Submitted:
02 October 2023
Posted:
04 October 2023
You are already at the latest version
Abstract
High production cost is one of the major factors that limit the market growth of polyhydroxyalkanoates (PHAs) as a biopolymer. Improving PHA synthesis performance and utilizing low grade feedstock are two logical strategies for reducing the cost. As an oleaginous yeast, Y. lipolytica has a high carbon flux through acetyl-CoA (main PHB precursor) which makes it a desired cell factory for PHB biosynthesis. Two different metabolic pathways introduced into Y. lipolytica PO1f in this study for enhancing PHB biosynthesis, along with glucose and acetate as co-substrates. Results showed that NBC pathway led to more than 11% PHB accumulation, while there was non-detectable PHB accumulations using ABC pathway. Further modifications were done using peroxisomal compartmentalization and gene-dose overexpression. Final strain showed up to 41% PHB of CDW with a growth rate of 0.227 h-1. Low growth rate was observed using acetate or glucose as sole carbon and energy source. The co-substrate strategy showed compensatory effect on overcoming inhibitory and toxic effects of both substrates at high concentrations. Cultivating the engineered strain in optimal co-substrate condition predicted by RSM led to 83.4 g/L biomass concentration and 31.7 g/L PHB.
Keywords:
Acetate
; Gene dosage effect
; Acetoacetyl-CoA synthase
; Compartmentalization
; Substrate inhibition
1. Introduction
The increasingly dependence on plastics has led to the generation of a large amount of plastic wastes, underscoring a critical need for biodegradable material as a sustainable replacement of the petroleum-based plastics [1]. There are several alternatives for fossil fuel-based plastics including: poly lactic acid (PLA), bio-polyethylene, polyhydroxyalkanoates (PHAs), and protein-based bioplastics [2]. Among different types of PHAs, polyhydroxybutyrate (PHB) is a naturally occurring polymer which is produced by many different bacterial and archaea species. The recognized advantages and special characteristics of PHB include sustainability, biocompatibility (non-toxic), biodegradability, versatility (using injection molding, extrusion, and film blowing as conventional techniques in plastic processing), customizability (possibility of modifying characteristics such as characteristics such as flexibility, tensile strength, and degradation rate), good resistance to UV radiations and insolubility in water. PHB is therefore considered as one of the alternatives that has most potential for the current plastic market [3] with different applications from food packaging to textiles, cosmetics, and biomedical implants.
Some of the microbial species are widely studied as natural producers of PHB, such as Cupriavidus necator (formerly Ralstonia eutropha), Azotobacter vinelandii, Aeromonas hydrophila, Pseudomonas putida, and Alcaligenes eutrophus [4]. To be a suitable microbial cell factory for producing PHB the selected species should have the following properties. First, it should be a generally recognized as safe (GRAS) microorganism; second, it should be a fast growing organism with significant PHB accumulation, to make the scale-up process feasible; and third, it should be able to use different and low cost substrates to make an affordable bioprocess [5]. However, among the PHB natural producers, very few have all these characteristics.
These challenges relying on the natural PHB producers have led to various attempts using genetic modification of some most robust GRAS microbial cell factories such as Saccharomyces cerevisiae, Pichia pastoris, Corynebacterium glutamicum, Yarrowia lipolytica, and Zymomonas mobilis [6]. Among these potential cell factories, Y. lipolytica is uniquely advantageous for PHB biosynthesis. Y. lipolytica historical uses in food, feed, lipid production, and several other industrial applications proved that Y. lipolytica was generally nonpathogenic and does not pose significant health risks to humans or animals. Indeed the U.S. Food and Drug Administration (FDA) declare Y. lipolytica as a GARS microorganism which indicates the reason of such high interest toward this yeast in industrial biotechnology [7].
Y. lipolytica grows relatively fast with a doubling time of 1.7 h-1 [8] and can be cultivated at various scales, which makes it suitable for both research and large-scale bioproduct applications. Additional benefits of this yeast for PHB production include: 1) the avoidance of bacteriophage attacks at industrial levels; 2) tolerant to inhibitors and pH stresses; and 3) forming filamentous cells under late growth stage that facilitate gravity separation [9].
Y. lipolytica is an oleaginous yeast with a great potential for producing different biochemicals using nutrient limited media and its metabolism is well-established for biosynthesizing triglycerides (TGs) [10]. Therefore, Y. lipolytica has a high flux through acetyl-CoA which makes it a desired cell factory for those moleculars that require acetyl-CoA as intermediate. These products include TGs, carotenoids, fatty acid derivatives, organic acids, and PHAs [11]. Some efforts have been made to genetically modify this cell factory to produce PHB to using its unique characteristics. Generally, two strategies have been used to design PHB biosynthesis pathways in Y. lipolytica. First, the ABC pathway consists of three genes phaA, phaB and phaC. As a result of applying the ABC pathway from Ralstonia eutropha to Y. lipolytica, Li and colleagues successfully produced PHB up to 10.2% of cell dry weight (CDW) by using glucose as a substrate [12]. The second strategy, however, is to introduce only the phaC gene, because other PHB precursors, such as acetoacetyl-CoA, are naturally synthesized by Y. lipolytica. Using this strategy, in another study, the phaC gene from Pseudomonas aeruginosa contained a PTS1 peroxisomal signal introduced to Y. lipolytica. Its result was mcl-PHA accumulation up to 5% of CDW [13]. Similar results were observed by Rigouin and colleagues, using the mutagenesis of phaC which led to 25% mcl-PHA of CDW (w/w) [14].
Despite all these efforts the modified strains still have a low PHB titer which is the main bottleneck for the scale-up process. A main reason is that Y. lipolytica has a robust flux toward the lipogenesis pathway which leads to high lipid accumulation rather than PHB [15]. Therefore, harnessing the lipogenesis pathway toward PHB precursors seems to be an effective strategy to increase PHB titer in Y. lipolytica. Indeed, previous studies on redirecting malonyl-CoA (main intermediate for fatty acid synthesis) in sugarcane toward PHB biosynthesis showed higher PHB titer in mesophyll cells than ABC pathway. The NBC pathway (including NphT7, phaB, and phaC genes) redirects the lipogenesis pathway to PHB biosynthesis by catalyzing the condensation reaction of malonyl-CoA to acetoacetyl-CoA. In contrast to PhaA which catalyzes the condensation reaction reversibly, NphT7 catalyzes the condensation reaction irreversibly, resulting in a higher accumulation of PHB [16].
Another bottleneck for producing PHB using modified Y. lipolytica is selecting a cost-effective and sustainable substrate for large scale fermentation. Indeed, several efforts have been made to develop PHB biosynthesis using metabolically engineered Y. lipolytica strains. In most cases, glucose is used as the substrate. Also, several studies showed glycerol as another preferred substrate for producing biodiesel using Y. lipolytica [17].
However, there are many other substrates to consider as glucose and glycerol alternatives. These substrates should be considered not only due to their low cost, but also for other benefits such as sustainability and better conversion to products of interest [18]. Acetic acid and other organic acids derived from waste material show inhibitory effects to many microorganisms including E. coli and S. cerevisiae. Y. lipolytica on the other hand, can readily uptake and convert acetic acid. This making it possible to utilize a variety of wastes through the VFA route that are less efficiently utilized by other organisms [19]. Using wastes as feedstock to produce biodegradable plastics would provide the most sustainable solution for the plastic production dilemma [20]. Despite this, using acetic acid (generally VFAs) as only carbon source cannot satisfy the industrial demand for fast growth media with optimized PHAs production. The main reason for this deficiency is unbalanced reductive agents (as NADPH) and energy carriers (as ATP) produced during the metabolic pathway of acetic acid [21]. On the other hand, glucose showed a noticeable higher growth rate and biomass productivity than acetic acid. Therefore, co-substrate utilization is a reliable strategy for low-cost and efficient PHA production [22].
In the current study, two PHB pathways, ABC pathway and NBC pathway, were introduced into Y. lipolytica and compared for PHB biosynthesis. In order to further enhance the PHB titer, gene dosage effect and compartmentalization strategies were used to reach the final modified strain. Also, a co-substrate strategy was implemented using acetate as a cost-effective substrate and glucose as dopant. The strategy was evaluated for the effect of each substrate and their mix on growth rate and biomass PHB content. These results led to an optimized co-substrate medium for PHB production using the engineered strain. Furthermore, this study provides significant insights into metabolic engineering of Y. lipolytica for the biosynthesis of PHB.
2. Materials and Methods
2.1. Strain Construction
The auxotrophic strain Y. lipolytica PO1f (ATCC MYA-2613) was used as a microbial host for the introduced pathway. E. coli DH5α was used for cloning the genes and amplifying the constructed shuttle vectors. To develop constructs single genes are amplified using PCR and cloned using a cloning vector in E. coli. The list of primers used for PCR reaction shows in Table 1. Cloned genes are assembled in E. coli-Y. lipolytica shuttle vectors. Assembled cassettes include genes, biomarkers, both well-characterized promoter (TEF1, GPM, and FBA) and terminator (XPR2, OCT1, and LIP1) elements. Next, existing the target genes in expression cassettes, and integration of the assembled expression cassettes into Y. lipolytica genome were verified using colony PCR and markers.
To cultivate and screen transformed E. coli for constructed plasmids, Luria-Bertani (LB) medium (5 g/l yeast extract, 10 g/l Bacto tryptone, and 10 g/l NaCl) at 37 °C with ampicillin as a marker was used. Synthetic yeast nitrogen base (YNB) media consisting of 20 g/L glucose as carbon source, at 28 ℃ was used for screening the transformed PO1f host. Transformants were selected on a defined medium containing 6.7 g/l YNB without amino acids. In each step medium was supplemented with 0.77g/L CSM-Ura (Complete supplement mixture minus uracil, US-Biological, USA), 0.67 g/L CSM-leucine, and 0.3 g/L hygromycin [12,23,24] glucose based on introduced vector.
Y. lipolytica wild-type strains are unable to synthesize PHB naturally. However, there are two common strategies for introducing the PHB biosynthesis pathway to non-natural PHB producer organisms. To evaluating the efficiency of these pathways in Y. lipolytica two vectors designed for expressing the ABC PHB biosynthesis pathway and NBC pathway. These expression cassettes contained fragments (promoters, terminators, and other regulatory elements). The only difference was that the ABC cassette contained phaA gene which expressed β-ketothiolase (pZX22), while the NBC cassette contained NphT7 gene (pZX30).
To reach to the final strain, Y. lipolytica PO1f modified by introducing the NBC pathway contained following genes: phaB (from R. eutropha), phaC1437 which encoding a mutant Pseudomonas sp, and NphT7 (acetoacetyl-CoA synthase) gene from Streptomyces sp. [12]. Then modified strain subjected to sequential transformations using integrative vectors. These modifications include increase the copy number of NBC PHB pathway genes [25]. Also bktb (β-ketothiolase) gene from R. eutropha was introduced to modified strain to catalyze the condensation of acyl-CoA and acetyl-CoA into 3-ketoacyl-CoA molecules to copolymer synthesis in new strain [26]. PHB synthase tagged with carboxyl-terminal amino acid sequence serine-lysine-leucine (SKL) to peroxisomal compartmentalization of synthesized PHB [27]. The map of constructed designed pathway shown in Figure 1A. These vectors are named pCV35, pCV88, and pCV342 which are shown in Figure 1B, Figure 1C, and Figure 1D respectively [28,29]. During a sequential transformation Y. lipolytica PO1f was integrated with pCV35 (PTEF-NphT7-Txpr2, PGMP-PhaC1437-Toct1, PFBA-PhaB-Tlip1, leu2 maker), and the result strain was transformed with linearized plasmid, pCV88 (PTEF-NphT7-TXPR2, PGMP-PhaC1437-TOCT1, PFBA-PhaB-TLIP1, ura3 maker). This current strain subjected to future transformation by linearized plasmid pCV342 (PTEF1N-PhaB-Txpr2, PTEF1N- PhaC1437_SKL-Txpr2, PTEF1N-BktB-Txpr2, hph marker). Therefore, there were two copies of PhaC1437, one copy of PhaC1437_SKL, two copies of NphT7, two copies of PhaB, and one copy of BktB in the Y. lipolytica PHB32 as the final strain.
2.2. Cultures for Y. lipolytica cultivation and PHA Production
Inoculants were prepared from single colonies in 25 mL of YPD medium containing 10 g/L yeast extract, 20g/L peptone and 20g/L glucose (dextrose) and incubated for 48 h at 28 ℃ with 150 rpm agitation. YNB medium was used for cell growth and PHB production in flask experiments. The YNB medium contained 1.7 g/L YNB w/o AA and 5 g/L NH4CL. Seven different levels of glucose (0, 5, 10, 20, 50, 100, and 150 g/L) and five different levels of sodium acetate (0, 10, 20, 50, and 100 g/L) were tested as substrate. 250 mL flasks were inoculated by 0.5% (v/v) of prepared seed of the engineered strain and incubated at 28 ℃ for 96 h. To produce PHB in the fermenter, a 1 L bioreactor was employed. The base medium was YPD, and the bioreactor operated at optimum concentrations of glucose and sodium acetate. The bioreactor was inoculated with 1% (v/v) prepared seeds and fermentation was done at 28 ℃ at a constant pH of 7. After 24 hours of batch culture and achieving an increased cell concentration, the bioreactor was switched to fed-batch culture. During this phase a constant flow of YPD containing carbon sources were injected into bioreactor. This fed-batch system was maintained with glucose and sodium acetate at their optimum concentration for an additional 48 hours. Filtered air was sparged into bioreactor with a flow rate of 2.5 L/min. To maintain the air saturation at 20%, flat blade used with adjusted agitation rate between 150 to 500 rpm.
2.3. Growth Rate and PHB Content Analysis
To detect and quantify the growth rate of the engineered strain, optical density (OD) of the culture at 600 nm was used for determining the cell dry weight (CDW). To measure the intracellular PHB content, 2 mL of culture were centrifuged at 5000 g for 5 min. Then, settled cell pellets were washed with deionized water and dried at 55 ℃ overnight. The dried biomass was digested by using a methanolysis solution containing 97% methanol and 3% H2SO4, 18M (v/v) for determined the PHB content [12]. During the transesterification reaction, PHB polymers were monomerized to butanoic acid, 3hydroxy-, methyl ester.
As a brief description, 1.5 mL of methanolysis solution and 3 mL of chloroform were added to each sample and then incubated at 95 ℃ for at least 6 h. After cooling the samples to room temperature, 1.5 mL deionized water was added to each sample which created two layers. The upper layer contained water, methanol and hydrophilic cell components; and lower layer contained chloroform and hydrophobic cell components including digested PHB (Figure 2). The separated chloroform layer was analyzed by Agilent 7890A gas chromatograph, equipped with HP-5MS 30m X 250µm X 0.25µm column, and coupled by 5975C mass detector. The temperature was held at 40 ℃ for 3 min, then ramped from 40 ℃ to 280 ℃ at 10 ℃ min-1 and held for 10 min.
2.4. Statistical Analysis
For each designed experiment, data were collected from three biologically independent experiments. SAS® 9.4M7 software was used for analyzing experimental data and calculating parameters such as mean value, standard deviation, and standard errors. Also, in order to evaluate the synergistic effects of both glucose, sodium acetate and the effects of their high concentration on Y. lipolytica growth and PHB content, response surface methodology RSM analysis was used [30]. RSM analysis was done using SAS RSREG procedure [31]. For the statistical analysis, linear, second-degree polynomials, and quadratic models were used to describe the relationship between the input variables and the responses. Two-factor ANOVA analysis was used for evaluating the interaction of two independent variables (glucose and sodium acetate) on the responses (growth rate and PHB content). Using SAS GML procedure, the F-value was calculated for detecting the interactions between the variables. In this analysis two factors were considered as cross factors and a completely randomized design (CRD) was used for evaluating the interactions between these two factors.
3. Results
3.1. Metabolic Pathway Design
Comparing the results of ABC PHB biosynthesis pathway (Figure 3A) and NBC pathway (Figure 3B) showed that redirecting the lipogenesis pathway toward PHB production using NBC pathway is a more effective strategy for PHB biosynthesis in Y. lipolytica. As it could be observed in Figure 3C, cultivating the transformed PO1f host strain by pZX30 vector (in YPD media with 30 g/L glucose) led to accumulation of PHB up to 11% of CDW. However, there was no detectable PHB synthesis in transformed strain with pZX22 plasmid. Therefore, the NBC pathway was selected as an efficient design for PHB biosynthesis in Y. lipolytica. One reason for high efficiency of NBC pathway in Y. lipolytica is high carbon flux toward the lipogenesis pathway in this yeast. This characteristic results a significant synthesis of malonyl-CoA as a main intermediate for fatty acids production [32]. Introducing the NphT7 redirects the Y. lipolytica metabolic pathway from lipogenesis to producing more acetoacetyl-CoA as PHB precursor. Considering that no natural replicative vectors (episome) have ever been observed in Y. lipolytica, designed vectors contain chromosomal centromere (cEN) fragments to transient expression of these heterologous genes [33].
The enzymatic reactions which are employed to catalyze the acetoacetyl-CoA synthesis were the main difference between these pathways. Synthesis of acetoacetyl-CoA is usually catalyzed by acetoacetyl-CoA thiolase via the condensation of two molecules of acetyl-CoA [34]. In the first pathway phaA, phaB, and phaC were used as basic PHB production pathway which is called ABC pathway. PhaA (β-ketothiolase) is used for catalyzing the acetyl-CoA condensation reaction to produce acetoacetyl-CoA as precursor for PHB biosynthesis [12]. However, in a study McQualter and colleagues [16] found that using the acetoacetyl-CoA synthase in place of a β-ketothiolase enhances PHB production in transgenic sugarcane. This study showed that introducing the NBC PHB pathway (containing NphT7, phaB, and phaC) instead of ABC PHB pathway into the sugarcane plant increased PHB content in mesophyll cells up to 11.8% of its CDW. It was more than twice the ABC pathway. Studies showed that PhaA preferentially catalysis thiolysis (breaking down the acetoacetyl-CoA to acetyl-CoA) compared to synthesis of acetoacetyl-CoA [35]. On the other hand, NphT7 catalysis acetoacetyl-CoA synthesis in an more energy-favored reaction [34]. Indeed, high equilibrium constant value (Km) of β-ketothiolase to acetyl-CoA (up to 1.1mM) compared to low Km value of NphT7 to acetyl-CoA (0.068mM) and malonyl-CoA (0.028mM) shows that NphT7 is a much more effective enzyme for catalyzing the condensation reaction for producing acetoacetyl-CoA compared to PhaA [16]. On the other hand, PhaA catalytic behavior is highly reversible which lead to high tendency of this enzyme catalyzing the thiolysis reaction and breaking down the acetoacetyl-CoA into two acetyl-CoA molecules, while NphT7 more shows irreversible behavior on catalyzing the condensation reaction [36].
The resulted strain from the sequential transformation showed a significant increase in the PHB content compared to strain modified by pZX30, without any detectable effect on growth rate (Figure 4A). Sequential transformation led to a strain capable of accumulating PHB up to 41% of CDW which is remarkably higher than 11% PHB titer in strain contains the basic PHB biosynthesis pathway (Figure 4B). Indeed, previous studies prove the gene dosage effect on the expression of desired genes in Y. lipolytica host cells. In a study on gene-dose-dependent overexpression of isocitric lyase encoding gene (ICL1), multiple copies of the ICL1 were introduced to Y. lipolytica which led to high levels of ICL1 expression in transformant. This over expression resulted in a significant shift of citric acid/isocitric acid ratio, more than 30% reduction in isocitric acid proportions without affecting the total amount of produced acid [37]. In another study a similar strategy was used for the same objective (increasing the citric acid/isocitric acid ratio), this time by increasing the aconitase encoding gene (ACO1) copy numbers. The recombinant Y. lipolytica strain showed a high-level of aconitase expression which resulted in increasing the citric acid/isocitric acid ratio from 41% to 68% [38]. Studies on the effect of gene dosage for overproduction of β-farnesene and heterologous lipase showed the same results [39,40].
3.2. Co-substrate Strategy for PHB Production
Using acetic acid as the only carbon source led to low growth rate and low biomass productivity which is a serious issue on industrial production of PHAs. Efficient growth of Y. lipolytica requires a balanced supply of ATP, acetyl-CoA and NADPH near to 1:1:2 ratio [41]. Single acetate substrate is very efficient in producing ATP and acetyl-CoA. However, it is an intensive ATP consumer process through oxidative pentose phosphate pathway (Ox-PPP) to produce NADPH from acetate. It is the reason that growing the modified Y. lipolytica on acetate was much slower than glucose [41]. On the other hand, using glucose as the sole energy and carbon source led to significantly higher growth rate and PHB content. Nevertheless, the cost of glucose seems to be a serious obstacle to feasibility of using it as the sole substrate in a cost-effective bioprocess for PHB production [42]. As a result, a co-substrate strategy with a mix of glucose and sodium acetate was evaluated to find the feasibility of this method as a solution for efficiency and cost dilemma. These results showed that high concentrations of both glucose and acetate have inhibitory effects on engineered strain’s growth rate. 10 g/L of sodium acetate (7.2 g/L acetate) as sole carbon source led to growth rate of 0.148 h-1. These results are similar to previous studies on W29 strain Y. lipolytica wild type that showed 0.16 h-1 growth rate cultivated on 5 g/L acetate [43]; but pretty higher than SZYL004, a modified strain for using xylose as substrate (by overexpressing the three native genes XR, XDH, and XK). SZYL004 couldn’t tolerate 75 mM of acetate (6.15 g/L) [44]. However, increasing the sodium acetate concentration to 50 g/L caused a 12% decrease in PHB32 strain growth rate. Further increasing of sodium acetate to 100 g/L (72 g/L acetate) significantly inhibited the engineered strain to grow. In an effort to modify Y. lipolytica to tolerate higher acetate concentration, recombinant strain (expressed acetyl-CoA synthase) tried to be adapted to 50 g/L acetate. The resulting ACS 5.0 strain (after genetic modification and culture adaptation), could grow effectively in 30 g/L of acetic acid, while 40 and 50 g/L acetate yet showed highly toxicity for this strain [45]. Considering that the current engineered strain (PHB32) is able to grow effectively on 36 g/L acetate, with a growth rate of 0.131 h-1, this strain could be comparable to other strains tolerating high acetate concentrations. In addition to high acetate tolerance, some studies had done to modifying PO1f Y. lipolytica strain to increase its growth rate using acetate as sole carbon and energy source. In one study overexpressing the two key enzymes in acetate metabolism (acetyl-CoA synthetase (ACS) and acetyl-CoA carboxylase (ACC1)), led to a strain showed 5.27 times more growth rate on acetate, and 5.39 times faster acetate consumption [46].
Using glucose as the sole carbon and energy source showed significantly higher growth rate. In a flask containing YNB and 10 g/L glucose a growth rate of 0.204 h-1 observed which was 38% higher than the growth rate using 10 g/L of sodium acetate. These results showed a little higher growth rate compared to previous study on cultivating strain Y. lipolytica H222 using glucose for citric acid production which is reached to 0.192 h-1 as the maximum specific growth rate [47]. However, PHB32 grow slower than Y. lipolytica IBT 446 with 0.24 h-1 of growth rate, using same submerged cultivation system with glucose as the sole carbon source [48]. In a comparison between four different strains of Y. lipolytica including A-101, Wratislavia 1.31, Wratislavia AWG7, and Wratislavia K1 for citric acid production from glucose, results showed 0.19 h-1, 0.23 h-1, 0.36 h-1, and 0.23 h-1 of growth rate respectively. These results showed that strain Wratislavia AWG7 had a higher growth rate compared to our engineered strain [49]. Increasing the glucose concentration from 10 g/L to 150 g/L showed a negative effect on growth rate. In 100 g/L of glucose, a growth rate of 0.163 h-1 was observed which is 20% lower than the growth rate at 10 g/L of glucose. Such substrate inhibition effects observed in other studies which showed that increasing the glucose concentration from 50 g/L to 200 g/L lead to an intensive depletion in growth rate from 0.17 h-1 to 0.055 h-1 in H222 strain [47]. Similar inhibitory effect on growth rate and lipid productivity in PO1f strain observed by increasing the glucose concentration to 160 g/L which suggested as a result of osmotic stress [50].
Using co-substrate strategy with glucose and sodium acetate showed a synergistic effect on growth rate. In 10 g/L of both glucose and sodium acetate, highest growth rate of 0.227 h-1 was observed. Such an ameliorative effect on bio-oil production using these two substrates simultaneously observed in Y. lipolytica NCYC 2904. However, this study showed that increasing 5 g/L glucose to media didn’t lead to biomass productivity improvement [51]. Similar study was conducted on using agricultural wastes containing a mixture of stearin, glycerol, and glucose as co-substrates. In the study Y. lipolytica ACA-DC 50109 was modified for producing of a cocoa-butter-like lipid. The results demonstrated that co-substrate media could be an effective strategy for such value added products [52]. Our results showed that in flasks with 100 g/L of sodium acetate as sole substrate, there was not any growth after 6 days, while adding 5 g/L glucose to such flasks led to 0.033 h-1 of growth rate. Similarly, in flasks with 50 g/L sodium acetate as mono-substrate a 0.131 h-1 growth rate observed while using co-substrate strategy with same sodium acetate concentration and 5 g/L glucose as dopant caused a 40% increase in growth rate up to 0.184 h-1.
Metabolism, stress-signaling pathways, and pH homeostasis are main factors affecting how yeast respond to acetic acid stress [53]. Among other physiological features, cell membrane transporters play an important role in yeast tolerance to high acetate concentrations [54]. Indeed, if the environmental pH is lower than acetic acid pKa (4.76), acetic acid exists mainly in the undissociated form which could be easily diffused into cell membrane and cytosol consequently [55]. Increasing the extracellular concentrations of acetate also could lead to high passive transport of acetate into cytoplasm [56]. It results intracellular acidification and lead to growth and metabolic activity inhibition, and oxidative damages. In this condition pH homeostasis stimulates the proton pumps to prevent further intracellular proton accumulation. Such active transport is a high energy-consuming process, which consume up to 20% of the produced ATP [57].
Our results showed a similar compensatory effect of acetate in high concentrations of glucose. It was observed that adding 10 g/L of sodium acetate in all initial concentrations of glucose significantly increased the growth rate. For example, while 100 g/L of glucose had some inhibitory effect on engineered strain, adding 10 g/L sodium acetate increased its growth rate up to 15%. Co-substrate strategy using multiple carbon and energy sources can balance key biosynthetic components. Synthetic pathways in cells are generally complex processes which are involving various types of molecules as their building blocks. Considering the nature of metabolism, each pathway showed different degrees of efficiency on converting the various substrates to the products. In the co-substrate concept, glucose is commonly considered as an energy deficient substrate (high carbon-to-energy ratio). This deficiency could be compensated by simultaneous use of a substrate (such as C1 substrates) which has a lower carbon-to-energy ratio as convenient energy donors [59]. For instance, in a study on gluconate doping in Y. lipolytica results showed that co-utilizing of this substrate caused an accelerated acetate lipogenesis by providing NADPH through pentose cycle [41]. Main reason for toxicity of glucose in high concentrations is the resulting osmostress which seems that acetate has a compensatory effect on it [60]. Previous studies showed that acetate could increase the intracellular proline concentration which is considered as an effective cellular osmoprotectant agent [61]. Another study on the effect of acetate on Lens culinaris showed that acetate increases the cellular accumulation of proline up to 24% and also it led to increasing the ascorbic acid content more than 2 times and catalase activity up to 1.5 times which both considered antioxidant agents [62]. Such results suggest that co-utilization of acetate has a compensatory effect for glucose toxicity in high concentrations because it triggers cells to accumulate proline with osmoprotective effect and increase the antioxidants’ concentrations and activity.
The interaction between glucose and acetate may play a key role in a co-substrate strategy as indicated in primary analysis. In order to reveal such interaction between acetate and glucose on modified strain’s growth rate, a two-factor design using General Linear Model (GLM) was employed. The resulting ANOVA analysis (Table 2) showed that acetate and glucose concentration had significant interactive effects on Y. lipolytica growth rate.
As shown in Table 2, both glucose (p<0.0001) and acetate (p<0.0001) had significant effects on biomass concentration. Also, these results suggest that glucose and acetate had a significant interaction (FGlucose*Acetate=105.0, p<0.0001) on engineered Y. lipolytica strain growth rate and biomass productivity.
PHB synthesized by engineered Y. lipolytica showed to be a growth associated product which was accumulated in produced biomass simultaneously with the yeast growth. Therefore, in order to find the optimal medium for the PHB production in a fed-batch system, response surface methodology (RSM) was used. The 3D surface plot and 2D contour plot shown in Figure 5 are graphical representations of statistical regression and growth rate prediction. This statistical modeling and 2D contour plot suggested 50 g/L glucose and 10 to 20 g/L sodium acetate as the optimum concentrations of these substrates using in a co-substrate strategy (Figure 5A). 3D surface plot predicted that increasing the concentration of both glucose and acetate have inhibitory effects on this strain’s growth rate.
3.3. Bioreactor Production of PHB
During the aerobic fermentation using Y. lipolytica PHB32 strain (in a controlled pH and co-substrate medium), fed-batch system used for maintaining the substrates concentration on their optimal level (Figure 6A). Initial concentration of glucose and sodium acetate set at 50 g/L and 20 g/L based on RSM optimal prediction (Figure 6B). Using this system, the CDW reached to 83.4 g/L with a noticeable high PHB titer of 31.7 g/L (Figure 6C).
Results showed that the polymer content in fed-batch culture and flask experiment was almost same. This fact suggested that changing the pH and also nutrient starvation during flask cultivation (as main inhibitory parameters in batch culture) did not affect the expression of the introduced PHB pathway. Similar results in previous studies suggest that PHB accumulation capacity in engineered Y. lipolytica is mainly restricted by expression level of introduced heterologous PHB pathway [12]. In our study, PHB content in the bioreactor experiment using YPD medium reached to 38% of CDW in modified Y. lipolytica PHB32. These results are significantly higher than that previously engineered Y. lipolytica CAB strain using ABC pathway [12]. Y. lipolytica PHB32 showed noticeably high capacity for PHB production not only for its high PHB titer, but also high biomass productivity even using minimal YNB medium. In an attempt, instead of using either ABC or NBC pathway, just PHA synthase (phaC) of Pseudomonas aeruginosa was introduced to this yeast which led to 2% PHA accumulation in produced biomass. Result of Haddouce and colleagues study showed that between six acyl-CoA oxidase (Aox) isoenzymes, Aox3p has a key role on converting C9:0 and C13:0 fatty acids into PHA [63]. Similar study used variant PhaC as the single heterologous gene for synthesizing medium-chain-length (mcl) PHA in engineered Y. lipolytica. Up to 25% (g/g) of PHA accumulation observed in some of these PhaC variants. In this study some of the modified strains were capable to synthesis mcl-PHA with an average molar mass of 316,000 g/mol which has a high potential in packaging applications [14]. Multicopy integration PhaC1 (P. aeruginosa PAO1) into Y. lipolytica led to an engineered strain which could accumulate up to 5% mcl-PHA in its biomass. In this study the highest biomass concentration reached to 21.9 g/L of CDW using YNB medium [13]. Among these strategies NBC pathway seems to be a more efficient pathway for PHB biosynthesis in Y. lipolytica.
4. Conclusion
NBC PHB pathway was introduced into the Y. lipolytica genome. The resulting engineered strain showed to be capable of accumulating PHB up to 41 % of CDW in shake flask culture using glucose and acetate as substrate. These results suggest that the NBC PHB pathway is a more efficient strategy compared to the ABC PHB pathway or introducing PhaC as a single gene pathway in Y. lipolytica. Using acetate or glucose as sole carbon and energy source showed slow growth rate and inhibitory effects in high concentrations. Co-substrate strategy using both substrates simultaneously increased the modified strain’s growth rate significantly. Simultaneous use of glucose and acetate showed an alleviative effect for substrate inhibition which suggests co-substrate strategy as a reasonable method for cultivating Y. lipolytica in high concentrations of substrate. For bench-scale production of PHB co-substrate strategy with a constant pH in a fed-batch fermentation system was used. During the fermentation, substrates’ concentrations maintained at optimal condition (based on RSM results). Bioreactor cultivation led to 83.4 g/L biomass and 31.7 g/L PHB concentration which is shows noticeably higher performance in PHB productivity using Y. lipolytica compared to previous studies.
Author Contributions
Conceptualization, M.T. and S.C.; writing—original draft preparation, M.T. and S.S.; writing—review and editing, S.C., M.T., and S.S.; supervision, S.C.; project administration, S.C.; funding acquisition, S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the USDA Small Business Innovation Research (SBIR) program and WSU.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- OECD Plastic Waste Generation in the United States in 2019, with Projections for 2030 and 2060 (in Million Metric Tons) [Graph]. Available online: https://www.statista.com/statistics/1339223/us-plastic-waste-generation-outlook/ (accessed on 8 September 2023).
- Karan, H.; Funk, C.; Grabert, M.; Oey, M.; Hankamer, B. Green Bioplastics as Part of a Circular Bioeconomy. Trends Plant Sci. 2019, 24, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Sehgal, R.; Gupta, R. Polyhydroxyalkanoate (PHA): Properties and Modifications. Polymer 2020, 212, 123161. [Google Scholar] [CrossRef]
- Vicente, D.; Proença, D.N.; Morais, P. V The Role of Bacterial Polyhydroalkanoate (PHA) in a Sustainable Future: A Review on the Biological Diversity. Int. J. Environ. Res. Public Health 2023, 20, 2959. [Google Scholar] [CrossRef] [PubMed]
- Abedi, E.; Hashemi, S.M.B. Lactic acid production – producing microorganisms and substrates sources-state of art. Heliyon 2020, 6. [Google Scholar] [CrossRef]
- Volk, M.J.; Tran, V.G.; Tan, S.-I.; Mishra, S.; Fatma, Z.; Boob, A.; Li, H.; Xue, P.; Martin, T.A.; Zhao, H. Metabolic Engineering: Methodologies and Applications. Chem. Rev. 2022, 123, 5521–5570. [Google Scholar] [CrossRef]
- Park, Y.-K.; Ledesma-Amaro, R. What Makes Yarrowia Lipolytica Well Suited for Industry? Trends Biotechnol. 2022. [CrossRef]
- Lindquist, M.R.; López-Núñez, J.C.; Jones, M.A.; Cox, E.J.; Pinkelman, R.J.; Bang, S.S.; Moser, B.R.; Jackson, M.A.; Iten, L.B.; Kurtzman, C.P.; et al. Irradiation of Yarrowia lipolytica NRRL YB-567 creating novel strains with enhanced ammonia and oil production on protein and carbohydrate substrates. Appl. Microbiol. Biotechnol. 2015, 99, 9723–9743. [Google Scholar] [CrossRef]
- Celińska, E. “Fight-Flight-or-Freeze”–How Yarrowia Lipolytica Responds to Stress at Molecular Level? Appl. Microbiol. Biotechnol. 2022, 106, 3369–3395. [Google Scholar] [CrossRef]
- Liu, Q.; Siloto, R.M.P.; Lehner, R.; Stone, S.J.; Weselake, R.J. Acyl-CoA: Diacylglycerol Acyltransferase: Molecular Biology, Biochemistry and Biotechnology. Prog. Lipid Res. 2012, 51, 350–377. [Google Scholar] [CrossRef]
- Li, Z.-J.; Wang, Y.-Z.; Wang, L.-R.; Shi, T.-Q.; Sun, X.-M.; Huang, H. Advanced Strategies for the Synthesis of Terpenoids in Yarrowia lipolytica. J. Agric. Food Chem. 2021, 69, 2367–2381. [Google Scholar] [CrossRef]
- Li, Z.-J.; Qiao, K.; Liu, N.; Stephanopoulos, G. Engineering Yarrowia lipolytica for poly-3-hydroxybutyrate production. J. Ind. Microbiol. Biotechnol. 2017, 44, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Qi, Q.; Madzak, C.; Lin, C.S.K. Exploring medium-chain-length polyhydroxyalkanoates production in the engineered yeast Yarrowia lipolytica. J. Ind. Microbiol. Biotechnol. 2015, 42, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Rigouin, C.; Lajus, S.; Ocando, C.; Borsenberger, V.; Nicaud, J.M.; Marty, A.; Avérous, L.; Bordes, F. Production and characterization of two medium-chain-length polydroxyalkanoates by engineered strains of Yarrowia lipolytica. Microb. Cell Factories 2019, 18, 99. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Mrozova, Z.; Thevenieau, F.; Le Dall, M.-T.; Hapala, I.; Papanikolaou, S.; Chardot, T.; Nicaud, J.-M. Control of Lipid Accumulation in the Yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 7779–7789. [Google Scholar] [CrossRef] [PubMed]
- McQualter, R.B.; Petrasovits, L.A.; Gebbie, L.K.; Schweitzer, D.; Blackman, D.M.; Chrysanthopoulos, P.; Hodson, M.P.; Plan, M.R.; Riches, J.D.; Snell, K.D.; et al. The use of an acetoacetyl-CoA synthase in place of a β-ketothiolase enhances poly-3-hydroxybutyrate production in sugarcane mesophyll cells. Plant Biotechnol. J. 2014, 13, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Yang, X.; Wang, H.; Rivero, C.P.; Li, C.; Cui, Z.; Qi, Q.; Lin, C.S.K. Robust succinic acid production from crude glycerol using engineered Yarrowia lipolytica. Biotechnol. Biofuels 2016, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, M.; Hussain, M.S.; Gambill, L.; Blenner, M. Alternative Substrate Metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 9, 1077. [Google Scholar] [CrossRef]
- Qin, Y.; Higashimori, A.; Wu, L.-J.; Hojo, T.; Kubota, K.; Li, Y.-Y. Phase separation and microbial distribution in the hyperthermophilic-mesophilic-type temperature-phased anaerobic digestion (TPAD) of waste activated sludge (WAS). Bioresour. Technol. 2017, 245, 401–410. [Google Scholar] [CrossRef]
- García-Depraect, O.; Bordel, S.; Lebrero, R.; Santos-Beneit, F.; Börner, R.A.; Börner, T.; Muñoz, R. Inspired by nature: Microbial production, degradation and valorization of biodegradable bioplastics for life-cycle-engineered products. Biotechnol. Adv. 2021, 53, 107772. [Google Scholar] [CrossRef]
- Chang, D.-E.; Shin, S.; Rhee, J.-S.; Pan, J.-G. Acetate Metabolism in a pta Mutant of Escherichia coli W3110: Importance of Maintaining Acetyl Coenzyme A Flux for Growth and Survival. J. Bacteriol. 1999, 181, 6656–6663. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Sabirova, J.; Soetaert, W.; Ki Carol Lin, S. Polyhydroxyalkanoates Production from Low-Cost Sustainable Raw Materials. Curr. Chem. Biol. 2012, 6, 14–25. [Google Scholar]
- Ji, Q.; Mai, J.; Ding, Y.; Wei, Y.; Ledesma-Amaro, R.; Ji, X.-J. Improving the homologous recombination efficiency of Yarrowia lipolytica by grafting heterologous component from Saccharomyces cerevisiae. Metab. Eng. Commun. 2020, 11, e00152. [Google Scholar] [CrossRef] [PubMed]
- Bredeweg, E.L.; Pomraning, K.R.; Dai, Z.; Nielsen, J.; Kerkhoven, E.J.; Baker, S.E. A molecular genetic toolbox for Yarrowia lipolytica. Biotechnol. Biofuels 2017, 10, 1–22. [Google Scholar] [CrossRef]
- Shi, X.; Park, H.M.; Kim, M.; Lee, M.-E.; Jeong, W.-Y.; Chang, J.; Cho, B.-H.; Han, S.O. Isopropanol biosynthesis from crude glycerol using fatty acid precursors via engineered oleaginous yeast Yarrowia lipolytica. Microb. Cell Factories 2022, 21, 1–11. [Google Scholar] [CrossRef]
- Slater, S.; Houmiel, K.L.; Tran, M.; Mitsky, T.A.; Taylor, N.B.; Padgette, S.R.; Gruys, K.J. Multiple β-Ketothiolases Mediate Poly (β-Hydroxyalkanoate) Copolymer Synthesis in Ralstonia Eutropha. J. Bacteriol. 1998, 180, 1979–1987. [Google Scholar] [CrossRef] [PubMed]
- DeLoache, W.C.; Russ, Z.N.; Dueber, J.E. Towards repurposing the yeast peroxisome for compartmentalizing heterologous metabolic pathways. Nat. Commun. 2016, 7, 11152. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, S.; Xiong, X. Metabolic Engineering of Non-carotenoid-Producing Yeast Yarrowia lipolytica for the Biosynthesis of Zeaxanthin. Front. Microbiol. 2021, 12, 699235. [Google Scholar] [CrossRef]
- Ye, R.W.; Sharpe, P.L.; Zhu, Q. Bioengineering of Oleaginous Yeast Yarrowia Lipolytica for Lycopene Production. Microb. carotenoids from fungi methods Protoc. 2012, 153–159. [Google Scholar]
- Tourang, M.; Baghdadi, M.; Torang, A.; Sarkhosh, S. Optimization of carbohydrate productivity of Spirulina microalgae as a potential feedstock for bioethanol production. Int. J. Environ. Sci. Technol. 2017, 16, 1303–1318. [Google Scholar] [CrossRef]
- Amir, W.M.; Shafiq, M.; Mokhtar, K.; Aleng, N.A.; Rahim, H.A. RSREG (SAS). J. Mod. Appl. Stat. Methods 2016, 15, 855–867. [Google Scholar] [CrossRef]
- Liu, H.; Marsafari, M.; Deng, L.; Xu, P. Understanding lipogenesis by dynamically profiling transcriptional activity of lipogenic promoters in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2019, 103, 3167–3179. [Google Scholar] [CrossRef] [PubMed]
- Larroude, M.; Rossignol, T.; Nicaud, J.-M.; Ledesma-Amaro, R. Synthetic biology tools for engineering Yarrowia lipolytica. Biotechnol. Adv. 2018, 36, 2150–2164. [Google Scholar] [CrossRef] [PubMed]
- Okamura, E.; Tomita, T.; Sawa, R.; Nishiyama, M.; Kuzuyama, T. Unprecedented acetoacetyl-coenzyme A synthesizing enzyme of the thiolase superfamily involved in the mevalonate pathway. Proc. Natl. Acad. Sci. 2010, 107, 11265–11270. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.J.; Rock, C.O. The Claisen condensation in biology. Nat. Prod. Rep. 2002, 19, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Chau, T.H.T.; Nguyen, A.D.; Lee, E.Y. Engineering type I methanotrophic bacteria as novel platform for sustainable production of 3-hydroxybutyrate and biodegradable polyhydroxybutyrate from methane and xylose. Bioresour. Technol. 2022, 363, 127898. [Google Scholar] [CrossRef]
- Förster, A.; Jacobs, K.; Juretzek, T.; Mauersberger, S.; Barth, G. Overexpression of the ICL1 gene changes the product ratio of citric acid production by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2007, 77, 861–869. [Google Scholar] [CrossRef]
- Holz, M.; Förster, A.; Mauersberger, S.; Barth, G. Aconitase overexpression changes the product ratio of citric acid production by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2009, 81, 1087–1096. [Google Scholar] [CrossRef]
- Zhou, Q.; Jiao, L.; Qiao, Y.; Wang, Y.; Xu, L.; Yan, J.; Yan, Y. Overexpression of GRAS Rhizomucor miehei lipase in Yarrowia lipolytica via optimizing promoter, gene dosage and fermentation parameters. J. Biotechnol. 2019, 306, 16–23. [Google Scholar] [CrossRef]
- Shi, T.; Li, Y.; Zhu, L.; Tong, Y.; Yang, J.; Fang, Y.; Wang, M.; Zhang, J.; Jiang, Y.; Yang, S. Engineering the Oleaginous Yeast Yarrowia Lipolytica for Β-farnesene Overproduction. Biotechnol. J. 2021, 16, 2100097. [Google Scholar] [CrossRef]
- Park, J.O.; Liu, N.; Holinski, K.M.; Emerson, D.F.; Qiao, K.; Woolston, B.M.; Xu, J.; Lazar, Z.; Islam, M.A.; Vidoudez, C.; et al. Synergistic substrate cofeeding stimulates reductive metabolism. Nat. Metab. 2019, 1, 643–651. [Google Scholar] [CrossRef]
- Bhola, S.; Arora, K.; Kulshrestha, S.; Mehariya, S.; Bhatia, R.K.; Kaur, P.; Kumar, P. Established and Emerging Producers of PHA: Redefining the Possibility. Appl. Biochem. Biotechnol. 2021, 193, 3812–3854. [Google Scholar] [CrossRef]
- Park, Y.-K.; Dulermo, T.; Ledesma-Amaro, R.; Nicaud, J.-M. Optimization of odd chain fatty acid production by Yarrowia lipolytica. Biotechnol. Biofuels 2018, 11, 158. [Google Scholar] [CrossRef]
- Konzock, O.; Zaghen, S.; Norbeck, J. Tolerance of Yarrowia lipolytica to inhibitors commonly found in lignocellulosic hydrolysates. BMC Microbiol. 2021, 21, 1–10. [Google Scholar] [CrossRef]
- Narisetty, V.; Prabhu, A.A.; Bommareddy, R.R.; Cox, R.; Agrawal, D.; Misra, A.; Haider, M.A.; Bhatnagar, A.; Pandey, A.; Kumar, V. Development of Hypertolerant Strain of Yarrowia lipolytica Accumulating Succinic Acid Using High Levels of Acetate. ACS Sustain. Chem. Eng. 2022, 10, 10858–10869. [Google Scholar] [CrossRef]
- Chen, L.; Yan, W.; Qian, X.; Chen, M.; Zhang, X.; Xin, F.; Zhang, W.; Jiang, M.; Ochsenreither, K. Increased Lipid Production in Yarrowia lipolytica from Acetate through Metabolic Engineering and Cosubstrate Fermentation. ACS Synth. Biol. 2021, 10, 3129–3138. [Google Scholar] [CrossRef]
- Moeller, L.; Strehlitz, B.; Aurich, A.; Zehnsdorf, A.; Bley, T. Optimization of Citric Acid Production from Glucose byYarrowia lipolytica. Eng. Life Sci. 2007, 7, 504–511. [Google Scholar] [CrossRef]
- Workman, M.; Holt, P.; Thykaer, J. Comparing cellular performance of Yarrowia lipolytica during growth on glucose and glycerol in submerged cultivations. AMB Express 2013, 3, 58. [Google Scholar] [CrossRef]
- Rywińska, A.; Rymowicz, W.; Żarowska, B.; Skrzypiński, A. Comparison of Citric Acid Production from Glycerol and Glucose by Different Strains of Yarrowia Lipolytica. World J. Microbiol. Biotechnol. 2010, 26, 1217–1224. [Google Scholar] [CrossRef]
- Blazeck, J.; Hill, A.; Liu, L.; Knight, R.; Miller, J.; Pan, A.; Otoupal, P.; Alper, H.S. Harnessing Yarrowia lipolytica lipogenesis to create a platform for lipid and biofuel production. Nat. Commun. 2014, 5, 3131. [Google Scholar] [CrossRef]
- Pereira, A.S.; Lopes, M.; Miranda, S.M.; Belo, I. Bio-oil production for biodiesel industry by Yarrowia lipolytica from volatile fatty acids in two-stage batch culture. Appl. Microbiol. Biotechnol. 2022, 106, 2869–2881. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Accumulation of a Cocoa-Butter-Like Lipid by Yarrowia lipolytica Cultivated on Agro-Industrial Residues. Curr. Microbiol. 2003, 46, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Guaragnella, N.; Stirpe, M.; Marzulli, D.; Mazzoni, C.; Giannattasio, S. Acid Stress Triggers Resistance to Acetic Acid-Induced Regulated Cell Death throughHog1Activation Which RequiresRTG2in Yeast. Oxidative Med. Cell. Longev. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Piper, P.; Mahé, Y.; Thompson, S.; Pandjaitan, R.; Holyoak, C.; Egner, R.; Mühlbauer, M.; Coote, P.; Kuchler, K. The pdr12 ABC transporter is required for the development of weak organic acid resistance in yeast. EMBO J. 1998, 17, 4257–4265. [Google Scholar] [CrossRef] [PubMed]
- van Beilen, J.W.A.; de Mattos, M.J.T.; Hellingwerf, K.J.; Brul, S. Distinct Effects of Sorbic Acid and Acetic Acid on the Electrophysiology and Metabolism of Bacillus subtilis. Appl. Environ. Microbiol. 2014, 80, 5918–5926. [Google Scholar] [CrossRef]
- Axe, D.D.; Bailey, J.E. Transport of lactate and acetate through the energized cytoplasmic membrane of Escherichia coli. Biotechnol. Bioeng. 1995, 47, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Guaragnella, N.; Bettiga, M. Acetic acid stress in budding yeast: From molecular mechanisms to applications. Yeast 2021, 38, 391–400. [Google Scholar] [CrossRef]
- Chung, K.-J.; Hwang, S.-K.; Hahn, B.-S.; Kim, K.-H.; Kim, J.-B.; Kim, Y.-H.; Yang, J.-S.; Ha, S.-H. Authentic seed-specific activity of the Perilla oleosin 19 gene promoter in transgenic Arabidopsis. Cell Biol. Morphog. 2007, 27, 29–37. [Google Scholar] [CrossRef]
- Liu, N.; Santala, S.; Stephanopoulos, G. Mixed Carbon Substrates: A Necessary Nuisance or a Missed Opportunity? Curr. Opin. Biotechnol. 2020, 62, 15–21. [Google Scholar] [CrossRef]
- Gomar-Alba, M.; Morcillo-Parra, M.Á.; Olmo, M. lí del Response of Yeast Cells to High Glucose Involves Molecular and Physiological Differences When Compared to Other Osmostress Conditions. FEMS Yeast Res. 2015, 15, fov039. [Google Scholar] [CrossRef]
- Rahman, M.; Mostofa, M.G.; Rahman, A.; Islam, R.; Keya, S.S.; Das, A.K.; Miah, G.; Kawser, A.Q.M.R.; Ahsan, S.M.; Hashem, A.; et al. Acetic acid: a cost-effective agent for mitigation of seawater-induced salt toxicity in mung bean. Sci. Rep. 2019, 9, 15186. [Google Scholar] [CrossRef]
- Hossain, S.; Abdelrahman, M.; Tran, C.D.; Nguyen, K.H.; Chu, H.D.; Watanabe, Y.; Fujita, M.; Tran, L.-S.P. Modulation of osmoprotection and antioxidant defense by exogenously applied acetate enhances cadmium stress tolerance in lentil seedlings. Environ. Pollut. 2022, 308, 119687. [Google Scholar] [CrossRef] [PubMed]
- Haddouche, R.; Delessert, S.; Sabirova, J.; Neuvéglise, C.; Poirier, Y.; Nicaud, J.-M. Roles of Multiple Acyl-CoA Oxidases in the Routing of Carbon Flow towards β-Oxidation and Polyhydroxyalkanoate Biosynthesis in Yarrowia Lipolytica. FEMS Yeast Res. 2010, 10, 917–927. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A) Designed pathway for producing PHB from acetyl-CoA as the main precursor. B) Map of pCV35 shuttle vector containing NphT7, phaC1437, and phaB genes; LEU as auxotrophic biomarker; TEF, GPM, and FBA promoters; XPR2, OCT1, and LIP terminators. C) Map of pCV88 shuttle vector containing similar genes, promoters, and terminators to pCV35; with URA3 as auxotrophic biomarker. D) Map of pCV342 shuttle vector containing phaB, phaC1437-SKL, and BktB genes; hygromycin as antibiotic biomarker; TEF promoter and XPR2 terminator.
Figure 1.
A) Designed pathway for producing PHB from acetyl-CoA as the main precursor. B) Map of pCV35 shuttle vector containing NphT7, phaC1437, and phaB genes; LEU as auxotrophic biomarker; TEF, GPM, and FBA promoters; XPR2, OCT1, and LIP terminators. C) Map of pCV88 shuttle vector containing similar genes, promoters, and terminators to pCV35; with URA3 as auxotrophic biomarker. D) Map of pCV342 shuttle vector containing phaB, phaC1437-SKL, and BktB genes; hygromycin as antibiotic biomarker; TEF promoter and XPR2 terminator.
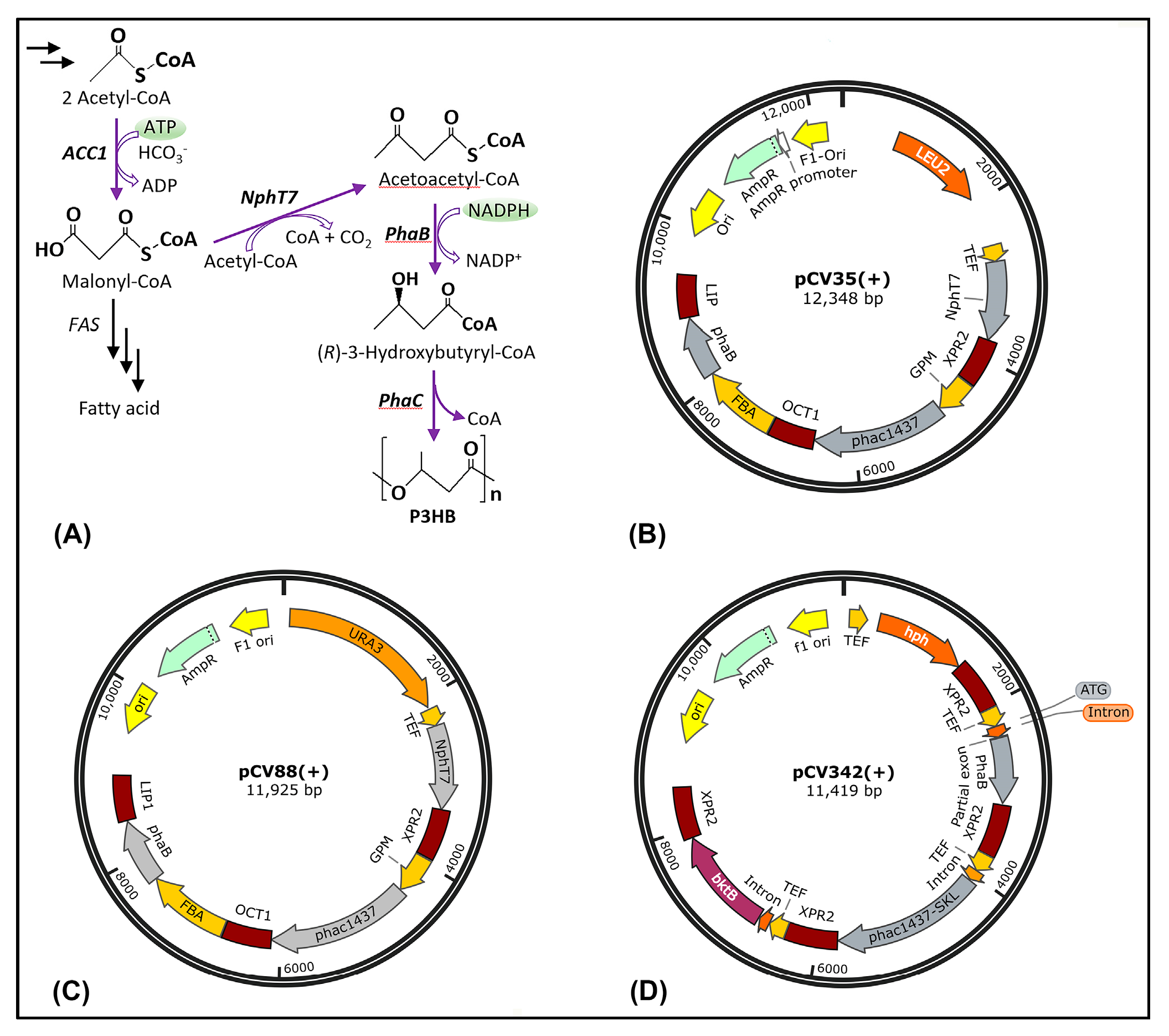
Figure 2.
Transesterification of PHB polymer using methanolysis solution. .
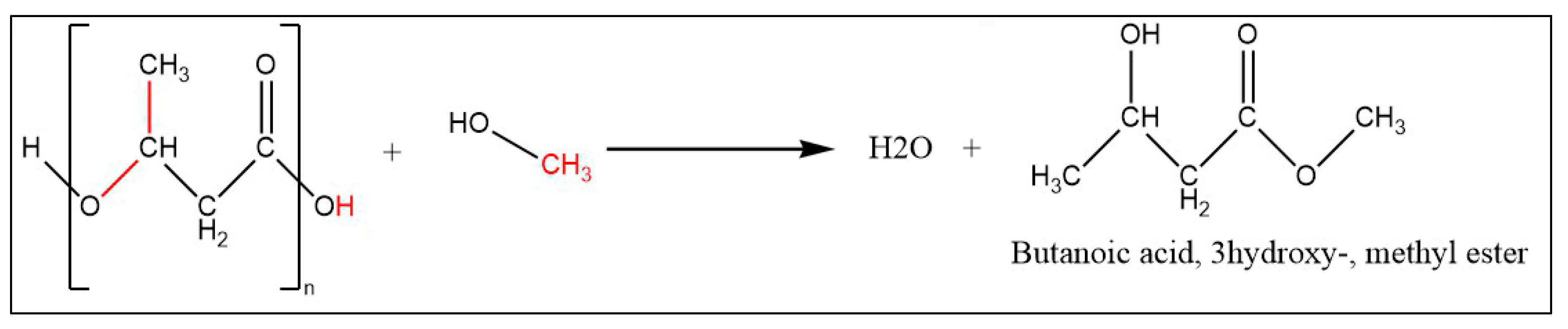
Figure 3.
A) Map of pZX22 vector containing ABC pathway, B) Map of pZX30 vector containing NBC pathway, C) Comparing Biomass concentration and PHB accumulation in modified Y. lipolytica strains using ABC and NBC pathways.
Figure 3.
A) Map of pZX22 vector containing ABC pathway, B) Map of pZX30 vector containing NBC pathway, C) Comparing Biomass concentration and PHB accumulation in modified Y. lipolytica strains using ABC and NBC pathways.
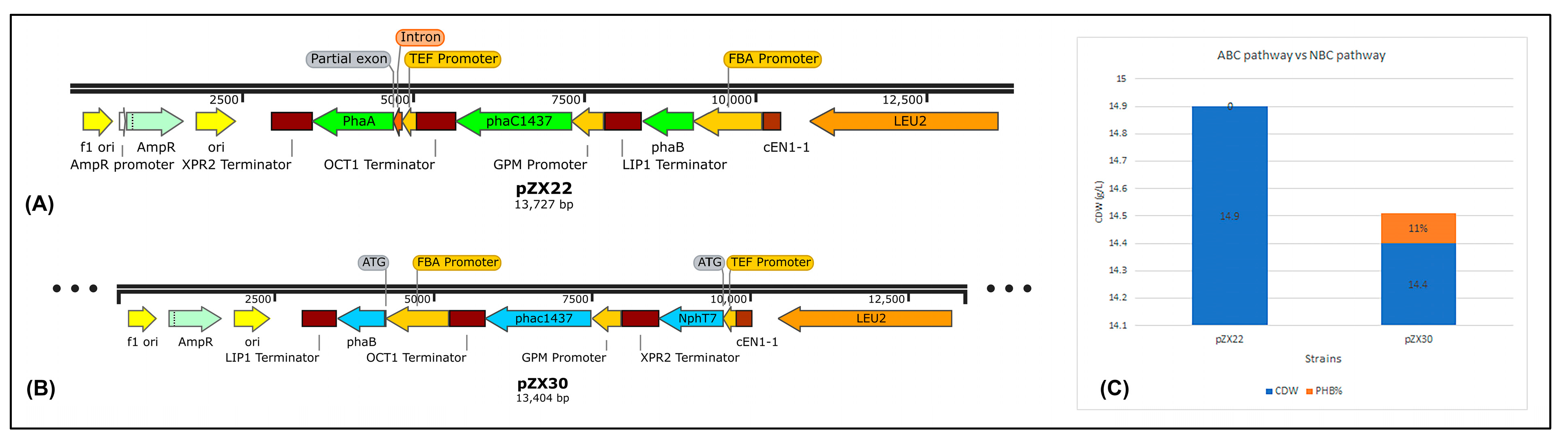
Figure 4.
A) Comparing the growth rate, and B) PHB content; in strain pZX30 (containing only NBC pathway) and strain PHB32 (result of sequential transformation) cultivated on YNB containing 50g/L glucose.
Figure 4.
A) Comparing the growth rate, and B) PHB content; in strain pZX30 (containing only NBC pathway) and strain PHB32 (result of sequential transformation) cultivated on YNB containing 50g/L glucose.
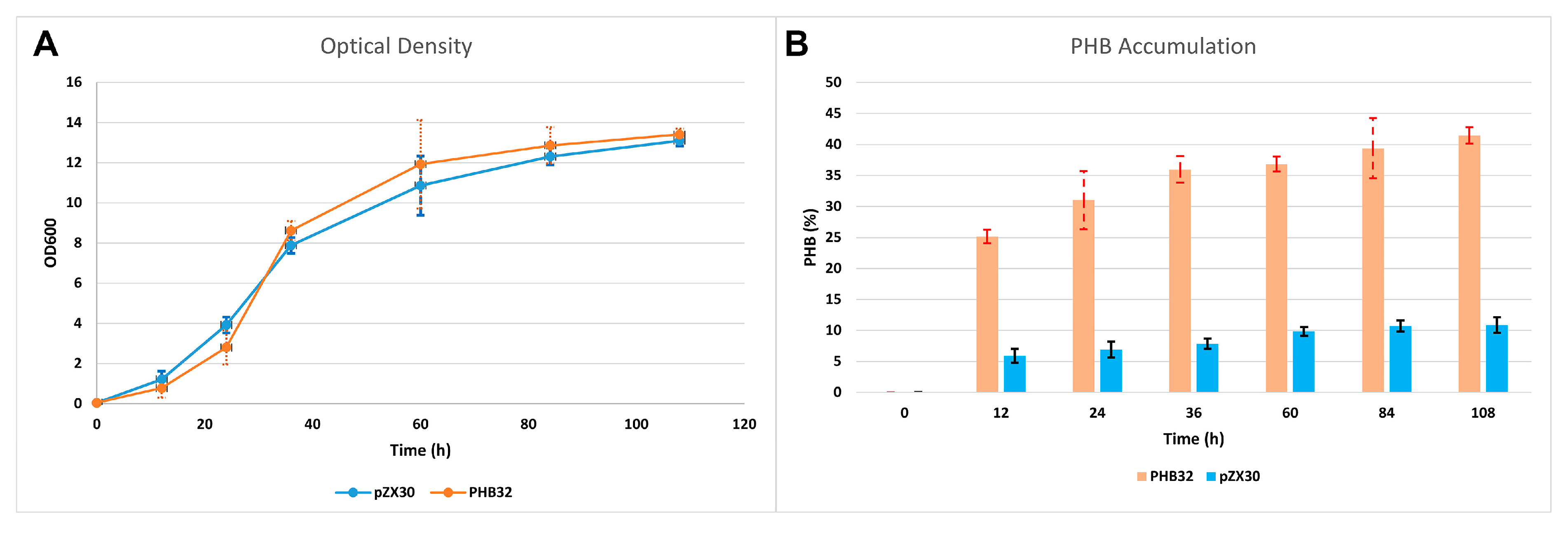
Figure 5.
A) 2D contour plot indicating the optimum levels of substrates for maximum growth rate, B) 3D surface plot shows the effect of glucose and sodium acetate on modified strain’s growth rate.
Figure 5.
A) 2D contour plot indicating the optimum levels of substrates for maximum growth rate, B) 3D surface plot shows the effect of glucose and sodium acetate on modified strain’s growth rate.
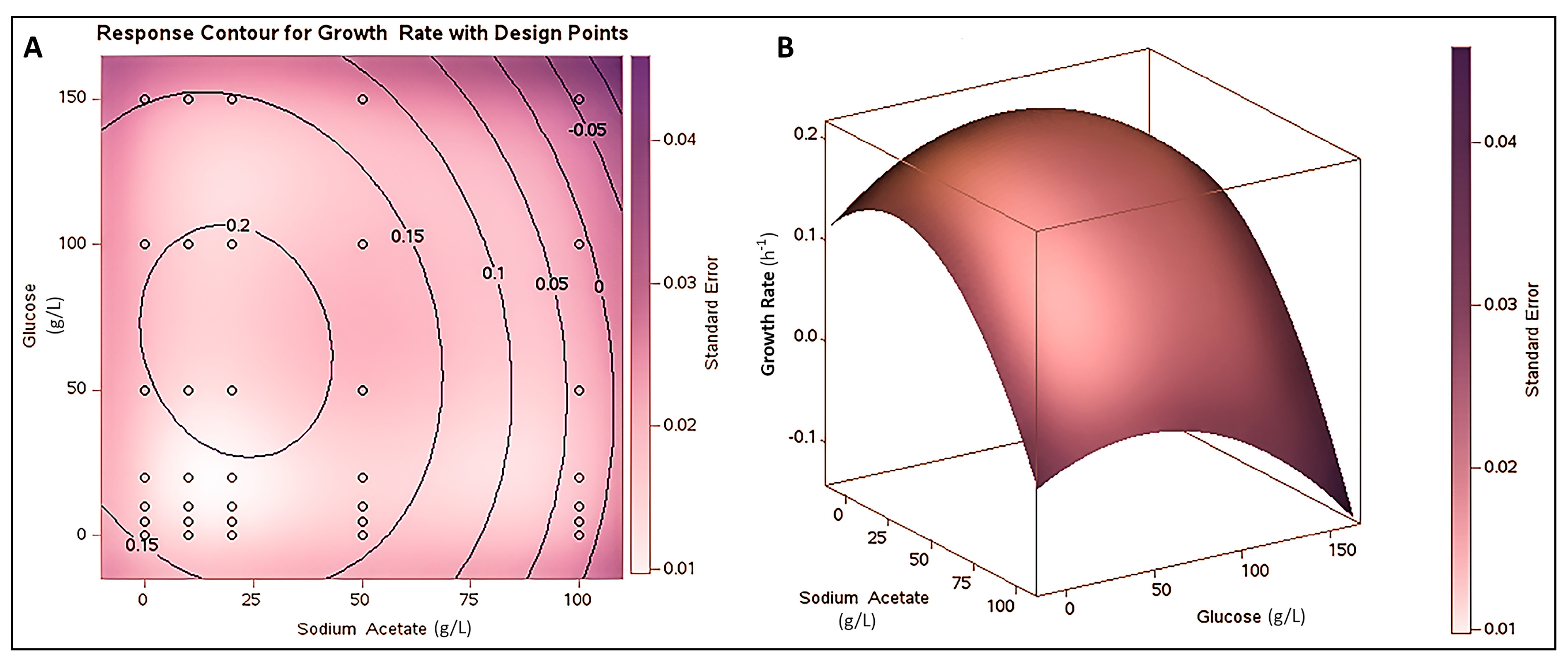
Figure 6.
A) Schematic diagram of the fed-batch bioreactor, B) Bioreactor before inoculation and fermentation, C) Bioreactor at the end of fermentation using Y. lipolytica PHB32.
Figure 6.
A) Schematic diagram of the fed-batch bioreactor, B) Bioreactor before inoculation and fermentation, C) Bioreactor at the end of fermentation using Y. lipolytica PHB32.
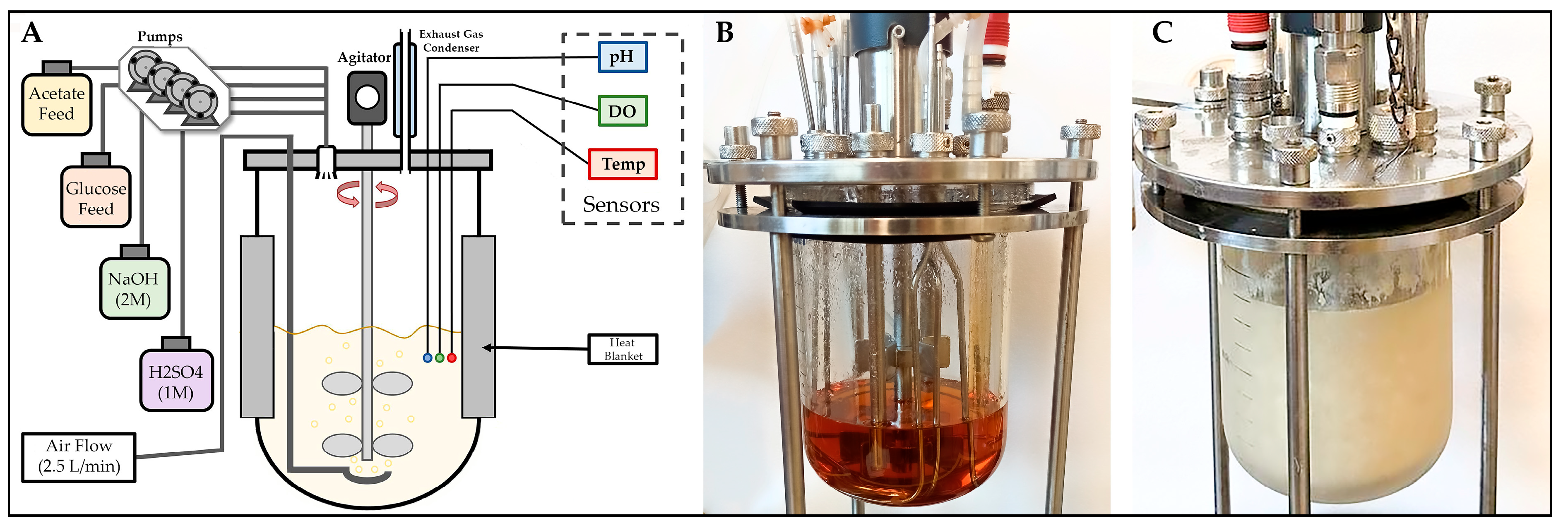

Table 1.
Oligonucleotides used as primers.
| Primers | Sequences (5'‒3') | GC (%) | Length (mer) | Molecular Mass (Da) |
|---|---|---|---|---|
| phaA-F | CCCGGGTTACTTTCTCTCGACGGCC | 64 | 25 | 7560.9 |
| phaA-R | AAGCTTATGACCGACGTGGTGATCGTGTCC | 53 | 30 | 9238.1 |
| phaB-F | CCCGGGAGAGTCGACCTGCAGTTAGCCCATGTGC | 65 | 34 | 10444.8 |
| phaB-R | AAGCTTATGACCCAGCGAATCGCCTACGTCACC | 55 | 33 | 10027.5 |
| phaC1437-F | CCCGGGTTATCGCTCGTGGACGTAAGTGCC | 63 | 30 | 9215.0 |
| phaC1437-R | AAGCTTATGTCTAACAAGTCTAACGACGAGC | 42 | 31 | 9512.3 |
| NphT7-F | CCCGGGTTACCACTCAATCAGGGCAAAGGAAGC | 58 | 33 | 10141.6 |
| NphT7-R | AAGCTTACTGATGTGCGATTTCGAATTATTGGAAC | 37 | 35 | 10800.1 |
| bktB-F | AAGCTTATGACCCGAGAGGTGGTGGTCGTGTCCGG | 60 | 35 | 10884.1 |
| bktB-R | CCCGGGTTAAATTCGTTCGAAGATGGCGGCAATGCC | 56 | 36 | 11101.3 |
| phaC1437-SKL-F | AAGCTTATGTCTAACAAGTCTAACGACGAGCTGAAGTACC | 43 | 40 | 12297.1 |
| phaC1437-SKL-R | CCCGGGTTACAGCTTAGATCGCTCGTGGACGTAAGTGCCAGG | 60 | 42 | 12971.4 |
Table 2.
ANOVA results for two-factor GLM between glucose and acetate on Y. lipolytica PHB32 growth rate.
Table 2.
ANOVA results for two-factor GLM between glucose and acetate on Y. lipolytica PHB32 growth rate.
| Source | SS | df | MS | F-value | P-value |
|---|---|---|---|---|---|
| Glucose | 0.110 | 6 | 0.0183 | 591.4 | <0.0001 |
| Acetate | 0.459 | 4 | 0.1148 | 3702 | <0.0001 |
| Glucose*Acetate | 0.078 | 24 | 0.00325 | 105.0 | <0.0001 |
| Model | 0.647 | 34 | 0.01904 | 614.0 | <0.0001 |
| Error | 0.002 | 70 | 0.00003 | ||
| Corrected Total | 0.6495 | 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



