Submitted:
09 October 2023
Posted:
11 October 2023
You are already at the latest version
Abstract
Chrysin, a flavonoid compound, has attracted interest as a therapeutic agent due to its anti-inflammatory, antidiabetic, antidepressant, and particularly its anticancer properties. Although studies have presented findings regarding the anticancer properties of chrysin, research on its molecular mechanisms of action remains largely insufficient. This research aimed to deeply investigate chrysin's effects on the breast cancer cell line, MDA-MB-231, in terms of proliferation, invasion, colony formation, and apoptosis. The XTT test results confirmed chrysin's cytotoxic effect on MDA-MB-231 cells, indicating a 48-hour IC50 value of 115.77 µM. Chrysin induced apoptosis in cells, as evidenced by Annexin V assay, and also reduced their colony-forming and invasion. Gene expression analyses showed elevated levels of APC, AXIN1, AXIN2, GSK3A, GSK3B, CK1α, CTNNB1, as well as apoptosis-related genes CASP3, -7, -9, and BAX. This increase was corroborated by the observed rise in protein levels of caspase 3/7 and GSK3B. Moreover, molecular docking results showed that chrysin interacted with genes in the WNT/β-catenin pathway and exhibited drug-like ADME properties. In conclusion, chrysin exhibits potential anticancer effects against MDA-MB-231 cells. It is hoped that these findings will advance preclinical and clinical studies on chrysin's potential in breast cancer treatment.
Keywords:
antiproliferation
; apoptosis
; breast cancer
; chrysin
; WNT/β-catenin pathway
1. Introduction
Breast cancer is a type of cancer predominantly affecting women and stands as the second-leading cause of cancer-related deaths [1]. In breast cancer treatment, chemotherapeutic drugs are among the most effective instruments, with commonly used drugs including doxorubicin, paclitaxel, docetaxel, and carboplatin [2]. These drugs are often given in combinations, which makes it challenging for cancer cells to develop resistance to the drug. However, the search for therapeutic agents that can be used in treatment continues due to high side effects. In recent years, natural products like flavonoids have become interesting in cancer treatment because they are less toxic and, therefore, cause fewer side effects [3].
Chrysin (5,7-dihydroxyflavone) is recognized as a natural flavonoid compound especially found in several plants, notably in blue passionflower varieties such as Passiflora caerulea and Passiflora incarnata and as well as in the plant Oroxylum indicum. Furthermore, both bee propolis and honey are sources of chrysin [4]. Both ethnobotanically and pharmacologically, plants containing chrysin are known to exhibit anxiolytic [5], anti-inflammatory [6], neuroprotective [7], antidiabetic [8], anti-asthmatic [9], antidepressant [10], and anticancer properties [11]. In addition, when the pharmacokinetic interactions of chrysin with other drugs were examined, it has been reported that it can be used as a neuroprotective, renal protective, and hepatoprotective agent [12]. In both laboratory and animal studies, chrysin has demonstrated its ability to inhibit tumor growth by inducing apoptosis, regulating the cell cycle, and limiting angiogenesis, invasion, and metastasis [13,14,15,16]. It also interferes with various cellular responses including intracellular reactive oxygen species (ROS) and cell differentiation. On a molecular level, it has been reported that it reduces the expression of pro-inflammatory cytokines and other inflammatory mediators by inhibiting nuclear factor-kappa B (NF-κB), signal transducer and activator of transcription 3 (STAT3), nitric oxide/poly (ADP-Ribose) polymerase-1 (NO/PARP-1), extracellular signal-regulated kinase/ nuclear factor erythroid 2-related factor 2 (ERK/NRF2) and mitogen-activated protein kinase (MAPK) pathways [17,18,19].
Considering its range of anti-tumor properties and a reliable safety profile, chrysin is increasingly being recognized for its potential in oncological treatments [20]. However, many questions remain about precisely how chrysin inhibits the growth of cancer cells. Recent research indicates that the wingless int-1/beta-catenin (WNT/β-catenin) pathway, which involves a series of proteins regulating cell growth and division, plays a pivotal role in cell proliferation, migration, and invasion across various cancer types [21,22]. The present study hypothesizes that the effect of chrysin on inducing apoptosis in the ER-negative breast cancer cells (MDA-MB-231) might be via the crucial WNT/β-catenin pathway. If chrysin can modulate this pathway's interactions, it could be one of the primary mechanisms behind chrysin's anticancer effect. This hypothesis could contribute to a better understanding of chrysin's potential therapeutic treatment. It may also pave the way for new strategies on how modulating this pathway can be used in cancer treatment. This research provides a detailed analysis of chrysin's potential anti-proliferative, apoptotic, and anti-invasive effects on MDA-MB-231 cells, underscoring the importance of the WNT/β-catenin signaling pathway as a therapeutic target in this process.
2. Results
2.1. Chrysin inhibits the proliferation of MDA-MB-231 cells.
The XTT assay was used to assess the cytotoxic effects of various concentrations (5, 10, 15, 20, 30, 40, 50, 75, 100 ve 150 µM) of chrysin and durations (24h and 48h) of exposure on MDA-MB-231 cells. Figure 1A shows that treating the MDA-MB-231 cell line with chrysin concentrations of 5, 10, 15, 20, 30, 40, 50, 75, 100, and 150 µM for 24 and 48 hours led to a dose- and time-dependent decrease in cell viability. After treating the MDA-MB-231 cell line with chrysin for 48 hours, a decrease in cell viability was observed starting from a concentration of 75 µM. Additionally, the half-maximal inhibitory concentration (IC50) was calculated to be 115.77 µM. Since cytotoxic activity was determined between 100 and 150 µM, subsequent experiments were conducted at a dose concentration of 150 µM. Based on the findings from the colony analysis, the ability of MDA-MB-231 cells to form colonies was significantly diminished following 7 days treatment with chrysin (Figure 1B). For the control group, the number of colonies for MDA-MB-231 cells was recorded as 265 ± 12.31, while the chrysin-treated group showed a reduced count of 123.04 ± 6.42 (*** p < 0.0001). Overall, both XTT and colony formation analyses indicate that chrysin effectively suppressed the proliferation of MDA-MB-231 cells.
2.2. Chrysin reduced the invasive potential of MDA-MB-231 cells.
Using a Matrigel invasion chamber, the effect of chrysin on the invasion capabilities of MDA-MB-231 cells was analyzed. After treating the cells for 48 hours with chrysin, a significant reduction in cell invasion was detected when compared with the control group. The number of invasive cells in the control was found to be 165 ± 5.48, while for the MDA-MB-231 cells subjected to chrysin, the count decreased to 57 ± 2.14 (***p < 0.0001) (Figure 2A).
2.3. Chrysin induced apoptotic cell death by WNT/β-catenin pathway in MDA-MB-231 cells.
In the detection of chrysin's apoptotic effect on MDA-MB-231 cells, according to the Annexin V assay, the total apoptotic cell percentage for chrysin was determined to be 32.46%, while it was 49.81% for the control. The values for early apoptosis, late apoptosis, and dead cells are shown in Figure 1D (** p < 0.001). To confirm the results of the Annexin V assay, a caspase 3/7 activity analysis was conducted. There was a significant increase in caspase 3/7 activity for chrysin-treated cells compared to the control. based on the (Figure 2B) (*p < 0.05).
In addition, in MDA-MB-231 cells treated with chrysin, the level of GSK3B, which plays a central role in regulating the stability and activation of β-catenin, increased compared to the control, while the level of β-catenin showed a significant decrease. These findings indicate that chrysin stimulates the apoptosis of MDA-MB-231 cells and that, following chrysin treatment, GSK3β may promote the proteasomal degradation of β-catenin by phosphorylating it (Figure 2B) (*p < 0.05).
2.4. Chrysin effects the expression profiles of genes associated with apoptosis and the WNT/β-catenin pathway in MDA-MB-231 cells.
After treating MDA-MB-231 cells with chrysin, significant modulations were observed in genes involved in the WNT/β-catenin pathway using RT-qPCR. Chrysin treatment in MDA-MB-231 cells led to significant modulations in the genes associated with the WNT/β-catenin pathway. Specifically, there was an upregulation in APC (66.60 fold; p = 0.000625), AXIN1 (27.14 fold; p = 0.002701), AXIN2 (2.43 fold; p = 0.000280), CTNNB1 (23.79 fold; p = 0.00224), LRP5 (12.50 fold; p = 0.001958), LRP6 (21.00 fold; p = 0.00874), TCF7 (5.31 fold; p = 0.00213), and LEF1 (12.97 fold; p = 0.00071). In contrast, genes such as GSK3A (-20.86 fold; p = 0.00096), GSK3B (-9.21 fold; p = 0.00088), C5NKA1 (-34.12 fold; p = 0,00056), and c-MYC (-2.57 fold; p = 0.0057) showed reduced expression. However, alterations in DVL1, DVL2, and CASP8 gene levels were not statistically significant. Additionally, there was a pronounced increase in the expression levels of apoptotic genes, namely BAX (1.2 fold; p = 0.00198), CASP3 (13.38 fold; p = 0.000803), CASP7 (7.13 fold; p = 0.000803) and CASP9 (8.38 fold; p = 0.00237). On the contrary, the expression of the BCL-2 gene, which is anti-apoptotic experienced a decrease (-4.62 fold; p = 0.00034) (Figure 3).
2.5. Computational ADME and molecular docking evaluations suggest a significant role for chrysin in the WNT/β-catenin signaling pathway.
The SMILES data offers significant insights into the potential of chrysin as a drug candidate. Preliminary results indicate that chrysin is efficiently absorbed from the gastrointestinal tract, can cross the blood-brain barrier, and complies with the druglikeness rules set by Lipinski, Ghose, Veber, Egan, and Muegge. According to the molecular docking analysis, the ensuing binding energies were observed to be -7.4 kcal/mol for APC, -7.6 kcal/mol for c-MYC, -7.9 kcal/mol for GSK3α, -7.1 kcal/mol for GSK3β, -9 kcal/mol for DVL1, -6.5 kcal/mol for AXIN, and -6.9 kcal/mol for LRP6 (Figure 4).
3. Discussion
For many years, plant-derived metabolites have been at the forefront of research into potential therapeutic agents for the treatment or prevention of cancer. In this study, the anticancer activity of the chrysin metabolite against MDA-MB-231 cells was investigated. Initially, based on antiproliferative effect results, chrysin displayed cytotoxic activity against MDA-MB-231 cells for 48 hours with an IC50 dose of 115.77 μM. The potential therapeutic efficacy of chrysin was further investigated in more detail through apoptosis, colony formation, cell invasion, and gene and protein expression studies related to the WNT/β-catenin signaling pathway. Additionally, computational ADME and molecular docking evaluations were conducted to assess chrysin's potential as a drug candidate. It was determined that chrysin, at a dose of 150 μM, stimulated cell death and reduced invasion in MDA-MB-231 cells, supporting its antiproliferative effect. Furthermore, the roles of apoptosis and the WNT/β-catenin pathway in bringing about this effect were revealed. There are other studies that support the anticancer activity of chrysin. While studies related to chrysin in the MDA-MB-231 cell line, an ER-negative cell line, are quite limited, it has been reported that chrysin inhibits the proliferation of cancer cells. In a study examining the cytotoxic activity of chrysin against MDA-MB-231 cells, researchers reported that cyrysin inhibited cell proliferation in a dose-dependent (100 μM) and time-dependent (48 hours) manner. However, it did not lead to apoptotic cell death. Additionally, they noted that the increase in PPARα mRNA expression is likely related to cell proliferation following chrysin treatment. In this study, apart from the expression information of PPARα, -β, and γ mRNA, there is no molecular and cellular data [23]. Another study identified the cytotoxic activity of extracts from Scutellaria species and metabolites including chrysin against the MDA-MB-231 cell line as 100 μM and determined that chrysin stimulated cell apoptosis in the G2 phase in the U87 cell line. Apart from this, the study did not include molecular data analysis, and only the apoptotic effect of chrysin in U87 cells was demonstrated [24]. Lastly, Sun et al. (2012) concluded their study by demonstrating that the treatment of 40 μM chrysin led to a notable inhibition of cell growth in MDA-MB-231 cells [25]. Besides chrysin has also demonstrated cytotoxic activity against various other cell lines. Specifically, its effects were observed at 14.2 μM in Hela cell lines over 72h [26], 40 mM in PC-3 cells over 24 [27], 20-80 μM in human bladder cancer cell lines t-24 and 5637 over 24h [28], and 25 μM in human mucoepidermoid carcinoma MC-3 cells over 24h [29].
Apoptosis represents the essential molecular action responsible for the anticancer effects of various potential anticancer agents, including chrysin. After the treatment of chrysin, the results of apoptosis in present study showed an increase in caspase 3/7 activity in MDA-MB-231 cells, as well as an increase in the expression of CASP3, -7, and -9 mRNAs, and a decrease in CASP8 and anti-apoptotic BCL-2 mRNA levels, suggesting that apoptosis might have occurred via the intrinsic pathway. However, only the level of CASP8 is insufficient to state that chrysin does not stimulate apoptosis via the extrinsic pathway. The induction of cellular apoptosis by chrysin has also been demonstrated in KYSE-510 [30] and A549 [31] cells. Furthermore, reports indicating that chrysin increases the BAX/BCL-2 ratio in A549 cells and triggers the activity of caspase-3 and -9, leading to cell death, are in line with the findings of this study [31]. Moreover, Al-Oudat and colleagues (2019) asserted that their synthesized chrysin analog triggered apoptosis in MDA-MB-231 cells through the activation of caspase 3/7 [32]. Lastly, Jung and collaborators (2022) reported that chrysin induced apoptosis and autophagy in the MC-3 cell line via activation of the MAPK pathway. They demonstrated that the viability of MC-3 cells significantly decreased after 24 hours of treatment with chrysin beyond 25 μM. Based on the data obtained from Annexin V/propidium iodide (PI) staining, analyzed using flow cytometry, exposure to varying concentrations of chrysin (0, 50, and 100 μM) resulted in apoptosis rates of 6.6%, 12.5%, and 16.5%, respectively [29]. According to the flow cytometry results of present study, the percentages of early apoptosis and late apoptosis in MDA-MB-231 cells treated with chrysin were determined as 3.12% and 46.69%, respectively. The reason for the difference in the apoptosis rate from Jung et al. (2022) could be due to the use of different cells, and differences in the treatment dose and duration.
The MDA-MB-231 cell line is frequently associated with the activation of the WNT/β-catenin pathway in various literature. In this cell line, the stabilization and accumulation of β-catenin in the cell nucleus have been observed to increase cell proliferation, invasion, and metastasis [22,33]. Additionally, specific compounds or drugs targeting the WNT/β-catenin pathway have been able to suppressed the growth and metastatic potential of MDA-MB-231 cells [20,34]. In this context, this signaling pathway is crucial due to its potential as a potential therapeutic focus in managing triple-negative breast cancer. In the WNT/β-catenin signaling pathway, β-catenin is located in the cytoplasm bound to the APC, AXIN, and GSK3 protein complex. When the WNT ligand binds to its receptor, the activated DVL disrupts the function of this complex, and DVL's binding to β-catenin facilitates the translocation and stabilization of β-catenin to the nucleus. Consequently, this leads to an increase in the transcription of target genes in the nucleus, such as c-MYC, triggering the cell's growth, survival, differentiation, and proliferation. A study has presented that chrysin increases the expression of GSK3B, promotes phosphorylation of β-catenin in Ser33/37 in the cytoplasm, leading to the degradation of β-catenin and attenuation of the WNT/β-catenin signaling pathway in a hepatocarcinogenesis rat model [35]. In the present study, chrysin significantly increased the GSK3B protein level while decreasing the β-catenin level in MDA-MB-231 cells. Additionally, due to the chrysin’s potential to mediate the inhibition of the WNT/β-catenin pathway, changes in the expression levels of genes involved in this pathway have been investigated. As a result of the RT-qPCR analysis, a significant increase in the expression of key genes in this pathway, namely APC, AXIN1, AXIN2, GSK3A, GSK3B, CK1α, and CTNNB1, suggests that the WNT/β-catenin pathway could be suppressed by chrysin. This increase in expression of these genes is associated with the activation of the "destruction complex" which inhibits the pathway's activation. The activation of the destruction complex leads to the phosphorylation of β-catenin and its proteasomal degradation, ensuring that β-catenin remains at low levels in the cytoplasm. On the other hand, the observed decrease in genes LRP5, LRP6, TCF7, LEF1, and c-MYC, which facilitate the activation of the pathway, is another evidence supporting this suppression. This indicates that the WNT ligand cannot bind to LRP receptors, and β-catenin, which normally becomes stable in the cytoplasm and translocates to the nucleus to interact with transcription factors like TCF7 and LEF1, cannot initiate the expression of target genes like c-MYC. Based on these findings, it can be concluded that chrysin might successfully inhibit the WNT/β-catenin pathway in MDA-MB-231 cells, steering them towards apoptosis. These results suggest that the WNT/β-catenin pathway is an essential target in evaluating chrysin as a potential therapeutic agent.
4. Materials and Methods
4.1. Cell culture and reagents
MDA-MB-231 breast cancer cells (ATCC® HTB-26TM) were cultivated in an incubator set at 37°C under a 5% CO2 atmosphere, using RPMI-1640 medium supplemented with 10% heat-treated fetal bovine serum (FBS) and 1% antibiotic solution of streptomycin/penicillin. As the cell growth reached between 80-90% confluence, subculturing was done. The growth medium underwent replacement every 2 to 3 days. Chrysin was purchased from Sigma-Aldrich Co. (Sigma, St. Louis, MO, USA, #C80105) and subsequently dissolved in 0.1% dimethyl sulfoxide (DMSO) for later use.
4.2. Cell proliferation assay
Using the XTT [2,3-bis (2-Methoxy-4-nitro-5-sulfophenyl) 2H tetrazolium- 5-carboxanilide] assay, the cell viability of MDA-MB-231 cells were assessed. The cells were placed into 96-well plates using RPMI-1640 medium with a concentration of 1 x 103 cells for each well. The cells were incubated at 37°C in a 5% CO2 environment for periods of 24 and 48 hours to detect cell proliferation. Following the addition of 50 μL of XTT solution into each well, the plates were then allowed to incubate for 4 hours at 37°C, shielded from light. Living cells generated soluble formazan, the absorbances of which were measured using an ELISA plate reader. The measurement was recorded at 450 nm while using 630 nm as the reference wavelength. The viability percentage was determined using the adjusted absorbance values in the following manner: Viability (%) = Absorbance of the experimental well/Absorbance of the control well × 100.
4.3. RNA isolation and real time quantitative PCR analysis (RT-qPCR)
RNA was isolated using the RiboEx reagent (GeneAll) as directed by the manufacturer, and was then stored at −80°C. The RNA concentrations were determined using a nanodrop, quantified in nM, and then adjusted to 1 μg for each RNA sample. Following this, cDNA was synthesized with the iScriptTM cDNA synthesis kit (BioRad, #1708890) as per the provided instructions. For the quantitative assessment of gene expression, we focused on APC, AXIN1, AXIN2, GSK3A, GSK3B, CK1α, CTNNB1, LRP5, LRP6, DVL1, DVL2, TCF7, LEF1, and c-MYC genes associated with the WNT/β-catenin pathway, as well as CASP3, -7, -8, -9, BAX and BCL-2 genes related to the apoptotic pathway. SyberGreen was used as DNA-binding molecule. The primer sequences were created using IDT PrimerQuest (http://eu.idtdna.com/home/home.aspx). Details are provided in Table 1.
4.4. Caspase 3/7 activation
Using the AmpliteTM colorimetric caspase 3/7 assay kit (AAT Bioquest, Sunnyvale, CA, USA, #13507) the activity levels of caspase 3/7 were determined in cell treated with chrysin. Cells, seeded in 96 well plates, were incubated for 24 hours before treatment and for an additional 48 hours post-treatment. After treatment, cells were lysed, and the reaction buffer was added, followed by the caspase 3/7 substrates in 96 well plate. The plate was incubated in the dark at room temperature for 1 hour, then centrifuged at 800 rpm for 2 minutes, and the absorbance increase is measured at 490 nm using a ELISA plate reader. The change in caspase activity was determined by comparing the readings from the chrysin-treated samples against the untreated controls.
4.5. Flow cytometry analysis
To assess the rate of apoptosis and to determine between necrotic, early and late apoptotic stages, the Muse™ Annexin V&Dead Cell from Merck Millipore, Burlington, MA, USA was employed. First, MDA-MB-231 cells were seeded to a concentration of 1x106 cells per mL. Subsequently, reagents from the Muse™ Annexin V&Dead Cell kit were introduced to the cell mixture. This was then left to sit for 20 minutes at room temperature, away from light.
4.6. Cell colony formation assay
To assess the effect of chrysin on the colony forming ability of MDA-MB-231 cells, a colony formation assay was conducted. Initially, cells were plated at a density of 1x103 cells per well in 6-well plates and incubated for a day. Post-incubation, they underwent chrysin treatment and were then subjected to subculturing every alternate day. After a 7-day period, the media were rinsed with PBS. The cells were subsequently fixed with 100% methanol at -20°C. After this, colonies from both the control and treated groups were stained with 1.0% crystal violet for 10 minutes and then imaged using an inverted microscope. The colony forming ability was deduced by comparing the number of colonies across the groups [36].
4.7. Cell invasion assay
The invasion potential of MDA-MB-231 cells after chrysin treatment was evaluated using Corning BioCoat™ Matrigel® Invasion Chambers, USA, in accordance with the manufacturer's instructions. Cells were suspended at a concentration of 2.5×104 cells/mL and introduced into the top chambers, with the bottom chambers filled with growth medium enhanced with 5% FBS, to act as a chemoattractant. After 22 hours, non-invading cells were removed from the top chamber using a cotton swab, while the invading cells were fixed with 100% methanol and stained with 0.5% crystal violet for 2 minutes each. Using a light microscope, the stained cells were visualized, images captured with the ZEN software (Carl-Zeiss-Promenade Jena, Germany) and invasion numbers analyzed using same software.
4.8. GSK3B and β-katenin activity assay
GSK3B is one of the key proteins responsible for the stability of β-catenin when the WNT pathway is in an inactive state or is suppressed, inhibiting the entry of β-catenin into the cell nucleus and its gene expression. To determine the GSK3B levels, human phospho-GSK3B ELISA (MyBioSource Inc. CA, USA, #MBS9511030) was used, following manufacturer instructions. Briefly, MDA-MB-231 cells were cultivated until they reached 80-90% confluency and were then treated with chrysin. Following the incubation, 100 μL of cell lysate from each sample was transferred to an ELISA plate coated for capture purposes and was further incubated for 2 hours. After this period, the plate was washed, and 100 μL of 1X Streptavidin-HRP variant was added to each well and incubated for another 30 minutes. At the end of this duration, 100 μL of Stop Solution was added, and the absorbance of the samples was measured at a wavelength of 450 nm using an ELISA plate reader.
For the detection of β-catenin levels, the highly sensitive β-catenin ELISA kit (Enzo Life Sciences, NY, USA, #P35222), capable of detecting as little as 33.8 pg/mL β-catenin, was applied in accordance with the manufacturer's instructions. Briefly, samples were mixed with the assay buffer and incubated at room temperature for 1 hour. Subsequently, the primary antibody was introduced and incubated for an additional hour. Following five plate washes, 100 μL of a blue conjugate was added. After another five washes, 100 μL of a soluble substrate was introduced for a 30 minute incubation period. The reaction was stopped by adding a stop solution, and optical readings were then taken at 450 nm.
4.8. Computational ADME and molecular docking evaluation of chrysin's role in the WNT/β-catenin signaling pathway
The pharmacokinetic parameters of chrysin, encompassing absorption, distribution, metabolism, and excretion, were assessed using the in silico ADME analysis through the SwissADME tool (http://www.swissadme.ch/index.php#). The SMILES data for the compound was sourced from PubChem (https://pubchem.ncbi.nlm.nih.gov/compound/5281607). Besides, from the gene expression analyses, notable expressions were identified for genes such as AXIN (PDB ID 1DK8), APC (PDB ID 5IZ6), c-MYC (PDB ID 6AU4), LRP6 (PDB ID 6GBI), GSK3A (PDB ID 7SXF), DVL1 (PDB ID 6TTK), and GSK3B (PDB ID 1O9U). The three-dimensional protein structures of these genes were downloaded from the RCSB protein database (https://www.rcsb.org/). Molecular docking studies were then performed to understand the binding interactions between these protein structures and chrysin using the in silico platform (cadd.labshare.cn/cb-dock2/php/blinddock.php).
4.9. Statistical Analysis
All experiments were conducted in triplicate, and the results were presented as the mean ± standard deviation (SD). Statistical differences between groups were assessed using the Student's t-test and one-way ANOVA via GraphPad Prism software (version 10.0.2; GraphPad Software, Inc., La Jolla, CA, USA). To identify statistically significant alterations in mRNA expressions among the groups, an online analysis tool (RT² Profiler™ PCR Array Data Analysis) was utilized, with a p - value less than 0.05 considered indicative of statistical significance.
5. Conclusion
It was demonstrated in this study that chrysin effectively inhibits the invasion and colony formation abilities of MDA-MB-231 cells, induces cell apoptosis at both cellular and molecular levels, and suppresses the WNT/β-catenin pathway. Moreover, according to in situ data, chrysin exhibits drug-like ADME properties. Molecular docking results further revealed that chrysin can energetically and specifically interact with genes in the WNT/β-catenin pathway where we detected expression changes. These findings bolster the potential therapeutic implications of chrysin and its modulatory activities on these genes. It is hoped that this study will lay the groundwork for data that will enable the use of chrysin in pre-clinical and clinical settings against breast cancer in the future.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The author would like to thank Assoc. Prof. İlknur Çınar Ayan and Assoc. Prof. Ebru Güçlü for their contributions throughout the related research process.
Conflicts of Interest
The author declares no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- World Health Organization. (2020). Breast cancer: Prevention and control.
- National Cancer Institute. (2021). Physician Data Query (PDQ). Breast Cancer Treatment – Health Professional Version. Accessed at https://www.cancer.gov/types/breast/hp/breast-treatment-pdq on August 10, 2021. 10 August.
- Shahbaz, M.; Naeem, H.; Imran, M.; Hassan, H.U.; Alsagaby, S.A.; Al Abdulmonem, W.; Waqar, A.B.; Ghorab, A.H.; Abdelgawad, M.A.; Ghoneim, M.M.; et al. Chrysin a promising anticancer agent: recent perspectives. Int. J. Food Prop. 2023, 26, 2294–2337. [Google Scholar] [CrossRef]
- Mani, R.; Natesan, V. Chrysin: Sources, beneficial pharmacological activities, and molecular mechanism of action. Phytochemistry 2018, 145, 187–196. [Google Scholar] [CrossRef]
- Zanoli, P.; Avallone, R.; Baraldi, M. Behavioral characterisation of the flavonoids apigenin and chrysin. Fitoterapia 2000, 71, S117–S123. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Ali, N.; Rashid, S.; Jain, T.; Nafees, S.; Tahir, M.; Khan, A.Q.; Lateef, A.; Khan, R.; Hamiza, O.O.; et al. Alleviation of hepatic injury by chrysin in cisplatin administered rats: Probable role of oxidative and inflammatory markers. Pharmacol. Rep. 2014, 66, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.C.; Antunes, M.S.; Filho, C.B.; Del Fabbro, L.; de Gomes, M.G.; Goes, A.T.R.; Donato, F.; Prigol, M.; Boeira, S.P.; Jesse, C.R. Flavonoid Chrysin prevents age-related cognitive decline via attenuation of oxidative stress and modulation of BDNF levels in aged mouse brain. Pharmacol. Biochem. Behav. 2015, 134, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Rani, N.; Bharti, S.; Bhatia, J.; Nag, T.; Ray, R.; Arya, D.S. Chrysin, a PPAR-γ agonist improves myocardial injury in diabetic rats through inhibiting AGE-RAGE mediated oxidative stress and inflammation. Chem.-Biol. Interact. 2016, 250, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Wadibhasme, P.G.; Ghaisas, M.M.; Thakurdesai, P.A. Anti-asthmatic potential of chrysin on ovalbumin-induced bronchoalveolar hyperresponsiveness in rats. Pharm. Biol. 2011, 49, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Filho, C.; Jesse, C.; Donato, F.; Giacomeli, R.; Del Fabbro, L.; Antunes, M.d.S.; de Gomes, M.; Goes, A.; Boeira, S.; Prigol, M.; et al. Chronic unpredictable mild stress decreases BDNF and NGF levels and Na+,K+-ATPase activity in the hippocampus and prefrontal cortex of mice: Antidepressant effect of chrysin. Neuroscience 2015, 289, 367–380. [Google Scholar] [CrossRef]
- Kasala, E.R.; Bodduluru, L.N.; Madana, R.M.; V, A.K.; Gogoi, R.; Barua, C.C. Chemopreventive and therapeutic potential of chrysin in cancer: mechanistic perspectives. Toxicol. Lett. 2015, 233, 214–225. [Google Scholar] [CrossRef]
- Garg, A.; Chaturvedi, S. A Comprehensive Review on Chrysin: Emphasis on Molecular Targets, Pharmacological Actions and Bio-pharmaceutical Aspects. Curr. Drug Targets 2022, 23, 420–436. [Google Scholar] [CrossRef]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic Effects of Chrysin in Human Cancer Cell Lines. Int. J. Mol. Sci. 2010, 11, 2188–2199. [Google Scholar] [CrossRef]
- Yang, B.; Huang, J.; Xiang, T.; Yin, X.; Luo, X.; Huang, J.; Luo, F.; Li, H.; Li, H.; Ren, G. Chrysin inhibits metastatic potential of human triple-negative breast cancer cells by modulating matrix metalloproteinase-10, epithelial to mesenchymal transition, and PI3K/Akt signaling pathway. J. Appl. Toxicol. 2014, 34, 105–112. [Google Scholar] [CrossRef]
- Talebi, M.; Talebi, M.; Farkhondeh, T.; Simal-Gandara, J.; Kopustinskiene, D.M.; Bernatoniene, J.; Samarghandian, S. Emerging cellular and molecular mechanisms underlying anticancer indications of chrysin. Cancer Cell Int. 2021, 21, 214. [Google Scholar] [CrossRef] [PubMed]
- Sassi, A.; Fredon, M.; Cotte, A.K.; Fuselier, C.; Schneider, C.; Martiny, L.; Monchaud, D.; Chekir-Ghedira, L.; Aires, V.; Delmas, D. Chrysin-Induced Regression of Angiogenesis via an Induction of DNA Damage Response and Oxidative Stress in In Vitro and In Vivo Models of Melanoma. Cells 2023, 12, 1561. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Jung, J. Upregulation of G Protein-Coupled Estrogen Receptor by Chrysin-Nanoparticles Inhibits Tumor Proliferation and Metastasis in Triple Negative Breast Cancer Xenograft Model. Front. Endocrinol. 2020, 11, 560605. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Sun, K.; Wang, X.; Pan, H.; Zhu, J.; Ji, X.; Li, X. Chrysin suppresses proliferation, migration, and invasion in glioblastoma cell lines via mediating the ERK/Nrf2 signaling pathway. Drug Des. Dev. Ther. 2018, ume 12, 721–733. [Google Scholar] [CrossRef]
- Saleh, D.O.; El-Nasr, N.M.A.; Fayez, A.M.; Ahmed, K.A.; Mohamed, R.A. Uro-protective role of chrysin against cyclophosphamide-induced hemorrhagic cystitis in rats involving the turning-off NF-κB/P38-MAPK, NO/PARP-1 and STAT-3 signaling cascades. Chem. Interactions 2023, 382, 110585. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, E.R.; Ang, H.L.; Asnaf, S.E.; Zabolian, A.; Saleki, H.; Yavari, M.; Esmaeili, H.; Zarrabi, A.; Ashrafizadeh, M.; Kumar, A.P. Broad-Spectrum Preclinical Antitumor Activity of Chrysin: Current Trends and Future Perspectives. Biomolecules 2020, 10, 1374. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Guan, H.; Fang, L.; Yang, Y.; Zhu, X.; Yuan, J.; Wu, J.; Li, M. MicroRNA-374a activates Wnt/β-catenin signaling to promote breast cancer metastasis. J. Clin. Investig. 2013, 123, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Fu, Q.; Li, S.; Li, J.; Liu, S.; Wang, Z.; Su, Z.; Song, J.; Lu, D. Emetine exhibits anticancer activity in breast cancer cells as an antagonist of Wnt/β-catenin signaling. Oncol. Rep. 2019, 42, 1735–1744. [Google Scholar] [CrossRef]
- Hong, T.B.; Rahumatullah, A.; Yogarajah, T.; Ahmad, M.; Yin, K.B. Potential Effects of Chrysin on MDA-MB-231 Cells. Int. J. Mol. Sci. 2010, 11, 1057–1069. [Google Scholar] [CrossRef]
- Parajuli, P.; Joshee, N.; Rimando, A.M.; Mittal, S.; Yadav, A.K. In vitroAntitumor Mechanisms of VariousScutellariaExtracts and Constituent Flavonoids. Planta Medica 2009, 75, 41–48. [Google Scholar] [CrossRef]
- Sun, L.-P.; Chen, A.-L.; Hung, H.-C.; Chien, Y.-H.; Huang, J.-S.; Huang, C.-Y.; Chen, Y.-W.; Chen, C.-N. Chrysin: A Histone Deacetylase 8 Inhibitor with Anticancer Activity and a Suitable Candidate for the Standardization of Chinese Propolis. J. Agric. Food Chem. 2012, 60, 11748–11758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, X.; Qu, L.; Wu, J.; Cui, R.; Zhao, Y. Chrysin and its phosphate ester inhibit cell proliferation and induce apoptosis in Hela cells. Bioorganic Med. Chem. 2004, 12, 6097–6105. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Afshari, J.T.; Davoodi, S. Chrysin reduces proliferation and induces apoptosis in the human prostate cancer cell line pc-3. Clinics 2011, 66, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tong, Y.; Ying, J.; Lei, Z.; Wan, L.; Zhu, X.; Ye, F.; Mao, P.; Wu, X.; Pan, R.; et al. Chrysin induces cell growth arrest, apoptosis, and ER stress and inhibits the activation of STAT3 through the generation of ROS in bladder cancer cells. Oncol. Lett. 2018, 15, 9117–9125. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.-H.; Lee, J.-H.; Han, S.-H.; Woo, J.-S.; Choi, E.-Y.; Jeon, S.-J.; Han, E.-J.; Jung, S.-H.; Park, Y.-S.; Park, B.-K.; et al. Chrysin Induces Apoptosis via the MAPK Pathway and Regulates ERK/mTOR-Mediated Autophagy in MC-3 Cells. Int. J. Mol. Sci. 2022, 23, 15747. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, X.-H.; Wang, Z.-J. Cytotoxicity of flavones and flavonols to a human esophageal squamous cell carcinoma cell line (KYSE-510) by induction of G2/M arrest and apoptosis. Toxicol. Vitr. 2009, 23, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, S.H.; Zafaryab; Nafees, S. ; Khan, A.; Ahmad, I.; Bin Hafeez, Z.; Alam Rizvi, M. Chrysin Sensitizes Human Lung Cancer Cells to Tumour Necrosis Factor Related Apoptosis-Inducing Ligand (TRAIL) Mediated Apoptosis. Asian Pac. J. Cancer Biol. 2019, 4, 27–33. [Google Scholar] [CrossRef]
- Al-Oudat, B.A.; Alqudah, M.A.; Audat, S.A.; Al-Balas, Q.A.; El-Elimat, T.; Hassan, M.A.; Frhat, I.N.; Azaizeh, M.M. Design, synthesis, and biologic evaluation of novel chrysin derivatives as cytotoxic agents and caspase-3/7 activators. Drug Des. Dev. Ther. 2019, ume 13, 423–433. [Google Scholar] [CrossRef]
- King, T.D.; Suto, M.J.; Li, Y. The wnt/β-catenin signaling pathway: A potential therapeutic target in the treatment of triple negative breast cancer. J. Cell. Biochem. 2012, 113, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Gangrade, A.; Pathak, V.; Augelli-Szafran, C.E.; Wei, H.-X.; Oliver, P.; Suto, M.; Buchsbaum, D.J. Preferential Inhibition of Wnt/β-Catenin Signaling by Novel Benzimidazole Compounds in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2018, 19, 1524. [Google Scholar] [CrossRef]
- Khan, M.S.; Halagowder, D.; Devaraj, S.N. Methylated chrysin induces co-ordinated attenuation of the canonical Wnt and NF-kB signaling pathway and upregulates apoptotic gene expression in the early hepatocarcinogenesis rat model. Chem. Interactions 2011, 193, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Ayan, I. .; Güçlü, E.; Vural, H.; Dursun, H.G. Piceatannol induces apoptotic cell death through activation of caspase-dependent pathway and upregulation of ROS-mediated mitochondrial dysfunction in pancreatic cancer cells. Mol. Biol. Rep. 2022, 49, 11947–11957. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) The cytotoxic effect of chrysin on MDA-MB-231 cell line. Cells were treated with control and chrysin (5-150 µM) for 24 and 48h. XTT cell proliferation assay was used for the detection of IC50 values. The dose and control group was subjected to least three independent experiments. (B) The effects of chrysin on colony numbers were evaluated using the colony formation assay in the MDA-MB-231 cell line (***p < 0.0001). (C) Determination of necrotic, early, and late apoptotic cells in MDA-MB-231 cell line after 48-hour treatment with chrysin using Annexin V (**p < 0.001).
Figure 1.
(A) The cytotoxic effect of chrysin on MDA-MB-231 cell line. Cells were treated with control and chrysin (5-150 µM) for 24 and 48h. XTT cell proliferation assay was used for the detection of IC50 values. The dose and control group was subjected to least three independent experiments. (B) The effects of chrysin on colony numbers were evaluated using the colony formation assay in the MDA-MB-231 cell line (***p < 0.0001). (C) Determination of necrotic, early, and late apoptotic cells in MDA-MB-231 cell line after 48-hour treatment with chrysin using Annexin V (**p < 0.001).
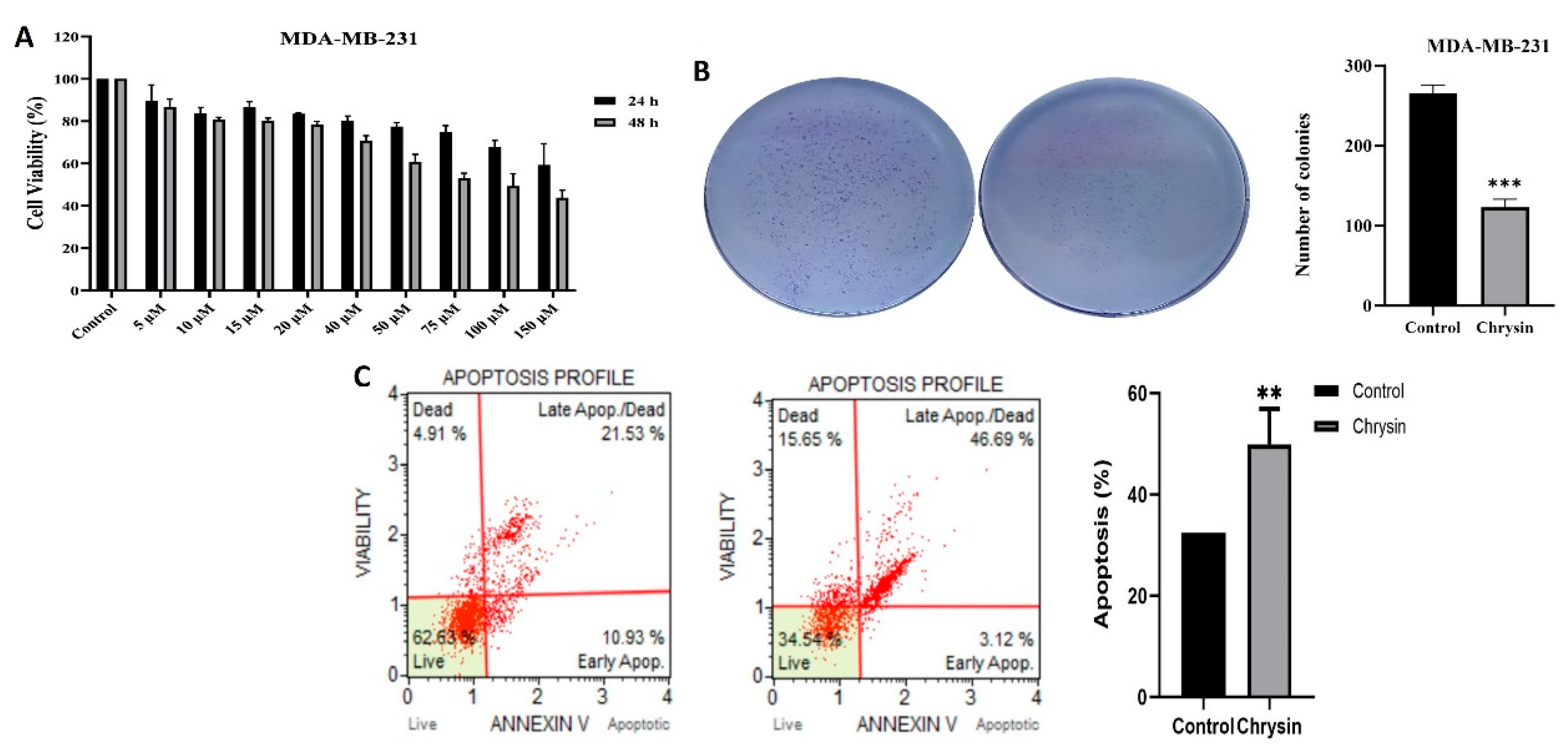
Figure 2.
(A) Invasion percentages determined by matrigel assay in MDA-MB-231 cells after the treatment of chrysin (***p < 0.0001). (B) ELISA results for changes in caspase 3/7 activity and GSK3B and β-catenin levels (*p < 0.05).
Figure 2.
(A) Invasion percentages determined by matrigel assay in MDA-MB-231 cells after the treatment of chrysin (***p < 0.0001). (B) ELISA results for changes in caspase 3/7 activity and GSK3B and β-catenin levels (*p < 0.05).
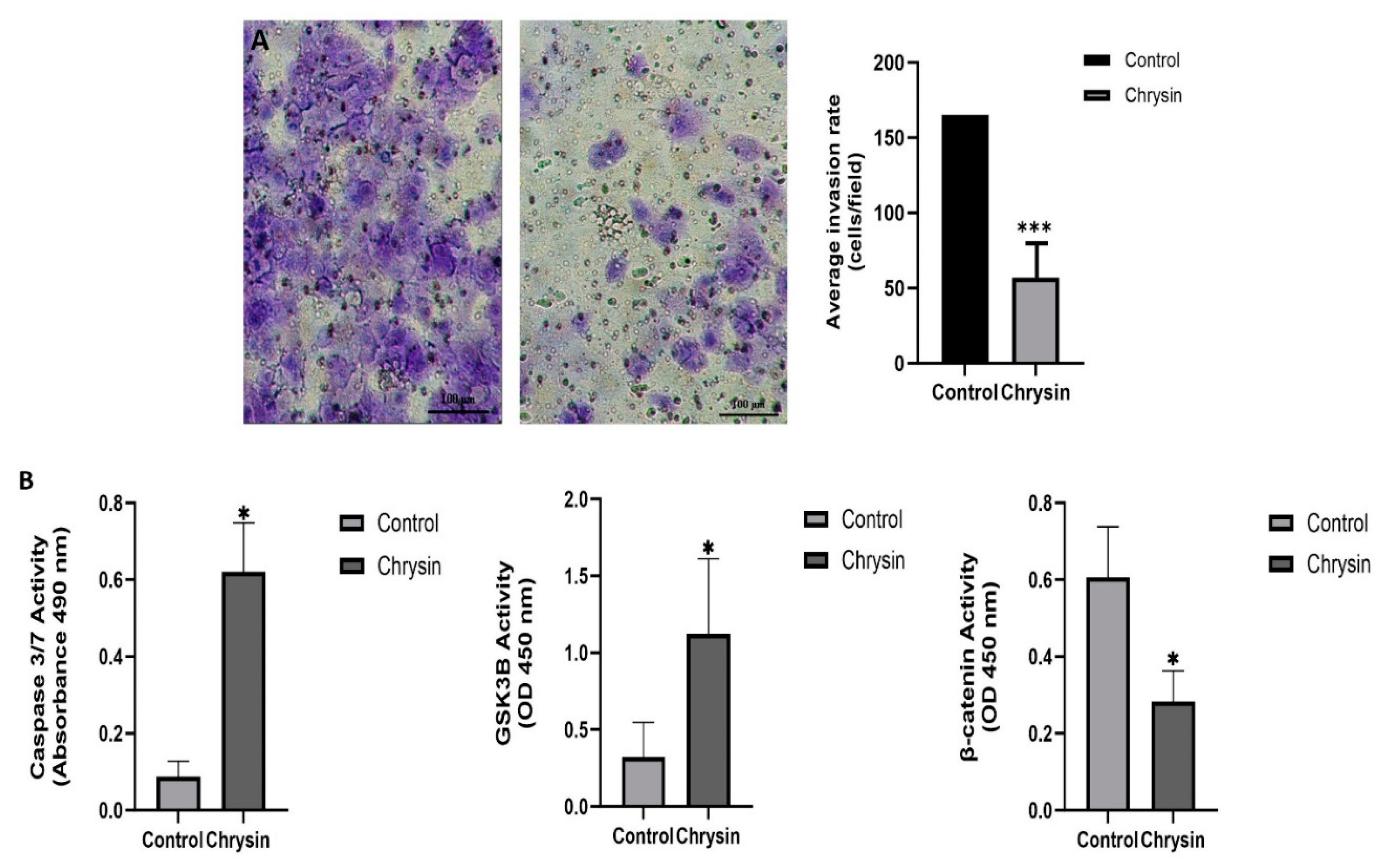
Figure 3.
Fold increase and decrease in the expression of genes related to apoptosis and WNT/β-catenin signaling pathway in MDA-MB-231 cells after chrysin treatment. Genes (DVL1 and DVL2) that did not show statistically significant expression changes are not included (*p < 0.05).
Figure 3.
Fold increase and decrease in the expression of genes related to apoptosis and WNT/β-catenin signaling pathway in MDA-MB-231 cells after chrysin treatment. Genes (DVL1 and DVL2) that did not show statistically significant expression changes are not included (*p < 0.05).
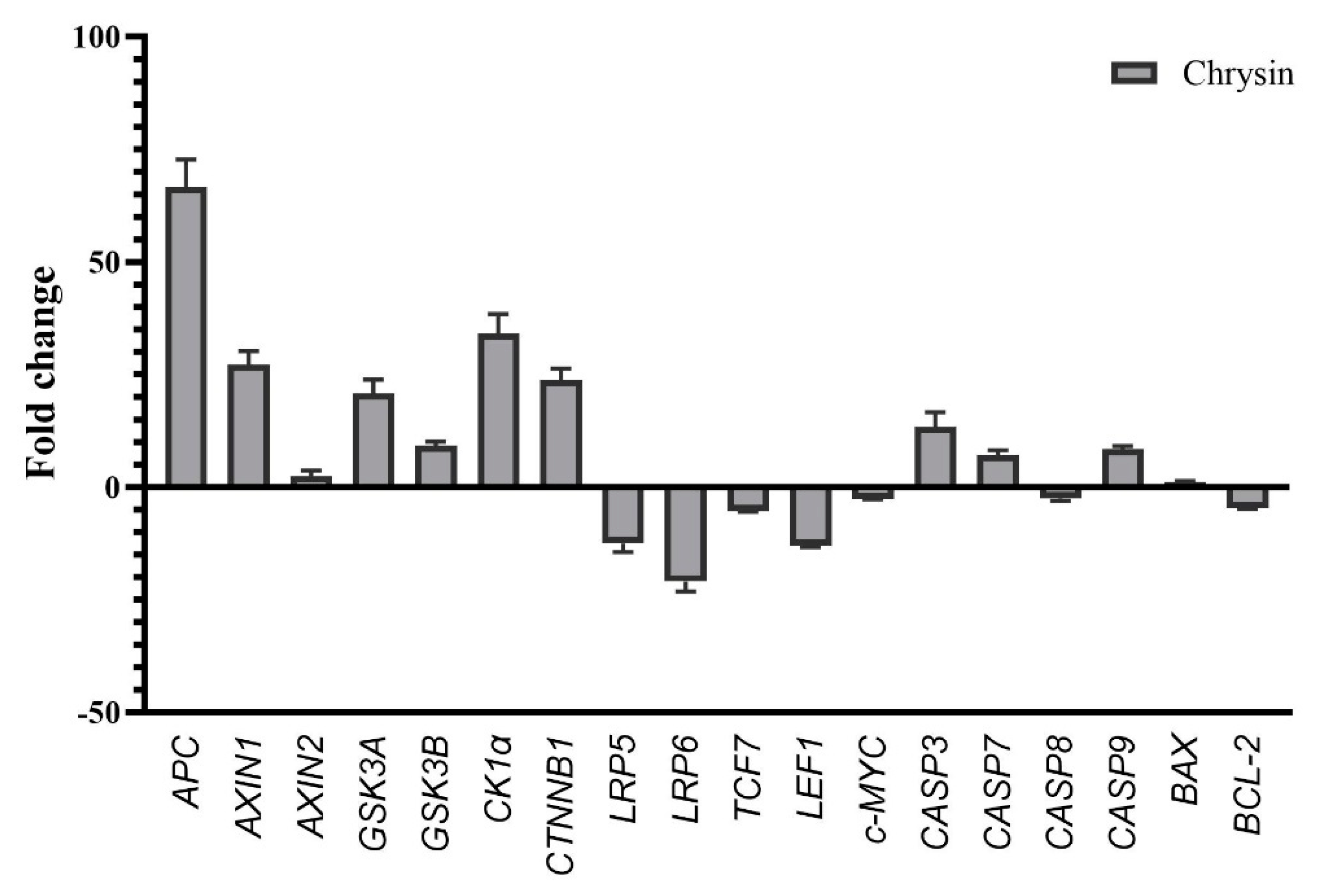
Figure 4.
SWISSADME results of chyrsin for evaluation of drug properties (1) and 3D Interaction of chyrsin with 1DK8 (2), 1O9U (2), 5IZ6 (3), 6AU4 (4), 7SXF (5), 6TTK (6) and 6GBI (7).
Figure 4.
SWISSADME results of chyrsin for evaluation of drug properties (1) and 3D Interaction of chyrsin with 1DK8 (2), 1O9U (2), 5IZ6 (3), 6AU4 (4), 7SXF (5), 6TTK (6) and 6GBI (7).
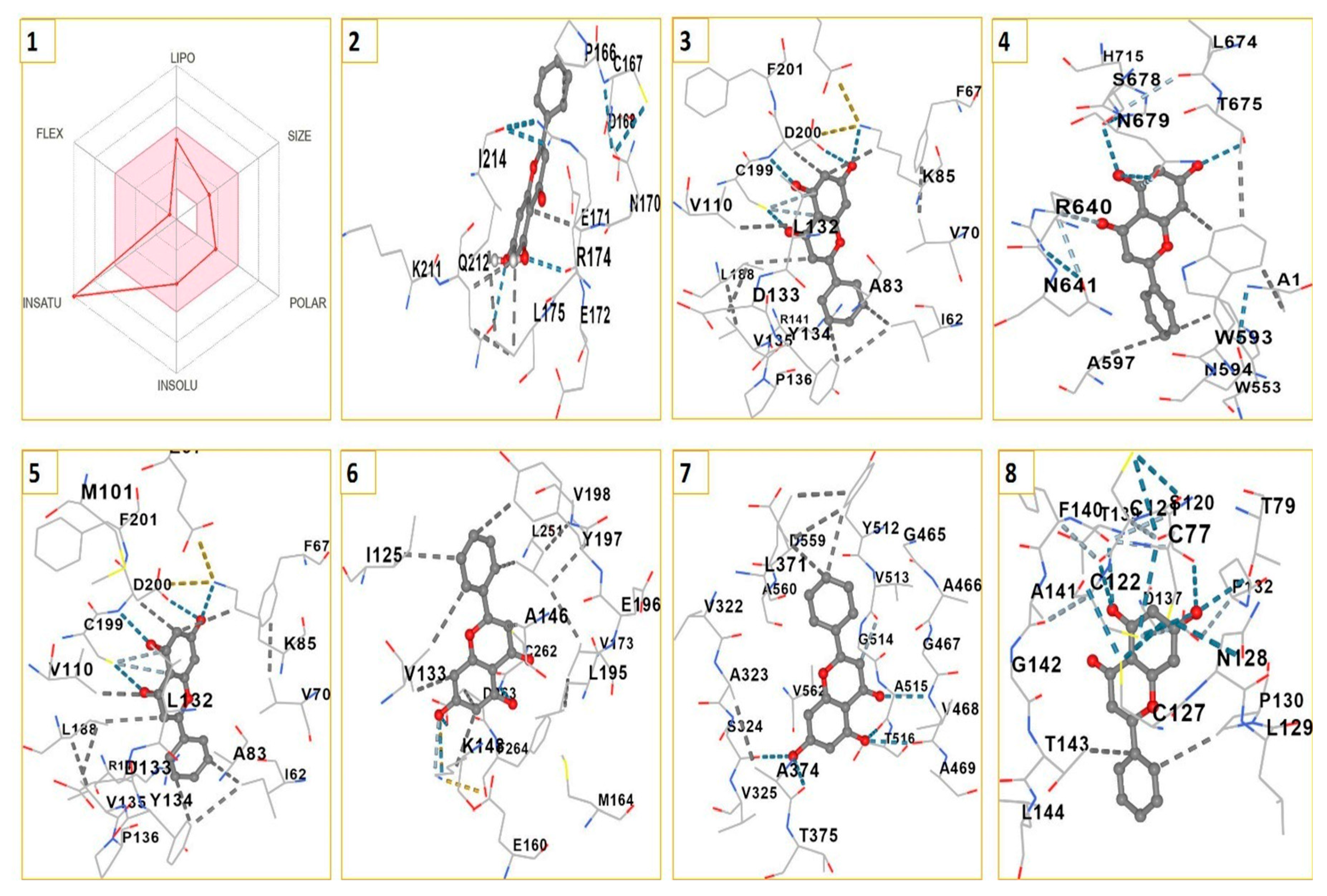

Table 1.
Primer sequences used RT-qPCR analysis.
| Gene | Gene Primer sequence | |
|---|---|---|
| CASP3 | F | GAGCCATGGTGAAGAAGGAATA |
| R | TCAATGCCACAGTCCAGTTC | |
| CASP7 | F | CGAAACGGAACAGACAAAGATG |
| R | TTAAGAGGATGCAGGCGAAG | |
| CASP8 | F | GCCCAAACTTCACAGCATTAG |
| R | GTGGTCCATGAGTTGGTAGATT | |
| CASP9 | F | CGACCTGACTGCCAAGAAA |
| R | CATCCATCTGTGCCGTAGAC | |
| BAX | F | GGAGCTGCAGAGGATGATTG |
| R | GGCCTTGAGCACCAGTTT | |
| BCL2 | F | GTGGATGACTGAGTACCTGAAC |
| R | GAGACAGCCAGGAGAAATCAA | |
| APC | F | TGGTTGGCACTCTTACTTACC |
| R | GCCTGTGGTCCTCATTTGTA | |
| AXIN1 | F | GAGGTATGTGCAGGAGGTTATG |
| R | TCCTCTGCGATCTTGTCTCT | |
| AXIN2 | F | CTTATCGTGTGGGCAGTAAGA |
| R | GTTCTCGGGAAATGAGGTAGAG | |
| GSK3A | F | CCTGGACAAAGGTGTTCAAATC |
| R | CAGACATCGCAGTTCATCAAAG | |
| GSK3B | F | GAGAGCTCCAGATCATGAGAAAG |
| R | GAACATAGTCCAGCACCAGATTA | |
| CK1α | F | GGTATTGGGCGTCACTGTAATA |
| R | GAGAAAGATGGGTCCTGAGAAG | |
| CTNNB1 | F | CTTCACCTGACAGATCCAAGTC |
| R | CCTTCCATCCCTTCCTGTTTAG | |
| LRP5 | F | CGTACAGGCCCTACATCATTC |
| R | GTCCGAGTTCAAATCCAGGTAG | |
| LRP6 | F | CCGAGTCAGAACCTGGAAATAC |
| R | CTCCAACTGATCTCCCATCTAATC | |
| DVL1 | F | GACTCATCCGGAAGCACAAA |
| R | GACATGGTGGAGTCGGTTATG | |
| DVL2 | F | CCTTCTCTGAGCAGTGCTATTAC |
| R | GCAGGGTATTGGTAGGAGAAAG | |
| TCF7 | F | ACTCTTCCCGGACAAACTTC |
| R | CAGATTGAAGGCGGAGTAGAC | |
| LEF1 | F | GATCACACCCGTCACACATC |
| R | ACCCGGAGACAAGGGATAAA | |
| GAPDH | F | TGAACGGGAAGCTCACTGG |
| R | TCCACCACCCTGTTGCTGTA | |
| ACTN | F | TGTGAACAAAGCGCTGGA |
| R | ATGGCGAACCTAAGGATGATG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



