
Submitted:
29 September 2023
Posted:
29 September 2023
You are already at the latest version
Abstract
This review delves into the intricate relationship between folate (Vitamin B9) intake, especially its synthetic form, folic acid, and its implications on health and disease. While folate plays a pivotal role in the one-carbon cycle, essential for DNA synthesis, repair, and methylation, concerns arise from its excessive intake. The literature underscores potential deleterious effects, such as an increased risk of carcinogenesis, disturbances in DNA methylation, and impacts on embryogenesis, pregnancy outcomes, neurodevelopment, and disease risk. Notably, these consequences stretch beyond the immediate effects, potentially influencing future generations through epigenetic reprogramming. We probe into the molecular mechanisms underlying these effects, including accumulation of unmetabolized folic acid, Vitamin B12 dependent mechanisms, altered one carbon metabolism, altered methylation patterns, and interactions with critical receptors and signaling pathways. Furthermore, we emphasize differences in the effects and mechanisms mediated by folic acid compared to natural folate. Given the widespread folic acid supplementation, it is imperative to further research its optimal intake levels, and the molecular pathways impacted by its excessive intake, ensuring the health and well-being of the global population.
Keywords:
Unmetabolized folic acid (UMFA)
; folic acid
; one-carbon cycle
; Methylation
; epigenetic
; carcinogenesis
; aging
; neurodevelopment
; embryogenesis
; folate
1. Introduction
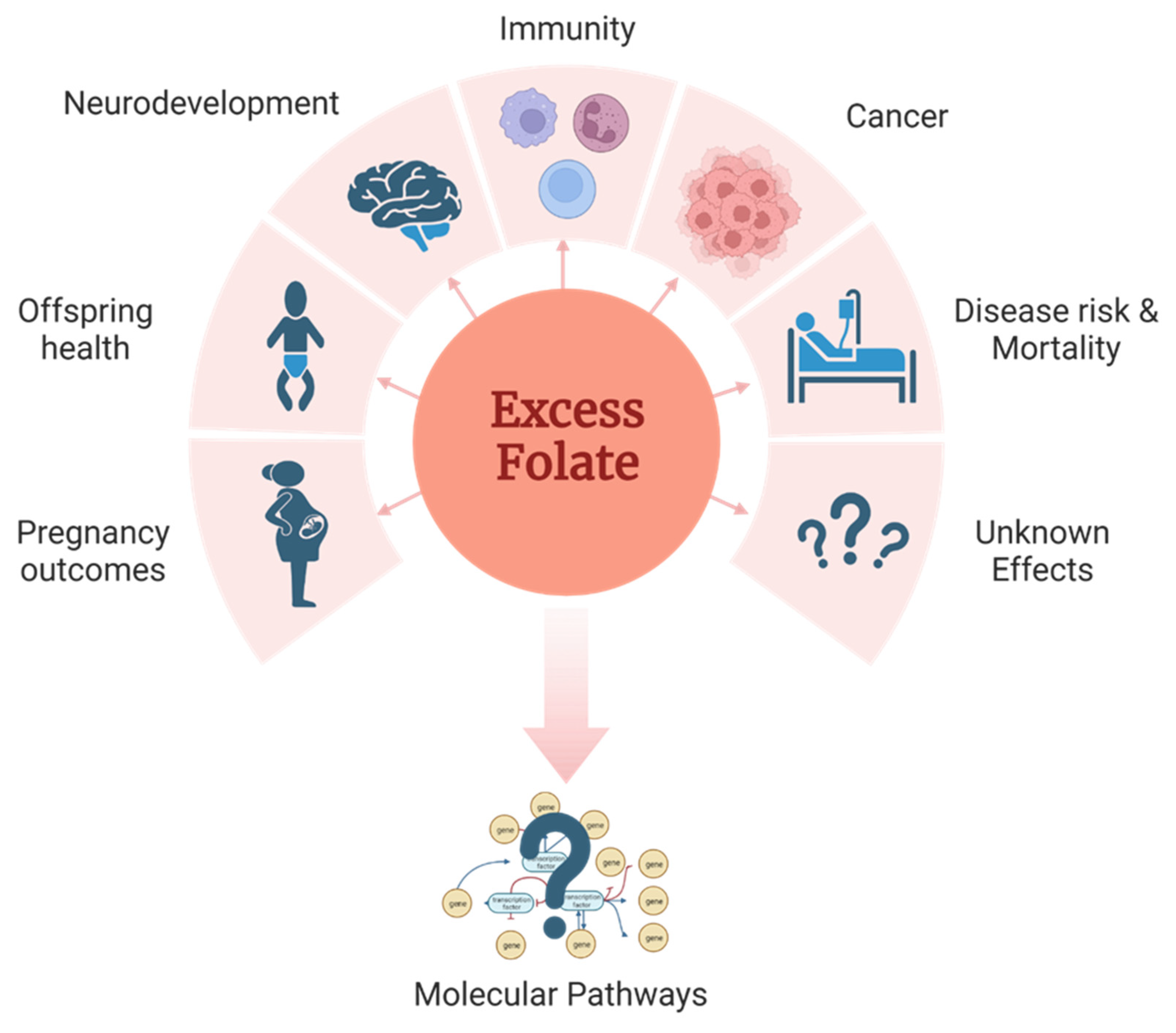
It is estimated that up to a third of the US population is chronically exposed to high levels of dietary folate from a combination of voluntary supplementation and food fortification with folic acid. Eukaryotic cells lack the capability to synthesize folate de novo, establishing folate as an essential nutrient for higher organisms and humans. While the benefits of adequate folate intake are well documented, there is mounting evidence linking its excess with deleterious effects on human health by impacting cancer, immunity, birth outcomes, cardiovascular disease, and overall mortality. The association between excess folate and these negative outcomes is known, but the causative mechanisms are not. Therefore, understanding and unraveling the mechanisms by which excess folate from natural sources, supplementation, enrichment, and fortification of the US food supply could negatively impact health and disease is of critical importance. Recent reports focused on the areas of nutrition, folate metabolism, cancer, and aging each conclude that there are substantial knowledge gaps relating to the effects of excess folate on human health [1,2,3]. Therefore, there is a need to understand how excess folate from either the fully oxidized folic acid used in supplementation or naturally occurring reduced folate forms impact health, and to determine the underlying biological mechanisms.
This review aims to provide a narrative summary of the existing literature describing the potential effects of excess folate and discussing the possible molecular mechanisms underlying these effects. A comprehensive perspective on the potential impacts of excessive folic acid intake on health and disease is currently lacking. Additionally, there is no consensus regarding the potentially harmful role of excess folic acid supplementation, and there is a need to reconcile conflicting reports to identify gaps and areas requiring further research. By addressing some of these gaps, our aim is to contribute to a better understanding of the complex relationship between folate intake and human health (Figure 1).
2. Defining excess folate
Determining what constitutes excessive folate intake is an elusive task since there is a lack of dose response data, and comprehensive documentation of adverse effects at high folate intake are not established [1]. The recommended daily allowance (RDA) of folic acid varies depending on age, sex, and life stage. Adult men and women are recommended 400mcg of dietary folate equivalent (DFE), increasing to 500 mcg and 600 mcg DFE for breastfeeding and pregnant women respectively. The Tolerable Upper Intake Level (UL) for folic acid was set at 1,000 mcg per day for adults. However, the determination of folic acid UL relied on limited and low-quality observational case-study data [4]. Generally, folate exposure at dosages exceeding the adult UL (1000 ug/day) are considered excessive, However, this criterion does not consider chronic exposure to elevated folate intake that approach the UL and the inherent variability and mutations in folate cycle enzymes that are prevalent in humans [5,6].
Folic acid (FA), the reduced synthetic form of folate, is used almost universally in fortification and supplementation as well as when conducting most laboratory research. Women of childbearing age taking prenatal vitamins as well as children and adults taking vitamins high in folic acid are likely to exceed the age adjusted UL of FA intake due to a combination of food fortification and supplementation [7,8]. FA supplementation is linked to elevated circulating unmetabolized folic acid (UMFA) in a dose dependent manner [9]. UMFA occurs due to saturation of mechanisms needed to convert folic acid into the active reduced forms in the intestine and liver [10]. Specifically, UMFA accumulation is associated with the saturation of the rate limiting one carbon enzyme dihydrofolate reductase (DHFR). Some evidence supports the use of UMFA as marker for excess folate in humans as UMFA levels generally increases in response to supplemental folic acid intake [11,12], however, newer sensitive methods are able to detect UMFA in majority of samples irrespective of folic acid intake [13,14]. A growing body of literature linking excess FA and UMFA to potential adverse health outcomes exists, However, evidence of causality between excess FA, UMFA and various adverse effects remains inconclusive[2].
Mandatory fortification in combination with supplementation with FA led to more than doubling of USA population serum folate concentration in the last thirty years [14,15,16]. In the USA, up to 35% of the population are taking folic acid containing supplements and roughly 5 percent of the population exceed the UL of folate intake. The average folate intake is estimated to be 813mcg/day for men and 724mcg/day for women [17]. The Canadian health measures survey identified that up to 40% of participants had High folate levels (defined as RBC folate >1360 nmol/ L) [18,19,20]. Unmetabolized folic acid was detected in over 95% of Americans participating in the National Health and Nutrition Examination Survey (NHANES), with significantly higher UMFA levels recorded among supplement users and older individuals[14,21]. Similarly, high levels of FA and UMFA was found in pregnant Canadian women and umbilical cord blood, with over 97% of pregnant Canadian women having detectable plasma UMFA [22,23], and up to 26% of pregnant women exceeded the UL of FA [24].
The distinction between natural folate and synthetic folic acid is an important consideration when evaluating the potential risks of excess intake. Natural folate, mainly in the form of 5-methyl-tetrahydrofolate (5-m-THF) and to a lesser extent, 5-formyl-tetrahydrofolate (5-f-THF), are biologically active forms that do not necessitate activation by dihydrofolate reductase (DHFR) as folic acid does. Natural folate intake is constrained by the limited amount present in food sources, as well as its lower bioavailability and stability. In contrast, large doses of synthetic folic acid can be easily consumed through food fortification and supplementation, making folic acid the primary contributor to elevated folate pools in the human population. It has been suggested that the deleterious effects of excess folate could be related to factors such as low vitamin B12 status, high levels of folic acid or UMFA, or elevated total folate pools. However, evidence linking these factors to causation of deleterious effects or specific molecular mechanisms remains limited [25]. Owing to incomplete data and a lack of scientific consensus regarding the effects of excess folate, as well as the absence of accepted physiological markers linked to such effects, determining what constitutes excess folate is challenging. For this review, we will discuss folate intake exceeding the WHO recommended level of 400 µg/day and approaching or exceeding the Upper Limit (UL) of 1 mg/day. We will also draw comparisons to studies reporting conservative levels of FA supplementation. Additionally, we will examine results from human, animal, and cell culture studies that reported elevated serum, RBC folate, and/or the presence of high levels of unmetabolized folic acid.
In rodent studies, excess folate could be defined as exceeding the accepted standard baseline supplementation of 2 mg/kg diet. However, the optimal folate requirements for animals may be lower, and the activity of rate limiting one-carbon metabolism enzymes, such as dihydrofolate reductase (DHFR), are typically much higher in rodents compared to humans [26,27,28,29]. Chow diets fed to laboratory rodents can contain a variable amount of FA ranging from 2-15mg/kg diet and averaging about 8 mg/kg [28]. The reason for supplementing rodent diets with excessive amount of FA is not clear, However, we can hypothesize that suppliers of research animals and their diets defaulted to higher levels of folate supplementation to ensure rapid growth and large litter sizes.
In cell culture studies, cells are subjected to elevated levels of folates that greatly exceed the physiological levels found in tissues in order to support growth and proliferation. In human plasma, the typical physiological levels of natural folate range from 150-450 nM, while standard cell culture media contains 2,200 nM (RPMI) or 9000 nM (DMEM) of the non-physiological synthetic folic acid [30,31]. This implies that most studies utilizing standard cell culture conditions could be confounded by the supraphysiological concentrations of folic acid [32,33].
Limited research has been conducted using physiologically relevant levels of natural folate in place of folic acid. Natural folates are inherently less stable and bioavailable than folic acid, which creates technical and logistical challenges for their integration into basic research. Additionally, the widespread use of the terms “folate” and “folic acid” interchangeably has led to the misconception that these forms are equivalent. In this review, “folate” refers to all folate species, synthetic and natural, found physiologically and in foods, while “folic acid” denotes the fully oxidized synthetic folate used in supplementation and food fortification.
3. Health impact of excess folate
3.1. Pregnancy and birth related outcomes
The primary goal of the folic acid food fortification mandates that were instituted in various countries around the world is to reduce the incidence of births with neural tube defects. Folate adequacy for preventing NTD’s is critical in the first 4 weeks of gestation. Folate supplementation in the following weeks, postnatally, and during breastfeeding could potentially provide benefits to maternal and fetal health with varying and inconsistent outcomes [34]. Supplementation at the WHO recommended levels of 0.4 mg/day in the periconceptual period protects against NTD’s with largely positive health outcomes reported. However, FA intake at higher doses, especially when approaching or surpassing the upper intake limit and administered for extended durations are at times associated with deleterious pregnancy and offspring outcomes. A comprehensive review on the effects of FA supplementation on gestation and long-term health is reviewed elsewhere by Silva et al. [34]. Common prenatal vitamins in the USA typically include an FA dose of 1mg/ day with some prescribed regiments using doses up to 5mg FA/day. On average, women taking prenatal vitamins in the are likely to surpass the UL of 1000ug folate per day due to a combination of supplementation and dietary intake [35], leading to elevated levels of total folate and UMFA in maternal and fetal samples [36,37].
The impact of FA supplementation and specifically high dose FA on non NTD’s pregnancy outcomes is not clear [34]. The link between FA supplementation and oral cleft incidence is inconsistent with studies reporting a protective effect [38,39,40,41], no clear association [42,43], or an increase in oral cleft births [44]. In one study, high FA (4mg/day) did not compromise fetal growth or provide additional protection against orofacial clefts compared to 0.4mg/day [42]. Another study that used the same dose of 4 mg FA /day in early pregnancy saw increased maternal UMFA, and serum folate, with no significant effect on RBC folate or evidence of altered one carbon metabolism [11].A Cochrane review found no conclusive evidence that FA supplementation improved focused pregnancy outcomes such as preterm birth, still birth, anemia, serum and RBC folate levels [45]. In a Greek cohort, 5mg FA/day was found to be protective against preterm birth and low birth weight [46]. FA supplementation provided benefits in reducing the incidence of congenital heart defects irrespective of the timing and dosage during pregnancy, with beneficial association attributed to high dose of FA (5 and 6 mg/day) [47,48].
High folate concentration was linked to an increased risk of gestational diabetes mellitus (GDM) in Chinese cohorts [49,50]. Specifically, Higher FA dose as well as the timing and duration of the supplementation was associated with an increased risk for the development of gestational diabetes mellitus [51,52,53,54], with a significant GDM association when high FA was combined with an imbalance in vitamin B12 levels [55]. In a Spanish cohort where pregnant women were supplemented with FA in doses exceeding 1 mg/day, an association was found with decreased birth length and an increase of the risk of low birth weight [56]. Excess FA intake in late pregnancy was linked to allergy and respiratory problems in children [34]. Increased incidence of position plagiocephaly was identified in a nested case-control study performed in the Netherlands in correlation to periconceptional high doses of FA [57]. Although both favorable and unfavorable pregnancy outcomes have been observed in relation to excess folate, more research is required to identify the appropriate levels of folate supplementation needed to achieve optimal reproductive health.
3.2. Disease risk in offspring
Folate intake plays a critical role in the availability of methyl donors necessary for epigenetic programming during conception, gestation, and early development. The paradigm that maternal exposure to various stressors, dietary factors, and nutrient excess or deficiency during the prenatal period and early development can modulate the risk for developing diseases later in life is well accepted [58]. As the scientific knowledge advances, much of the etiology behind these changes has been attributed to epigenetic modifications and imprinting and is influenced by one carbon metabolism [58,59,60]. Folate and its reduced intermediates play a crucial role in the one carbon cycle and in regulating the processes involved in methylation and epigenetic modifications. High maternal FA supplementation can negatively impact health outcomes of the progeny by altering embryonic development [61], and modulating the expression of key genes involved in growth [62]. Excess FA supplementation may potentially alter gene expression and lead to aberrant epigenetic programming in the progeny[63,64]. Understanding the link between folate status in the parents, offspring, and epigenetic changes can further our understanding of the origins of adult diseases [65,66,67].
Numerous human and animal studies identified associations between excess folate intake and disease risk in offspring, though the outcomes have been inconsistent. In the Indian subcontinent, where expecting mothers are often prescribed folic acid doses as high as 5mg/day, high folate concentrations during pregnancy were linked to insulin resistance in children [68]. Results from a UK cohort of pregnant women revealed that FA supplementation after the 12th week of pregnancy impacts DNA methylation in the offspring with significant changes in the methylation of regions that regulate the insulin like growth factor 2 in infants potentially altering susceptibility to chronic diseases [65]. A systematic review by Xie et al., aimed at exploring the association between maternal folate status and the risk of obesity and insulin resistance in offspring, was unable to reach a definitive conclusion due to inconsistencies in data collected form animal and human studies [69].
While the clinical significance of altered methylation in response to excess folate in humans is not clear, animal studies provided a more robust link between epigenetic changes and maternal folate status. In rodents, maternal and post weaning supplementation with FA significantly impacted global and gene specific methylation in the offspring [70]. Alteration of DNA methylation and imprinting patterns was partially responsible for deleterious transgenerational effects observed when either folate deficient or excess folate diets were fed to female mice [71]. Excessive FA during pregnancy induced obesity and altered the epigenetic profile of the offspring in rats [72,73], and led to metabolic dysfunction in late adulthood [74]. Similarly, other studies found that excess perinatal folic acid supplementation caused insulin resistance, dyslipidemia, disrupted glucose metabolism and hepatic fat metabolism in both mice[75] and rat offspring [76,77,78]. In mice, excess maternal folic acid supplementation during pregnancy led to altered offspring cardiac gene expression [79].
FA intake during the periconceptual period, as well as throughout the second and third trimesters, can induce epigenetic alterations, leading to changes in the expression of genes related to brain development and function [80,81,82]. Rodent studies have shown that exposure to maternal FA supplementation can modify offspring DNA methylation profiles and gene expression in the brain, resulting in behavioral and morphological changes [83,84,85,86]. Further research is necessary to determine the effects of maternal excess folate on health and disease risk in children.
The developmental origins of health and disease hypothesis suggest that fetal programming is influenced by maternal nutrition, among other factors. Moreover, growing evidence indicates that paternal environment and diet significantly impact progeny’s health and risk of developing disease through epigenetic changes in sperm [87]. High doses of paternal folic acid supplementation was associated with increased gestational duration in births conceived through assisted reproduction [88]. Male offspring of mice fed a high-folate diet had a decreased sperm count, altered methylation in key genes, increased postnatal mortality in offspring, and reduced litter size [89]. Similarly, high-dose folic acid resulted in sperm DNA hypomethylation in both men and mice [90]. While moderate folic acid supplementation was beneficial in reducing some assisted reproduction-induced DNA methylation variance, high-dose supplementation produced deleterious effects with sex-specific outcomes [91]. These findings highlight the critical need to examine the epigenetic effects of excess dietary folate more carefully, as the influence of both maternal and paternal folate supplementation can potentially increase children’s disease susceptibility later in life.
3.3. Neurodevelopment
Excess folic acid supplementation during pregnancy is linked to both beneficial and detrimental effects on neurodevelopmental outcomes in children. Although FA supplementation is beneficial during the pre-conceptual period and early weeks of gestation by preventing neural tube defects, continued supplementation may significantly impact children’s neurodevelopment. FA supplementation at the WHO-recommended level of 400µg/d beyond the first trimester was associated with benefits on neurocognitive development in children, with positive effects on cognitive development observed at ages 7 and 11 [92,93]. In another study where high dose FA supplementation was implemented, cord blood analysis revealed significant changes in DNA methylation, exhibiting sex-specific and loci-specific alterations in genes related to neurodevelopment [94]. Doses exceeding the upper limit (≥1000 µg/d) during the periconceptual phase were linked to impaired neurocognitive development in children [95,96]. Both low (<400 µg/d) and excess FA (>1000µg/d) maternal supplementation were associated with attentional dysfunction in children aged 4-5 years[95]. High-dose FA (>5mg/day) supplementation in pregnant mothers led to poor psychomotor development in their children compared to those receiving 0.4-1mg/day [97]. In a small Greek cohort (n=58), high-dose FA supplementation (5mg/day) in early pregnancy improved children’s vocabulary development, verbal comprehension, and communication skills, while doses exceeding 5mg/day showed no benefit [46].
The relationship between FA supplementation and autism spectrum disorders (ASD) is complex and marked by contradictory findings. Recent systematic reviews found inconclusive evidence associating FA supplementation and ASD [96]. However, some studies did find a significantly increased risk of ASD with elevated maternal plasma FA [96] and linked synthetic folic acid supplementation during pregnancy to an increased autism risk [98]. Some suggested that excess FA leading to unmetabolized folic acid (UMFA) can potentially heighten ASD risk [80]. Moderate prenatal FA supplementation of at least 400µg from diet and supplements was associated with a decreased ASD risk in children [84,85,92,93], while excess FA corresponded with lower cognitive development and increased ASD risk [94,96,99]. This suggests a U-shaped relationship where both inadequate and high FA exposure could increase ASD risk [100].
Animal models offer valuable insights into the effects of excess FA on neurodevelopmental outcomes. In these models, excess FA during the prenatal period is associated with adverse impacts on behavior, memory, embryonic growth, methyl metabolism, and neurodevelopment [100]. In rodent studies, excess maternal FA supplementation resulted in deleterious behavioral abnormalities in offspring, including anxiety-like behavior, hyperactivity, impaired reversal learning, and seizure susceptibility [96,99,101]. The collected evidence indicates that the influence of excess FA on the type of behavioral abnormalities in offspring is sex-specific [83,84,86,102]. Both deficient and excess FA during pregnancy impaired cortical neurodevelopment in mice offspring[103]. In another study, moderate FA supplementation led to behavioral changes and sex-dependent alterations in one-carbon metabolism [104]. High-dose FA supplementation in mice caused neural tube defects and disrupted embryonic development [84,105,106]. Neurodevelopmental toxicity in male mouse offspring’s brain was observed when a 2.5-fold FA requirement was provided one week before mating and continued through pregnancy and lactation [107]. In rats, high FA doses administered periconceptually decreased the offspring seizure threshold [106]. High FA intake (20mg/kg) impaired rat dam’s memory and Na+, K+ - ATPase activity in the pup’s hippocampus [108]. High exposure to maternal FA in utero resulted in DNA methylation-dependent changes in the expression of hypothalamic genes involved in food intake and weight regulation [73]. These findings underscore the intricate relationship between excess FA and neurodevelopmental outcomes, emphasizing the need for more comprehensive research to elucidate the underlying mechanisms and their implications for optimizing human health.
3.4. Immune function and allergies.
Folate plays a crucial role in maintaining a healthy immune system, influencing both innate and adaptive immune responses. Adequate folate status is essential for optimal immune function, while deficiency or excess of folate can lead to immune dysregulation. Folate is required for the production and maintenance of fast proliferating white blood cells and is necessary for the maturation of lymphocytes. Folate plays a role in regulating inflammation and the production of pro-inflammatory cytokines [109]. Folate deficiency has been associated with impaired immune cell function and increased levels of tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), which can contribute to impaired immunity, chronic inflammation, and the development of inflammatory diseases [110]. There is evidence that folate status is linked to the development and progression of inflammation and autoimmune diseases [111,112,113]. However, the impact of FA supplementation on inflammatory markers and associated diseases in humans remains inconclusive [114].
Excessive folate intake can negatively impact innate immunity. In postmenopausal women, high levels of folic acid intake through supplements and a folate-rich diet were found to reduce NK cell cytotoxicity [115]. Unmetabolized folic acid (UMFA) was inversely associated with NK cell cytotoxicity in the same study [115]. Prenatal and periconceptual FA supplementation and particularly excess FA was associated with an increased risk of childhood asthma in several studies [116,117,118,119,120,121], However, others could not find a clear association [122,123,124,125]. Interestingly, late pregnancy FA supplementation as well as the extended duration of the supplementation (over 6 months) was associated with the highest risk of childhood asthma [63,121]. Similarly, FA supplementation in the first trimester of pregnancy correlated with a higher probability and severity of bronchiolitis compared to non-supplemented mothers in the USA [126]. FA supplementation of pregnant women was associated with an increased risk of wheeze and lower respiratory tract infection in the offspring up to 18 months of age in a Norwegian cohort [127]. Conversely, others reported that high plasma folate in mid pregnancy was linked to decreased odds of wheezing at age 3 [128]. In a recent systematic review and meta-analysis, Chen et al. concluded that FA intake can be a risk factor for respiratory tract allergies in infants and children, however, a significant association was found only for doses not exceeding 400 ug/day, while the risk effect decreased with increasing FA dose. Moreover, the association between FA supplements and children’s risk of allergic diseases was significantly increased only in countries without FA fortification. This increase in risk was more pronounced in cases where mothers used folic acid supplements [129]. Others reported that high folate and B12 levels during pregnancy were associated with a higher prevalence of atopic dermatitis in children [130], while third trimester FA supplementation was associated with eczema in children but no other allergic outcomes [131].
In preclinical models, high dose FA (50mg/kg-diet) administered intraperitoneally to a murine model of colitis improved associated inflammation [132], however, others found that a high methyl donor diets including 5mg/kg-diet FA increased susceptibility to colitis in offspring [133,134]. Similarly, excess FA supplementation to rat dams (20mg/kg diet) exacerbated weight gain and inflammation [135]. Our own research demonstrated that high folic acid exposure negatively affected human lymphocyte stress response, genomic stability, and DNA methylation in both in-vitro and in-vivo conditions [136,137].The relationship between FA supplementation and immune-related outcomes is complex, with varying results depending on factors such as supplementation timing, dosage, food fortification, and more. Further research is needed to elucidate the potential mechanisms underlying these associations and to determine the optimal FA supplementation strategies for minimizing immune-related risks.
3.5. Carcinogenesis
Due to the importance of folate cycle in DNA replication and repair, inadequate consumption of folate is considered an independent risk factor for cancer. A low folate diet leads to uracil misincorporation into the DNA leading to single and double stranded breaks [138]. On the other hand, excess folate may increase cancer risk in the presence of preneoplastic lesions [4]. In rapidly proliferating pre-neoplastic or transformed cells, excess folate could help sustain high rates of growth by providing thymidylate and purines for DNA synthesis.
Colorectal cancer (CRC) is the second most prevalent cancer in western societies, and it is closely associated with folate intake. Following the fortification mandate, a small increase in the incidence of colorectal cancer in the USA and Canada was observed [139]. This association has proven controversial as data from human and animal studies yielded contradictory results. Higher folate intake in natural and synthetic forms was associated with decreased risk of CRC in women participating in the nurses’ health study [140]. Others found that vitamin B12 and folic acid supplementation significantly increased CRC incidence [139] and increased overall cancer incidence and mortality [141]. Similarly, a meta-analysis found a borderline significant increase of overall cancer risk with FA supplementation[142]. Another independent meta-analysis of RCT’s did not find any significant effect of FA supplementation on overall cancer or colorectal cancer risk [143]. Supplementation of FA at 1mg/d was associated with increased multiplicity of colorectal adenomas, advanced lesions, and increased risk of prostate cancer in some studies [144,145,146]. However, In a recent systematic review, High folate intake was associated with reduced risk of colorectal cancer is the USA and Europe, particularly among people with middle or high alcohol consumption [147].
The association of folate intake with the incidence of non-CRC cancers is also inconsistent and is reviewed elsewhere [148]. Higher intake of folate was associated with a reduced risk of squamous cell carcinoma of the head and neck [141], esophageal [149,150,151], oral [152], pancreatic [153,154], and bladder cancers [155,156]. We should note that the higher intake in these studies were unlikely to represent excess folate intake and the analysis was not stratified to isolate the effects of intake that approach or exceed the UL. No clear association between high folate intake and lung cancer was observed in a meta-analysis [157], However, a study from Poland found that high systemic folate levels (RBC folate concentrations above 506.5 nmol/l) increased lung cancer risk in heavy smokers[158]. High intake of folate was not protective against ovarian cancer [159]. Despite an overall marginally negative association between endometrial cancer and folate intake, at higher intake levels ranging between 205-987mcg/day, cancer risk increased in a dose dependent manner suggesting a threshold effect [160].
Conflicting associations were found between folate intake and breast cancer risk [161,162,163]. In a meta-analysis of by Chen et al., a U-Shaped relationship was observed in prospective studies where moderate folate intake (153-400mcd/day) decreased breast cancer risk compared to <153mcg/day, while intake >400mcg had no effect [162]. In another analysis by Zhang et al., moderate folate intake between 200-320 mcg/day lowered the risk of breast cancer, while a significantly increased risk was observed with a daily folate intake exceeding 400mcg [161]. recent studies concluded that high intake of one carbon related vitamins including folic acid may be protective against estrogen-receptor-negative/progesterone receptor negative tumors and in individuals with moderate to high alcohol consumption [163,164].
Studies in rodents revealed a complex and sometimes contradictory relationship between folic acid intake and tumorigenesis. The difference in models, timing, duration, dose, and experimental design led to inconsistent findings in rodents. The impact of folic acid supplementation on colorectal cancer in rodents was reviewed in details elsewhere [165]. Notably, in some rodent studies that used high doses of folic acid (8mg FA kg−1), supplementation led to increased occurrence of preneoplastic lesions and CRC [149,150,151]. Increased proliferation of colorectal epithelia and intake of folate after the development of neoplastic lesions was hypothesized to be responsible for the observed increased risk. In an inducible breast cancer mouse model, high FA (10 mg kg−1) significantly increased tumor volume compared to baseline diets (2mg/kg)[166].
Moderately high FA supplementation (5mg FA kg−1) prior to and during pregnancy and lactation in rats increases the risk and multiplicity of mammary adenocarcinomas in the offspring and reduced global DNA methylation [167]. In another rat model, FA supplementation (ranging from 5-10mg FA kg−1) promoted the progression of established mammary tumors [168]. Restricting FA in the diet lowered the incidence of neuroblastoma in the offspring of a predisposed pregnant mice model receiving a low FA diet [169]. In a chemically inducible prostatitis model, maternal folic acid supplementation at a dose of 2mg/kg in rats led to a heightened incidence and severity of inflammation in their offspring [140]. This is especially relevant to the observation that folic acid supplementation was associated with increased risk of prostate cancer in humans [170]. However, further research is needed to validate the effects and explore the underlying mechanisms.
In cell culture studies, FA depletion and supplementation enhanced the transformation of keratinocytes infected by the human papilloma virus [171,172]. Furthermore, depletion and over-supplementation of FA led to increased progression, migration, and invasion of the hepatocarcinoma HepG2 cells [173]. In human lymphoblastoid cell lines, we found that excess FA can mimic folate restriction by similarly impacting oxidative stress response, DNA damage repair, and DNA methylation [136]. The relationship between excess folic acid intake and carcinogenesis presents complex and often contradictory findings. Further research is necessary, particularly to investigate the effects of high levels of folate intake, and better understand the underlying mechanisms and potential risks. It is crucial to consider human genetic polymorphisms, genetic backgrounds, diets, environment, smoking, and other factors that may confound study outcomes. By addressing these factors, future studies can help elucidate the optimal levels of folate intake for various populations and contribute to more accurate public health recommendations.
3.6. Morbidity and mortality.
Numerous studies have sought to determine the relationship between high folate intake, chronic disease risk, and overall mortality, but the results remain inconclusive. A systematic review by Colapinto et al. observed a negative association between high folate levels and adverse health outcomes. However, the variability in study methods, designs, and high folate cutoffs prevented definitive conclusions [18]. A large UK population cohort (n=115664) found that higher dietary folate intake correlated with lower CVD risk and overall mortality[174], and a similar trend was noted in a recent NHANES survey data analysis [175]. Other studies, however, did not find a significant association between high folic acid intake and overall mortality [176,177]. A prospective analysis of 2011-2014 NHANES data revealed a higher mortality risk associated with increased serum folate levels, such as 5-mTHF, UMFA, non-methyl folate, and MeFox (a biologically inactive degradation product of 5-mTHF), as well as with 5-mTHF insufficiency [178]. Another cohort study based on 1999-2010 NHANES data reported a modestly elevated all-cause and cardiovascular mortality in participants with high folate levels (>40 nmol/L) [179].
The impact of high folate levels on overall mortality remains unclear, but some studies have offered insights into the relationship between folate intake and non-cancer health outcomes, particularly metabolic and cardiovascular diseases. A recent systematic review and meta-analysis found that folate supplementation improved fasting glucose, insulin resistance, and insulin levels but had no effect on diabetes or HbA1c [180]. A large meta-analysis of randomized controlled trials did not establish a connection between folic acid supplementation and reduced cardiovascular risk, although a modest benefit was observed in stroke prevention[181]. However, high folate intake was associated with an increased risk of thyroid and endocrine disorders [5,181]. Studies in rats comparing the effects of 5-m-THF and folic acid supplementation on metabolism showed that high doses of either form led to distinct adverse and harmful effects [182]. Research comparing the effects of high intakes of natural and synthetic folate on health and disease is of particular significance, given that very few comparative studies have been conducted in human and animal models. Future investigations into the consequences of excess folate should consider the impacts of supplementation with natural folates, such as 5-m-THF and 5-formyl-THF, in comparison to folic acid.
Folic acid supplementation is commonly used to treat hyperhomocysteinemia, a condition characterized by elevated homocysteine levels [178,183]. Homocysteine is a risk factor for stroke, cardiovascular disease, cognitive function, and adverse pregnancy outcomes [184,185,186,187]. In two separate studies, folic acid supplementation at 0.5 and 5 mg per day were similarly effective at reducing homocysteine levels[188,189]. A dose-finding interventional trial concluded that long-term doses as low as 0.2 mg of folic acid or lower effectively lowered homocysteine levels, with no further significant reduction observed at higher doses[190]. However, a high dose of folic acid (5 mg/day) for three months failed to normalize homocysteine levels in over 40% of participants in a recent study [191].
The relationship between high folate intake, chronic disease risk, and overall mortality remains unclear due to inconsistent results from various studies. Caution must be exercised in interpreting these findings, as the studies employed different methodologies and high folate cutoffs. Given the equivocal results, additional research is needed to better understand the complex relationship between high folate intake and its potential impact on chronic disease risk and overall mortality.
4. Mechanisms Underlying the Adverse Effects of Excess Folate Intake
Several potential mechanisms have been proposed to explain the effects of excess folate. In this section, we will discuss known potential molecular and genetic mechanisms that are hypothesized to mediate the deleterious effects observed in human and animal studies. (Figure 2)
4.1. Excess folate and B12 dependent mechanisms
The aging population is particularly at risk for vitamin B12 deficiency, and there is concern that folic acid (FA) supplementation might mask and exacerbate the severe, potentially irreversible neurological damage caused by B12 deficiency [183,184,192,193]. Excess FA can conceal hematological and neurological symptoms of B12 deficiency, worsening the condition and delaying diagnosis, leading to irreversible morbidities [3,194]. Reynolds et al.’s comprehensive review highlighted evidence that long-term exposure to excess FA, even at doses below the upper limit (0.5-1 mg), combined with B12 deficiency, can lead to neurological harm and cognitive decline [192]. FA supplementation may increase the demand for vitamin B12, exacerbating its depletion, pernicious anemia, and associated neurological and hematological decline [195,196,197]. Following fortification mandates in countries like the United States and Australia, epidemiological studies found a link between excess folic acid and cognitive decline in older adults at risk of B12 deficiency [198,199,200,201]. However, there is still no consensus regarding the role of excess FA in potentiating neurological and cognitive impairment in individuals with low or deficient B12 status [202,203].
A B-vitamin imbalance resulting from high FA combined with inadequate B12 may lead to a functional folate deficiency, contributing to insulin resistance and gestational diabetes mellitus [204]. In pregnant rats, high FA supplementation combined with a B12-deficient diet led to oxidative stress in both the mothers and pups, compared to the same supplementation with adequate B12 status [205]. Excess FA supplementation alongside B12 deficiency during pregnancy and lactation resulted in diet-dependent metabolic impairment in female rat offspring [206]. Similar studies in rats demonstrated that adequate B12 levels are required to mitigate the potential harmful effects of high FA on DNA methylation and neurodevelopment in offspring [207,208].
Mechanistically, the paradoxical combination of excess folate with B12 deficiency leads to unique metabolic situation known as the 5-Methyltetrahydrofolate Trap. The primary enzymatic reaction implicated in the methyl trap is catalyzed by methionine synthase (MTR), a vitamin B12 (cobalamin)-dependent enzyme. MTR catalyzes the conversion of homocysteine to methionine, with 5-m-THF acting as the methyl donor. During this process, 5-m-THF transfers a methyl group to cobalamin, generating methylcobalamin, which subsequently transfers the methyl group to homocysteine, yielding methionine and tetrahydrofolate (THF). THF is essential for purine and pyrimidine synthesis, which are required for DNA and RNA production. In situations of vitamin B12 deficiency, MTR activity is compromised, leading to an accumulation of 5-m-THF, as the enzyme is unable to utilize it as a methyl donor, leading to the phenomenon known as the methyl trap (Figure 3). This sequestration of folate in the form of 5-m-THF results in a functional folate deficiency, as the intracellular pool of THF is depleted. The reduced availability of THF impairs the synthesis of purines and pyrimidines, ultimately affecting DNA and RNA synthesis. Consequently, this disruption in nucleotide synthesis manifests as megaloblastic anemia, characterized by large, immature erythrocytes. Additionally, the vitamin B12 deficiency and the subsequent impairment of MTR activity leads to an accumulation of homocysteine, as it cannot be converted to methionine. Elevated homocysteine levels have been associated with an increased risk of cardiovascular disease, neurodegenerative disorders, and other adverse health outcomes.
The complex interplay between excess folate and B12 deficiency presents a significant challenge in understanding the potential consequences on health, particularly in vulnerable populations such as the elderly and pregnant women. The 5-MTHF trap phenomenon illustrates the critical need for a balanced intake of these essential B-vitamins and highlights the importance of monitoring their levels to prevent detrimental health outcomes. Nevertheless, further research is required to study the long-term effects of this phenomenon and determine the optimal balance of these nutrients. Such studies will be instrumental in establishing evidence-based guidelines and devising effective strategies to prevent, diagnose, and manage the health consequences arising from the interplay between excess folate and B12.
4.2. Mechanisms Related to the One-Carbon Cycle
The one-carbon metabolism plays a crucial role in numerous cellular processes, including DNA synthesis, repair, and methylation. Excess folic acid can disrupt the balance of this metabolic cycle, leading to various adverse health effects. The mechanisms related to the one-carbon cycle can be further divided into the following categories:
4.2.1. Pseudo-MTHFR Deficiency
MTHFR is an enzyme that catalyzes the conversion of 5,10-methylenetetrahydrofolate (5,10-MTHF) to 5-methyltetrahydrofolate (5-MTHF), the biologically active form of folate. This conversion is a critical step in the folate metabolic pathway, as 5-MTHF participates in the re-methylation of homocysteine to methionine. high folic acid intake can lead to an accumulation of unmetabolized folic acid in the bloodstream. The presence of excess folic acid could competitively inhibit the enzymes involved in the conversion of other forms of folate, such as dihydrofolate reductase (DHFR), which converts dietary folic acid to dihydrofolate (DHF) and subsequently to tetrahydrofolate (THF). This competitive inhibition can exacerbate an existing MTHFR deficiency by further disrupting the folate metabolic pathway, leading to a functional folate deficiency despite adequate folate intake. Polymorphisms in the MTHFR gene are associated with reduced enzyme activity and increased health risks. In humans C677T and A1298C polymorphism are common with an estimated 20-40% of the US population harboring one or both mutations [6]. Nevertheless, MTHFR inhibition by excess folate is not exclusively linked to cases where genetic polymorphisms are present. Even in the absence of known MTHFR polymorphisms, elevated levels of folic acid can still result in a pseudo-MTHFR deficiency, which hinders the conversion of 5,10-methylenetetrahydrofolate to 5-MTHF [61,107,209,210,211]. This pseudo-deficiency can disrupt the one-carbon cycle and negatively affect methylation reactions, potentially leading to various health issues, including abnormal embryonic growth and neurodevelopment [210].
4.2.2. Altered folate cycle and Methylation Patterns
Folate is vital for the generation of S-adenosylmethionine (SAM), the primary methyl donor for DNA methylation. However, excess folic acid can disrupt the balance of SAM and its demethylated product, S-adenosylhomocysteine (SAH). This disruption may lead to global DNA hypomethylation, resulting in genomic instability, altered gene expression, and an increased risk of diseases. Excess folic acid intake can cause an imbalance in the distribution of folate coenzymes and derivatives, leading to disruptions in the one-carbon cycle thereby impeding methylation reactions, embryonic growth, and neurodevelopment [210]. High FA levels were linked to dysregulation of intracellular one carbon metabolism [212]. Both deficient and excess maternal folic acid in mice have been shown to cause alterations in offspring’s folate metabolism, favoring DNA synthesis over methylation reactions and leading to cytoarchitectural defects and behavioral abnormalities [103]. Similarly, both folate deficiency and excess supplementation similarly compromised folate dependent pathways, resulting DNA damage, cell cycle arrest, and impaired hematopoiesis in mice [213]. Excessive FA led to abnormal cardiac development and defects in folate metabolism similarly to folate depletion using the antifolate drug methotrexate in a zebrafish model. Interestingly, both FA excess and insufficiency resulted in similar phenotypic and genotypic changes impacting methylation levels and expression of cardiac development related genes [214]. Both excess paternal FA supplementation and FA depletion in mice resulted in a similarly deleterious transgenerational outcomes potentially stemming from altered methylation of imprinted sperm genes [89]. Similarly, paternal FA supplementation in chicken induced transgenerational changes in glucose and lipid metabolism [210]. Excess FA administered during pregnancy, lactation, and post weaning impacted DNA methylation, gene expression, behavior, neural development, and disrupted folate metabolism in mice offspring [83,84,85,215,216,217].
4.2.3. Accumulation of Unmetabolized Folic Acid (UMFA)
High intake of folic acid can result in the accumulation of UMFA in the bloodstream, as the body’s natural capacity to convert folic acid to its active form, 5-methyltetrahydrofolate (5-MTHF), becomes saturated [1,218]. This excess UMFA can compete with and inhibit key enzymes in the one-carbon cycle, such as dihydrofolate reductase (DHFR) and methylenetetrahydrofolate reductase (MTHFR) [219]. As a result, essential reactions in the cycle can be disrupted, affecting DNA synthesis and repair, as well as methylation processes. However, UMFA is not known to accumulate in biological tissues and inside cellular compartments. Unlike reduced folate, UMFA has low affinity for the reduced folate carrier transporter (RFC) and is not a substrate for the enzyme folate polyglutamate synthase (FPGS) [220,221]. Since UMFA cannot participate in one carbon cycle metabolism and binds weekly to folate enzymes, it is also plausible that the potential effects of UMFA could be mediated by non-canonical pathways independent of one carbon metabolism.
4.3. Non-Canonical pathways
4.3.1. Folic acid specific pathways
Folic acid is the fully oxidized form of folate. It is a pre-vitamin that requires a 2-step reduction by DHFR to THF, the metabolically active form participating in the one carbon cycle. However, FA can bind to and be transported into tissues by the folate receptor (FOLR) and the proton coupled folate transporters (PCFT). Excess intake of FA is linked to saturation of the capacity of DHFR leading to accumulation of UMFA. DHFR activity in humans is variable and limited compared to lab rodents [218]. Unlike natural folates that are polyglutamated and retained within the cells, Folic acid in its unmetabolized form is not known to accumulate in tissues. Studies have shown that UMFA circulates at a low levels in individuals not taking supplementation, with more sustained elevation of UMFA recorded in adults ingesting 200ug to 400ug of FA daily [10]. Higher levels of UMFA generally correlate with increased folic acid intake [222,223]. Intakes of FA that surpass the UL are associated with an increased likelihood of having elevated UMFA levels [224,225,226,227,228].
The presence of UMFA is of interest as a potential causative factor in mediating the deleterious effects of excess FA. However, there are significant gaps in understanding the mechanisms involved [229]. one possible mechanism of interest involves the role of folate receptor (FOLR) in binding FA and mediating its transport and triggering potential downstream effectors. Folate receptors have much higher affinity to FA compared to natural folates. The FOLRα facilitates the intake of FA through receptor meditated endocytosis. FOLRα is ubiquitously expressed in the epithelia of normal tissues (kidney, placenta, blood-brain barrier), as well as being overexpressed in many cancers such as ovarian, breast, and colon cancers [230]. The role of FOLRα in mediating tumorigenesis as well as the associated physiological function in cancer are not clear [166,231]. In the presence of FA, FOLRα could translocate into the nucleus and function as a transcription factor impacting genes responsible for cellular pluripotency [222,223]. In fact, FA bound FOLR may function as signaling molecule activating downstream targets such as the pro-oncogene STAT3, extracellular signal-regulated kinases (ERK) kinases, and cellular Sarcoma (SRC) pathways [224,225,226,227,228].
Folic acid, but not reduced folate were linked to regulation of neurogenesis in the ventral hippocampus in rats [229]. Folic acid could also be involved in modulating endothelial NO synthase activity via post translational modifications [230]. A study in piglets found that a high FA diet altered the expression of liver proteins involved in oxidative responses, metabolic regulation, and cancer pathways compared to a control diet (30mg FA/kg vs 1.3 mg FA/kg diet) [232]. Periconceptual excess FA led to upregulated Fos expression in mice [83], and activation of β-catenin (Wnt pathway) in the brain of male mouse offspring [108]. In studies that compared the effects of supplementing excess synthetic and natural folate in rats, the authors observed contrasting metabolic shifts leading to weight gain, leptin dysregulation, and increased food intake int the female offspring when dams where fed excess 5-m-THF compared to excess FA, with FA significantly impacting neurotransmitter signaling genes [182,233]. Taken together, these studies suggest the existence of non-canonical pathways that could mediate the effects of excess folate, both synthetic and natural independently of one carbon cycle. Additional exploratory work and research is needed to fully elucidate the mechanisms involved.
4.3.2. Mechanisms related to elevated folate
Determining the specific effects of excess folic acid intake from elevated folate status is a difficult task. In individuals with elevated folate status the widespread use of folic acid supplementation contributes significantly to folate intake, compared to natural dietary sources. Naturally occurring folate in unfortified foods generally is characterized by its limited content, stability, and bioavailability compared to synthetic folic acid. Consequently, individuals who consume excessive amounts of folate are more likely to be supplemented with synthetic folic acid, potentially surpassing their relative intake of natural dietary folate. Determining whether exogenous folic acid intake or the resulting elevated folate status is responsible for the observed effects is a significant challenge. Nonetheless, studies investigating the relationship between high folate levels and adverse health outcomes managed to identify possible underlying mechanisms.
One area that has received significant research attention pertains to the effects of excess folate on carcinogenesis. The precise causal mechanism underlying folic acid’s role in promoting carcinogenesis remains unclear. Excess folate could elevate cancer risk when preneoplastic lesions are present [4]. In rapidly proliferating pre-neoplastic or transformed cells, an abundance of folate could support accelerated growth by supplying thymidylate and purines for DNA synthesis [234]. Furthermore, folate availability has a direct and substantial influence on DNA methylation. Disturbances in folate levels, arising from either low or high folate status, can lead to hypo- or hypermethylation, resulting in epigenetic instability [235,236,237]. Disruption of normal DNA methylation patterns can lead to the dysregulation of gene expression and genomic instability, ultimately contributing to the initiation and progression of cancer. DNA hypomethylation is considered an early event in colon carcinogenesis [238]. Excess FA is capable of inducing hypomethylation in cultured human cells [239], and alteration in methyl metabolism in mice [104,209]. Work in multiple models suggest that both folate deficiency and excess FA can disrupt the folate cycle and the connected methionine cycle leading to dysregulated methylation [103,210,240,241]. In our investigations involving human lymphocytes, we observed that high folic acid intake relative to natural folate, can influence DNA methylation, disrupt genomic stability, and diminish the capacity to withstand with oxidative stress [27,136]. Excess folic acid supplemented to cancer cell organoid, can counteract methionine dependency, thus promoting cancer stem cell formation [242]. In mice, a high FA diet promoted hepatocellular carcinoma development through stabilization of the methionine cycle enzyme MATIIα. In prostate cancer, folate was found to be directly involved in oncogenic signaling pathways, specifically the PI3K-AKT-mTOR cascade [243]. In our study exploring the impact of folate restriction on colon carcinogenesis, we observed a significant increase in mTOR signaling in mice supplemented with folic acid compared to those without supplementation [26].
The role of excess folate in genomic methylation and epigenetic reprogramming is not limited to cancer etiology. As previously mentioned, excessive folate intake during pregnancy has been linked to both adverse and favorable birth outcomes, as well as an elevated risk of disease in offspring, as demonstrated in both human and animal studies. Folic acid supplementation impacts one-carbon metabolism during embryogenesis, which in turn leads to changes in genome wide and gene specific methylation patterns [82]. In rodent models, excess maternal folic acid supplementation has been demonstrated to alter methylation and expression of imprinted genes, histone modifications, and heterochromatin assembly, However the underlying epigenetic mechanism is still unknown [71,82]. In humans, maternal FA supplementation was linked to changes in DNA methylation of genes related to brain development with beneficial effects on cognitive development [80,92,93]. The in-vivo and in-vitro mechanisms through which excess maternal FA supplementation can influence neurological development were reviewed elsewhere [244], detailing behavioral, morphological, and molecular changes in the offspring’s brain exposed to FA over-supplementation. A recent investigation into some of the underlying mechanisms discovered significant changes in DNA methylation of neurodevelopmental genes, which play a crucial role in transcriptional regulation [245]. Notably, these modified epigenetic patterns persisted throughout an individual’s life, as demonstrated in the Aberdeen Folic Acid Supplementation Trial [246]. Although both animal and human studies have identified a correlation between excessive maternal FA intake and metabolic abnormalities in offspring, a clear causative link with the modified epigenetic landscape has not yet been established [68,76,77,78,247,248]. Further research is required to establish if epigenetic reprogramming is partially or wholly responsible for both the beneficial and adverse effects of excess folate.
Both folate deficiency and excess are known to impact the immune system and its function which could result in increased susceptibility to inflammation, chronic diseases, and cancer. Lymphocytes are especially sensitive to changes in the folate pool due to high proliferation rate, making them a robust biomarker for folate status [138,251]. Excess folic acid intake is also linked to impaired natural killer cell activity among postmenopausal women, healthy adults, and aged mice [115,249,250]. Both folate deficiency and excess impaired metabolism and hematopoiesis in mice [213]. Our findings in human lymphocytes aligned with these observations, as we identified impaired DNA repair and oxidative stress response under conditions of excess folic acid and high folic acid to natural folate intake ratio, both in-vitro and in-vivo settings, respectively [136,137]. Others found that folic acid supplementation could promote a proinflammatory transcriptomic milieu in the colon [251,252], and, that excess FA increased inflammation in response to high fat diet in rats [253]. It remains an open question whether the observed perturbations in the immune system and inflammatory response are responsible for some of the deleterious effects of excess folate.
Recently, the one-carbon cycle has been identified as a central regulator of organismal aging [254,255]. Metabolomic analysis of long-lived C. elegans and mouse models revealed common signatures and regulation of the one-carbon folate cycle. In a robust analysis, Annibal et al. demonstrated that supplementing long-lived C. elegans mutants with physiological levels of folate partially abrogated the lifespan extension, while RNAi knockdown of the key enzyme DHFR extended it in the wild-type worms [254]. This work highlighted the folate cycle as a shared causal mechanism for longevity. In our preliminary work with mice, we observed increased longevity in wild-type C57/Bl6 mice on a folic acid-restricted diet compared to those receiving folic acid supplemented diet (86% vs. 70% survival rate at 690 days, respectively). Similarly, our pilot work revealed that exposing C. elegans nematodes to the DHFR inhibitor methotrexate significantly increased the worm’s lifespan, while supplementing the media with either folic acid, 5-formyl-THF (leucovorin), or 5-methyl-THF significantly shortened lifespan in the wildtype N2 worms in a dose-dependent manner.
5. Conclusions
As researchers, including ourselves, strive to unravel the conserved and intricate dynamics governing the relationship between folate, health, and longevity, a crucial question arises: could exposing the human population to excessive levels of folate potentially accelerate aging and increase the risk of disease?
The key to answering this question lies in identifying causal underlying mechanisms that can robustly link excess folate intake to the observed detrimental effects. Achieving this goal requires substantial research efforts and collaboration across multiple scientific disciplines to identify and address the existing knowledge gaps. The importance of pursuing such research cannot be overstated, as these gaps in knowledge may be unknowingly and insidiously compromising the health of hundreds of millions of people in the USA and worldwide who are consuming excess folate.
Author Contributions
writing—original draft preparation, A.M.F.; writing—review and editing, A.M.F., A.R.H; All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maruvada, P.; Stover, P.J.; Mason, J.B.; Bailey, R.L.; Davis, C.D.; Field, M.S.; Finnell, R.H.; Garza, C.; Green, R.; Gueant, J.-L., et al. Knowledge gaps in understanding the metabolic and clinical effects of excess folates/folic acid: a summary, and perspectives, from an NIH workshop. The American Journal of Clinical Nutrition 2020, 112, 1390-1403. [CrossRef]
- Field, M.S.; Stover, P.J. Safety of folic acid. Annals of the New York Academy of Sciences 2018, 1414, 59-71. [CrossRef]
- Selhub, J.; Rosenberg, I.H. Excessive folic acid intake and relation to adverse health outcome. Biochimie 2016, 126, 71-78. [CrossRef]
- Crider, K.S.; Bailey, L.B.; Berry, R.J. Folic acid food fortification-its history, effect, concerns, and future directions. Nutrients 2011, 3, 370-384. [CrossRef]
- Shulpekova, Y.; Nechaev, V.; Kardasheva, S.; Sedova, A.; Kurbatova, A.; Bueverova, E.; Kopylov, A.; Malsagova, K.; Dlamini, J.C.; Ivashkin, V. The Concept of Folic Acid in Health and Disease. Molecules 2021, 26, 3731. [CrossRef]
- Menezo, Y.; Elder, K.; Clement, A.; Clement, P. Folic Acid, Folinic Acid, 5 Methyl TetraHydroFolate Supplementation for Mutations That Affect Epigenesis through the Folate and One-Carbon Cycles. Biomolecules 2022, 12, 197. [CrossRef]
- Wilson, R.D.; O’Connor, D.L. Maternal folic acid and multivitamin supplementation: International clinical evidence with considerations for the prevention of folate-sensitive birth defects. Prev Med Rep 2021, 24, 101617. [CrossRef]
- P, M.; Pj, S.; Jb, M.; Rl, B.; Cd, D.; Ms, F.; Rh, F.; C, G.; R, G.; Jl, G., et al. Knowledge gaps in understanding the metabolic and clinical effects of excess folates/folic acid: a summary, and perspectives, from an NIH workshop. The American journal of clinical nutrition 2020, 112, 1390-1403. [CrossRef]
- Obeid, R.; Kirsch, S.H.; Dilmann, S.; Klein, C.; Eckert, R.; Geisel, J.; Herrmann, W. Folic acid causes higher prevalence of detectable unmetabolized folic acid in serum than B-complex: a randomized trial. European Journal of Nutrition 2016, 55, 1021-1028. [CrossRef]
- Sweeney, M.; McPartlin, J.; Scott, J. Folic acid fortification and public health: Report on threshold doses above which unmetabolised folic acid appear in serum. BMC Public Health 2007, 10.1186/1471-2458-7-41. [CrossRef]
- Murphy, M.S.Q.; Muldoon, K.A.; Sheyholislami, H.; Behan, N.; Lamers, Y.; Rybak, N.; White, R.R.; Harvey, A.L.J.; Gaudet, L.M.; Smith, G.N., et al. Impact of high-dose folic acid supplementation in pregnancy on biomarkers of folate status and 1-carbon metabolism: An ancillary study of the Folic Acid Clinical Trial (FACT). Am J Clin Nutr 2021, 113, 1361-1371. [CrossRef]
- Palchetti, C.Z.; Paniz, C.; De Carli, E.; Marchioni, D.M.; Colli, C.; Steluti, J.; Pfeiffer, C.M.; Fazili, Z.; Guerra-Shinohara, E.M. Association between Serum Unmetabolized Folic Acid Concentrations and Folic Acid from Fortified Foods. Journal of the American College of Nutrition 2017, 36, 572-578. [CrossRef]
- Williams, B.A.; Mayer, C.; McCartney, H.; Devlin, A.M.; Lamers, Y.; Vercauteren, S.M.; Wu, J.K.; Karakochuk, C.D. Detectable Unmetabolized Folic Acid and Elevated Folate Concentrations in Folic Acid-Supplemented Canadian Children With Sickle Cell Disease. Front Nutr 2021, 8, 642306. [CrossRef]
- Pfeiffer, C.M.; Sternberg, M.R.; Fazili, Z.; Yetley, E.A.; Lacher, D.A.; Bailey, R.L.; Johnson, C.L. Unmetabolized folic acid is detected in nearly all serum samples from US children, adolescents, and adults. J Nutr 2015, 145, 520-531. [CrossRef]
- Pfeiffer, C.M.; Johnson, C.L.; Jain, R.B.; Yetley, E.A.; Picciano, M.F.; Rader, J.I.; Fisher, K.D.; Mulinare, J.; Osterloh, J.D. Trends in blood folate and vitamin B-12 concentrations in the United States, 1988 2004. Am J Clin Nutr 2007, 86, 718-727.
- Pfeiffer, C.M.; Hughes, J.P.; Lacher, D.A.; Bailey, R.L.; Berry, R.J.; Zhang, M.; Yetley, E.A.; Rader, J.I.; Sempos, C.T.; Johnson, C.L. Estimation of Trends in Serum and RBC Folate in the U.S. Population from Pre- to Postfortification Using Assay-Adjusted Data from the NHANES 1988–2010. The Journal of Nutrition 2012, 142, 886-893. [CrossRef]
- Bailey, R.L.; Dodd, K.W.; Gahche, J.J.; Dwyer, J.T.; McDowell, M.A.; Yetley, E.A.; Sempos, C.A.; Burt, V.L.; Radimer, K.L.; Picciano, M.F. Total folate and folic acid intake from foods and dietary supplements in the United States: 2003-2006. Am J Clin Nutr 2010, 91, 231-237. [CrossRef]
- Colapinto, C.K.; O’Connor, D.L.; Sampson, M.; Williams, B.; Tremblay, M.S. Systematic review of adverse health outcomes associated with high serum or red blood cell folate concentrations. Journal of Public Health 2016, 38, e84-e97. [CrossRef]
- Colapinto, C.K.; O’Connor, D.L.; Dubois, L.; Tremblay, M.S. Prevalence and correlates of high red blood cell folate concentrations in the Canadian population using 3 proposed cut-offs. Applied Physiology, Nutrition, and Metabolism 2015, 40, 1025-1030. [CrossRef]
- Colapinto, C.K.; O’Connor, D.L.; Tremblay, M.S. Folate status of the population in the Canadian Health Measures Survey. Canadian Medical Association Journal 2011, 183, E100-E106. [CrossRef]
- Fazili, Z.; Sternberg, M.R.; Potischman, N.; Wang, C.-Y.; Storandt, R.J.; Yeung, L.; Yamini, S.; Gahche, J.J.; Juan, W.; Qi, Y.P., et al. Demographic, Physiologic, and Lifestyle Characteristics Observed with Serum Total Folate Differ Among Folate Forms: Cross-Sectional Data from Fasting Samples in the NHANES 2011–2016. The Journal of Nutrition 2019, 150, 851-860. [CrossRef]
- Plumptre, L.; Tammen, S.A.; Sohn, K.J.; Masih, S.P.; Visentin, C.E.; Aufreiter, S.; Malysheva, O.; Schroder, T.H.; Ly, A.; Berger, H., et al. Maternal and Cord Blood Folate Concentrations Are Inversely Associated with Fetal DNA Hydroxymethylation, but Not DNA Methylation, in a Cohort of Pregnant Canadian Women. J Nutr 2020, 150, 202-211. [CrossRef]
- Plumptre, L.; Masih, S.P.; Ly, A.; Aufreiter, S.; Sohn, K.-J.; Croxford, R.; Lausman, A.Y.; Berger, H.; O’Connor, D.L.; Kim, Y.-I. High concentrations of folate and unmetabolized folic acid in a cohort of pregnant Canadian women and umbilical cord blood. The American Journal of Clinical Nutrition 2015, 102, 848-857. [CrossRef]
- Gómez, M.F.; Field, C.J.; Olstad, D.L.; Loehr, S.; Ramage, S.; McCargar, L.J.; Team, t.A.S. Use of micronutrient supplements among pregnant women in Alberta: results from the Alberta Pregnancy Outcomes and Nutrition (APrON) cohort. Maternal & Child Nutrition 2015, 11, 497-510. [CrossRef]
- Maruvada, P.; Stover, P.J.; Mason, J.B.; Bailey, R.L.; Davis, C.D.; Field, M.S.; Finnell, R.H.; Garza, C.; Green, R.; Gueant, J.L., et al. Knowledge gaps in understanding the metabolic and clinical effects of excess folates/folic acid: a summary, and perspectives, from an NIH workshop. Am J Clin Nutr 2020, 112, 1390-1403. [CrossRef]
- Fardous, A.M.; Beydoun, S.; James, A.A.; Ma, H.; Cabelof, D.C.; Unnikrishnan, A.; Heydari, A.R. The Timing and Duration of Folate Restriction Differentially Impacts Colon Carcinogenesis. Nutrients 2022, 14, 16.
- Alnabbat, K.I.; Fardous, A.M.; Shahab, A.; James, A.A.; Bahry, M.R.; Heydari, A.R. High Dietary Folic Acid Intake Is Associated with Genomic Instability in Peripheral Lymphocytes of Healthy Adults. Nutrients 2022, 14. [CrossRef]
- Reeves, P.G. Components of the AIN-93 Diets as Improvements in the AIN-76A Diet. The Journal of Nutrition 1997, 10.1093/jn/127.5.838s. [CrossRef]
- Council, N.R. Nutrient Requirements of Laboratory Animals,: Fourth Revised Edition, 1995; The National Academies Press: Washington, DC, 1995 . [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N., et al. HMDB 4.0: the human metabolome database for 2018. Nucleic Acids Research 2017, 46, D608-D617. [CrossRef]
- Fazili, Z.; Pfeiffer, C.M.; Zhang, M. Comparison of Serum Folate Species Analyzed by LC-MS/MS with Total Folate Measured by Microbiologic Assay and Bio-Rad Radioassay. Clinical Chemistry 2007, 53, 781-784. [CrossRef]
- Lee, W.D.; Pirona, A.C.; Sarvin, B.; Stern, A.; Nevo-Dinur, K.; Besser, E.; Sarvin, N.; Lagziel, S.; Mukha, D.; Raz, S., et al. Tumor Reliance on Cytosolic versus Mitochondrial One-Carbon Flux Depends on Folate Availability. Cell Metab 2021, 33, 190-198.e196. [CrossRef]
- López, J.M.; Outtrim, E.L.; Fu, R.; Sutcliffe, D.J.; Torres, R.J.; Jinnah, H.A. Physiological levels of folic acid reveal purine alterations in Lesch-Nyhan disease. Proc Natl Acad Sci U S A 2020, 117, 12071-12079. [CrossRef]
- Silva, C.; Keating, E.; Pinto, E. The impact of folic acid supplementation on gestational and long term health: Critical temporal windows, benefits and risks. Porto Biomed J 2017, 2, 315-332. [CrossRef]
- Ledowsky, C.; Mahimbo, A.; Scarf, V.; Steel, A. Women Taking a Folic Acid Supplement in Countries with Mandatory Food Fortification Programs May Be Exceeding the Upper Tolerable Limit of Folic Acid: A Systematic Review. Nutrients 2022, 14. [CrossRef]
- Cheng, T.L.; Mistry, K.B.; Wang, G.; Zuckerman, B.; Wang, X. Folate Nutrition Status in Mothers of the Boston Birth Cohort, Sample of a US Urban Low-Income Population. American Journal of Public Health 2018, 108, 799-807. [CrossRef]
- Page, R.; Robichaud, A.; Arbuckle, T.E.; Fraser, W.D.; Macfarlane, A.J. Total folate and unmetabolized folic acid in the breast milk of a cross-section of Canadian women. The American Journal of Clinical Nutrition 2017, 105, 1101-1109. [CrossRef]
- Wu, J.; Zheng, Q.; Huang, Y.-Q.; Wang, Y.; Li, S.; Lu, D.-W.; Shi, B.; Chen, H.-Q. Significant Evidence of Association Between Polymorphisms in ZNF533, Environmental Factors, and Nonsyndromic Orofacial Clefts in the Western Han Chinese Population. DNA and Cell Biology 2010, 30, 47-54. [CrossRef]
- Wilcox, A.J.; Lie, R.T.; Solvoll, K.; Taylor, J.; McConnaughey, D.R.; Abyholm, F.; Vindenes, H.; Vollset, S.E.; Drevon, C.A. Folic acid supplements and risk of facial clefts: national population based case-control study. Bmj 2007, 334, 464. [CrossRef]
- Bille, C.; Olsen, J.; Vach, W.; Knudsen, V.K.; Olsen, S.F.; Rasmussen, K.; Murray, J.C.; Andersen, A.M.N.; Christensen, K. Oral clefts and life style factors — A case-cohort study based on prospective Danish data. European Journal of Epidemiology 2007, 22, 173-181. [CrossRef]
- Jia, Z.-L.; Shi, B.; Chen, C.-H.; Shi, J.-Y.; Wu, J.; Xu, X. Maternal malnutrition, environmental exposure during pregnancy and the risk of non-syndromic orofacial clefts. Oral Diseases 2011, 17, 584-589. [CrossRef]
- Wehby, G.; Félix, T.; Goco, N.; Richieri-Costa, A.; Chakraborty, H.; Souza, J.; Pereira, R.; Padovani, C.; Moretti-Ferreira, D.; Murray, J. High Dosage Folic Acid Supplementation, Oral Cleft Recurrence and Fetal Growth. International Journal of Environmental Research and Public Health 2013, 10, 590-605. [CrossRef]
- Little, J.; Gilmour, M.; Mossey, P.A.; Fitzpatrick, D.; Cardy, A.; Clayton-Smith, J.; Fryer, A.E. Folate and Clefts of the Lip and Palate—A U.K.-Based Case-Control Study: Part I: Dietary and Supplemental Folate. The Cleft Palate-Craniofacial Journal 2008, 45, 420-427. [CrossRef]
- Rozendaal, A.M.; Van Essen, A.J.; Te Meerman, G.J.; Bakker, M.K.; Van Der Biezen, J.J.; Goorhuis-Brouwer, S.M.; Vermeij-Keers, C.; De Walle, H.E.K. Periconceptional folic acid associated with an increased risk of oral clefts relative to non-folate related malformations in the Northern Netherlands: a population based case-control study. European Journal of Epidemiology 2013, 28, 875-887. [CrossRef]
- Lassi, Z.S.; Salam, R.A.; Haider, B.A.; Bhutta, Z.A. Folic acid supplementation during pregnancy for maternal health and pregnancy outcomes. Cochrane Database of Systematic Reviews 2013, 10.1002/14651858.CD006896.pub2. [CrossRef]
- Chatzi, L.; Papadopoulou, E.; Koutra, K.; Roumeliotaki, T.; Georgiou, V.; Stratakis, N.; Lebentakou, V.; Karachaliou, M.; Vassilaki, M.; Kogevinas, M. Effect of high doses of folic acid supplementation in early pregnancy on child neurodevelopment at 18 months of age: the mother-child cohort ‘Rhea’ study in Crete, Greece. Public health nutrition 2012, 15, 1728-1736. [CrossRef]
- Csáky-Szunyogh, M.; Vereczkey, A.; Kósa, Z.; Gerencsér, B.; Czeizel, A.E. Risk Factors in the Origin of Congenital Left-Ventricular Outflow-Tract Obstruction Defects of the Heart: A Population-Based Case–Control Study. Pediatric Cardiology 2014, 35, 108-120. [CrossRef]
- Vereczkey, A.; Kósa, Z.; Csáky-Szunyogh, M.; Czeizel, A.E. Isolated atrioventricular canal defects: Birth outcomes and risk factors: A population-based hungarian case–control study, 1980–1996. Birth Defects Research Part A: Clinical and Molecular Teratology 2013, 97, 217-224. [CrossRef]
- Xie, K.; Xu, P.; Fu, Z.; Gu, X.; Li, H.; Cui, X.; You, L.; Zhu, L.; Ji, C.; Guo, X. Association of maternal folate status in the second trimester of pregnancy with the risk of gestational diabetes mellitus. Food Science & Nutrition 2019, 7, 3759-3765. [CrossRef]
- Zhu, B.; Ge, X.; Huang, K.; Mao, L.; Yan, S.; Xu, Y.; Huang, S.; Hao, J.; Zhu, P.; Niu, Y., et al. Folic Acid Supplement Intake in Early Pregnancy Increases Risk of Gestational Diabetes Mellitus: Evidence From a Prospective Cohort Study. Diabetes Care 2016, 39, e36-e37. [CrossRef]
- Cheng, G.; Sha, T.; Gao, X.; He, Q.; Wu, X.; Tian, Q.; Yang, F.; Tang, C.; Wu, X.; Xie, Q., et al. The Associations between the Duration of Folic Acid Supplementation, Gestational Diabetes Mellitus, and Adverse Birth Outcomes based on a Birth Cohort. International Journal of Environmental Research and Public Health 2019, 16, 4511.
- Huang, L.; Yu, X.; Li, L.; Chen, Y.; Yang, Y.; Yang, Y.; Hu, Y.; Zhao, Y.; Tang, H.; Xu, D., et al. Duration of periconceptional folic acid supplementation and risk of gestational diabetes mellitus. Asia Pac J Clin Nutr 2019, 28, 321-329. [CrossRef]
- Petersen, J.M.; Parker, S.E.; Benedum, C.M.; Mitchell, A.A.; Tinker, S.C.; Werler, M.M. Periconceptional folic acid and risk for neural tube defects among higher risk pregnancies. Birth Defects Res 2019, 111, 1501-1512. [CrossRef]
- Chen, X.; Zhang, Y.; Chen, H.; Jiang, Y.; Wang, Y.; Wang, D.; Li, M.; Dou, Y.; Sun, X.; Huang, G., et al. Association of Maternal Folate and Vitamin B12 in Early Pregnancy With Gestational Diabetes Mellitus: A Prospective Cohort Study. Diabetes Care 2021, 44, 217-223. [CrossRef]
- Li, S.; Hou, Y.; Yan, X.; Wang, Y.; Shi, C.; Wu, X.; Liu, H.; Zhang, L.; Zhang, X.; Liu, J., et al. Joint effects of folate and vitamin B12 imbalance with maternal characteristics on gestational diabetes mellitus. Journal of Diabetes 2019, 11, 744-751. [CrossRef]
- Pastor-Valero, M.; Navarrete-Muoz, E.M.; Rebagliato, M.; Iñiguez, C.; Murcia, M.; Marco, A.; Ballester, F.; Vioque, J. Periconceptional folic acid supplementation and anthropometric measures at birth in a cohort of pregnant women in Valencia, Spain. British Journal of Nutrition 2011, 105, 1352-1360. [CrossRef]
- Michels, A.; Bakkali, N.E.; Bastiaenen, C.H.; De Bie, R.A.; Colla, C.G.; Van der Hulst, R.R. Periconceptional Folic Acid Use and the Prevalence of Positional Plagiocephaly. Journal of Craniofacial Surgery 2008, 19, 37-39. [CrossRef]
- James, P.; Sajjadi, S.; Tomar, A.S.; Saffari, A.; Fall, C.H.D.; Prentice, A.M.; Shrestha, S.; Issarapu, P.; Yadav, D.K.; Kaur, L., et al. Candidate genes linking maternal nutrient exposure to offspring health via DNA methylation: a review of existing evidence in humans with specific focus on one-carbon metabolism. Int J Epidemiol 2018, 47, 1910-1937. [CrossRef]
- Callinan, P.A.; Feinberg, A.P. The emerging science of epigenomics. Hum Mol Genet 2006, 15 Spec No 1, R95-101. [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N Engl J Med 2008, 359, 61-73. [CrossRef]
- Mikael, L.G.; Deng, L.; Paul, L.; Selhub, J.; Rozen, R. Moderately high intake of folic acid has a negative impact on mouse embryonic development. Birth Defects Research Part A: Clinical and Molecular Teratology 2013, 97, 47-52. [CrossRef]
- Barua, S.; Chadman, K.K.; Kuizon, S.; Buenaventura, D.; Stapley, N.W.; Ruocco, F.; Begum, U.; Guariglia, S.R.; Brown, W.T.; Junaid, M.A. Increasing maternal or post-weaning folic acid alters gene expression and moderately changes behavior in the offspring. PLoS ONE 2014, 10.1371/journal.pone.0101674. [CrossRef]
- Whitrow, M.J.; Moore, V.M.; Rumbold, A.R.; Davies, M.J. Effect of Supplemental Folic Acid in Pregnancy on Childhood Asthma: A Prospective Birth Cohort Study. American Journal of Epidemiology 2009, 170, 1486-1493. [CrossRef]
- Burdge, G.C.; Lillycrop, K.A. Folic acid supplementation in pregnancy: Are there devils in the detail? Br J Nutr 2012, 108, 1924-1930. [CrossRef]
- Haggarty, P.; Hoad, G.; Campbell, D.M.; Horgan, G.W.; Piyathilake, C.; McNeill, G. Folate in pregnancy and imprinted gene and repeat element methylation in the offspring. The American Journal of Clinical Nutrition 2012, 97, 94-99. [CrossRef]
- Hoyo, C.; Murtha, A.P.; Schildkraut, J.M.; Forman, M.R.; Calingaert, B.; Demark-Wahnefried, W.; Kurtzberg, J.; Jirtle, R.L.; Murphy, S.K. Folic acid supplementation before and during pregnancy in the Newborn Epigenetics STudy (NEST). BMC Public Health 2011, 10.1186/1471-2458-11-46. [CrossRef]
- Hoyo, C.; Murtha, A.P.; Schildkraut, J.M.; Jirtle, R.L.; Demark-Wahnefried, W.; Forman, M.R.; Iversen, E.S.; Kurtzberg, J.; Overcash, F.; Huang, Z., et al. Methylation variation at IGF2 differentially methylated regions and maternal folic acid use before and during pregnancy. Epigenetics 2011, 6, 928-936. [CrossRef]
- Krishnaveni, G.V.; Veena, S.R.; Karat, S.C.; Yajnik, C.S.; Fall, C.H.D. Association between maternal folate concentrations during pregnancy and insulin resistance in Indian children. Diabetologia 2014, 57, 110-121. [CrossRef]
- Xie, R.H.; Liu, Y.J.; Retnakaran, R.; Macfarlane, A.J.; Hamilton, J.; Smith, G.; Walker, M.C.; Wen, S.W. Maternal folate status and obesity/insulin resistance in the offspring: a systematic review. International Journal of Obesity 2016, 40, 1-9. [CrossRef]
- Sie, K.K.Y.; Li, J.; Ly, A.; Sohn, K.-J.; Croxford, R.; Kim, Y.-I. Effect of maternal and postweaning folic acid supplementation on global and gene-specific DNA methylation in the liver of the rat offspring. Molecular Nutrition & Food Research 2013, 57, 677-685. [CrossRef]
- Ly, L.; Chan, D.; Landry, M.; Angle, C.; Martel, J.; Trasler, J. Impact of mothers’ early life exposure to low or high folate on progeny outcome and DNA methylation patterns. Environmental Epigenetics 2020, 6. [CrossRef]
- Hammoud, R.; Pannia, E.; Kubant, R.; Wasek, B.; Bottiglieri, T.; Malysheva, O.V.; Caudill, M.A.; Anderson, G.H. Choline and Folic Acid in Diets Consumed during Pregnancy Interact to Program Food Intake and Metabolic Regulation of Male Wistar Rat Offspring. The Journal of Nutrition 2021, 151, 857-865. [CrossRef]
- Cho, C.E.; Sánchez-Hernández, D.; Reza-López, S.A.; Huot, P.S.; Kim, Y.I.; Anderson, G.H. High folate gestational and post-weaning diets alter hypothalamic feeding pathways by DNA methylation in Wistar rat offspring. Epigenetics 2013, 8, 710-719. [CrossRef]
- Tojal, A.; Neves, C.; Veiga, H.; Ferreira, S.; Rodrigues, I.; Martel, F.; Calhau, C.; Negrão, R.; Keating, E. Perigestational high folic acid: impact on offspring’s peripheral metabolic response. Food & Function 2019, 10, 7216-7226. [CrossRef]
- Kintaka, Y.; Wada, N.; Shioda, S.; Nakamura, S.; Yamazaki, Y.; Mochizuki, K. Excessive folic acid supplementation in pregnant mice impairs insulin secretion and induces the expression of genes associated with fatty liver in their offspring. Heliyon 2020, 6, e03597. [CrossRef]
- Morakinyo, A.O.; Samuel, T.A.; Awobajo, F.O.; Oludare, G.O.; Mofolorunso, A. High-Dose Perinatal Folic-Acid Supplementation Alters Insulin Sensitivity in Sprague-Dawley Rats and Diminishes the Expression of Adiponectin. J Diet Suppl 2019, 16, 14-26. [CrossRef]
- Huang, Y.; He, Y.; Sun, X.; He, Y.; Li, Y.; Sun, C. Maternal high folic acid supplement promotes glucose intolerance and insulin resistance in male mouse offspring fed a high-fat diet. International Journal of Molecular Sciences 2014, 10.3390/ijms15046298. [CrossRef]
- Keating, E.; Correia-Branco, A.; Araújo, J.R.; Meireles, M.; Fernandes, R.; Guardão, L.; Guimarães, J.T.; Martel, F.; Calhau, C. Excess perigestational folic acid exposure induces metabolic dysfunction in post-natal life. Journal of Endocrinology 2015, 224, 245-259. [CrossRef]
- Caldwell, P.T.; Manziello, A.; Howard, J.; Palbykin, B.; Runyan, R.B.; Selmin, O. Gene expression profiling in the fetal cardiac tissue after folate and low-dose trichloroethylene exposure. Birth Defects Research Part A: Clinical and Molecular Teratology 2010, 88, 111-127. [CrossRef]
- Caffrey, A.; Irwin, R.E.; McNulty, H.; Strain, J.J.; Lees-Murdock, D.J.; McNulty, B.A.; Ward, M.; Walsh, C.P.; Pentieva, K. Gene-specific DNA methylation in newborns in response to folic acid supplementation during the second and third trimesters of pregnancy: epigenetic analysis from a randomized controlled trial. Am J Clin Nutr 2018, 107, 566-575. [CrossRef]
- Irwin, R.E.; Pentieva, K.; Cassidy, T.; Lees-Murdock, D.J.; McLaughlin, M.; Prasad, G.; McNulty, H.; Walsh, C.P. The interplay between DNA methylation, folate and neurocognitive development. Epigenomics 2016, 8, 863-879. [CrossRef]
- Liu, H.Y.; Liu, S.M.; Zhang, Y.Z. Maternal Folic Acid Supplementation Mediates Offspring Health via DNA Methylation. Reprod Sci 2020, 27, 963-976. [CrossRef]
- Chu, D.; Li, L.; Jiang, Y.; Tan, J.; Ji, J.; Zhang, Y.; Jin, N.; Liu, F. Excess Folic Acid Supplementation Before and During Pregnancy and Lactation Activates Fos Gene Expression and Alters Behaviors in Male Mouse Offspring. Front Neurosci 2019, 13, 313. [CrossRef]
- Barua, S.; Chadman, K.K.; Kuizon, S.; Buenaventura, D.; Stapley, N.W.; Ruocco, F.; Begum, U.; Guariglia, S.R.; Brown, W.T.; Junaid, M.A. Increasing Maternal or Post-Weaning Folic Acid Alters Gene Expression and Moderately Changes Behavior in the Offspring. PLOS ONE 2014, 9, e101674. [CrossRef]
- Barua, S.; Kuizon, S.; Brown, W.T.; Junaid, M.A. DNA Methylation Profiling at Single-Base Resolution Reveals Gestational Folic Acid Supplementation Influences the Epigenome of Mouse Offspring Cerebellum. Frontiers in Neuroscience 2016, 10. [CrossRef]
- Huot, P.S.; Ly, A.; Szeto, I.M.; Reza-López, S.A.; Cho, D.; Kim, Y.I.; Anderson, G.H. Maternal and postweaning folic acid supplementation interact to influence body weight, insulin resistance, and food intake regulatory gene expression in rat offspring in a sex-specific manner. Appl Physiol Nutr Metab 2016, 41, 411-420. [CrossRef]
- Soubry, A. POHaD: why we should study future fathers. Environmental Epigenetics 2018, 4. [CrossRef]
- Martín-Calvo, N.; Mínguez-Alarcón, L.; Gaskins, A.J.; Nassan, F.L.; Williams, P.L.; Souter, I.; Hauser, R.; Chavarro, J.E. Paternal preconception folate intake in relation to gestational age at delivery and birthweight of newborns conceived through assisted reproduction. Reprod Biomed Online 2019, 39, 835-843. [CrossRef]
- Ly, L.; Chan, D.; Aarabi, M.; Landry, M.; Behan, N.A.; MacFarlane, A.J.; Trasler, J. Intergenerational impact of paternal lifetime exposures to both folic acid deficiency and supplementation on reproductive outcomes and imprinted gene methylation. Molecular Human Reproduction 2017, 23, 461-477. [CrossRef]
- Aarabi, M.; Christensen, K.E.; Chan, D.; Leclerc, D.; Landry, M.; Ly, L.; Rozen, R.; Trasler, J. Testicular MTHFR deficiency may explain sperm DNA hypomethylation associated with high dose folic acid supplementation. Hum Mol Genet 2018, 27, 1123-1135. [CrossRef]
- Rahimi, S.; Martel, J.; Karahan, G.; Angle, C.; Behan, N.A.; Chan, D.; Macfarlane, A.J.; Trasler, J.M. Moderate maternal folic acid supplementation ameliorates adverse embryonic and epigenetic outcomes associated with assisted reproduction in a mouse model. Human Reproduction 2019, 34, 851-862. [CrossRef]
- McNulty, H.; Rollins, M.; Cassidy, T.; Caffrey, A.; Marshall, B.; Dornan, J.; McLaughlin, M.; McNulty, B.A.; Ward, M.; Strain, J.J., et al. Effect of continued folic acid supplementation beyond the first trimester of pregnancy on cognitive performance in the child: a follow-up study from a randomized controlled trial (FASSTT Offspring Trial). BMC Medicine 2019, 17, 196. [CrossRef]
- Caffrey, A.; McNulty, H.; Rollins, M.; Prasad, G.; Gaur, P.; Talcott, J.B.; Witton, C.; Cassidy, T.; Marshall, B.; Dornan, J., et al. Effects of maternal folic acid supplementation during the second and third trimesters of pregnancy on neurocognitive development in the child: an 11-year follow-up from a randomised controlled trial. BMC Medicine 2021, 19, 73. [CrossRef]
- Valera-Gran, D.; Navarrete-Muñoz, E.M.; Garcia de la Hera, M.; Fernández-Somoano, A.; Tardón, A.; Ibarluzea, J.; Balluerka, N.; Murcia, M.; González-Safont, L.; Romaguera, D., et al. Effect of maternal high dosages of folic acid supplements on neurocognitive development in children at 4-5 y of age: the prospective birth cohort Infancia y Medio Ambiente (INMA) study. Am J Clin Nutr 2017, 106, 878-887. [CrossRef]
- Compañ Gabucio, L.M.; García de la Hera, M.; Torres Collado, L.; Fernández-Somoano, A.; Tardón, A.; Guxens, M.; Vrijheid, M.; Rebagliato, M.; Murcia, M.; Ibarluzea, J., et al. The Use of Lower or Higher Than Recommended Doses of Folic Acid Supplements during Pregnancy Is Associated with Child Attentional Dysfunction at 4–5 Years of Age in the INMA Project. Nutrients 2021, 13, 327.
- Egorova, O.; Myte, R.; Schneede, J.; Hägglöf, B.; Bölte, S.; Domellöf, E.; Ivars A’roch, B.; Elgh, F.; Ueland, P.M.; Silfverdal, S.A. Maternal blood folate status during early pregnancy and occurrence of autism spectrum disorder in offspring: a study of 62 serum biomarkers. Mol Autism 2020, 11, 7. [CrossRef]
- Valera-Gran, D.; García de la Hera, M.; Navarrete-Muñoz, E.M.; Fernandez-Somoano, A.; Tardón, A.; Julvez, J.; Forns, J.; Lertxundi, N.; Ibarluzea, J.M.; Murcia, M., et al. Folic Acid Supplements During Pregnancy and Child Psychomotor Development After the First Year of Life. JAMA Pediatrics 2014, 168, e142611-e142611. [CrossRef]
- Wiens, D.; Desoto, M. Is High Folic Acid Intake a Risk Factor for Autism?—A Review. Brain Sciences 2017, 7, 149. [CrossRef]
- Raghavan, R.; Riley, A.W.; Volk, H.; Caruso, D.; Hironaka, L.; Sices, L.; Hong, X.; Wang, G.; Ji, Y.; Brucato, M., et al. Maternal Multivitamin Intake, Plasma Folate and Vitamin B12 Levels and Autism Spectrum Disorder Risk in Offspring. Paediatric and Perinatal Epidemiology 2018, 32, 100-111. [CrossRef]
- Liu, X.; Zou, M.; Sun, C.; Wu, L.; Chen, W.-X. Prenatal Folic Acid Supplements and Offspring’s Autism Spectrum Disorder: A Meta-analysis and Meta-regression. J Autism Dev Disord 2022, 52, 522-539. [CrossRef]
- Raghavan, R.; Selhub, J.; Paul, L.; Ji, Y.; Wang, G.; Hong, X.; Zuckerman, B.; Fallin, M.D.; Wang, X. A prospective birth cohort study on cord blood folate subtypes and risk of autism spectrum disorder. Am J Clin Nutr 2020, 112, 1304-1317. [CrossRef]
- Yang, X.; Sun, W.; Wu, Q.; Lin, H.; Lu, Z.; Shen, X.; Chen, Y.; Zhou, Y.; Huang, L.; Wu, F., et al. Excess Folic Acid Supplementation before and during Pregnancy and Lactation Alters Behaviors and Brain Gene Expression in Female Mouse Offspring. Nutrients 2021, 14, 66. [CrossRef]
- Harlan De Crescenzo, A.; Panoutsopoulos, A.A.; Tat, L.; Schaaf, Z.; Racherla, S.; Henderson, L.; Leung, K.-Y.; Greene, N.D.E.; Green, R.; Zarbalis, K.S. Deficient or Excess Folic Acid Supply During Pregnancy Alter Cortical Neurodevelopment in Mouse Offspring. Cerebral Cortex 2020, 31, 635-649. [CrossRef]
- Cosín-Tomás, M.; Luan, Y.; Leclerc, D.; Malysheva, O.V.; Lauzon, N.; Bahous, R.H.; Christensen, K.E.; Caudill, M.A.; Rozen, R. Moderate Folic Acid Supplementation in Pregnant Mice Results in Behavioral Alterations in Offspring with Sex-Specific Changes in Methyl Metabolism. Nutrients 2020, 12, 1716.
- Henzel, K.S.; Ryan, D.P.; Schröder, S.; Weiergräber, M.; Ehninger, D. High-dose maternal folic acid supplementation before conception impairs reversal learning in offspring mice. Scientific Reports 2017, 7, 3098. [CrossRef]
- Girotto, F.; Scott, L.; Avchalumov, Y.; Harris, J.; Iannattone, S.; Drummond-Main, C.; Tobias, R.; Bello-Espinosa, L.; Rho, J.M.; Davidsen, J., et al. High dose folic acid supplementation of rats alters synaptic transmission and seizure susceptibility in offspring. Scientific Reports 2013, 3, 1465. [CrossRef]
- Pickell, L.; Brown, K.; Li, D.; Wang, X.L.; Deng, L.; Wu, Q.; Selhub, J.; Luo, L.; Jerome-Majewska, L.; Rozen, R. High intake of folic acid disrupts embryonic development in mice. Birth Defects Res A Clin Mol Teratol 2011, 91, 8-19. [CrossRef]
- Wu, Q.; Zhou, D.; Wu, R.; Shi, R.; Shen, X.; Jin, N.; Gu, J.; Gu, J.H.; Liu, F.; Chu, D. Excess folic acid supplementation before and during pregnancy and lactation activates β-catenin in the brain of male mouse offspring. Brain Res Bull 2022, 178, 133-143. [CrossRef]
- Cianciulli, A.; Salvatore, R.; Porro, C.; Trotta, T.; Panaro, M.A. Folic Acid Is Able to Polarize the Inflammatory Response in LPS Activated Microglia by Regulating Multiple Signaling Pathways. Mediators Inflamm 2016, 2016, 5240127. [CrossRef]
- Lisboa, J.V.C.; Ribeiro, M.R.; Luna, R.C.P.; Lima, R.P.A.; Nascimento, R.; Monteiro, M.; Lima, K.Q.F.; Fechine, C.; Oliveira, N.F.P.; Persuhn, D.C., et al. Food Intervention with Folate Reduces TNF-α and Interleukin Levels in Overweight and Obese Women with the MTHFR C677T Polymorphism: A Randomized Trial. Nutrients 2020, 12. [CrossRef]
- Jones, P.; Lucock, M.; Scarlett, C.J.; Veysey, M.; Beckett, E.L. Folate and Inflammation – links between folate and features of inflammatory conditions. Journal of Nutrition & Intermediary Metabolism 2019, 18, 100104. [CrossRef]
- Pan, Y.; Liu, Y.; Guo, H.; Jabir, M.S.; Liu, X.; Cui, W.; Li, D. Associations between Folate and Vitamin B12 Levels and Inflammatory Bowel Disease: A Meta-Analysis. Nutrients 2017, 9, 382.
- Lazzerini, P.E.; Capecchi, P.L.; Selvi, E.; Lorenzini, S.; Bisogno, S.; Galeazzi, M.; Laghi Pasini, F. Hyperhomocysteinemia, inflammation and autoimmunity. Autoimmunity Reviews 2007, 6, 503-509. [CrossRef]
- Asbaghi, O.; Ashtary-Larky, D.; Bagheri, R.; Moosavian, S.P.; Nazarian, B.; Afrisham, R.; Kelishadi, M.R.; Wong, A.; Dutheil, F.; Suzuki, K., et al. Effects of Folic Acid Supplementation on Inflammatory Markers: A Grade-Assessed Systematic Review and Dose–Response Meta-Analysis of Randomized Controlled Trials. Nutrients 2021, 13, 2327.
- Troen, A.M.; Mitchell, B.; Sorensen, B.; Wener, M.H.; Johnston, A.; Wood, B.; Selhub, J.; McTiernan, A.; Yasui, Y.; Oral, E., et al. Unmetabolized folic acid in plasma is associated with reduced natural killer cell cytotoxicity among postmenopausal women. J Nutr 2006, 136, 189-194. [CrossRef]
- Crider, K.S.; Cordero, A.M.; Qi, Y.P.; Mulinare, J.; Dowling, N.F.; Berry, R.J. Prenatal folic acid and risk of asthma in children: a systematic review and meta-analysis. Am J Clin Nutr 2013, 98, 1272-1281. [CrossRef]
- Whitrow, M.J.; Moore, V.M.; Rumbold, A.R.; Davies, M.J. Effect of supplemental folic acid in pregnancy on childhood asthma: a prospective birth cohort study. Am J Epidemiol 2009, 170, 1486-1493. [CrossRef]
- Zetstra-van der Woude, P.A.; De Walle, H.E.; Hoek, A.; Bos, H.J.; Boezen, H.M.; Koppelman, G.H.; de Jong-van den Berg, L.T.; Scholtens, S. Maternal high-dose folic acid during pregnancy and asthma medication in the offspring. Pharmacoepidemiol Drug Saf 2014, 23, 1059-1065. [CrossRef]
- Veeranki, S.P.; Gebretsadik, T.; Mitchel, E.F.; Tylavsky, F.A.; Hartert, T.V.; Cooper, W.O.; Dupont, W.D.; Dorris, S.L.; Hartman, T.J.; Carroll, K.N. Maternal Folic Acid Supplementation During Pregnancy and Early Childhood Asthma. Epidemiology 2015, 26, 934-941. [CrossRef]
- Yang, L.; Jiang, L.; Bi, M.; Jia, X.; Wang, Y.; He, C.; Yao, Y.; Wang, J.; Wang, Z. High dose of maternal folic acid supplementation is associated to infant asthma. Food Chem Toxicol 2015, 75, 88-93. [CrossRef]
- Chu, S.; Zhang, J. Periconceptional folic acid supplementation is a risk factor for childhood asthma: a case-control study. BMC Pregnancy and Childbirth 2022, 22. [CrossRef]
- Magdelijns, F.J.; Mommers, M.; Penders, J.; Smits, L.; Thijs, C. Folic acid use in pregnancy and the development of atopy, asthma, and lung function in childhood. Pediatrics 2011, 128, e135-144. [CrossRef]
- Martinussen, M.P.; Risnes, K.R.; Jacobsen, G.W.; Bracken, M.B. Folic acid supplementation in early pregnancy and asthma in children aged 6 years. Am J Obstet Gynecol 2012, 206, 72.e71-77. [CrossRef]
- Trivedi, M.K.; Sharma, S.; Rifas-Shiman, S.L.; Camargo, C.A., Jr.; Weiss, S.T.; Oken, E.; Gillman, M.W.; Gold, D.R.; DeMeo, D.L.; Litonjua, A.A. Folic Acid in Pregnancy and Childhood Asthma: A US Cohort. Clin Pediatr (Phila) 2018, 57, 421-427. [CrossRef]
- Vereen, S.; Gebretsadik, T.; Johnson, N.; Hartman, T.J.; Veeranki, S.P.; Piyathilake, C.; Mitchel, E.F.; Kocak, M.; Cooper, W.O.; Dupont, W.D., et al. Association Between Maternal 2nd Trimester Plasma Folate Levels and Infant Bronchiolitis. Maternal and Child Health Journal 2019, 23, 164-172. [CrossRef]
- Veeranki, S.P.; Gebretsadik, T.; Dorris, S.L.; Mitchel, E.F.; Hartert, T.V.; Cooper, W.O.; Tylavsky, F.A.; Dupont, W.; Hartman, T.J.; Carroll, K.N. Association of Folic Acid Supplementation During Pregnancy and Infant Bronchiolitis. American Journal of Epidemiology 2014, 179, 938-946. [CrossRef]
- Håberg, S.E.; London, S.J.; Stigum, H.; Nafstad, P.; Nystad, W. Folic acid supplements in pregnancy and early childhood respiratory health. Archives of Disease in Childhood 2009, 94, 180-184. [CrossRef]
- Roy, A.; Kocak, M.; Hartman, T.J.; Vereen, S.; Adgent, M.; Piyathilake, C.; Tylavsky, F.A.; Carroll, K.N. Association of prenatal folate status with early childhood wheeze and atopic dermatitis. Pediatr Allergy Immunol 2018, 29, 144-150. [CrossRef]
- Chen, Z.; Xing, Y.; Yu, X.; Dou, Y.; Ma, D. Effect of Folic Acid Intake on Infant and Child Allergic Diseases: Systematic Review and Meta-Analysis. Frontiers in Pediatrics 2021, 8. [CrossRef]
- Kiefte-de Jong, J.C.; Timmermans, S.; Jaddoe, V.W.V.; Hofman, A.; Tiemeier, H.; Steegers, E.A.; de Jongste, J.C.; Moll, H.A. High Circulating Folate and Vitamin B-12 Concentrations in Women During Pregnancy Are Associated with Increased Prevalence of Atopic Dermatitis in Their Offspring. The Journal of Nutrition 2012, 142, 731-738. [CrossRef]
- Dunstan, J.A.; West, C.; McCarthy, S.; Metcalfe, J.; Meldrum, S.; Oddy, W.H.; Tulic, M.K.; D’Vaz, N.; Prescott, S.L. The relationship between maternal folate status in pregnancy, cord blood folate levels, and allergic outcomes in early childhood. Allergy 2012, 67, 50-57. [CrossRef]
- Kominsky, D.J.; Keely, S.; MacManus, C.F.; Glover, L.E.; Scully, M.; Collins, C.B.; Bowers, B.E.; Campbell, E.L.; Colgan, S.P. An Endogenously Anti-Inflammatory Role for Methylation in Mucosal Inflammation Identified through Metabolite Profiling. The Journal of Immunology 2011, 186, 6505-6514. [CrossRef]
- Schaible, T.D.; Harris, R.A.; Dowd, S.E.; Smith, C.W.; Kellermayer, R. Maternal methyl-donor supplementation induces prolonged murine offspring colitis susceptibility in association with mucosal epigenetic and microbiomic changes. Human Molecular Genetics 2011, 20, 1687-1696. [CrossRef]
- Mir, S.A.; Nagy-Szakal, D.; Dowd, S.E.; Szigeti, R.G.; Smith, C.W.; Kellermayer, R. Prenatal Methyl-Donor Supplementation Augments Colitis in Young Adult Mice. PLOS ONE 2013, 8, e73162. [CrossRef]
- Pannia, E.; Cho, C.E.; Kubant, R.; Sánchez-Hernández, D.; Huot, P.S.P.; Chatterjee, D.; Fleming, A.; Anderson, G.H. A high multivitamin diet fed to Wistar rat dams during pregnancy increases maternal weight gain later in life and alters homeostatic, hedonic and peripheral regulatory systems of energy balance. Behavioural Brain Research 2015, 278, 1-11. [CrossRef]
- Alnabbat, K.I.; Fardous, A.M.; Cabelof, D.C.; Heydari, A.R. Excessive Folic Acid Mimics Folate Deficiency in Human Lymphocytes. Current Issues in Molecular Biology 2022, 44, 1452-1462.
- Alnabbat, K.I.; Fardous, A.M.; Shahab, A.; James, A.A.; Bahri, M.R.; Heydari, A.R. High Dietary Folic Acid Intake is Associated with Genomic instability in Peripheral Lymphocytes of Healthy Adults. Preprints 2022. [CrossRef]
- Courtemanche, C.; Huang, A.C.; Elson-Schwab, I.; Kerry, N.; Ng, B.Y.; Ames, B.N. Folate deficiency and ionizing radiation cause DNA breaks in primary human lymphocytes: a comparison. FASEB J 2004, 18, 209-211. [CrossRef]
- Mason, J.B.; Dickstein, A.; Jacques, P.F.; Haggarty, P.; Selhub, J.; Dallal, G.; Rosenberg, I.H. A temporal association between folic acid fortification and an increase in colorectal cancer rates may be illuminating important biological principles: a hypothesis. Cancer Epidemiol Biomarkers Prev 2007, 16, 1325-1329. [CrossRef]
- F, W.; K, W.; Y, L.; R, S.; Y, W.; X, Z.; M, S.; L, L.; Sa, S.-W.; El, G., et al. Association of folate intake and colorectal cancer risk in the postfortification era in US women. The American journal of clinical nutrition 2021, 114, 49-58. [CrossRef]
- Ebbing, M.; Bønaa, K.H.; Nygård, O.; Arnesen, E.; Ueland, P.M.; Nordrehaug, J.E.; Rasmussen, K.; Njølstad, I.; Refsum, H.; Nilsen, D.W., et al. Cancer incidence and mortality after treatment with folic acid and vitamin B12. Jama 2009, 302, 2119-2126. [CrossRef]
- Wien, T.N.; Pike, E.; Wisloff, T.; Staff, A.; Smeland, S.; Klemp, M. Cancer risk with folic acid supplements: a systematic review and meta-analysis. BMJ Open 2012, 2, e000653. [CrossRef]
- Oliai Araghi, S.; Kiefte-de Jong, J.C.; van Dijk, S.C.; Swart, K.M.A.; van Laarhoven, H.W.; van Schoor, N.M.; de Groot, L.; Lemmens, V.; Stricker, B.H.; Uitterlinden, A.G., et al. Folic Acid and Vitamin B12 Supplementation and the Risk of Cancer: Long-term Follow-up of the B Vitamins for the Prevention of Osteoporotic Fractures (B-PROOF) Trial. Cancer Epidemiol Biomarkers Prev 2019, 28, 275-282. [CrossRef]
- Cole, B.F.; Baron, J.A.; Sandler, R.S.; Haile, R.W.; Ahnen, D.J.; Bresalier, R.S.; McKeown-Eyssen, G.; Summers, R.W.; Rothstein, R.I.; Burke, C.A., et al. Folic Acid for the Prevention of Colorectal Adenomas. JAMA 2007, 10.1001/jama.297.21.2351. [CrossRef]
- Figueiredo, J.C.; Grau, M.V.; Haile, R.W.; Sandler, R.S.; Summers, R.W.; Bresalier, R.S.; Burke, C.A.; McKeown-Eyssen, G.E.; Baron, J.A. Folic acid and risk of prostate cancer: Results from a randomized clinical trial. Journal of the National Cancer Institute 2009, 10.1093/jnci/djp019. [CrossRef]
- Liss, M.A.; Ashcraft, K.; Satsangi, A.; Bacich, D. Rise in serum folate after androgen deprivation associated with worse prostate cancer-specific survival. Urol Oncol 2020, 38, 682.e621-682.e627. [CrossRef]
- Fu, H.; He, J.; Li, C.; Deng, Z.; Chang, H. Folate intake and risk of colorectal cancer: a systematic review and up-to-date meta-analysis of prospective studies. Eur J Cancer Prev 2023, 32, 103-112. [CrossRef]
- Pieroth, R.; Paver, S.; Day, S.; Lammersfeld, C. Folate and Its Impact on Cancer Risk. Current Nutrition Reports 2018, 7, 70-84. [CrossRef]
- Song, J.; Sohn, K.J.; Medline, A.; Ash, C.; Gallinger, S.; Kim, Y.I. Chemopreventive effects of dietary folate on intestinal polyps in Apc+/- Msh2-/- Mice. Cancer Research 2000.
- Le Leu, R.K.; Young, G.P.; McIntosh, G.H. Folate deficiency reduces the development of colorectal cancer in rats. Carcinogenesis 2000, 21, 2261-2265. [CrossRef]
- Lindzon, G.M.; Medline, A.; Sohn, K.J.; Depeint, F.; Croxford, R.; Kim, Y.I. Effect of folic acid supplementation on the progression of colorectal aberrant crypt foci. Carcinogenesis 2009, 30, 1536-1543. [CrossRef]
- Galeone, C.; Edefonti, V.; Parpinel, M.; Leoncini, E.; Matsuo, K.; Talamini, R.; Olshan, A.F.; Zevallos, J.P.; Winn, D.M.; Jayaprakash, V., et al. Folate intake and the risk of oral cavity and pharyngeal cancer: a pooled analysis within the International Head and Neck Cancer Epidemiology Consortium. Int J Cancer 2015, 136, 904-914. [CrossRef]
- Tio, M.; Andrici, J.; Cox, M.R.; Eslick, G.D. Folate intake and the risk of upper gastrointestinal cancers: a systematic review and meta-analysis. J Gastroenterol Hepatol 2014, 29, 250-258. [CrossRef]
- Fu, H.; Zeng, J.; Liu, C.; Gu, Y.; Zou, Y.; Chang, H. Folate Intake and Risk of Pancreatic Cancer: A Systematic Review and Updated Meta-Analysis of Epidemiological Studies. Dig Dis Sci 2021, 66, 2368-2379. [CrossRef]
- Gu, Y.; Zeng, J.; Zou, Y.; Liu, C.; Fu, H.; Chang, H. Folate Intake and Risk of Urothelial Carcinoma: A Systematic Review and Meta-Analysis of Epidemiological Studies. Nutr Cancer 2022, 74, 1593-1605. [CrossRef]
- He, H.; Shui, B. Folate intake and risk of bladder cancer: a meta-analysis of epidemiological studies. Int J Food Sci Nutr 2014, 65, 286-292. [CrossRef]
- Zhang, Y.F.; Zhou, L.; Zhang, H.W.; Hou, A.J.; Gao, H.F.; Zhou, Y.H. Association between folate intake and the risk of lung cancer: a dose-response meta-analysis of prospective studies. PLoS One 2014, 9, e93465. [CrossRef]
- Stanisławska-Sachadyn, A.; Borzyszkowska, J.; Krzemiński, M.; Janowicz, A.; Dziadziuszko, R.; Jassem, J.; Rzyman, W.; Limon, J. Folate/homocysteine metabolism and lung cancer risk among smokers. PLOS ONE 2019, 14, e0214462. [CrossRef]
- Li, C.; Chen, P.; Hu, P.; Li, M.; Li, X.; Guo, H.; Li, J.; Chu, R.; Zhang, W.; Wang, H. Folate intake and MTHFR polymorphism C677T is not associated with ovarian cancer risk: evidence from the meta-analysis. Mol Biol Rep 2013, 40, 6547-6560. [CrossRef]
- Du, L.; Wang, Y.; Zhang, H.; Zhang, H.; Gao, Y. Folate intake and the risk of endometrial cancer: A meta-analysis. Oncotarget 2016, 7, 85176-85184. [CrossRef]
- Zhang, Y.F.; Shi, W.W.; Gao, H.F.; Zhou, L.; Hou, A.J.; Zhou, Y.H. Folate intake and the risk of breast cancer: a dose-response meta-analysis of prospective studies. PLoS One 2014, 9, e100044. [CrossRef]
- Chen, P.; Li, C.; Li, X.; Li, J.; Chu, R.; Wang, H. Higher dietary folate intake reduces the breast cancer risk: a systematic review and meta-analysis. Br J Cancer 2014, 110, 2327-2338. [CrossRef]
- Zeng, J.; Gu, Y.; Fu, H.; Liu, C.; Zou, Y.; Chang, H. Association Between One-carbon Metabolism-related Vitamins and Risk of Breast Cancer: A Systematic Review and Meta-analysis of Prospective Studies. Clin Breast Cancer 2020, 20, e469-e480. [CrossRef]
- Zeng, J.; Wang, K.; Ye, F.; Lei, L.; Zhou, Y.; Chen, J.; Zhao, G.; Chang, H. Folate intake and the risk of breast cancer: an up-to-date meta-analysis of prospective studies. Eur J Clin Nutr 2019, 73, 1657-1660. [CrossRef]
- Rosati, R.; Ma, H.; Cabelof, D.C. Folate and colorectal cancer in rodents: A model of DNA repair deficiency. 2012; 10.1155/2012/105949.
- Hansen, M.F.; Jensen, S.Ø.; Füchtbauer, E.-M.; Martensen, P.M. High folic acid diet enhances tumour growth in PyMT-induced breast cancer. British Journal of Cancer 2017, 116, 752-761. [CrossRef]
- Ly, A.; Lee, H.; Chen, J.; Sie, K.K.Y.; Renlund, R.; Medline, A.; Sohn, K.J.; Croxford, R.; Thompson, L.U.; Kim, Y.I. Effect of maternal and postweaning folic acid supplementation on mammary tumor risk in the offspring. Cancer Research 2011, 71, 988-997. [CrossRef]
- Deghan Manshadi, S.; Ishiguro, L.; Sohn, K.-J.; Medline, A.; Renlund, R.; Croxford, R.; Kim, Y.-I. Folic Acid Supplementation Promotes Mammary Tumor Progression in a Rat Model. PLOS ONE 2014, 9, e84635. [CrossRef]
- Been, R.A.; Ross, J.A.; Nagel, C.W.; Hooten, A.J.; Langer, E.K.; DeCoursin, K.J.; Marek, C.A.; Janik, C.L.; Linden, M.A.; Reed, R.C., et al. Perigestational dietary folic acid deficiency protects against medulloblastoma formation in a mouse model of nevoid basal cell carcinoma syndrome. Nutr Cancer 2013, 65, 857-865. [CrossRef]
- Rycyna, K.J.; Bacich, D.J.; O’Keefe, D.S. Opposing roles of folate in prostate cancer. Urology 2013, 82, 1197-1203. [CrossRef]
- Savini, C.; Yang, R.; Savelyeva, L.; Göckel-Krzikalla, E.; Hotz-Wagenblatt, A.; Westermann, F.; Rösl, F. Folate Repletion after Deficiency Induces Irreversible Genomic and Transcriptional Changes in Human Papillomavirus Type 16 (HPV16)-Immortalized Human Keratinocytes. Int J Mol Sci 2019, 20. [CrossRef]
- Xiao, S.; Tang, Y.S.; Kusumanchi, P.; Stabler, S.P.; Zhang, Y.; Antony, A.C. Folate Deficiency Facilitates Genomic Integration of Human Papillomavirus Type 16 DNA In Vivo in a Novel Mouse Model for Rapid Oncogenic Transformation of Human Keratinocytes. J Nutr 2018, 148, 389-400. [CrossRef]
- Sharma, R.; Ali, T.; Kaur, J. Folic acid depletion as well as oversupplementation helps in the progression of hepatocarcinogenesis in HepG2 cells. Sci Rep 2022, 12, 16617. [CrossRef]
- Zhang, B.; Dong, H.; Xu, Y.; Xu, D.; Sun, H.; Han, L. Associations of dietary folate, vitamin B6 and B12 intake with cardiovascular outcomes in 115664 participants: a large UK population-based cohort. Eur J Clin Nutr 2022, 10.1038/s41430-022-01206-2. [CrossRef]
- Bo, Y.; Xu, H.; Zhang, H.; Zhang, J.; Wan, Z.; Zhao, X.; Yu, Z. Intakes of Folate, Vitamin B6, and Vitamin B12 in Relation to All-Cause and Cause-Specific Mortality: A National Population-Based Cohort. Nutrients 2022, 14. [CrossRef]
- Li, Y.; Huang, T.; Zheng, Y.; Muka, T.; Troup, J.; Hu, F.B. Folic Acid Supplementation and the Risk of Cardiovascular Diseases: A Meta-Analysis of Randomized Controlled Trials. J Am Heart Assoc 2016, 5. [CrossRef]
- Bønaa, K.H.; Njølstad, I.; Ueland, P.M.; Schirmer, H.; Tverdal, A.; Steigen, T.; Wang, H.; Nordrehaug, J.E.; Arnesen, E.; Rasmussen, K. Homocysteine Lowering and Cardiovascular Events after Acute Myocardial Infarction. New England Journal of Medicine 2006, 354, 1578-1588. [CrossRef]
- Liu, M.; Zhang, Z.; Zhou, C.; Li, Q.; He, P.; Zhang, Y.; Li, H.; Liu, C.; Liang, M.; Wang, X., et al. Relationship of several serum folate forms with the risk of mortality: A prospective cohort study. Clin Nutr 2021, 40, 4255-4262. [CrossRef]
- Peng, Y.; Dong, B.; Wang, Z. Serum folate concentrations and all-cause, cardiovascular disease and cancer mortality: A cohort study based on 1999–2010 National Health and Nutrition Examination Survey (NHANES). International Journal of Cardiology 2016, 219, 136-142. [CrossRef]
- Zhao, J.V.; Schooling, C.M.; Zhao, J.X. The effects of folate supplementation on glucose metabolism and risk of type 2 diabetes: a systematic review and meta-analysis of randomized controlled trials. Ann Epidemiol 2018, 28, 249-257.e241. [CrossRef]
- Yang, H.T.; Lee, M.; Hong, K.S.; Ovbiagele, B.; Saver, J.L. Efficacy of folic acid supplementation in cardiovascular disease prevention: an updated meta-analysis of randomized controlled trials. Eur J Intern Med 2012, 23, 745-754. [CrossRef]
- Pannia, E.; Hammoud, R.; Simonian, R.; Arning, E.; Ashcraft, P.; Wasek, B.; Bottiglieri, T.; Pausova, Z.; Kubant, R.; Anderson, G.H. [6S]-5-Methyltetrahydrofolic Acid and Folic Acid Pregnancy Diets Differentially Program Metabolic Phenotype and Hypothalamic Gene Expression of Wistar Rat Dams Post-Birth. Nutrients 2020, 13, 48. [CrossRef]
- Dose-dependent effects of folic acid on blood concentrations of homocysteine: a meta-analysis of the randomized trials. Am J Clin Nutr 2005, 82, 806-812. [CrossRef]
- Chen, Y.; Liu, R.; Zhang, G.; Yu, Q.; Jia, M.; Zheng, C.; Wang, Y.; Xu, C.; Zhang, Y.; Liu, E. Hypercysteinemia promotes atherosclerosis by reducing protein S-nitrosylation. Biomed Pharmacother 2015, 70, 253-259. [CrossRef]
- Kalita, J.; Kumar, G.; Bansal, V.; Misra, U.K. Relationship of homocysteine with other risk factors and outcome of ischemic stroke. Clin Neurol Neurosurg 2009, 111, 364-367. [CrossRef]
- Bonetti, F.; Brombo, G.; Magon, S.; Zuliani, G. Cognitive Status According to Homocysteine and B-Group Vitamins in Elderly Adults. J Am Geriatr Soc 2015, 63, 1158-1163. [CrossRef]
- Yajnik, C.S.; Chandak, G.R.; Joglekar, C.; Katre, P.; Bhat, D.S.; Singh, S.N.; Janipalli, C.S.; Refsum, H.; Krishnaveni, G.; Veena, S., et al. Maternal homocysteine in pregnancy and offspring birthweight: epidemiological associations and Mendelian randomization analysis. Int J Epidemiol 2014, 43, 1487-1497. [CrossRef]
- den Heijer, M.; Brouwer, I.A.; Bos, G.M.; Blom, H.J.; van der Put, N.M.; Spaans, A.P.; Rosendaal, F.R.; Thomas, C.M.; Haak, H.L.; Wijermans, P.W., et al. Vitamin supplementation reduces blood homocysteine levels: a controlled trial in patients with venous thrombosis and healthy volunteers. Arterioscler Thromb Vasc Biol 1998, 18, 356-361. [CrossRef]
- van der Griend, R.; Biesma, D.H.; Haas, F.J.; Faber, J.A.; Duran, M.; Meuwissen, O.J.; Banga, J.D. The effect of different treatment regimens in reducing fasting and postmethionine-load homocysteine concentrations. J Intern Med 2000, 248, 223-229. [CrossRef]
- Tighe, P.; Ward, M.; McNulty, H.; Finnegan, O.; Dunne, A.; Strain, J.; Molloy, A.M.; Duffy, M.; Pentieva, K.; Scott, J.M. A dose-finding trial of the effect of long-term folic acid intervention: implications for food fortification policy. Am J Clin Nutr 2011, 93, 11-18. [CrossRef]
- Tian, H.; Tian, D.; Zhang, C.; Wang, W.; Wang, L.; Ge, M.; Hou, Q.; Zhang, W. Efficacy of Folic Acid Therapy in Patients with Hyperhomocysteinemia. Journal of the American College of Nutrition 2017, 36, 528-532. [CrossRef]
- Reynolds, E.H. What is the safe upper intake level of folic acid for the nervous system? Implications for folic acid fortification policies. European Journal of Clinical Nutrition 2016, 70, 537-540. [CrossRef]
- Bailey, R.L.; Jun, S.; Murphy, L.; Green, R.; Gahche, J.J.; Dwyer, J.T.; Potischman, N.; McCabe, G.P.; Miller, J.W. High folic acid or folate combined with low vitamin B-12 status: potential but inconsistent association with cognitive function in a nationally representative cross-sectional sample of US older adults participating in the NHANES. The American Journal of Clinical Nutrition 2020, 112, 1547-1557. [CrossRef]
- Paul, L.; Selhub, J. Interaction between excess folate and low vitamin B12 status. Mol Aspects Med 2017, 53, 43-47. [CrossRef]
- Will, J.J.; Mueller, J.F.; Brodine, C.; Kiely, C.E.; Friedman, B.; Hawkins, V.R.; Dutra, J.; Vilter, R.W. Folic acid and vitamin B12 in pernicious anemia; studies on patients treated with these substances over a ten year period. J Lab Clin Med 1959, 53, 22-38.
- Lear, A.A.; Castle, W.B. Supplemental folic acid therapy in pernicious anemia: the effect on erythropoiesis and serum vitamin B12 concentrations in selected cases. J Lab Clin Med 1956, 47, 88-97.
- Bok, J.; Faber, J.G.; De Vries, J.A.; Kroese, W.F.; Nieweg, H.O. The effect of pteroylglutamic acid administration on the serum vitamin B12 concentration in pernicious anemia in relapse. J Lab Clin Med 1958, 51, 667-671.
- Moore, E.M.; Ames, D.; Mander, A.G.; Carne, R.P.; Brodaty, H.; Woodward, M.C.; Boundy, K.; Ellis, K.A.; Bush, A.I.; Faux, N.G., et al. Among vitamin B12 deficient older people, high folate levels are associated with worse cognitive function: combined data from three cohorts. J Alzheimers Dis 2014, 39, 661-668. [CrossRef]
- Morris, M.S.; Selhub, J.; Jacques, P.F. Vitamin B-12 and folate status in relation to decline in scores on the mini-mental state examination in the framingham heart study. J Am Geriatr Soc 2012, 60, 1457-1464. [CrossRef]
- Morris, M.S.; Jacques, P.F.; Rosenberg, I.H.; Selhub, J. Folate and vitamin B-12 status in relation to anemia, macrocytosis, and cognitive impairment in older Americans in the age of folic acid fortification. Am J Clin Nutr 2007, 85, 193-200. [CrossRef]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Hebert, L.E.; Scherr, P.A.; Schneider, J.A. Dietary folate and vitamin B12 intake and cognitive decline among community-dwelling older persons. Arch Neurol 2005, 62, 641-645. [CrossRef]
- van Gool, J.D.; Hirche, H.; Lax, H.; Schaepdrijver, L. Fallacies of clinical studies on folic acid hazards in subjects with a low vitamin B(12) status. Crit Rev Toxicol 2020, 50, 177-187. [CrossRef]
- Selhub, J.; Miller, J.W.; Troen, A.M.; Mason, J.B.; Jacques, P.F. Perspective: The High-Folate-Low-Vitamin B-12 Interaction Is a Novel Cause of Vitamin B-12 Depletion with a Specific Etiology-A Hypothesis. Adv Nutr 2022, 13, 16-33. [CrossRef]
- Maher, A.; Sobczyńska-Malefora, A. The Relationship Between Folate, Vitamin B12 and Gestational Diabetes Mellitus With Proposed Mechanisms and Foetal Implications. J Family Reprod Health 2021, 15, 141-149. [CrossRef]
- Roy, S.; Kale, A.; Dangat, K.; Sable, P.; Kulkarni, A.; Joshi, S. Maternal micronutrients (folic acid and vitamin B12) and omega 3 fatty acids: Implications for neurodevelopmental risk in the rat offspring. Brain and Development 2012, 34, 64-71. [CrossRef]
- Henderson, A.M.; Tai, D.C.; Aleliunas, R.E.; Aljaadi, A.M.; Glier, M.B.; Xu, E.E.; Miller, J.W.; Verchere, C.B.; Green, T.J.; Devlin, A.M. Maternal folic acid supplementation with vitamin B(12) deficiency during pregnancy and lactation affects the metabolic health of adult female offspring but is dependent on offspring diet. Faseb j 2018, 32, 5039-5050. [CrossRef]
- Kulkarni, A.; Dangat, K.; Kale, A.; Sable, P.; Chavan-Gautam, P.; Joshi, S. Effects of Altered Maternal Folic Acid, Vitamin B12 and Docosahexaenoic Acid on Placental Global DNA Methylation Patterns in Wistar Rats. PLOS ONE 2011, 6, e17706. [CrossRef]
- Sable, P.; Dangat, K.; Kale, A.; Joshi, S. Altered brain neurotrophins at birth: consequence of imbalance in maternal folic acid and vitamin B12 metabolism. Neuroscience 2011, 190, 127-134. [CrossRef]
- Christensen, K.E.; Mikael, L.G.; Leung, K.Y.; Levesque, N.; Deng, L.; Wu, Q.; Malysheva, O.V.; Best, A.; Caudill, M.A.; Greene, N.D., et al. High folic acid consumption leads to pseudo-MTHFR deficiency, altered lipid metabolism, and liver injury in mice. Am J Clin Nutr 2015, 101, 646-658. [CrossRef]
- Bahous, R.H.; Jadavji, N.M.; Deng, L.; Cosin-Tomas, M.; Lu, J.; Malysheva, O.; Leung, K.Y.; Ho, M.K.; Pallas, M.; Kaliman, P., et al. High dietary folate in pregnant mice leads to pseudo-MTHFR deficiency and altered methyl metabolism, with embryonic growth delay and short-term memory impairment in offspring. Hum Mol Genet 2017, 26, 888-900. [CrossRef]
- Cornet, D.; Clement, A.; Clement, P.; Menezo, Y. High doses of folic acid induce a pseudo-methylenetetrahydrofolate syndrome. SAGE Open Med Case Rep 2019, 7, 2050313x19850435. [CrossRef]
- Smith, D.E.C.; Hornstra, J.M.; Kok, R.M.; Blom, H.J.; Smulders, Y.M. Folic acid supplementation does not reduce intracellular homocysteine, and may disturb intracellular one-carbon metabolism. Clinical Chemistry and Laboratory Medicine (CCLM) 2013, 51, 1643-1650. [CrossRef]
- Henry, C.J.; Nemkov, T.; Casas-Selves, M.; Bilousova, G.; Zaberezhnyy, V.; Higa, K.C.; Serkova, N.J.; Hansen, K.C.; D’Alessandro, A.; DeGregori, J. Folate dietary insufficiency and folic acid supplementation similarly impair metabolism and compromise hematopoiesis. Haematologica 2017, 102, 1985-1994. [CrossRef]
- Han, X.; Wang, B.; Jin, D.; Liu, K.; Wang, H.; Chen, L.; Zu, Y. Precise Dose of Folic Acid Supplementation Is Essential for Embryonic Heart Development in Zebrafish. Biology 2021, 11, 28. [CrossRef]
- Barua, S.; Kuizon, S.; Ted Brown, W.; Junaid, M.A. High Gestational Folic Acid Supplementation Alters Expression of Imprinted and Candidate Autism Susceptibility Genes in a sex-Specific Manner in Mouse Offspring. Journal of Molecular Neuroscience 2016, 58, 277-286. [CrossRef]
- Barua, S.; Kuizon, S.; Chadman, K.K.; Flory, M.J.; Brown, W.T.; Junaid, M.A. Single-base resolution of mouse offspring brain methylome reveals epigenome modifications caused by gestational folic acid. Epigenetics and Chromatin 2014, 10.1186/1756-8935-7-3. [CrossRef]
- Luan, Y.; Cosín-Tomás, M.; Leclerc, D.; Malysheva, O.V.; Caudill, M.A.; Rozen, R. Moderate Folic Acid Supplementation in Pregnant Mice Results in Altered Sex-Specific Gene Expression in Brain of Young Mice and Embryos. Nutrients 2022, 14. [CrossRef]
- Bailey, S.W.; Ayling, J.E. The extremely slow and variable activity of dihydrofolate reductase in human liver and its implications for high folic acid intake. Proc Natl Acad Sci U S A 2009, 106, 15424-15429. [CrossRef]
- Wu, S.; Guo, W.; Li, X.; Liu, Y.; Li, Y.; Lei, X.; Yao, J.; Yang, X. Paternal chronic folate supplementation induced the transgenerational inheritance of acquired developmental and metabolic changes in chickens. Proceedings of the Royal Society B: Biological Sciences 2019, 286, 20191653. [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. Cell Metabolism Review One-Carbon Metabolism in Health and Disease. Cell Metabolism 2017, 10.1016/j.cmet.2016.08.009. [CrossRef]
- Moran, R.G. Roles of folylpoly-gamma-glutamate synthetase in therapeutics with tetrahydrofolate antimetabolites: an overview. Semin Oncol 1999, 26, 24-32.
- Mohanty, V.; Shah, A.; Allender, E.; Siddiqui, M.R.; Monick, S.; Ichi, S.; Mania-Farnell, B.; G. McLone, D.; Tomita, T.; Mayanil, C.S. Folate Receptor Alpha Upregulates Oct4, Sox2 and Klf4 and Downregulates miR-138 and miR-let-7 in Cranial Neural Crest Cells. STEM CELLS 2016, 34, 2721-2732. [CrossRef]
- Boshnjaku, V.; Shim, K.W.; Tsurubuchi, T.; Ichi, S.; Szany, E.V.; Xi, G.; Mania-Farnell, B.; McLone, D.G.; Tomita, T.; Mayanil, C.S. Nuclear localization of folate receptor alpha: a new role as a transcription factor. Sci Rep 2012, 2, 980. [CrossRef]
- Mohanty, V.; Siddiqui, M.R.; Tomita, T.; Mayanil, C.S. Folate receptor alpha is more than just a folate transporter. Neurogenesis (Austin) 2017, 4, e1263717. [CrossRef]
- Frigerio, B.; Bizzoni, C.; Jansen, G.; Leamon, C.P.; Peters, G.J.; Low, P.S.; Matherly, L.H.; Figini, M. Folate receptors and transporters: biological role and diagnostic/therapeutic targets in cancer and other diseases. J Exp Clin Cancer Res 2019, 38, 125. [CrossRef]
- Hansen, M.F.; Greibe, E.; Skovbjerg, S.; Rohde, S.; Kristensen, A.C.; Jensen, T.R.; Stentoft, C.; Kjær, K.H.; Kronborg, C.S.; Martensen, P.M. Folic acid mediates activation of the pro-oncogene STAT3 via the Folate Receptor alpha. Cell Signal 2015, 27, 1356-1368. [CrossRef]
- Zhang, X.M.; Huang, G.W.; Tian, Z.H.; Ren, D.L.; Wilson, J.X. Folate stimulates ERK1/2 phosphorylation and cell proliferation in fetal neural stem cells. Nutr Neurosci 2009, 12, 226-232. [CrossRef]
- Kuo, C.T.; Chang, C.; Lee, W.S. Folic acid inhibits COLO-205 colon cancer cell proliferation through activating the FRα/c-SRC/ERK1/2/NFκB/TP53 pathway: in vitro and in vivo studies. Sci Rep 2015, 5, 11187. [CrossRef]
- Qiu, W.; Gobinath, A.R.; Wen, Y.; Austin, J.; Galea, L.A.M. Folic acid, but not folate, regulates different stages of neurogenesis in the ventral hippocampus of adult female rats. J Neuroendocrinol 2019, 31, e12787. [CrossRef]
- Taylor, S.Y.; Dixon, H.M.; Yoganayagam, S.; Price, N.; Lang, D. Folic acid modulates eNOS activity via effects on posttranslational modifications and protein-protein interactions. Eur J Pharmacol 2013, 714, 193-201. [CrossRef]
- Köbel, M.; Madore, J.; Ramus, S.J.; Clarke, B.A.; Pharoah, P.D.; Deen, S.; Bowtell, D.D.; Odunsi, K.; Menon, U.; Morrison, C., et al. Evidence for a time-dependent association between FOLR1 expression and survival from ovarian carcinoma: implications for clinical testing. An Ovarian Tumour Tissue Analysis consortium study. Br J Cancer 2014, 111, 2297-2307. [CrossRef]
- Liu, J.; Yao, Y.; Yu, B.; Mao, X.; Huang, Z.; Chen, D. Effect of maternal folic acid supplementation on hepatic proteome in newborn piglets. Nutrition 2013, 29, 230-234. [CrossRef]
- Pannia, E.; Hammoud, R.; Kubant, R.; Sa, J.Y.; Simonian, R.; Wasek, B.; Ashcraft, P.; Bottiglieri, T.; Pausova, Z.; Anderson, G.H. High Intakes of [6S]-5-Methyltetrahydrofolic Acid Compared with Folic Acid during Pregnancy Programs Central and Peripheral Mechanisms Favouring Increased Food Intake and Body Weight of Mature Female Offspring. Nutrients 2021, 13. [CrossRef]
- Palmer, A.M.; Kamynina, E.; Field, M.S.; Stover, P.J. Folate rescues vitamin B(12) depletion-induced inhibition of nuclear thymidylate biosynthesis and genome instability. Proc Natl Acad Sci U S A 2017, 114, E4095-e4102. [CrossRef]
- Jennings, B.A.; Willis, G. How folate metabolism affects colorectal cancer development and treatment; a story of heterogeneity and pleiotropy. Cancer Letters 2015, 356, 224-230. [CrossRef]
- Pufulete, M.; Emery, P.W.; Sanders, T.A.B. Folate, DNA methylation and colo-rectal cancer. Proceedings of the Nutrition Society 2003, 10.1079/pns2003265. [CrossRef]
- Hanley, M.P.; Aladelokun, O.; Kadaveru, K.; Rosenberg, D.W. Methyl donor deficiency blocks colorectal cancer development by affecting key metabolic pathways. Cancer Prevention Research 2020, 10.1158/1940-6207.CAPR-19-0188. [CrossRef]
- Ryan, B.M.; Weir, D.G. Relevance of folate metabolism in the pathogenesis of colorectal cancer. Journal of Laboratory and Clinical Medicine 2001, 138, 164-176. [CrossRef]
- Charles, M.A.; Johnson, I.T.; Belshaw, N.J. Supra-physiological folic acid concentrations induce aberrant DNA methylation in normal human cells in vitro. Epigenetics 2012, 7, 689-694. [CrossRef]
- Ortbauer, M.; Ripper, D.; Fuhrmann, T.; Lassi, M.; Auernigg-Haselmaier, S.; Stiegler, C.; Konig, J. Folate deficiency and over-supplementation causes impaired folate metabolism: Regulation and adaptation mechanisms in Caenorhabditis elegans. Mol Nutr Food Res 2016, 60, 949-956. [CrossRef]
- Han, X.; Wang, B.; Jin, D.; Liu, K.; Wang, H.; Chen, L.; Zu, Y. Precise Dose of Folic Acid Supplementation Is Essential for Embryonic Heart Development in Zebrafish. Biology (Basel) 2021, 11. [CrossRef]
- Zgheib, R.; Battaglia-Hsu, S.F.; Hergalant, S.; Quéré, M.; Alberto, J.M.; Chéry, C.; Rouyer, P.; Gauchotte, G.; Guéant, J.L.; Namour, F. Folate can promote the methionine-dependent reprogramming of glioblastoma cells towards pluripotency. Cell Death Dis 2019, 10, 596. [CrossRef]
- Kaittanis, C.; Andreou, C.; Hieronymus, H.; Mao, N.; Foss, C.A.; Eiber, M.; Weirich, G.; Panchal, P.; Gopalan, A.; Zurita, J., et al. Prostate-specific membrane antigen cleavage of vitamin B9 stimulates oncogenic signaling through metabotropic glutamate receptors. J Exp Med 2018, 215, 159-175. [CrossRef]
- Murray, L.K.; Smith, M.J.; Jadavji, N.M. Maternal oversupplementation with folic acid and its impact on neurodevelopment of offspring. Nutrition Reviews 2018, 76, 708-721. [CrossRef]
- Ondičová, M.; Irwin, R.E.; Thursby, S.J.; Hilman, L.; Caffrey, A.; Cassidy, T.; McLaughlin, M.; Lees-Murdock, D.J.; Ward, M.; Murphy, M., et al. Folic acid intervention during pregnancy alters DNA methylation, affecting neural target genes through two distinct mechanisms. Clin Epigenetics 2022, 14, 63. [CrossRef]
- Richmond, R.C.; Sharp, G.C.; Herbert, G.; Atkinson, C.; Taylor, C.; Bhattacharya, S.; Campbell, D.; Hall, M.; Kazmi, N.; Gaunt, T., et al. The long-term impact of folic acid in pregnancy on offspring DNA methylation: follow-up of the Aberdeen Folic Acid Supplementation Trial (AFAST). Int J Epidemiol 2018, 47, 928-937. [CrossRef]
- Li, Z.; Gueant-Rodriguez, R.M.; Quilliot, D.; Sirveaux, M.A.; Meyre, D.; Gueant, J.L.; Brunaud, L. Folate and vitamin B12 status is associated with insulin resistance and metabolic syndrome in morbid obesity. Clin Nutr 2018, 37, 1700-1706. [CrossRef]
- Yajnik, C.S.; Deshpande, S.S.; Jackson, A.A.; Refsum, H.; Rao, S.; Fisher, D.J.; Bhat, D.S.; Naik, S.S.; Coyaji, K.J.; Joglekar, C.V., et al. Vitamin B12 and folate concentrations during pregnancy and insulin resistance in the offspring: the Pune Maternal Nutrition Study. Diabetologia 2008, 51, 29-38. [CrossRef]
- Paniz, C.; Bertinato, J.F.; Lucena, M.R.; De Carli, E.; Amorim, P.M.D.S.; Gomes, G.W.; Palchetti, C.Z.; Figueiredo, M.S.; Pfeiffer, C.M.; Fazili, Z., et al. A Daily Dose of 5 mg Folic Acid for 90 Days Is Associated with Increased Serum Unmetabolized Folic Acid and Reduced Natural Killer Cell Cytotoxicity in Healthy Brazilian Adults. The Journal of Nutrition 2017, 147, 1677-1685. [CrossRef]
- Sawaengsri, H.; Wang, J.; Reginaldo, C.; Steluti, J.; Wu, D.; Meydani, S.N.; Selhub, J.; Paul, L. High folic acid intake reduces natural killer cell cytotoxicity in aged mice. J Nutr Biochem 2016, 30, 102-107. [CrossRef]
- Kadaveru, K.; Protiva, P.; Greenspan, E.J.; Kim, Y.I.; Rosenberg, D.W.; K, K.; P, P.; Ej, G.; Yi, K.; Dw, R., et al. Dietary methyl donor depletion protects against intestinal tumorigenesis in Apc(Min/+) mice. Cancer Prevention Research 2012, 5, 911-920. [CrossRef]
- Protiva, P.; Mason, J.B.; Liu, Z.; Hopkins, M.E.; Nelson, C.; Marshall, J.R.; Lambrecht, R.W.; Pendyala, S.; Kopelovich, L.; Kim, M., et al. Altered folate availability modifies the molecular environment of the human colorectum: implications for colorectal carcinogenesis. Cancer prevention research (Philadelphia, Pa.) 2011, 4, 530-543. [CrossRef]
- Kelly, K.B.; Kennelly, J.P.; Ordonez, M.; Nelson, R.; Leonard, K.; Stabler, S.; Gomez-Munoz, A.; Field, C.J.; Jacobs, R.L. Excess Folic Acid Increases Lipid Storage, Weight Gain, and Adipose Tissue Inflammation in High Fat Diet-Fed Rats. Nutrients 2016, 8. [CrossRef]
- Annibal, A.; Tharyan, R.G.; Schonewolff, M.F.; Tam, H.; Latza, C.; Auler, M.M.K.; Antebi, A. Regulation of the one carbon folate cycle as a shared metabolic signature of longevity. Nature Communications 2021, 12, 1-14. [CrossRef]
- Liu, Y.J.; Janssens, G.E.; McIntyre, R.L.; Molenaars, M.; Kamble, R.; Gao, A.W.; Jongejan, A.; Weeghel, M.V.; MacInnes, A.W.; Houtkooper, R.H. Glycine promotes longevity in Caenorhabditis elegans in a methionine cycle-dependent fashion. PLoS Genet 2019, 15, e1007633. [CrossRef]
Figure 2.
Mechanisms of excess folate.
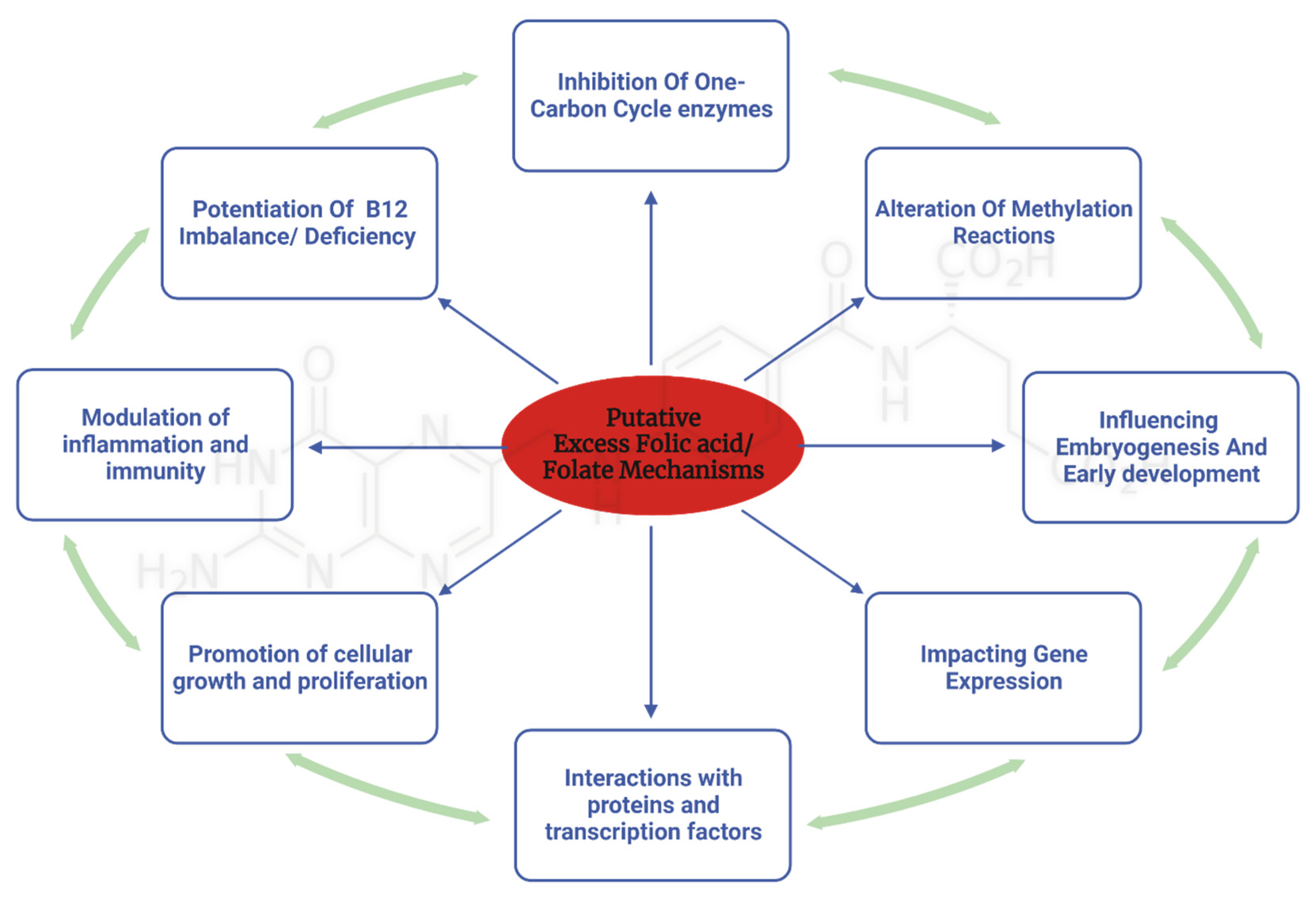
Figure 3.
molecular effects of excess folate on one carbon metabolism.
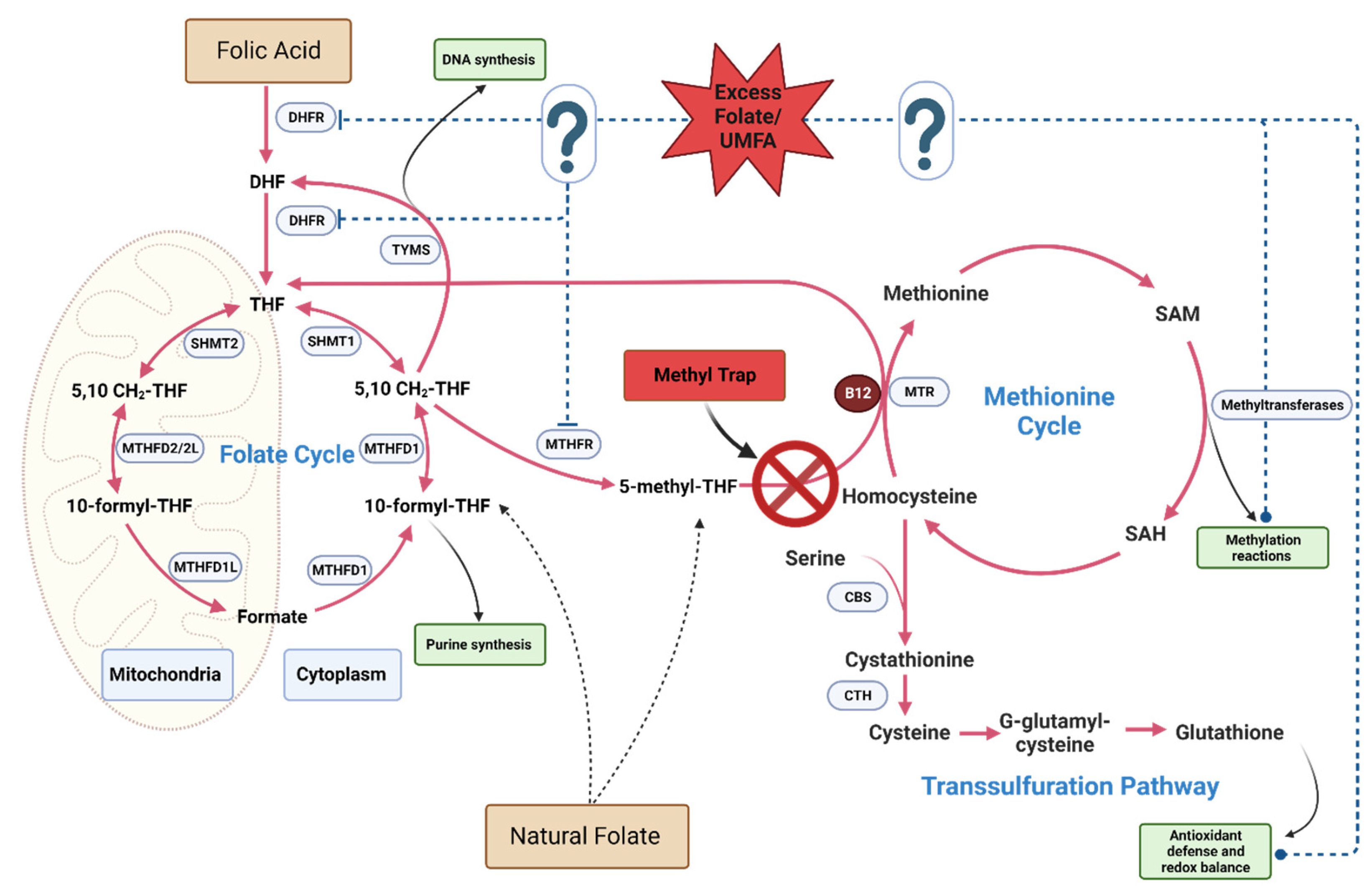
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



