
Submitted:
29 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
Coxsackievirus B3 (CVB3) is one of the most common pathogens that cause myocarditis in humans. However, a sufficient therapeutic drug was not developed yet. Caboxamycin, a benzoxazole antibiotic, isolated from the culture broth of the marine strain Streptomyces sp., SC0774, displayed an antiviral effect in the CVB3-infected HeLa cells and CVB3-induced myocarditis mouse model. Caboxamycin significantly inhibited CVB3 VP1 production and cleavage of translation factor eIF4G1 from CVB3-infected HeLa cells at a 100 μg/ml concentration. Moreover, virus RNA amplification was dramatically reduced by caboxamycin treatment in HeLa cells. The caboxamycin cardiac protective effect was observed on the CVB3-mediated myocarditis mice model. The caboxamycin-treated mice survival was significantly increased compared to untreated mice (treated vs untreated, 100 vs 45 %). Heart histologic findings showed that myocardium damage and inflammation were significantly decreased by caboxamycin treatment. Caboxamycin directly inhibited caspase3 activity and cardiac myocytes apoptosis. These results suggested that caboxamycin strongly suppressed cardiac myocyte apoptosis and effectively inhibited the proliferation of CVB3 from in vitro and in vivo conditions. Furthermore, this study proposed that caboxamycin could potentially be applied in developing a new antiviral drug for CVB3-induced myocarditis.
Keywords:
coxsackievirus B3
; myocarditis
; apoptosis
; caboxamycin
; Streptomyces sp. SC0774
1. Introduction
Enteroviruses of the Picornaviridae family, particularly Coxsackievirus B3 (CVB3), have been considered one of the most common etiological agents of myocarditis. Although most of enterovirus infections are subclinical, acute myocardial inflammation can induce severe arrhythmias and sudden cardiac death, as well as lead to the development of chronic myocarditis and dilated cardiomyopathy [1,2,3]. Previous studies reported the antiviral effects of several candidate antiviral reagents in the experimental CVB3 acute murine myocarditis model in the Balb/C strain [4]. For instance, IL-1 receptor antagonist (IL-1Ra) trap, Coxsackievirus and Adenovirus receptor (CAR) trap gene therapy, and 3C protease inhibitor (3CPI) were tested in Balb/C strain as a model of acute severe myocarditis [5,6,7,8]. The implanted micro-osmotic pumps deliver water-insoluble 3CPI in 100% dimethyl sulfoxide in a subcutaneous Balb/C mouse. The result demonstrated that 3CPI effectively suppressed CVB3-induced myocardial damage and mortality [7]. However, there are many limitations to solve developing therapeutic drugs for clinical application.
Viruses cause a variety of human pathogeneses, including cancer, and play an important role in human disease. Although improvements in vaccination and drug development are made, numerous viruses lack preventive vaccines and effective antiviral medicines, which are frequently affected by viral escape mutants. As a result, discovering new antiviral drugs is crucial. Herbal medicines and natural products are a rich resource for discovering new antiviral drugs [9]. Marine-derived actinomycetes have been considered an essential source of structurally distinct secondary metabolites with various biological activities including antiviral, anti-tumor, antimicrobial and anticancer [10,11]. HPLC-UV guided fractionation of the culture broth of Streptomyces sp. strain SC0774, isolated from marine sediment of the Antarctic, has led to the discovery of caboxamycin (1) (Figure 1). This study describes the isolation of caboxamycin (1) and demonstrates its antiviral activity against CVB3.
2. Results and Discussion
Identification of Compound 1. Compound 1 was isolated as a white powder with
a pseudomolecular ion peak at the m/z = 256.23 [M+H]+ in LRMS spectroscopic data. The 1H NMR spectrum of compound 1 displayed seven aromatic protons at δH 8.09 (d, J = 8.4 Hz, 1H, H-7′), 8.02 (d, J = 8.4 Hz, 1H, H-5′), 7.81 (d, J = 7.9 Hz, 1H, H-6), 7.45 (t, J = 9.4 Hz, 1H, H-6′), 7.44 (t, J = 9.4 Hz, 1H, H-4), 7.13 (d, J = 7.9 Hz, 1H, H-3), and 7.02 (t, J = 9.4 Hz, 1H, H-5). Moreover, the 13C NMR spectrum of 1 displayed seven quaternary carbons at δC 164.1 (C-8′), 158.9 (C-2′), 158.8 (C-2), 149.6 (C-1′), 140.1 (C-3′), 120.5 (C-4′) and 110.0 (C-1) and seven methines at δC 134.5 (C-6′), 128.1 (C-7′), 127.7 (C-5′), 124.9 (C-4), 119.8 (C-5), 119.7 (C-3), 115.5 (C-6). Finally, compound 1 was identified as caboxamycin (Figure 1A) based on a comparison of its NMR data to the literature [14]. Caboxamycin (1) was first isolated from the extracts of the marine Streptomyces sp. NTK 937. Compound 1 possessed the benzoxazole scaffold as the core center as shown in Figure 1 and was reported to exhibited antibiotic activity against Gram-positive bacteria Bacillus subtilis and Staphylococcus lentus, as well as the yeast Candida glabrata [12,13]. In addition, 1 also possessed moderate cytotoxic effect against gastric adenocarcinoma (AGS), hepatocellular carcinoma (Hep G2) and breast carcinoma cells (MCF7).
Caboxamycin Strongly Inhibit Coxsackievirus B3 Replication. The antiviral effect of caboxamycin (1) was observed in HeLa cells with CVB3 infection. CVB3 was infected with 100 to 0.1 ug/ml serial diluted 1 for 16 hours, and then the cell survival was measured by CCK-8 kit. HeLa cell survival was strongly preserved by 1 treatment compared to untreated (Figure 2A). In addition, 1 showed very weak cell toxicity in HeLa cells. But 100 μg/ml of 1 showed about 20% cell death (Figure 2B). High dose of chemical has some cytotoxicity, so it is not a serious problem to apply the development of a new antiviral component based on caboxamycin.
The CVB3 replication was observed by western blot analysis and quantitative RT-PCR of viral capsid protein (VP1). During replication, CVB3 produces protease 2A protein to cleave their polyprotein to generate viral particles. This protease 2A breaks down the host translation initiation factor eIF4G1. Caboxamycin (1) treatment dramatically suppressed eIFG1 cleavage and VP1 viral capsid protein production (Figure 3A and B). Virus replication is initiated from RNA genome amplification. Especially, negative-strand RNA amplification is the major step of CVB3 replication. 100, 10 ug/ml of 1 treatment inhibited positive and negative-strand RNA genome amplification (Figure 3C and D). These results showed that 1 has a strong antiviral effect on both genome amplification and viral protein production.
Caboxamycin treatment inhibited apoptosis and improved the survival cell signaling pathway. Virus infection activates the apoptosis pathway and kills the cells. This pathway was regulated by activating Bcl-2 family proteins such as pro-apoptotic BAD (Bcl-2 associated Agonist of cell death), and BAX (Bcl-2 Antagonist X) activation. We tested the apoptosis inhibition by treatment of 1 in CVB3-infected HeLa cells. However, pro-apoptotic BAD and BAX transcription levels were dramatically increased by CVB3 infection but were not affected by 1 treatment. Moreover, a high dose of 1 enhanced apoptosis gene transcription (Figure 4A and B). This apoptosis activation may be beneficial for antiviral effects in CVB3-infected cells. In contrast, the PARP (Poly ADP-ribose polymerase) cleavage, Bcl-2, and p38 phosphorylation were significantly decreased by 100 ug/ml treatment of 1 (Figure 4C and D). These results implied that apoptotic cell signaling was dramatically inhibited and virus-infected cell survival improved. The inhibition of CVB3 replication can reduce apoptotic activity, but BAX and BAD mRNA expression level was not decreased from the 1 treatment.
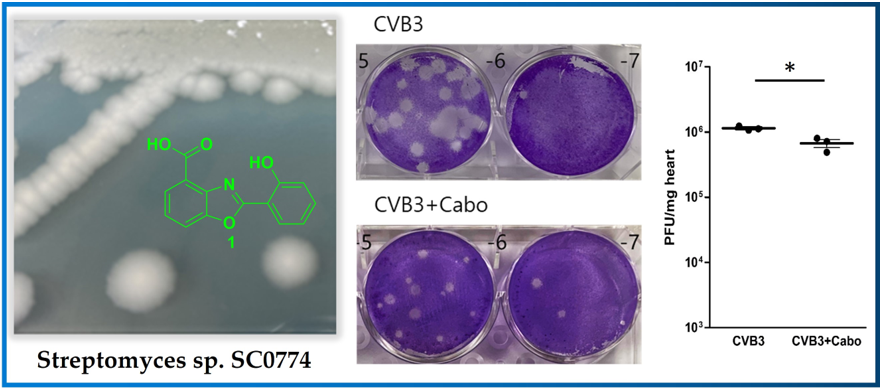
Caboxamycin reduced cardiac inflammation and virus replication in the CVB3-induced myocarditis mice model. Caboxamycin was administered in the CVB3-induced myocarditis mice model. The animal experiment was performed following the experiment design (Figure 5A). The six-weeks-old male mice (about 20 g in weight) were infected intraperitoneally with 2 x 105 PFU of CVB3 (CVB3) or with caboxamycin (CVB3+Cabo). The mice survival rate was preserved in the caboxamycin-treated group compared to the untreated group (CVB3 Vs CVB3+Cabo; 45 Vs 100%, p<0.08). Heart virus titer was measured by PFU assay. The overall result showed that heart virus proliferation was significantly decreased in CVB3+cabo group compared to CVB3 group (CVB3 Vs CVB3+Cabo: 1143000±87369 Vs 670233±161678, p<0.05) (Figure 5B, and C).
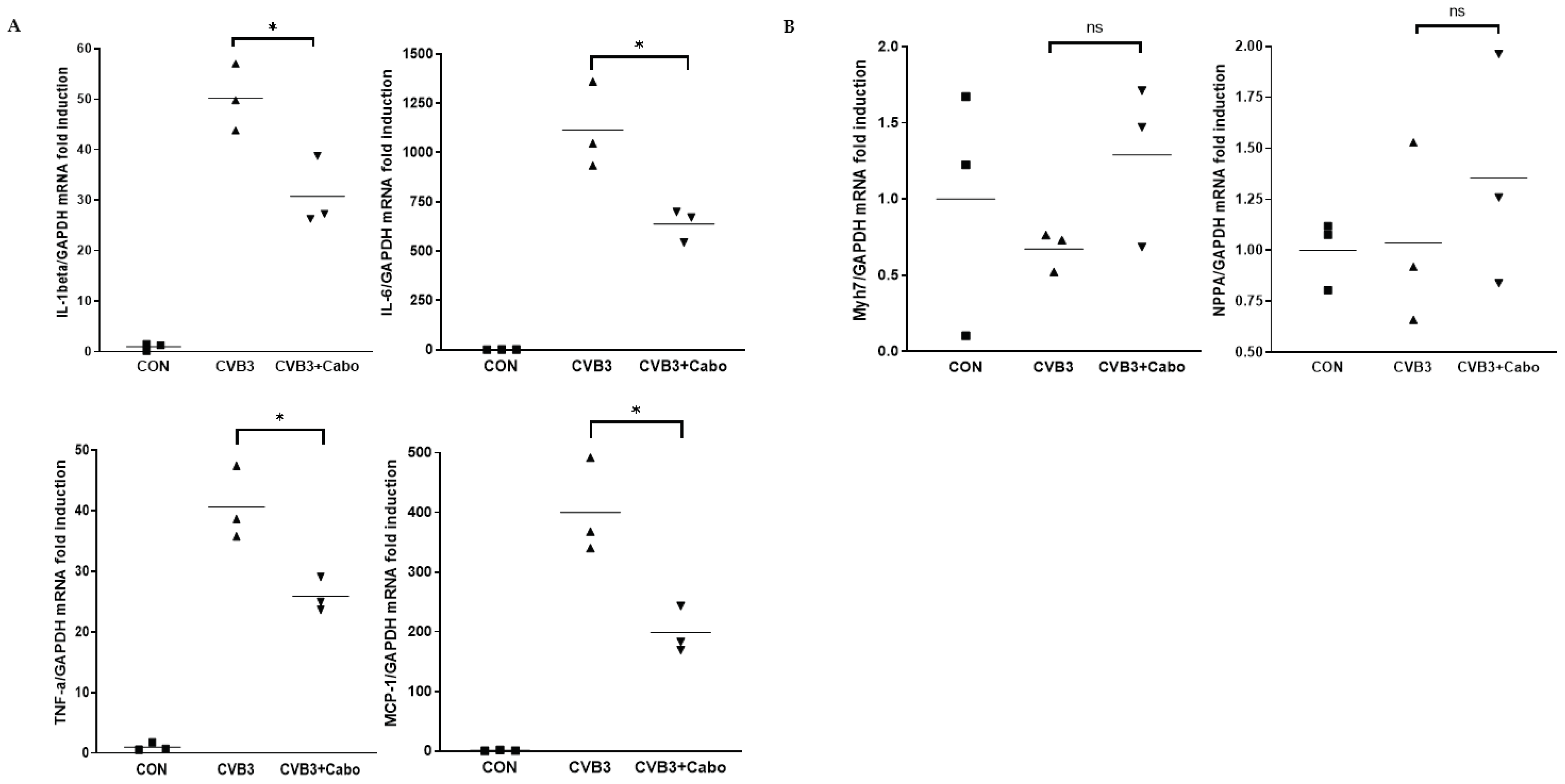
Heart inflammation was observed by hematoxylin and eosin (H&E) stains of day 3 and 12 post-infection mice hearts. NIH-ImageJ software quantified the focal inflammation area in two or three fields of each heart. The inflammation percent area was dramatically decreased by caboxamycin treatment compared to CVB3 group (Figure 6A, and B). CVB3 infection activate innate immunity and enhance inflammatory cell infiltrate on the virus infected damaged myocyte area [14,15]. Heart inflammatory cytokine mRNA expression was observed by quantitative RT-PCR using day 3 pi mice heart with caboxamycin treatment (CVB3+Cabo). IL-1beta, IL-6, TNF-alpha, and MCP-1 were significantly decreased by caboxamycin treated mice heart compared to CVB3-infected mice heart (Figure 7A). In addition, myocardium damage marker Myh7 and NPPA were little changed by caboxamycin treatment, respectively. However, it was not significantly different between two groups (Figure 7B). These results demonstrated that the inhibition of virus replication by caboxamycin treatment was effective to reduce inflammatory cell infiltration and cytokine production in CVB3-infected mice heart. However, the myocardium damage still not occurred at the early stage of CVB3 infection.
Viral myocarditis is an inflammatory process that causes damage to cardiac muscle cells [16]. The inhibitory effect of caboxamycin in CVB3 infection was tested. The result described that treatment with caboxamycin significantly suppressed CVB3 infection and cell mortality in HeLa cells. In addition, caboxamycin reduced apoptosis and cell death to HeLa cells by inhibiting virus replication and capsid protein (VP1) production. The cleavage of eIF4G1, a transcription initiation factor cleaved by viral protease 2A [17], was significantly decreased by caboxamycin treatment. CVB3 is a positive single-strand RNA virus, and producing a negative-strand RNA is essential for viral genome amplification [18]. caboxamycin treatment strongly inhibited negative strand RNA amplification of CVB3. Enterovirus replication occurs by activating ERK, a cell signaling molecule [19]. MAPK (mitogen-activated protein kinases) p38 activity was significantly suppressed following the drug concentration, and the cells remained viable. However, high concentration of caboxamycin has a cytotoxicity, therefore we should carefully define this reason by further experiment.
3. Materials and Methods
General Experimental Procedures. Low-resolution LC/MS measurements were performed using the Agilent Technologies 1260 quadrupole mass-spectrometer (Agilent Technologies, Santa Clara, CA, USA) and Waters Micromass-ZQ 2000 MS system (Waters Corp, Milford, MA, USA) using a reversed-phase column (Phenomenex Luna C-18 (2), 50 mm × 4.6 mm, 5 µm, 100 Å) at a flow rate of 1.0 mL/min at the National Research Facilities and Equipment Center (NanoBioEnergy Materials Center) at Ewha Womans University [20]. 1H and 2D NMR spectra were recorded at 400 MHz in CDCl3 using a solvent signal as internal standard on Varian Inova spectrometers (Bruker, Billerica, MA, USA). 13C NMR spectrum was acquired at 100 MHz on the Varian Inova spectrometer. Open-column chromatography was performed on C-18 resin (40-63 μm, ZEO prep 90) with a gradient solvent of water (H2O) and methanol (MeOH). The fractions obtained from open column chromatography were subsequently purified by reversed-phase high-performance liquid chromatography (HPLC) using a Phenomenex Luna C-18 (2), 100 Å, 250 nm × 10 mm, 5 μm column with a mixture of acetonitrile (CH3CN) and H2O at a flow rate of 2.0 mL min−1.
Collection and Phylogenetic Analysis of Strain SC0774. The marine-derived actinomycete strain SC0774 was isolated from marine sediment sample collected in Antarctic. The strain SC0774 was identified as Streptomyces sp. with 100% similarity to that of Streptomyces griseolus, based on the NCBI blast analysis of the partial 16S rRNA. The gene sequence data are available from Genebank (deposit MW132412.1).
Fermentation, Extraction, and Isolation. The strain SC0774 was cultured in 80 L of 2.5 L Ultra Yield Flasks, with each flask containing 1 L of SYP SW medium (10 g/L of Soluble starch, 2 g/L of yeast extract, 4 g/L of peptone, 139 g/L of sea salt in 1 L of distilled water) at 27°C with shaking at 120 rpm for 7 days. After a culture period of 7 days, the culture medium was extracted with ethyl acetate (EtOAc), yielding a total of 80 L of extract, which was concentrated in a rotary vacuum evaporator to yield 4.0 g of crude extract. The crude extract was fractionated into 8 fractions using reverse-phase C18 flash chromatography with a stepwise gradient elution of 80% of H2O in MeOH to 100% MeOH. The sixth fraction, 80% MeOH in H2O, was purified by reversed-phase HPLC (Phenomenex Luna C-18 (2), 250 × 100 mm, 2.0 mL/min, 5 μm, 100 Å, UV = 210 nm) using isocratic condition with 45% aqueous CH3CN, to yield 4.9 mg of caboxamycin (1).
Caboxamycin (1): 1H (400 MHz, CDCl3); δH 8.09 (d, J = 8.4 Hz, 1H), 8.02 (d, J = 8.4 Hz, 1H), 7.81 (d, J = 7.9 Hz, 1H), 7.45 (t, J = 9.4 Hz, 1H), 7.44 (t, J = 9.4 Hz, 1H), 7.13 (d, J = 7.9 Hz, 1H), 7.02 (t, J = 9.4 Hz, 1H), 13C NMR (100 MHz, CDCl3); δC 164.1, 158.9, 158.8, 149.6, 140.1, 134.5, 128.1, 127.7, 124.9, 120.5, 119.8, 119.7, 115.5, 110.0, LRMS m/z = 256.23 [M+H]+
Viruses and Cells. Coxsackievirus B3 (CVB3) was derived from the infectious cDNA copy of the cardiotropic CVB3-H3 was amplified in HeLa cells [5]. The HeLa cells, and mice tissue virus titer were determined by plaque forming unit (PFU) assay [7]. HeLa cells were cultured in Dulbecco’s Modified Eagle’s medium (DMEM, Welgene, Inc. Gyeongsan-si, Korea) supplemented with 5% fetal bovine serum (FBS) and 1% penicillin-streptomycin solution (Welgene, Inc.) solution at 37°C.
Screening of optimization antiviral compounds. The antiviral activity was observed following the previous reports. Briefly, HeLa cells cultured on a 96-well plate were infected with 20 μL of CVB3-H3 (106 PFU/mL) and treated with individual compounds, which were serially diluted from 100 μg/mL to 0.1 μg/mL. At 16 hours of post-infection, cell survival was measured with the addition of 8 μL of Cell Counting Kit 8 (CCK-8; Dojindo Molecular Technologies, Inc. Rockville, MD) reagent [5].
Myocarditis mouse model and drug administration. The protocols used in this study conformed to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85–23, revised 1996). Six-week-old male Balb/C mice were infected on day 0 by intraperitoneal injection with 2x105 plaque-forming units (PFU) of CVB3. Mice were euthanized via cervical dislocation and sera and various organs (heart, liver, spleen, and pancreas) were collected on days 7, and 12. The virus titer was measured by PFU assay from collected heart and pancreas. Antiviral effects of caboxamycin (100 μg/ml in 100μl saline or saline alone) were administrated intraperitoneally from day 0 post-infection for five consecutive days (CVB3+caboxamycin group (CVB3+Cabo), n = 10; CVB3 group (CVB3); n=10). The heart, livers, and pancreas of mice were taken out and evaluated on days 3, 7, and 12 post infection (p.i). Before analysis, mice were injected with Evans blue dye 14 hours before being examined. The heart inflammation and myocardium damage were observed by histologic analysis. All procedures were reviewed and approved by the Institutional Animal Care and Use Committee of Samsung Biomedical Research Institute (SBRI, #20191227002). SBRI is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC International) and abides by the Institute of Laboratory Animal Resources (ILAR) guide.
Total RNA extraction and quantitative real-time PCR. RNA was isolated from virus-infected mice hearts. Total RNA was extracted using the TRIzol reagent (ThermoFisher Scientific, Cambridge, MA, USA) to the manufacturer’s protocol. Complementary DNA (cDNA) was synthesized using 1ug RNA through a reverse transcription reaction using an oligo-dT primer for RNA quantification. Real-time PCR quantitative RNA or DNA analyses were performed in an ABI Sequence Detection System using the SYBR green fluorescence quantification system (Applied Biosystems, Waltham, MA, USA). The standard PCR conditions were 95°C for 10 minutes, then 40 cycles at 95°C (30 seconds), and 60°C (60 seconds), followed by a standard denaturation curve. All samples were tested in duplicate. The primer sequences shown in the were shown in Table S1.
Western blot analysis. Protein was extracted from frozen heart samples or cultured HeLa cells using LIPA buffer [50 mM Tris (pH 8.0), 0.1% SDS, 1% NP40, 150 mM NaCl, 0.5 % Sodium-deoxycholate] or PBS lysis buffer [1x PBS buffer, 1% Triton-x100], respectively. Approximately 20 μg of protein was separated on 10% SDS-PAGE, transferred to nitrocellulose membranes, and immunoblotted in 5% skim milk using standard methods. The membranes were probed with the primary antibodies anti-eIF4G1, enterovirus-VP1 (ThermoFisher Scientific), phosphor-p38, BCL-2, and GAPDH (Cell Signaling, Danvers, MA, USA). Detection was performed using an ECL solution (Intron Biotech, Inc. Seongnam-si, Korea), and band intensities were quantified by NIH-ImageJ software [21].
Histopathology and Immunohistochemistry. The hearts, pancreas, and liver were fixed in 10 % formalin, embedded in paraffin, and staining with hematoxylin and eosin (H&E) was performed using 10 μm paraffin-embedded sections as described previously [21]. Inflammatory cell infiltration was observed under a light microscope. Stained tissue images were taken and processed using a light microscope (Olympus Co. San Jose, CA, USA).
Statistical Analysis. All data were analyzed by Prism9 software (GraphPad Software, San Diego, CA, USA) and were presented as the means ± standard deviation (SD). For analyzing the statistical significance between the two groups, a student's t-test was used. For analyzing statistical significance between multiple groups, a one-way ANOVA was used.
4. Conclusions
Caboxamycin (1), a benzoxazole class of natural products, was isolated from the marine bacterium Streptomyces sp. SC0774. 1 inhibited apoptosis and inflammatory cell activation associated with replication and proliferative effects of CVB3, an effective agent of viral myocarditis. From this result, 1 was expected to be used to develop a new therapeutic drug for CVB3.
Supplementary Materials
The following are available online at www.mdpi.com/xxx/s1, Table S1. Real-time PCR primer sequences; Figure S1-S2. 1D NMR spectral of caboxamycin.
Author Contributions
H-.G.K. Data curation, Investigation, Writing – original draft.; P.F.H. Data curation, Investigation, Writing – original draft.; Y.-J.L. Investigation.; H.-E.J. Investigation.; B-.K.L. Conceptualization, Methodology, Writing – review & editing, Supervision.; S-.J.N Conceptualization, Methodology, Writing – review & editing, Supervision. All authors have agreed to the published version of the manuscript.
Funding
This study was supported by a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: HV22C0160, B.K.L); grant of the National Research Foundation (NRF) of Korea provided by the Korean Government (No. NRF-2022R1F1A1063986, B.K.L).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Mrs. Chae-Ok Gil for the help with the molecular biology experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Feldman, A.M.; McNamara, D. Myocarditis. N Engl J Med 2000, 343, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.F. Viral myocarditis. A review. Am J Pathol 1980, 101, 425–484. [Google Scholar]
- Huber, S.A.; Lodge, P.A. Coxsackievirus B-3 myocarditis. Identification of different pathogenic mechanisms in DBA/2 and Balb/c mice. Am J Pathol 1986, 122, 284–291. [Google Scholar] [PubMed]
- Herskowitz, A.; Beisel, K.W.; Wolfgram, L.J.; Rose, N.R. Coxsackievirus B3 murine myocarditis: wide pathologic spectrum in genetically defined inbred strains. Hum Pathol 1985, 16, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.K.; Choe, S.C.; Shin, J.O.; Ho, S.H.; Kim, J.M.; Yu, S.S.; Kim, S.; Jeon, E.S. Local expression of interleukin-1 receptor antagonist by plasmid DNA improves mortality and decreases myocardial inflammation in experimental coxsackieviral myocarditis. Circulation 2002, 105, 1278–1281. [Google Scholar] [CrossRef]
- Lim, B.K.; Choi, J.H.; Nam, J.H.; Gil, C.O.; Shin, J.O.; Yun, S.H.; Kim, D.K.; Jeon, E.S. Virus receptor trap neutralizes coxsackievirus in experimental murine viral myocarditis. Cardiovasc Res 2006, 71, 517–526. [Google Scholar] [CrossRef]
- Yun, S.H.; Lee, W.G.; Kim, Y.C.; Ju, E.S.; Lim, B.K.; Choi, J.O.; Kim, D.K.; Jeon, E.S. Antiviral activity of coxsackievirus B3 3C protease inhibitor in experimental murine myocarditis. J Infect Dis 2012, 205, 491–497. [Google Scholar] [CrossRef]
- Lim, B.K.; Yun, S.H.; Ju, E.S.; Kim, B.K.; Lee, Y.J.; Yoo, D.K.; Kim, Y.C.; Jeon, E.S. Soluble coxsackievirus B3 3C protease inhibitor prevents cardiomyopathy in an experimental chronic myocarditis murine model. Virus Res 2015, 199, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.T.; Hsu, W.C.; Lin, C.C. Antiviral natural products and herbal medicines. J Tradit Complement Med 2014, 4, 24–35. [Google Scholar] [CrossRef]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: marine actinomycete bacteria. Nat Chem Biol 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr Opin Microbiol 2006, 9, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, C.; Schneider, K.; Bruntner, C.; Irran, E.; Nicholson, G.; Bull, A.T.; Jones, A.L.; Brown, R.; Stach, J.E.; Goodfellow, M.; Beil, W.; Kramer, M.; Imhoff, J.F.; Sussmuth, R.D.; Fiedler, H.P. Caboxamycin, a new antibiotic of the benzoxazole family produced by the deep-sea strain Streptomyces sp. NTK 937. J Antibiot (Tokyo) 2009, 62, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Losada, A.A.; Cano-Prieto, C.; Garcia-Salcedo, R.; Brana, A.F.; Mendez, C.; Salas, J.A.; Olano, C. Caboxamycin biosynthesis pathway and identification of novel benzoxazoles produced by cross-talk in Streptomyces sp. NTK 937. Microb Biotechnol 2017, 10, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.T. Jr. Myocarditis. N Engl J Med 2009, 360, 1526–1538. [Google Scholar] [CrossRef]
- Fairweather, D.; Yusung, S.; Frisancho, S.; Barrett, M.; Gatewood, S.; Steele, R.; Rose, N.R. IL-12 receptor beta 1 and Toll-like receptor 4 increase IL-1 beta- and IL-18-associated myocarditis and coxsackievirus replication. J Immunol 2003, 170, 4731–4737. [Google Scholar] [CrossRef]
- Klingel, K.; Kandolf, R. The role of enterovirus replication in the development of acute and chronic heart muscle disease in different immunocompetent mouse strains. Scand J Infect Dis Suppl 1993, 88, 79–85. [Google Scholar]
- Castello, A.; Alvarez, E.; Carrasco, L. Differential cleavage of eIF4GI and eIF4GII in mammalian cells. Effects on translation. J Biol Chem 2006, 281, 33206–33216. [Google Scholar] [CrossRef]
- Lim, B.K.; Yun, S.H.; Gil, C.O.; Ju, E.S.; Choi, J.O.; Kim, D.K.; Jeon, E.S. Foreign gene transfer to cardiomyocyte using a replication-defective recombinant coxsackievirus B3 without cytotoxicity. Intervirology 2012, 55, 201–209. [Google Scholar] [CrossRef]
- Opavsky, M.A.; Martino, T.; Rabinovitch, M.; Penninger, J.; Richardson, C.; Petric, M.; Trinidad, C.; Butcher, L.; Chan, J.; Liu, P.P. Enhanced ERK-1/2 activation in mice susceptible to coxsackievirus-induced myocarditis. J Clin Invest 2002, 109, 1561–1569. [Google Scholar] [CrossRef]
- Han, J.Y.; Jeong, H.I.; Park, C.W.; Yoon, J.; Ko, J.; Nam, S.J.; Lim, B.K. Cholic acid attenuates ER stress-induced cell death in coxsackievirus-B3 infection. J Microbiol Biotechnol 2018, 28, 109–114. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, H.H.; Kim, J.H.; Park, J.H.; Jeon, E.S.; Lim, B.K. Protein Kinase B2 (PKB2/AKT2) Is Essential for Host Protection in CVB3-Induced Acute Viral Myocarditis. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of caboxamycin (1).
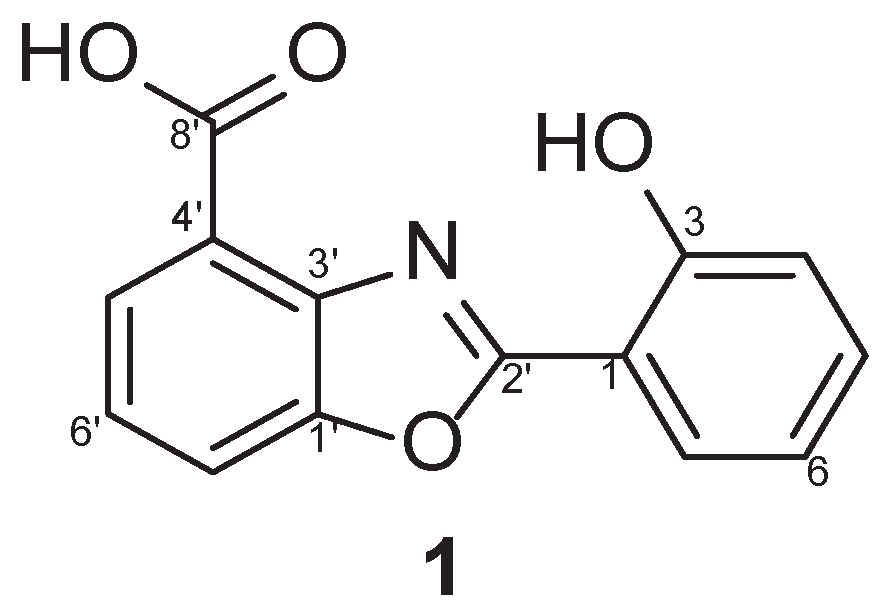
Figure 2.
(A) HeLa cell was treated with serially diluted caboxamycin (1) (100 to 0.1 ug/ml) with or without CVB3 infection for 18hr, then a microplate reader measured cell survival. Caboxamycin significantly improved virus-infected cell survival. (B) Caboxamycin cell toxicity was observed from HeLa cell survival. The purified drug showed weak cytotoxicity at the high concentration. All data are presented as the mean ± standard deviation (SD) from 3 independent experiments. ** P < 0.01 by two-tailed Student’s t-test.
Figure 2.
(A) HeLa cell was treated with serially diluted caboxamycin (1) (100 to 0.1 ug/ml) with or without CVB3 infection for 18hr, then a microplate reader measured cell survival. Caboxamycin significantly improved virus-infected cell survival. (B) Caboxamycin cell toxicity was observed from HeLa cell survival. The purified drug showed weak cytotoxicity at the high concentration. All data are presented as the mean ± standard deviation (SD) from 3 independent experiments. ** P < 0.01 by two-tailed Student’s t-test.
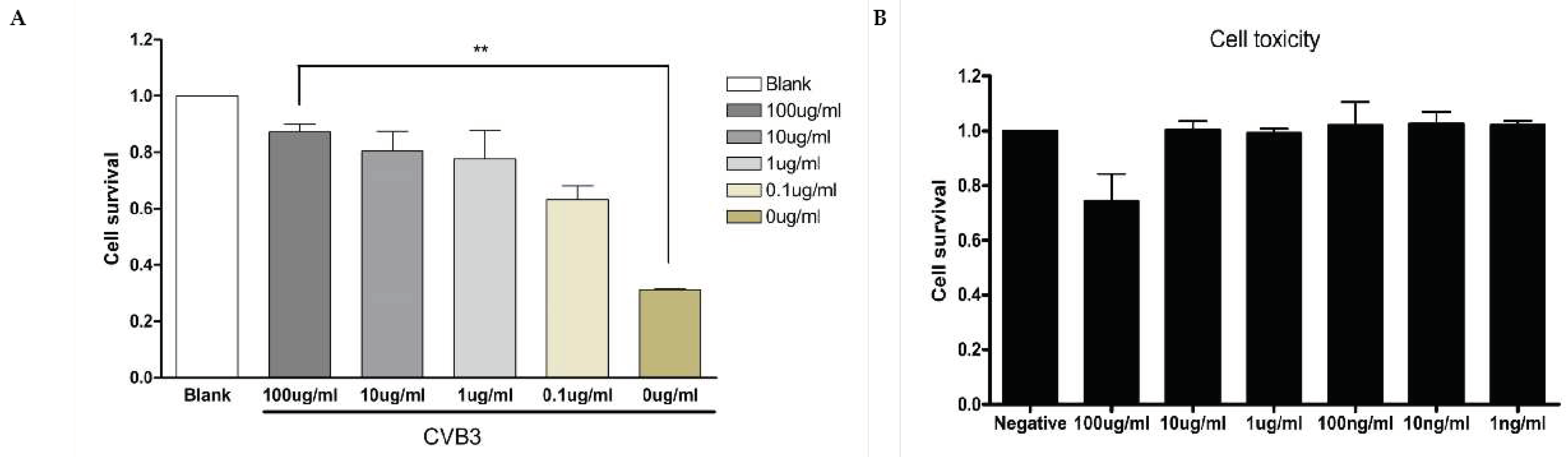
Figure 3.
(A) HeLa cells infected by CVB3 with serially diluted caboxamycin (1), then protein and RNA were extracted at 16hr pi. The extracted protein was applied for western blot analysis and probed by antibody eIF4G1, enterovirus-VP1, and GAPDH as a protein loading control. Cleaved eIF4G1 (arrow) and viral capsid protein VP1 decreased in 100 ug/ml of caboxamycin (1) treatment. (C) The extracted RNA was applied for quantitative RT-PCR, primer pair identified the negative and positive-strand RNA genome of CVB3. (B,D) Western blot and RT-PCR results were quantitated by NIH-ImageJ software. All data are presented as the mean ± SD. * p < 0.05 by two-tailed Student’s t-test.
Figure 3.
(A) HeLa cells infected by CVB3 with serially diluted caboxamycin (1), then protein and RNA were extracted at 16hr pi. The extracted protein was applied for western blot analysis and probed by antibody eIF4G1, enterovirus-VP1, and GAPDH as a protein loading control. Cleaved eIF4G1 (arrow) and viral capsid protein VP1 decreased in 100 ug/ml of caboxamycin (1) treatment. (C) The extracted RNA was applied for quantitative RT-PCR, primer pair identified the negative and positive-strand RNA genome of CVB3. (B,D) Western blot and RT-PCR results were quantitated by NIH-ImageJ software. All data are presented as the mean ± SD. * p < 0.05 by two-tailed Student’s t-test.
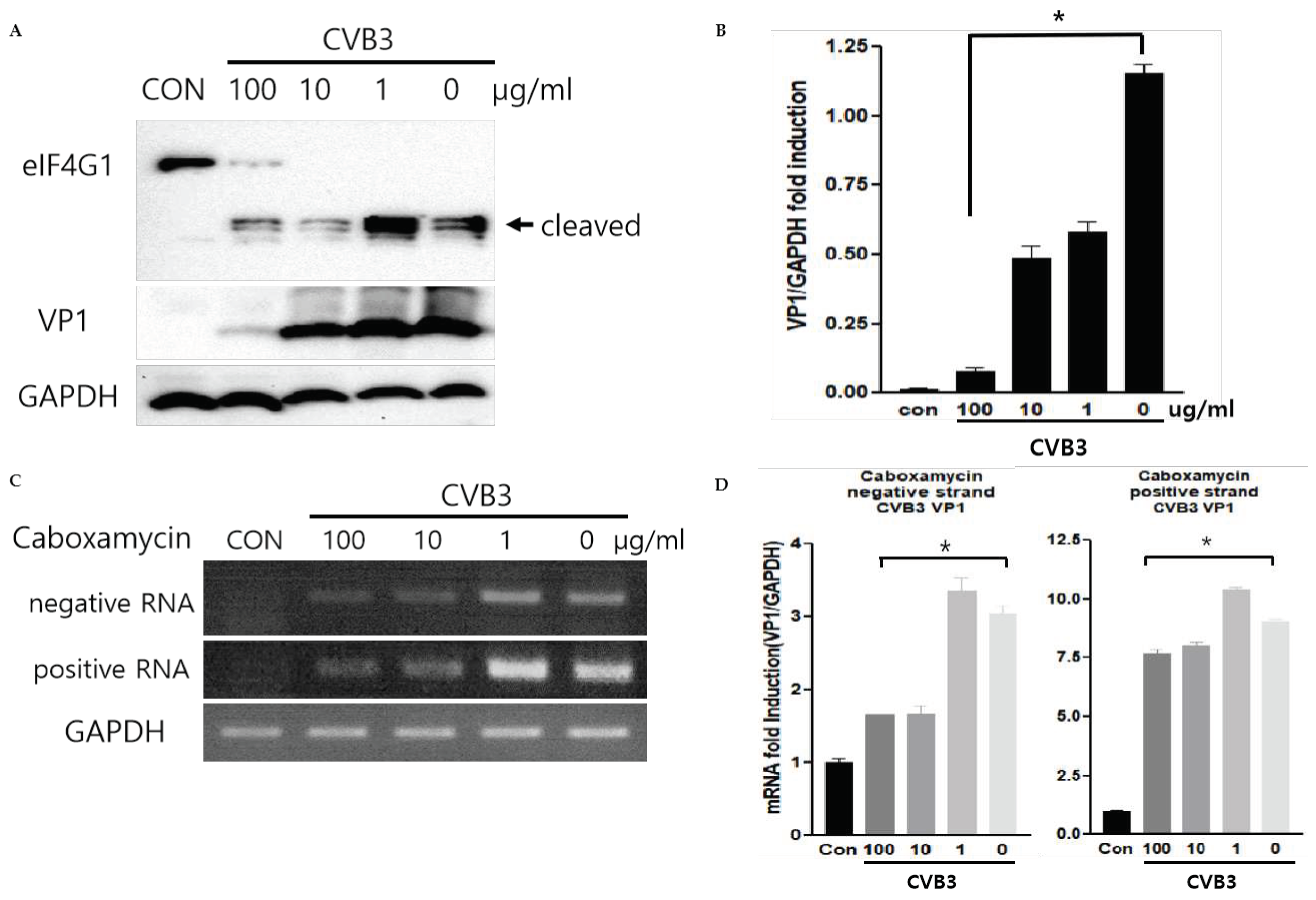
Figure 4.
(A,B) The RNA was extracted from HeLa cells after being treated with serially diluted caboxamycin (1) in CVB3 infection. Bad and Bax mRNA expression levels were quantified by quantitative RT-PCR (mean ± SD; n=3). (C,D) The protein levels of cleaved PARP, Bcl-2, phospho- p38, and GAPDH expression were observed by western blot analysis. The band intensity was quantitated by NIH-ImageJ software. All data are presented as the mean ± SD. * p < 0.05; ** P < 0.01 by two-tailed Student’s t-test.
Figure 4.
(A,B) The RNA was extracted from HeLa cells after being treated with serially diluted caboxamycin (1) in CVB3 infection. Bad and Bax mRNA expression levels were quantified by quantitative RT-PCR (mean ± SD; n=3). (C,D) The protein levels of cleaved PARP, Bcl-2, phospho- p38, and GAPDH expression were observed by western blot analysis. The band intensity was quantitated by NIH-ImageJ software. All data are presented as the mean ± SD. * p < 0.05; ** P < 0.01 by two-tailed Student’s t-test.
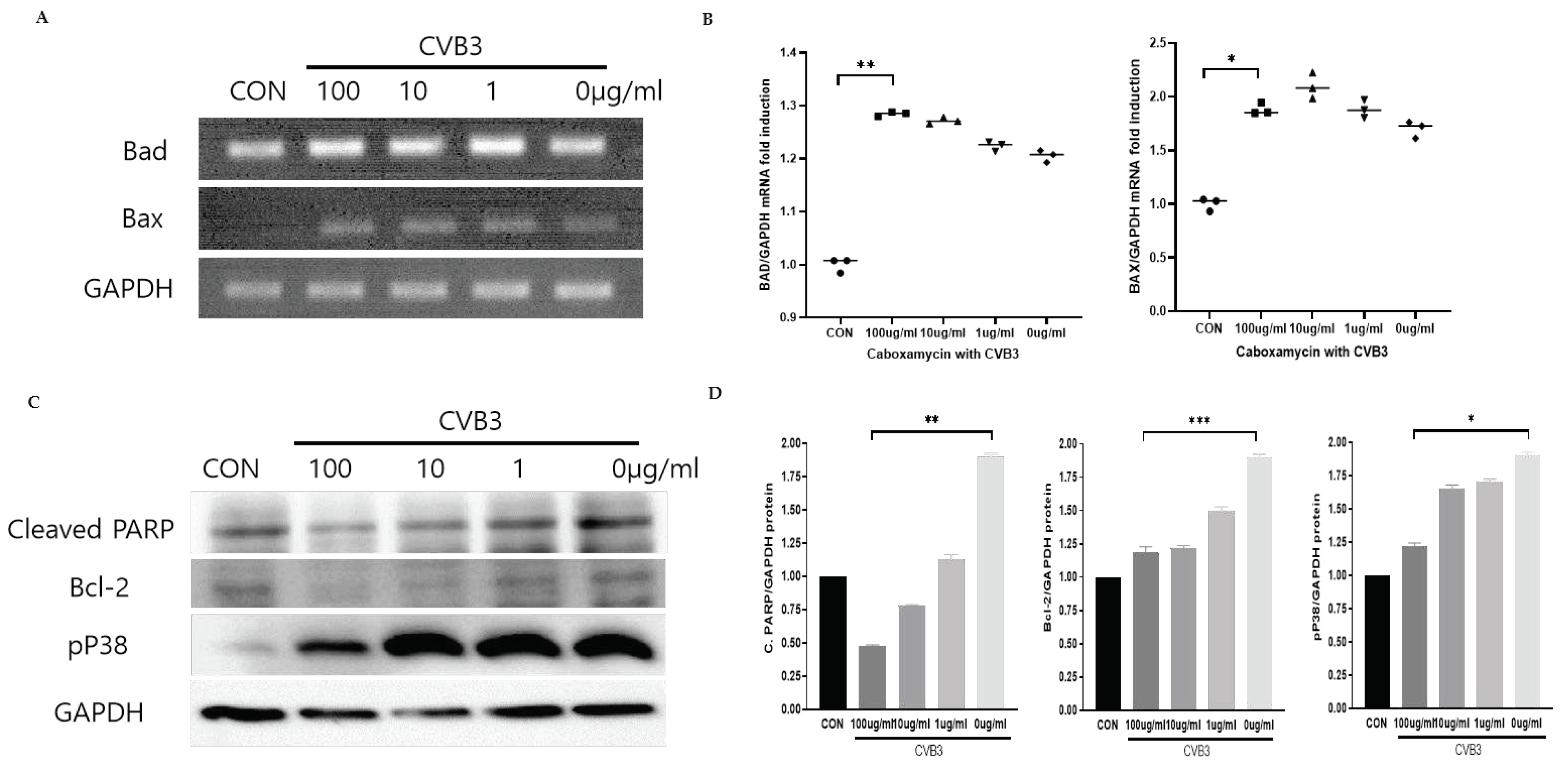
Figure 5.
(A) CVB3 was injected intraperitoneally to 6week old ages of male Balb/C mice (CVB3, n=10) or caboxamycin treatment (CVB3+Cabo, n=10). (B) Mice survival rate was recorded at day 14 pi. (C) The heart was lysed at day 5 pi. Tissue lysates were subjected to plaque forming unit (PFU) assay to measure tissue virus titers. All data are the mean ± SD. *P<0.05 by two-tailed Student’s t-test.
Figure 5.
(A) CVB3 was injected intraperitoneally to 6week old ages of male Balb/C mice (CVB3, n=10) or caboxamycin treatment (CVB3+Cabo, n=10). (B) Mice survival rate was recorded at day 14 pi. (C) The heart was lysed at day 5 pi. Tissue lysates were subjected to plaque forming unit (PFU) assay to measure tissue virus titers. All data are the mean ± SD. *P<0.05 by two-tailed Student’s t-test.
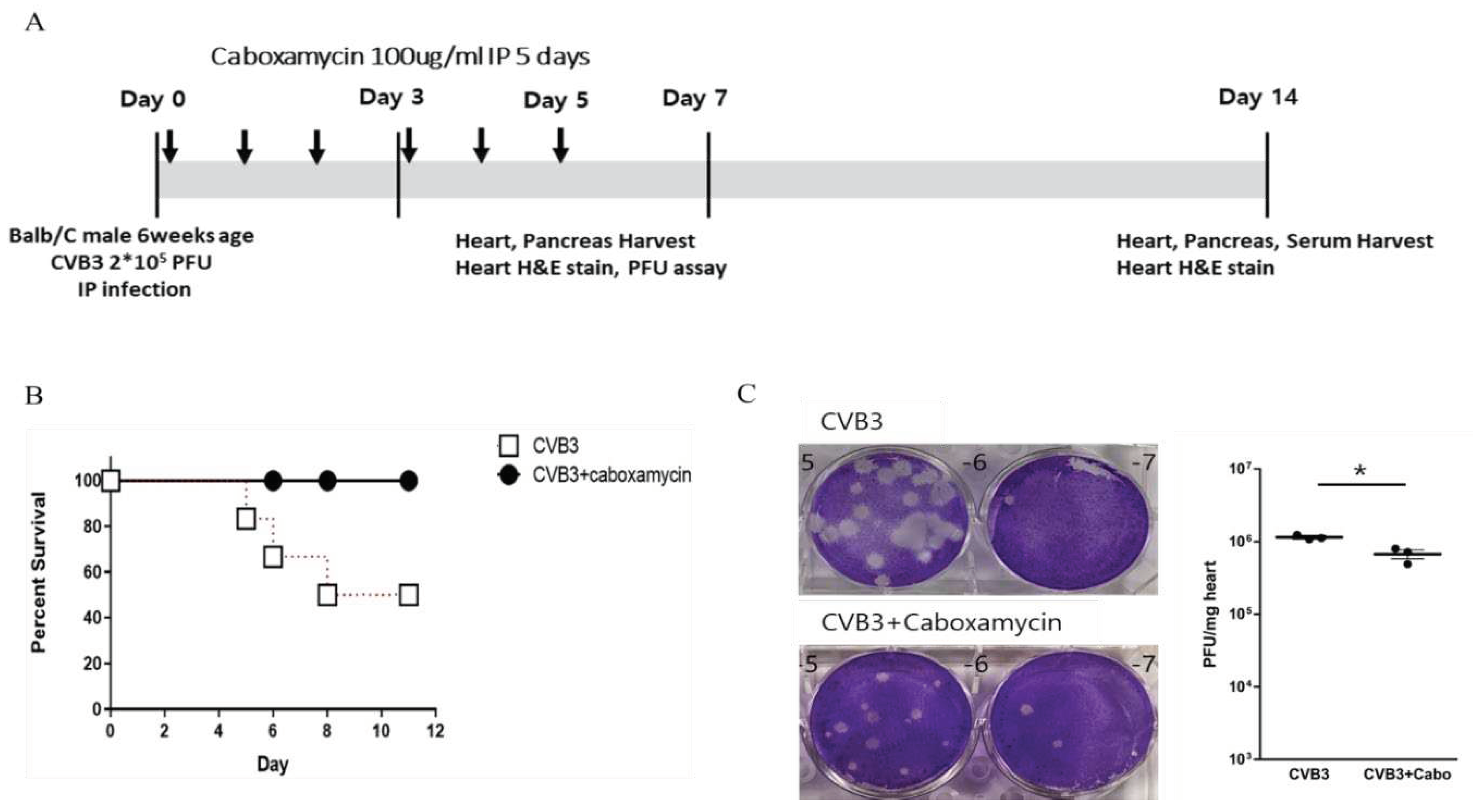
Figure 6.
(A) CVB3-infected mice heart inflammation was observed by the heart histological finding. Mice were sacrificed on day 3 and 12 pi, and then collected the serum and heart. Histological findings of the heart showed inflammatory cell infiltration using Hematoxylin & Eosin (H&E) staining. (B) Quantification of inflammation area (%) of the heart (CVB3 Vs CVB3+Cabo, n=3). All data are presented as the mean ± SD. ****P<0.0001 by two-tailed Student’s t-test.
Figure 6.
(A) CVB3-infected mice heart inflammation was observed by the heart histological finding. Mice were sacrificed on day 3 and 12 pi, and then collected the serum and heart. Histological findings of the heart showed inflammatory cell infiltration using Hematoxylin & Eosin (H&E) staining. (B) Quantification of inflammation area (%) of the heart (CVB3 Vs CVB3+Cabo, n=3). All data are presented as the mean ± SD. ****P<0.0001 by two-tailed Student’s t-test.
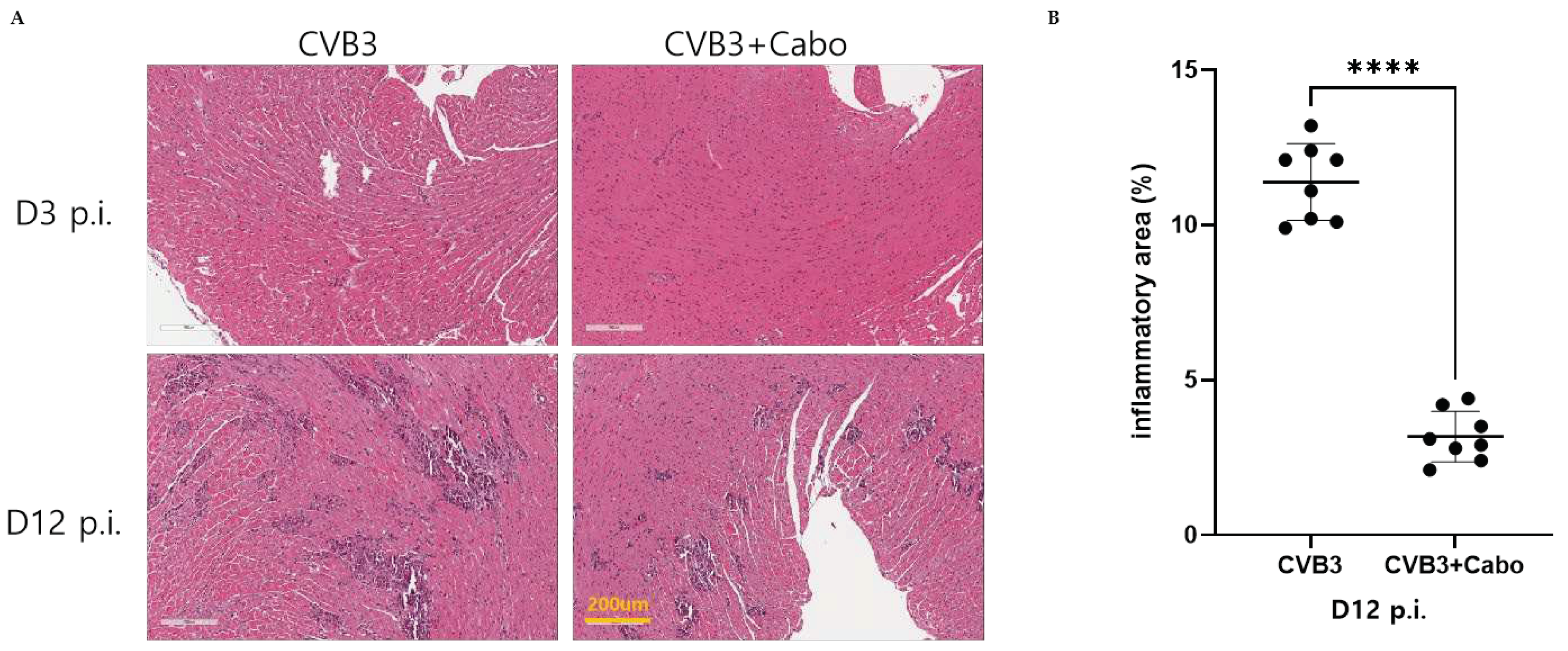
Figure 7.
(A and B) RNA was extracted from the mice heart of each group (n=3 each group) and applied for quantitative RT-PCR. Primer was identified for inflammation marker (IL-1beta, IL-6, TNF-alpha, and MCP-1) and heart damage marker (Myh7 and NPPA). The mRNA levels showed by individual dot graph. All data are the mean ± SD from independent experiments. NS>0.05, *P < 0.05 by two-tailed Student’s t-test.
Figure 7.
(A and B) RNA was extracted from the mice heart of each group (n=3 each group) and applied for quantitative RT-PCR. Primer was identified for inflammation marker (IL-1beta, IL-6, TNF-alpha, and MCP-1) and heart damage marker (Myh7 and NPPA). The mRNA levels showed by individual dot graph. All data are the mean ± SD from independent experiments. NS>0.05, *P < 0.05 by two-tailed Student’s t-test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



