
Submitted:
25 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
The effect of high temperature (HT) stress on nicotine biosynthesis in Nicotiana attenuata was examined. Nicotine content was measured in mature leaves, young sink leaves and in roots from well-watered plants grown at 25 °C as controls and from plants exposed to 38 °C and 43 °C temperatures applied for 24, 48, 72 and 96-hr duration. At 38 °C, all leaf nicotine levels were significantly less than control plants for up to 72-hr exposure but rose sharply thereafter to levels significantly greater than controls with 96-hr exposure. In contrast, plants exposed to 43 °C never exhibited a reduction in leaf nicotine content and showed a significant increase in content with just 48-hr exposure. When radioactive 11CO2 was administered to plants, HT stress significantly reduced fixation. Furthermore, radiocarbon flux analysis revealed that ‘new’ C partitioning (as 11C) into the [11C]-amino acid pool was significantly reduced with HT stress as were yields of [11C]-aspartic acid, an important amino acid in nicotine biosynthesis, and its beta-amido counterpart [11C]-asparagine significantly reduced. In contrast, [12C]-aspartic acid levels appeared unaffected at 38 °C but were significantly elevated at 43 °C relative to controls. Additionally, [12C]-asparagine levels were significantly elevated at both stress temperatures. Finally, leaf total soluble protein (TSP) was reduced 39% with long exposures to both stress temperatures. However, Ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) which was 41% TSP appeared unaffected by HT stress. Altogether, these results support the theory that plant proteins other than Rubisco degrade at elevated temperatures freeing up essential amino acids supporting the nicotine biosynthetic machinery, and while feedback might downregulate de novo amino acid synthesis from ‘new’ C resources, more than likely plant C/N rebalancing caused by HT reductions in C input restricted N uptake, which in turn, limited this process.
Keywords:
heat stress
; nicotine biosynthesis
; radiocarbon flux analysis
; carbon-11
; protein degradation
; L-Asparaginase activity
1. Introduction
Global warming is a real phenomenon that has been documented over the last 100 years [1,2]. With this trend in global warming, mean ambient temperatures are expected to rise 1.0 to 1.7 °C by 2050 and 4.0 to 5.0 °C by the end of the 21st century [3,4,5] at which point, high temperature stress will become one of the most prominent abiotic stresses confronted by the plant kingdom with major impacts to agricultural productivity [6,7,8,9,10].
While most studies concerning climate change have been concerned with plant growth and development, we recognize that many higher plants have evolved various strategies that allow them to adapt to certain environmental stresses. One important adaptation mechanism lies in their ability to synthesize specialized secondary metabolites. Alkaloids represent a group of very specialized secondary metabolites that play a major role in how certain plants can cope with defending themselves against pathogens and generalist herbivores [11,12,13,14]. These substrates often possess unique biological activities and even strong cytotoxic characteristics. Nicotiana, a member of the Solanaceous family, produces several pyrimidine-based alkaloids where nicotine is the most dominant in this class of specialized metabolite. Nicotine is biosynthesized primarily in the roots of the plant [15,16,17,18], after which it is transported through the xylem where it accumulates in leaf vacuoles to protect areas of the plant most vulnerable to attack [19,20].
Nicotine biosynthesis has been extensively investigated over the years, leading to a clear picture of its contributing pathways [21]. However, nicotine production is complicated involving multiple steps with two contributing pathways (Figure 1).
Here, the portion involving the formation of a pyrrolidine ring initiates with the formation of putrescine. Putrescine can be produced by two routes: either directly from ornithine involving the ornithine decarboxylase enzyme, or indirectly from arginine involving arginine decarboxylase. In the latter, arginine is decarboxylated to agmatine, which is subsequently hydrolyzed to N-carbamoylputrescine by agmatine iminohydrolase and further to putrescine by N-carbamoylputrescine amidohydrolase. Then, putrescine is converted to N-methylputrescine by putrescine N-methyltransferase where it is later deaminated oxidatively by N-methylputrescine oxidase to 4-methylaminobutanal and spontaneously cyclizes to N-methylpyrrolinium cation that contains the pyrrolidine ring.
In the other portion of the pathway aspartic acid is converted to α-iminosuccinic acid, then reacted with glycerol-3-phosphate to form the pyridine ring of quinolinic acid. Nicotinic acid is then formed by the same enzymes involved in nicotinamide adenine dinucleotide (NAD) biosynthesis, including aspartate oxidase, quinolinic acid synthase, and quinolinic acid phosphoribosyl transferase. While the exact nature of the metabolite derived from nicotinic acid is not known it is thought that nicotinic acid needs to be reduced to 3,6-dihydronicotinic acid before condensation with N-methylpyrrolinium cation to form nicotine.
Many abiotic conditions including water availability [22], light intensity and UV radiation [23], ambient temperature [24,25] as well as other environmental factors including nutrient availability are known to influence plant secondary metabolism. These conditions can induce changes at the level of gene expression, signal hormone transduction and enzyme activity, which will alter the content of specialized metabolites such as alkaloids [26].
Little is known about the specific effects of heat stress (HT) on the metabolic regulation of plant alkaloids largely because published works are too few and inconsistent. For example, the role of temperature on indole alkaloid content was studied in Catharanthus roseus hairy root cultures, where it was shown that by lowering the temperature the level of these alkaloids increased significantly [27]. Specifically, the expression of genes involved in alkaloid biosynthesis was highest in C. roseus plants at 12 °C. In that same work, nicotine production in Nicotiana tabacum was claimed to be significantly reduced at 30 °C when compared to 25 °C. In contrast to these early observations, more recent studies have shown that elevated temperatures promoted nicotine biosynthesis. In one study, investigators found that HT treatment induced nicotine biosynthesis in Nicotiana tabacum which was linked to the rapid accumulation of the suspected signaling agent, hydrogen sulfide. These same studies showed that HT triggered the biosynthesis of jasmonic acid, a plant hormone that is also linked to promoting nicotine biosynthesis [28]. In another study, HT induced expression of the tobacco transcription factor NtMYC2a that upregulated jasmonic acid biosynthesis by stimulating the transcription of genes associated with JA biosynthesis, including lipoxygenase, allene oxide synthase, allene oxide cyclase, and 12-oxophytodienodate reductase [29]. This action activated the biosynthesis of nicotine and related pyridine alkaloids in tobacco by up-regulating the expression of putrescine N-methyltransferase 1 (NtPMT1), which encoded putrescine N-methyltransferase to catalyze nicotine formation. In yet another more recent study, carbon monoxide signaling was linked to the increased expression of NtPMT1, via promotion of NtMYC2a binding to the G-box region of its promoter all leading to increased nicotine production with HT stress [30].
While these recent past studies provide compelling arguments for HT induction of the putrescine pathway in nicotine biosynthesis, there has not been a study that addresses whether HT stress impacts the aspartate pathway contributing to nicotinic acid biosynthesis and nicotine production. To explore the underlying mechanisms of metabolic regulation here, we undertook a series of studies in Nicotiana attenuata where plants were subjected to temperature stresses at 38 °C and 43 °C and examined for temporal changes in tissue nicotine content. Furthermore, we leveraged the power of radiocarbon flux measurements using radioactive carbon-11 to gain insight into the effects of HT stress on aspartate regulation.
2. Results
2.1. Acute and Chronic Heat Stress Alters Tissue Nicotine Content
We examined the effects of heat stresses at 38 °C (Figure 2A-C) and 43 °C (Figure 2D-F) applied across timelines spanning 24-hr to 96-hr of exposure at each temperature. Data was collated showing how temperature and time altered the nicotine content in mature leaves, young sink leaves and in root tissues.
Mature leaf nicotine content was initially reduced at 38 °C to 1.74 ± 0.12 mg g-1DW with 24-hr exposure and to 1.92 ± 0.46 mg g-1DW with 48-hr exposures relative to 25 °C control levels of 3.64 ± 0.54 mg g-1DW. However, these levels increased sharply and significantly by 96-hr exposure to 6.66 ± 0.58 mg g-1DW. Young sink leaves possessed less nicotine than fully expanded mature leaves harvested from the same plants. Even so, young leaves exhibited the same behavior as their mature leaf counterparts when exposed to 38 °C. Here levels dropped significantly to 0.45 ± 0.08 mg g-1DW with 24-hr exposure and 0.56 ± 0.15 mg g-1DW with 48-hr exposures relative to 25 °C control levels of 1.82 ± 0.19 mg g-1DW, but then started to rise slightly with 72-hr exposure to 1.56 ± 0.16 mg g-1DW and increase even more sharply and significantly to 4.29 ± 0.30 mg g-1DW with 96-hr exposure.
In contrast to data collected at 38 °C, mature leaf nicotine content of plants exposed to 43 °C never dropped below control values of 3.64 ± 0.54 mg g-1DW but rather rose gradually to 5.49 ± 0.77 mg g-1DW with 24-hr exposure, to 5.84 ± 1.31 mg g-1DW with 48-hr exposure, to 7.38 ± 0.96 mg g-1DW with 72-hr exposure and finally to 11.34 ± 0.71 mg g-1DW with 96-hr exposure. We also note that the final mature leaf nicotine content with 96-hr exposure to 43 °C temperature was significantly higher than that at 38 °C.
Identical trends were observed at 43 °C when examining changes in nicotine content in the young sink leaves. Here, a gradual but significant rise in content was observed as a function of exposure time to a final level of 6.42 ± 0.44 mg g-1DW at 96-hr exposure. Unlike their mature leaf counterparts, this rise in nicotine content at 43 °C was not significantly different to that at 38 °C.
Roots exhibited identical behavior at both elevated temperatures where significant downward trends in nicotine content were observed with increasing exposure from 1.34 ± 0.19 g-1DW to 0.74 ± 0.08 g-1DW.
2.2. Effect of Heat Stress on 11CO2 Fixation and ‘New’ Carbon Partitioning (as 11C) into Soluble Leaf Extracts
Using radioactive 11C administered to plants as 11CO2 we examined the effect of applied heat stresses on the photosynthetic machinery of the plant by measuring the extent of tracer fixation. Radiotracer was administered to plants after 48-hr exposure to heat stress temperatures of 38 °C and 43 °C and compared to levels for control plants grown at 25 °C. Results in Figure 3A show that leaf fixation of radiotracer decreased by these treatments. Specifically, fixation decreased increased significantly from 64.97 ± 2.66 percent of 11C radioactivity within the pulse (% fixation) for 25 °C control plants to 50.28 ± 3.70 % at 38 °C and 45.46 ± 2.90 % at 43 °C. Our radiotracer method also enabled measurement of ‘new’ carbon (as 11C) partitioning between soluble and insoluble fractions (Figure 3B). Here results showed that ‘new’ carbon partitioning into the soluble extract fraction increased significantly to 54.95 ± 3.72 % and 50.65 ± 5.58 % of total 11C activity assimilated by the leaf at 38 °C and 43 °C, respectively, relative to an average level observed for control plants of 38.57 ± 4.58 %.
2.3. Effect of Heat Stress on ‘New’ Carbon Partitioning (as 11C) into Amino Acids
The same tracer provided insight into the state of de novo amino acid biosynthesis. Results in Figure 4A showed that ‘new’ carbon partitioning into the total free amino pool decreased significantly to levels of 3.14 ± 0.56 % and 2.78 ± 1.06 % of total 11C-activity assimilated by the leaf at 38 °C and 43 °C, respectively, relative to control levels of 6.10 ± 0.99 %. Likewise, results in Figure 4B showed that ‘new’ carbon partitioning into [11C]-aspartic acid decreased significantly to levels of 0.69 ± 0.07 % and 0.41 ± 0.28 % of total 11C radiotracer assimilated by the leaf at 38 °C and 43 °C, respectively, relative to control levels of 1.49 ± 0.12 % as did levels of [11C]-asparagine drop (Figure 4C) to levels of 0.23 ± 0.03 % and 0.31 ± 0.09 %, respectively, relative to control levels of 1.17 ± 0.39 %.
2.4. Effect of Heat Stress on the Endogenous Concentrations of [12C]-Aspartic Acid and [12C]-Asparagine
Here, we examined the effect of heat stress on the endogenous pools of aspartic acid and asparagine amino acids in the same leaf tissue where radiocarbon fluxes were measured as well as in root tissues of same plants. Data was collected with 48-hr exposure to heat stress temperatures of 38 °C and 43 °C and compared to levels for control plants grown at 25 °C. Results in Figure 5A showed that the average endogenous leaf concentration of aspartic acid at 38 °C was 3.46 ± 1.79 µmoles per gram fresh weight of tissue (µmole g-1FW) which was not significantly different from the 5.63 ± 0.49 µmole g-1FW level observed for controls. In contrast, the leaf level of aspartic acid rose significantly to at 43 °C to 28.52 ± 1.60 µmole g-1FW. Results in Figure 5B showed that the average endogenous leaf concentration of asparagine rose significantly with applied heat stresses to 40.65 ± 4.39 µmole g-1FW and 102.85 ± 13.35 µmole g-1FW for 38 °C and 43 °C, respectively, relative to a 24.46 ± 5.14 µmole g-1FW level for controls.
In sharp contrast to leaves, root tissues presented a very different picture of how heat stress affected aspartic acid and asparagine levels. Results in Figure 5C showed that endogenous concentrations of aspartic acid dropped significantly to levels of 1.43 ± 0.28 µmole g-1FW and 1.01 ± 0.38 µmole g-1FW at 38 °C and 43 °C, respectively, relative to control levels of 4.91 ± 0.54 µmole g-1FW. In Figure 5D that the endogenous concentration of 38.22 ± 2.58 µmole g-1FW for asparagine at 38 °C was significantly higher than the 30.37 ± 4.97 µmole g-1FW level of controls, but then turned down sharply and significantly to 2.31 ± 0.55 µmole g-1FW at 43 °C. Overall, concentrations of aspartic acid and asparagine in roots was significantly less than those see in mature leaves where these amino acids are synthesized.
2.5. Effect of Heat Stress on Leaf Total Protein and Rubisco Protein Content
Finally, we examined the influence of heat stress on leaf soluble total protein content and leaf Rubisco content. Results in Figure 6A&B showed significant downward trends in the total soluble protein (TSP) content dropping 39% from a level of 116.68 ± 2.95 mg protein per gram dry weight (mg g-1DW) for 25 °C controls to 72.07 ± 3.380 mg g-1DW and 70.77 ± 3.71 mg g-1DW with 96-hr exposures to applied stresses at 38 °C and 43 °C, respectively. Contrary to this, leaf Rubisco which amounted to ~41% TSP decreased only slightly from the 47.22 ± 2.06 mg g-1DW level for 25 °C controls, but that downward trend in content with applied heat stress and exposure was not significant.
3. Discussion
The present work brings to light answers to why there has been so much inconsistency in past reporting of HT stress effects on nicotine biosynthesis [27,28,29,30]. Our work points to both temperature of the HT stress and the duration of exposure are equally important factors to consider when reporting results. The fact that we observed an initial decline in foliar nicotine levels at moderate 38 °C temperatures with short exposures to the stress, followed by significant increases in foliar nicotine with longer exposures tells us that the regulation of nicotine biosynthesis is complex involving several regulating factors.
In agreement with past HT stress studies [31,32,33,34], our work showed that leaf photosynthesis decreased significantly with applied stresses resulting in decreased C input. With the decreased availability of ‘new’ C resources, more was seen to partition into soluble metabolite pools. However, not so surprisingly our radiocarbon flux measurements showed a significant reduction in ‘new’ carbon partitioning (as 11C) into the total amino acid pool, and most particularly, a reduction in ‘new’ carbon partitioning into [11C]-aspartic acid and [11C]-asparagine amino acids that are relevant to nicotine biosynthesis.
Typically, sequestration of nitrogen (N) in higher plants is balanced by the plant’s C input or photosynthetic capacity [35]. This behavior has been clearly demonstrated in our past radiotracer experiments where young poplar saplings were tested for radioactive 13NO3- acquisition using Positron Emission Tomography [36]. In these studies, 13NO3- acquisition was examined along with leaf photosynthesis, measured by infrared gas exchange, and showed that changing trends in C input were tightly coupled with 13NO3- acquisition, and vice versa. Hence a lower photosynthetic capacity with HT stress should manifest in reduced N availability via nitrate uptake.
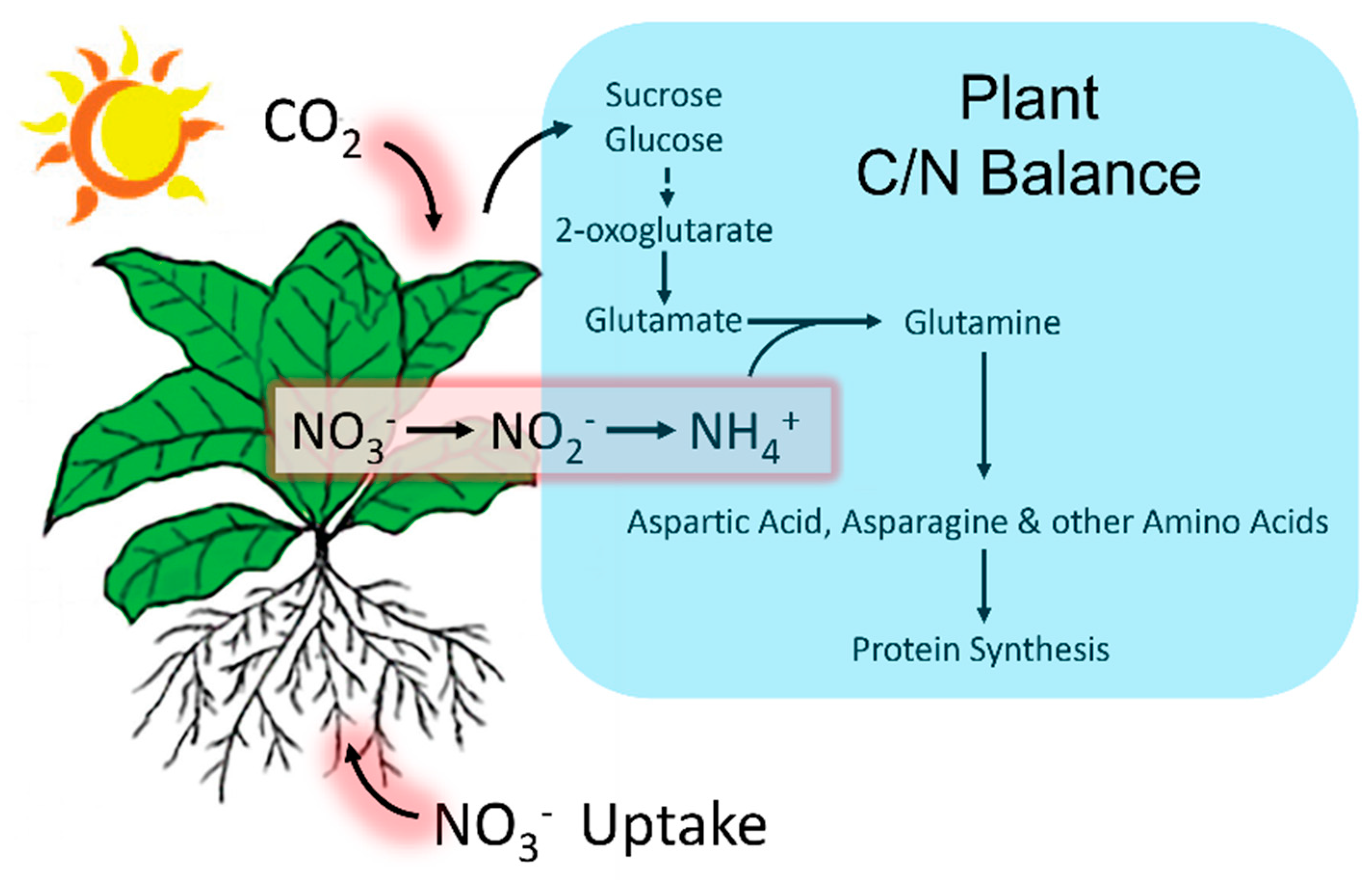
Shown pictorially in Figure 7, the reduction in 11CO2 fixation seen in the present work with applied HT stress likely reduced nitrate assimilation, (though not measured in the present work) which caused the reduction in ‘new’ carbon (as 11C) partitioning into amino acid synthesis.
In most higher plants, asparagine, and glutamine are the most abundant amino acids found in the plant vasculature although all amino acids are capable of transporting from sites of their biosynthesis to distal tissues where they are used in plant metabolism [37,38,39,40,41]. With shortfalls in the supply of [11C]-amino acids and particularly [11C]-asparagine, we questioned what was supplying the aspartic acid needed to support increased nicotine biosynthesis from HT stress.
Figure 7.
Plant tight coordination of C and N metabolism. CO2 fixation and N uptake occur in the leaf and the root systems, respectively. 2-oxoglutarate, an important intermediate product of C metabolism, serves as the C-skeleton for the synthesis of glutamate. Leaf ammonium (NH4+) derives primarily from root nitrate (NO3-) assimilation and transport aboveground where it is reduced and incorporated to glutamate and glutamine. Other amino acids including aspartic acid and asparagine are synthesized by using NH4+ donated from glutamate and glutamine [42]. These amino acids then provide the basic building blocks for protein synthesis.
Figure 7.
Plant tight coordination of C and N metabolism. CO2 fixation and N uptake occur in the leaf and the root systems, respectively. 2-oxoglutarate, an important intermediate product of C metabolism, serves as the C-skeleton for the synthesis of glutamate. Leaf ammonium (NH4+) derives primarily from root nitrate (NO3-) assimilation and transport aboveground where it is reduced and incorporated to glutamate and glutamine. Other amino acids including aspartic acid and asparagine are synthesized by using NH4+ donated from glutamate and glutamine [42]. These amino acids then provide the basic building blocks for protein synthesis.
Our examination of the endogenous pools of leaf aspartic acid and asparagine revealed a different story from the radiotracer results with possible insight into a mechanism for N remobilization. Here, 38 °C HT stress did not influence the endogenous pool of leaf aspartic acid, although the 43 °C stress did elevate its content significantly relative to controls. Furthermore, the pool of leaf asparagine was significantly elevated with application of both HT stresses. Since de novo biosynthesis of these amino acids were downregulated with HT stress, there needs to be another source maintaining and even increasing their content in tissues.
It is well established that many plant proteins will undergo degradation when subjected to HT stress [10,44,45]. In turn, this action will free up many α-amino acids that comprise the protein backbone. We examined the extent of protein degradation in the present work by measuring the content of both leaf total soluble protein (TSP) as well as Rubisco protein as a function of applied HT stresses and the different exposure times to these stresses. As noted earlier, TSPs decreased by nearly the same amount when subjected to either 38 °C or 43 °C stresses and these reductions became significant with greater than 24-hr exposures. Also, Rubisco which is typically the most abundant leaf protein (comprising 41% TSP in the present work) was not seen to degrade significantly with temperature nor exposure time. This behavior is consistent with prior reports of HT stress on Rubisco protein content [33,34] where reduction in photosynthetic capacity was attributed to loss of Rubisco activase and not to Rubisco protein content. Thus, the supply of essential ‘free’ α-amino acids supporting nicotine biosynthesis was rapid and identical for both stress temperatures and derived from ‘other’ regulatory proteins, and not Rubisco.
Even so, the rapid turnover of protein to free up α-amino acids at both HT stresses does not explain the initial drop observed in foliar nicotine content with short exposures at the lower 38 °C temperature. Hence, there must be another regulatory control mechanism underpinning the process. Some insight into its nature comes from our examination of root endogenous pools of aspartic acid and asparagine. As expected, the increased tug on the root aspartic acid pool when the nicotine biosynthesis was upregulated by HT stress manifested in a significant reduction in its root content. We suspect that the supply train feeding this aspartic acid pool may derive, in part, from the competitive conversion of recently ‘freed’ asparagine from protein degradation to aspartic acid. This action is driven by L-Asparaginase (EC 3.5.1.1), an enzyme that catalyzes asparagine’s hydrolysis yielding aspartic acid and ammonia in the process [46]. On closer inspection, our results show that root asparagine at 38 °C was significantly higher than control levels. As leaf protein degradation was prominent even at this temperature, foliar asparagine, ‘freed’ from this process, likely translocated to the roots causing the elevated levels observed in those tissues. We noted earlier that asparagine is one of the most abundant mobile amino acids found in plant vasculature so likely, what was remobilized in leaf tissue would supply the roots with this amino acid. Interestingly, at 43 °C, asparagine levels suddenly dropped to levels significantly less than controls. To rationalize this drastic shift in metabolism, we suggest that L-Asparaginase function may be strongly coupled to temperature and its state of activation may have accounted for the delayed response in root asparagine conversion to aspartic acid, and ultimately accounted for the delayed response observed in the HT induction of nicotine biosynthesis at 38 °C. Indeed, because of its high commercial value [47] with applications in the food industry and as an anti-cancer drug in the treatment of lymphoblastic leukemia, researchers have examined the thermostability of L-Asparaginase and found its activity is strongly correlated with temperature [48].
Lastly, we note that while nicotine production appeared to ramp up over time with exposure to applied HT stresses, root nicotine content continually dropped. Past work has identified several transport proteins in N. tabacum that likely play an important role in the cellular trafficking of nicotine [19,20]. Of course, once in the xylem, this alkaloid will quickly travel to aboveground tissues relying on the water transpiration stream to drive its translocation aboveground. Our data suggests that roots do not retain much of the nicotine alkaloid they produce but rather quickly ship it aboveground. This seems logical from a plant defense perspective.
4. Materials and Methods
4.1. Plant Growth
Nicotiana attenuata seeds (Accension W6 59297; USDA Agricultural Research Services, Pullman, WA, USA) were germinated in ProMix (Strawberry Hill Farms, Columbia, MO, USA) and after 10 days seedlings were transplanted to individual 3-inch pots filled with ProMix and supplemented with Osmocote™ Smart Release Plant Food (Scotts Co., Marysville, OH, USA) for continued growth in a commercial growth chamber (Model PGC-15, Percival Scientific, Perry, IA, USA) under 12-hr photoperiods at 500 μmol m-2 s-1 light intensity at constant temperature of 25 °C and relative humidity (RH, 60%). Heat stress studies were conducted in 5-wk-old plants. Plants were transferred from their control growth chamber to a second chamber elevated to either 38 °C or 43 °C while maintaining the same light intensity and relative humidity. Stress treatment duration was for 24, 48, 72 or 96-hr. Pots were placed in trays that held a surplus of water that was wicked up through the ProMix to maintain a constant well-watered status throughout the stress treatment.
4.2. Nicotine Analysis
After stress treatment, mature leaves and young sink leaves were harvested from individual plants and air dried for 1-week at 70 °C. Roots were also harvested, washed rigorously in water and air dried. Dried tissues were placed in pre-weighed 1.5 mL Eppendorf™ tubes equipped with a stainless-steel ball bearing. Sufficient tissue was introduced to each sample tube to ensure at least 500 mg of ground tissue would be available after the grinding process. Samples were ground to a powder using a ball mill grinder (Retsch, Inc., Newtown, PA, USA) and exact tissue masses recorded. Ground tissue was mixed with 0.5 mL 1M NaOH and 0.5 mL petroleum ether and the contents shook for 15-minutes using the ball mill grinder adjusted to a lower oscillation setting (10 oscillation s-1). The contents were centrifuged at 14,000 × g for 5-min and the upper organic layer removed. The organic extraction process was repeated twice more to yield a total of 1.5 mL of petroleum ether when all the extractions were combined. The contents were placed in a vacuum centrifuge (Labconco, Inc., Kansas City, MO, USA) set for 14,000 × g, 80 °C and a 30-min duration. This process evaporated the petroleum ether. Samples were then reconstituted in 100 µL methanol, vortex mixed and 1 µL aliquots were injected into a flame-ionization Hewlett Packard 5890A gas chromatograph equipped with a 30-m long capillary column, 0.53 mm I.D., 0.25 µm thick film of Rtx®-Wax (Restek, Inc., Bellefonte, PA, USA) with a 10 mL min-1 helium flowrate and a 1:50 sample split. At injection the initial column temperature was maintained at 100 °C and was programmed to increase to 220 °C at a 3 °C min-1 rate. The sample injector was maintained at 250 °C and the flame ionization detector at 350 °C. Chromatographic peaks for nicotine were recorded at a 7.4-min retention time and quantified using PeakSimple™ chromatography software v4.88 (SRI, Inc., Torrance, CA, USA) against a commercial nicotine standard (Sigma Aldrich, St. Louis, MO, USA).
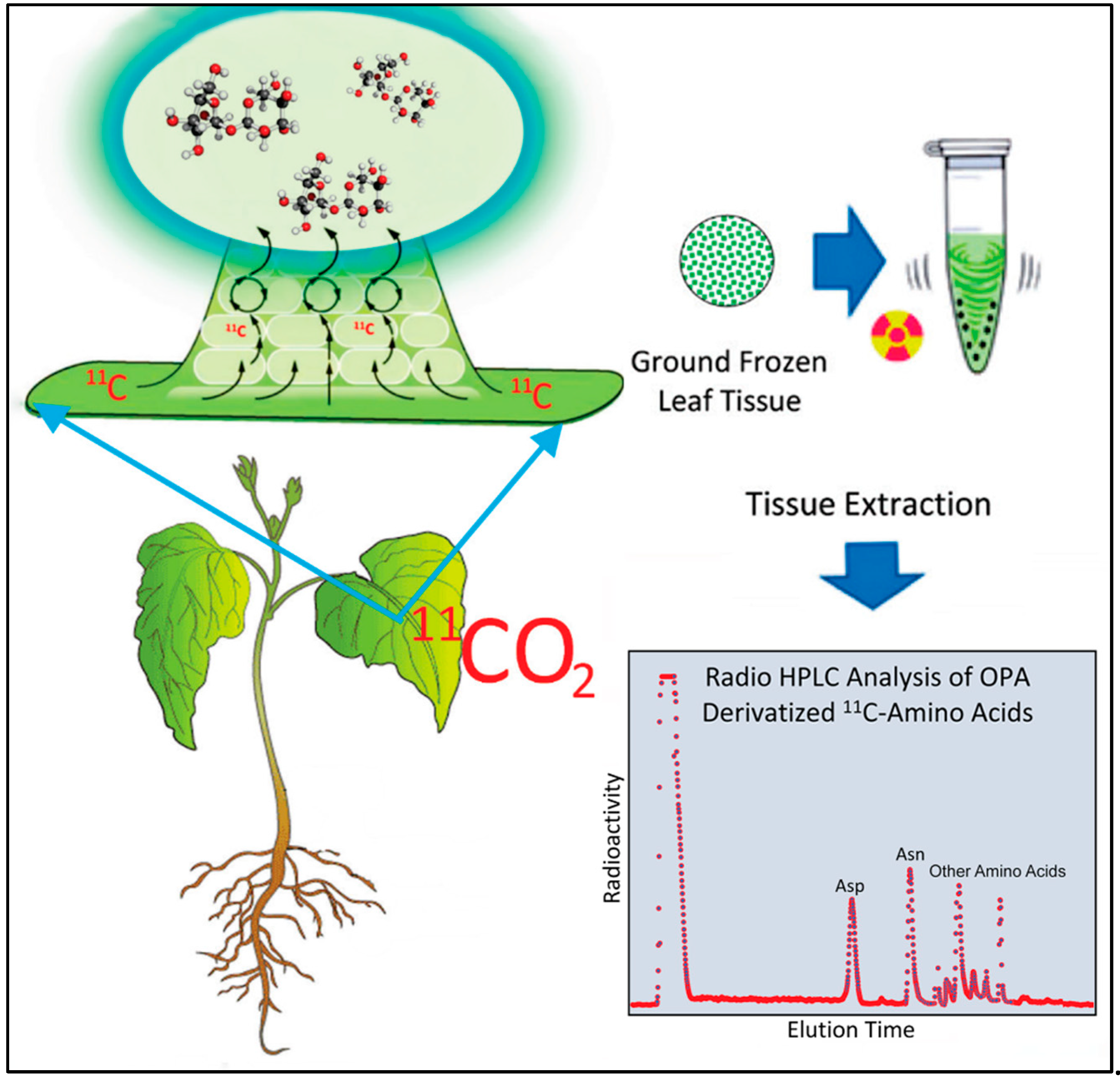
4.3. Production and Administration of Radioactive 11CO2
11CO2 (t½ 20.4 min) was produced on the GE PETtrace Cyclotron located at the Missouri Research Reactor Center using high-pressure research grade N2 gas target irradiated with a 16.4 MeV proton beam to generate 11C via the 14N(p,α)11C nuclear transformation [49,50]. The 11CO2 was trapped on the molecular sieve, desorbed, and quickly released into an air stream at 200 mL min-1 as a discrete pulse for labeling a leaf affixed within a 5 x 10 cm lighted (560 µmol m-2 s-1) leaf cell to ensure a steady level of fixation. The load leaf affixed within the cell was pulse-fed 11CO2 for 1 min, then chased with normal air for the duration of exposure [51]. A PIN diode radiation detector (Carroll Ramsey Associates, Berkeley, CA USA) attached to the bottom of the leaf cell enabled continuous measurement of radioactivity levels within the cell during the initial pulse and in the minutes directly following to give information on 11CO2 fixation. The attached leaf was incubated for 20-min after which exposed tissue was harvested for analysis.
4.4.[11/12. C]-Aspartic Acid and [11/12C]-Asparagine Analyses
Exposed leaf tissue (~500 mg) was flash frozen in liquid nitrogen, ground to a fine powder in pre-weighed 1.5 mL Eppendorf™ tubes using a ball mill grinder, then weighed for exact tissue mass and extracted in 1 mL methanol: water (60:40 v/v) using a cell disruptor (Branson, Bransonic 32; Sigma-Aldrich Corp) set at 100% amplitude for 2-minute duration. Afterwards, samples were centrifuged @ 14,000 × g for 2-minutes to separate insoluble and soluble portions. Supernatant was removed and placed in a separate Eppendorf™ where a 20 µL aliquot was removed for counting [11C]-radioactivity using a NaI gamma counter. The pellet was also counted for [11C]-radioactivity on the same instrument. Radioactivity data was decay corrected back to the end of bombardment, or end of cyclotron beam. A visual of the workflow here can be found in Figure 8. The combined radioactivity of the fraction corrected soluble extract, plus pellet provided a measure of total [11C]-radioactivity from which to base [11C]-amino acid yields.
[11/12C]-Aspartic acid and [11/12C]-asparagine were analyzed using pre-column OPA(o-phthaldehyde) derivatization and quantified by gradient radio HPLC (Sonntek, Inc. Upper Saddle River, NJ 07458, USA) using a Phenomenex Luna 5µm C18(2), 100 mm x 4.6 mm inner diameter column heated to 30 °C and mobile phase system comprised of Solvent A (30 mM KH2PO4; 0.4% (v/v) tetrahydrofuran; pH 6.8), Solvent B (1:1 acetonitrile: water) starting at 100% of A at injection and ramping to 50:50 of A:B within 20 minutes at a flow rate of 1.5 mL min-1. This method enabled baseline separation of aspartic acid and asparagine in less than 6 minutes but left many of the remaining amino acids unresolved. On-line fluorescence detection (340 nm excitation, 450 nm emission; Hitachi LaChrom Elite L-2485; Sonntek, Inc.) was used for quantification of the OPA-derivatized peaks that were associated with [12C]-aspartic acid and [12C]-asparagine and a NaI gamma radiation detector (Eckert and Ziegler, Inc., Wilmington, MA, USA) enabled direct measurement of the [11C]-labelled amino acids. Data was acquired using PeakSimple™ chromatography software v4.88 (SRI, Inc., Torrance, CA, USA) and compared to standard calibration curves constructed for aspartic acid and asparagine providing absolute amounts for these amino acids in units of µmol per gram fresh weight (g-1FW) tissue. Radioactive metabolite peaks for [11C]-aspartic acid and [11C]-asparagine were quantified using the same software, corrected for radioactive decay, and corrected for differences in efficiencies of the gamma counter NaI detector and the HPLC’s NaI detector. As noted earlier, the other radioactive amino acids were unresolved with our HPLC method but were recorded as a sum of their contributions to the radiolabeled amino acid pool. After corrections, individual [11C]-aspartic acid and [11C]-asparagine data was reported as percent total [11C]-radioactivity within the plant as was the total [11C]-amino acid fraction reported as percent total [11C]-radioactivity.
4.5. Protein Analysis
Mature leaf tissues were harvested after the different stress treatments were applied and tissues were stored at -80 °C until analyzed. For protein extraction, samples were placed in a freeze dryer (FreezeZone 1 Dryer, Labconco Corp., Kansas City, MO, USA) and lyophilized. Once dried these samples were ground to a fine powder using the ball mill grinder, weighed in 100 mg amounts (± 1 mg) and placed into 1.5 mL Eppendorf™ tubes. Leaf pigments and other confounding leaf metabolites were first removed by extracting with samples using 1 mL cold acetone (5% trichloroacetic acid, w/v) following published procedures [52]. Samples were then sonicated for 15-min and centrifuged at 14,000 × g for 10-min to separate the protein pellet. Once the supernatant was removed, the pellet was washed three more times with cold acetone to remove lingering trichloroacetic acid. Samples were dried at ambient temperature under vacuum. Once dried, total proteins were extracted in 1 mL of 50 mM Tris–Cl buffer (pH 8.8) containing (1% v/v) plant protease inhibitors (Plant Protease Inhibitor Cocktail, Sigma Aldrich). Liquefied tissues were clarified with centrifugation at 14,000 × g for 10 min, and supernatant was removed and placed into a clean tube for analysis.
Following published a recent procedure [53] describing selective Rubisco fractionation, we mixed 500 µL of protein extract with an equal volume (50% v/v) of 100 mM CaCl2·2H2O and sodium phytate (Sigma Aldrich, St. Louis, MO, USA) to selectively precipitate the Rubisco protein. After addition of the fractionation agents, gentle mixing and incubation, solutions were centrifuged at 16,100 × g for 10-min and the supernatant removed. The protein precipitate was washed with cold acetone and resuspended in 50 mM Tris–Cl buffer (pH 8.8) containing (1% v/v) plant protease inhibitors. The solution was clarified by centrifugation and the resolubilized Rubisco protein collected for analysis.
Following published procedures for the Bradford Assay (Sigma Aldrich Technical Report B6916) 100 µL aliquots of protein extract were removed from each sample tube and mixed with 3 mL of Bradford Reagent (Sigma Aldrich, Inc., St. Louis, MO, USA) in 10 mL glass vials [54]. Samples were incubated at room temperature for 45-min then transferred to optical cuvettes for measuring their absorbance at 595 nm using an Evolution 201 UV/VIS spectrophotometer (ThermoFisher Scientific Inc., Waltham, MA, USA). Absorbances were correlated with a standard curve created from measurements using a commercial protein standard (1 mg mL-1) of bovine serum albumin (Sigma Aldrich, Inc., St. Louis, MO, USA).
4.6. Statistical Analysis
Data was subjected to one-way analysis of variance (ANOVA) in R using SigmaPlot 14.5. Tukey’s HSD test was used for post hoc correction of comparisons of stress treatments to unstressed control plants. Significance was set at a level of P < 0.05.
5. Conclusions
In summary, this work clearly demonstrates that nicotine biosynthesis is inducible with HT stress. However, regulatory control of the nicotine pathway, at least regarding aspartic acid’s contribution, is not straight forward. With short exposure times at 38 °C foliar tissues had less nicotine content than 25 °C controls, but this level increased over time of exposure to the stress. At 43 °C, foliar nicotine levels were consistently elevated over those of control plants. Our application of radiocarbon flux analysis provided mechanistic insight into the reasons for this difference. With HT stress, plant C input via photosynthesis declines significantly. Plants adapt to this by rebalancing their N uptake to their C input. Hence, we surmise there is less N available to supply amino acid biosynthesis which accounted for the reduction in radiolabeled amino acids. To supply nicotine biosynthesis, we discovered that protein degradation at HT was likely remobilizing these essential amino acids. We also note that the delayed response in the HT induction of nicotine biosynthesis with short exposure to 38 °C temperatures likely was due to the temperature dependence of L-Asparaginase enzyme responsible for converting asparagine into aspartic acid in the root tissues.
From an ecological perspective, the stresses imposed by climate change will naturally weaken a plant. To survive, plants might elevate their defense posture. At least for N. attenuata, we see an upregulation in chemical defense synthesis helping to protect the plant from herbivores while in a weakened state of a C/N resource deficiency.
Author Contributions
Experimental design and supervision - R.A.F.; data collection and analysis – S.W, A.P., R.N., M.J.S., R.A.F.; initial draft - R.A.F.; final draft - all authors.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data needed to evaluate the conclusions in the paper are present in the main text.
Acknowledgments
We would like to thank the leadership team and support staff of the Missouri Research Reactor Center for providing financial and infrastructure support for this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Keller, C.F. Global warming 2007: An update to global warming: the balance of evidence and its policy implications. Sci. World J. 2007, 7, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Schnoor, J.L. The IPCC fourth assessment. Environ. Sci. Technol. 2007, 41, 1503. [Google Scholar] [CrossRef] [PubMed]
- De Costa, W.A.J.M. A review of the possible impacts of climate change on forests in the humid tropics. J Natl Sci Found Sri 2011, 39, 281–302. [Google Scholar] [CrossRef]
- Jones, N. Climate assessments: 25 years of the IPCC. Nature 2013, 501, 298–299. [Google Scholar] [CrossRef] [PubMed]
- Kintisch, E. Climate science. For researchers, IPCC leaves a deep impression. Science 2013, 342, 24. [Google Scholar]
- Deryng, D.; Conway, D.; Ramankutty, N.; Price, J. , Warren, R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 2014, 9, 1–13. [Google Scholar]
- Horton, D.; Johnson, N.C.; Singh, D.; Swain, D.; Rajaratnam, B.; Diffenbaugh, N. Contribution of changes in atmospheric circulation patterns to extreme temperature trends. Nat. Cell Biol. 2015, 522, 465–469. [Google Scholar]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; Durand, J.L.; Elliott, J.; Ewert, F.; Janssens, I.A.; Li, T.; Lin, E.; Liu, Q.; Martre, P.; Müller, C.; Peng, S.; Peñuelas, J.; Ruane, A.C.; Wallach, D.; Wang, T.; Wu, D.; Liu, Z.; Zhu, Y.; Zhu, Z.; Asseng, S. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. (USA) 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Delmotte, M.V.; Zhai, P.; Pörtner, H.O.; Roberts, D.; Skea, J.; Shukla, P.R.; Pirani, A. ; Moufouma-Oki,W.; Péan, C., Pidcock, R., Connors, S., Matthews, J.B.R., Chen, Y., Zhou, X., Gomis, M.I., Lonnoy, E., Maycock, T., Tignor, M., Eds.; Waterfield, T. (Eds.) IPCC Global Warming of 1.5 °C—An IPCC Special Report on the Impacts of Global Warming of 1.5 C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways; IPPC: Geneva, Switzerland, 2018; Volume 2. [Google Scholar]
- Akter, N.; Islam, M.R. Heat stress effects and management in wheat: A review. Agron. Sustain. Dev. 2017, 37, 37–54. [Google Scholar]
- Baldwin, I.T. The alkaloidal responses of wild tobacco to real and simulated herbivory. Oecologia 1988, 77, 378–381. [Google Scholar]
- Baldwin, I.T. Mechanism of damage-induced alkaloid production in wild tobacco. J Chem. Ecol. 1989, 15, 1661–1680. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T. Jasmonate-induced responses are costly but benefit plants under attack in native populations. Proc. Nat’l. Acad. Sci. (USA) 1998, 95, 8113–8118. [Google Scholar] [CrossRef] [PubMed]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural Products (Secondary Metabolites). Biochemistry & Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Maryland, 2000; pp. 1250–1318. [Google Scholar]
- Bedewitz, M.A.; Góngora-Castillo, E.; Uebler, J.B.; Gonzales-Vigil, E.; Wiegert-Rininger, K.E.; Childs, K.L.; Hamilton, J.P.; Vaillancourt, B.; Yeo, Y.S.; Chappell, J.; DellaPenna, D.; Jones, A.D.; Buell, C.R.; Barry, C.S. A root-expressed L-phenylalanine: 4-hydroxyphenylpyruvate aminotransferase is required for tropane alkaloid biosynthesis in Atropa belladonna. Plant Cell 2014, 26, 3745–3762. [Google Scholar] [CrossRef] [PubMed]
- Biastoff, S. , Brandt, W., Dräger, B. Putrescine N-methyltransferase—The start for alkaloids. Phytochem 2009, 70, 1708–1718. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, C.; Cordeiro, A.; Alcázar, R.; Borrell, A.; Culiañez-Macià, F.A.; Tiburcio, A.F.; Altabella, T. Localization of arginine decarboxylase in tobacco plants. Physiol. Plantarum 2004, 120, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Katoh, A.; Ohki, H.; Inai, K.; Hashimoto, T. Molecular regulation of nicotine biosynthesis. Plant Biotechnol. 2005, 22, 389–392. [Google Scholar] [CrossRef]
- Shitan, N.; Minami, S.; Morita, M.; Hayashida, M.; Ito, S.; Takanashi, K.; Omote, H.; Moriyama, Y.; Sugiyama, A.; Goossens, A.; Moriyasu, M.; Yazaki, K. Involvement of the leaf-specific multidrug and toxic compound extrusion (MATE) transporter Nt-JAT2 in vacuolar sequestration of nicotine in Nicotiana tabacum. PLoS ONE 2014, 9, e108789. [Google Scholar] [CrossRef]
- Shitan, N.; Hayashida, M.; Yazaki, K. Translocation and accumulation of nicotine via distinct spatio-temporal regulation of nicotine transporters in Nicotiana tabacum. Plant Signaling & Behavior 2015, 10, e1035852. [Google Scholar]
- Leete, E.; Siegfried, K.J. The biogenesis of nicotine. III. Further observations on the incorporation of ornithine into the pyrrolidine ring. J American Chem. Soc. 1957, 19, 4529–4531. [Google Scholar]
- Tso, T.C. Organic metabolism - Alkaloids. In Production, Physiology and Biochemistry of the Tobacco Plant; Ideals Inc.: Beltsville, Maryland, USA, 1993. [Google Scholar]
- Voipio, I.; Autio, J. Responses of red-leaved lettuce to light intensity, UV-A radiation, and root zone temperature. Acta Horticulture 1995, 399, 183–190. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Hoppula, K.I.; Nestby, R.; Verheul, M.J.; Kar-jalainen, R.O. Influence of fertilization, mulch color, early forcing, fruit order, planting date, shading, growing environment, and genotype on the contents of selected phenolics in strawberry (Fragaria ananassa Duch.) fruits. J Agricultural and Food Chemistry 2006, 54, 2614–2620. [Google Scholar] [CrossRef]
- Coolong, T.W.; Randle, W.M. The influence of root zone temperature on growth and flavour precursors in Allium cepa L. J Horticultural Sci. and Biotech. 2006, 81, 199–204. [Google Scholar] [CrossRef]
- Ziegler, J.; Facchini, P.J. Alkaloid biosynthesis: metabolism and trafficking. Annu. Rev. Plant Biol. 2008, 59, 735–69. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, L.; Laakso, S.; Rosenqvist, H. The effect of temperature on growth, indole alkaloid accumulation and lipid composition of Catharanthus roseus cell suspension cultures. Plant Cell Rep. 1992, 11, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, Q.; Zhang, X.; Li, R.; Jia, Y.; Ef, A.-A.; Jia, A.; Hu, L.; Hu, X. Hydrogen sulfide mediates nicotine biosynthesis in tobacco (Nicotiana tabacum) under high temperature conditions. Plant Physiol. and Biochem. 2016, 104, 174–179. [Google Scholar] [CrossRef]
- Yang, L.; Li, J.; Ji, J.; Li, P.; Yu, L.; Abd-Allah, E.F.; Luo, Y.; Hu, L.; Hu, X. High Temperature Induces expression of tobacco transcription factor NtMYC2a to regulate nicotine and JA biosynthesis. Front. Physiol. 2016, 7, 465. [Google Scholar] [CrossRef]
- Cheng, T.; Hu, L.; Wang, P.; Yang, X.; Peng, Y.; Lu, Y.; Chen, J.; Shi, J. Carbon monoxide potentiates high temperature-induced nicotine biosynthesis in tobacco. Int. J. Mol. Sci. 2018, 19, 188. [Google Scholar] [CrossRef]
- Salvucci, M.E.; Crafts-Brandner, S.J. Mechanism for deactivation of Rubisco under moderate heat stress. Physiol. Plantarum 2004, 122, 513–519. [Google Scholar] [CrossRef]
- Schrader, S.M.; Wise, R.R.; Wacholtz, W.F.; Ort, D.R.; Sharkey, T.D. Thylakoid membrane responses to moderately high leaf temperature in Pima cotton. Plant, Cell & Environ. 2004, 27, 725–735. [Google Scholar]
- Perdomo, J.A.; Capó-Bauçà, S.; Carmo-Silva, E.; Galmés, J. Rubisco and Rubisco activase play an important role in the biochemical limitations of photosynthesis in rice, wheat, and maize under high temperature and water deficit. Front. Plant Sci. 2017, 8, 490. [Google Scholar] [CrossRef]
- Degen, G.E.; Orr, D.J.; Carmo-Silva, E. Heat-induced changes in the abundance of wheat Rubisco activase isoforms. New Phytol. 2021, 229, 1298–1311. [Google Scholar] [CrossRef]
- Zheng, Z.-L. Carbon and nitrogen nutrient balance signaling in plants. Plant Signaling & Behavior 2009, 4, 584–591. [Google Scholar]
- Ferrieri, R.A.; Herman, E.; Babst, B.; Schueller, M.J. Managing the Soil Nitrogen Cycle in Agroecosystem. In Soil Nitrogen uses and Environmental Impacts; Lai, R., Stewart, B.A., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, Chapter 14; pp. 343–360.
- Lohaus, G.; Burba, M.; Heldt, H.W. Comparison of the contents of sucrose and amino acids in the leaves, phloem sap and taproots of high and low sugar-producing hybrids of sugar beet (Beta vulgaris L.). J. Exp. Bot. 1994, 45, 1097–1101. [Google Scholar] [CrossRef]
- Rentsch, D.; Schmidt, S.; Tegeder, M. Transporters for uptake and allocation of organic nitrogen compounds in plants. FEBS Lett. 2007, 581, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Hammes, U.Z. The way out and in: phloem loading and unloading of amino acids. Curr. Opinion in Plant Biol. 2018, 43, 16–21. [Google Scholar] [CrossRef]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino acid transporters in plants: Identification and function. Plant 2020, 9, 972. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Zhai, Y.; Guo, M.; Wang, H.; Lu, J.; Liu, J.; Zhang, C.; Gong, Z.; Lu, M. Autophagy, a conserved mechanism for protein degradation, responds to heat, and other abiotic stresses in Capsicum annuum L. Front. Plant Sci. 2016, 7, 131. [Google Scholar] [CrossRef]
- Su, T.; Yang, M.; Wang, P.; Zhao, Y.; Ma, C. Interplay between the ubiquitin proteasome system and ubiquitin-mediated autophagy in plants. Cells 2020, 9, 2219. [Google Scholar] [CrossRef]
- Zhang, Y.; Min, H.; Shi, C.; Xia, G.; Lai, Z. Transcriptome analysis of the role of autophagy in plant response to heat stress. PLoS ONE 2021, 16, e0247783. [Google Scholar] [CrossRef]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Annals. Applied Biol. 2006, 150, 1–26. [Google Scholar] [CrossRef]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A comprehensive review on L-Asparaginase and its applications. Applied Biochem. and Biotech. 2016, 178, 900–923. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Xu, S.; Zhang, H.; Xu, M.; Yang, T.; Wang, L.; Qian, H.; Zhang, H.; Fang, H.; Osire, T.; Rao, Z.; Yang, S. Simultaneous cell disruption and semi-quantitative activity assays for high-throughput screening of thermostable L-Asparaginases. Scientific Reports 2018, 8, 7915. [Google Scholar] [CrossRef]
- Ferrieri, R.A.; Wolf, A.P. The chemistry of positron-emitting nucleogenic (hot) atoms with regard to preparation of labeled compounds of practical utility. Radiochim. Acta. 1983, 34, 69–83. [Google Scholar] [CrossRef]
- Ferrieri, R.A. Production and application of synthetic precursors labeled with carbon-11 and fluorine-18. In Handbook of Radiopharmaceuticals: Radiochemistry and Applications; Welch, M.J., Redvanly, C.S., Eds.; John Wiley & Sons, Ltd.: Chichester, England, 2003. [Google Scholar]
- Ferrieri, R.A.; Gray, D.W.; Babst, B.A.; Schueller, M.J.; Schlyer, D.J.; Thorpe, M.R.; Orians, C.M.; Lerdau, M. Use of carbon-11 in Populus shows that exogenous jasmonic acid increases biosynthesis of isoprene from recently fixed carbon. Plant, Cell & Environ. 2005, 25, 591–602. [Google Scholar]
- Wu, X.; Xiong, E.; Wang, W.; Scali, M.; Cresti, M. Universal sample preparation method integrating trichloroacetic acid/acetone precipitation with phenol extraction for crop proteomic analysis. Nature Protocols 2014, 9, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B.; Natarajan, S.S. A rapid method for depletion of Rubisco from soybean (Glycine max) leaf for proteomic analysis of lower abundance proteins. Phytochem. 2009, 70, 1958–1964. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Figure 1.
Biosynthetic pathway for nicotine production.
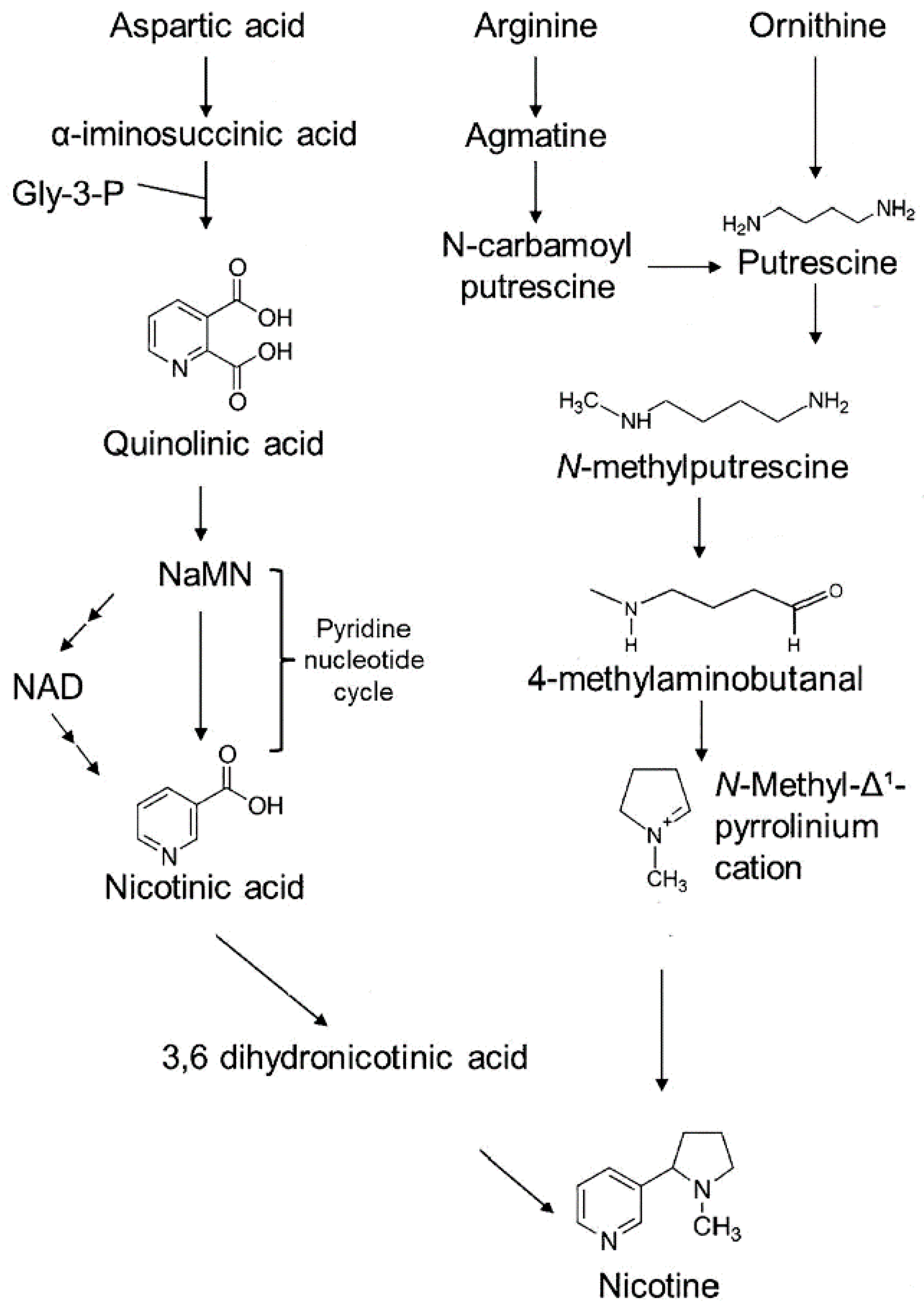
Figure 2.
The effect of heat stresses on nicotine content presented as milligram of nicotine per gram dry weight tissue (mg g-1DW). Heat stresses were examined at 38 °C (Panels 2A-C) and 43 °C (Panels 2D-F) applied across exposure timelines spanning 24-hr to 96-hr. Tissues examined included mature expanded leaves, young developing leaves, and roots. All data points are shown for at least N=6 replicates for each timepoint. The 25 °C control data average is shown as the dashed line in each panel. Average values for each data set are presented as black horizontal bars with levels of significance set as: *, P < 0.05; **, P < 0.01; ***, P < 0.001. Best fit trendlines were applied to data within each panel.
Figure 2.
The effect of heat stresses on nicotine content presented as milligram of nicotine per gram dry weight tissue (mg g-1DW). Heat stresses were examined at 38 °C (Panels 2A-C) and 43 °C (Panels 2D-F) applied across exposure timelines spanning 24-hr to 96-hr. Tissues examined included mature expanded leaves, young developing leaves, and roots. All data points are shown for at least N=6 replicates for each timepoint. The 25 °C control data average is shown as the dashed line in each panel. Average values for each data set are presented as black horizontal bars with levels of significance set as: *, P < 0.05; **, P < 0.01; ***, P < 0.001. Best fit trendlines were applied to data within each panel.
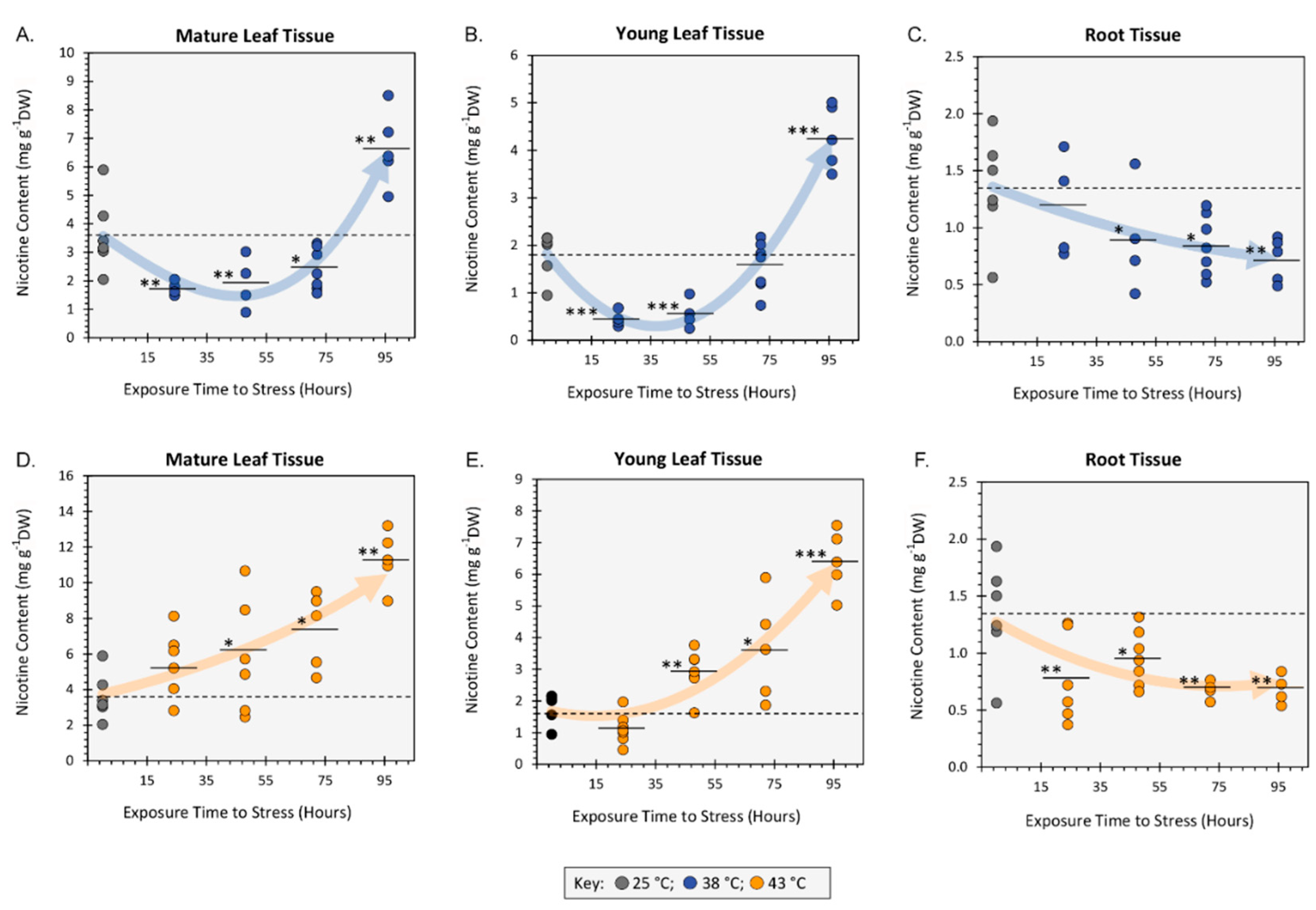
Figure 3.
The effect heat stresses of 38 °C and 43 °C applied for 48-hr on 11CO2 fixation in mature leaf tissue and the partitioning of ‘new’ carbon (as 11C) into soluble and insoluble fractions. All data bars represent average values ± SE percent of total 11C activity assimilated by the leaf for N = 3-5 replicates. Panel A: percent 11CO2 fixation (leaves were normalized to a standard mass). Panel B: percent distribution of 11C into the soluble fraction of leaf extracts. Levels of significance were set as: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
Figure 3.
The effect heat stresses of 38 °C and 43 °C applied for 48-hr on 11CO2 fixation in mature leaf tissue and the partitioning of ‘new’ carbon (as 11C) into soluble and insoluble fractions. All data bars represent average values ± SE percent of total 11C activity assimilated by the leaf for N = 3-5 replicates. Panel A: percent 11CO2 fixation (leaves were normalized to a standard mass). Panel B: percent distribution of 11C into the soluble fraction of leaf extracts. Levels of significance were set as: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
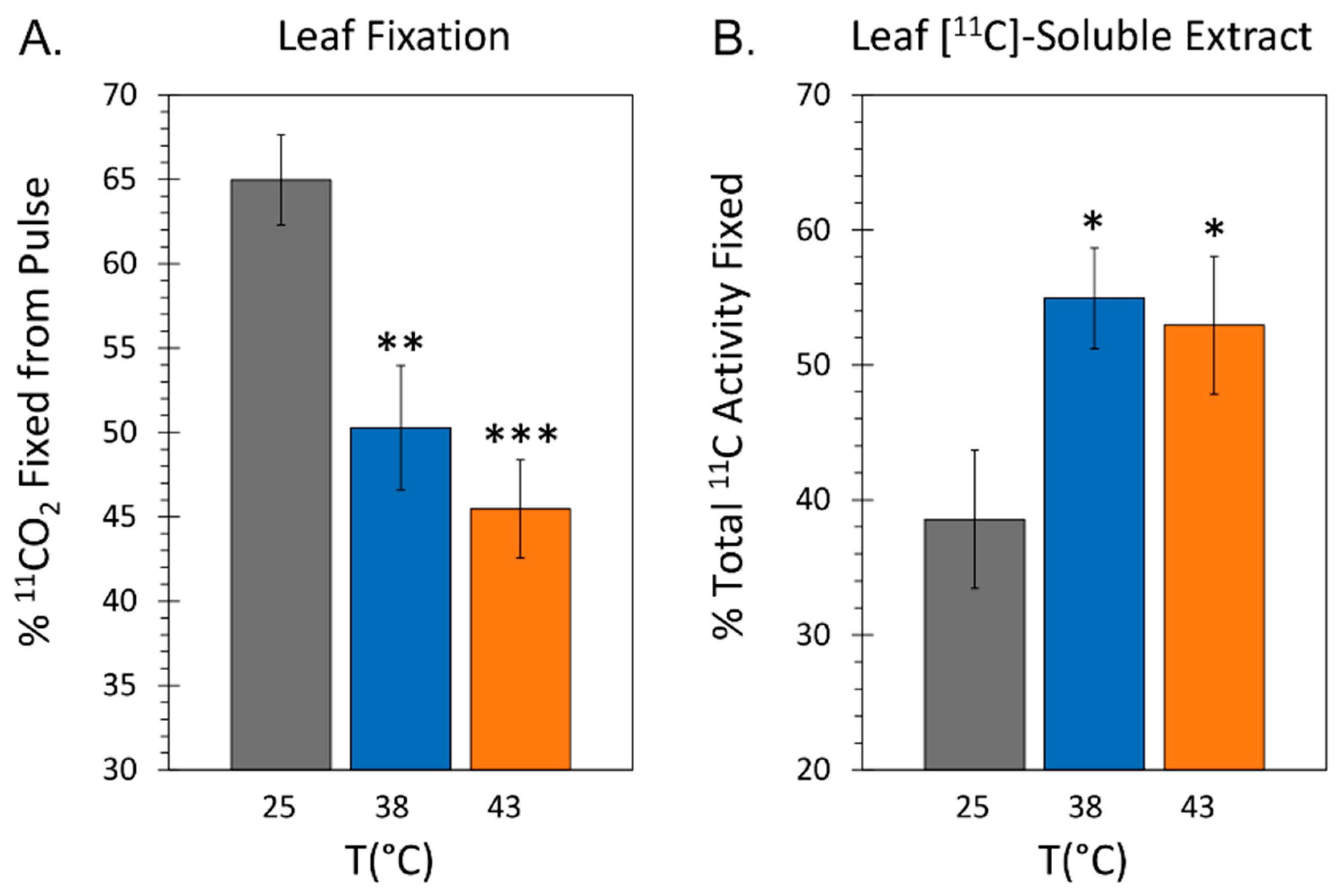
Figure 4.
The effect heat stresses of 38 °C and 43 °C applied for 48-hr on the metabolic partitioning of ‘new’ carbon (as 11C) in mature leaf tissue. All data bars represent average values ± SE percent of total 11C activity assimilated by the leaf vis 11CO2 fixation for N = 3-5 replicates. Panel A: percent of 11C partitioning into the total amino acid pool. Panel B: percent of 11C partitioning into [11C]-aspartic acid. Panel C: percent of 11C partitioning into [11C]-asparagine. Levels of significance set as: *, P < 0.05; **, P < 0.01.
Figure 4.
The effect heat stresses of 38 °C and 43 °C applied for 48-hr on the metabolic partitioning of ‘new’ carbon (as 11C) in mature leaf tissue. All data bars represent average values ± SE percent of total 11C activity assimilated by the leaf vis 11CO2 fixation for N = 3-5 replicates. Panel A: percent of 11C partitioning into the total amino acid pool. Panel B: percent of 11C partitioning into [11C]-aspartic acid. Panel C: percent of 11C partitioning into [11C]-asparagine. Levels of significance set as: *, P < 0.05; **, P < 0.01.
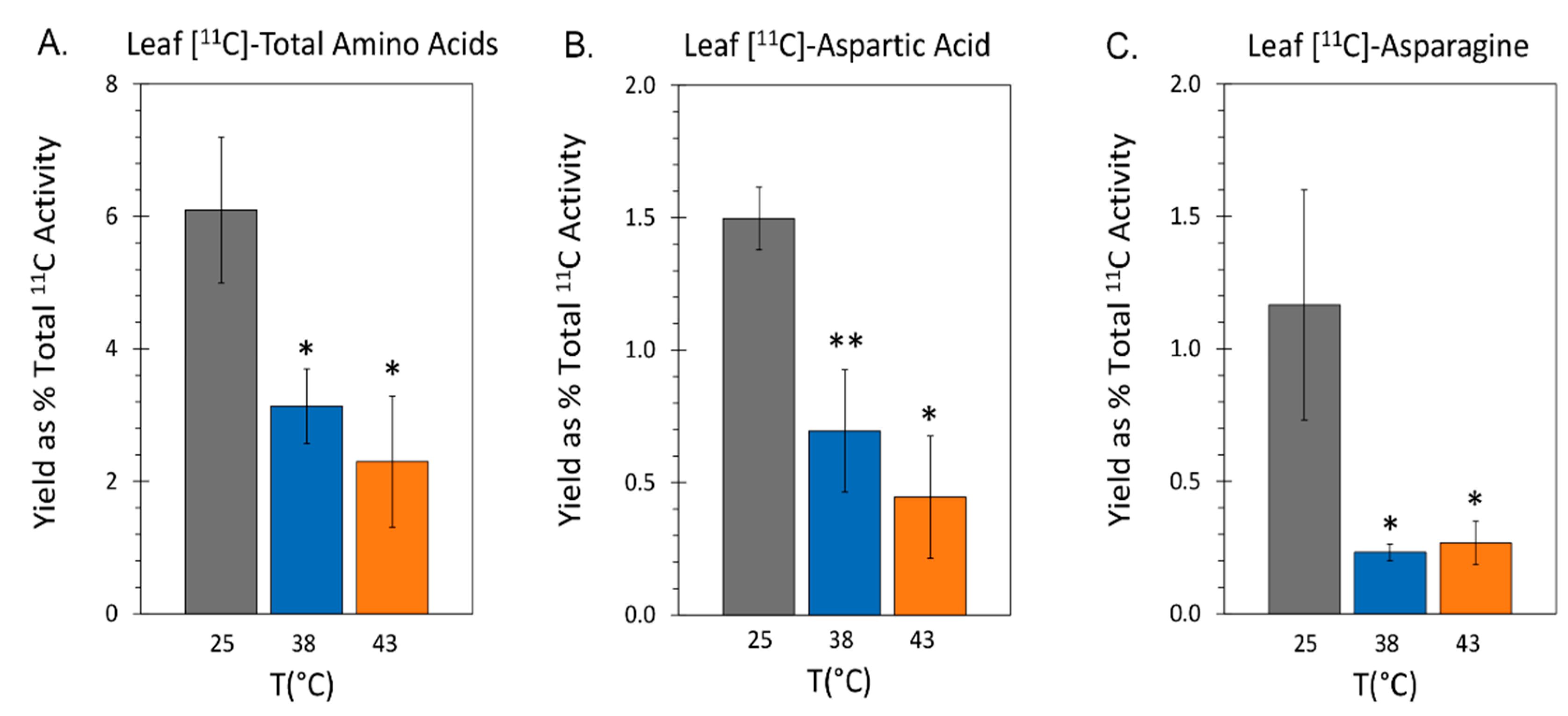
Figure 5.
The effect heat stresses of 38 °C and 43 °C applied for 48-hr on the endogenous concentrations of aspartic acid and asparagine amino acids in mature leaves and roots. All data bars represent average values ± SE in micromoles of metabolite per gram fresh weight of tissue (µmole g-1FW) for N = 3-5 replicates. Panels A&B reflect levels of aspartic acid and asparagine in leaves. Panels C&D reflect levels of aspartic acid and asparagine in roots. Levels of significance set as: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
Figure 5.
The effect heat stresses of 38 °C and 43 °C applied for 48-hr on the endogenous concentrations of aspartic acid and asparagine amino acids in mature leaves and roots. All data bars represent average values ± SE in micromoles of metabolite per gram fresh weight of tissue (µmole g-1FW) for N = 3-5 replicates. Panels A&B reflect levels of aspartic acid and asparagine in leaves. Panels C&D reflect levels of aspartic acid and asparagine in roots. Levels of significance set as: *, P < 0.05; **, P < 0.01; ***, P < 0.001.
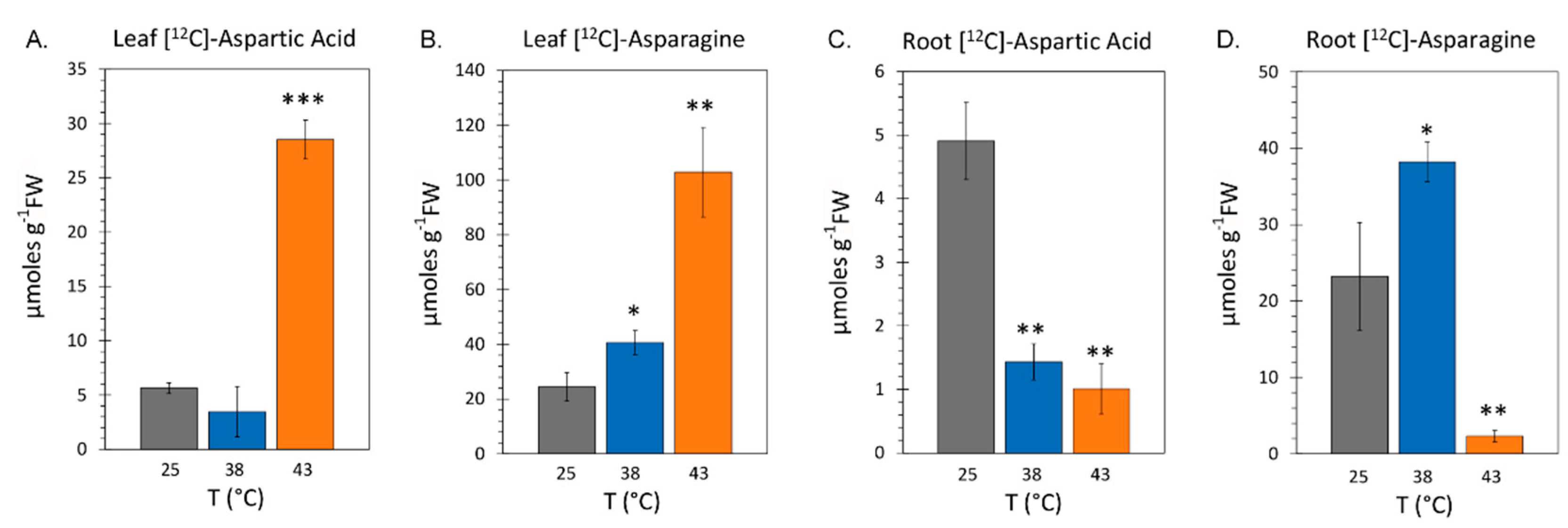
Figure 6.
The effect heat stresses of 38 °C (Panel A) and 43 °C (Panel B) spanning a timeline of exposure from 24-hr to 96-hr on total soluble leaf protein content and Rubisco protein content presented in milligrams protein per gram dry weight tissue (mg g-1DW). All protein data points are shown for N = 3-6 replicates at each timepoint. Average total protein content and Rubisco protein content for 25 °C control plants are shown as horizontal dashed lines. Average values from applied heat stresses are shown by the solid black horizontal bars with levels of significance shown as: **, P < 0.01; ***, P < 0.001.
Figure 6.
The effect heat stresses of 38 °C (Panel A) and 43 °C (Panel B) spanning a timeline of exposure from 24-hr to 96-hr on total soluble leaf protein content and Rubisco protein content presented in milligrams protein per gram dry weight tissue (mg g-1DW). All protein data points are shown for N = 3-6 replicates at each timepoint. Average total protein content and Rubisco protein content for 25 °C control plants are shown as horizontal dashed lines. Average values from applied heat stresses are shown by the solid black horizontal bars with levels of significance shown as: **, P < 0.01; ***, P < 0.001.
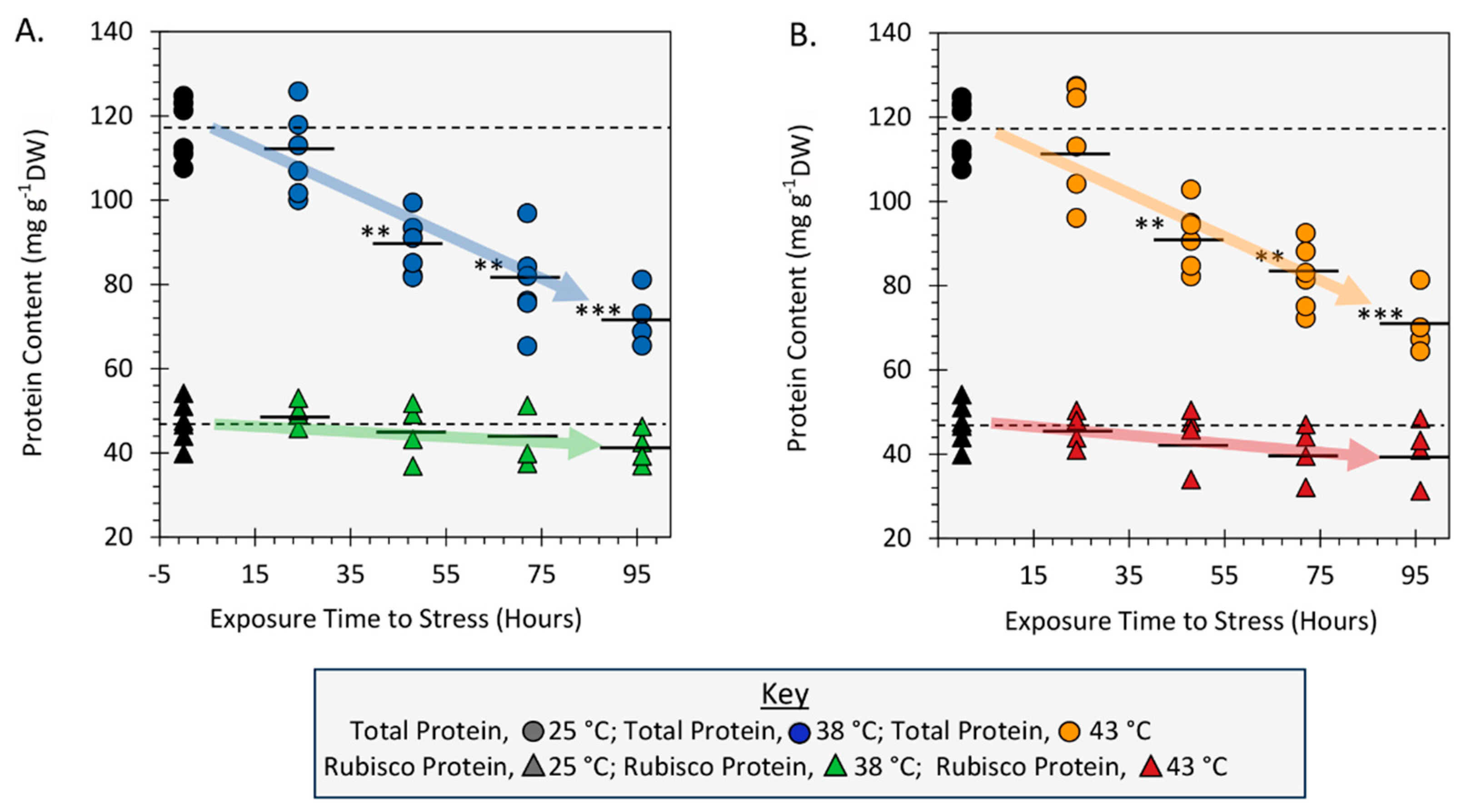
Figure 8.
Visual workflow for [11C]-radiotracer analysis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



