Submitted:
06 October 2023
Posted:
06 October 2023
You are already at the latest version
Abstract
The purported cognitive benefits associated with nicotine and its metabolites in the brain are a matter of debate. In this review, the impact of the pharmacologically active metabolite of a nicotine derivative produced by bacteria (6-hydroxy-L-nicotine) on memory, oxidative stress, and the activity of the cholinergic system in the brain was examined. A search in the PubMed, Science Direct, Web of Science, and Google Scholar databases, limiting entries to those no later than July 2023, was conducted. The search focused specifically on articles pertaining to nicotine metabolites, memory, oxidative stress, cholinergic system activity, as well as enzymes or pathways related to nicotine degradation in bacteria. The preliminary search resulted in 696 articles, and following the application of exclusion criteria, 185 articles were deemed eligible for inclusion. This review focuses on experimental studies supporting nicotine catabolism in bacteria, and the chemical and pharmacological activities of nicotine and its metabolite 6-hydroxy-L-nicotine.
Keywords:
nicotine
; metabolite
; bacteria
; 6-hydroxy-L-nicotine
; memory
; oxidative stress
; degradation.
1. Introduction
Nicotine (NIC) is a psychoactive substance found in tobacco and is the primary addictive component of cigarettes. Its effects on the human body are complex and can be both stimulating and relaxing. However, it is important to note that while NIC is often associated with tobacco use, it can also be consumed through other means such as e-cigarettes, nicotine replacement therapy (NRT) products, and smokeless tobacco. NIC is highly addictive and can lead to dependence, making it challenging for individuals to quit smoking or using nicotine-containing products [1]. Moreover, NIC has cognitive-enhancing effects, such as improved attention, memory, and cognitive function. This has led to studies exploring its potential therapeutic use in conditions like Alzheimer's (AD) and Parkinson's disease (PD) [2]. Some studies suggest that NIC may have neuroprotective effects, which means it could protect the brain from damage. However, the balance between potential benefits and risks is complex [3,4].
NIC catabolism in mammalian cells primarily occurs in the liver. The liver enzymes play a crucial role in breaking down nicotine into various metabolites. The major enzyme responsible for NIC metabolism is cytochrome P450 2A6 (CYP2A6). The primary metabolite formed through this pathway is cotinine. Cotinine is further metabolized to trans-3'-hydroxycotinine and other secondary metabolites. These metabolites are eventually eliminated from the body through urine [5]. Research on the relationship between cotinine, the primary metabolite of nicotine, and memory is an area of interest, particularly in the context of cognitive function and neurological disorders. Some studies suggest that cotinine may have cognitive-enhancing effects, potentially improving memory and cognitive function [6,7]. Cotinine has been investigated for its potential neuroprotective effects, and such protection may contribute to improved cognitive outcomes [8].
Research on the direct relationship between NIC metabolites produced by bacteria, memory, oxidative stress, and cholinergic system activity is limited. Therefore, the primary aim of this review is to identify the NIC catabolism in bacteria as a source of metabolites with possible neurological effects and to search for possible explanations of the mechanisms that mediate brain function.
2. Methodology
2.1. Search strategy
The current review was based on different electronic databases such as PubMed, Science Direct, Web of Science, and Google Scholar by using several keywords: nicotine [Title/Abstract] AND bacteria [Title/Abstract] AND (degrad* [Title/Abstract] OR catabol* [Title/Abstract] OR metab* [Title/Abstract]), nicotine [Title/Abstract] AND metabolite [Title/Abstract] and nicotine metabolite [Title/Abstract] AND memory [Title/Abstract]. For the selection of articles, the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guidelines were employed [9]. Two examiners (M.M. and L.H.) individually reviewed the title, abstract, and if needed, the complete text, according to the established criteria for inclusion and exclusion.
2.2. Inclusion and Exclusion Criteria
All experimental studies including the effects of NIC metabolites as compared to control on memory, oxidative stress, and gene expression in different animal models, as well as reporting enzymes or pathways for NIC degradation in bacteria were included until July 2023. The search was limited to texts in English and original articles. The following exclusion criteria were applied: 1) conference abstracts, books, and book chapters; 2) non-English articles; 3) reviews, systematic reviews, meta-analyses, and letters.
2.3. Data extraction
From the included articles, the data about NIC, metabolite synthesis, the type of articles, the mechanism, dose, duration, and route of metabolite administration, enzymes or pathways for NIC degradation in bacteria were extracted. The electronic search of the specified databases identified 696 articles, out of which 185 studies fulfilled the requirements for inclusion (Figure 1).
3. Nicotine
3.1. Chemical and pharmacological properties of nicotine
Nicotine (NIC, (S)-3-(1-methyl-2-pyrrolidinyl) pyridine) is an alkaloid present in the leaves of the tobacco plant (Nicotiana tabacum L.), where it constitutes up to 2 - 8% of the dry weight of the leaves, and to a lesser extent in other plants of the Solanaceae family such as eggplants, tomatoes, and potatoes, where it acts as a natural insecticide [10].
From a chemical standpoint, NIC is a heterocyclic compound composed of a pyridine ring and a pyrrolidine ring (each ring being a tertiary amine). It has an active center (appearing as R and S stereoisomers) and is relatively soluble in water. However, it prefers to separate in organic solvents, making it easily extractable from aqueous solutions. Only 0.1 - 0.6% of the total NIC content in tobacco appears in the form of the (R) isomer, with the majority being the (S) isomer. In its pure state, the compound is a pale yellow or dark brown liquid, and when heated, it acquires a fishy odor. The alkaloid is a weak base, with a melting point of - 79°C and a boiling point of 247°C [11]. The acidity constant (pKa) value of the nitrogen atom in the pyridine nucleus is 3.04, while the pKa value of the pyrrolidine nitrogen is 7.84 at 25°C. With these characteristics, approximately 23% of NIC is non-ionized at physiological pH, allowing it to rapidly traverse biological membranes [10]. Absorption of NIC can occur through the oral cavity, skin, lungs, urinary bladder, and gastrointestinal tract. A key factor regulating NIC passage through biological membranes is the pH of the environment [10]. In an acidic environment, NIC is ionized and does not readily cross membranes. For instance, due to the acidic pH of gastric juice, NIC is minimally absorbed when ingested and reaches the stomach. Tobacco smoke resulting from pipes and cigars is more alkaline (pH ≥ 6.5), and consequently, NIC is mostly non-ionized, being well absorbed in the oral cavity [10,12]. Cigarette smoke is more acidic (pH = 5.5 - 6) and significantly reduces NIC absorption from the oral cavity (as NIC is ionized at this pH), requiring smokers to inhale cigarette smoke into the lungs.
The primary route of NIC absorption is through the pulmonary alveoli. The alkaline fluid (pH = 7.4) on the inner surface of the lungs can buffer the acidic smoke to a neutral pH on the extensive alveolar surface, allowing NIC to be absorbed into the pulmonary circulation (the lungs can also act as a reservoir for NIC) [10,12]. In the blood (at physiological pH of 7.4), NIC is approximately 69% ionized and 31% non-ionized, with its binding to plasma proteins being less than 5% [10]. NIC can easily cross the blood-brain barrier (BBB) through passive diffusion and probably with the assistance of a transporter (the exact mechanisms are still unknown). Chronic NIC administration does not appear to influence its absorption kinetics in the brain [13]. The plasma half-life (t1/2) of NIC is approximately 2 h, while in the brain, the t1/2 of NIC is 10 min. The half-life represents the time it takes for 50% of the maximum dose of NIC in the brain to decrease, as NIC is distributed to other high-affinity compartments in the body (such as the liver, kidneys, spleen, and lungs) [10]. When the t1/2 of NIC was determined through urinary excretion, it was longer (~ 11 h). This can be explained by the slow release of NIC from body tissues [10].
NIC is extensively metabolized in the liver into six major metabolites: nicotine-glucuronide, nicotine N-oxide, nornicotine, nicotine isomethonium ion, cotinine, and 2-hydroxynicotinine. The predominant pathway during presystemic metabolism (first-pass metabolism) leads to the formation of cotinine (in humans, 70 - 80% of NIC is metabolized into cotinine), which may have relevance in the various neurobiological effects of smoking, as it is a possible ligand for nicotinic acetylcholine receptors (nAChRs) [10]. Several enzymes are involved in NIC metabolism in the liver, such as some cytochrome P450 enzymes (subtypes CYP2A6, CYP2B6, and CYP2D6), flavin monooxygenase 3 (FMO3), N-methyltransferase, aldehyde oxidase, and UDP-glucuronosyltransferase [10]. NIC excretion that is not metabolized in the urine accounts for approximately 5% of total elimination [14]. Animal studies have suggested that NIC is metabolized to a small extent in extrahepatic tissues (e.g., kidneys, lungs, brain) [10]. The rate of NIC metabolism is influenced by various factors such as age, sex, food consumption, ethnicity, liver or kidney diseases, pregnancy, tobacco ingredients (e.g., menthol, which inhibits both CYP2A6 and UDP-glucuronosyltransferase activity), or medication (e.g., contraceptive use) [10,14]. The actual tobacco smoke influences NIC metabolism. NIC elimination is slower (resulting in a tendency for a longer NIC t1/2) in smokers compared to non-smokers [10,14]. This phenomenon can be explained by the effect of β-nicotyrine (a component of smoke with inhibitory activity on CYP2A6), as well as the reduced expression of CYP2A6 protein and mRNA levels in the liver caused by NIC [10].
3.2. The cognitive effects of nicotine
The brain's cholinergic nicotinic system is involved in several aspects of major mental disorders such as AD, PD, attention-deficit hyperactivity disorder (ADHD), and schizophrenia. Both clinical and animal studies support the role of nicotinic acetylcholine receptors (nAChRs) in learning, memory, and cognition. NIC is the prototypic agonist of nAChRs, which could enhance cognition through direct effects on attention and interaction with presynaptic nAChRs to facilitate the release of neurotransmitters involved in memory and learning: acetylcholine (ACh), glutamate, dopamine, norepinephrine, serotonin, and gamma-aminobutyric acid (GABA). Several studies have demonstrated that NIC enhances attention and improves the learning process. Additionally, NIC and nicotinic derivatives have been shown to possess neuroprotective effects, likely mediated by stimulation of α7 nAChRs. To assess the effectiveness of NIC and its potential for use in treatments to enhance cognitive performance, this alkaloid has been administered to patients with cognitive disorders through patches and injections. This strategy has been employed to demonstrate the efficacy of NIC treatment in various populations, including normal non-smoking adults, AD patients, schizophrenic patients, and adults with ADHD. Animal models have also been utilized to determine the efficacy of NIC treatment for enhancing cognitive functions and to ascertain the mechanism of action of this alkaloid. Ultimately, clinical and animal studies have provided insights into the development of NIC-based treatments for cognitive dysfunction [15].
3.3. Short-term cognitive effects of nicotine
Although numerous studies have investigated the acute effects of NIC administration on cognition, the results are contradictory. NIC appears to enhance cognition in several domains, particularly memory. This was demonstrated by Shu et al. [16], who investigated the effect of acute NIC treatment on lipopolysaccharide (LPS) - induced impairment of fear memory reconsolidation and the underlying mechanism. The authors found that the stimulatory effects of acute NIC treatment are limited to improving memory deficits induced by inflammation or other stimuli. However, some studies do not show cognitive improvement effects due to NIC. Grus and Hromatko [17] evaluated the acute administration of NIC on 22 occasional smokers aged 19-29 years. Attention, working memory, and visuospatial reasoning were assessed using a within-subjects design with a control setting. The authors concluded that, at least among young, occasional smokers, smoking does not affect cognition and the claims of its improvement are probably a result of some sort of cognitive bias. The contradictory results of these two studies may be explained by different NIC doses and administration routes used. Indeed, a higher dose of NIC is needed to achieve a facilitated response [18]. Although an appropriate dose of NIC is essential to achieve a beneficial effect, high doses can have adverse effects on cognition. Poltavski et al. [19] investigated the effect of different doses administered through NIC patches (7, 14, and 21 mg) on cognitive performance. They found an inverted U-shaped relationship between different NIC doses and cognitive performance. Indeed, as previously emphasized by Newhouse et al. [20], moderate NIC consumption can lead to optimal performance, while low or high consumption can affect performance. These studies suggest that only individuals performing below the optimal level can benefit from NIC administration. However, in cases where individuals have optimal performance, NIC administration will affect cognition, as shown in the study conducted by Grundey et al. [21]. This idea is supported by studies indicating that NIC can act as a cognitive enhancer in individuals with low performance, such as those with AD, schizophrenia, or ADHD [20]. In an innovative study, Potter and Newhouse [22] investigated how acute NIC administration, in the form of a patch (7 mg administered for 45 min), affects various cognitive functions in 15 non-smoking adults diagnosed with ADHD. After NIC administration, participants showed improved behavioral inhibition and a non-significant trend toward improved recognition memory. In conclusion, acute NIC administration supports cognition, but this effect seems to be limited to the domains of attention and memory. Moreover, this effect appears to vary depending on the NIC dose.
3.4. Effects of nicotine on neuroinflammation
Aging is associated with changes in the immune system that generally promote pro-inflammatory cytokines and the process of neuroinflammation in the brain [23]. Neuroinflammation has been implicated in the pathophysiology of several age-related neurodegenerative disorders, such as AD or PD [24,25]. Aging-related neuroinflammation reduces neuronal plasticity, with long-term effects on cognitive function [26]. Preclinical studies have shown that the administration of NIC reduces neuroinflammation in the brain [27]. This action may be mediated by the systemic effects of NIC administration in preventing the proliferation of T cells in peripheral tissue and their infiltration into the brain. Additionally, NIC modifies the production of TNF-α, IL-1β, IL-6, MIP-2/CXCL2, MIP 1α/CCL3, and eotaxin/CCL11 in T helper cells (Th cells). All these factors can counteract inflammation [28]. Microglia are the resident macrophages of the brain and serve to mediate the innate immunity of the nervous system. Studies have demonstrated that NIC administration significantly reduces microglial activation. Considering that the degeneration of cholinergic neurons with aging is accompanied by increased microglial activation, this suggests a mechanism of neuroprotection [24]. Furthermore, NIC reduces the production of certain inflammatory cytokines (such as IL-6 and TNF-α) in astrocytes, thereby mitigating neuroinflammation in the brain [29].
The reduction of the central nervous system (CNS) neuroinflammation induced by NIC is mediated by nAChRs [28]. Among all the receptors that mediate the anti-inflammatory effects of NIC, the α7 subtype deserves special attention [30]. A study conducted in 2003 by Wang et al. [31] demonstrated, for the first time, that the α7 subunit of nAChRs plays an important role in suppressing cytokine production in response to NIC stimulation. After treating wild-type mice and those with the gene for the α7 subtype of nAChRs deactivated with a bacterial endotoxin called LPS, the level of TNF-α in the serum was significantly higher in genetically modified mice than in wild-type ones. Stimulation with NIC or ACh had no effect on TNF-α expression in LPS-treated peritoneal macrophages obtained from mice with the α7 nAChRs gene deactivated. This suggests that α7 nAChRs are essential for blocking cytokine synthesis through the cholinergic anti-inflammatory pathway [31]. In another study by De Simone et al. [32], it was found that in LPS-activated microglial cells, the interaction between NIC and α7 nAChRs led to a significant activation of cyclooxygenase 2 (COX-2) expression and prostaglandin E2 synthesis, but there was also a moderate (or absent) effect on nitric oxide, IL-1β, and IL-10. Activation of α7 nAChRs expressed by microglia and T cells results in a temporary increase in intracellular Ca2+ levels in these cells, which subsequently leads to a decrease in the phosphorylation of the mitogen-activated protein kinase (MAPK), p38, and p44 and, consequently, a reduction in the expression of pro-inflammatory cytokines [33]. Additionally, activation of α7 nAChRs in monocytes or macrophages leads to unfavorable effects in the neuroinflammation process: (1) prevention of IκB phosphorylation, an inhibitor of nuclear factor-kappa B (NF-κB) transcription factor, (2) activation of adenylate cyclase 6, and (3) recruitment of Janus kinase 2 (JAK2). All these initiate cascades of interactions that ultimately deactivate the NF-κB signaling pathway and reduce the expression of pro-inflammatory cytokines. Furthermore, the results of an experiment conducted by Nizri et al. [34] suggested that α7 nAChR activation by NIC has immunomodulatory properties, suppressing the reactions of Th1 and Th17 cells, but not Th2 cells. Moreover, treatment with NIC (2 mg/kg for 28 days) significantly suppressed clinical symptoms of experimental autoimmune encephalomyelitis and inflammatory infiltration in the CNS in mice. In another in vivo study, it was demonstrated that intraperitoneal administration of NIC (0.2, 0.4, and 0.8 mg/kg) blocked the expression (at the mRNA level) of pro-inflammatory cytokines induced by LPS, which had previously been injected intracerebroventricularly (i.c.v.) in rats to induce neuroinflammation [35]. Furthermore, it appears that the anti-inflammatory effect of NIC was blocked by the administration of methyllycaconitine (MLA), an α7 nAChR antagonist, but not by dihydro-β-erythroidine (DHβE), an α4β2 nAChR antagonist, suggesting that NIC's inhibitory effect on pro-inflammatory cytokines is due to its action on α7 nAChRs. In conclusion, the anti-inflammatory characteristics of NIC make it a promising agent in preventing or mitigating age-induced neuroinflammation in the brain [26].
3.5. Effects of nicotine on apoptosis
Apoptosis, or programmed cell death, is an energy-dependent suicidal process in which the targeted cell is eliminated without the inflammation typically seen in necrotic degeneration [36,37]. While apoptosis is an essential element of brain development, aberrant or pathological apoptosis has been associated with several neurodegenerative disorders [37]. Furthermore, it has been demonstrated that brain aging renders it more vulnerable to apoptotic-induced neuronal injuries, which can lead to age-related cognitive impairments [26].
Since NIC prevents apoptosis, it has been termed a “survival agonist” [38]. A growing body of evidence has indicated that NIC protects neurons against apoptosis through both caspase-dependent and caspase-independent pathways [39]. NIC administration inhibits the activation of caspases 3, 8, and 9, thus blocking the caspase-dependent pathway [38,40]. Additionally, NIC prevents the release of apoptosis-inducing factors from mitochondria and their translocation into the nucleus, which can be mediated by the activation of α7 nAChRs [39]. Evidence suggests that the α7 nAChR receptor is not the sole subtype involved in the anti-apoptotic effects of NIC. The α4β2 subtype, which has a wider distribution in the brain and a higher affinity for NIC, can similarly mediate these anti-apoptotic effects [41]. On the other hand, there are studies that have reported contradictory results. In an experiment conducted by Hritcu et al. [42] on Wistar rats, chronic NIC treatment induced DNA fragmentation accompanied by an increase in caspase 3 activity in neurons located in the temporal cortex, suggesting an intensification of apoptosis. These findings support the results of the Jang et al. [43] group, which showed that exposure of male rats to NIC for 3 days led to a 110% increase in caspase 3 activity in the dentate gyrus. The anti-apoptotic effects mediated by NIC can also be achieved through the activation of the MAPK cascade and extracellular signal-regulated kinase (ERK) 2, which plays an important role in regulating cell growth and apoptosis [44]. Although one study suggested that changes in the anti-apoptotic protein Bcl-2 levels might not be involved in the anti-apoptotic effects of NIC [44], a more recent study demonstrated that Bcl-2 is involved in the anti-apoptotic effects of NIC through the α7 nAChRs/JAK2/signal transducer and activator of transcription 3 (STAT3)/NF-κB/Bcl-2 signalling pathway in neurons [45]. Moreover, NIC reduces neuronal nitric oxide synthase (nNOS) activity and nitric oxide production, which may contribute to its anti-apoptotic effects [44]. It appears that NIC when administered in appropriate doses for a mature brain, can halt age-induced neuronal apoptosis and thereby reduce cognitive impairments [26].
3.6. Effects of nicotine on neurotrophic factors
Neurotrophic factors are members of a protein family that includes brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), and glial cell-derived neurotrophic factor (GDNF). Together, these factors play a significant role in the development, differentiation, survival, and function of neurons [46]. Normally, the production of neurotrophic factors decreases over time with brain aging [47]. Evidence has suggested that these factors, especially BDNF and downstream pathways, could represent new and interesting therapeutic targets for treating cognitive deficits and age-associated brain changes [48].
Numerous studies have shown that NIC can have neurotrophic effects and, in conjunction with nAChRs, can play a crucial role in neuron development and maturation [49]. NIC activates α7 nAChRs and can increase NGF expression through NF-κB-dependent pathways [50]. Indeed, NIC increases nuclear translocation and transcriptional activity of NF-κB and enhances p65 binding to the NGF gene promoter region, ultimately leading to increased NGF expression [50]. Additionally, NIC increases TrkA receptor mRNA expression, which mediates NGF effects in neurons [51]. Subsequently, NGF exerts neuroprotective effects by promoting synaptic plasticity while attenuating glutamate-induced excitotoxicity [52]. Studies have also shown positive effects of NGF on learning and memory, further supporting its neuroprotective effects [53]. In addition to these effects mentioned above, there is evidence that certain doses of NIC can increase BDNF levels in the hippocampus and neocortex [54]. Administration of α-bungarotoxin (α-BTX), a selective antagonist of α7 nAChRs, reduces BDNF mRNA expression in the brain, suggesting that this NIC-induced increase in BDNF levels may be mediated through the α7 nAChR receptor [55]. Other studies have shown that BDNF plays a significant role in memory trace formation in the hippocampus and can impact long-term potentiation (LTP) [26,56]. Similar effects of NIC have been reported for GDNF levels, a cytokine that has been shown to enhance memory in animal models [26]. Consequently, it appears that NIC, in a dose-dependent manner and through its positive effects on neurotrophins, may improve memory and learning deficits that can occur as part of brain aging [26].
3.7. Effects of nicotine on amyloid beta peptide (Aβ)
Considerable evidence has shown that Aβ and its aggregated forms are contributors to brain aging [57]. Specifically, animals with accelerated senescence exhibit higher levels of the amyloid-beta precursor protein (APP) and Aβ associated with learning and memory impairments at younger ages [58]. Promising preclinical studies have indicated that injection of anti-Aβ antibodies reduces cognitive deficits in these animals [59], while recent clinical trials with similar antibodies have failed to demonstrate beneficial changes in AD patients [60]. Accumulated evidence has demonstrated that both short-term and long-term NIC treatment significantly reduces Aβ deposits and plaque accumulations in the brains of transgenic mice [61]. This reduction in Aβ plaque density includes both parenchymal and vascular deposits. Several mechanisms have been suggested to account for this phenomenon. Particularly, NIC administration increases the total amount of APP in the cerebrospinal fluid (CSF), which likely hampers amyloidogenesis due to enhanced clearance. However, it remains unclear whether the effects of NIC on Aβ clearance are direct [62] or associated with increased CSF flow. Additionally, NIC might favor the breakdown of amyloid fibrils, thereby interfering with Aβ plaque buildup [63,64]. The improved cholinergic functions resulting from NIC's agonism towards nAChRs could also contribute to Aβ deposition reduction, with specific involvement of α7 subtype receptors. It has been suggested that a direct interaction between Aβ and α7 nAChRs leads to increased Aβ-induced MAPK activation and subsequent phosphorylation of cAMP response element-binding protein (CREB) with an attenuating effect downstream on Aβ deposition [63,65]. Furthermore, chronic NIC treatment might exert neuroprotective influence against pre- and postsynaptic injuries caused by Aβ oligomers or amyloidosis in the pre-plaque formation stage. This effect is thought to be mediated by the interaction between α7 nAChRs and the phosphatidylinositol-3-kinase (PI3K)/Akt signaling pathway at pre- and postsynaptic elements [66]. Additionally, α7 nAChR activation through NIC administration activates the Wnt/β-catenin signaling pathway, which is considered to play a major role in protection against Aβ aggregates in the brain [67]. Considering all these studies, it is highly plausible that NIC diminishes Aβ plaque burden and oligomer concentration in the aging brain, thereby exerting neuroprotective effects against Aβ-induced lesions and cognitive impairments [68].
3.8. Effects of nicotine on oxidative stress
Considerable evidence has shown that oxidative stress is a phenomenon resulting from an imbalance between the production of reactive oxygen species (ROS) and antioxidants, such as free radical scavenging systems [37]. Oxidative stress, particularly oxidative damage to the brain induced by iron, appears to play a crucial role in triggering neuronal death and is thus implicated in many age-related neurodegenerative disorders, such as AD and PD [69]. Furthermore, due to the brain's high oxygen metabolism and its limited regenerative capacity [69,70], oxidative stress is considered a significant factor in the brain aging process and the cognitive and functional impairments associated with it [71]. Although NIC's properties regarding oxidative stress and neuroprotection are controversial and may be complicated by inverted U-shaped dose-response curves [14,72], several studies have reported the antioxidant effects of NIC on neurodegenerative disorders such as AD and PD [73]. It has been demonstrated that NIC administration, under certain circumstances, can reduce ROS-induced lipid peroxidation both in vivo and in vitro [72,74]. This could result from NIC's ability to chelate Fe2+ through the nitrogen atom in its pyridine nucleus, thereby inhibiting the Fenton reaction involved in hydroxyl free radical formation [74]. Other studies have confirmed NIC's iron-chelating capacity and its prevention of the Fenton reaction. It has also been suggested that NIC can bind to Fe2+ in the pro-inflammatory enzyme thromboxane synthase, thereby inhibiting its function [75,76]. On the other hand, some studies have not shown a negative effect of NIC on ROS generation and lipid peroxidation [72,76]. Indeed, in certain circumstances, NIC administration interferes with the mitochondrial respiratory chain, leading to ROS production and oxidative stress [72]. Moreover, NIC has been shown to increase the levels of malondialdehyde (MDA) and lactate dehydrogenase activity, which can trigger lipid peroxidation [77]. Additionally, NIC is a substrate for cytochrome P450 enzymes, which could lead to intracellular oxidative stress [72]. A pro-oxidant activity of NIC was described by Hritcu et al. in 2009 [42]. Following an in vivo experiment on male Wistar rats, chronic treatment with NIC (0.3 mg/kg, i.p. for 7 consecutive days) led to a decrease in antioxidant enzyme activities superoxide dismutase (SOD) and glutathione peroxidase (GPX), and an increase in MDA and ROS levels [42]. These conflicting studies may reflect various factors involved, such as NIC dosage (high or low), enantiomer choice, and specific effects in brain regions [77]. NIC's effects on oxidative stress are dose-dependent, as antioxidant effects can be observed at low doses (10 µM), while oxidative stress exacerbation occurs at high doses (1 - 10 mM) [72]. However, a concentration of NIC as low as 0.8 µM was reported to induce oxidative stress [78]. Moreover, it has been reported that NIC-induced changes in the expression of antioxidant-related genes differ between brain regions [77].
4. 6-hydroxy-L-nicotine
4.1. Nicotine catabolism in bacteria as a source of 6-hydroxy-L-nicotine
NIC is a naturally produced plant alkaloid present in the leaves and stems of plants from the Solanaceae family. During the plant’s life cycle, nicotine ends up in the soil together with the dead plant material, offering a specific carbon and nitrogen source for the soil bacteria. It is hence no surprise that this ecological niche was occupied by microorganisms that have evolved genes and pathways to utilize this compound to sustain cell growth. Bacteria that have evolved NIC degradation pathways include strains of Paenarthrobacter [79,80], Arthrobacter [81,82,83], Pseudomonas [84,85,86,87,88,89,90,91,92,93], Agrobacterium [94], Nocardioides [95,96], Ochrobactrum [97,98,99], Shinella [100,101], Acinetobacter [102], Bacillus [103,104], Sphingomonas [105] Pusilimonas [106] Rhodococus [96,107,108] and Ensifer [109].
Among these NIC-degrading bacteria (NDB), three main NIC degradation pathways have been described based on the identified metabolic intermediates: the pyridine pathway, the pyrrolidine pathway, and a variation of the pyridine and pyrrolidine pathway – the VPP pathway [110].
Figure 2.
The nicotine catabolic pathways producing 6-hydroxy-L-nicotine (6HLN) as an intermediate: pyridine pathway in Paenarthrobacter nicotinovorans ATCC 49919 (blue) and the VPP pathway in Agrobacterium tumefaciens S33. Bold letters indicate enzymes: NDH - nicotine dehydrogenase; 6HLNO - 6-hydroxy-L-nicotine oxidase; 6HDNO - 6-hydroxy-D-nicotine oxidase; KDH - ketone dehydrogenase; DHPONH - 2,6-dihydroxypseudooxynicotine hydrolase; DHPH – 2,6-dihydroxypyridine-3-hydroxylase NBOR – nicotine blue oxidoreductase; MABO - γ-N-methylaminobutyrate oxidase; FolD - methylene-tetrahydrofolate dehydrogenase/cyclohydrolase; PurU - formyl-tetrahydrofolate deformylase; MAO - monoamine-oxidase; AO – amine-oxidase; SsaDH - succinic semialdehyde dehydrogenase; PKC – putative polyketide cyclase; NIT - w-amidase; Ndo - nicotine hydroxylase; Pno - 6-hydroxypseudooxynicotine oxidase; Sapd - 3-succinoylsemialdehyde-pyridne dehydrogenase; Hsh - 6-hydroxy-3-succinoylpyridine 3-monooxygenase; Hpo - 2,5-dihydroxypyridine 5,6-dioxygenase; Nfo - N-formylmaleamate deformylase; Ami - maleamate amidohydrolase; Iso - maleate cis/trans-isomerase; CAPS and smaller fond indicates the intermediates: L-6-HMM – 6-hydroxy-L-methylmyosmine; D-6-HMM – 6-hydroxy-D-methylmyosmine; 6-HPON – 6-hydroxy-pseudooxynicotine; 2,6-DHPON – 2,6-dihydroxypseudooxynicotine; 2,6-DHP – 2,6-dihydoxypyridine; CH3-4-GABA - γ-N-methylaminobutyrate ; 2,3,6-THP - 2,3,6-trihydroxypyridine; NB -nicotine blue, 4,4‘,5,5‘-tetrahydroxy-3,3‘-diazadiphenoquinone-(2,2‘); CH2TH4 - methylenetetrahydrofolate; GABA - γ-aminobutyric acid ; SSA - succinic semialdehyde, alpha-KGA - a-keto-glutaramate; alpha-KG - a-keto-glutarate; 6HSAP - 6-hydroxy-3-succinoylsemialdehyde pyridine; HSP - 6-hydroxy-3-succinoyl pyridine; 2,5-DHP – 2,5-dihydoxypyridine.
Figure 2.
The nicotine catabolic pathways producing 6-hydroxy-L-nicotine (6HLN) as an intermediate: pyridine pathway in Paenarthrobacter nicotinovorans ATCC 49919 (blue) and the VPP pathway in Agrobacterium tumefaciens S33. Bold letters indicate enzymes: NDH - nicotine dehydrogenase; 6HLNO - 6-hydroxy-L-nicotine oxidase; 6HDNO - 6-hydroxy-D-nicotine oxidase; KDH - ketone dehydrogenase; DHPONH - 2,6-dihydroxypseudooxynicotine hydrolase; DHPH – 2,6-dihydroxypyridine-3-hydroxylase NBOR – nicotine blue oxidoreductase; MABO - γ-N-methylaminobutyrate oxidase; FolD - methylene-tetrahydrofolate dehydrogenase/cyclohydrolase; PurU - formyl-tetrahydrofolate deformylase; MAO - monoamine-oxidase; AO – amine-oxidase; SsaDH - succinic semialdehyde dehydrogenase; PKC – putative polyketide cyclase; NIT - w-amidase; Ndo - nicotine hydroxylase; Pno - 6-hydroxypseudooxynicotine oxidase; Sapd - 3-succinoylsemialdehyde-pyridne dehydrogenase; Hsh - 6-hydroxy-3-succinoylpyridine 3-monooxygenase; Hpo - 2,5-dihydroxypyridine 5,6-dioxygenase; Nfo - N-formylmaleamate deformylase; Ami - maleamate amidohydrolase; Iso - maleate cis/trans-isomerase; CAPS and smaller fond indicates the intermediates: L-6-HMM – 6-hydroxy-L-methylmyosmine; D-6-HMM – 6-hydroxy-D-methylmyosmine; 6-HPON – 6-hydroxy-pseudooxynicotine; 2,6-DHPON – 2,6-dihydroxypseudooxynicotine; 2,6-DHP – 2,6-dihydoxypyridine; CH3-4-GABA - γ-N-methylaminobutyrate ; 2,3,6-THP - 2,3,6-trihydroxypyridine; NB -nicotine blue, 4,4‘,5,5‘-tetrahydroxy-3,3‘-diazadiphenoquinone-(2,2‘); CH2TH4 - methylenetetrahydrofolate; GABA - γ-aminobutyric acid ; SSA - succinic semialdehyde, alpha-KGA - a-keto-glutaramate; alpha-KG - a-keto-glutarate; 6HSAP - 6-hydroxy-3-succinoylsemialdehyde pyridine; HSP - 6-hydroxy-3-succinoyl pyridine; 2,5-DHP – 2,5-dihydoxypyridine.
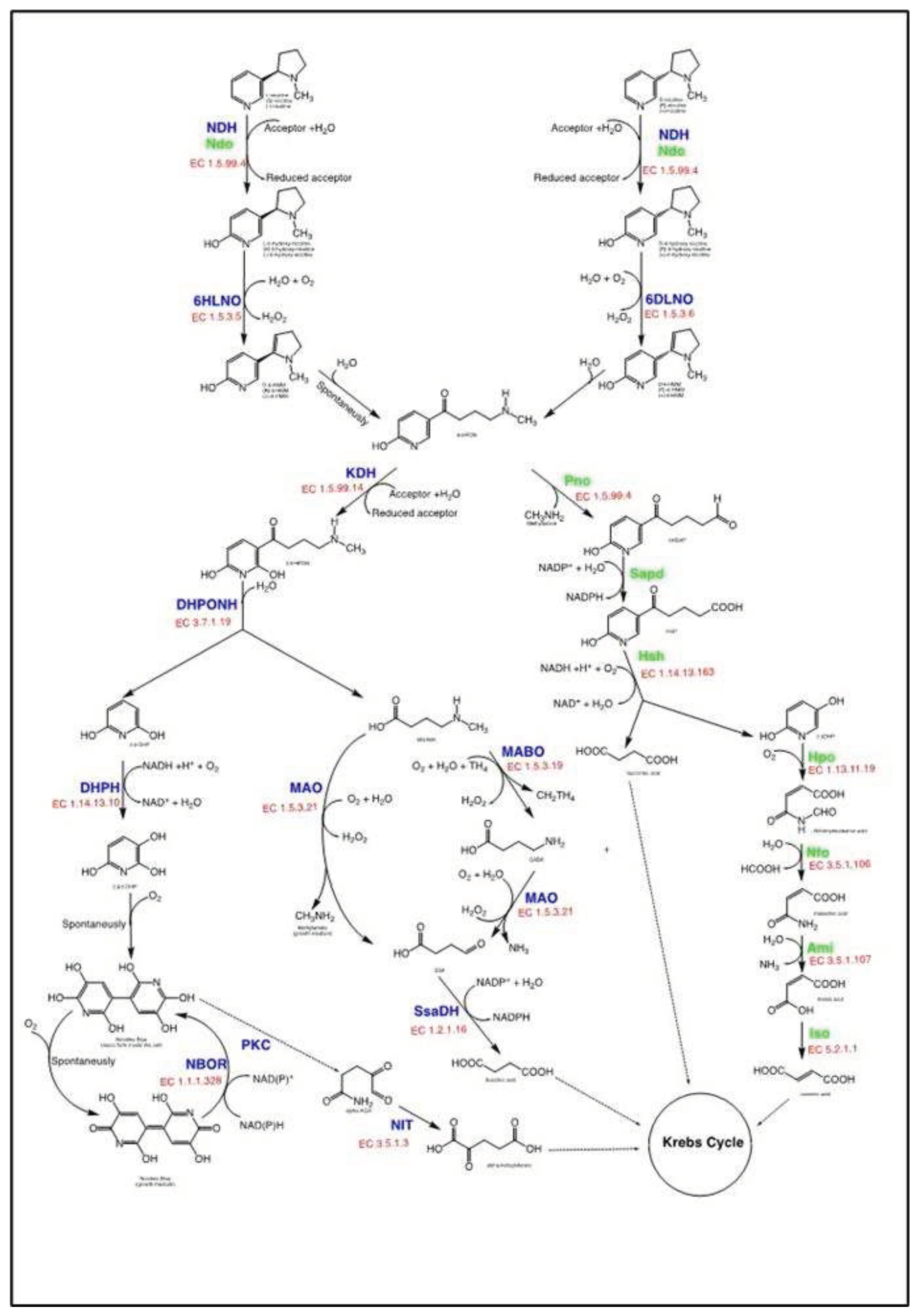
In the pyridine pathway, NIC is first attacked at the C6 of the pyridine ring by hydroxylation [111] forming 6-hydroxy-nicotine, while in the pyrrolidine pathway, the pyrrolidine ring of nicotine is either dehydrogenated [112,113], demethylated [89,92,93] or hydroxylated in C2 [92] The VPP pathway shares the first steps with the pyridine pathway and the final steps with the pyrrolidine pathway [114,115]. Only the strains harboring the pyridine pathway and the VPP pathway produce 6HLN as a metabolic intermediate (Figure 2) and have relevance for the current work.
4.1.1. The pyridine pathway for nicotine degradation
All strains that harbor the pyridine catabolic pathway are Gram-positive and belong to the Actinobacteria. The NIC catabolic gene cluster (nic-genes) in Paenarthrobacter nicotinovorans (former Arthrobater nicotinovorans) and other Arthrobacters are located on a plasmid [116,117] while in Rhodococcus and Nocardioides sp. the nic-genes are located on the chromosome [117].
By far, the most well-studied pathway is the one encoded by the pAO1 plasmid from P. nicotinovorans ATCC 49919 [118] Not only that the complete genome of the strain was sequenced [80] but also the nicotine-induced proteome of the strain is available [119,120,121] On the pAO1 megaplasmid, the nic-genes cluster is flanked by integrases and consists of a 49 kb catabolic transposon [122] that was shown to be spreading the nic-genes from/to chromosomes and plasmids in soil bacteria [116] It has been suggested that this catabolic transposon has been transferred into pAO1 from the chromosome of a bacterium possibly related to Rhodococcus, Arthrobacter, or Penarthrobacter [116,117,123].
The transposon is organized into several gene modules [116] responsible for the catabolism of nicotine to α-ketoglutarate, succinate, methylamine, and the characteristic blue pigment known as nicotine blue (NB or 4,4′,5,5′-tetrahydroxy-3,3′-diazadiphenoquinone-(2,2′)) (Figure 2). While NB and methylamine accumulate in the growth medium, α-ketoglutarate and succinate are integrated into the Krebs cycle and support cell growth [119,121]. In P. nicotinovorans ATCC49919, NIC is taken up by the cell by facilitated diffusion, but the gene encoding the permease responsible for the importation is still unknown [124]. The first step of the pathway happening inside the cell is the hydroxylation of NIC at position C6 of the pyridine ring. This step is encoded by nicotine-dehydrogenase (nicotine:acceptor 6-oxidoreductase (hydroxylating), EC 1.5.99.4, NDH). The enzyme is a trimeric metalloprotein consisting of a 14.9 kDa subunit containing an iron-sulfur cluster, a 30 kDa subunit with a flavin adenine dinucleotide (FAD) - binding domain, and an 87.7 kDa subunit containing the molybdopterin site, respectively [111,125]. The enzyme can act on both the naturally abundant (S)-enantiomer and the less abundant (R)-enantiomer of NIC, with retention of configuration in both cases.
The (R/S) 6-hydroxy-nicotine is further oxidized by two strictly stereospecific enzymes. (S)-6-hydroxy-nicotine oxidase (6-hydroxy-L-nicotine oxidase, 6HLNO, EC 1.5.3.5) is a member of the monoamine oxidase (MAO) family of proteins and contains non-covalently bound FAD. Long thought to perform an oxidation of the C2–C3 bond of the pyrrolidine, it was shown and generally accepted that that the enzyme catalyzes oxidation of the C2–N bond forming 6-hydroxy-N-methylmyosmine. The product amine is unstable and forms 6-hydroxypseudooxynicotine following nonenzymatic hydrolysis [126,127]. (R)-6-hydroxynicotine oxidase (6-hydroxy-D-nicotine oxidase, 6HLNO, EC 1.5.3.6) belongs to the p-cresol methylhydroxylase-vanillyl-alcohol oxidase family and contains a FAD covalently bound to the protein through a C8 α-histidyl linkage [128]. As in the case of 6HLNO, 6HDNO also performs oxidation of the C2–C3 bond of the pyrrolidine followed by nonenzymatic hydrolysis of 6-hydroxy-N-methylmyosmine to form 6-hydroxypseudooxynicotine.
The next enzyme in the NIC pyridine catabolic pathway in P. nicotinovorans ATCC49919 is 6-hydroxypseudooxynicotine dehydrogenase (ketone dehydrogenase, KDH, EC 1.5.99.14) which catalyzes a new hydroxylation of the pyridyl ring to form 2,6-dihydroxypseudooxynicotine. The enzyme is similar to NDH, also being a trimeric metalloprotein with the large subunit containing molybdopterin cytosine dinucleotide (MCD) [129]. The requirement for MCD of NDH and KDH is highly consistent with the presence of the pAO1 plasmid of several genes coding for proteins involved in the uptake of molybdenum and biosynthesis of the molybdopterin cofactor [130]. The product of the KDH reaction is 2,6-dihydroxypseudooxynicotine which is further cleaved to 2,6-dihydropyridine and N-methylaminobutyrate by 2,6-dihydroxypseudooxynicotine hydrolase (DHPON hydrolase, EC 3.7.1.19). The enzyme belongs to the α/β hydrolase family that catalyzes a broad range of hydrolase and lyase reactions and its structure is known [131].
All steps from NIC to 2,6-dihydroxypyridine and N-methylaminobutyrate are part of what is called the upper NIC pathway. The two products go separate ways in the lower nicotine pathway: 2,6-dihydroxypyridine is converted to blue pigment and N-methylaminobutyrate is integrated into the Krebs cycle.
This integration of N-methylaminobutyrate can be done in two different ways, depending on the C substrate available to the cell. One direction is oxidative demethylation and the other is oxidative deamination to form succinate semialdehyde and methylamine. In the oxidative demethylation, the substrate is converted to hydrogen peroxide, a methyl group, and 4-aminobutryate by a 4-methylaminobutanoate oxidase (formaldehyde-forming) (γ-N-methylaminobutyrate demethylating oxidase, MABO, EC 1.5.3.19) that contains covalently bound FAD [132]. In the oxidative deamination pathway, N-methylaminobutyrate is the substrate for 4-methylaminobutanoate oxidase (methylamine-forming) (γ-N-methylaminobutyrate oxidase, MAO, EC 1.5.3.21) an flavoprotein with noncovalently-bound FAD that converts it to semialdehyde and methylamine. The latter compound is excreted in the medium by a metabolic valve comprising a two-component small multidrug resistance pump and is known to accumulate during nicotine catabolism [133]. MAO can also deaminate 4-aminobutryate to form succinic semialdehyde. So, either way, N-methylaminobutyrate is metabolized, always succinic semialdehyde is formed and is further converted to succinic acid by an NADP+-dependent succinate semialdehyde dehydrogenase (SsaDH, EC 1.2.1.16) [122]. The microorganism was shown to select the demethylation or the deamination pathway depending on the C availability. When C sources are scarce and only nicotine is available, the demethylation pathway is preferred as it has the advantage over the deamination that one extra methyl group is generated and can be used for growth [121].
The second product of the upper pathway, 2,6-dihydroxypyridine is further hydroxylated by a 2,6-dihydroxypyridine 3-monooxygenase (2,3-Dihydroxypyridine 3-hydroxylase, 2,6-DHPH, EC 1.14.13.10) a FAD-containing protein with known structure [134]. The product of the reaction, 2,3,6-trihydroxypyridine, can oxidatively dimerize to form nicotine blue which accumulates in the growth medium.
In terms of transcriptional factors involved in the regulation of this pathway, two major regulators have been described. One is PmfR, an activator of an operon containing the mabO, maO, and ssaDH genes [135,136]. Although the recognition sequence for PmfR has been described, its effector molecule that modulates its binding to DNA is still unknown. The second factor is the HdnoR repressor sensible to nicotine and other nicotine derivatives and controlling the expression of the 6hdno gene [137].
4.1.2. The VPP pathway for nicotine degradation
Phylogenetic analysis showed that the VPP pathway is more closely related to the pyrrolidine pathway, and both are found predominantly in Gram-negative bacteria [138]. The strains that harbor the VPP catabolic pathway are mostly Alphaproteobacteria such as Ochrobactrum sp. SJY1 [139], Shinella sp. HZN7 [140,141], Sphingomonas melonis TY [138], Agrobacterium tumefaciens S33 [142] or Gammaproteobacteria, such as Pseudomonas geniculate N1 [143]. Albeit the pathway is well characterized in most of the mentioned strains, the available complete genome and transcriptome of A. tumefaciens S33 allowed the identification of most of the genes involved in various steps of VPP in this microorganism [144].
As in the pyridine pathway, the VPP pathway also debuts the hydroxylation of NIC with the formation of 6-hydroxy-nicotine. The reaction is catalyzed by, nicotine:acceptor 6-oxidoreductase (hydroxylating) (Ndo, nicotine hydroxylase, EC 1.5.99.4), a member of the molybdopterin enzymes family. Unlike the P. nicotinovorans ATCC 49919 counterpart, Ndo is known to form a complex [142] with 6-hydroxypseudooxynicotine oxidase (Pno, EC 1.5.99.14), a flavoprotein also containing a 4Fe/4S [145]. Albeit 6LHNO activity has been detected when nicotine is present in the growth media of A. tumefaciens S33, the pure enzyme has not been yet isolated [146]. The gene has nevertheless been knocked out [147] and a similar enzyme has been isolated from another strain harboring the same VPP pathway [148].
The catalytic products of Pno are methylamine and 6-hydroxy-3-succinoylsemialdehyde pyridine, the latter compound marking the point where the similarities with the pyridine pathway ends, and the pyrrolidine pathway starts. The semialdehyde is converted into 6-hydroxy-3-succinoyl pyridine by an aldehyde dehydrogenase that has not been yet experimentally isolated [144], but putative gene sharing 32% identity to an NADP+-dependent 3-succinoylsemialdehyde-pyridne dehydrogenase from Pseudomonas sp. HZN6 has been identified in the A. tumefaciens S33 genome [149]. The resulting pyridine derivate is further processed by a specific hydroxylase (Hsh, 6-hydroxy-3-succinoylpyridine 3-monooxygenase, EC 1.14.13.163) containing FAD which performs an oxidative decarboxylation and converts it in a NADH-dependent manner to 2,5-dihydroxypyridine and succinate. 2,5-dihydroxypyridine ring is opened in a Fe (II)-dependent manner by a 2,5-dihydroxypyridine 5,6-dioxygenase (Hpo, EC 1.13.11.9).
In the final steps of the pathway, N-formylmaleamic acid is then converted into maleamic and formic acid by N-formylmaleamate deformylase (Nfo, EC 3.5.1.106). A maleamate amidohydrolase (Ami, EC 3.5.1.107) further converts the maleamic acid into maleic acid and ammonia. Finally, maleate cis/trans-isomerase (Iso, EC 5.2.1.1) catalyzes the isomerization of maleic acid into fumaric acid, which is channeled to the Krebs cycle for further catabolism.
4.2. Applications of NDB for 6-hydroxy-L-nicotine production from nicotine-containing waste
NDBs and their NIC-degrading pathways are a promising solution for a neglected environmental problem. The tobacco industry responsible for the massive manufacture of cigars, cigarettes, snuff, chewing tobacco and other tobacco products including some e-juice formulations produces large amounts of powdery solid or liquid wastes containing high concentrations of NIC [150]. This waste is released into the environment leading to contamination of the groundwater with NIC [151] and causing a serious threat not only to the ecological balance of soil and water but also to human health [152,153]. Although physical and chemical methods to degrade NIC from tobacco are available, these are expensive and still involve the use of harmful solvents [154]. Bioremediation and bioconversion of the toxic NIC-containing wastes using NDB are environmentally friendly strategies for either decontamination [155,156,157,158,159,160,161,162] or, even more appealing, for converting the waste into green chemicals.
The proof of concept has been demonstrated by the production of 6-hydroxy-3-succinoyl-pyridine. A biotransformation technology using Pseudomonas sp. has been used to produce 1.45 g/L 6-hydroxy-3-succinoyl-pyridine. from 3 g/L of NIC in 5 h with 3.4 g/L of cells in a 5-L, with an overall yield of 43.8% (w/w). As the reaction was done in water, HSP could be easily purified from the reaction without the need for extensive purification steps [163]. In a further development of the technology, a genetically engineered strain of P. putida S16 was employed, and a 3.7-fold higher accumulation of 6-hydroxy-3-succinoyl-pyridine was observed. The recombinant strain had the equivalent hsh gene deleted by homologous recombination and was able to convert both NIC as well as a crude tobacco-waste extract (~8.7% (w/v) NIC) obtained by steam distillation [164].
Another green chemical that can be obtained from NIC-containing waste by using NDBs is 3-succinoyl-pyridine. The biotransformation reaction also makes use of the same P. putida S16 strain and is again genetically engineered to accumulate 3-succinoyl-pyridine by deleting specific genes. Aqueous NIC solution and a crude suspension of the tobacco waste were both successfully used and a maximum yield of 54.2% was obtained [165].
Such biotransformation technologies are also available for 6HLN. The strains of choice this time are A. tumefaciens S33 and P. nicotinovorans ATCC 49919. In both strains, 6HLN is an intermediate that accumulates temporally in the growth media as it is converted by 6HLNO to 6-hydroxy-N-methylmyosmine / 6-hydroxypseudooxynicotine. To achieve higher 6HLN concentrations, two different approaches have been employed. In P. nicotinovorans ATCC 49919, the NDH enzyme was overexpressed and the 6HLNO activity was inhibited by using metal ions resulting in a 5-fold accumulation of the product compared with the wild-type strain [166]. In A. tumefaciens S33 the 6hlno gene was knocked out by using homologous recombination and the molar conversion reached approximately 98%.
4.3. The behavioral effects of 6-hydroxy-L-nicotine
Previous studies have shown that 6HLN is a neurologically active substance, stimulating higher cognitive functions such as memory and learning and improving the mood, by reducing the anxiety- and depressive-like state (Table 1). Hritcu et al. [167] have shown that chronic administration of 6HLN (0.3 mg/kg, b.w., i.p. for 7 consecutive days) to normal Wistar rats improves locomotor activity and spatial memory, especially the short-term and working memory, without affecting the long-term memory in specific hippocampal-dependent assays, such as Y-maze and radial arm-maze (RAM) tasks. Due to these cognitive-improving effects, 6HLN was further tested for the ability to mitigate the cognitive and non-cognitive symptoms of AD condition using different animal models. In 2015, Hritcu et al. [168] administered 6HLN (0.3 mg/kg, b.w., i.p. with 30 min before testing) to a rat model of AD induced by scopolamine (SCOP, 0.7 mg/kg, b.w., i.p., with 24 h before testing), a competitive antagonist of muscarinic acetylcholine receptors (mAChRs). It has been shown that 6HLN improves the spatial working memory in the Y-maze task and enhances the working and reference memory in the RAM task of rats with SCOP-induced memory deficits. In other studies, performed on rats [169,170], 6HLN (0.3 mg/kg, b.w., i.p.) was compared to NIC (0.3 mg/kg, b.w., i.p.) in terms of its ability to alleviate the memory impairment and anxious and [171] depressive behavior induced by chlorisondamine (CHL, 10 mg/kg, b.w., i.p.), a nAChRs antagonist. It was found that both 6HLN and NIC alone or in combination with CHL enhance memory in specific behavioral paradigms such as Y-maze and RAM and mitigate anxiety and depression in the elevated plus maze test and Porsolt’s forced swimming test respectively. In a study performed by Boiangiu et al. [171], 6HLN and its precursor were chronically administered (0.3 and 0.7 mg/kg, b.w., i.p. for 33 days) to a rat model of AD induced by brain infusion of amyloid beta peptide fragment 25-35 (Aβ25-35) and the memory performances were assessed in Y-maze, RAM, and novel object recognition (NOR) tasks. Both compounds exhibited a promnesic effect, ameliorating the Aβ25-35-induced impairment of spatial recognition memory, working and reference memory, and recognition memory. Also, 6HLN administration positively regulated the Bdnf, Arc, and IL-1β gene expression in the Aβ25-35-treated rat's hippocampus. Therefore, 6HLN might support neuroprotection, increase synaptic plasticity and memory consolidation, and diminish neuroinflammation leading thus to the cognitive-enhancing effects noticed in behavioral tasks [171]. Recently, the effects of 6HLN on cognition and anxiety-like behavior were investigated in a zebrafish (Danio rerio) model of AD induced by immersion in SCOP [172]. It has been shown that acute treatment with 6HLN (1 and 2 mg/L, for 3 min) to zebrafish pre-treated with SCOP (100 µM, for 30 min) improves spatial memory and recognition memory in Y-maze task and NOR task respectively and reduces the anxiety in the novel tank diving test (NTT). Consistent with the results obtained from the studies performed on rats, 6HLN upregulated the expression of Bdnf, Npy, and Egr1 genes in the SCOP-treated zebrafish brain, thus indicating the supportive role of 6HLN in neuroprotection, cognition, memory, and learning. Taken together, all these in vivo studies suggest that 6HLN is a neurologically active molecule that could be therapeutically used to improve cognition and mood in healthy or AD-diseased individuals.
4.4. Effects of 6-hydroxy-L-nicotine on acetylcholinesterase activity
ACh is one of the most important neurotransmitters in the central cholinergic system, originating in the basal forebrain from both the basal nucleus of Meynert and the medial septal area. It binds specifically to mAChRs and nAChRs and is critically involved in learning and memory. Acetylcholinesterase (AChE) is a serine-hydrolase that stops the ACh action by hydrolyzing it in acetate ions and choline and represents a marker of cholinergic neuron loss in the brain region [173,174,175,176]. The cholinergic hypothesis of AD states that the reduction of ACh synthesis is the main cause of the disease [177]. Therefore, the ACh level elevation in the brain by inhibiting the AChE biological activity represents a therapeutic approach to AD [178]. Previous in vivo studies investigated the effect of 6HLN on AChE activity in the brain of animal models of AD. Thus, it has been shown that chronic treatment with 6HLN decreases the AChE activity in the hippocampus of Aβ25-35-treated rats[171]. Similarly, the acute administration of 6HLN to SCOP-treated zebrafish clearly inhibited the AChE activity in the brain [172]. This anti-AChE profile of 6HLN could increase the bioavailability of ACh in the brain and ameliorate the AD-like condition.
4.5. Effects of 6-hydroxy-L-nicotine on oxidative stress
The antioxidant properties of 6HLN and NIC were compared initially using computational methods. Quantitative Structure-Activity Relationship (QSAR) modeling suggested that 6HLN could be a better antioxidant than NIC [179]. This comparison was also performed in vitro using the Ferric Reducing Ability of Plasma (FRAP) assay [180]. Both NIC and 6HLN were tested for their ability to inhibit the formation of Fe2+ from Fe3+ at three different concentrations: 15, 30, and 45 mM. It was found that the antioxidant potential of 6HLN was better compared to NIC. This is consistent with the fact that the antioxidant activity associated with the compound's chemical structure is dependent on the number of active hydroxyl and amino groups included. A compound is more active if it has more active groups in the molecule [180], and 6HLN has an additional hydroxyl group compared to NIC.
The antioxidant effects of 6HLN were demonstrated in vivo by Hritcu et al. [167]. They evaluated the activity of antioxidant defense enzymes, such as superoxide dismutase (SOD) and glutathione peroxidase (GPX), and measured the level of malondialdehyde (MDA), the major lipid peroxidation product in the brain of male Wistar rats. Chronic administration of 6HLN significantly increased the specific activity of SOD and GPX enzymes and decreased the MDA level in the temporal cortex of the animals [167]. Also, it was demonstrated that 6HLN increases the SOD, GPX, and catalase (CAT) specific activities, reduces the MDA level, and increases the reduced glutathione (GSH) content in the hippocampal homogenates of SCOP- or CHL-treated rats, suggesting thus that 6HLN could be a potent compound with potential applications in AD therapy [168,169]. Boiangiu et al. [171] assessed the antioxidant potential of chronic administration of 6HLN in the hippocampus of a rat model of AD induced by intracerebroventricular infusion of Aβ25-35 peptide. Their findings showed that 6HLN reduces the Aβ25-35-induced oxidative stress by increasing the SOD, CAT, and GPX activity and GSH content and lowering the MDA and carbonylated proteins levels in the rat hippocampus. Recently, the impact of 6HLN on brain oxidative status was evaluated in a zebrafish model of AD induced by immersion in SCOP [172]. Consistent with the results obtained on rats, 6HLN clearly reduced the SCOP-induced oxidative stress in the zebrafish brain by intensifying the SOD, CAT, and GPX activities, increasing the GSH content, and reducing the MDA and carbonylated proteins levels. Additionally, it was found that 6HLN positively regulates the Nrf2a gene expression in the brain of the zebrafish. This gene encodes a transcription factor that regulates several antioxidants and cytoprotective genes involved in the protection against reactive oxygen species cytotoxicity [181]. Therefore, the improvement of brain oxidative status by 6HLN noticed in the in vivo studies might involve specific gene expression.
4.6. The proposed mechanism of action for 6-hydroxy-L-nicotine
To date, there is limited amount of experimental data to explain the mechanism underlying the neurobiological effects of 6HLN. However, due to the structural resemblance towards NIC, its precursor molecule, we consider that 6HLN might exert its effects on the CNS in a similar manner to that of NIC.
Both clinical and animal studies support the role of nAChRs in memory, learning, and cognition. The nAChRs are transmembrane pentameric ligand-gated ion channels that are present in both CNS and PNS. These receptors are composed by combining different α and β subunits leading to different pharmacological and kinetic properties of the receptor [182,183,184]. The α7 and α4β2 subtypes of nAChRs are present in the hippocampus and involved in memory formation and cognitive function and are also affected in AD [182]. NIC is the prototypic agonist of nAChRs and could improve cognition by interacting with the presynaptic nAChRs facilitating the release of the neurotransmitters involved in memory and learning processes, such as ACh, glutamate, dopamine, norepinephrine, serotonin, and GABA [185]. Previous in silico studies have shown that 6HLN could interact with nAChR receptors. Using molecular docking experiments, Mihasan et al. [179] and Boiangiu et al. [171] assessed the potential of 6HLN to bind to α4β2 nAChRs (3α:2β stoichiometry) and Acetylcholine Binding Protein (AChBP), a protein produced by glial cells of Lymnaea stagnalis and which possess the pharmacological properties of α7 nAChRs. At that time, the AChBP structure was used as a 3D model to provide details regarding the potential mechanism of action of various α7 nAChRs ligands. It has been suggested that 6HLN might bind to α4β2 nAChRs and AChBP with similar or higher affinity compared to NIC, due to an extra hydrogen bond formed between the hydroxyl group of 6HLN and the Y204 residue of the binding cavity of α4β2 nAChRs or M114 residue of AChBP respectively. Also, the comparative analysis of the theoretical binding energies of 6HLN and NIC to α4β2 nAChRs revealed that both compounds might show an affinity towards the binding site located at the α-β interface instead of α-α interface [171].
In vivo studies performed on rats and zebrafish showed that 6HLN exposure improves memory, learning cognition, and mood, and decreases brain oxidative stress. In addition, these phenotypes were accompanied by specific gene expressions. Thus, the upregulation of the Bdnf, Arc, Egr1, and Npy gene expression could explain the cognitive improvement, while the upregulation of the Nrf2a gene could explain the antioxidant properties of 6HLN in the brain of the animals [171,172]. Taking into consideration the available data, we believe that nAChRs positive modulation by 6HLN could trigger a downstream cellular pathway which may lead to specific gene expression and the occurrence of the discussed biological effects.
5. Conclusion
While the primary source of NIC for humans is tobacco, some bacteria can metabolize NIC or produce various derivatives in the process. The relationship between NIC derivatives from bacteria and cognition, oxidative stress, and cholinergic system activity is an interesting and complex topic. Conversely, there is corroborating evidence indicating that 6HLN, a NIC metabolite from bacteria, displays pharmacological activity in the brains of animal models, all the while avoiding the manifestation of adverse effects. Taken together, we contend that this biologically active metabolite holds promise for therapeutic applications in alleviating the symptoms associated with dementia conditions.
Author Contributions
Conceptualization, H.L., M.M., and R.S.B.; methodology, M.M., R.S.B., I.B., I.H., and H.L.; formal analysis, M.M., R.S.B., I.B., I.H., and H.L.; investigation, H.L., M.M., and R.S.B.; resources, H.L.; data curation, M.M., H.L., and R.S.B.; writing—original draft preparation, H.L., M.M., and R.S.B.; writing—review and editing, H.L., M.M., and R.S.B.; supervision, H.L. and M.M.; funding acquisition, H.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant of the Ministry of Research, Innovation and Digitization, CNCS-UEFISCDI, project number PN-III-P4-PCE-2021-1692, within PNCDI III.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Picciotto, M.R.; Kenny, P.J. Mechanisms of Nicotine Addiction. Cold Spring Harb Perspect Med 2021, 11. [Google Scholar] [CrossRef]
- Newhouse, P.; Kellar, K.; Aisen, P.; White, H.; Wesnes, K.; Coderre, E.; Pfaff, A.; Wilkins, H.; Howard, D.; Levin, E.D. Nicotine Treatment of Mild Cognitive Impairment: A 6-Month Double-Blind Pilot Clinical Trial. Neurology 2012, 78. [Google Scholar] [CrossRef]
- Ruan, Y.; Xie, Z.; Liu, Q.; Zhang, L.; Han, X.; Liao, X.; Liu, J.; Gao, F. Nicotine and Menthol Independently Exert Neuroprotective Effects against Cisplatin- or Amyloid- Toxicity by Upregulating Bcl-Xl via JNK Activation in SH-SY5Y Cells. Biocell 2021, 45. [Google Scholar] [CrossRef]
- Xue, M.Q.; Liu, X.X.; Zhang, Y.L.; Gao, F.G. Nicotine Exerts Neuroprotective Effects against β-Amyloid-Induced Neurotoxicity in SH-SY5Y Cells through the Erk1/2-P38-JNK-Dependent Signaling Pathway. Int J Mol Med 2014, 33. [Google Scholar] [CrossRef]
- Bao, Z.; He, X.Y.; Ding, X.; Prabhu, S.; Hong, J.Y. Metabolism of Nicotine and Cotinine by Human Cytochrome P450 2A13. Drug Metabolism and Disposition 2005, 33. [Google Scholar] [CrossRef]
- Sadigh-Eteghad, S.; Vatandoust, S.M.; Mahmoudi, J.; Rahigh Aghsan, S.; Majdi, A. Cotinine Ameliorates Memory and Learning Impairment in Senescent Mice. Brain Res Bull 2020. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, V.; Zeitlin, R.; Burgess, S.; Patel, S.; Barman, A.; Thakur, G.; Mamcarz, M.; Wang, L.; Sattelle, D.B.; Kirschner, D.A.; et al. Cotinine Reduces Amyloid-β Aggregation and Improves Memory in Alzheimer’s Disease Mice. Journal of Alzheimer’s Disease 2011, 24, 817–835. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Adam, B.L.; Terry, A. V. Evaluation of Nicotine and Cotinine Analogs as Potential Neuroprotective Agents for Alzheimer’s Disease. Bioorg Med Chem Lett 2014. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372. [Google Scholar] [CrossRef]
- Benowitz, N.L.; Hukkanen, J.; Jacob, P. Nicotine Chemistry, Metabolism, Kinetics and Biomarkers. Handb Exp Pharmacol 2009, 192, 29–60. [Google Scholar] [CrossRef]
- Gurusamy, R.; Natarajan, S. Current Status on Biochemistry and Molecular Biology of Microbial Degradation of Nicotine. The Scientific World Journal 2013, 2013. [Google Scholar] [CrossRef]
- Metz, C.N.; Gregersen, P.K.; Malhotra, A.K. Metabolism and Biochemical Effects of Nicotine for Primary Care Providers. Medical Clinics of North America 2004, 88, 1399–1413. [Google Scholar] [CrossRef]
- Lockman, P.R.; McAfee, G.; Geldenhuys, W.J.; Van Der Schyf, C.J.; Abbruscato, T.J.; Allen, D.D. Brain Uptake Kinetics of Nicotine and Cotinine after Chronic Nicotine Exposure. Journal of Pharmacology and Experimental Therapeutics 2005, 314, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Matta, S.G.; Balfour, D.J.; Benowitz, N.L.; Boyd, R.T.; Buccafusco, J.J.; Caggiula, A.R.; Craig, C.R.; Collins, A.C.; Damaj, M.I.; Donny, E.C.; et al. Guidelines on Nicotine Dose Selection for in Vivo Research. Psychopharmacology (Berl) 2007, 190, 269–319. [Google Scholar] [CrossRef]
- Valentine, G.; Sofuoglu, M. Cognitive Effects of Nicotine: Recent Progress. Curr Neuropharmacol 2017, 15, 403. [Google Scholar] [CrossRef]
- Shu, H.; Wang, M.; Song, M.; Sun, Y.; Shen, X.; Zhang, J.; Jin, X. Acute Nicotine Treatment Alleviates LPS-Induced Impairment of Fear Memory Reconsolidation through AMPK Activation and CRTC1 Upregulation in Hippocampus. International Journal of Neuropsychopharmacology 2021, 23, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Grus, A.; Hromatko, I. Acute Administration of Nicotine Does Not Enhance Cognitive Functions. Arh Hig Rada Toksikol 2019, 70, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Belluzzi, J.D.; Wang, R.; Leslie, F.M. Acetaldehyde Enhances Acquisition of Nicotine Self-Administration in Adolescent Rats. Neuropsychopharmacology 2005, 30, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Poltavski, D. V.; Petros, T. V.; Holm, J.E. Lower but Not Higher Doses of Transdermal Nicotine Facilitate Cognitive Performance in Smokers on Gender Non-Preferred Tasks. Pharmacol Biochem Behav 2012, 102, 423–433. [Google Scholar] [CrossRef]
- Newhouse, P.A.; Potter, A.; Singh, A. Effects of Nicotinic Stimulation on Cognitive Performance. Curr. Opin. Pharmacol. 2004, 4, 36–46. [Google Scholar] [CrossRef]
- Grundey, J.; Amu, R.; Ambrus, G.G.; Batsikadze, G.; Paulus, W.; Nitsche, M.A. Double Dissociation of Working Memory and Attentional Processes in Smokers and Non-Smokers with and without Nicotine. Psychopharmacology (Berl) 2015, 232, 2491–2501. [Google Scholar] [CrossRef] [PubMed]
- Potter, A.S.; Newhouse, P.A. Acute Nicotine Improves Cognitive Deficits in Young Adults with Attention-Deficit/Hyperactivity Disorder. Pharmacol Biochem Behav 2008, 88, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.A.; Banks, W.A. Age-Associated Changes in the Immune System and Blood–Brain Barrier Functions. Int J Mol Sci 2019, 20, 1632. [Google Scholar] [CrossRef] [PubMed]
- Shytle, R.D.; Mori, T.; Townsend, K.; Vendrame, M.; Sun, N.; Zeng, J.; Ehrhart, J.; Silver, A.A.; Sanberg, P.R.; Tan, J. Cholinergic Modulation of Microglial Activation by A7 Nicotinic Receptors. J Neurochem 2004, 89, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Zenaro, E.; Pietronigro, E.; Bianca, V. Della; Piacentino, G.; Marongiu, L.; Budui, S.; Turano, E.; Rossi, B.; Angiari, S.; Dusi, S.; et al. Neutrophils Promote Alzheimer’s Disease-like Pathology and Cognitive Decline via LFA-1 Integrin. Nat Med 2015, 21, 880–886. [Google Scholar] [CrossRef]
- Majdi, A.; Kamari, F.; Vafaee, M.S.; Sadigh-Eteghad, S. Revisiting Nicotine’s Role in the Ageing Brain and Cognitive Impairment. Rev Neurosci 2017, 28, 767–781. [Google Scholar] [CrossRef]
- Hao, J.; Simard, A.R.; Turner, G.H.; Wu, J.; Whiteaker, P.; Lukas, R.J.; Shi, F.D. Attenuation of CNS Inflammatory Responses by Nicotine Involves A7 and Non-A7 Nicotinic Receptors. Exp Neurol 2011, 227, 110–119. [Google Scholar] [CrossRef]
- Wei, P.; Liu, Q.; Li, D.; Zheng, Q.; Zhou, J.; Li, J. Acute Nicotine Treatment Attenuates Lipopolysaccharide-Induced Cognitive Dysfunction by Increasing BDNF Expression and Inhibiting Neuroinflammation in the Rat Hippocampus. Neurosci Lett 2015, 604, 161–166. [Google Scholar] [CrossRef]
- Sadigh-Eteghad, S.; Majdi, A.; Mahmoudi, J.; Golzari, S.E.J.; Talebi, M. Astrocytic and Microglial Nicotinic Acetylcholine Receptors: An Overlooked Issue in Alzheimer’s Disease. J Neural Transm 2016, 123, 1359–1367. [Google Scholar] [CrossRef]
- Han, Y.; Lau, Y.-L. Nicotine, an Anti-Inflammation Molecule. Inflamm Cell Signal 2014, 1. [Google Scholar] [CrossRef]
- Wang, H.; Yu, M.; Ochani, M.; Amelia, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, N.; Ulloa, L.; et al. Nicotinic Acetylcholine Receptor A7 Subunit Is an Essential Regulator of Inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef]
- De Simone, R.; Ajmone-Cat, M.A.; Carnevale, D.; Minghetti, L. Activation of A7 Nicotinic Acetylcholine Receptor by Nicotine Selectively Up-Regulates Cyclooxygenase-2 and Prostaglandin E2 in Rat Microglial Cultures. J Neuroinflammation 2005, 2. [Google Scholar] [CrossRef]
- Razani-Boroujerdi, S.; Boyd, R.T.; Dávila-García, M.I.; Nandi, J.S.; Mishra, N.C.; Singh, S.P.; Pena-Philippides, J.C.; Langley, R.; Sopori, M.L. T Cells Express A7-Nicotinic Acetylcholine Receptor Subunits That Require a Functional TCR and Leukocyte-Specific Protein Tyrosine Kinase for Nicotine-Induced Ca2+ Response. The Journal of Immunology 2007, 179, 2889–2898. [Google Scholar] [CrossRef]
- Nizri, E.; Irony-Tur-Sinai, M.; Lory, O.; Orr-Urtreger, A.; Lavi, E.; Brenner, T. Activation of the Cholinergic Anti-Inflammatory System by Nicotine Attenuates Neuroinflammation via Suppression of Th1 and Th17 Responses. J Immunol 2009, 183, 6681–6688. [Google Scholar] [CrossRef]
- Tyagi, E.; Agrawal, R.; Nath, C.; Shukla, R. Cholinergic Protection via A7 Nicotinic Acetylcholine Receptors and PI3K-Akt Pathway in LPS-Induced Neuroinflammation. Neurochem Int 2010, 56, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T. Apoptosis and Its Functional Significance in Molluscs. Apoptosis 2010, 15, 313–321. [Google Scholar] [CrossRef]
- Majdi, A.; Mahmoudi, J.; Sadigh-Eteghad, S.; Golzari, S.E.J.; Sabermarouf, B.; Reyhani-Rad, S. Permissive Role of Cytosolic PH Acidification in Neurodegeneration: A Closer Look at Its Causes and Consequences. J Neurosci Res 2016, 94, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Tizabi, Y.; Manaye, K.F.; Taylor, R.E. Nicotine Blocks Ethanol-Induced Apoptosis in Primary Cultures of Rat Cerebral Cortical and Cerebellar Granule Cells. Neurotox Res 2005, 7, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Mechawar, N.; Krantic, S.; Quirion, R. A7 Nicotinic Receptor Activation Reduces β-Amyloid-Induced Apoptosis by Inhibiting Caspase-Independent Death through Phosphatidylinositol 3-Kinase Signaling. J Neurochem 2011, 119, 848–858. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, B. Nicotine Attenuates Beta-Amyloid Peptide-Induced Neurotoxicity, Free Radical and Calcium Accumulation in Hippocampal Neuronal Cultures. Br J Pharmacol. 2004, 141, 746–754. [Google Scholar] [CrossRef]
- Hejmadi, M. V.; Dajas-Bailador, F.; Barns, S.M.; Jones, B.; Wonnacott, S. Neuroprotection by Nicotine against Hypoxia-Induced Apoptosis in Cortical Cultures Involves Activation of Multiple Nicotinic Acetylcholine Receptor Subtypes. Molecular and Cellular Neuroscience 2003, 24, 779–786. [Google Scholar] [CrossRef]
- Hritcu, L.; Ciobica, A.; Gorgan, L. Nicotine-Induced Memory Impairment by Increasing Brain Oxidative Stress. Cent Eur J Biol 2009, 4. [Google Scholar] [CrossRef]
- Jang, M.H.; Shin, M.C.; Jung, S.B.; Lee, T.H.; Bahn, G.H.; Kwon, Y.K.; Kim, E.H.; Kim, C.J. Alcohol and Nicotine Reduce Cell Proliferation and Enhance Apoptosis in Dentate Gyrus. Neuroreport 2002, 13, 1509–1513. [Google Scholar] [CrossRef] [PubMed]
- Garrido, R.; Mattson, M.P.; Hennig, B.; Toborek, M. Nicotine Protects against Arachidonic-Acid-Induced Caspase Activation, Cytochrome c Release and Apoptosis of Cultured Spinal Cord Neurons. J Neurochem 2001, 76, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Marrero, M.B.; Bencherif, M. Convergence of Alpha 7 Nicotinic Acetylcholine Receptor-Activated Pathways for Anti-Apoptosis and Anti-Inflammation: Central Role for JAK2 Activation of STAT3 and NF-ΚB. Brain Res 2009, 1256, 1–7. [Google Scholar] [CrossRef]
- De Azevedo Cardoso, T.; Mondin, T.C.; Wiener, C.D.; Marques, M.B.; Fucolo, B.D.Á.; Pinheiro, R.T.; De Souza, L.D.M.; Da Silva, R.A.; Jansen, K.; Oses, J.P. Neurotrophic Factors, Clinical Features and Gender Differences in Depression. Neurochem Res 2014, 39, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Erraji-Benchekroun, L.; Underwood, M.D.; Arango, V.; Galfalvy, H.; Pavlidis, P.; Smyrniotopoulos, P.; Mann, J.J.; Sibille, E. Molecular Aging in Human Prefrontal Cortex Is Selective and Continuous throughout Adult Life. Biol Psychiatry 2005, 57, 549–558. [Google Scholar] [CrossRef]
- Pourmemar, E.; Majdi, A.; Haramshahi, M.; Talebi, M.; Karimi, P.; Sadigh-Eteghad, S. Intranasal Cerebrolysin Attenuates Learning and Memory Impairments in D-Galactose-Induced Senescence in Mice. Exp Gerontol 2017, 87, 16–22. [Google Scholar] [CrossRef]
- Ferrea, S.; Winterer, C. Neuroprotective and Neurotoxic Effects of Nicotine. Pharmacopsychiatry 2009, 42, 255–265. [Google Scholar] [CrossRef]
- Wongtrakool, C.; Grooms, K.; Bijli, K.M.; Crothers, K.; Fitzpatrick, A.M.; Hart, C.M. Nicotine Stimulates Nerve Growth Factor in Lung Fibroblasts through an NFκB-Dependent Mechanism. PLoS One 2014, 9, e109602. [Google Scholar] [CrossRef]
- Garrido, R.; King-Pospisil, K.; Son, K.W.; Hennig, B.; Toborek, M. Nicotine Upregulates Nerve Growth Factor Expression and Prevents Apoptosis of Cultured Spinal Cord Neurons. Neurosci Res 2003, 47, 349–355. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, R.; Toledano, A.; Álvarez, M.I.; Turégano, L.; Colman, O.; Rosés, P.; Gómez de Segura, I.; De Miguel, E. Chronic Nicotine Administration Increases NGF-like Immunoreactivity in Frontoparietal Cerebral Cortex. J Neurosci Res 2003, 73, 708–716. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, R.; Garcia, A.A.; Braschi, C.; Capsoni, S.; Maffei, L.; Berardi, N.; Cattaneo, A. Intranasal Administration of Nerve Growth Factor (NGF) Rescues Recognition Memory Deficits in AD11 Anti-NGF Transgenic Mice. Proc Natl Acad Sci U S A 2005, 102, 3811–3816. [Google Scholar] [CrossRef] [PubMed]
- Czubak, A.; Nowakowska, E.; Kus, K.; Burda, K.; Metelska, J.; Baer-Dubowska, W.; Cichocki, M. Influences of Chronic Venlafaxine, Olanzapine and Nicotine on the Hippocampal and Cortical Concentrations of Brain-Derived Neurotrophic Factor (BDNF). Pharmacol Rep 2009, 61, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Freedman, R.; Wetmore, C.; Stromberg, I.; Leonard, S.; Olson, L. α-Bungarotoxin Binding to Hippocampal Interneurons: Immunocytochemical Characterization and Effects on Growth Factor Expression. Journal of Neuroscience 1993, 13, 1965–1975. [Google Scholar] [CrossRef]
- Yamada, K.; Mizuno, M.; Nabeshima, T. Role for Brain-Derived Neurotrophic Factor in Learning and Memory. Life Sci 2002, 70, 735–744. [Google Scholar] [CrossRef]
- Rodrigue, K.M.; Kennedy, K.M.; Devous, M.D.; Rieck, J.R.; Hebrank, A.C.; Diaz-Arrastia, R.; Mathews, D.; Park, D.C. β-Amyloid Burden in Healthy Aging: Regional Distribution and Cognitive Consequences. Neurology 2012, 78, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Zahs, K.R.; Ashe, K.H. β-Amyloid Oligomers in Aging and Alzheimer’s Disease. Front Aging Neurosci 2013, 5, 51139. [Google Scholar] [CrossRef]
- Banks, W.A.; Kumar, V.B.; Farr, S.A.; Nakaoke, R.; Robinson, S.M.; Morley, J.E. Impairments in Brain-to-Blood Transport of Amyloid-β and Reabsorption of Cerebrospinal Fluid in an Animal Model of Alzheimer’s Disease Are Reversed by Antisense Directed against Amyloid-β Protein Precursor. Journal of Alzheimer’s Disease 2011, 23, 599–605. [Google Scholar] [CrossRef]
- Holmes, C.; Boche, D.; Wilkinson, D.; Yadegarfar, G.; Hopkins, V.; Bayer, A.; Jones, R.W.; Bullock, R.; Love, S.; Neal, J.W.; et al. Long-Term Effects of Aβ42 Immunisation in Alzheimer’s Disease: Follow-up of a Randomised, Placebo-Controlled Phase I Trial. The Lancet 2008, 372, 216–223. [Google Scholar] [CrossRef]
- Hellström-Lindahl, E.; Court, J.; Keverne, J.; Svedberg, M.; Lee, M.; Marutle, A.; Thomas, A.; Perry, E.; Bednar, I.; Nordberg, A. Nicotine Reduces Aβ in the Brain and Cerebral Vessels of APPsw Mice. European Journal of Neuroscience 2004, 19, 2703–2710. [Google Scholar] [CrossRef]
- Utsuki, T.; Shoaib, M.; Holloway, H.W.; Ingram, D.K.; Wallace, W.C.; Haroutunian, V.; Sambamurti, K.; Lahiri, D.K.; Greig, N.H. Nicotine Lowers the Secretion of the Alzheimer’s Amyloid β-Protein Precursor That Contains Amyloid β-Peptide in Rat. Journal of Alzheimer’s Disease 2002, 4, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, A.; Hellström-Lindahl, E.; Lee, M.; Johnson, M.; Mousavi, M.; Hall, R.; Perry, E.; Bednar, I.; Court, J. Chronic Nicotine Treatment Reduces β-Amyloidosis in the Brain of a Mouse Model of Alzheimer’s Disease (APPsw). J Neurochem 2002, 81, 655–658. [Google Scholar] [CrossRef]
- Ono, K.; Hasegawa, K.; Yamada, M.; Naiki, H. Nicotine Breaks down Preformed Alzheimer’s β-Amyloid Fibrils in Vitro. Biol Psychiatry 2002, 52, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Dineley, K.T.; Westerman, M.; Bui, D.; Bell, K.; Ashe, K.H.; Sweatt, J.D. β-Amyloid Activates the Mitogen-Activated Protein Kinase Cascade via Hippocampal A7 Nicotinic Acetylcholine Receptors: In Vitro and in Vivo Mechanisms Related to Alzheimer’s Disease. Journal of Neuroscience 2001, 21, 4125–4133. [Google Scholar] [CrossRef] [PubMed]
- Sadigh-Eteghad, S.; Talebi, M.; Farhoudi, M.; Golzari, S.E.J.; Sabermarouf, B.; Mahmoudi, J. Beta-Amyloid Exhibits Antagonistic Effects on Alpha 7 Nicotinic Acetylcholine Receptors in Orchestrated Manner. Journal of Medical Hypotheses and Ideas 2014, 8, 49–52. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Godoy, J.A.; Vargas, J.Y.; Arrazola, M.S.; Rios, J.A.; Carvajal, F.J.; Serrano, F.G.; Farias, G.G. Nicotine Prevents Synaptic Impairment Induced by Amyloid-β Oligomers through A7-Nicotinic Acetylcholine Receptor Activation. Neuromolecular Med 2013, 15, 549–569. [Google Scholar] [CrossRef] [PubMed]
- Majdi, A.; Kamari, F.; Vafaee, M.S.; Sadigh-Eteghad, S. Revisiting Nicotine’s Role in the Ageing Brain and Cognitive Impairment. Rev Neurosci 2017, 28, 767–781. [Google Scholar] [CrossRef]
- Ward, R.J.; Zucca, F.A.; Duyn, J.H.; Crichton, R.R.; Zecca, L. The Role of Iron in Brain Ageing and Neurodegenerative Disorders. Lancet Neurol 2014, 13, 1045. [Google Scholar] [CrossRef]
- Daugherty, A.M.; Raz, N. Appraising the Role of Iron in Brain Aging and Cognition: Promises and Limitations of MRI Methods. Neuropsychol Rev 2015, 25, 272–287. [Google Scholar] [CrossRef]
- Haddadi, M.; Jahromi, S.R.; Sagar, B.K.C.; Patil, R.K.; Shivanandappa, T.; Ramesh, S.R. Brain Aging, Memory Impairment and Oxidative Stress: A Study in Drosophila Melanogaster. Behavioural Brain Research 2014, 259, 60–69. [Google Scholar] [CrossRef]
- GuanZ. Z.; YuW.F.; NordbergA.; Guan, Z.-Z.; Yu, W.-F.; Nordberg, A. Dual Effects of Nicotine on Oxidative Stress and Neuroprotection in PC12 Cells. Neurochem Int. 2003, 43, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, V.; Flora, S.J.S. Effect of Nicotine Pretreatment on Arsenic-Induced Oxidative Stress in Male Wistar Rats. Hum Exp Toxicol 2013, 32, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Soto-Otero, R.; Méndez-Alvarez, E.; Hermida-Ameijeiras, A.; López-Real, A.M.; Labandeira-García, J.L. Effects of (-)-Nicotine and (-)-Cotinine on 6-Hydroxydopamine-Induced Oxidative Stress and Neurotoxicity: Relevance for Parkinson’s Disease. Biochem Pharmacol 2002, 64, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Goerig, M.; Ullrich, V.; Schettler, G.; Foltis, C.; Habenicht, A. A New Role for Nicotine: Selective Inhibition of Thromboxane Formation by Direct Interaction with Thromboxane Synthase in Human Promyelocytic Leukaemia Cells Differentiating into Macrophages. Clin Investig 1992, 70, 239–243. [Google Scholar] [CrossRef]
- Linert, W.; Bridge, M.H.; Huber, M.; Bjugstad, K.B.; Grossman, S.; Arendash, G.W. In Vitro and in Vivo Studies Investigating Possible Antioxidant Actions of Nicotine: Relevance to Parkinson’s and Alzheimer’s Diseases. Biochim Biophys Acta Mol Basis Dis 1999, 1454, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Nesil, T.; Cao, J.; Yang, Z.; Chang, S.L.; Li, M.D. Nicotine Mediates Expression of Genes Related to Antioxidant Capacity and Oxidative Stress Response in HIV-1 Transgenic Rat Brain. J Neurovirol 2016, 22, 114–124. [Google Scholar] [CrossRef]
- C.L. Crowley-Weber C. Crowley, H. Bernstein, C. Bernstein, H. Garewal, C.M. Payne, K.D. Nicotine Increases Oxidative Stress, Activates NF-KappaB and GRP78, Induces Apoptosis and Sensitizes Cells to Genotoxic/Xenobiotic Stresses by a Multiple Stress Inducer, Deoxycholate: Relevance to Colon Carcinogenesis. Chem. Biol. Interact. 2003, 145, 53–66.
- Igloi, G.; Brandsch, R. Arthrobacter Nicotinovorans PAO1 Megaplasmid Sequence,Strain ATCC 49919. GenBank 2002. [Google Scholar]
- El-Sabeh, A.; Honceriu, I.; Kallabi, F.; Boiangiu, R.-S.; Mihasan, M. Complete Genome Sequences of Two Closely Related Paenarthrobacter Nicotinovorans Strains. Microbiol Resour Announc 2022, 11. [Google Scholar] [CrossRef]
- Ruan, A.; Min, H.; Zhu, W. Studies on Biodegradation of Nicotine by Arthrobacter Sp. Strain HF-2. J Environ Sci Health B 2006, 41. [Google Scholar] [CrossRef]
- Yao, Y.; Tang, H.; Ren, H.; Yu, H.; Wang, L.; Xu, P. Genome Sequence of a Nicotine-Degrading Strain of Arthrobacter. J Bacteriol 2012, 194. [Google Scholar] [CrossRef]
- Ruan, A.; Gao, Y.; Fang, C.; Xu, Y. Isolation and Characterization of a Novel Nicotinophilic Bacterium, Arthrobacter Sp. ARF-1 and Its Metabolic Pathway. Biotechnol Appl Biochem 2018, 65. [Google Scholar] [CrossRef]
- Tang, H.; Yu, H.; Tai, C.; Huang, K.; Liu, Y.; Wang, L.; Yao, Y.; Wu, G.; Xu, P. Genome Sequence of a Novel Nicotine-Degrading Strain, Pseudomonas Geniculata N1. J Bacteriol 2012, 194. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Huang, K.; Wang, W.; Nie, X.; Jiang, Y.; Li, P.; Liu, S.; Xu, P.; Tang, H. Physiological and Biochemical Characterization of a Novel Nicotine-Degrading Bacterium Pseudomonas Geniculata N1. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Wei, H.; Lei, L.; Xia, Z.; Liu, S.; Liu, P.; Liu, X. Characterisation of a Novel Aerobic Nicotine-Biodegrading Strain of Pseudomonas Putida. Ann Microbiol 2008, 58. [Google Scholar] [CrossRef]
- Pan, D.; Sun, M.; Wang, Y.; Lv, P.; Wu, X.; Li, Q.X.; Cao, H.; Hua, R. Characterization of Nicotine Catabolism through a Novel Pyrrolidine Pathway in Pseudomonas Sp. S-1. J Agric Food Chem 2018, 66. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.Y.; Yu, Q.; Lei, L.P.; Wu, Y.P.; Ren, K.; Li, Y.; Zou, C.M. A Novel Nicotine-Degrading Bacterium Pseudomonas Fluorescens Strain 1206. Appl Biochem Microbiol 2019, 55. [Google Scholar] [CrossRef]
- Ruan, A.; Min, H.; Peng, X.; Huang, Z. Isolation and Characterization of Pseudomonas Sp. Strain HF-1, Capable of Degrading Nicotine. Res Microbiol 2005, 156. [Google Scholar] [CrossRef]
- Raman, G.; Mohan, K.N.; Manohar, V.; Sakthivel, N. Biodegradation of Nicotine by a Novel Nicotine-Degrading Bacterium, Pseudomonas Plecoglossicida TND35 and Its New Biotransformation Intermediates. Biodegradation 2014, 25. [Google Scholar] [CrossRef]
- Qiu, J.; Ma, Y.; Zhang, J.; Wen, Y.; Liu, W. Cloning of a Novel Nicotine Oxidase Gene from Pseudomonas Sp. Strain HZN6 Whose Product Nonenantioselectively Degrades Nicotine to Pseudooxynicotine. Appl Environ Microbiol 2013, 79. [Google Scholar] [CrossRef]
- Wang, H.H.; Yin, B.; Peng, X.X.; Wang, J.Y.; Xie, Z.H.; Gao, J.; Tang, X.K. Biodegradation of Nicotine by Newly Isolated Pseudomonas Sp. CS3 and Its Metabolites. J Appl Microbiol 2012, 112. [Google Scholar] [CrossRef]
- Chen, C.; Li, X.; Yang, J.; Gong, X.; Li, B.; Zhang, K.Q. Isolation of Nicotine-Degrading Bacterium Pseudomonas Sp. Nic22, and Its Potential Application in Tobacco Processing. Int Biodeterior Biodegradation 2008, 62. [Google Scholar] [CrossRef]
- Wang, S.N.; Liu, Z.; Xu, P. Biodegradation of Nicotine by a Newly Isolated Agrobacterium Sp. Strain S33. J Appl Microbiol 2009, 107. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhang, Y.; Zhao, L.; He, Q.; Jiang, J.; Hong, Q.; He, J. Isolation and Characterization of the Cotinine-Degrading Bacterium Nocardioides Sp. Strain JQ2195. J Hazard Mater 2018, 353. [Google Scholar] [CrossRef] [PubMed]
- Cobzaru, C.; Ganas, P.; Mihasan, M.; Schleberger, P.; Brandsch, R. Homologous Gene Clusters of Nicotine Catabolism, Including a New ω-Amidase for α-Ketoglutaramate, in Species of Three Genera of Gram-Positive Bacteria. Res Microbiol 2011, 162. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.F.; Xia, Z.Z.; Yao, J.C.; Feng, Z.; Li, D.H.; Liu, T.; Cheng, G.J.; He, D.L.; Li, X.H. Functional Analysis of the Ocne Gene Involved in Nicotine-Degradation Pathways in Ochrobactrum Intermedium SCUEC4 and Its Enzymatic Properties. Can J Microbiol 2021, 67. [Google Scholar] [CrossRef]
- Yu, H.; Tang, H.; Zhu, X.; Li, Y.; Xu, P. Molecular Mechanism of Nicotine Degradation by a Newly Isolated Strain, Ochrobactrum Sp. Strain SJY1. Appl Environ Microbiol 2015, 81. [Google Scholar] [CrossRef]
- Yuan, Y.J.; Lu, Z.X.; Wu, N.; Huang, L.J.; Lü, F.X.; Bie, X.M. Isolation and Preliminary Characterization of a Novel Nicotine-Degrading Bacterium, Ochrobactrum Intermedium DN2. Int Biodeterior Biodegradation 2005, 56. [Google Scholar] [CrossRef]
- Qiu, J.; Yang, Y.; Zhang, J.; Wang, H.; Ma, Y.; He, J.; Lu, Z. The Complete Genome Sequence of the Nicotine-Degrading Bacterium Shinella Sp. HZN7. Front Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Jiang, H.J.; Ma, Y.; Qiu, G.J.; Wu, F.L.; Chen, S.L. Biodegradation of Nicotine by a Novel Strain Shinella Sp. HZN1 Isolated from Activated Sludge. J Environ Sci Health B 2011, 46. [Google Scholar]
- Wang, M.; Yang, G.; Wang, X.; Yao, Y.; Min, H.; Lu, Z. Nicotine Degradation by Two Novel Bacterial Isolates of Acinetobacter Sp. TW and Sphingomonas Sp. TY and Their Responses in the Presence of Neonicotinoid Insecticides. World J Microbiol Biotechnol 2011, 27. [Google Scholar] [CrossRef]
- Jiang, Y.; Gong, J.; Chen, Y.; Hu, B.; Sun, J.; Zhu, Y.; Xia, Z.; Zou, C. Biodegradation of Nicotine and TSNAs by Bacterium Sp. Strain J54. Iran J Biotechnol 2021, 19, 20–27. [Google Scholar] [CrossRef]
- Zhang, K.; Yin, M.; Lei, S.; Zhang, H.; Yin, X.; Niu, Q. Bacillus Sp. YC7 from Intestines of Lasioderma Serricorne Degrades Nicotine Due to Nicotine Dehydrogenase. AMB Express 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhi, X.Y.; Qiu, J.; Shi, L.; Lu, Z. Characterization of a Novel Nicotine Degradation Gene Cluster Ndp in Sphingomonas Melonis TY and Its Evolutionary Analysis. Front Microbiol 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wen, R.; Qiu, J.; Hong, J.; Liu, M.; Zhang, D. Biodegradation of Nicotine by a Novel Strain Pusillimonas. Res Microbiol 2015, 166. [Google Scholar] [CrossRef]
- Gong, X.W.; Yang, J.K.; Duan, Y.Q.; Dong, J.Y.; Zhe, W.; Wang, L.; Li, Q.H.; Zhang, K.Q. Isolation and Characterization of Rhodococcus Sp. Y22 and Its Potential Application to Tobacco Processing. Res Microbiol 2009, 160. [Google Scholar] [CrossRef]
- Ganas, P.; Sachelaru, P.; Mihasan, M.; Igloi, G.L.; Brandsch, R. Two Closely Related Pathways of Nicotine Catabolism in Arthrobacter Nicotinovorans and Nocardioides Sp. Strain JS614. Arch Microbiol 2008, 189. [Google Scholar] [CrossRef]
- Lei, L.; Zhang, W.; Wei, H.; Xia, Z.; Liu, X. Characterization of a Novel Nicotine-Degrading Ensifer Sp. Strain N7 Isolated from Tobacco Rhizosphere. Ann Microbiol 2009, 59, 247–252. [Google Scholar] [CrossRef]
- Liu, G.; Wang, W.; He, F.; Zhang, P.; Xu, P.; Tang, H. Structural Insights into 6-Hydroxypseudooxynicotine Amine Oxidase from Pseudomonas Geniculata N1, the Key Enzyme Involved in Nicotine Degradation. Appl Environ Microbiol 2020, 86. [Google Scholar] [CrossRef]
- Grether-Beck, S.; Igloi, G.L.; Pust, S.; Schilz, E.; Decker, K.; Brandsch, R. Structural Analysis and Molybdenum-dependent Expression of the PAO1-encoded Nicotine Dehydrogenase Genes of Arthrobacter Nicotinovorans. Mol Microbiol 1994, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Qiu, J.; Chen, D.; Ye, J.; Wang, Y.; Tong, L.; Jiang, J.; Chen, J. Characterization and Genome Analysis of a Nicotine and Nicotinic Acid-Degrading Strain Pseudomonas Putida JQ581 Isolated from Marine. Mar Drugs 2017, 15. [Google Scholar] [CrossRef]
- Xia, Z.; Zhang, W.; Lei, L.; Liu, X.; Wei, H.L. Genome-Wide Investigation of the Genes Involved in Nicotine Metabolism in Pseudomonas Putida J5 by Tn5 Transposon Mutagenesis. Appl Microbiol Biotechnol 2015, 99. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, X.; Liu, X.; Wu, W.; Xu, P.; Tang, H. Cloning and Characterization the Nicotine Degradation Enzymes 6-Hydroxypseudooxynicotine Amine Oxidase and 6-Hydroxy-3-Succinoylpyridine Hydroxylase in Pseudomonas Geniculata N1. Int Biodeterior Biodegradation 2019, 142. [Google Scholar] [CrossRef]
- Li, H.; Xie, K.; Yu, W.; Hu, L.; Huang, H.; Xie, H.; Wang, S. Nicotine Dehydrogenase Complexed with 6-Hydroxypseudooxynicotine Oxidase Involved in the Hybrid Nicotine-Degrading Pathway in Agrobacterium Tumefaciens S33. Appl Environ Microbiol 2016, 82. [Google Scholar] [CrossRef] [PubMed]
- Brandsch, R.; Mihasan, M. A Soil Bacterial Catabolic Pathway on the Move: Transfer of Nicotine Catabolic Genes between Arthrobacter Genus Megaplasmids and Invasion by Mobile Elements. J Biosci 2020, 45. [Google Scholar] [CrossRef]
- El-Sabeh, A.; Mlesnita, A.M.; Munteanu, I.T.; Honceriu, I.; Kallabi, F.; Boiangiu, R.S.; Mihasan, M. Characterisation of the Paenarthrobacter Nicotinovorans ATCC 49919 Genome and Identification of Several Strains Harbouring a Highly Syntenic Nic-Genes Cluster. BMC Genomics 2023, 24. [Google Scholar] [CrossRef]
- Igloi, G.L.; Brandsch, R. Sequence of the 165-Kilobase Catabolic Plasmid PAO1 from Arthrobacter Nicotinovorans and Identification of a PAO1-Dependent Nicotine Uptake System. J Bacteriol 2003, 185, 1976–1986. [Google Scholar] [CrossRef]
- Mihǎşan, M.; Boiangiu, R.Ş.; Guzun, D.; Babii, C.; Aslebagh, R.; Channaveerappa, D.; Dupree, E.; Darie, C.C.; Mihǎşan, M. Time-Dependent Analysis of Paenarthrobacter Nicotinovorans PAO1 Nicotine-Related Proteome. ACS Omega 2021, 6. [Google Scholar] [CrossRef]
- Mihăşan, M.; Babii, C.; Aslebagh, R.; Channaveerappa, D.; Dupree, E.J.; Darie, C.C. Exploration of Nicotine Metabolism in Paenarthrobacter Nicotinovorans PAO1 by Microbial Proteomics. Adv Exp Med Biol 2019, 1140. [Google Scholar] [CrossRef]
- Mihăşan, M.; Babii, C.; Aslebagh, R.; Channaveerappa, D.; Dupree, E.; Darie, C.C. Proteomics Based Analysis of the Nicotine Catabolism in Paenarthrobacter Nicotinovorans PAO1. Sci Rep 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Chiribau, C.B.; Mihasan, M.; Ganas, P.; Igloi, G.L.; Artenie, V.; Brandsch, R. Final Steps in the Catabolism of Nicotine Deamination versus Demethylation of γ-N-Methylaminobutyrate. FEBS Journal 2006, 273. [Google Scholar] [CrossRef]
- Mihasan, M.; Brandsch, R. PAO1 of Arthrobacter Nicotinovorans and the Spread of Catabolic Traits by Horizontal Gene Transfer in Gram-Positive Soil Bacteria. J Mol Evol 2013, 77. [Google Scholar] [CrossRef] [PubMed]
- Ganas, P.; Brandsch, R. Uptake of L-Nicotine and of 6-Hydroxy-L-Nicotine by Arthrobacter Nicotinovorans and by Escherichia Coli Is Mediated by Facilitated Diffusion and Not by Passive Diffusion or Active Transport. Microbiology (N Y) 2009, 155. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, W.; König, K.; Andreesen, J.R. Nicotine Dehydrogenase from Arthrobacter Oxidans: A Molybdenum-Containing Hydroxylase. FEMS Microbiol Lett 1988, 52. [Google Scholar] [CrossRef]
- Fitzpatrick, P.F.; Chadegani, F.; Zhang, S.; Roberts, K.M.; Hinck, C.S. Mechanism of the Flavoprotein l -Hydroxynicotine Oxidase: Kinetic Mechanism, Substrate Specificity, Reaction Product, and Roles of Active-Site Residues. Biochemistry 2016, 55. [Google Scholar] [CrossRef]
- Yildiz, I.; Yildiz, B.S. Mechanistic Study of L-6-Hydroxynicotine Oxidase by DFT and ONIOM Methods. J Mol Model 2021, 27. [Google Scholar] [CrossRef]
- Koetter, J.W.A.; Schulz, G.E. Crystal Structure of 6-Hydroxy-D-Nicotine Oxidase from Arthrobacter Nicotinovorans. J Mol Biol 2005, 352. [Google Scholar] [CrossRef]
- Sachelaru, P.; Schiltz, E.; Brandsch, R. A Functional MobA Gene for Molybdopterin Cytosine Dinucleotide Cofactor Biosynthesis Is Required for Activity and Holoenzyme Assembly of the Heterotrimeric Nicotine Dehydrogenases of Arthrobacter Nicotinovorans. Appl Environ Microbiol 2006, 72. [Google Scholar] [CrossRef]
- Menéndez, C.; Otto, A.; Igloi, G.; Nick, P.; Brandsch, R.; Schubach, B.; Böttcher, B.; Brandsch, R. Molybdate-Uptake Genes and Molybdopterin-Biosynthesis Genes on a Bacterial Plasmid. Characterization of MoeA as a Filament-Forming Protein with Adenosinetriphosphatase Activity. Eur J Biochem 1997, 250. [Google Scholar] [CrossRef]
- Schleberger, C.; Sachelaru, P.; Brandsch, R.; Schulz, G.E. Structure and Action of a C‒C Bond Cleaving α/β-Hydrolase Involved in Nicotine Degration. J Mol Biol 2007, 367. [Google Scholar] [CrossRef]
- Chiribau, C.B.; Mihasan, M.; Ganas, P.; Igloi, G.; Artenie, V.; Brandsch, R. Final Steps in the Catabolism of Nicotine - Deamination versus Demethylation of Gamma-N-Methylaminobutyrate. Febs Journal 2006, 273, 1528–1536. [Google Scholar] [CrossRef]
- Ganas, P.; Mihasan, M.; Igloi, G.L.; Brandsch, R. A Two-Component Small Multidrug Resistance Pump Functions as a Metabolic Valve during Nicotine Catabolism by Arthrobacter Nicotinovorans. Microbiology (N Y) 2007, 153. [Google Scholar] [CrossRef] [PubMed]
- Treiber, N.; Schulz, G.E. Structure of 2,6-Dihydroxypyridine 3-Hydroxylase from a Nicotine-Degrading Pathway. J Mol Biol 2008, 379, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Chiribau, C.B.; Sandu, C.; Igloi, G.L.; Brandsch, R. Characterization of PmfR, the Transcriptional Activator of the PAO1-Borne PurU-MabO-FolD Operon of Arthrobacter Nicotinovorans. J Bacteriol 2005, 187, 3062–3070. [Google Scholar] [CrossRef] [PubMed]
- Mihasan, M.; Chiribau, C.-B.; Friedrich, T.; Artenie, V.; Brandsch, R. An NAD(P)H-Nicotine Blue Oxidoreductase Is Part of the Nicotine Regulon and May Protect Arthrobacter Nicotinovorans from Oxidative Stress during Nicotine Catabolism. Appl Environ Microbiol 2007, 73, 2479–2485. [Google Scholar] [CrossRef] [PubMed]
- Sandu, C.; Chiribau, C.B.; Brandsch, R. Characterization of HdnoR, the Transcriptional Repressor of the 6-Hydroxy-D-Nicotine Oxidase Gene of Arthrobacter Nicotinovorans PAO1, and Its DNA-Binding Activity in Response to L- and D-Nicotine Derivatives. J Biol Chem 2003, 278, 51307–51315. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhi, X.Y.; Qiu, J.; Shi, L.; Lu, Z. Characterization of a Novel Nicotine Degradation Gene Cluster Ndp in Sphingomonas Melonis TY and Its Evolutionary Analysis. Front Microbiol 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Tang, H.; Zhu, X.; Li, Y.; Xu, P. Molecular Mechanism of Nicotine Degradation by a Newly Isolated Strain, Ochrobactrum Sp. Strain SJY1. Appl Environ Microbiol 2015, 81, 272–281. [Google Scholar] [CrossRef]
- Qiu, J.; Yang, Y.; Zhang, J.; Wang, H.; Ma, Y.; He, J.; Lu, Z. The Complete Genome Sequence of the Nicotine-Degrading Bacterium Shinella Sp. HZN7. Front Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Ma, Y.; Wei, Y.; Qiu, J.; Wen, R.; Hong, J.; Liu, W. Isolation, Transposon Mutagenesis, and Characterization of the Novel Nicotine-Degrading Strain Shinella Sp. HZN7. Appl Microbiol Biotechnol 2014, 98, 2625–2636. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xie, K.; Yu, W.; Hu, L.; Huang, H.; Xie, H.; Wang, S. Nicotine Dehydrogenase Complexed with 6-Hydroxypseudooxynicotine Oxidase Involved in the Hybrid Nicotine-Degrading Pathway in Agrobacterium Tumefaciens S33. Appl Environ Microbiol 2016, 82, 1745. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, X.; Liu, X.; Wu, W.; Xu, P.; Tang, H. Cloning and Characterization the Nicotine Degradation Enzymes 6-Hydroxypseudooxynicotine Amine Oxidase and 6-Hydroxy-3-Succinoylpyridine Hydroxylase in Pseudomonas Geniculata N1. Int Biodeterior Biodegradation 2019, 142, 83–90. [Google Scholar] [CrossRef]
- Huang, H.; Yu, W.; Wang, R.; Li, H.; Xie, H.; Wang, S. Genomic and Transcriptomic Analyses of Agrobacterium Tumefaciens S33 Reveal the Molecular Mechanism of a Novel Hybrid Nicotine-Degrading Pathway. Sci Rep 2017, 7. [Google Scholar] [CrossRef]
- Wang, R.; Yi, J.; Shang, J.; Yu, W.; Li, Z.; Huang, H.; Xie, H.; Wang, S. 6-Hydroxypseudooxynicotine Dehydrogenase Delivers Electrons to Electron Transfer Flavoprotein during Nicotine Degradation by Agrobacterium Tumefaciens S33. Appl Environ Microbiol 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, H.; Xie, K.; Xu, P. Identification of Nicotine Biotransformation Intermediates by Agrobacterium Tumefaciens Strain S33 Suggests a Novel Nicotine Degradation Pathway. Appl Microbiol Biotechnol 2012, 95, 1567–1578. [Google Scholar] [CrossRef]
- Yu, W.; Wang, R.; Li, H.; Liang, J.; Wang, Y.; Huang, H.; Xie, H.; Wang, S. Green Route to Synthesis of Valuable Chemical 6-Hydroxynicotine from Nicotine in Tobacco Wastes Using Genetically Engineered Agrobacterium Tumefaciens S33. Biotechnol Biofuels 2017, 10, 288. [Google Scholar] [CrossRef]
- Qiu, J.; Wei, Y.; Ma, Y.; Wen, R.; Wen, Y.; Liu, W. A Novel (S)-6-Hydroxynicotine Oxidase Gene from Shinella Sp. Strain HZN7. Appl Environ Microbiol 2014, 80, 5552–5560. [Google Scholar] [CrossRef]
- Qiu, J.; Ma, Y.; Wen, Y.; Chen, L.; Wu, L.; Liu, W. Functional Identification of Two Novel Genes from Pseudomonas Sp. Strain HZN6 Involved in the Catabolism of Nicotine. Appl Environ Microbiol 2012, 78, 2154–2160. [Google Scholar] [CrossRef]
- Novotny, T.E.; Zhao, F. Consumption and Production Waste: Another Externality of Tobacco Use. Tob Control 1999, 8, 75–80. [Google Scholar] [CrossRef]
- Civilini, M. Nicotine Decontamination of Tobacco Agro-Industrial Waste and Its Degradation by Micro-Organisms. Waste Management & Research 1997, 15, 349–358. [Google Scholar] [CrossRef]
- Tang, H.; Wang, L.; Meng, X.; Lanying, M.; Wang, S.; He, X.; Wu, G.; Xu, P. Novel Nicotine Oxidoreductase-Encoding Gene Involved in Nicotine Degradation by Pseudomonas Putida Strain S16. Appl Environ Microbiol 2009, 75, 772–778. [Google Scholar] [CrossRef]
- Qiu, J.; Wei, Y.; Ma, Y.; Wen, R.; Wen, Y.; Liu, W. A Novel (S)-6-Hydroxynicotine Oxidase Gene from Shinella Sp. Strain HZN7. Appl Environ Microbiol 2014, 80, 5552. [Google Scholar] [CrossRef]
- Lenkey, A.A.; Hills, S. Nicotine Removal Process and Product Produced Thereby 1987.
- Wang, M.; Yang, G.; Min, H.; Lv, Z.; Jia, X. Bioaugmentation with the Nicotine-Degrading Bacterium Pseudomonas Sp. HF-1 in a Sequencing Batch Reactor Treating Tobacco Wastewater: Degradation Study and Analysis of Its Mechanisms. Water Res 2009, 43, 4187–4196. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Zhu, C.; Shu, M.; Sun, K.; Zhao, L.; Wang, C.; Ye, Z.; Chen, J. Degradation of Nicotine in Tobacco Waste Extract by Newly Isolated Pseudomonas Sp. ZUTSKD. Bioresour Technol 2010, 101, 6935–6941. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.S.; Wang, L.J.; He, H.Z.; Wang, M.Z. Effect of Transient Nicotine Load Shock on the Performance of Pseudomonas Sp. HF-1 Bioaugmented Sequencing Batch Reactors. J Chem 2016, 2016. [Google Scholar] [CrossRef]
- Wang, X.; Tang, L.; Yao, Y.; Wang, H.; Min, H.; Lu, Z. Bioremediation of the Tobacco Waste-Contaminated Soil by Pseudomonas Sp. HF-1: Nicotine Degradation and Microbial Community Analysis. Appl Microbiol Biotechnol 2013, 97, 6077–6088. [Google Scholar] [CrossRef]
- Wang, M.Z.; He, H.Z.; Zheng, X.; Feng, H.J.; Lv, Z.M.; Shen, D.S. Effect of Pseudomonas Sp. HF-1 Inoculum on Construction of a Bioaugmented System for Tobacco Wastewater Treatment: Analysis from Quorum Sensing. Environmental Science and Pollution Research 2014, 21, 7945–7955. [Google Scholar] [CrossRef]
- Briški, F.; Kopčić, N.; Ćosić, I.; Kučić, D.; Vuković, M. Biodegradation of Tobacco Waste by Composting: Genetic Identification of Nicotine-Degrading Bacteria and Kinetic Analysis of Transformations in Leachate. Chemical Papers 2012, 66, 1103–1110. [Google Scholar] [CrossRef]
- Mandić, N.; Lalević, B.; Raičević, V.; Radojičić, V. Impact of Composting Conditions on the Nicotine Degradation Rate Using Nicotinophilic Bacteria from Tobacco Waste. International Journal of Environmental Science and Technology 2023, 20, 7787–7798. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, X.; Chu, P.; Shi, H.; Wang, R.; Li, J.; Zheng, S. Cultivation and Application of Nicotine-Degrading Bacteria and Environmental Functioning in Tobacco Planting Soil. Bioresour Bioprocess 2023, 10, 1–11. [Google Scholar] [CrossRef]
- Wang, S.N.; Xu, P.; Tang, H.Z.; Meng, J.; Liu, X.L.; Qing, C. “Green” Route to 6-Hydroxy-3-Succinoyl-Pyridine from (S)-Nicotine of Tobacco Waste by Whole Cells of a Pseudomonas Sp. Environ Sci Technol 2005, 39, 6877–6880. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Tang, H.; Xu, P. Green Strategy from Waste to Value-Added-Chemical Production: Efficient Biosynthesis of 6-Hydroxy-3-Succinoyl-Pyridine by an Engineered Biocatalyst. Sci Rep 2014, 4, 5397. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, P.; Tang, H. Sustainable Production of Valuable Compound 3-Succinoyl-Pyridine by Genetically Engineering Pseudomonas Putida Using the Tobacco Waste. Sci Rep 2015, 5, 16411. [Google Scholar] [CrossRef] [PubMed]
- Mihalache, G.; Babii, C.; Stefan, M.; Motei, D.; Marius, M. Steps towards an Arthrobacter Nicotinovorans Based Biotechnology for Production of 6-Hidroxy-Nicotine. In Proceedings of the 16 th INTERNATIONAL MULTIDISCIPLINARY SCIENTIFIC GEOCONFERENCE ; 2016; pp. 341–346. [Google Scholar]
- Hrițcu, L.; Ștefan, M.; Brandsch, R.; Mihășan, M. 6-Hydroxy-l-Nicotine from Arthrobacter Nicotinovorans Sustain Spatial Memory Formation by Decreasing Brain Oxidative Stress in Rats. J Physiol Biochem 2013, 69, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Hritcu, L.; Stefan, M.; Brandsch, R.; Mihasan, M. Enhanced Behavioral Response by Decreasing Brain Oxidative Stress to 6-Hydroxy-l-Nicotine in Alzheimer’s Disease Rat Model. Neurosci Lett 2015, 591, 41–47. [Google Scholar] [CrossRef]
- Hritcu, L.; Ionita, R.; Motei, D.E.; Babii, C.; Stefan, M.; Mihasan, M. Nicotine versus 6-Hydroxy-l-Nicotine against Chlorisondamine Induced Memory Impairment and Oxidative Stress in the Rat Hippocampus. Biomedicine & Pharmacotherapy 2017, 86, 102–108. [Google Scholar] [CrossRef]
- Ioniță, R.; Valu, V.M.; Postu, P.A.; Cioancă, O.; Hrițcu, L.; Mihasan, M. 6-Hydroxy-l-Nicotine Effects on Anxiety and Depression in a Rat Model of Chlorisondamine. Farmacia 2017, 65. [Google Scholar]
- Boiangiu, R.S.; Mihasan, M.; Gorgan, D.L.; Stache, B.A.; Petre, B.A.; Hritcu, L. Cotinine and 6-Hydroxy-L-Nicotine Reverses Memory Deficits and Reduces Oxidative Stress in Aβ25-35-Induced Rat Model of Alzheimer’s Disease. Antioxidants 2020, 9, 768. [Google Scholar] [CrossRef]
- Boiangiu, R.S.; Mihasan, M.; Gorgan, D.L.; Stache, B.A.; Hritcu, L. Anxiolytic, Promnesic, Anti-Acetylcholinesterase and Antioxidant Effects of Cotinine and 6-Hydroxy-L-Nicotine in Scopolamine-Induced Zebrafish (Danio Rerio) Model of Alzheimer’s Disease. Antioxidants 2021, 10, 212. [Google Scholar] [CrossRef]
- Thapa, S.; Lv, M.; Xu, H. Acetylcholinesterase: A Primary Target for Drugs and Insecticides. Mini-Reviews in Medicinal Chemistry 2017, 17, 1665–1676. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.S.; Girgis, A.S.; Prakash, A.; Khanna, L.; Khanna, P.; Shalaby, E.M.; Fawzy, N.G.; Jain, S.C. Protective Effects of Aporosa Octandra Bark Extract against D-Galactose Induced Cognitive Impairment and Oxidative Stress in Mice. Heliyon 2018, 4, e00951. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liao, C.; Ge, F.; Ao, J.; Liu, T. Acetylcholine Bidirectionally Regulates Learning and Memory. Journal of Neurorestoratology 2022, 10. [Google Scholar] [CrossRef]
- Sam, C.; Bordoni, B. Physiology, Acetylcholine; 2021.
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Khachaturian, A.S.; Vergallo, A.; Farlow, M.R.; Snyder, P.J.; Giacobini, E.; Khachaturian, Z.S. Revisiting the Cholinergic Hypothesis in Alzheimer’s Disease: Emerging Evidence from Translational and Clinical Research. J Prev Alzheimers Dis 2019, 6, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Cholinesterase Inhibitors as Alzheimer’s Therapeutics (Review). Mol Med Rep 2019, 20, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Mihășan, M.; Căpățînă, L.; Neagu, E.; Ștefan, M.; Hrițcu, L. In-Silico Identification of 6-Hydroxy-L-Nicotine as a Novel Neuroprotective Drug. Rom Biotechnol Lett 2013, 18, 8333–8340. [Google Scholar]
- Mocanu, E.M.; Mazarachi, A.L.; Mihasan, M. In Vitro Stability and Antioxidant Potential of the Neuprotective Metabolite 6-Hydroxy-Nicotine. JOURNAL OF EXPERIMENTAL AND MOLECULAR BIOLOGY 2018, 19, 53–58. [Google Scholar]
- Yamazaki, H.; Tanji, K.; Wakabayashi, K.; Matsuura, S.; Itoh, K. Role of the Keap1/Nrf2 Pathway in Neurodegenerative Diseases. Pathol Int 2015, 65, 210–219. [Google Scholar] [CrossRef]
- Taly, A.; Corringer, P.-J.; Guedin, D.; Lestage, P.; Changeux, J.-P. Nicotinic Receptors: Allosteric Transitions and Therapeutic Targets in the Nervous System. Nat Rev Drug Discov 2009, 8, 733–750. [Google Scholar] [CrossRef]
- Lombardo, S.; Maskos, U. Role of the Nicotinic Acetylcholine Receptor in Alzheimer’s Disease Pathology and Treatment. Neuropharmacology 2015, 96, 255–262. [Google Scholar] [CrossRef]
- Posadas, I.; López-Hernández, B.; Ceña, V. Nicotinic Receptors in Neurodegeneration. Curr Neuropharmacol 2013, 11, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, A.H.; Levin, E.D. Cognitive Effects of Nicotine. Biol Psychiatry 2001, 49, 258–267. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The PRISMA flow diagram of the search and selection of the included studies.
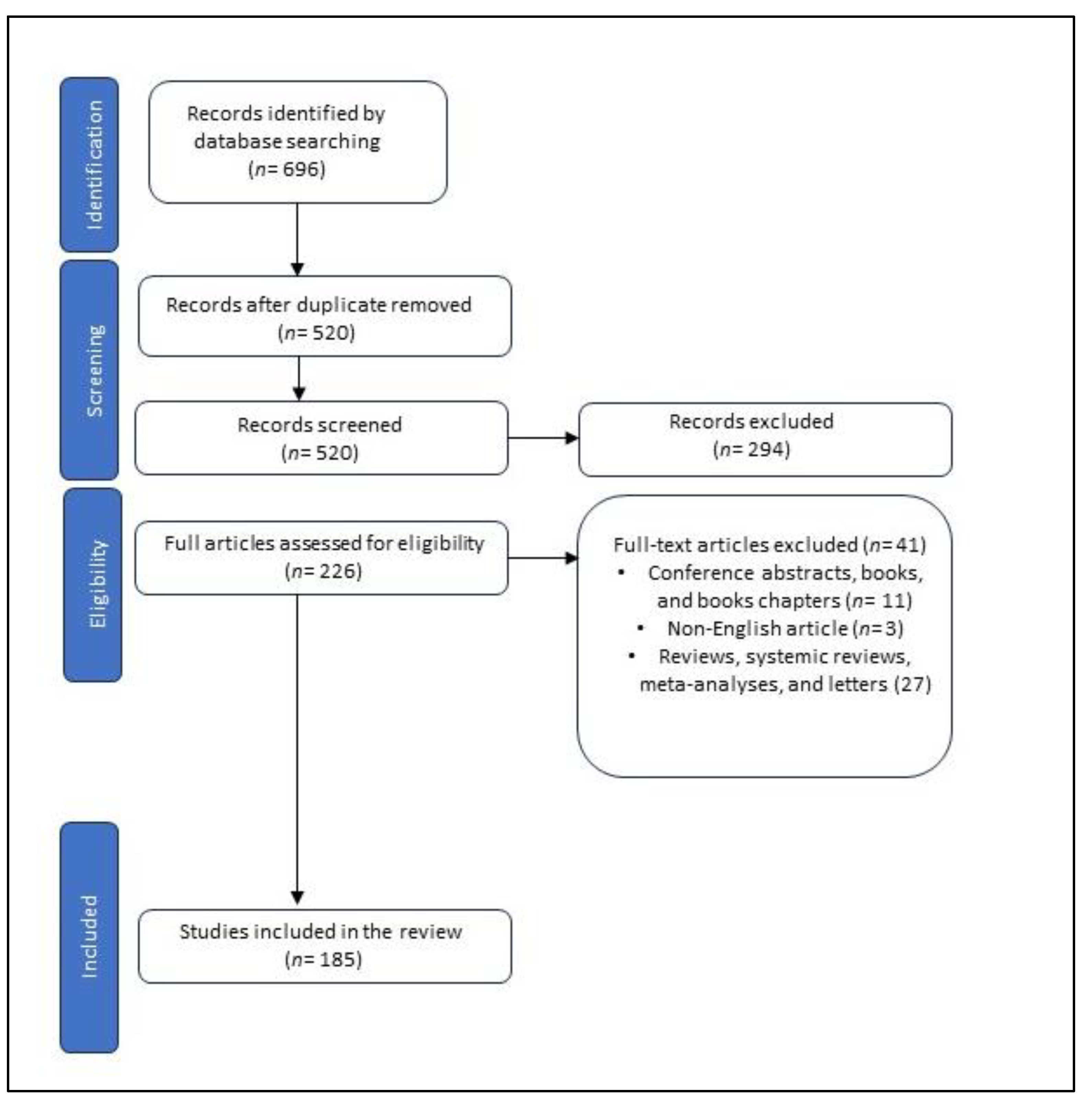

Table 1.
The effects of 6-hydroxy-L-nicoine (6HLN) on animal behavior in specific tasks
| Model | Model inducer (dose, route of administration, time of exposure) | Dose of 6HLN, route of administration and time of exposure | Behavioral task | Phenotype | References |
|---|---|---|---|---|---|
| Ratus norvegicus | |||||
| Normal | - | 0.3 mg/kg, b.w., i.p., for 7 consecutive days |
Y-maze | Improves short-term memory acquisition and increases locomotor activity | [167] |
| Normal | - | 0.3 mg/kg, b.w., i.p., for 7 consecutive days |
Radial arm-maze | Improves working memory. no effect on long-term memory |
[167] |
| AD | SCOP (0.7 mg/kg, b.w., i.p., 24 h before testing) | 0.3 mg/kg, b.w., i.p., 30 min before testing |
Y-maze | Improves spatial working memory. no effect on locomotor activity |
[168] |
| AD | SCOP (0.7 mg/kg, b.w., i.p., 24 h before testing) | 0.3 mg/kg, b.w., i.p., 30 min before testing |
Radial arm-maze | Improves spatial memory formation | [168] |
| AD | CHL (10 mg/kg, b.w., i.p., 24 h before testing) | 0.3 mg/kg, b.w., i.p., 30 min before testing |
Y-maze | Enhances spatial memory formation | [169] |
| AD | CHL (10 mg/kg, b.w., i.p., 24 h before testing) | 0.3 mg/kg, b.w., i.p., 30 min before testing |
Radial arm maze | Improves short- and long-term memory | [169] |
| AD | CHL (10 mg/kg, b.w., i.p., 24 h before testing) | 0.3 mg/kg, b.w., i.p., 30 min before testing |
Elevated plus maze | Anxiolytic properties (increased time and entries in open arms) | [170] |
| AD | CHL (10 mg/kg, b.w., i.p., 24 h before testing) | 0.3 mg/kg, b.w., i.p., 30 min before testing |
Forced swimming | Anti-depressant effect (reduce the immobility period) | [170] |
| AD | Aβ25-35 peptide fragment (0.5 mg/ml, 4 µl, i.c.v.) | 0.3 and 0.7 mg/kg, b.w., i.p., for 33 days | Y-maze | Improves spatial recognition memory. Restored the normal locomotor activity. |
[171] |
| AD | Aβ25-35 peptide fragment (0.5 mg/ml, 4 µl, i.c.v.) | 0.3 and 0.7 mg/kg, b.w., i.p., for 33 days |
Radial arm maze | Rescue short- and long-term memory | [171] |
| AD | Aβ25-35 peptide fragment (0.5 mg/ml, 4 µl, i.c.v.) | 0.3 and 0.7 mg/kg, b.w., i.p., for 33 days | Novel object recognition | Improves recognition memory | [171] |
| Danio rerio | |||||
| AD | SCOP (100 µM, immersion, 30 before testing) | 1 and 2 mg/L, immersion, for 3 min |
Y-maze | Rescue spatial recognition memory. Enhances locomotor activity. |
[172] |
| AD | SCOP (100 µM, immersion, 30 before testing) | 1 and 2 mg/L, immersion, for 3 min |
Novel object recognition | Improves recognition memory. | [172] |
| AD | SCOP (100 µM, immersion, 30 before testing) | 1 and 2 mg/L, immersion, for 3 min | Novel tank diving test | Reduces anxiety-like behavior. Induces hyperactivity. |
[172] |
Table legend: AD – Alzheimer’s disease; SCOP – scopolamine; CHL – chlorisondamine; i.p. – intraperitoneal, i.c.v. – intracerebroventricular; b.w. – body weight.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



