
Submitted:
04 October 2023
Posted:
09 October 2023
You are already at the latest version
Abstract
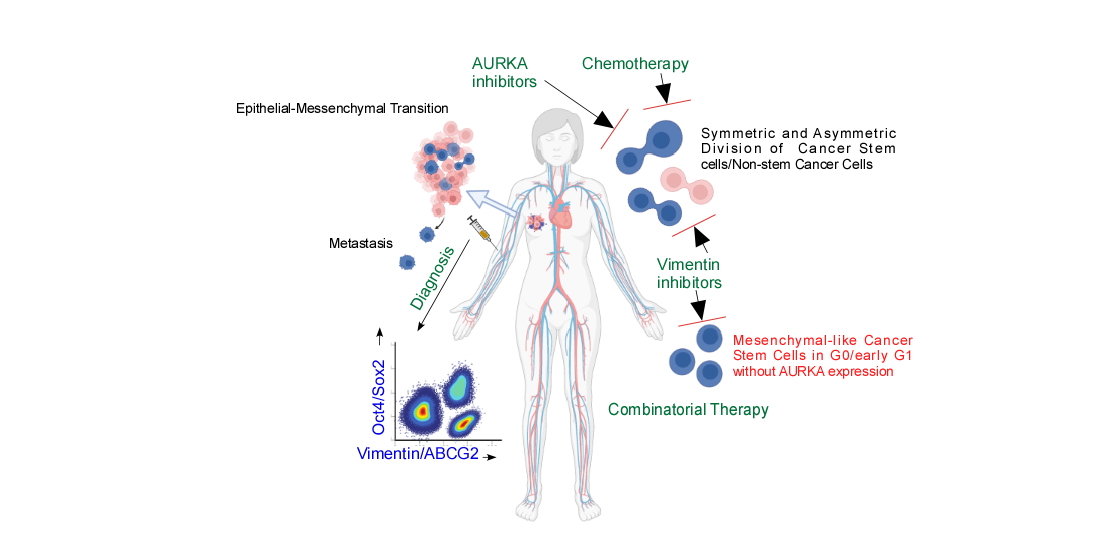
Aurora Kinase A being overexpressed in majority of cancers, appear to be an attractive therapeutic target. However, a Phase III clinical trial of Alisertib, a selective AURKA inhibitor, resulted in no better response compared to the comparator arm of chemotherapeutic regimen raising question regarding ability of the same to target the undetectable stem cell functions. In silico analysis indicated regulation of AURKA by the stemness factors Oct4 and Sox2. TCGA data indicated positive correlation of each of the factors with AURKA which were eventually validated in cell lines, patient tissues and blood by flow cytometry along with Oct4 binding on AURKA promoter being detected by ChIP assay. However, indirect immunofluorescence and cell cycle analyses indicated proliferation-independent AURKA functions during asymmetric cell division, a characteristic feature of stem cells. Thorough screening of the AURKA positive cells in patient samples denoted epithelial to mesenchymal transition and significant upregulation of Vimentin, a mesenchymal marker and ABCG2, a drug resistance marker under Oct4 or Sox2 influence. Overall, our study demonstrated combinatorial selection Oct4, Sox2, AURKA, Vimentin and ABCG2 for diagnostics and intervention of circulating breast cancer stem cells as a blood-based, cost-effective and simple approach which will be beneficial in reducing relapse.
Keywords:
Oct4
; Sox2
; AURKA
; Vimentin
; breast cancer
; Alisertib
; epithelial to mesenchymal transition
; asymmetric cell division
; NUMB
; stem cell
; diagnostics
; intervention
; chemotherapy
Introduction:
Breast cancer is the leading cause of cancer-related death in women with the age and sex-standardized incidence rate of ~47.8% as per GLOBOCAN 2020. There is about 0.5% increase in the incidence rate per year globally despite decrease in death rates, which is associated with improved awareness and screening resulting in early detection as per American Cancer Society [1]. However, about 30% of patients with apparent clinical remission present with disease recurrence in course of time as observed up to 24 years of follow-up studies [2]. In fact, often the actual statistics of patients showing clinical remission remains only an estimate since a majority presents for the initial detection at a considerably late age with a median age of diagnosis about 62 years while the global life expectancy itself is about 73 years [3]. Therefore, considering most of the time the minor initial symptoms being ignored by the patients and costly scans associated in the follow-up procedures, a cost-effective and simple detection system seems extremely necessary, which can be incorporated into yearly check-up.
With the discovery of cancer stem cells (CSCs) in 1994 [4], multifarious approaches have been made to identify cancer stem cell specific markers [5,6] to develop effective screening strategies without success. Disease recurrence has been shown to be resulting from this apparently unidentifiable population which happens to be plastic, low-maintenance, chemo –refractory, radiation-resistant and with the ability for self-renewal as well as differentiation [7,8,9]. The phenotypic switches selected by the stem cell niche contribute to change of CSC marker profile thereby failing the diagnostic protocols. Eventually, this population escapes the chemotherapeutic regimen due to their quiescent nature and chemotherapy being aimed at targeting the proliferating cells; leading to metastasis and disease recurrence after apparent clinical remission.
Importantly, breast cancer stem cells (BCSCs) have been shown to impart therapeutic resistance not only against chemotherapy through modulation of PI3K/Akt signaling, NFκB signaling etc. [10], but also against targeted endocrine therapy through manipulation of Notch/HIF/integrin/Akt signaling [11] implying the need to develop effective BCSC diagnostics and targeting strategies. For this purpose, identification of the changing marker profile of BCSCs during progression of the disease appears inevitable.
Aurora Kinase A (AURKA) gene amplification has been reported in basal-like breast cancer phenotypes [12]. Enhanced transcriptional activity by positive regulators like FOXM1, EGF, [13] TRAP200/MED1 complex, β-catenin/TCF-4 complex [14], Myc or inactivation of transcriptional repressors [15] etc have been also reported thereby presenting the same as an attractive therapeutic target [16] as well as a probable diagnostic marker. AURKA is a serine threonine kinase responsible for spindle organization, assembly and other mitotic functions [17] as well as non-mitotic functions [18] like regulation of cell motility, senescence and DNA repair etc. AURKA has also been reported to be involved in BCSC self renewal through induction of c-Myc, Sox2 and Nanog and enrichment of CD44hi/CD24low subpopulation [19]. Overexpression of AURKA in invasive breast cancer has been reported already [20] in 94% of cases irrespective of histological subtypes and AURKA protein expression has been already proven to be a better prognostic marker than Ki67 in ER-positive breast cancer cases [21] although not clinically implemented yet in practice. However, recently a phase III clinical trial of Alisertib as a single agent, reported no better prognosis in a 960-day timeframe compared to the comparator arm consisting of Gemcitabin/Pralatrexate/Romidepsin in relapsed or refractory peripheral T-cell lymphoma [22] proving that AURKA expression may be dispensible during dissemination of the breast cancer stem cells (BCSCs).
Our study was initially aimed at finding out the possible role of stemness factors in regulation of AURKA which was confirmed by in silico and in vitro cell line studies. Oct4 and Sox2 were found to be associated with regulation of AURKA and correlative expression of these stemness factors and AURKA were identified in patient tissues and blood. Interestingly, in an attempt to identify AURKA-mediated Oct4 or Sox2 functions, breast cancer cells were found to undergo epithelial-to-mesenchymal transition under G0/G1 phase. An insight into stem cell-specific asymmetric cell division of the breast cancer cells in vitro demonstrated differential AURKA distribution in the daughter cells along with phosphorylated-NUMB involving one differentiating breast cancer cell having colocalization of NUMB and AURKA and the other without any. The daughter cell without AURKA is expected to escape the AURKA inhibitors if the same chooses to remain at G0. Intriguingly, in patient blood, Oct4 or Sox2 and AURKA were not found to coexpress in the same cells although being positively correlated in terms of percent positive cells indicating the Oct4 or Sox2 positive cells without AURKA expression in blood may be in a phase like G0. These cells were found to be mesenchymal (Vimentin expressing) and positive for drug resistant marker ABCG2 confirming that the circulating Oct4+/Sox2+Vimentin+ABCG2+ cells are mesenchymal –like breast cancer cells and therefore extremely important for diagnostics and therapeutics.
Results:
AURKA is transcriptionally upregulated and associated with relapse in breast cancer:
To ensure that the AURKA protein overexpression mentioned earlier is supported by transcript expression as well in breast cancer, we performed retrospective analysis of 1085 invasive breast carcinoma samples and 112 paired breast samples from TCGA database (Figure 1A). Results demonstrated significant upregulation of the transcript in tumours compared to normal tissues across all the molecular subtypes hinting towards deregulatory events occurring at AURKA promoter of tumour samples. When the TCGA data was screened for survival analysis, this high AURKA expression as expected from the link to the stemness, was inversely proportional to the relapse free survival (RFS ) of the patients (Figure 1B) demonstrating the need for understanding the regulation of AURKA by the stemness factors.
Oct4 and Sox2 regulate AURKA expression which colocalizes with drug-resistance marker ABCG2 and phospho-NUMB:
To validate the positive correlation at protein level, we attempted silencing of Oct4 or Sox2 in MCF-7 and MDA MB-231 breast cancer cells which express both the stemness factors [23,24] and checked AURKA expression individually by flow cytometry. Results (Figure 2B) confirmed positive regulation of AURKA by Oct4 and Sox2 indicating that AURKA is indeed a downstream target whether or not the BCSCs depend on AURKA for surviving the therapies. Gating strategy for identification of Oct4/Sox2 +ve cells have been shown in Supplementary Data 1. ChIP assays confirmed binding of Oct4, but not Sox2 about 2kb upstream of AURKA TSS (Figure 2C). ChIP primers were designed spanning the regions designated in Figure 1C. In breast cancer patient tissues, Oct4, Sox2 and AURKA expressions appeared to be positively correlated to each other, but not necessarily in the same cells as demonstrated when per cell expression (mean fluorescence intensities i.e MFI) was considered (Figure 3A). Percentages of Oct4/Sox2/AURKA positive cells have been included in Supplementary Data 3. In peripheral blood however, percentages of Oct4 or Sox2 positive cells were significantly correlated to percentages of AURKA positive cells (Figure 3B).
AURKA colocalizes with drug-resistant marker ABCG2 and phospho-NUMB in asymmetrically dividing breast cancer cells:
In order to ensure the role of AURKA in stem cell functions, we decided to analyse one of the specific stem cell characteristics-the asymmetric cell division. In asymmetrically dividing BCSCs (Figure 3C), AURKA was found to be colocalized with multi-drug resistance marker ABCG2 in one of the daughter cells indicating the mentioned daughter cell retains the BCSC properties while the other might differentiate into non-stem cancer cell (NSCC). Accordingly, we questioned whether AURKA-ABCG2 colocalization is associated with NUMB [25], which is reportedly phosphorylated by AURKA for eventual destabilization of p53 during self-renewal. Indeed, AURKA was found to be colocalized with phospho-NUMB as well, in indirect immunofluorecence study (Figure 4A) and their physical interaction was confirmed in a patient sample (Figure 4B) through co-immunoprecipitation not in tumour, but in the adjacent normal counterpart. Importantly, AURKA and phospho-NUMB colocalized not only during asymmetric cell divisions, but symmetric divisions as well in the cell lines.
Oct4/Sox2 is associated with proliferation-independent AURKA functions during epithelial to mesenchymal transition:
AURKA being regulated by Oct4/Sox2, targeting Oct4/Sox2 seems to be an attractive way to inhibit stem cell division, also due to confirmation of its involvement in asymmetric cell division. However, silencing Oct4/Sox2 only impacted G0/G1 (Figure 5A) indicating that the same may not be involved in regulating AURKA-mediated cell proliferation, but possibly cell fate. Interestingly, AURKA positive cells in the breast cancer tissues, were positive for SLUG, TWIST, N-cadherin and Vimentin; among which, Vimentin demonstrated extremely high MFI in the AURKA-positive tumour cells (Figure 5B). When patient peripheral blood was analysed, both Oct4 (Figure 5C) and Sox2 (Figure 5D) were found to be positively correlated with Vimentin either through MFI (Oct4) or through percent positive cells (Sox2) suggesting Oct4/Sox2 regulated AURKA functions are associated with epithelial to mesenchymal transition.
Vimentin in addition to AURKA, has to be included as a marker for tracking or targeting drug-resistant Oct4/Sox2 positive BCSCs:
Both low ABCG2 and high ABCG2 expressing breast cancer cells in tumour (Figure 5E) and patient blood (Figure 5F) were found to be Vimentin positive in contrast to AURKA for every CD44/CD24 profile, indicating Vimentin may prove to be a very important marker to identify drug-resistant BCSCs which may or may not express AURKA. When peripheral blood was screened, Vimentin was found to be co expressed in both Oct4 and Sox2 positive cells in Ductal Carcinoma in situ (Figure 6A), Invasive Ductal Carcinoma (Figure 6B) and healthy donors (Figure 6C), while AURKA was found to be induced only in the follow-up blood of the IDC patient post surgery, chemotherapy and radiation. Compared to the healthy donors, Vimentin expression was found to be much higher in DCIS. While total percentages of Oct4/Sox2/AURKA expressing cells were positively correlated in patient blood (Figure 3B), comparison of MFIs indicated this correlation of Oct4/Sox2 with AURKA was not applicable inside the same cell (Figure 6D). When tissue and peripheral blood from a single patient were analysed, in the tissue, most of the Vimentin positive cells were under CD44+CD24+ profile and ABCG2 positive; while the AURKA positive cells had diverse profiles with or without ABCG2 expression and increased CD44+CD24- cells in tumour (Figure 7A) as reported earlier. The peripheral blood of as well demonstrated similar profile of ABCG2 positive cells (Figure 7B). On the other hand, low-ABCG2 and high-ABCG2 expressing cells had different CD44/CD24 profile and AURKA/Vimentin profile (Figure 8) with low-ABCG2 cells demonstrating diverse CD44/CD24/AURKA/Vimentin profiles and high ABCG2 being primarily Vimentin+CD24+.
Discussion:
AURKA being over-expressed in refractory T-cell lymphoma, failure of Alisertib to improve prognosis was quite startling. However, although in a different model system, AURKA, is also overexpressed in breast cancer and therefore expected to respond to Alisertib. As evident from our findings, the drug resistance marker ABCG2 which have been recently reported to remain uninhibited by Alisertib [26], is expressed not only by the AURKA-positive BCSC-like cells but also by the Vimentin-positive cells irrespective of AURKA expression. In the patient samples, the ABCG2+ve cells were found to be sustaining low or high MFI of the mentioned drug exporter pump. The cells with high MFI mostly had CD44+CD24+ or CD44-CD24+ profile and the ones with low MFI had diverse profiles including the classical CD44+CD24-profile for stem cells as mentioned before. However, 100% of the Vimentin+ve, but not AURKA+ve population was found to be ABCG2+ve implying Vimentin expression as a better predictor of ABCG2-mediated drug resistance compared to AURKA and also being a surface marker, easily targetable or sortable for ex vivo culture, in case intervention is to be planned. Also, the low ABCG2 cells demonstrated more AURKA positivity compared to Vimentin in contrast to the high ABCG2 cells which demonstrated mostly Vimentin+AURKA- profile thereby implying AURKA targeting will be including the ABCG2-low dividing population with diverse CD44/CD24 profiles while Vimentin targeting will be including the non-dividing, high ABCG2 cells undergoing EMT.
Notably, AURKA-expressing cells are apparently under active cell-cycle i.e they are destined to divide. Apart from this dividing population, cancer tissues in general, retain a dormant population of BCSCs which are not under active cell-cycle, better explained as in the G0 phase. BCSCs in this dormant state maintain only the minimal survival mechanisms and demonstrate a phenotype unrecognizable to the host immune system. Due to this survival strategy, these cells remain radio resistant for lack of hypoxic environment [27], chemoresistant due to lack of division [28] and untraceable due to plasticity [29] and changing surface marker profile [30]. The above mentioned Vimentin positive population also includes Oct4/Sox2 positive cells with or without AURKA expression i.e the dividing and non-dividing population of potential BCSCs. Again in AURKA-expressing cells, AURKA may be carrying out the kinase-dependent and independent functions [31], which can be identified as cells under S-M and G1 phases responding to catalytic and allosteric inhibitors of AURKA respectively. Under Oct4/Sox2 silencing, AURKA was found to be downregulated along with significant reduction in cells under G0/G1; supporting kinase-independent functions of AURKA to be affected under Oct4/Sox2 targeting and G1 commitment of cells facing fluctuations in Oct4/Sox2 levels [32]. Importantly, the study unravelling this G1 commitment also indicated the cell to be differentiation biased. Our immunofluorescence data revealed colocalization of AURKA with phospho-NUMB in dividing cells supporting the manipulative role [33] of Oct4/Sox2 in differentiating progeny through AURKA induction which can be targeted by the AURKA or NUMB inhibitors, if not by the chemotherapeutic agents. The chemotherapeutic regimen on the other hand will be targeting the AURKA+ve progenies under active cell cycle phases i.e cells under G1 (cyclophosphamides), S (antimetabolites), G2 (alkylating agents) and M (taxanes and vinca alkaloids) upto mitotic exit. However, the AURKA-ve progeny, for example the Oct4/Sox2+ve cells in patient blood will be capable of adapting therapy escaping mechanisms for surviving under the mentioned treatments for example through the ABCG2 expression. The present findings are encouraging enough to propose Vimentin as a notable target, given the fact that Vimentin is expressed in the ABCG2 expressing cells irrespective of AURKA expression. Vimentin expression has already been reported to be induced in neural stem cells exiting G0 [34]; suggesting that Oct4/Sox2+ve cells surrounding the tumour are undergoing EMT and as represented by Vimentin, may be exiting quiescence to enter G1. Additionally, targeting ABCG2 may exhibit toxicity owing to the expression of the same in the epithelium of small intestine, colon, liver canalicular cells, ducts and lobules of mammary tissue [35]. Vimentin targeting reduces the toxicity significantly by selecting only the dividing messenchymal cells [36]. Vimentin-targeting compounds like Withaferin A [37], Simvastatin [38], Silibinin [39], Resveratrol [40], Fluvastatin [41], FiVe1 [42] etc. are under investigation and therefore can be proposed to be combined with AURKA inhibitors for effective remission.
AURKA-NUMB interaction in adjacent normal tissue was another interesting observation of our study; justifying deprivation of the tumour-suppressor activities of both NUMB and p53 within the cells surrounding the tumours as a result of AURKA functions. Interestingly, the antibodies used in our CoIP represented phosphorylated AURKA at threonine-288 i.e the active conformation for kinase activity and NUMB phosphorylated at serine-276, which is responsible for mobilization of the protein [43]. Strong interaction between the mentioned proteins furthermore aligns with previous observations regarding involvement of G-protein coupled receptor (GPCR)/Protein Kinase C (PKC) in the adjacent normal tissues to mediate NUMB phosphorylation by AURKA [44]. Moreover, cancer stem cell markers have been shown to be enriched in the adjacent normal tissues in triple negative breast cancer [45] and the mentioned cells have been reported to possess DNA repair capability thereby withstanding chemotherapy and radiation. Although in a different tissue model, in colon cancer, tumor adjacent ‘normal’ tissues have been compared with healthy tissues which indicated similarity of both the populations in terms of morphology, clonogenic abilities and differentiation except the tumour adjacent normal tissue being changed at molecular level [46] resulting in EMT enrichment. Supporting the mentioned report, in our TNBC sample with enhanced AURKA-NUMB interaction in the adjacent normal part, Vimentin expression was induced more in Oct4 and Sox2+ve cells (54.5% and 90.8% respectively) compared to tumour counterpart (33.7% and 87.7% respectively). However, the adjacent normal Oct4+ve and Sox2+ve cells were Vimentin-low (MFI: 951 and 1694 respectively) compared to the tumour counterpart (MFI:15027 and 3195 respectively) suggesting quantitative expression of EMT markers should not be considered while identifying the cancer stem cells. Therefore, before deciding surgical margins, multiple specimens need to be collected from the tumour periphery during biopsy and molecular analyses for drug resistance markers, EMT and stemness markers need to be performed for comparing the same with healthy counterpart keeping in mind the rationality of complete remission after surgery.
Our brief report connects some missing links between breast cancer stem cell research and applicable therapeutic regimen which not only identify the targetable population under the control of the ‘undruggable’ stemness factors, but also suggest potential combinatorial therapies for metastatic breast cancer. Also, tracking the mentioned quiescent population in patient peripheral blood suggests a cost-effective approach for diagnosis of metastatic breast cancer and potential risk assessment for relapse. It is worth mentioning that there are instances of Oct4+Vimentin+ cells being positively correlated with tumour grade, lymph node metastasis and infiltration of surrounding tissues and vasculogenic mimicry in gall bladder adenocarcinoma [47]. On the other hand, isoforms of Oct4 have been identified [48] in bone-marrow derived messenchymal stem cells (BM MSC) which are recruited to tumour microenvironment for effective immune suppression as reported in colorectal cancer [49]. Importantly, BM MSCs may also be mobilized to blood [50] post administration of GM-CSF in cancer patients which is used as adjuvant post chemotherapy or in combination with vaccines for anti-tumorigenic immune stimulation [51]; thereby pointing out the necessity to analyse the complete profile of the Oct4 positive cells from patient blood before isolation and ex vivo coculture for the purpose of studying possible of pro-tumourigenic activities. In any case, Oct4+Vimentin+ or Sox2+Vimentin+ cells are not expected to be ABCG2+ irrespective of CD44/CD24 profile, thereby confirming the mentioned Oct4+Vimentin+ABCG2+ or Sox2+Vimentin+ABCG2+ cells to be traceable CSCs in patient peripheral blood. On the other hand, allosteric inhibition of AURKA is needed, to include the non-dividing chemotherapy resistant Oct4+/Sox2+ cells which are yet to differentiate and therefore not expressing Vimentin. Another area of further study involves possible extracellular transport of Sox2 which happen to participate in AURKA transcriptional upregulation with Oct4 without DNA binding. Extracellular transportation of Oct4 [52] has been already documented, however there is till-date, no such data regarding Sox2. The mentioned studies will be able to explain the underlying reason for obtaining positive correlation of Oct4/Sox2 expressing cells with AURKA-expressing cells without having similar correlation in MFI. Lastly, we were unable to study the coexpression of Oct4, Sox2 and ABCG2 in the same cells due to lack of primary tagged antibodies. Elaborate analysis is needed to obtain the actual picture of CSC plasticity from tumour initiation to distant metastasis. In this regards, Nanog and ALDH1 expression and ex vivo mammosphere formation of the identified Oct4+/Sox2+Vimentin+ABCG2+ populations are also inevitable.
Materials and Methods:
Gene expression analysis:
For comparing AURKA transcript expression in tumour vs. normal samples from TCGA data, GEPIA2 platform was used. In the BRCA dataset, separated box plot was generated across the histological subtypes with Log2FC cutoff 1, p-value cut off 0.01 and jitter size 0.4.
Survival analysis:
For calculation of relapse-free survival (RFS) in AURKA-high or AURKA-low breast cancer patients, Kaplan Meier Plotter for breast cancer has been used [53]. The dataset selected, was 208079_s_at; patients were split using auto select best cutoff and JetSet best probe set was used as option.
Promoter analysis in silico:
DNA sequence upto 2kb upstream of AURKA (Gene ID: 6790) TSS was obtained from NCBI and used for analysis of transcription factor binding sites in ALGGEN-PROMO.
Correlation analyses:
Correlation of Oct4/Sox2 transcript expression with AURKA transcript expression were performed in GEPIA2 using Pearson Correlation coefficient using non-log scale for calculation and log scale axis for visualization. TCGA tumour as well as normal dataset was used for Oct4/AURKA and TCGA tumour and GTeX normal data sets were used for Sox2/AURKA. 1085 invasive breast carcinoma samples and 112 paired breast samples from TCGA database were considered. In case where the number of corresponding normal samples was not statistically fit to compare with the tumour samples GTeX normal sample data was used, which included 179 normal samples.
Cell lines and transfections:
Autheniticated breast cancer cell lines MCF-7 and MDAMB 231 were procured from NCCS, Pune, maintained in MEM (HIMEDIA) supplemented with 10% FBS (Gibco) with antibiotics Penicillin, Streptomycin and Gentamycin in a humidified CO2 incubator at 37 0C for maximum 20 passages. Prior to transfection, serum containing media was replaced with Opti-MEM (Gibco) without antibiotics and conditioned for 24 hrs. pLKO.1 Sox2 3H b was a gift from Matthew Meyerson (Addgene plasmid # 26352; http://n2t.net/addgene:26352; RRID:Addgene_26352); LL - hOCT4i -2 was a gift from George Daley (Addgene plasmid # 12197; http://n2t.net/addgene:12197; RRID:Addgene_12197); pLKO.1 puro was a gift from Bob Weinberg (Addgene plasmid # 8453; http://n2t.net/addgene:8453; RRID:Addgene_8453). Plasmid DNA and Lipofectamine Stem Transfection Reagent (Invitrogen); were diluted in Opti-MEM and added to the cells. Transfected cells were maintained in culture for at least 72 hrs and eventually the efficiency of transfection was assessed by flow-cytometry.
Flow cytometric analyses:
Breast cancer tissue samples were minced and treated with type IV collagenase until single-cell suspension was obtained. In case of blood, CTCs were isolated using Histopaque-1077 [54] and following manufacturer’s (Sigma) procedure. Cell lines were trypsinized to obtain single cells. Eventually, the cells were fixed and permeabilized in chilled acetone for 20min, washed in PBS, blocked using 1% FBS, treated with antibodies and analysed in BD FACS Verse flow cytometer.
Cell Cycle:
Cells were fixed in acetone [55], washed and treated with RNAse at 37 0C for 1hr. Next, the cells were incubated in propidium iodide (Sigma-Aldrich) for 15 minutes, washed and flow cytometric analyses were performed.
Chromatin Immunoprecipitation:
Cells were fixed using 1% formaldehyde (HIMEDIA) to the media. Excess formaldehyde was quenched with glycine and the media was aspirated off. Cells were subsequently lysed in lysis buffer containing protease inhibitor cocktail and sonication was performed. Lysates were then precleared using Protein A/G plus Agarose beads (SANTA CRUZ BIOTECHNOLOGY).Next, a portion was stored in -20°C freezer as input and to the rest, antibodies were added and kept in a rotator overnight at 4°C. Next day, the DNA-protein-antibody complexes were pulled down with beads, washed in low salt, high salt and LiCl buffers and eluted. Reverse crosslinking was performed at 650C overnight using 5M NaCl and subsequent proteinase K (SRL) treatment was done. Finally, DNA was purified by phenol chloroform isoamyl alcohol extraction, resuspended in nuclease free water (Millipore Sigma) and proceeded for PCR. Sequences of the ChIP primers have been included in Supplementary Data 3.
Indirect Immunofluorescence:
Cells were grown on sterilized and poly L-lysine coated cover slips in cell culture plates. On the day of processing, the cover slips were washed in chilled PBS and fixed in methanol: acetone (1:1) for 30 min in -20°C. Next the cover slips were washed in PBS three times and blocked with 1% FBS for 1hr at room temperature. After blocking, the cells were incubated overnight in antibody-containing PBS as per the antibody manufacturer’s recommended dilution and again, washed in PBS. Finally, the cells were stained in DAPI, coverslips were mounted in DPX and assessed in fluorescence microscope (Leica) under 20X magnification.
Co-immunoprecipitation:
Protease inhibitors and antibodies (2 μg/100-200μg of protein) were added to tissue lysates and incubated at 40C overnight. The IP suspensions were then transferred to a column containing filter coated with Protein-A Sepharose beads and left at 40C overnight. The next day, bead-antibody-protein congugates were washed with 1X and 0.1X IP Buffer. The purified protein complexes were then eluted with IP Elution Buffer and qualitatively analysed for expression of the interacting protein candidates by western blotting.
Statistics
All the statistical analyses were performed using the Graphpad-Prism software. All the data have been acquired in replicates-as mentioned in the legends, placed in the Prism for graphical representation and statistical analysis was performed in for each graph on a case-to-case basis using a statistical method recommended by the software itself. The specific tests for calculating P-values/R-values have have been mentioned in the concerned figure legends.
Author Contributions
DS: designed the study, lead the team in performing experiments and acquiring data, analysed the data and wrote the manuscript; DT: performed the experiments; EM and SD: participated in performing some of the experiments; AR and SS: acquisition of surgical tissues and blood; SM: collaborative designing of the study, senior advisory and supervision, participation in preparing the final draft of the manuscript.
Study approval
Human Ethical Committee approval for the study was obtained prior to patient data acquisition from the institute and the study was performed following Uniform Requirements for Manuscripts Submitted to Biomedical Journals, issued by the International Committee for Medical Journal Editors (ICMJE). Written informed consents were obtained from all the patients prior to participation. Patient details have been included in Supplementary Data 3.
Acknowledgement
We are grateful to the Director, Chittaranjan National Cancer Institute (CNCI) and Dr. Madhumita Roy, former HOD, Department of Environmental Carcinogenesis and Toxicology, CNCI for the infrastructural support provided for pursuing this study, Mr. Archishmaan Ghosh and Ms. Shalini Upadhyay for helping in data acquisitions. We would also like to thank the Council of Scientific and Industrial Research (CSIR), Govt. of India for the fellowships of Dr. Debomita Sengupta and Ms. Salini Das and their contingencies; and the Ministry of Health and Family Welfare, Govt. of India for Institutional Funding.
Declaration
The authors have declared that no conflict of interest exists.
References
- Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017, 67, 7–30.
- Colleoni M, Sun Z, Price KN, Karlsson P, Forbes JF, Thürlimann B, et al. Annual Hazard Rates of Recurrence for Breast Cancer During 24 Years of Follow-Up: Results From the International Breast Cancer Study Group Trials I to V. J Clin Oncol [Internet]. 2016, 34, 927–35. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4933127/ (accessed on 9 January 2023).
- Roser M, Ortiz-Ospina E, Ritchie H. Life Expectancy Our World in Data [Internet]. Available online: https://ourworldindata.org/life-expectancy (accessed on 10 January 2023).
- Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature. 1994, 367, 645–8.
- Zhao W, Li Y, Zhang X. Stemness-Related Markers in Cancer. Cancer Transl Med [Internet]. 2017, 3, 87–95. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5737740/ (accessed on 10 January 2023).
- Walcher L, Kistenmacher A-K, Suo H, Kitte R, Dluczek S, Strauß A, et al. Cancer Stem Cells—Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Frontiers in Immunology [Internet]. 2020, 11, 1280. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2020.01280 (accessed on 14 December 2021).
- Qin S, Jiang J, Lu Y, Nice EC, Huang C, Zhang J, et al. Emerging role of tumor cell plasticity in modifying therapeutic response. Sig Transduct Target Ther [Internet]. Nature Publishing Group; 2020, 5, 1–36. Available online: https://www.nature.com/articles/s41392-020-00313-5 (accessed on 10 January 2023).
- Zhou H-M, Zhang J-G, Zhang X, Li Q. Targeting cancer stem cells for reversing therapy resistance: mechanism, signaling, and prospective agents. Sig Transduct Target Ther [Internet]. Nature Publishing Group; 2021, 6, 1–17. Available online: https://www.nature.com/articles/s41392-020-00430-1 (accessed on 10 January 2023).
- Arnold CR, Mangesius J, Skvortsova I-I, Ganswindt U. The Role of Cancer Stem Cells in Radiation Resistance. Front Oncol [Internet]. 2020, 10, 164. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7044409/ (accessed on 10 January 2023).
- Zheng Q, Zhang M, Zhou F, Zhang L, Meng X. The Breast Cancer Stem Cells Traits and Drug Resistance. Front Pharmacol [Internet]. 2021, 11, 599965. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7876385/ (accessed on 1 March 2023).
- Mavingire N, Campbell P, Wooten J, Aja J, Davis MB, Loaiza-Perez A, et al. Cancer stem cells: Culprits in endocrine resistance and racial disparities in breast cancer outcomes. Cancer Lett. 2021, 500, 64–74.
- Staff S, Isola J, Jumppanen M, Tanner M. Aurora-A gene is frequently amplified in basal-like breast cancer. Oncol Rep. 2010, 23, 307–12.
- den Hollander J, Rimpi S, Doherty JR, Rudelius M, Buck A, Hoellein A, et al. Aurora kinases A and B are up-regulated by Myc and are essential for maintenance of the malignant state. Blood [Internet]. 2010, 116, 1498–505. [CrossRef]
- Tanaka M, Ueda A, Kanamori H, Ideguchi H, Yang J, Kitajima S, et al. Cell-cycle-dependent regulation of human aurora A transcription is mediated by periodic repression of E4TF1. J Biol Chem. 2002, 277, 10719–26.
- Wu C-C, Yang T-Y, Yu C-TR, Phan L, Ivan C, Sood AK, et al. p53 negatively regulates Aurora A via both transcriptional and posttranslational regulation. Cell Cycle [Internet]. 2012, 11, 3433–42. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3466554/ (accessed on 2 March 2023).
- Du R, Huang C, Liu K, Li X, Dong Z. Targeting AURKA in Cancer: molecular mechanisms and opportunities for Cancer therapy. Molecular Cancer [Internet]. 2021, 20, 15. [CrossRef]
- Nikonova AS, Astsaturov I, Serebriiskii IG, Dunbrack RL, Golemis EA. Aurora A kinase (AURKA) in normal and pathological cell division. Cell Mol Life Sci. 2013, 70, 661–87.
- Bertolin G, Tramier M. Insights into the non-mitotic functions of Aurora kinase A: more than just cell division. Cell Mol Life Sci [Internet]. 2020, 77, 1031–47. [CrossRef]
- Yang N, Wang C, Wang Z, Zona S, Lin S-X, Wang X, et al. FOXM1 recruits nuclear Aurora kinase A to participate in a positive feedback loop essential for the self-renewal of breast cancer stem cells. Oncogene [Internet]. 2017, 36, 3428–40. Available online: https://www.nature.com/articles/onc2016490 (accessed on 10 December 2021).
- Tanaka T, Kimura M, Matsunaga K, Fukada D, Mori H, Okano Y. Centrosomal kinase AIK1 is overexpressed in invasive ductal carcinoma of the breast. Cancer Res. 1999, 59, 2041–4.
- Ali HR, Dawson S-J, Blows FM, Provenzano E, Pharoah PD, Caldas C. Aurora kinase A outperforms Ki67 as a prognostic marker in ER-positive breast cancer. Br J Cancer [Internet]. Nature Publishing Group; 2012, 106, 1798–806. Available online: https://www.nature.com/articles/bjc2012167 (accessed on 5 April 2023).
- O’Connor OA, Özcan M, Jacobsen ED, Roncero JM, Trotman J, Demeter J, et al. Randomized Phase III Study of Alisertib or Investigator’s Choice (Selected Single Agent) in Patients With Relapsed or Refractory Peripheral T-Cell Lymphoma. J Clin Oncol. 2019, 37, 613–23.
- Hu J, Qin K, Zhang Y, Gong J, Li N, Lv D, et al. Downregulation of transcription factor Oct4 induces an epithelial-to-mesenchymal transition via enhancement of Ca2+ influx in breast cancer cells. Biochemical and Biophysical Research Communications [Internet]. 2011, 411, 786–91. Available online: https://www.sciencedirect.com/science/article/pii/S0006291X11012447 (accessed on 2 March 2023).
- Wu F, Zhang J, Wang P, Ye X, Jung K, Bone KM, et al. Identification of two novel phenotypically distinct breast cancer cell subsets based on Sox2 transcription activity. Cell Signal. 2012, 24, 1989–98.
- Siddique HR, Feldman DE, Chen C-L, Punj V, Tokumitsu H, Machida K. NUMB phosphorylation destabilizes p53 and promotes self-renewal of tumor-initiating cells by a NANOG-dependent mechanism in liver cancer. Hepatology. 2015, 62, 1466–79.
- Vagiannis D, Zhang Y, Budagaga Y, Novotna E, Skarka A, Kammerer S, et al. Alisertib shows negligible potential for perpetrating pharmacokinetic drug-drug interactions on ABCB1, ABCG2 and cytochromes P450, but acts as dual-activity resistance modulator through the inhibition of ABCC1 transporter. Toxicol Appl Pharmacol. 2022, 434, 115823.
- Arnold CR, Mangesius J, Skvortsova I-I, Ganswindt U. The Role of Cancer Stem Cells in Radiation Resistance. Front Oncol [Internet]. 2020, 10, 164. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7044409/ (accessed on 30 September 2023).
- Brown JA, Schober M. Cellular quiescence: How TGFβ protects cancer cells from chemotherapy. Mol Cell Oncol [Internet]. 2018, 5, e1413495. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5821413/ (accessed on 30 September 2023).
- Chen W, Dong J, Haiech J, Kilhoffer M-C, Zeniou M. Cancer Stem Cell Quiescence and Plasticity as Major Challenges in Cancer Therapy. Stem Cells Int [Internet]. 2016, 2016, 1740936. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4932171/ (accessed on 30 September 2023).
- Walcher L, Kistenmacher A-K, Suo H, Kitte R, Dluczek S, Strauß A, et al. Cancer Stem Cells—Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front Immunol [Internet]. 2020, 11, 1280. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7426526/ (accessed on 30 September 2023).
- Guarino Almeida E, Renaudin X, Venkitaraman AR. A kinase-independent function for AURORA-A in replisome assembly during DNA replication initiation. Nucleic Acids Res [Internet]. 2020, 48, 7844–55. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7430631/ (accessed on 1 May 2023).
- Strebinger D, Deluz C, Friman ET, Govindan S, Alber AB, Suter DM. Endogenous fluctuations of OCT4 and SOX2 bias pluripotent cell fate decisions. Mol Syst Biol [Internet]. 2019, 15, e9002. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6759502/ (accessed on 1 May 2023).
- Gómez-López S, Lerner RG, Petritsch C. Asymmetric cell division of stem and progenitor cells during homeostasis and cancer. Cell Mol Life Sci [Internet]. 2014, 71, 575–97. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3901929/ (accessed on 10 April 2023).
- Morrow CS, Porter TJ, Xu N, Arndt ZP, Ako-Asare K, Heo HJ, et al. Vimentin Coordinates Protein Turnover at the Aggresome during Neural Stem Cell Quiescence Exit. Cell Stem Cell [Internet]. 2020, 26, 558–568e9. Available online: https://linkinghub.elsevier.com/retrieve/pii/S1934590920300187 (accessed on 1 May 2023).
- Mo W, Zhang J-T. Human ABCG2: structure, function, and its role in multidrug resistance. Int J Biochem Mol Biol [Internet]. 2011, 3, 1–27. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3325772/ (accessed on 9 September 2023).
- Strouhalova K, Přechová M, Gandalovičová A, Brábek J, Gregor M, Rosel D. Vimentin Intermediate Filaments as Potential Target for Cancer Treatment. Cancers (Basel) [Internet]. 2020, 12, 184. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7017239/ (accessed on 9 September 2023).
- Bargagna-Mohan P, Hamza A, Kim Y, Khuan Abby Ho Y, Mor-Vaknin N, Wendschlag N, et al. The tumor inhibitor and antiangiogenic agent withaferin A targets the intermediate filament protein vimentin. Chem Biol. 2007, 14, 623–34.
- Bai F, Yu Z, Gao X, Gong J, Fan L, Liu F. Simvastatin induces breast cancer cell death through oxidative stress up-regulating miR-140-5p. Aging (Albany NY) [Internet]. 2019, 11, 3198–219. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6555469/ (accessed on 3 October 2023).
- Bosch-Barrera J, Corominas-Faja B, Cuyàs E, Martin-Castillo B, Brunet J, Menendez JA. Silibinin Administration Improves Hepatic Failure Due to Extensive Liver Infiltration in a Breast Cancer Patient. Anticancer Research [Internet]. International Institute of Anticancer Research; 2014, 34, 4323–7. Available online: https://ar.iiarjournals.org/content/34/8/4323 (accessed on 3 October 2023).
- Guo K, Feng Y, Zheng X, Sun L, Wasan HS, Ruan S, et al. Resveratrol and Its Analogs: Potent Agents to Reverse Epithelial-to-Mesenchymal Transition in Tumors Frontiers in Oncology [Internet]. 2021, 11. Available online: https://www.frontiersin.org/articles/10.3389/fonc.2021.644134 (accessed on 3 October 2023).
- Dorsch M, Kowalczyk M, Planque M, Heilmann G, Urban S, Dujardin P, et al. Statins affect cancer cell plasticity with distinct consequences for tumor progression and metastasis. Cell Reports [Internet]. 2021, 37, 110056. Available online: https://linkinghub.elsevier.com/retrieve/pii/S2211124721015424 (accessed on 3 October 2023).
- Bollong MJ, Pietilä M, Pearson AD, Sarkar TR, Ahmad I, Soundararajan R, et al. A vimentin binding small molecule leads to mitotic disruption in mesenchymal cancers. Proceedings of the National Academy of Sciences [Internet]. Proceedings of the National Academy of Sciences; 2017, 114, E9903–12. Available online: https://www.pnas.org/doi/10.1073/pnas.1716009114 (accessed on 3 October 2023).
- Smith CA, Lau KM, Rahmani Z, Dho SE, Brothers G, She YM, et al. aPKC-mediated phosphorylation regulates asymmetric membrane localization of the cell fate determinant Numb. EMBO J [Internet]. 2007, 26, 468–80. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1783459/ (accessed on 21 September 2023).
- Dho SE, Trejo J, Siderovski DP, McGlade CJ. Dynamic regulation of mammalian numb by G protein-coupled receptors and protein kinase C activation: Structural determinants of numb association with the cortical membrane. Mol Biol Cell. 2006, 17, 4142–55.
- Atkinson RL, Yang WT, Rosen DG, Landis MD, Wong H, Lewis MT, et al. Cancer stem cell markers are enriched in normal tissue adjacent to triple negative breast cancer and inversely correlated with DNA repair deficiency. Breast Cancer Research [Internet]. 2013, 15, R77. [CrossRef]
- Zhao Y, Guo M, Zhao F, Liu Q, Wang X. Colonic stem cells from normal tissues adjacent to tumor drive inflammation and fibrosis in colorectal cancer. Cell Communication and Signaling. 2023, 21, 186. [CrossRef]
- Zhang Y, Xu J, Xu Z, Wang Y, Wu S, Wu L, et al. Expression of vimentin and Oct-4 in gallbladder adenocarcinoma and their relationship with vasculogenic mimicry and their clinical significance. Int J Clin Exp Pathol [Internet]. 2018, 11, 3618–27. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6962874/ (accessed on 9 September 2023).
- Wang X, Dai J. Concise Review: Isoforms of OCT4 Contribute to the Confusing Diversity in Stem Cell Biology. Stem Cells [Internet]. 2010, 28, 885–93. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2962909/ (accessed on 9 September 2023).
- Nishikawa G, Kawada K, Nakagawa J, Toda K, Ogawa R, Inamoto S, et al. Bone marrow-derived mesenchymal stem cells promote colorectal cancer progression via CCR5. Cell Death Dis [Internet]. Nature Publishing Group; 2019, 10, 1–13. Available online: https://www.nature.com/articles/s41419-019-1508-2 (accessed on 9 September 2023).
- Kim J, Kim NK, Park SR, Choi BH. GM-CSF Enhances Mobilization of Bone Marrow Mesenchymal Stem Cells via a CXCR4-Medicated Mechanism. Tissue Eng Regen Med [Internet]. 2018, 16, 59–68. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6361095/ (accessed on 9 September 2023).
- Kumar A, Taghi Khani A, Sanchez Ortiz A, Swaminathan S. GM-CSF: A Double-Edged Sword in Cancer Immunotherapy. Front Immunol [Internet]. 2022, 13, 901277. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9294178/ (accessed on 9 September 2023).
- Rolf HJ, Niebert S, Niebert M, Gaus L, Schliephake H, Wiese KG. Intercellular Transport of Oct4 in Mammalian Cells: A Basic Principle to Expand a Stem Cell Niche? PLoS One [Internet]. 2012, 7, e32287. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3281129/ (accessed on 1 May 2023).
- Győrffy, B. Survival analysis across the entire transcriptome identifies biomarkers with the highest prognostic power in breast cancer Computational and Structural Biotechnology Journal [Internet]. Elsevier; 2021, 19, 4101–9. Available online: https://www.csbj.org/article/S2001-0370(21)00304-4/fulltext (accessed on 19 September 2023). [CrossRef] [PubMed]
- He W, Kularatne SA, Kalli KR, Prendergast FG, Amato RJ, Klee GG, et al. Quantitation of circulating tumor cells in blood samples from ovarian and prostate cancer patients using tumor-specific fluorescent ligands. Int J Cancer [Internet]. 2008, 123, 1968–73. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2778289/ (accessed on 24 May 2023).
- Carbonari M, Mancaniello D, Tedesco T, Fiorilli M. Flow acetone-staining technique: A highly efficient procedure for the simultaneous analysis of DNA content, cell morphology, and immunophenotype by flow cytometry Cytometry Part A [Internet]. 2008, 73A, 168–74. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/cyto.a.20521 (accessed on 24 May 2023).
Figure 1.
AURKA expression and relapse cases in breast cancer are associated with stemness factors: (A) Box-whisker plots demonstrating overexpression of AURKA transcripts in tumour samples compared to adjacent normal samples as per TCGA database (B) KM-plotter analysis denoting more relapse cases are associated with high-AURKA samples (C) Diagrammatic representation demonstrating stemness factor binding sites within 2 kb upstream of AURKA transcription start site (TSS). Accordingly, Oct4 and Sox2 binding sites have been marked red and yellow respectively.
Figure 1.
AURKA expression and relapse cases in breast cancer are associated with stemness factors: (A) Box-whisker plots demonstrating overexpression of AURKA transcripts in tumour samples compared to adjacent normal samples as per TCGA database (B) KM-plotter analysis denoting more relapse cases are associated with high-AURKA samples (C) Diagrammatic representation demonstrating stemness factor binding sites within 2 kb upstream of AURKA transcription start site (TSS). Accordingly, Oct4 and Sox2 binding sites have been marked red and yellow respectively.
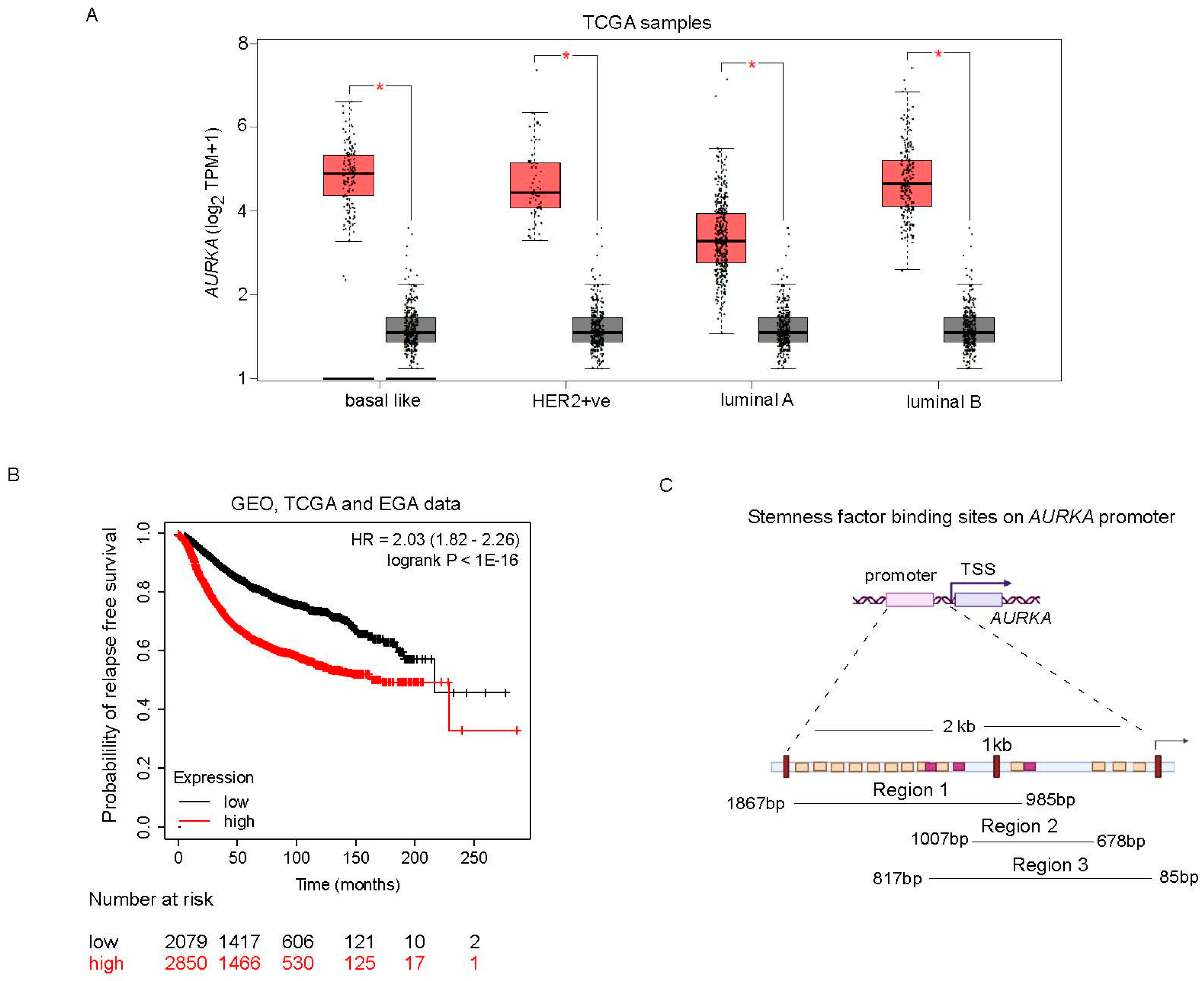
Figure 2.
AURKA transcript and protein expression are positively correlated with Oct4/Sox2 transcript and protein expression: (A) scatter plots demonstrating positive correlation of Oct4/Sox2 transcripts with AURKA transcripts as per TCGA data (B) Interleaved bar diagrams representing Oct4/Sox2 binding efficiency on AURKA promoter compared to IgG control; noteworthy, Oct4 binding to DNA is confirmed. But Sox2 is not. (C-D) Interleaved bar diagrams representing downregulation of AURKA under silencing of Oct4/Sox2 in MCF-7 (C) or MDA MB-231 (D) breast cancer cells by transfection as observed by flow cytometric analysis; left panels represent efficiency of transfection and the right panels represent mean fluorescent intensities (MFIs) of AURKA positive cells under vector control vs. shRNA transfection. The transfections have been performed in triplicates and P-values have been calculated using unpaired t-tests. Importantly, in MDA MB-231 our standard time of processing the shRNA transfected cells i.e 48 hr which worked for the rest of the experiments, could not reduce the percentage of cells expressing Oct4. However, the MFI of Oct4 in the transfected cells was reduced significantly, which was taken into consideration.
Figure 2.
AURKA transcript and protein expression are positively correlated with Oct4/Sox2 transcript and protein expression: (A) scatter plots demonstrating positive correlation of Oct4/Sox2 transcripts with AURKA transcripts as per TCGA data (B) Interleaved bar diagrams representing Oct4/Sox2 binding efficiency on AURKA promoter compared to IgG control; noteworthy, Oct4 binding to DNA is confirmed. But Sox2 is not. (C-D) Interleaved bar diagrams representing downregulation of AURKA under silencing of Oct4/Sox2 in MCF-7 (C) or MDA MB-231 (D) breast cancer cells by transfection as observed by flow cytometric analysis; left panels represent efficiency of transfection and the right panels represent mean fluorescent intensities (MFIs) of AURKA positive cells under vector control vs. shRNA transfection. The transfections have been performed in triplicates and P-values have been calculated using unpaired t-tests. Importantly, in MDA MB-231 our standard time of processing the shRNA transfected cells i.e 48 hr which worked for the rest of the experiments, could not reduce the percentage of cells expressing Oct4. However, the MFI of Oct4 in the transfected cells was reduced significantly, which was taken into consideration.
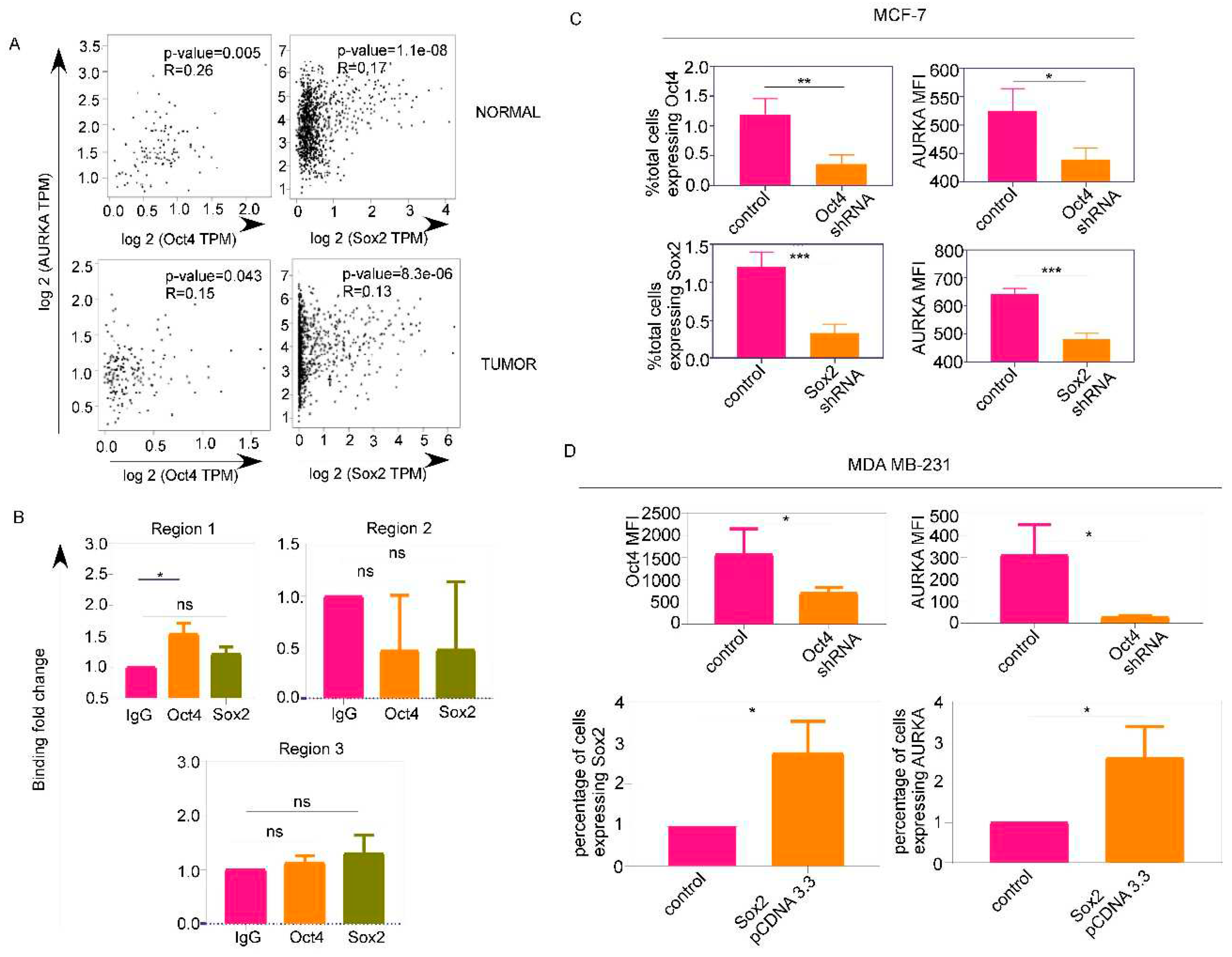
Figure 3.
AURKA expression is positively correlated with Oct4/Sox2 in patient samples and also associated with multi drug resistance marker ABCG2: (A) Rainbow heat map representing Oct4/Sox2 MFI and AURKA MFI in Oct4 [AURKA(O)]/Sox2 [AURKA(S)] cells in adjacent normal samples (AN) and tumour(T) samples as surgically dissected by clinicians (B) Scatter plot representing positive correlation of total AURKA positive cells with that of total Oct4/Sox2 positive cells in patient blood. Significant correlations have been indicated using 2-tailed tests and Pearson Correlation coefficient (C) Indirect immunoflurescence demonstrating colocalization of AURKA with ABCG2 in one of the progenies during asymmetric division, a characteristic feature of stem cells.
Figure 3.
AURKA expression is positively correlated with Oct4/Sox2 in patient samples and also associated with multi drug resistance marker ABCG2: (A) Rainbow heat map representing Oct4/Sox2 MFI and AURKA MFI in Oct4 [AURKA(O)]/Sox2 [AURKA(S)] cells in adjacent normal samples (AN) and tumour(T) samples as surgically dissected by clinicians (B) Scatter plot representing positive correlation of total AURKA positive cells with that of total Oct4/Sox2 positive cells in patient blood. Significant correlations have been indicated using 2-tailed tests and Pearson Correlation coefficient (C) Indirect immunoflurescence demonstrating colocalization of AURKA with ABCG2 in one of the progenies during asymmetric division, a characteristic feature of stem cells.
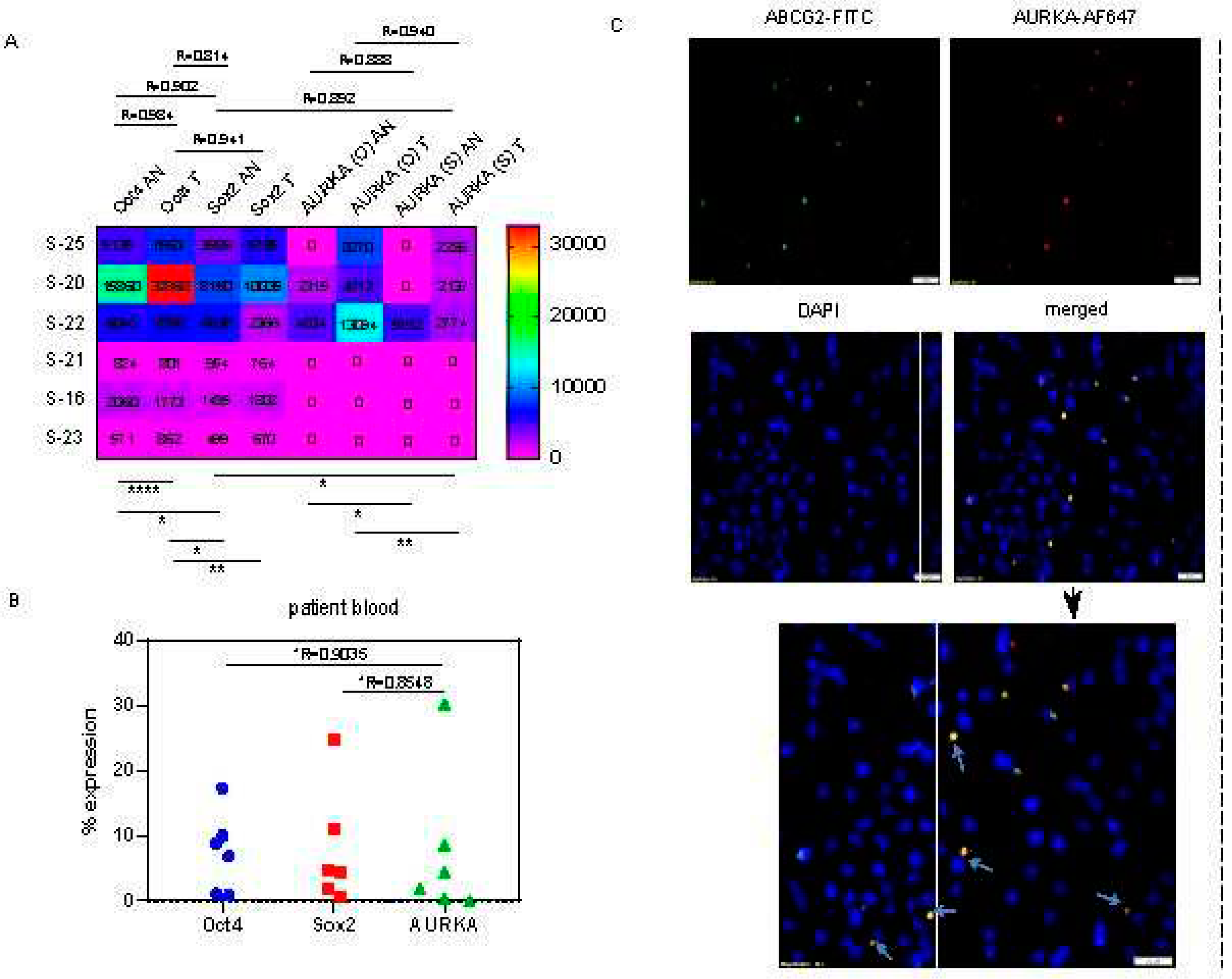
Figure 4.
Enzymatically active AURKA colocalizes with phospho-NUMB (Ser-276) during asymmetric cell division and their physical interaction is observable in the adjacent normal counterpart of TNBC tissue sample: (A) Indirect immunofluorescence demonstrating involvement of AURKA in stem cell division; notably, active phospho-AURKA (Thr-288) localization follows distribution pattern of phospho-NUMB (Ser-276), indicating that NUMB is phosphorylated by AURKA leading to mobilization of the same during decision of cell fate; colocalization is identifiable both in case of symmetric and asymmetric cell division (B) Co-immunoprecipitation demonstrating physical interaction of phospho-AURKA with phospho-NUMB in adjacent normal counterpart of a TNBC tissue sample.
Figure 4.
Enzymatically active AURKA colocalizes with phospho-NUMB (Ser-276) during asymmetric cell division and their physical interaction is observable in the adjacent normal counterpart of TNBC tissue sample: (A) Indirect immunofluorescence demonstrating involvement of AURKA in stem cell division; notably, active phospho-AURKA (Thr-288) localization follows distribution pattern of phospho-NUMB (Ser-276), indicating that NUMB is phosphorylated by AURKA leading to mobilization of the same during decision of cell fate; colocalization is identifiable both in case of symmetric and asymmetric cell division (B) Co-immunoprecipitation demonstrating physical interaction of phospho-AURKA with phospho-NUMB in adjacent normal counterpart of a TNBC tissue sample.
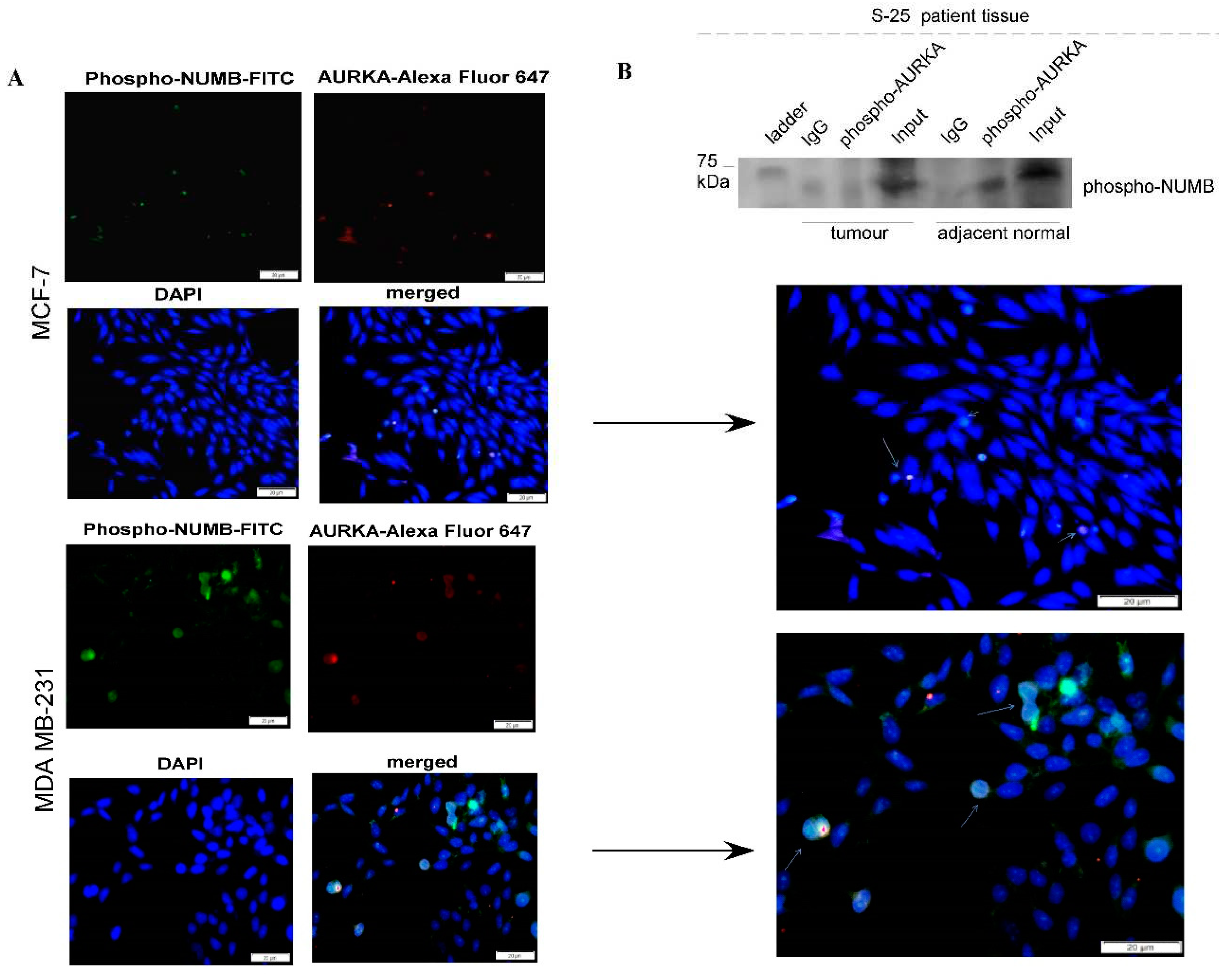
Figure 5.
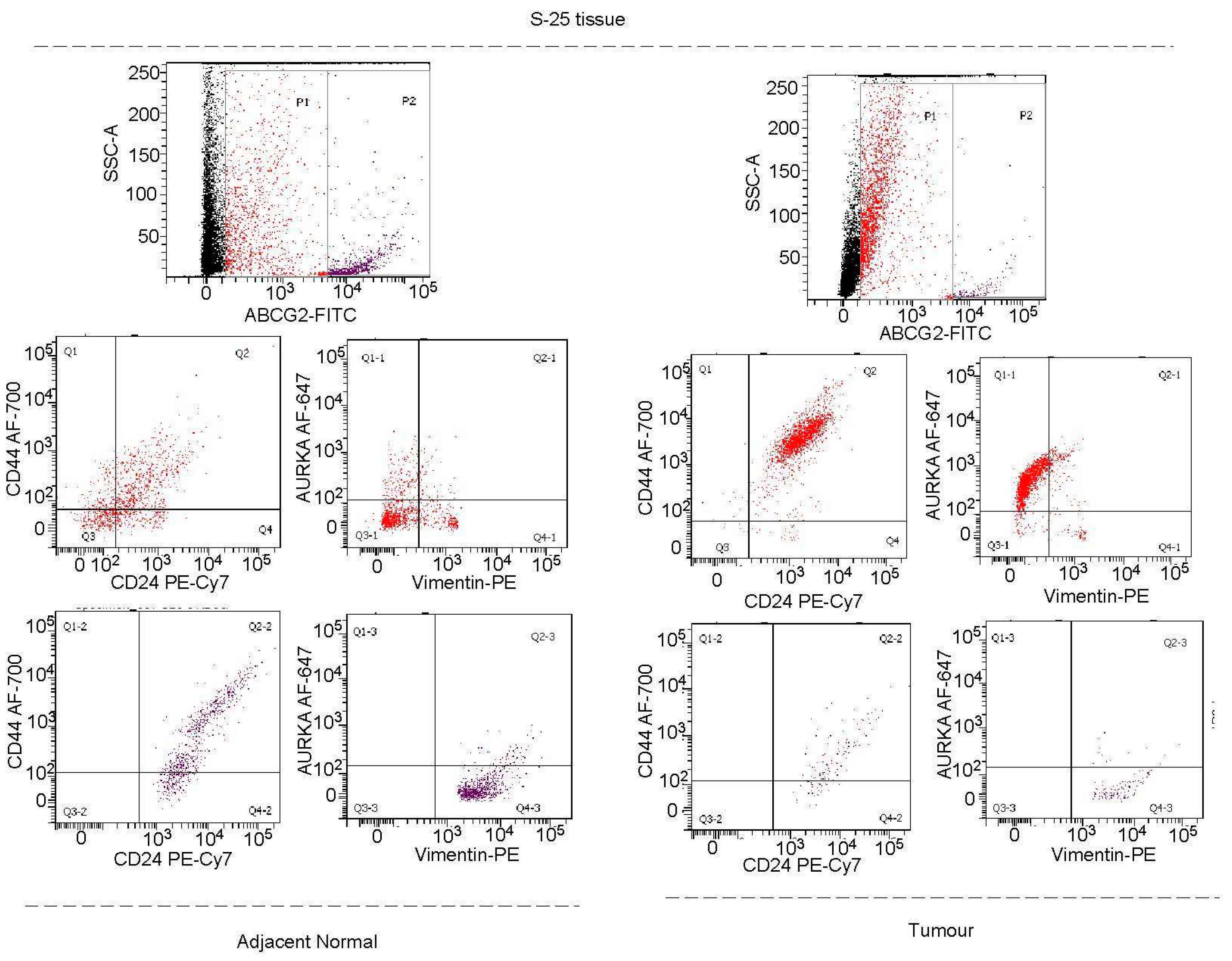
Oct4/Sox2-mediated AURKA regulation is associated with EMT and simultaneously drug resistance: (A) Scatter plots showing decrease in G0/G1 population under Oct4/Sox2 shRNA transfection (Figure 2C) compared to vector control in MCF-7; the cell cycle assay has been performed in duplicates and 2-way ANOVA has been used for calculation of P-values (B) Scatter plots representing MFI of different EMT markers in AURKA +ve cells from patient tissues (C-D) Scatter plots representing positive correlation of Oct4 (C) and Sox2 (D) with Vimentin expression in patient blood either at intracellular (Oct4) or intercellular (Sox2) level (E) Flow cytometric scatter plots demonstrating distribution of ABCG2-low (red) and ABCG2-high (violet) cells in a patient tissue (S-23) under CD44/CD24 profile (F) Flow cytometric scatter plot showing ABCG2+ve cells in the blood of the same patient (S-23) express Vimentin, but not AURKA.
Figure 5.
Oct4/Sox2-mediated AURKA regulation is associated with EMT and simultaneously drug resistance: (A) Scatter plots showing decrease in G0/G1 population under Oct4/Sox2 shRNA transfection (Figure 2C) compared to vector control in MCF-7; the cell cycle assay has been performed in duplicates and 2-way ANOVA has been used for calculation of P-values (B) Scatter plots representing MFI of different EMT markers in AURKA +ve cells from patient tissues (C-D) Scatter plots representing positive correlation of Oct4 (C) and Sox2 (D) with Vimentin expression in patient blood either at intracellular (Oct4) or intercellular (Sox2) level (E) Flow cytometric scatter plots demonstrating distribution of ABCG2-low (red) and ABCG2-high (violet) cells in a patient tissue (S-23) under CD44/CD24 profile (F) Flow cytometric scatter plot showing ABCG2+ve cells in the blood of the same patient (S-23) express Vimentin, but not AURKA.
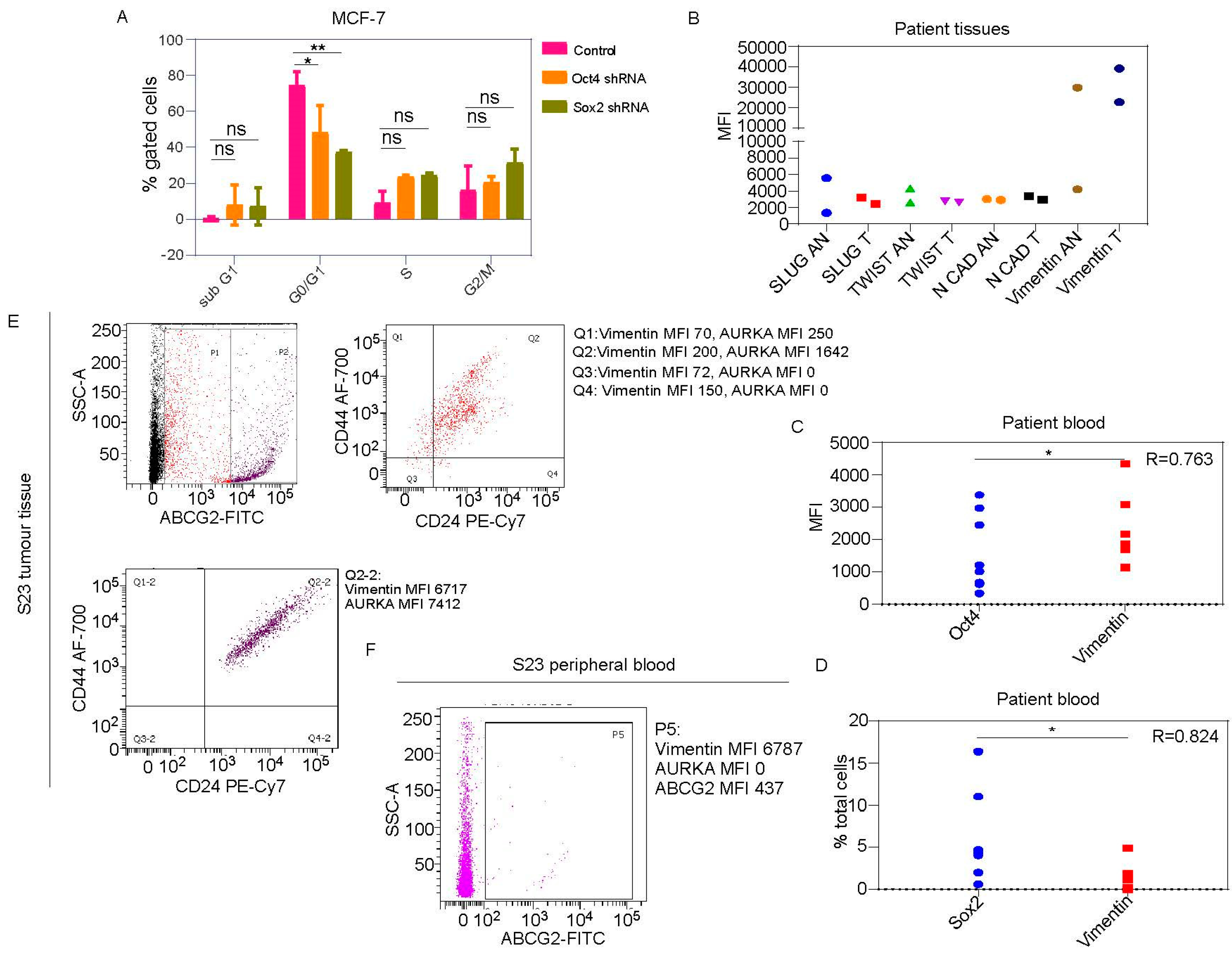
Figure 6.
Vimentin expression, but not AURKA expression can be relied on for tracking Oct4/Sox2+ve cells in healthy donor or cancer patient blood: (A-C) Flow cytometric scatter plots showing Oct4/Sox2 +ve cells in blood of DCIS patient (A), IDC patient (B) and healthy donors (C) positively express Vimentin, but not necessarily AURKA; AURKA in Oct4/Sox2+ve cells can be only detected in the follow-up blood of IDC patient (D) Rainbow heat map image representing status of AURKA expression in Oct4/Sox2 positive cells in patient blood; the MFIs of Oct4/Sox2/AURKA have been included in the respective cells of the heatmap. Notably, Oct4/Sox2+ve cells in patient peripheral blood (n=6) did not express AURKA, even if AURKA MFI in the PBMC was quite high suggesting the mesenchymal-like Oct4/Sox2+ve cells were probably in G0 phase or early G1 (Figure 5A) and might demonstrate chemoresistance or resistance to AURKA inhibitors.
Figure 6.
Vimentin expression, but not AURKA expression can be relied on for tracking Oct4/Sox2+ve cells in healthy donor or cancer patient blood: (A-C) Flow cytometric scatter plots showing Oct4/Sox2 +ve cells in blood of DCIS patient (A), IDC patient (B) and healthy donors (C) positively express Vimentin, but not necessarily AURKA; AURKA in Oct4/Sox2+ve cells can be only detected in the follow-up blood of IDC patient (D) Rainbow heat map image representing status of AURKA expression in Oct4/Sox2 positive cells in patient blood; the MFIs of Oct4/Sox2/AURKA have been included in the respective cells of the heatmap. Notably, Oct4/Sox2+ve cells in patient peripheral blood (n=6) did not express AURKA, even if AURKA MFI in the PBMC was quite high suggesting the mesenchymal-like Oct4/Sox2+ve cells were probably in G0 phase or early G1 (Figure 5A) and might demonstrate chemoresistance or resistance to AURKA inhibitors.
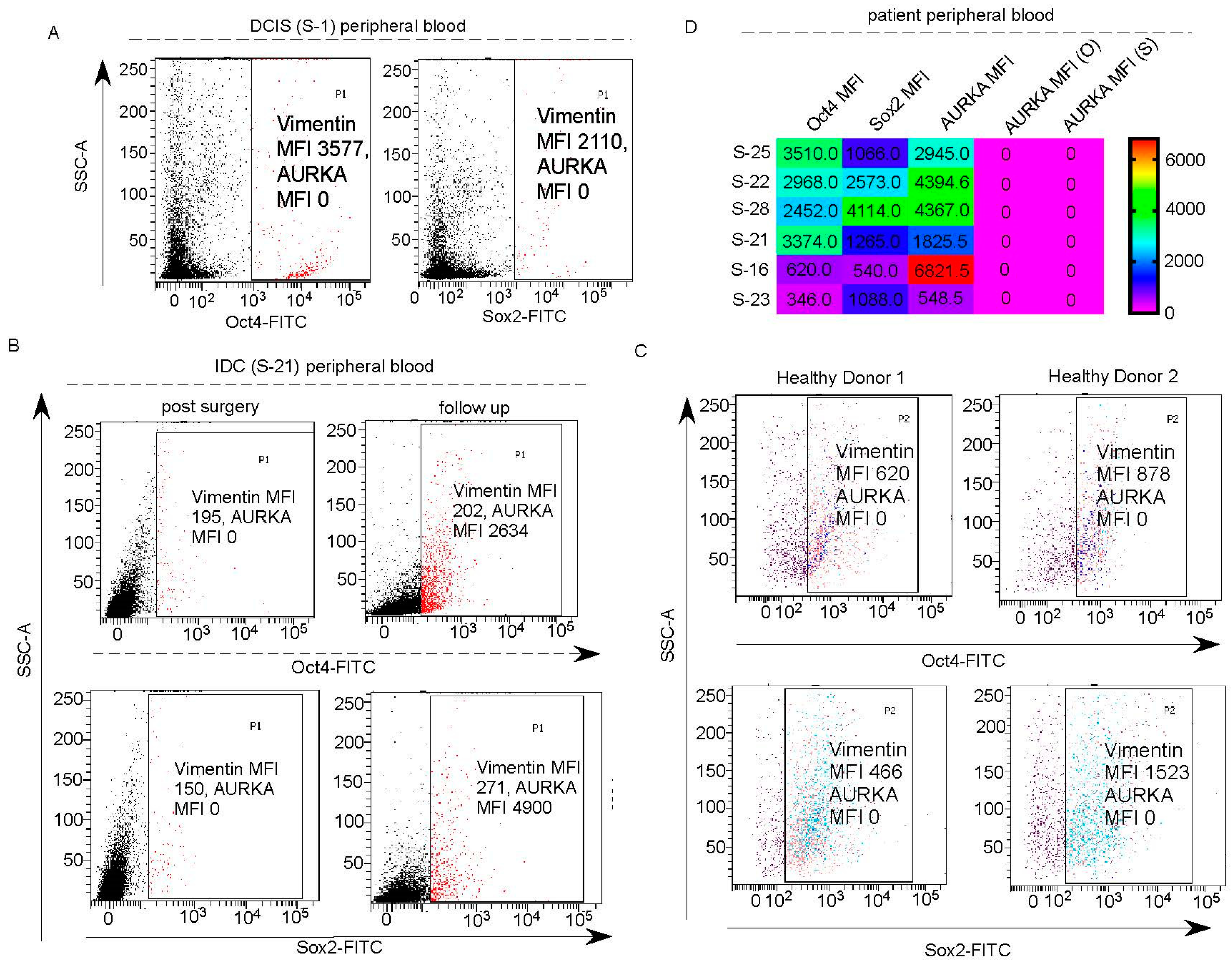
Figure 7.
Vimentin+ve cells, but not AURKA+ve cells in patient tumour and blood, are almost completely multi-drug resistant: Flow cytometric scatter plots showing almost 100% of Vimentin+ve cells in IDC patient tissue (A) and blood (B) are ABCG2+ve; however, the Vimentin+ve population primarily lies in CD44+CD24+ profile, while AURKA+ve cells although not necesarily ABCG2+ve, includes other CD44/CD24 profiles denoting necessity to analyse the AURKA+ cells whether or not ABCG2/Vimentin is expressed.
Figure 7.
Vimentin+ve cells, but not AURKA+ve cells in patient tumour and blood, are almost completely multi-drug resistant: Flow cytometric scatter plots showing almost 100% of Vimentin+ve cells in IDC patient tissue (A) and blood (B) are ABCG2+ve; however, the Vimentin+ve population primarily lies in CD44+CD24+ profile, while AURKA+ve cells although not necesarily ABCG2+ve, includes other CD44/CD24 profiles denoting necessity to analyse the AURKA+ cells whether or not ABCG2/Vimentin is expressed.
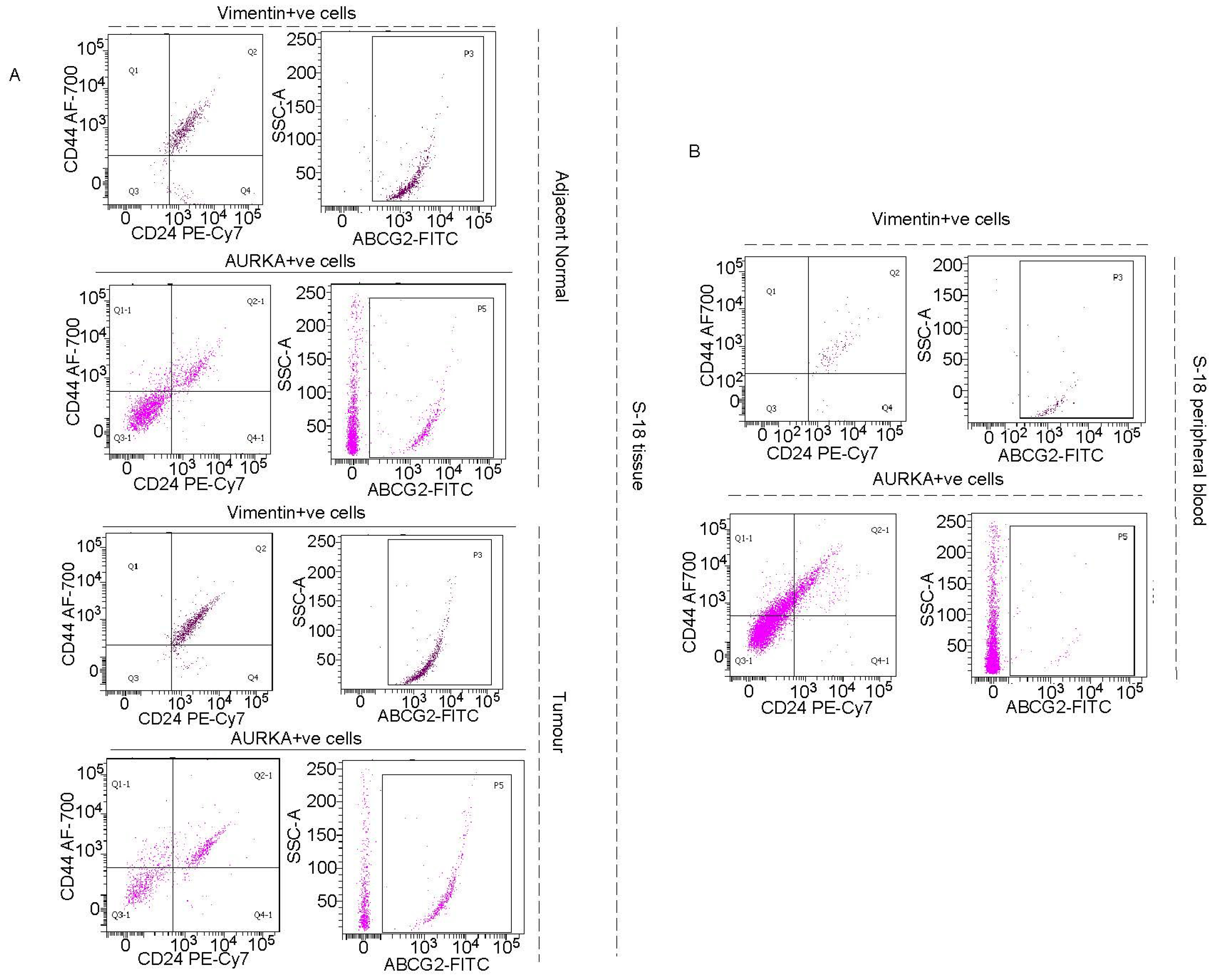
Figure 8.
CD44/CD24 profile changes with changes in ABCG2 expression with impact on AURKA and Vimentin expression in IDC: Flow cytometric scatter plots demonstrating AURKA expression has to be analysed in ABCG2+ve cells, since low-ABCG2 cells include AURKA+CD44+CD24-Vimentin- category. Notably, high-ABCG2 cells are essentially Vimentin+ve again.
Figure 8.
CD44/CD24 profile changes with changes in ABCG2 expression with impact on AURKA and Vimentin expression in IDC: Flow cytometric scatter plots demonstrating AURKA expression has to be analysed in ABCG2+ve cells, since low-ABCG2 cells include AURKA+CD44+CD24-Vimentin- category. Notably, high-ABCG2 cells are essentially Vimentin+ve again.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



