RT Kit (Toyobo, Japan) according to the manufacturers’ protocols. Each sample was run in duplicate with β-actin RNA as a control and quantified using the 2−ΔΔCt method [34]. The primers are listed in Table 2. The data were presented as the means ± standard errors. Statistical analysis of the expression levels of immune genes were carried out using the unpaired t-tests using the GraphPad Prism software (GraphPad Software, USA). Differences were considered statistically significant at p < 0.05, p < 0.01, or p < 0.001, depending on the experiment.
The spleen is one of the indicators for RBIV replication, since the RBIV MCP gene is easily detected in spleen tissue during RBIV infection [3,4,7,35]. To measure the absolute RBIV copy number, genomic DNA was extracted from whole spleen (20–150 mg) samples using an AccuPrep®Genomic DNA extraction kit (Bioneer) according to the manufacturer’s protocol. A standard curve was generated to determine the MCP gene copy number by quantitative PCR (qPCR) as previously described [25]. Virus copy number was determined from 1 μL of genomic DNA obtained from 100 μL of total genomic DNA.
3. Results
3.1. RBIV Levels and Fish Mortality
In the high susceptibility condition at 26 °C (group A), mortality was observed between 7 and 15 dpi. The final mortality was 100% (Figure 1). Clinical signs of RBIV disease were evident. The virus copy number gradually increased to 1.2 × 106 MCP gene copies/µL at 4 dpi and then quickly reached peak levels at 8 and 10 dpi (average range of 8.1 × 106 to 2.2 × 107 MCP gene copies/µL) (Figure 2).
In group B (23 °C reduced to 17 °C at 7 dpi), the virus copy number gradually increased until 7 dpi at 23 °C (average 7.1 × 105/µL). The copy number continued to increase even after the water temperature was reduced to 17 °C at 7 dpi, with maximum copy numbers at 22 dpi (average 1.12 × 107/µL) (Figure 2). Mortality was observed after 24, 25, 26, 27, and 35 dpi (one fish each day) with an estimated cumulative mortality of 4, 8, 13, 18, and 28%, respectively (Figure 1). The virus copy was reduced at 25 and 28 dpi (average 2.8 × 104 and 1.5 × 104/µL). However, the average virus copy number at 30 dpi (1.1 × 107/µL) and individual differences were high since complete inactivation of virus did not occur in the fish. Two fish displayed drastically reduced virus copy numbers (6.4 × 102 and 2.2 × 102/µL) and three fish displayed high virus copy numbers (8.2 × 105, 3.5 × 106, and 4.9 × 107/µL) (Figure 2).
In group C, all the fish survived without any clinical signs until 40 dpi (Figure 1). Virus copy numbers increased but remained relatively low with the highest occurrence at 20 and 25 dpi (average virus copy number of 2.7 × 104 and 1.8 × 104/µL, respectively). Virus copy numbers then gradually decreased until 40 dpi (below 102) (Figure 2).
3.2. IFN-related Genes and Their Expression at Different Stages of RBIV Progression
3.2.1. Expression in Group A
The induced levels of expression of the IRF1, IRF3, IRF4, IRF8, STAT1, Mx, ISG15, PKR, Viperin, GVIN1, IFI44, ISG56, and IFI30 genes varied (Figure 3). Similar gene expression trends were observed for several immune genes in the artificial infection experiment. Significantly higher IRF1, IRF3, STAT1, ISG15, Viperin, IFI44, and IRF56 expression levels were observed at 4 dpi (5.8-, 23.6-, 6.5-, 13.3-, 4.8-, 8.2-, and 47.7-fold, respectively; p < 0.05 and p < 0.01) and 8 dpi (18.9-, 52.2-, 8.5-, 79.9, 21.4, 21.7-, and 158.6-fold, respectively; p < 0.05, p < 0.01, and p < 0.001) compared with the control group (Figure 3A, B, E, G, I, K, and L). Slightly increased IRF4, IRF8, PKR, and GVIN1 gene expressions were observed at 4 dpi (1.3-, 1.3-, 2.1-, and 12.1-fold, respectively). Significantly increased expression of the IRF4, IRF8, PKR, and GVIN1 genes were observed at 8 dpi (3.8-, 3.5-, 5.2-, and 81.8-fold, respectively; p < 0.05 and p < 0.01) (C, D, H, and J). However, IRF1, IRF3, IRF4, IRF8, STAT1, ISG15, PKR, Viperin, GVIN1, IFI44, and IRF56 gene expressions decreased further starting at 10 dpi (1.6-, 4.1-, 0.3-, 0.9-, 0.5-, 4.5-, 0.5-, 2.7-, 7.7-, 0.9-, and 19.3-fold, respectively). These changes were not statistically significant. Subsequently, the RBIV copy number increased. The IFI30 expression level was changed <1.0-fold and was not statistically significant (Figure 3M). Interestingly, only the Mx mRNA expression was significantly prolonged in RBIV-infected fish at 4 dpi (40.1-fold), 8 dpi (49.8-fold), and 10 dpi (36.6-fold) (p < 0.01) (Figure 3F).
3.2.2. Expression in Group B
In group B, most of the IFN-related molecules were highly activated during the progression of RBIV in rock bream. A statistically significant increase in IRF1 expression was observed at 10 dpi (3.6-fold), 15 dpi (4.8-fold), 22 dpi (4.6-fold), and 25 dpi (3.5-fold) (p < 0.05 and p < 0.01) (Figure 4A). IRF3 exhibited significantly increased expression compared with the control at 15 dpi (19.4-fold), 20 dpi (14.1-fold), 22 dpi (10.4-fold), and 25 dpi (3.6-fold) (p < 0.05) (Figure 4B). Significantly higher (p < 0.05) IRF8 expression levels were observed at 20 dpi (5.6-fold), 22 dpi (2.2-fold), 25 dpi (2.3-fold), and 30 dpi (2.2-fold) compared with the control group (Figure 4D). A statistically significant increase in Mx expression was observed at 7 dpi (7.1-fold), 10 dpi (5.8-fold), 15 dpi (9.2-fold), 20 dpi (7.4-fold), 22 dpi (17.1-fold), and 25 dpi (3.8-fold) (p < 0.05) (Figure 4F). ISG15 exhibited significantly increased expression compared with the control (5.9-, 14.2-, 2.7-, 2.6-, 2.2-, and 6.0-fold) at 15, 20, 22, 25, 28, and 30 dpi, respectively (Figure 4G). The significantly increased expression levels of PKR ranged between 1.9- and 2.4-fold (15 to 28 dpi) in group B (Figure 4H). Significantly higher levels of STAT1 (3.1- and 4.5-fold at 15 and 20 dpi, respectively; p < 0.05), Viperin (5.6-, 8.8-, 3.9-, and 3.9-fold at 15, 20, 22, and 30 dpi, respectively; p < 0.05 and p < 0.01), GVIN1 (7.1-, 5.3-, and 9.3-fold at 10, 20, and 22 dpi, respectively; p < 0.01 and p < 0.001), IFI44 (3.4-, 9.3-, 4.5-, 2.0-, and 4.4-fold at 15, 20, 22, 25, and 30 dpi, respectively; p < 0.05 and p < 0.01) and ISG56 (2.6-, 4.0-, 4.6-, and 6.6-fold at 10, 15, 20 and 22 dpi, respectively; p < 0.05, p < 0.01, and p < 0.001) (Figure 4E, I, J, K, and L).
3.2.3. Expression in Group C
Significantly higher levels of IRF3 (15.4- and 11.9-fold at 7 and 10 dpi, respectively; p < 0.05), Mx (4.2-fold at 10 dpi; p < 0.05), ISG15 (6.0-, 8.4-, 10.4-, 9.8-, and 11.0-fold at 4, 7, 10, 15, and 20 dpi, respectively; p < 0.05), PKR (3.3-, 5.2-, and 6.0-fold at 2, 4, and 7 dpi, respectively; p < 0.05), and Viperin (2.8-fold at 10 dpi; p < 0.05) expression were observed (Figure 5B, F, G, H, and I). However, during the experimental period, the IRF1, IRF4, IRF8, STAT1, GVIN1, IFI44, ISG56, and IFI30 expression levels were not statistically significant (Figure 5A, C, D, E, J, K, L, and M).
4. Discussion
IFNs are released by a virus-infected cells. This causes nearby cells to heighten their antiviral defenses. Synthetic IFN inducers such as poly (I:C) and CpG ODNs drastically reduce mortality against fish viruses, including infectious pancreatic necrosis virus (IPNV), nervous necrosis virus (NNV), and viral hemorrhagic septicemia virus (VHSV) [36,37,38,39]. Therefore, IFN-mediated responses appear to activate an antiviral state to inhibit virus replication to protect fish from virus infection. However, in orange-spotted grouper (Epinephelus coioides), while ISG15 did not reportedly reduce the transcription of Singapore grouper iridovirus (SGIV), ISG15 significantly inhibited the replication of grouper nervous necrosis virus [40]. The findings indicate that the IFN-related response does not confer protection against all fish viruses, with possible virus-specific roles in viral infections of different fish species. Therefore, in the present study, the transcriptional response of IFN-related genes was evaluated to determine whether the important immune responses in RBIV-infected rock bream (high-, low-, and no-mortality conditions) could be activated and virus replication inhibited.
For group A, in the high susceptibility condition at 26 °C (100% mortality at 15 dpi), the virus replicated quickly, reaching its peak at 8 and 10 dpi (average range of 1.1 × 106 to 2.2 × 107/μL). At 4 dpi, IRF1, IRF3, STAT1, Mx, ISG15, Viperin, IFI44, and ISG56 were significantly activated. At 7 dpi, specific mortality began to be observed, and 12 IFN-related genes (IRF1, IRF3, IRF4, IRF8, STAT1, Mx, ISG15, PKR, Viperin, GVIN1, IFI44, and ISG56) had significantly elevated levels of expression at 8 dpi. However, at 10 dpi, while Mx expression levels remained significantly high, the expression levels of the IRF1, IRF3, IRF4, IRF8, ISG15, STAT1, PKR, Viperin, GVIN1, IFI44, and ISG56 genes decreased to basal levels after virus replication reached its peak (2.2 × 107/μL, which is a lethal level of virus copies). The present results indicate that, even though Mx-mediated antiviral immune response occurred quickly and was prolonged, the effect of IRF1, IRF3, IRF4, IRF8, STAT1, ISG15, PKR, Viperin, GVIN1, IFI44, and ISG56-mediated immune responses quickly decreased. In other words, IFN responses induced by RBIV infection were limited and were not able to active antiviral responses at later stages (10 dpi) in the high-mortality condition. These observations are similar to previous proteomic analyses of red blood cells from RBIV-infected rock bream, which revealed an ISG15 antiviral mechanism-related pathway, including the down-regulation of filamin B (FLNB), IFN regulatory factor 3 (IRF3), nucleoporin 35 (NUP35), tripartite motif-containing 25 (TRIM25), and karyopherin subunit alpha 3 (KPNA3) after RBIV infection at 26 °C (100% mortality condition) [41]. This phenomenon suggests that the upstream regions of most major IFN-related antiviral immune responses are not sufficient to activate downstream molecules and are suppressed in the high susceptibility condition, which ultimately leads to mortality of the infected rock bream. Also our previous study showed that the IFN-mediated antiviral responses was not sufficient for induction in rock bream following administration of immunostimulants: i) Mx, ISG15, and PKR expression levels were not significantly induced in the head kidney following CpG ODN 1668 administration [27]; and ii) rock bream individuals treated with poly (I:C) displayed high expression levels of IRF3, ISG15, and PKR genes in blood samples, whereas no significant up-regulation was observed in the spleen or head kidney [28]. Furthermore, in the aquaculture field condition, RBIV outbreaks typically occurred in rock bream when water temperature was between 23 and 27 °C [2]. In rock bream, while the CpG ODN 1668 and poly (I:C) did not induce antiviral immune responses with higher virus replication speed (i.e. water temperature of 26 °C and infection dose of 1.1 × 107), CpG ODN 1668 and poly (I:C) significantly induced high survival rates when the RBIV replication speed was reduced (i.e. water temperature of 20 °C and infection dose of 1.1 × 104) [5,6]. These findings suggest that IFN induction does not result in a strong antiviral response when rock bream are exposed to the high susceptibility condition.
Conversely, our results reveal that IFN and ISGs-related genes play an important role in the antiviral immune response, as well as survival of the rock bream in the low susceptibility condition (group B). This group of rock bream infected with RBIV and maintained for 7 days at 23 °C before the water temperature was reduced to 17 °C had mortality rates of 28%. In the virus replication patterns of group B, over 105/μL of RBIV MCP copies were detected in the spleen on 7 dpi and reached approximately 107/μL of MCP copies at 20 d and 22 dpi, then decreased after 25 dpi. The specific mortality was observed after 24 d, 25 d, 26 d, 27 d, and 35 dpi (one fish each day) with an estimated mortality of 4, 8, 13, 18, and 28%, respectively. Individual differences were high between 25 d and 30 dpi, 12 fish could drastically eliminate the virus (range 102–104/μL) and three fish exhibiting high RBIV copy number (range 105–107/μL). Thus, this time period (25 d to 30 dpi) was regarded as the recovery stage from infection. Hence, immune responses at approximately 20 d to 30 dpi may be important in evaluating factor(s) for fish survival or death. At 7 dpi, Mx gene expression was only significantly activated and then continuously elevated until 25 dpi. Between 10 d and 22 dpi, the remainder of the IFN and ISGs-related genes were significantly elevated in the head kidney of rock bream at several experimental points. Interestingly, at 20 d and 22 dpi, when virus replication is peaked at approximately 107/μL, most of the IFN-related gene (IRF1, IRF3, IRF8, STAT1, Mx, ISG15, PKR, Viperin, GVIN1, IFI44, and ISG56) were significantly expressed. However, those immune gene responses (except Mx) was not high when virus replication reached its peak (around 107/μL) in the group A. Moreover, while IFI30 was significantly expressed in the group B (low-mortality condition), IFI30 was not significantly induced in all of infection stages in group A (high-mortality condition). Furthermore, persistent and effective antiviral immune responses in the RBIV-infected rock bream were evident in the initial stages of infection and also at the later stage in the group B. Prolonged and elevated expression of IRF3 (at least 30 dpi), IRF8 (at least 30 dpi), ISG15 (at least 30 dpi), PKR (at least 28 dpi), Viperin (at least 30 dpi), and IFI44 (at least 30 dpi) was observed. The persistent expressions of these genes could be one of the reasons for the effective control of viral replication at the recovery stage in the group B. Therefore, we can conclude that the immune responses dependent on IRF3, IRF8, ISG15, PKR, Viperin, IFI44, and IFI30 are efficiently induced in rock bream, leading to survival of RBIV-infected fish.
The details of the interaction and activation of those genes in rock bream after RBIV infection remain to be elucidated. Similar to these results, other experimental studies on fish viral diseases have demonstrated the effect of water temperature-dependency on the antiviral immune responses of olive flounder (Paralichthys olivaceus) against hirame rhabdovirus (HIRRV) and VHSV [23,42], and observed more efficient IFN, Mx, and ISG15-related antiviral immune responses at 20 °C (non-susceptible) than at 10 °C or 15 °C (highly susceptible). This result in the ability of the fish to recover from virus infection.
On the other hand, in group C, rock bream did not show any mortality at the no-susceptibility water temperature of 17 °C. The RBIV copy number exceeded 104/μL in the spleen on 20 dpi, which then decreased to a minimum at 40 dpi (5.3 × 101/μL). These findings suggest that RBIV could replicate slowly in fish even under no-mortality conditions. Furthermore, in the no-mortality group, IRF3 (7 and 10 dpi), Mx (10 dpi), ISG15 (4, 7, 10, 15, and 20 dpi), PKR (2, 4, and 7 dpi), and Viperin (10 dpi) were significantly upregulated. While not significantly upregulated under RBIV replication, the expression levels of antiviral immune related genes (IRF1, IRF8, STAT1, GVIN1, IFI44, and ISG56) were higher than the control at certain times. Similarly, it has been reported that the HIRRV-infected olive flounder at 20°C (non-susceptible condition) induced efficient IFN, Mx, and ISG15-related immune responses and significantly reduced viral loads, and prevented HIRRV infection [23].
5. Conclusions
To conclude, in the present study, virus replication, mortality rates, and IFN-related gene expression levels were analyzed in high susceptibility (26 °C), low susceptibility (starting at 23 °C and then reduced to 17 °C), or no-susceptibility (17 °C) conditions of virus infection. In the low and no-susceptibility conditions, RBIV infection induced an advanced antiviral immune response, which remained elevated until the later infection stages. This response may help the complete elimination of the virus, allowing the fish to recover from viral infection.
Author Contributions
M-HJ performed experiments, analyzed data and wrote the manuscript.
Funding
This research was supported by a grant from Hanseo university (2023).
Institutional Review Board Statement
All rock bream experiments were carried out in strict accordance with the recommendations of Hanseo University.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data that support the findings of this study are available from the author, Myung-Hwa Jung, upon reasonable request.
Acknowledgments
This research was supported by a grant from Hanseo university (2023).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kurita, J.; Nakajima, K. Megalocytiviruses. Viruses. 2012, 4, 521–538. [Google Scholar] [CrossRef]
- Jung, S.J.; Oh, M.J. Iridovirus-like infection associated with high mortalities of striped beakperch, Oplegnathus fasciatus (Temminck et Schlegel), in southern coastal areas of the Korean peninsula. J Fish Dis. 2000, 23, 223–226. [Google Scholar] [CrossRef]
- Jung, M.H.; Jung, S.J.; Vinay, T.N.; Nikapitiya, C.; Kim, J.O.; Lee, J.H.; Lee, J.H.; Oh, M.J. Effects of water temperature on mortality in Megalocytivirus-infected rock bream Oplegnathus fasciatus (Temminck et Schlegel) and development of protective immunity. J Fish Dis. 2015, 38, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.H.; Lee, J.H.; Jung, S.J. Low pathogenicity of FLIV (flounder iridovirus) and the absence of cross-protection between FLIV and RBIV (rock bream iridovirus). J Fish Dis. 2016, 39, 1325–1333. [Google Scholar] [CrossRef]
- Jung, M.H.; Lee, J.H.; Ortega-Villaizan, M.; Perez, L.; Jung, S.J. Protective immunity against Megalocytivirus infection in rock bream (Oplegnathus fasciatus) following CpG ODN administration. Vaccine. 2017, 35, 3691–3699. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.H.; Jung, S.J. Protective immunity against rock bream iridovirus (RBIV) infection and TLR3-mediated type I interferon signaling pathway in rock bream (Oplegnathus fasciatus) following poly (I:C) administration. Fish Shellfish Immunol. 2017, 67, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.H.; Nikapitiya, C.; Vinay, T.N.; Lee, J.; Jung, S.J. Rock bream iridovirus (RBIV) replication in rock bream (Oplegnathus fasciatus) exposed for different time periods to susceptible water temperatures. Fish Shellfish Immunol. 2017, 70, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Gui, J.F. Molecular regulation of interferon antiviral response in fish. Dev Comp Immunol. 2012, 38, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Bergan, V.; Kileng, Ø.; Sun, B. Robertsen B. Regulation and function of interferon regulatory factors of Atlantic salmon. Mol Immunol. 2010, 47, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Mamane, Y.; Heylbroeck, C.; Génin, P.; Algarté, M.; Servant, M.J.; LePage, C.; DeLuca, C.; Kwon, H.; Lin, R. , Hiscott, J. Interferon regulatory factors: the next generation. Gene. 1999, 237, 1–14. [Google Scholar] [CrossRef]
- Yasuike, M.; Kondo, H.; Hirono, I.; Aoki, T. Identification and characterization of Japanese flounder, Paralichthys olivaceus interferon-stimulated gene 15 (Jf-ISG15). Comp Immunol Microb. 2011, 34, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Hirono, I.; Aoki, T. Cloning and analysis of expression of Mx cDNA in Japanese flounder, Paralichthys olivaceus. Dev Comp Immunol. 2000, 24, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.E. Antiviral actions of interferons. Clin Microbiol Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef] [PubMed]
- Hallen, L.C.; Burki, Y.; Ebeling, M.; Broger, C.; Siegrist, F.; Oroszlan-Szovik, K.; Bohrmann, U.; Certa, U.; Foser, S. Antiproliferative activity of the human IFN-α-inducible protein IFI44. J Interferon Cytokine Res. 2007, 27, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, B.; Phan, U.T.; Geuze, H.J.; Cresswell, P. Enzymatic reduction of disulfide bonds in lysosomes: characterization of a gamma-interferon-inducible lysosomal thiol reductase (GILT). Proc Natl Acad Sci. 2000, 97, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Landolfo, S.; Gribaudo, G.; Angeretti, A.; Gariglio, M. Mechanisms of viral inhibition by interferons. Pharmacol Therapeut. 1995, 65, 415–442. [Google Scholar] [CrossRef]
- Chen, Y.M.; Su, Y.L.; Lin, J.H.Y.; Yang, H.L.; Chen, T.Y. Cloning of an orange-spotted grouper (Epinephelus coioides) Mx cDNA and characterisation of its expression in response to nodavirus. Fish Shellfish Immunol. 2006, 20, 58–71. [Google Scholar] [CrossRef]
- Larsen, R.; Røkenes, T.P.; Robertsen, B. Inhibition of infectious pancreatic necrosis virus replication by Atlantic salmon Mx1 protein. J Virol. 2004, 78, 7938–7944. [Google Scholar] [CrossRef]
- Lin, C.H.; John, J.A.C.; Lin, C.H.; Chang, C.Y. Inhibition of nervous necrosis virus propagation by fish Mx proteins. Biochem Bioph Res Co. 2006, 351, 534–539. [Google Scholar] [CrossRef]
- Caipang, C.M.A.; Hirono, I.; Aoki, T. In vitro inhibition of fish rhabdoviruses by Japanese flounder, Paralichthys olivaceus Mx. Virology. 2003, 317, 373–382. [Google Scholar] [CrossRef]
- Avunje, S.; Kim, W.S.; Park, C.S.; Oh, M.J.; Jung, S.J. Toll-like receptors and interferon associated immune factors in viral haemorrhagic septicaemia virus-infected olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2011, 31, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Thanasaksiri, K.; Sakai, N.; Yamashita, H.; Hirono, I.; Kondo, H. Influence of temperature on Mx gene expression profiles and the protection of sevenband grouper, Epinephelus septemfasciatus, against red-spotted grouper nervous necrosis virus (RGNNV) infection after poly (I:C) injection. Fish Shellfish Immunol. 2014, 40, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tang, X.; Sheng, X.; Xing, J.; Zhan, W. The influence of temperature on viral replication and antiviral-related genes response in hirame rhabdovirus-infected flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2017, 68, 260–265. [Google Scholar] [CrossRef]
- Jung, M.H.; Jung, S.J. Gene expression regulation of the TLR9 and MyD88-dependent pathway in rock bream against rock bream iridovirus (RBIV) infection. Fish Shellfish Immunol. 2017, 70, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.H.; Nikapitiya, C.; Song, J.Y.; Lee, J.H.; Lee, J.H.; Oh, M.J.; Jung, S.J. Gene expression of pro- and anti-apoptotic proteins in rock bream (Oplegnathus fasciatus) infected with Megalocytivirus (family Iridoviridae). Fish Shellfish Immunol. 2014, 37, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Nikapitiya, C.; Jung, S.J.; Jung, M.H.; Song, J.Y.; Lee, J.H.; Lee, J.H.; Oh, M.J. Identification and Molecular Characterization of Z/ZE Lineage MHC Class I Heavy Chain Homologue and β2-microglobulin from Rock Bream Oplegnathus fasciatus. Fish Pathol. 2014, 49, 93–112. [Google Scholar] [CrossRef]
- Jung, M.H.; Jung, S.J. CpG ODN 1668 induce innate and adaptive immune responses in rock bream (Oplegnathus fasciatus) against rock bream iridovirus (RBIV) infection. Fish Shellfish Immunol. 2017, 69, 247–257. [Google Scholar] [CrossRef]
- Jung, M.H.; Jung, S.J. Innate immune responses against rock bream iridovirus (RBIV) infection in rock bream (Oplegnathus fasciatus) following poly (I:C) administration. Fish Shellfish Immunol. 2017, 71, 171–176. [Google Scholar] [CrossRef]
- Jung, M.H.; Nikapitiya, C.; Jung, S.J. DNA vaccine encoding myristoylated membrane protein (MMP) of rock bream iridovirus (RBIV) induces protective immunity in rock bream (Oplegnathus fasciatus). Vaccine. 2018, 6, 802–810. [Google Scholar] [CrossRef]
- Zenke, K.; Nam, Y.K.; Kim, K.H. Molecular cloning and expression analysis of double-stranded RNA-dependent protein kinase (PKR) in rock bream (Oplegnathus fasciatus). Veterinary Immunol Immunopathol. 2010, 133, 290–295. [Google Scholar] [CrossRef]
- Wan, Q.; Wicramaarachchi, W.N.; Whang, I.; Lim, B.S.; Oh, M.J.; Jung, S.J.; Kim, H.C.; Yeo, S.Y. , Lee, J.H. Molecular cloning and functional characterization of two duplicated two-cysteine containing type I interferon genes in rock bream Oplegnathus fasciatus. Fish Shellfish Immunol. 2012, 33, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Bathige, S.D.N.K.; Whang, I.; Umasuthan, N.; Lim, B.S.; Park, M.A.; Kim, E.; Park, H.C.; Lee, J.H. Interferon regulatory factors 4 and 8 in rock bream, Oplegnathus fasciatus: structural and expressional evidence for their antimicrobial role in teleosts. Fish Shellfish Immunol. 2012, 33, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Wickramaarachchi, W.N.; Wan, Q.; Lim, B,S. ; Jung, H.B.; De Zoysa, M.; Park, M.A.; Lee, J.H.; Hwang, I.S. Genomic characterization of interferon regulatory factor 5 from rock bream (Oplegnathus fasciatus) and its role in antiviral defense. Fish Shellfish Immunol. 2014, 37, 256–267. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jung, M.H.; Jung, S.J. Correlation of virus replication and spleen index in rock bream iridovirus infected rock bream Oplegnathus fasciatus. J Fish Pathol. 2019, 32, 1–8. [Google Scholar] [CrossRef]
- Kim, H.J.; Oseko, N.; Nishizawa, T.; Yoshimizu, M. Protection of rainbow trout from infectious hematopoietic necrosis (IHN) by injection of infectious pancreatic necrosis virus (IPNV) or poly (I:C). Dis Aquat Organ. 2009, 83, 105–113. [Google Scholar] [CrossRef]
- Oh, M.J.; Takami, I.; Nishizawa, T.; Kim, W.S.; Kim, C.S.; Kim, S.R.; Park, M.A. Field tests of Poly (I:C) immunization with nervous necrosis virus (NNV) in sevenband grouper, Epinephelus septemfasciatus (Thunberg). J Fish Dis. 2012, 35, 187–191. [Google Scholar] [CrossRef]
- Avunje. S.; Jung, S.J. Poly (I:C) and imiquimod induced immune responses and their effects on the survival of olive flounder (Paralichthys olivaceus) from viral haemorrhagic septicaemia. Fish Shellfish Immunol. 2017, 71, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y,J,; Kim, K,H. Effect of CpG-ODNs belonging to different classes on resistance of olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) and Miamiensis avidus (Ciliata; Scuticociliatia) infections. Aquaculture. 2012, 324, 39–43. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Y.; Cai, J.; Wei, S.; Ouyang, Z.; Qin, Q. Molecular cloning, expression and functional analysis of ISG15 in orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2013, 34, 1094–102. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.H.; Chico, V.; Ciordia, S.; Mena, M.C.; Jung, S.J.; Ortega-Villaizan, M.D.M. The Megalocytivirus RBIV Induces Apoptosis and MHC Class I Presentation in Rock Bream (Oplegnathus fasciatus) Red Blood Cells. Front Immunol. 2019, 10, 160. [Google Scholar] [CrossRef]
- Avunje, S.; Kim, W.S.; Oh, M.J.; Choi, I.; Jung, S.J. Temperature-dependent viral replication and antiviral apoptotic response in viral haemorrhagic septicaemia virus (VHSV)-infected olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2012, 32, 1162–1170. [Google Scholar] [CrossRef]
Figure 1.
Mortality patterns of fish intraperitoneally injected with RBIV (1.1 × 107 MCP gene copies, 100 μL/fish) at different water temperatures. i) Group A: high susceptibility condition and virus infection at 26 °C; ii) Group B: low susceptibility condition and virus infection at 23 °C, with reduction to 17 °C at 7 dpi. The mortality (%) of group B was calculated as: (number of dead fish/fish population parameter) × 100; iii) Group C: no-susceptibility condition and virus infection at 17 °C. The cumulative mortalities of groups A, B, and C published previously [24,25] are shown for reference.
Figure 1.
Mortality patterns of fish intraperitoneally injected with RBIV (1.1 × 107 MCP gene copies, 100 μL/fish) at different water temperatures. i) Group A: high susceptibility condition and virus infection at 26 °C; ii) Group B: low susceptibility condition and virus infection at 23 °C, with reduction to 17 °C at 7 dpi. The mortality (%) of group B was calculated as: (number of dead fish/fish population parameter) × 100; iii) Group C: no-susceptibility condition and virus infection at 17 °C. The cumulative mortalities of groups A, B, and C published previously [24,25] are shown for reference.
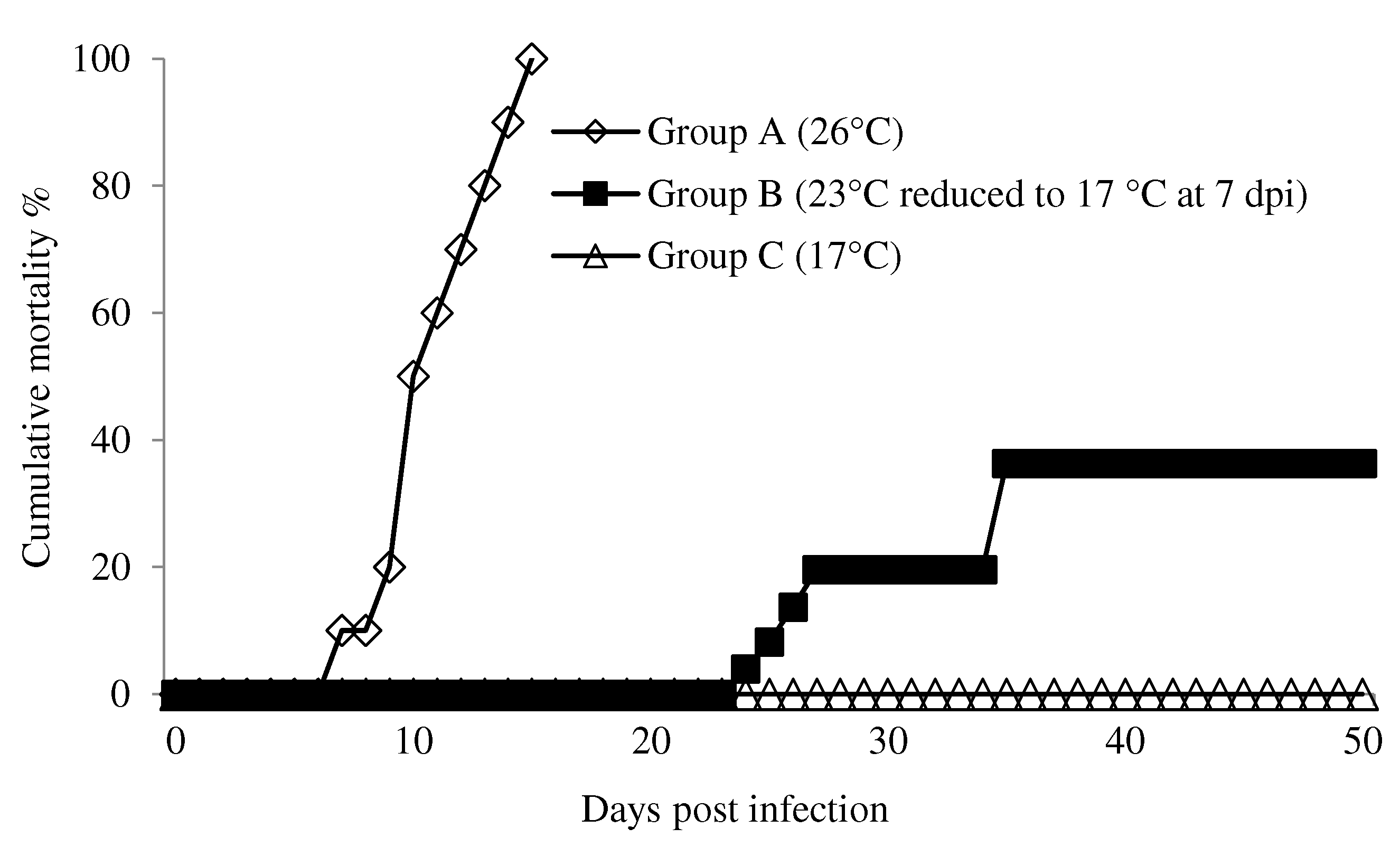
Figure 2.
Changes of absolute virus copy number of rock bream according to time after injection of RBIV (1.1 × 107 MCP gene copies per fish). In group A, fish maintained at 26 °C. In group B, fish were injected at 23 °C, and the water temperature was reduced to 17 °C at 7 d post-infection. In groups C, fish were maintained at 17 °C. One-way analysis of variance (ANOVA) was performed between conditions, with Tukey’s multiple comparison test. Different superscript letters denote significant differences (p < 0.05). Data are represented as individual values. Line represents mean value. Results of virus copy numbers are taken from Jung et al. [24,25].
Figure 2.
Changes of absolute virus copy number of rock bream according to time after injection of RBIV (1.1 × 107 MCP gene copies per fish). In group A, fish maintained at 26 °C. In group B, fish were injected at 23 °C, and the water temperature was reduced to 17 °C at 7 d post-infection. In groups C, fish were maintained at 17 °C. One-way analysis of variance (ANOVA) was performed between conditions, with Tukey’s multiple comparison test. Different superscript letters denote significant differences (p < 0.05). Data are represented as individual values. Line represents mean value. Results of virus copy numbers are taken from Jung et al. [24,25].
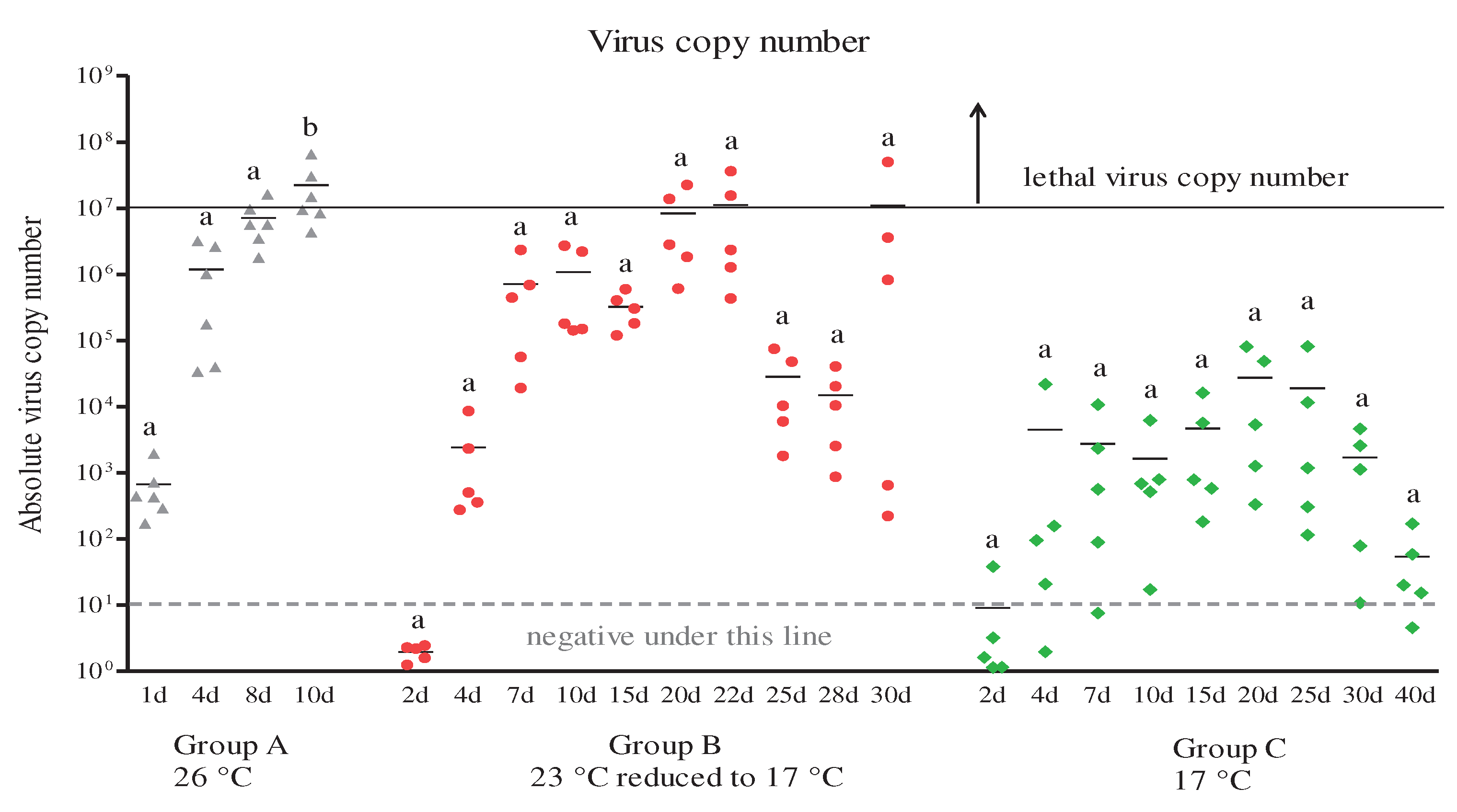
Figure 3.
Expression levels of interferon-related genes in head kidneys of RBIV-infected fish from group A (high susceptibility condition; 26 °C). Gene expression analysis, relative to control individuals (blue line) were evaluated by RT-qPCR. Control fish injected with PBS were maintained and relative expression was determined by normalizing with the expression of the β-actin internal control. Each experiment was performed in duplicate. Bars represent the standard error (SE) of the mean for six individuals, *p < 0.05 and **p < 0.01. A-IRF1; B-IRF3; C-IRF4; D-IRF6; E-STAT1; F-Mx; G-ISG15; H-PKR; I-Viperin; J-GVIN1; K-IFI44; L-ISG56, and IFI30. The virus copy number at each sampling point is plotted for reference. Results of virus copy numbers are from Jung et al. [24,25].
Figure 3.
Expression levels of interferon-related genes in head kidneys of RBIV-infected fish from group A (high susceptibility condition; 26 °C). Gene expression analysis, relative to control individuals (blue line) were evaluated by RT-qPCR. Control fish injected with PBS were maintained and relative expression was determined by normalizing with the expression of the β-actin internal control. Each experiment was performed in duplicate. Bars represent the standard error (SE) of the mean for six individuals, *p < 0.05 and **p < 0.01. A-IRF1; B-IRF3; C-IRF4; D-IRF6; E-STAT1; F-Mx; G-ISG15; H-PKR; I-Viperin; J-GVIN1; K-IFI44; L-ISG56, and IFI30. The virus copy number at each sampling point is plotted for reference. Results of virus copy numbers are from Jung et al. [24,25].
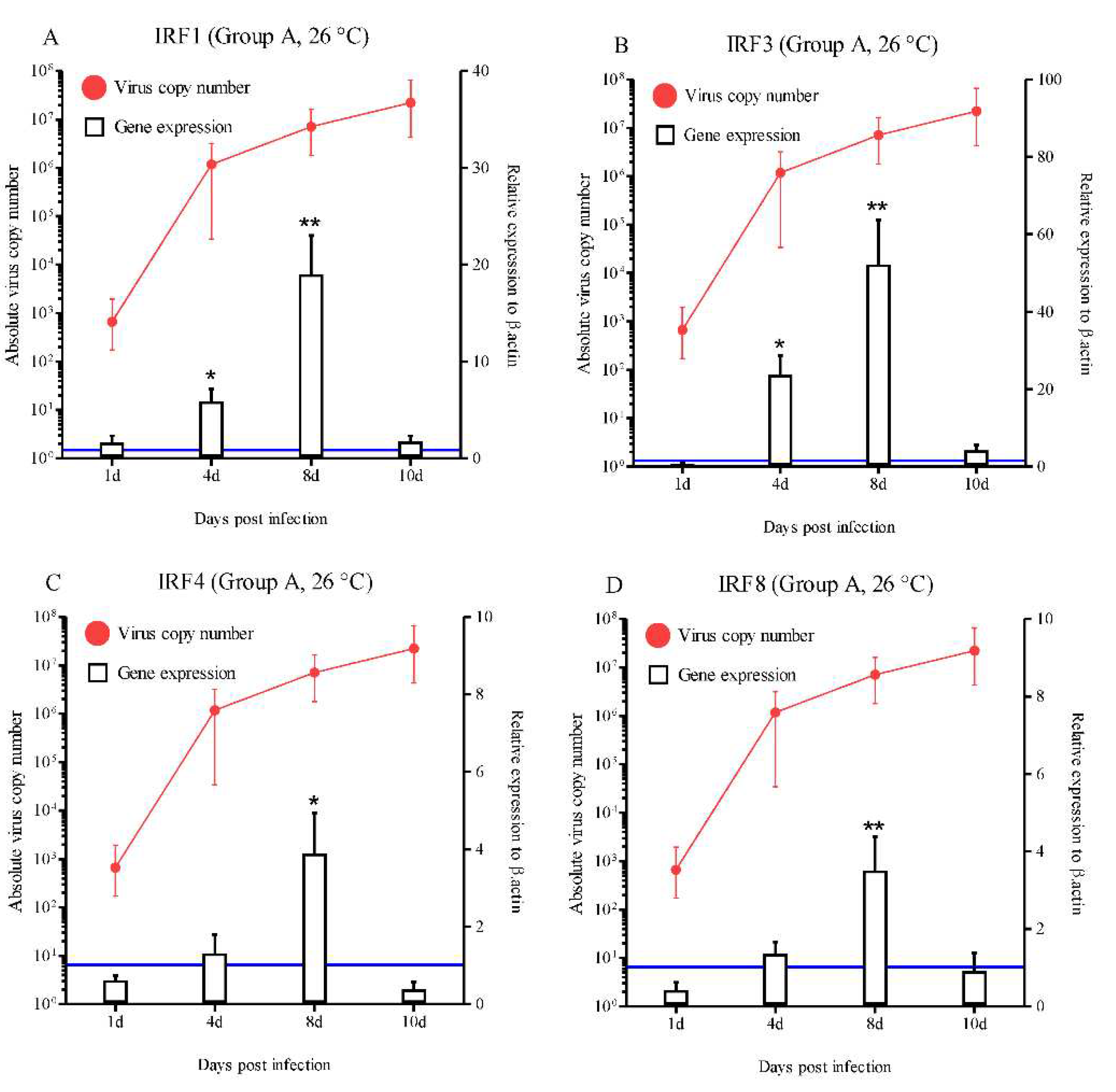
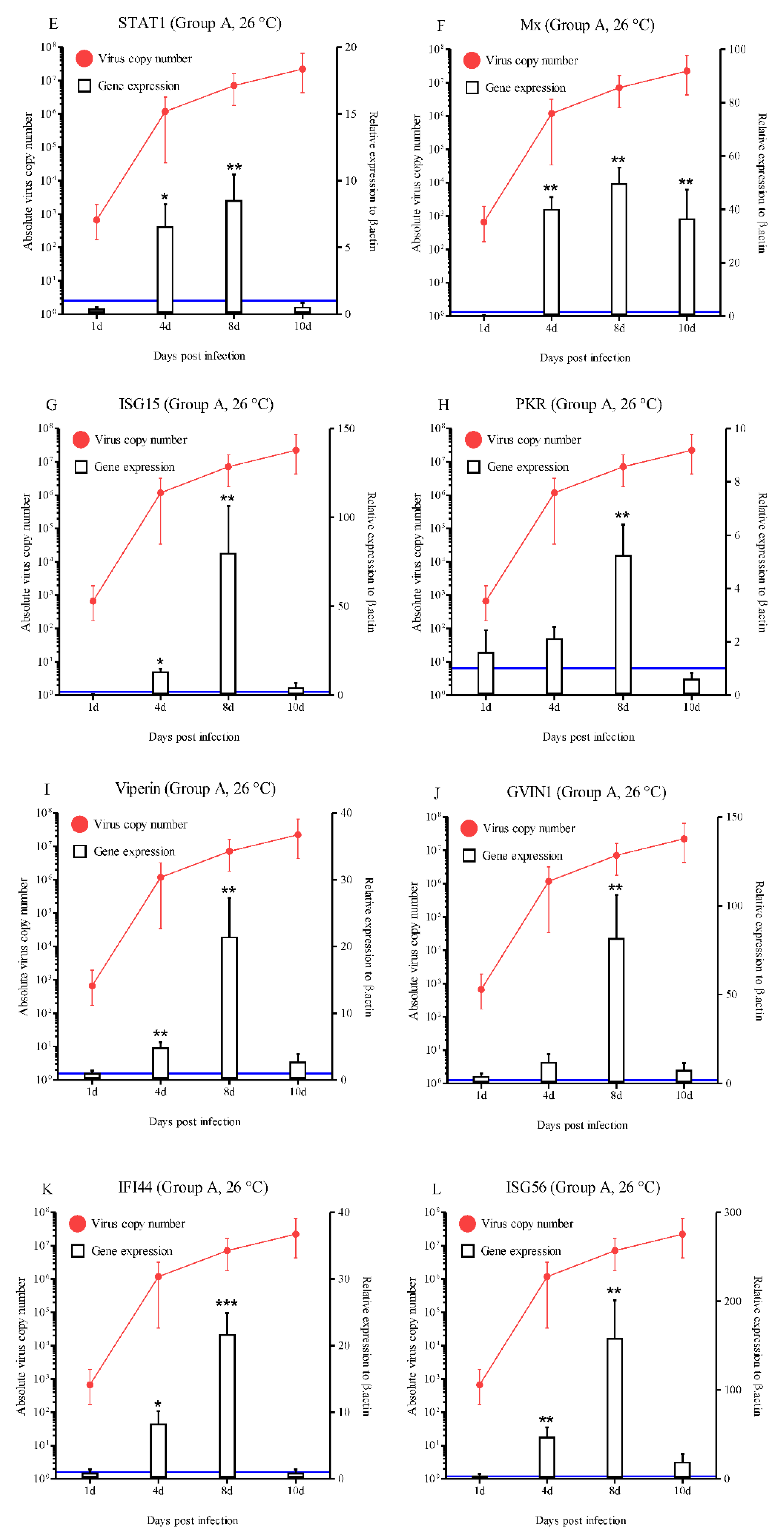
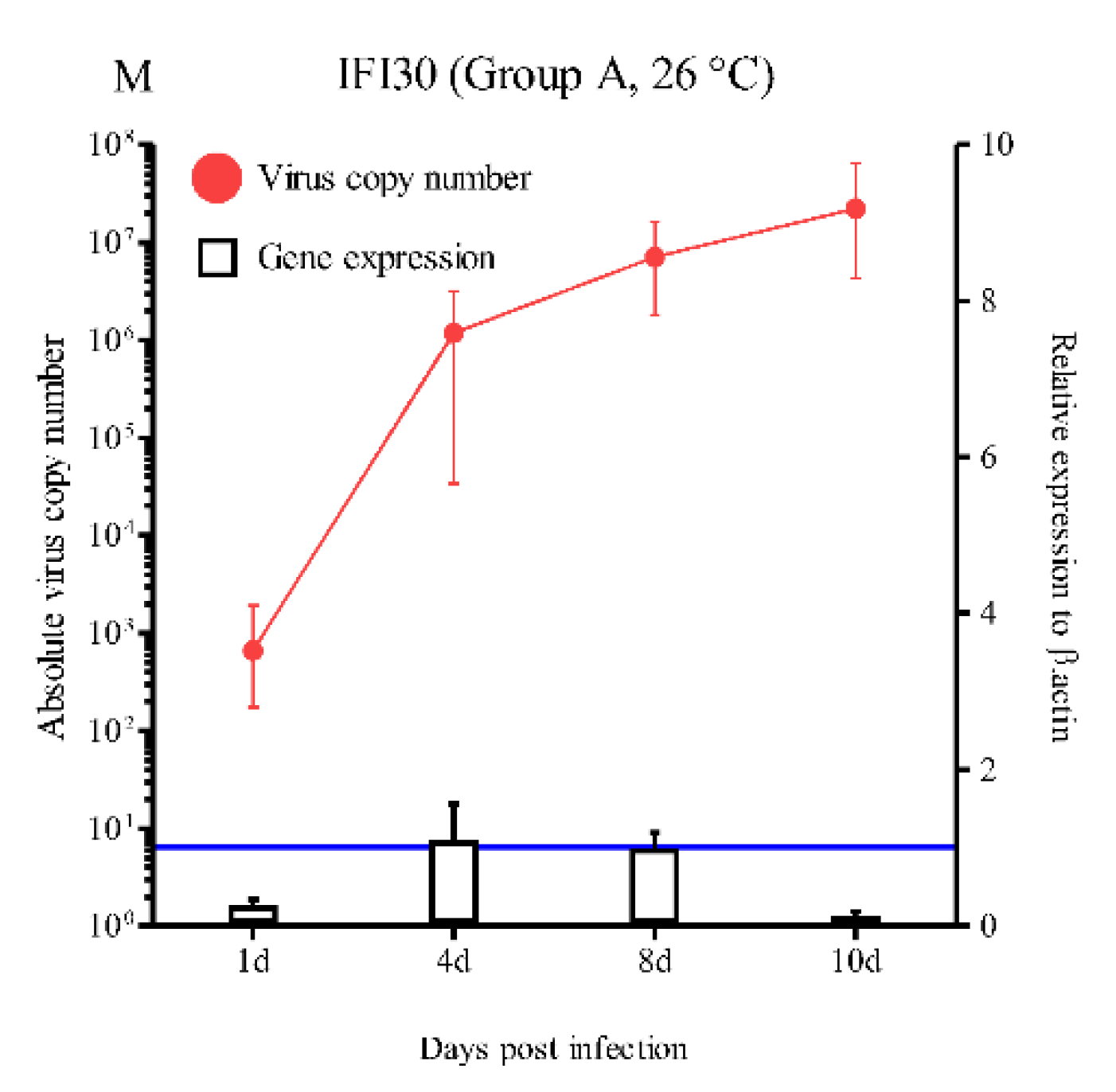
Figure 4.
Expressions levels of interferon-related genes in head kidneys of RBIV-infected fish from group B (low susceptibility condition; 23 °C reduced to 17 °C at 7 dpi). Gene expression analysis, relative to control individuals (blue line), evaluated by means of RT-qPCR. Control groups with PBS injection were maintained and relative expression was determined by normalizing with the expression of the β-actin internal control. Each experiment was performed in duplicate. Bars represent the standard error (SE) of the mean for five individuals, *p < 0.05, **p < 0.01, and ***p < 0.001. A-IRF1; B-IRF3; C-IRF4; D-IRF6; E-STAT1; F-Mx; G-ISG15; H-PKR; I-Viperin; J-GVIN1; K-IFI44; L-ISG56, and IFI30. The virus copy number at each sampling point is plotted for reference. Results of virus copy numbers are taken from Jung et al. [24,25].
Figure 4.
Expressions levels of interferon-related genes in head kidneys of RBIV-infected fish from group B (low susceptibility condition; 23 °C reduced to 17 °C at 7 dpi). Gene expression analysis, relative to control individuals (blue line), evaluated by means of RT-qPCR. Control groups with PBS injection were maintained and relative expression was determined by normalizing with the expression of the β-actin internal control. Each experiment was performed in duplicate. Bars represent the standard error (SE) of the mean for five individuals, *p < 0.05, **p < 0.01, and ***p < 0.001. A-IRF1; B-IRF3; C-IRF4; D-IRF6; E-STAT1; F-Mx; G-ISG15; H-PKR; I-Viperin; J-GVIN1; K-IFI44; L-ISG56, and IFI30. The virus copy number at each sampling point is plotted for reference. Results of virus copy numbers are taken from Jung et al. [24,25].
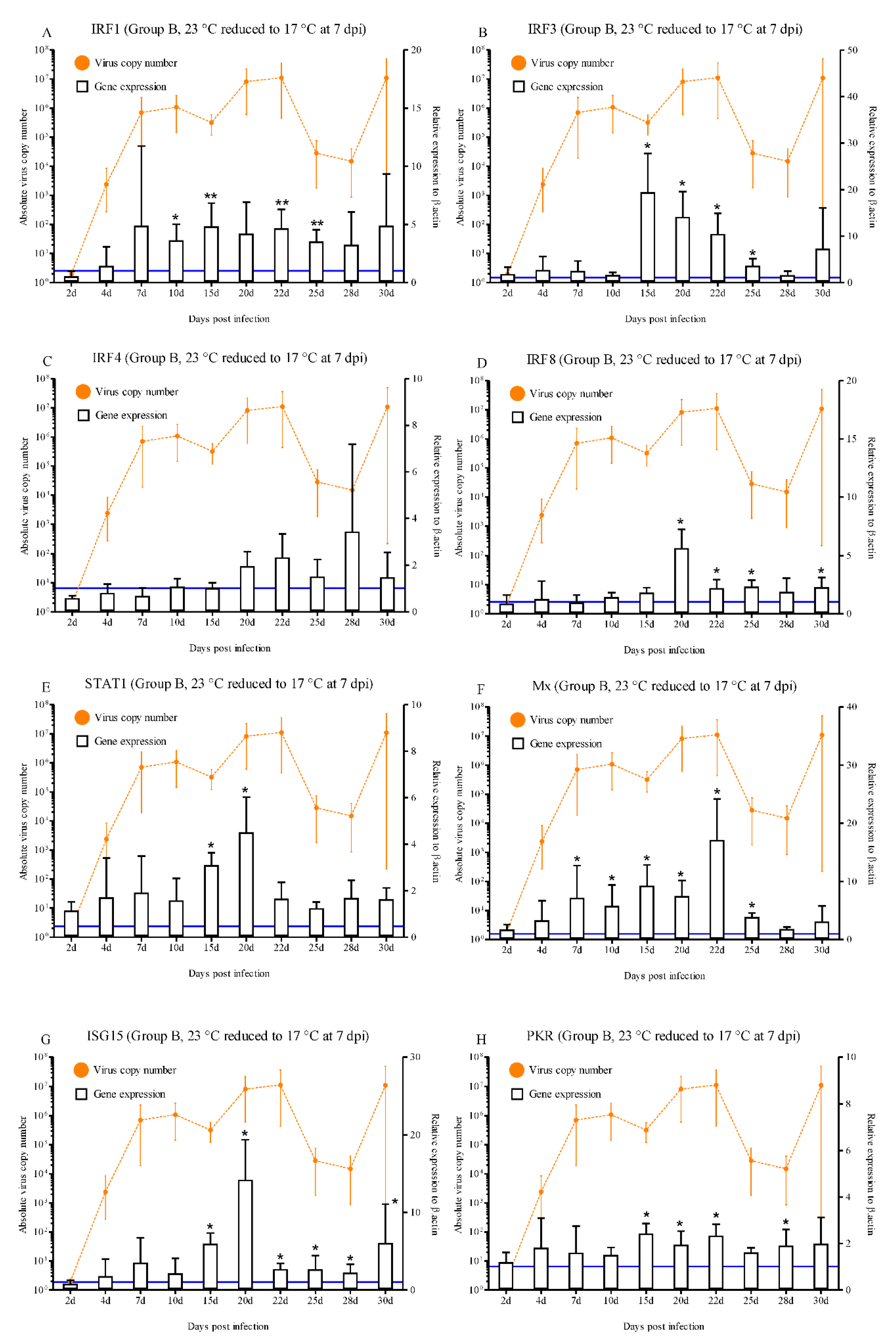
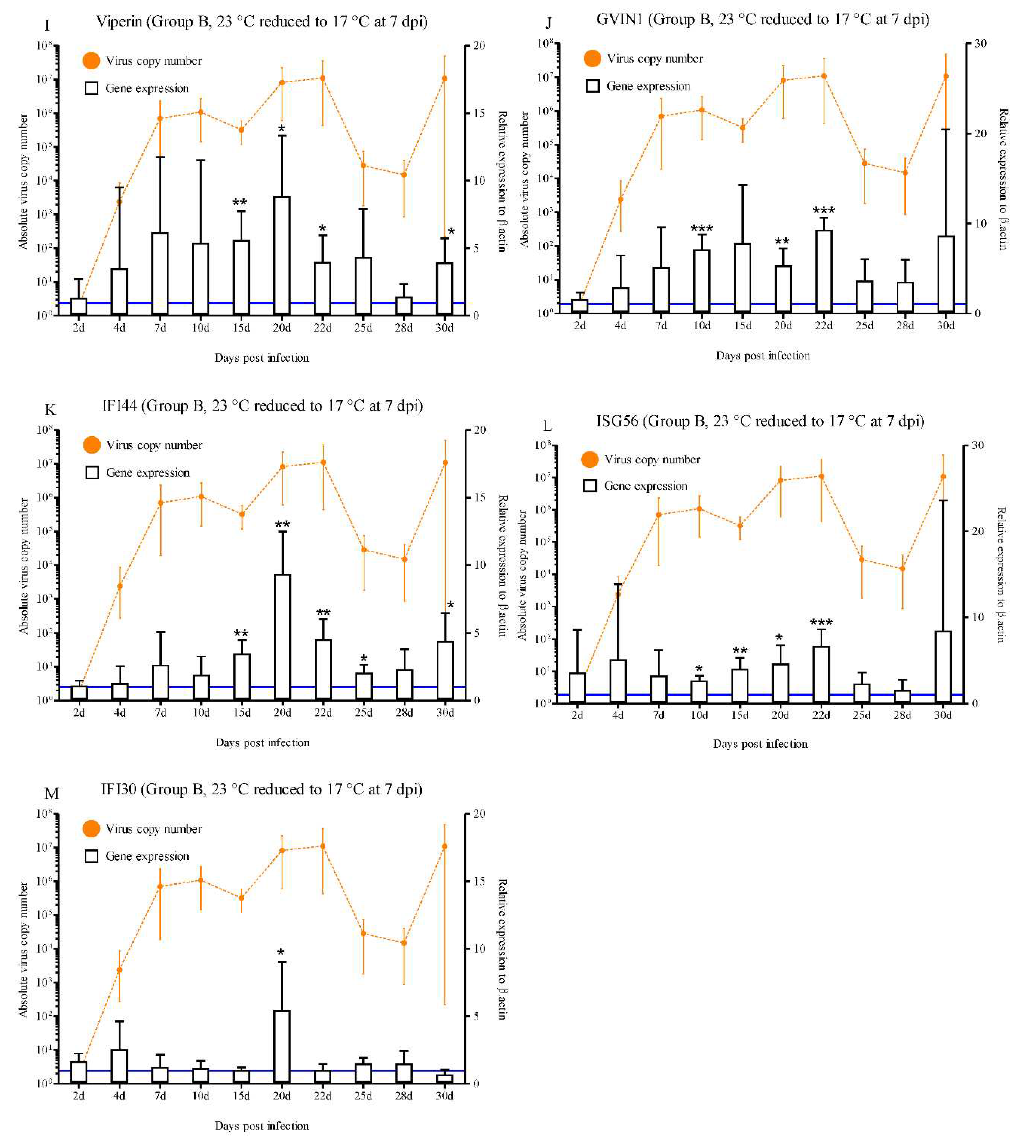
Figure 5.
Expressions levels of interferon-related genes in head kidneys of
RBIV-infected fish from group C (no-susceptibility condition; 17 °C). Gene
expression analysis, relative to control individuals (blue line), evaluated by
means of RT-qPCR. Control groups with PBS injection were maintained and
relative expression was determined by normalizing with the expression of the
β-actin internal control. Each experiment was performed in duplicate. Bars
represent the standard error (SE) of the mean for five individuals, *p
< 0.05. A-IRF1; B-IRF3; C-IRF4; D-IRF6; E-STAT1; F-Mx; G-ISG15; H-PKR;
I-Viperin; J-GVIN1; K-IFI44; L-ISG56, and IFI30. The virus copy number at each
sampling point is plotted for reference. Results of virus copy numbers are
taken from Jung et al. [24,25].
Figure 5.
Expressions levels of interferon-related genes in head kidneys of
RBIV-infected fish from group C (no-susceptibility condition; 17 °C). Gene
expression analysis, relative to control individuals (blue line), evaluated by
means of RT-qPCR. Control groups with PBS injection were maintained and
relative expression was determined by normalizing with the expression of the
β-actin internal control. Each experiment was performed in duplicate. Bars
represent the standard error (SE) of the mean for five individuals, *p
< 0.05. A-IRF1; B-IRF3; C-IRF4; D-IRF6; E-STAT1; F-Mx; G-ISG15; H-PKR;
I-Viperin; J-GVIN1; K-IFI44; L-ISG56, and IFI30. The virus copy number at each
sampling point is plotted for reference. Results of virus copy numbers are
taken from Jung et al. [24,25].
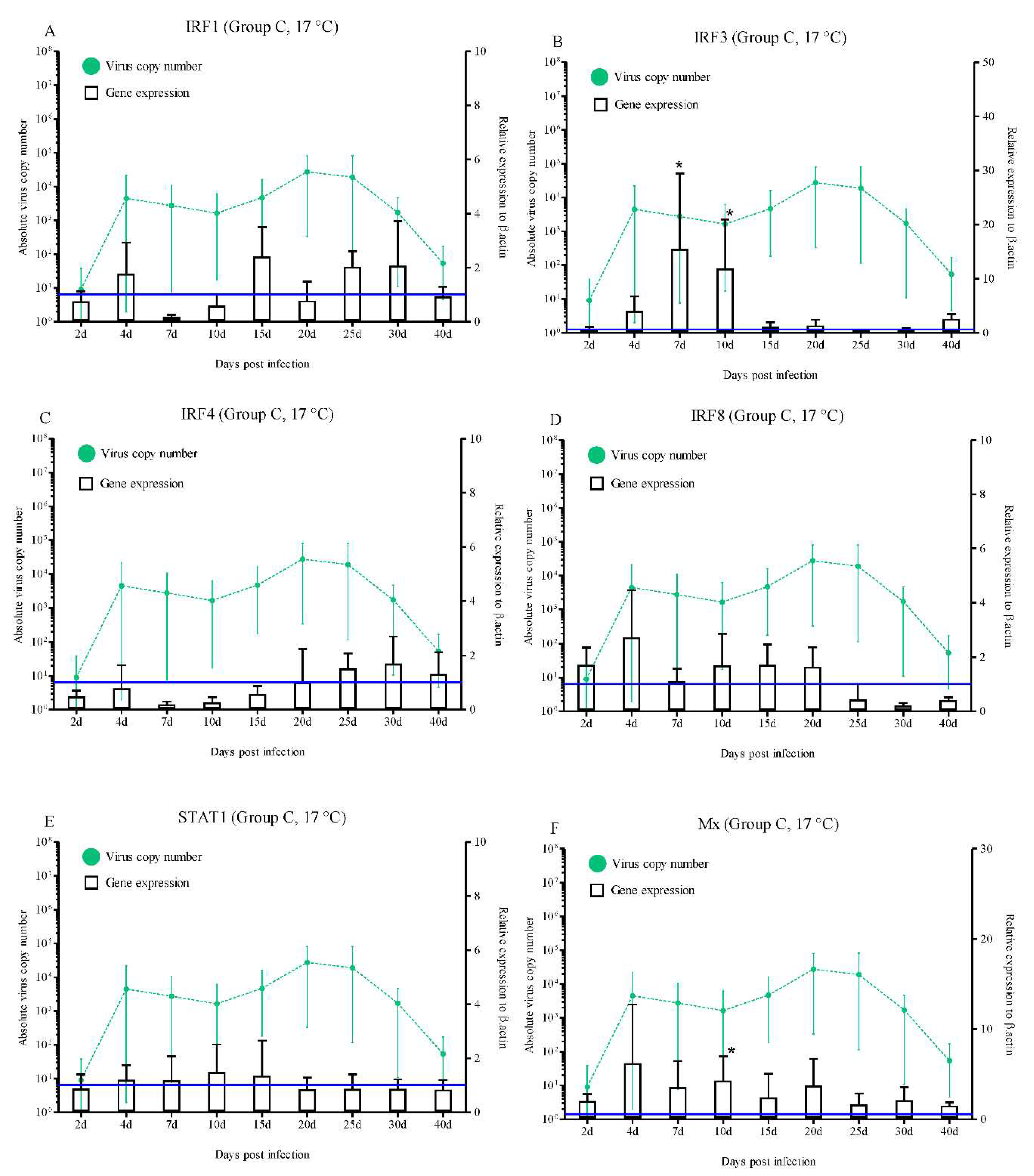
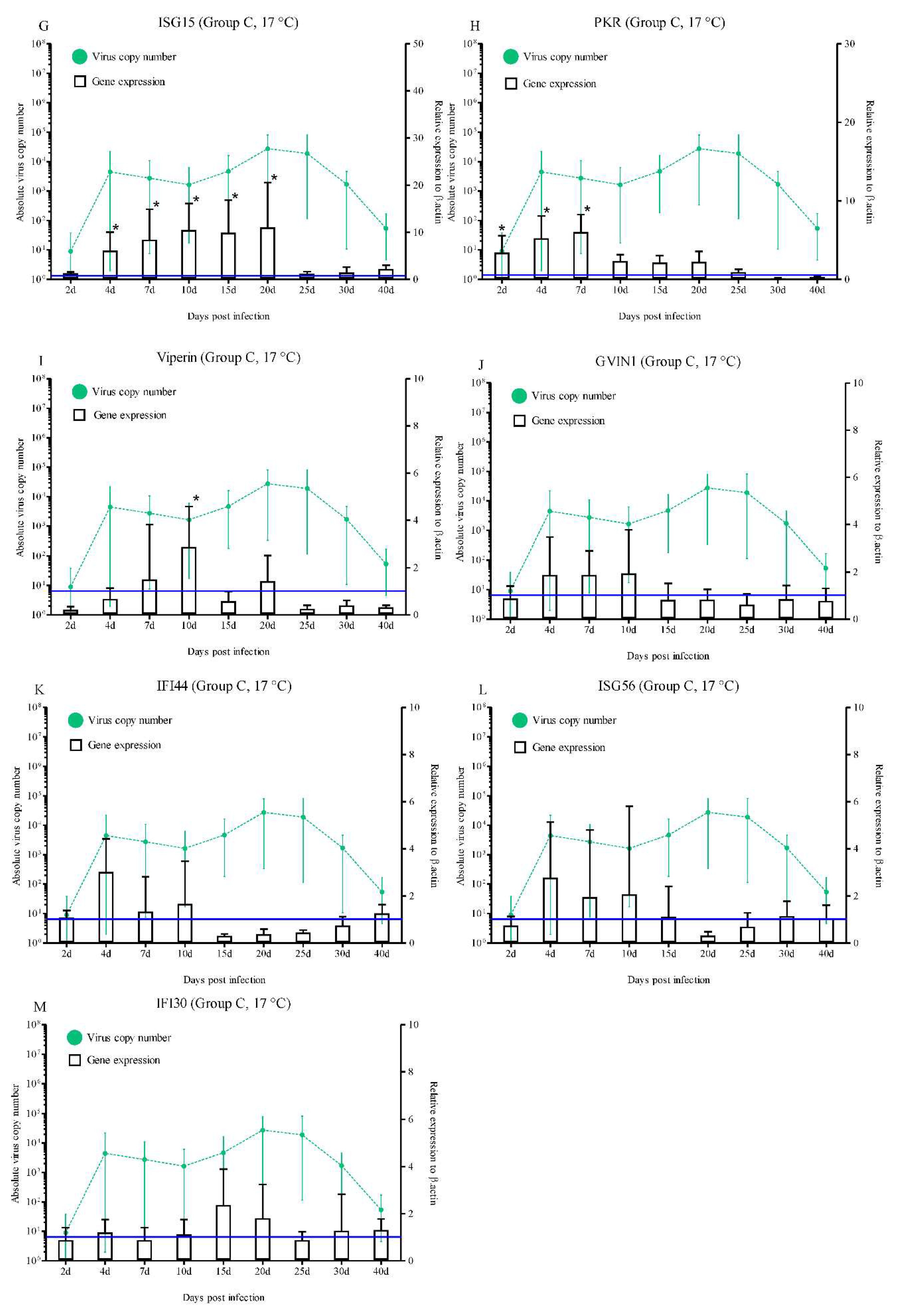

Table 1.
Experimental details of artificial infection.
| Group | Infection dose per fish |
Number of fish | Sampling times (dpi) | Mortality (%) |
| A (26 °C) | 1.1 × 107 | 50 | 1, 4, 8, and 10 | 100 |
| B (23 °C reduced to 17 °C at 7 dpi) | 1.1 × 107 | 60 | 2, 4, 7, 10, 15, 20, 22, 25, 28, and 30 | 28 |
| C (17 °C) | 1.1 × 107 | 50 | 2, 4, 7, 10, 15, 20, 25, 30, and 40 | 0 |
Table 2.
Primers used in this study.
| Name | Sequence (Forward) | Sequence (Reverse) | Accession number |
| β-actin | CAGGGAGAAGATGACCCAGA | CATAGATGGGCACTGTGTGG | FJ975145 |
| MCP | TGCACAATCTAGTTGAGGAGGTG | AGGCGTTCCAAAAGTCAAGG | AY849394 |
| IRF1 | CTGTGACCCAAAGACGTGGA | CTTGGTGGCCTTTGTTGACG | GQ903769.1 |
| IRF3 | GTGTCTAAAGGGACTGAACATCG | CCCTCAAACGTTACTGGATACTG | KF267453.1 |
| IRF4 | ATGGCTTATACGCTAAGCGCCTCT | GTTTGCATGGCTGTTCCTTCTCCA | JQ388475 |
| IRF8 | GGAGCTCTTCAGCGTGTACTAGA | CCTCCTTTATGTCTCCACTGTGA | JQ388478 |
| STAT1 | AGCTCAACATGCTGGCTGACAAAC | AAAGCTTTCTCGTTGGCGCTCTTG | KX714722 |
| Mx | GATCGCCTCTCCTGATGTTC | ACCACCAAGCTGATGGTTTC | FJ155359.1 |
| ISG15 | CTTTAAGACCAAGGTCCAATGC | GCCTCCACATTGTAGTCTGAAAG | BAJ16365.1 |
| PKR | GCTCAACAAATAGTCAGTGGAGT | CTCTGGTGACCAGACCAAAGTCC | FJ179396.1 |
| Viperin | TCAGCAGTACAAAGTGGCGT | TCCCCGTCTATCAGCAGACA | AB665349.1 |
| GVIN1 | TGCAGCTGGTCAAAGTGTCA | CGCTACCTGCCAACTCAAGA | MK674614 |
| IFI44 | GCAGTTCGAGCTCAGAAGGT | CGAGCCTGAGTCACATCCTC | MK674612 |
| ISG56 | GTGAGCGCCTTTAAGCACAG | AGGTACAGAGCAGCGAGGTA | MK674613 |
| IFI30 | GCTGCAGAGGGTTTCTCACT | ATTCCTGTTCCCCATGCTGG | GQ903766.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



