Submitted:
21 September 2023
Posted:
22 September 2023
You are already at the latest version
Abstract
Upper mesophilic temperature acclimation of halophilic, hydrogen-producing bacteria from salt fields was investigated in this study, along with the changes in microbial abundance during an-aerobic digestion (AD) process. Genomic approaches such as PCR-denaturing gradient gel elec-trophoresis (DGGE) and next-generation sequencing (NGS) were performed to profile the mi-crobial communities. During AD, there was a significant abundance of Halanaerobacter lacunarum at 48°C followed by the increase in hydrogen yield, signifying potential contribution from the halophile in the hydrogen production. A decrease in the dominance of H. lacunarum and Halan-aerobium fermentans at 42°C, likely due to an increase in other bacterial species, was noted but their dominance significantly increased at temperatures of 45°C and 48°C. This investigation provides valuable insights in highlighting the potential of Halanaerobium sp. and the other halo-philic bacteria to adapt under upper mesophilic temperature conditions and synthesizing hy-drogen. The findings in the present study also underscore the importance of optimizing temper-ature and pH conditions to maximize hydrogen yield during high-salt anaerobic digestion.
Keywords:
halophilic bacteria
; hydrogen production
; thermotolerance
; upper mesophilic temperature acclimation
; PCR-DGGE
; next-generation sequencing
; salt fields
1. Introduction
The contemporary energy landscape remains heavily reliant on fossil fuels, such as carbon-containing oil, coal, and natural gas. Over-reliance and overconsumption of these resources over the past century has led to a significant increase in carbon dioxide emissions. To address this environmental issue, alternative renewable energy sources, including biomass, solar photovoltaic, solar thermal, geothermal, wind, hydro, tidal, and wave power, have been explored. Among these, biomass energy offers advantages of low capital investment and relatively inexpensive operation and management. Consequently, developing efficient biomass utilization methods has become a priority to tackle energy depletion and environmental concerns.
Mitigating global warming and achieving carbon neutrality by 2050, in line with the United Nations framework convention on climate change, have been prioritized by governments across the globe. As part of these initiatives, there has been an increased emphasis on the promotion of biomass utilization, with objectives extending to the creation of recycling-oriented societies and revitalization of agriculture, forestry, and fishing communities. The Japanese government, for instance, established the Biomass Nippon Comprehensive Strategy in December 2002. The aim of this strategy is to promote the effective utilization of biomass resources. This goal is realized through the development of pertinent technologies, the provision of financial support, and the implementation of policy measures fostering sustainability. Moreover, the Kyoto Protocol, an international treaty adopted in February 2005, holds Japan accountable for the reduction of its greenhouse gas emissions. The protocol promotes the use of clean development mechanisms, endorsing renewable energy sources such as biomass. Japan’s innovative Biomass Town concept has enabled local governments to create self-sustaining circular economies. By harnessing biomass resources produced within their own regions for energy, fuel, and material production, these towns not only revitalize local communities but also contribute significantly to the reduction of carbon emissions. The New National Energy Strategy in Japan envisages a future with decreased dependency on oil within the transport sector. The strategy aims to lower this dependence to approximately 80% by 2030, achieved by encouraging the development and adoption of alternative fuels (biofuels, hydrogen, electric power), and by endorsing energy-efficient transportation technologies.
One of the most promising alternatives, especially when considering biomass, is the production of hydrogen. Biomass energy refers to the energy derived from organic materials, including plant and animal waste. As a renewable resource, biomass offers significant advantages over fossil fuels due to its abundance, carbon neutrality, and potential to mitigate greenhouse gas emissions [1,2]. Biomass feedstocks include agricultural and forestry residues, energy crops, algae, and organic waste from various industries [3,4]. Through processes such as direct combustion, gasification, pyrolysis, and anaerobic digestion, biomass can be converted into various forms of energy, including heat, electricity, and biofuels [5,6]. However, efficient conversion processes for biomass-derived energy remain an area for further investigation.
Hydrogen has several properties that make it a desirable energy carrier, such as its high energy content per unit mass (142 MJ/kg), non-toxic nature, and clean combustion, producing only water as a byproduct [7,8], [9]. Numerous conventional methodologies are used for hydrogen production, which include steam methane reforming, partial oxidation of hydrocarbons, coal gasification, and electrolysis of water. Despite their widespread utilization, these traditional techniques often necessitate high energy inputs and generate considerable greenhouse gas emissions [10,11]. This has inevitably led to an amplified interest in more sustainable means of hydrogen production, such as biological processes.
Continuing this exploration of sustainable methods, focus is placed on anaerobic decomposition - a biological procedure whereby organic matter is broken down by microorganisms in the absence of oxygen. This process results in the production of biogas (a mixture primarily composed of methane and carbon dioxide) and a nutrient-rich digestate, both of which can be productively repurposed. When contrasted with aerobic treatment, anaerobic digestion boasts numerous advantages such as lower energy consumption, diminished sludge production, and the generation of biogas that can serve as an energy resource [12,13]. This digestion process is typically characterized by four stages: hydrolysis, acidogenesis, acetogenesis, and methanogenesis, with each stage facilitated by groups of microorganisms [14,15,16]. Despite these benefits, there are still hurdles to overcome regarding the optimization of anaerobic treatment for specific substrates and applications. A particularly promising application is biohydrogen production via anaerobic digestion, which involves the metabolic conversion of organic substrates into hydrogen gas by fermentative bacteria [17]. Further investigation into this process, and the optimal conditions required for efficient hydrogen production by these microorganisms, is the primary objective of this study. Various factors, such as temperature, pH, substrate type, and microbial consortium, can influence the efficiency of hydrogen production [18,19]. Several types of substrates have been explored for biohydrogen production, including glucose, cellulose, lignocellulosic materials, and organic wastes [20,21]. Dark fermentation and photo fermentation are the two primary biological routes for biohydrogen production, with dark fermentation being more prevalent due to its ability to operate under mesophilic and thermophilic conditions [22]. However, further research is needed to enhance the efficiency and scalability of biohydrogen production through anaerobic fermentation.
Fermentation is a metabolic process in which microorganisms convert organic substrates into simpler compounds, generating energy in the form of ATP and producing various byproducts, including hydrogen gas [23,24]. Hydrogen production through fermentation is mainly attributed to the activity of hydrogenase enzymes, which catalyze the reversible reduction of protons to hydrogen [25]. Mixed culture systems and pure cultures of hydrogen-producing bacteria have been used in biohydrogen production studies [24]. The selection of appropriate microbial strains, optimization of operational parameters, and development of efficient bioreactor configurations are crucial for enhancing the biohydrogen production process [24,26]. Despite progress in this field, achieving high hydrogen yields and understanding the complex interactions between microbial consortia and operational parameters require further investigation.
Bioethanol and methane fermentation are generally considered promising biomass energy sources. However, hydrogen is particularly attractive because it produces only water during combustion, does not emit carbon dioxide, and has an energy density three times higher than that of fossil fuels. Although efficient production methods for hydrogen have not yet been established, developing such processes is of great importance.
In the context of our study, the potential of anaerobic fermentation using salt-tolerant hydrogen-producing unique avenue for inexpensive and efficient hydrogen generation. One of the most noteworthy findings is that, in environments with high salt concentrations and elevated temperatures, these bacteria actively produce hydrogen. Importantly, they also suppress the activity of methanogenic bacteria. This differential effect, where salt-tolerant hydrogen fermenting bacteria flourish while methanogenic activity is inhibited, ensures a more exclusive and potentially efficient production of hydrogen. Anaerobic hydrogen-producing bacteria rapidly generate hydrogen, which is then converted to methane by methanogenic bacteria. In typical anaerobic treatment processes, it is challenging to isolate and recover hydrogen. Nonetheless, in high-salt environments, the unique trait of these highly salt-tolerant hydrogen fermenting bacteria allows them to thrive, producing hydrogen without the interference of methane production. In contrast, methanogenic bacteria have a maximum salt tolerance of 7-8%, indicating their activity can be hindered at high salinities. This inhibition leads to the exclusive production of hydrogen.
Halophiles, microorganisms tailored for high-salt habitats, exhibit a classification spectrum based on their salt requirements, from mild to extreme halophily [27,28,29]. Their adaptation to such environments is anchored in specialized cellular mechanisms: production of compatible solutes, ion balance maintenance, and lipid membrane modification. Beyond mere survival, these adaptive strategies enable halophiles to engage in processes of industrial significance, notably within biotechnological applications such as biofuel generation, waste remediation, and enzyme extraction [27,28,30,31,32,33].
Recent research underscores the promise of halophiles in hydrogen production, an energy form gaining traction for its sustainability. In saline contexts, halophiles might subvert challenges introduced by methanogenic bacteria, amplifying standalone hydrogen yield [29,34]. In further study, biohydrogen production from glucose could be investigated to use highly salt-tolerant hydrogen-producing bacteria. The theoretical hydrogen yield in the butyric acid production pathway is 1.45 mol-H2/mol-glucose under optimal conditions (26% NaCl, 35°C, and pH 9)[29].
Our primary objective is to examine the high-temperature acclimation of salt-tolerant hydrogen fermentation flora from salt fields and analyze the flora during this process. By exploring the mechanisms behind high-temperature acclimation and microbial species interactions, we aim to contribute to the development of innovative, sustainable hydrogen production technologies. This research could help reduce fossil fuel reliance and support a sustainable, environmentally friendly energy transition.
2. Materials and Methods
The purpose of this study was to explore the biohydrogen production capabilities of the highly salt-tolerant Haloanaerobium sp. bacteria, which was isolated from soil samples collected in salt fields near Bangkok, Thailand. The primary objective was to acclimate this bacterial species to a variety of temperatures (42°C, 45°C, and 48°C) and pH conditions (7.0, 8.5, and 10.0), and in turn, investigate the potential optimizations for hydrogen production.
2.1. Preparation of Bacterial Cultures
Salt field soil samples were collected and maintained at a stable temperature of 37°C with a saturated salt concentration over a period of five years. This procedure was implemented to stimulate the growth of highly salt-tolerant bacteria, with a specific focus on the Haloanaerobium sp. The goal was to encourage the adaptation of these bacterial communities to a higher temperature range (42°C - 48°C), and concurrently observe the changes throughout this acclimation process.
2.2. Hydrogen Fermentation Experimentation
The experimental setup involved conducting hydrogen fermentation trials in a shaking incubator (model BT-100 or BT-200), utilizing 200 mL vials (Figure 1). These vials were chosen based on their ability to maintain anaerobic conditions effectively, their compact size and space-efficiency, as well as their suitability for parallel testing of various experimental conditions. The incubator was programmed to shake at a rate of 50 rpm.
2.3. Experimental Conditions: Temperature Acclimation and pH Optimization
The bacterial cultures and substrates were introduced into the vials, with the vapor phase subsequently replaced by nitrogen gas, thereby establishing an anaerobic environment. After the complete replacement of the gas phase with nitrogen was confirmed via gas chromatography, incubation commenced at the prescribed temperatures of 42°C, 45°C, and 48°C.
Following the successful acclimation of the bacterial cultures to elevated temperatures, the initial pH was then manipulated to levels of 7.0, 8.5, and 10.0 using 0.1 M solutions of sodium hydroxide and hydrochloric acid. Incubation was conducted at the three aforementioned temperatures for each pH condition.
A schematic representation of a vial during the incubation period is provided in Figure 1.
2.4. Substrate and inoculum preparation
The substrate solution contained NaHCO3 (2 g/L), K2HPO4 (2 g/L), yeast extract (1 g/L), glucose (10 g/L), NaCl (26 wt%), 2 mL/L of (NH4)2HPO4 (350 g/L), 10 mL/L of trace metal salt mixture (KCl- 75 g/L; NH4Cl-85 g/L; FeCl3・6H2O -42 g/L; MgCl2・6H2O- 81 g/L; MgSO4・7H2O - 25 g/L; CoCl2・6H2O-1.8 g/L ) and 1mL/L of CaCl2・2H2O 150 g/L [35]. Glucose served as the carbon source, yeast extract as the nitrogen source, and additional inorganic nutrients were provided. Sodium bicarbonate and dipotassium hydrogen phosphate acted as buffers to control rapid pH changes. The gaseous phase was replaced with nitrogen to maintain an anaerobic environment, and A critical step was the adjustment of salinity using NaCl to attain a saturated concentration of 26 wt% (351.35 g/L)[34]. The inoculum’s origin was soil from salt fields located near Bangkok, Thailand. This soil had been incubated under conditions of high salinity and anaerobicity for approximately five years.
2.5. Assessment of hydrogen production during anaerobic digestion process
Theoretical hydrogen production was calculated based on the butyric acid production pathway, and the hydrogen recovery rate was determined using actual and theoretical hydrogen volumes. Gas chromatography was used to continuously measure the total gas volume and composition throughout the incubation period. The volume of the generated biogas was measured using the glass syringe technique. Using a GC-8APT/TCD instrument (Shimadzu Corp., Kyoto, Japan) outfitted with a 60/80 activated charcoal mesh column (1.5 m x 3.0 mm internal diameter) and argon as the carrier gas, the samples’ H2, N2, CH4, and CO2 levels were analyzed using gas chromatography. During the analytical process, the injector, column, and detector temperatures were set at 50 °C, 60 °C, and 50 °C, respectively.
2.6. Genetic Analysis of Highly Salt-Tolerant Hydrogen-Producing Bacteria using Next Generation Sequencing (NGS)
DNA was extracted from samples using the ISOIL for Beads Beating kit (Nippon Gene), known for effectively procuring DNA from microbes with resilient cell walls. This process provided DNA samples of high quality, suitable for analyzing soil microbial community structure and biomass quantification. Following the DNA extraction, the samples then transferred to the Faculty of Medicine at Yamaguchi University in Japan for next-generation sequencing. NCBI BLAST was used to analyze the sequence. NGS was used to collect a wide range of genes from phylum to genus using 16S ribosomal ribonucleic acid (rRNA) gene amplicons using the Illumina MiSeq System. In the current study, 16S rRNA amplicon sequencing was conducted using two primer sets as follows [36]:
16S amplicon forward primer: TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG
16S amplicon reverse primer: GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC
2.6.1. PCR-DGGE and Sequencing
Following DNA extraction, the bacterial community’s composition during acclimation was analyzed using Denaturing Gradient Gel Electrophoresis (DGGE). PCR products and DGGE Marker II were loaded onto the prepared denaturing polyacrylamide gels. Post-electrophoresis at 60°C, the gels were stained with SYBR Gold Nucleic Acid Gel Stain to examine the bacterial community composition. Post-DGGE, PCR products were purified using the silica membrane method via the NucleoSpin® Gel and PCR Clean-up kit (MACHEREY-NAGEL). Sanger sequencing was performed on an ABI 3130XL instrument at Yamaguchi University’s Gene Experimental Facility, and a subsequent BLAST analysis helped assign the bands to the respective sequences.
2.6.2. Next-Generation Sequencing (NGS)
The NucleoSpin Soil kit (TaKaRa Bio Inc., Shiga, Japan) was employed to prepare DNA samples through unbiased lysis. The DNA samples were eluted with Elution Buffer SE after a streamlined process involving the addition of buffers, centrifugation, and vortexing. The samples were then analyzed for bacterial community composition and species diversity using next-generation sequencing, a service provided by the Gene Research Laboratory of Yamaguchi University.
3. Results
3.1. Thermal Responsiveness of Hydrogen Production
Hydrogen production was evaluated at three distinct temperature settings: 42°C, 45°C, and 48°C, as presented in Figure 2. The initial six days of cultivation displayed significant hydrogen gas production, which subsequently tapered off. Notably, the final hydrogen gas yields at 42°C and 45°C were almost identical, reaching 4.6 mL and 4.7 mL, equivalent to 0.185 mol H2/mol glucose and 0.19 mol H2/mol glucose, respectively. The hydrogen content hovered between 7.4 – 7.6% for both temperatures. Contrastingly, the hydrogen yield at 48°C substantially elevated, achieving 8.8 mL (0.256 mol H2/mol glucose). These observations indicate that Haloanaerobium sp. has an augmented hydrogen production capacity at elevated temperatures. Concurrently, the absence of methane production across all temperatures indicates the prevention of hydrogen consumption and complete control over methane production (Figure 3).
A notable observation from Figure 3 is the exclusive hydrogen production at 48°C, without any concurrent methane generation. This characteristic is particularly striking given the metabolic pathways involved in anaerobic digestion. Typically, hydrogen-producing bacteria convert organic matter into hydrogen and VFAs. These VFAs can then serve as substrates for methanogenic archaea, which generate methane. Thus, the complete absence of methane suggests a strong suppression or inactivity of the methanogenic microorganisms at this temperature.
3.2. Influence of pH Conditions on Hydrogen Production.
The potential impact of varying pH levels on hydrogen production was scrutinized at pH 7.0, 8.5, and 10.0 (refer to Figure 4). At the lower temperature range of 42°C and 45°C with a pH of 7.0, hydrogen gas yields were markedly superior compared to those at pH 8.5. Remarkably, a shift in this trend was observed at 48°C. Here, pH 8.5 manifested an initial lag in hydrogen production but later surpassed the yields at pH 7.0 as the cultivation progressed. In stark contrast, at pH 10.0, irrespective of the temperature setting, there was a complete absence of hydrogen production. Such patterns suggest that Haloanaerobium sp. thrives optimally and shows the best hydrogen production at neutral pH values.
3.3. PCR-DGGE and Next-Generation Sequencing (NGS) Analysis
The PCR-DGGE analysis, as shown in Figure 5 and Table 1, revealed distinct bacterial compositions at each temperature condition. Predominantly, bacteria closely related to Haloanaerobium sp., a hydrogen-producing bacterium, were observed. In addition, Halanaerobacter lacunarum was particularly pronounced at 48°C, not observed elsewhere.
The band profiles of soil samples incubated for 15 months, obtained by PCR-DGGE method, and the identification results based on the band sequence analysis are illustrated in Figure 5 and Table 1, respectively. As evident from Figure 5, the bacterial population composition differs across temperature conditions. Furthermore, Table 1 reveals that in each experimental system, bacteria closely related to Halanaerobium sp., known as hydrogen-producing bacteria, were dominant. A unique, strong band a, Halanaerobacter lacunarum, was obtained at 48°C, which was not observed at other temperatures. This indicates that H. lacunarum significantly contributes to hydrogen production at 48°C. Conversely, at 42°C and 45°C, the flora resembled the original 37°C flora, suggesting the medium-temperature bacteria became thermostable.
The 16S metagenome analysis, presented in Figure 6, identified Halanaerobacter lacunarum as the predominant species across all temperatures. Upon comparing the data from the 3-month and 1-year incubation periods, it was evident that the presence of H. lacunarum and H. fermentans decreased at 42°C, likely due to an increase in other species. In contrast, at 45°C and 48°C, the presence of H. lacunarum increased significantly. Comparing the samples after three months and one year of incubation, the dominance of the top two species, H. lacunarum and H. fermentans, decreased at 42°C, likely due to an increase in the dominance of other bacterial species. However, at 45°C and 48°C, the dominance of H. lacunarum increased by 3.8 and 6.9 percentage points, respectively. This could be attributed to H. lacunarum’s preference for temperatures above 45°C or a decrease in the number of other species surviving at higher temperatures.
Examining Figure 7 reveals intriguing patterns at the genus level within the microbial communities. Specifically, these communities are predominantly populated by halophilic bacteria such as Halanaerobacter and Halanaerobium. By contrast, genera such as Clostridium and Bacillus comprise less than 0.5% of the total microbial population. These observations highlight the critical role that environmental factors, particularly salinity and temperature, play in shaping the microbial communities involved in the anaerobic digestion process. In the present study, the halophilic genus Halanaerobacter demonstrated dominance in the microbial communities across all temperature conditions tested: 42 °C (80% abundance), 45 °C (89% abundance), and 48 °C (86% abundance). Notably, these high prevalence rates indicate that the growth and activity of Halanaerobacter are not significantly inhibited by fluctuations in temperature. Conversely, the abundance of the genus Halanaerobium appeared to be significantly affected by temperature conditions. At 42 °C, Halanaerobium represented 15% of the microbial community. However, its prevalence dropped sharply to 3.2% at 45 °C and even further to a mere 0.15% at 48 °C. These findings suggest that despite both Halanaerobacter and Halanaerobium being halophilic bacteria and thus subjected to similar salinity conditions, their respective capacities to tolerate varying temperature conditions differ. Such differences highlight the importance of considering both mesophilic and thermophilic capabilities when studying microbial communities within AD systems.
3.4. Functional Characterization of Halophilic Bacteria in High-Salt Soil Anaerobic Digestion
The primary enzymatic contributors to hydrogen production during high-salt soil anaerobic digestion (AD) were identified via the KEGG database, as summarized in Table 2.
Hydrogen production mainly occurred via acidogenesis and acetogenesis, involving the conversion of hydrolytic products into hydrogen and short-chain fatty acids (SCFAs). The enzyme xylan 1,4-beta-xylosidase (EC: 3.2.1.37), critical for the hydrolysis of hemicellulose, was abundant at 42°C but decreased significantly at higher temperatures.
The investigation also revealed a consistent presence of D-ribose-5-phosphate aldose-ketose isomerase (EC: 5.3.1.6) throughout hydrolysis. This enzyme, alongside xylan 1,4-beta-xylosidase, is produced by Halanaerobium which dominated the microbial communities in high-salt soil AD.
For acidogenesis, L-lactate dehydrogenase (EC: 1.1.1.27) was identified as a significant enzyme potentially secreted by Halanaerobium. Its inhibition might elevate hydrogen yield by maintaining pyruvate availability for conversion into formate, a primary hydrogen production substrate.
4. Discussion
This research delves deep into the role of temperature and pH on the hydrogen production capacities of the salt-tolerant bacterium, Halanaerobacter and Halanaerobium sp. The findings have pivotal implications for enhancing biogas production during upper mesophilic temperature incubation in a high salinity environment.
In the present study, the hydrogen yield was recorded at the range of 0.185 – 0.256 mol H2/mol glucose which was relatively low compared to previous study that also used high salt soil as the source of microorganisms and glucose as carbon source (0.66 – 1.15 mol H2/mol glucose at 26% salinity) [34]. However, note that the previous research required 2 years of acclimatization to achieve the high hydrogen yield, while during the early incubation, no hydrogen was produced at 26% salinity. The finding in the present study also confirmed previous result from Zheng et al. [37] that found deterioration in hydrogen yield along with the increase in salinity levels ( 0 – 30 g NaCl/L) with the lowest yield of 0.089 mol H2/mol sucrose at 30 g NaCl/L. Pierra et al. [38] also showed a significant decrease in hydrogen production at salinity levels around 19 – 29% and increased substantially at salinity above 38% (pH 8.5). Salinity at 26% level is considered a challenging environment for hydrogen-producing bacteria to grow as the known hydrogen producers, Clostridium survived at NaCl levels of 0 – 5 g NaCl/L and completely inhibited at higher salinity level [38,39], while Bacillus megaterium and Halanaerobium were mostly active in hypersaline conditions with up to 70% salinity and 150 g NaCl/L, respectively [40,41].
Temperature also plays an essential role in hydrogen production. The present study demonstrated significant surge in hydrogen production during the incubation at 48°C compared to incubation at 42°C and 45°C. This result confirmed the findings of research by Westerholm et al. [42], who discovered that the temperature range of 40–44 °C had a substantial adverse impact on the populations of both mesophilic and thermophilic microbes, which in turns, hindered biogas production. The plausible explanation for this phenomenon is that the temperature range may be over the upper limit for the growth of mesophiles but below the threshold required for the growth of thermophiles [43]. The substantial increase in hydrogen production at 48°C was also associated with the abundance of Halanaerobacter followed by the growth of Bacillus. Bacillus sp. has a thermotolerant characteristic due to its capability to grow in temperature of 20 – 90 °C with optimum pH of 7.5 and 10.5 [44].
The most important phase in the hydrogen production by microorganisms in environments with high salinity is the establishment of osmoadaptation qualities through salt accumulation in the cytoplasm and the production of low molecular weight organic molecules that act as macromolecule stabilizers [45]. Only few microorganisms were known to be able to survive high salinity environments and upper mesophilic temperature while produce hydrogen via intracellular accumulation of organic substrates pathway such as those that belong to the Bacillus, Clostridium, and Halanaerobium genus [29,38,40]. The existence of Halanaerobacter in the present study enlarges the list of microorganisms that possess osmoadaptation and thermotolerant properties. In a broader microbial perspective, the observed results bring into light the essential roles played by both Halanaerobacter and Halanaerobium in the realm of hydrogen production [37]. Particular haloarchaea were found to enhance metabolic rates during harsh environment, including Halanaerobium sp. These upper mesophilic microbes, when exposed to increased temperatures, activate specific metabolic processes, thus driving up hydrogen production. The unique thermal resilience of Halanaerobium sp. in sustaining elevated activity levels at these temperatures is of particular interest and warrants further investigation.
From a pH standpoint, optimal hydrogen production materialized under neutral conditions. However, a dip occurred at pH 8.5, plummeting further at pH 10. This parallels prior studies, underscoring the role of pH in biohydrogen production. The delayed yet magnified yield at 48°C and pH 8.5 presents an enthralling interaction of these factors, meriting comprehensive study.
Functional analysis via the KEGG database revealed hydrogen being chiefly generated via acidogenesis process by converting hydrolytic products into hydrogen and short-chain fatty acids (SCFAs), and through acetogenesis process by converting SCFAs into acetate and hydrogen [46,47]. Notably, xylan 1,4-beta-xylosidase enzyme (EC: 3.2.1.37) was found abundant in the high-salt soil AD during incubation at 42 °C while decreased significantly at 45 °C and 48 °C, signifying potential perturbations of this enzyme at the higher temperature. This enzyme, produced by several halophilic bacteria [48] and was one of the most important enzymes responsible in the hydrolysis of hemicellulose, especially in synthesizing 1,4-beta-D-xylan and H2O from D-xylose [49,50].
Additionally, we identified the prevalence of D-ribose-5-phosphate aldose-ketose isomerase (EC: 5.3.1.6), vital for the pentose phosphate pathway, throughout the hydrolysis process. This enzyme is in charge of catalyzing the isomerization between D-ribulose 5-phosphate and D-ribose 5-phosphate [51]. Both most abundant enzymes, xylan 1,4-beta-xylosidase and D-ribulose 5-phosphate and D-ribose 5-phosphate can be excreted by halophilic Halanaerobium which was found to dominate the microbial communities that were involved in the high salinity AD process, according to the KEGG database. Another enzyme that was potentially released by Halanaerobium was L-lactate dehydrogenase (EC: 1.1.1.27) which was associated with the acidogenesis process. L-lactate dehydrogenase (ldh) is the last enzyme involved in glycolysis which encourages the reciprocal conversion of pyruvate into lactate [52,53]. However, in production of hydrogen, the inhibition of ldh enzyme may lead to higher hydrogen yield due to less pyruvate being converted to lactate. Pyruvate is the key element in hydrogen production as it will be transformed into formate and use as the main substrate for hydrogen production through formate hydrogen pathway, in normal acidogenesis conditions. In deteriorated acidogenesis, pyruvate will be converted into lactate and the accumulation of lactate leads to the reduction of pH which is unfavorable for the growth of hydrogen-producing microorganisms [46,54].
Key insights include the adaptive genetic behavior of salt-tolerant, hydrogen-producing bacterial communities across temperature gradients. The behavior of Halanaerobacter lacunarum at 48°C, contrasted by Halanaerobium fermentans’s non-thermotolerance, accentuates the need for strategic microbial selection based on operational criteria.
Bioenergy’s sustainability remains a priority. Our research provides insights into the capabilities of salt-tolerant, hydrogen-fermenting bacteria across temperature and pH ranges. Optimal hydrogen yields were achieved at an initial pH of 7.0 and 42°C, but intriguingly, maximum recovery was observed at a high 48°C. This alludes to an inherent thermostability of the examined bacteria.
In-depth microbial analysis identified Halanaerobacter lacunarum as a central agent in hydrogen production at elevated temperatures. This accentuates the immense potential of extremophilic bacteria in biogas production. Furthermore, the Halanaerobium sp.’s resilience suggests vast unexplored potential, meriting further investigation into its metabolic pathways and adaptive mechanisms.
In summation, our study is more than a collection of findings. It signifies the untapped potential of extremophilic bacteria in biogas production. By modulating operational parameters, such as temperature and pH, we can maximize the efficiency of these microbial powerhouses. Our findings underscore the promise of sustainable bioenergy production using extremophilic bacteria, heralding a future of efficient, eco-friendly energy solutions.
5. Conclusions
In this study, we thoroughly explored hydrogen production by highly salt-tolerant, hydrogen-fermenting bacteria under a variety of temperature and pH conditions. We found that the optimal conditions for hydrogen recovery were an initial pH of 7.0 at a temperature of 42°C. However, intriguingly, the maximum hydrogen recovery was observed at an elevated temperature of 48°C, suggesting a potential thermostability among the bacteria studied. Our 16S metagenomic analysis pinpointed Halanaerobacter lacunarum as a crucial contributor to hydrogen production, particularly at high temperatures. Moreover, our microbial community analysis revealed that Halanaerobium sp., a mesophilic bacterium detected in our study, can perform hydrogen production even at temperatures exceeding its optimal growth temperature.
Collectively, these findings not only enhance our understanding of the potential of highly salt-tolerant hydrogen-fermenting bacteria in biogas production but also underscore the significance of optimizing operational conditions, such as temperature and pH, to maximize hydrogen yield. This work opens new avenues for bioenergy production using extremophilic bacteria, contributing to the broader efforts of sustainable and environmentally friendly energy solutions.
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, H.T.V. A.R. and T.I.; methodology, formal analysis and software, H.T.V., J.T. and G.A.W.S.; validation, J.T., T.I. and H.Y.; investigation, P.C., A.R. and T.I.; resources, T.I.; data curation, H.T.V. and G.A.W.S.; writing—original draft preparation, H.T.V.; writing—review and editing, T.I and J.T; visualization, H.T.V.; supervision, A.R., T.I. and H.Y.; project administration, T.I.; funding acquisition, T.I. and H.Y.; All authors have read and agreed to the published version of the manuscript.
Funding
JSPS KAKENHI Grant Number JP23K04089
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
This work was supported by JSPS KAKENHI Grant Number JP23K04089.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Owusu, P.A.; Asumadu-Sarkodie, S. A Review of Renewable Energy Sources, Sustainability Issues and Climate Change Mitigation. Cogent Eng. 2016, 3, 1167990. [Google Scholar] [CrossRef]
- Nguyen, V.G.; Nguyen-Thi, T.X.; Phong Nguyen, P.Q.; Tran, V.D.; Ağbulut, Ü.; Nguyen, L.H.; Balasubramanian, D.; Tarelko, W.; A. Bandh, S.; Khoa Pham, N.D. Recent Advances in Hydrogen Production from Biomass Waste with a Focus on Pyrolysis and Gasification. Int. J. Hydrogen Energy 2023. [Google Scholar] [CrossRef]
- Honorato-Salazar, J.A.; Sadhukhan, J. Annual Biomass Variation of Agriculture Crops and Forestry Residues, and Seasonality of Crop Residues for Energy Production in Mexico. Food Bioprod. Process. 2020, 119, 1–19. [Google Scholar] [CrossRef]
- Taneja, T.; Chopra, M.; Sharma, I. Bioenergy: Biomass Sources, Production, and Applications. Green Approach to Altern. Fuel a Sustain. Futur. 2023, 443–451. [Google Scholar] [CrossRef]
- Okolie, J.A.; Epelle, E.I.; Tabat, M.E.; Orivri, U.; Amenaghawon, A.N.; Okoye, P.U.; Gunes, B. Waste Biomass Valorization for the Production of Biofuels and Value-Added Products: A Comprehensive Review of Thermochemical, Biological and Integrated Processes. Process Saf. Environ. Prot. 2022, 159, 323–344. [Google Scholar] [CrossRef]
- Velvizhi, G.; Jacqueline, P.J.; Shetti, N.P.; K, L.; Mohanakrishna, G.; Aminabhavi, T.M. Emerging Trends and Advances in Valorization of Lignocellulosic Biomass to Biofuels. J. Environ. Manage. 2023, 345, 118527. [Google Scholar] [CrossRef]
- Acar, C.; Dincer, I. Review and Evaluation of Hydrogen Production Options for Better Environment. J. Clean. Prod. 2019, 218, 835–849. [Google Scholar] [CrossRef]
- Davoodabadi, A.; Mahmoudi, A.; Ghasemi, H. The Potential of Hydrogen Hydrate as a Future Hydrogen Storage Medium. iScience 2021, 24, 101907. [Google Scholar] [CrossRef]
- 2019.
- Osman, A.I.; Mehta, N.; Elgarahy, A.M.; Hefny, M.; Al-Hinai, A.; Al-Muhtaseb, A.H.; Rooney, D.W. Hydrogen Production, Storage, Utilisation and Environmental Impacts: A Review. Environ. Chem. Lett. 2022, 20, 153–188. [Google Scholar] [CrossRef]
- Abe, J.O.; Popoola, A.P.I.; Ajenifuja, E.; Popoola, O.M. Hydrogen Energy, Economy and Storage: Review and Recommendation. Int. J. Hydrogen Energy 2019, 44, 15072–15086. [Google Scholar] [CrossRef]
- Parvez, A.M.; Lewis, J.D.; Afzal, M.T. Potential of Industrial Hemp (Cannabis Sativa L.) for Bioenergy Production in Canada: Status, Challenges and Outlook. Renew. Sustain. Energy Rev. 2021, 141, 110784. [Google Scholar] [CrossRef]
- Tshemese, Z.; Deenadayalu, N.; Linganiso, L.Z.; Chetty, M. An Overview of Biogas Production from Anaerobic Digestion and the Possibility of Using Sugarcane Wastewater and Municipal Solid Waste in a South African Context. Appl. Syst. Innov. 2023, 6, 13. [Google Scholar] [CrossRef]
- Meegoda, J.N.; Li, B.; Patel, K.; Wang, L.B. A Review of the Processes, Parameters, and Optimization of Anaerobic Digestion. Int. J. Environ. Res. Public Health 2018, 15. [Google Scholar] [CrossRef]
- Maus, I.; Bremges, A.; Stolze, Y.; Hahnke, S.; Cibis, K.G.; Koeck, D.E.; Kim, Y.S.; Kreubel, J.; Hassa, J.; Wibberg, D.; et al. Genomics and Prevalence of Bacterial and Archaeal Isolates from Biogas-Producing Microbiomes. Biotechnol. Biofuels 2017, 10, 264. [Google Scholar] [CrossRef]
- Xu, R. ze; Fang, S.; Zhang, L.; Huang, W.; Shao, Q.; Fang, F.; Feng, Q.; Cao, J.; Luo, J. Distribution Patterns of Functional Microbial Community in Anaerobic Digesters under Different Operational Circumstances: A Review. Bioresour. Technol. 2021, 341, 125823. [Google Scholar] [CrossRef] [PubMed]
- El Bari, H.; Lahboubi, N.; Habchi, S.; Rachidi, S.; Bayssi, O.; Nabil, N.; Mortezaei, Y.; Villa, R. Biohydrogen Production from Fermentation of Organic Waste, Storage and Applications. Clean. Waste Syst. 2022, 3, 100043. [Google Scholar] [CrossRef]
- Pachapur, V.L.; Kutty, P.; Pachapur, P.; Brar, S.K.; Le Bihan, Y.; Galvez-Cloutier, R.; Buelna, G. Seed Pretreatment for Increased Hydrogen Production Using Mixed-Culture Systems with Advantages over Pure-Culture Systems. Energies 2019, 12, 530. [Google Scholar] [CrossRef]
- Wang, J.; Wan, W. Effect of Temperature on Fermentative Hydrogen Production by Mixed Cultures. Int. J. Hydrogen Energy 2008, 33, 5392–5397. [Google Scholar] [CrossRef]
- Bhatia, L.; Sarangi, P.K.; Singh, A.K.; Prakash, A.; Shadangi, K.P. Lignocellulosic Waste Biomass for Biohydrogen Production: Future Challenges and Bio-Economic Perspectives. Biofuels, Bioprod. Biorefining 2022, 16, 838–858. [Google Scholar] [CrossRef]
- Ayodele, D.T.; Ogunbiyi, O.D.; Akamo, D.O.; Otun, K.O.; Akinpelu, D.A.; Adegoke, J.A.; Fapojuwo, D.P.; Oladoye, P.O. Factors Affecting Biohydrogen Production: Overview and Perspectives. Int. J. Hydrogen Energy 2023, 48, 27513–27539. [Google Scholar] [CrossRef]
- Tsai, C.R.; Garcia, J.L.; Patel, B.K.; Cayol, J.L.; Baresi, L.; Mah, R.A. Haloanaerobium Alcaliphilum Sp. Nov., an Anaerobic Moderate Halophile from the Sediments of Great Salt Lake, Utah. Int. J. Syst. Bacteriol. 1995, 45, 301–307. [Google Scholar] [CrossRef]
- Bhatia, S.C. Biohydrogen. Adv. Renew. Energy Syst. 2014, 627–644. [Google Scholar] [CrossRef]
- Qu, X.; Zeng, H.; Gao, Y.; Mo, T.; Li, Y. Bio-Hydrogen Production by Dark Anaerobic Fermentation of Organic Wastewater. Front. Chem. 2022, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, W.; Ogata, H. Hydrogenases, Structure and Function. Encycl. Biol. Chem. Second Ed. 2013, 562–567. [Google Scholar] [CrossRef]
- Banu J, R.; Usman T M, M.; S, K.; Kannah R, Y.; K N, Y.; P, S.; Bhatnagar, A.; Kumar, G. A Critical Review on Limitations and Enhancement Strategies Associated with Biohydrogen Production. Int. J. Hydrogen Energy 2021, 46, 16565–16590. [Google Scholar] [CrossRef]
- De Lourdes Moreno, M.; Pérez, D.; García, M.T.; Mellado, E. Halophilic Bacteria as a Source of Novel Hydrolytic Enzymes. Life 2013, 3, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Mokashe, N.; Chaudhari, B.; Patil, U. Operative Utility of Salt-Stable Proteases of Halophilic and Halotolerant Bacteria in the Biotechnology Sector. Int. J. Biol. Macromol. 2018, 117, 493–522. [Google Scholar] [CrossRef] [PubMed]
- Taroepratjeka, D.A.H.; Imai, T.; Chairattanamanokorn, P.; Reungsang, A. Extremely Halophilic Biohydrogen Producing Microbial Communities from High-Salinity Soil and Salt Evaporation Pond. Fuels 2021. [Google Scholar] [CrossRef]
- Karray, F.; Ben Abdallah, M.; Kallel, N.; Hamza, M.; Fakhfakh, M.; Sayadi, S. Extracellular Hydrolytic Enzymes Produced by Halophilic Bacteria and Archaea Isolated from Hypersaline Lake. Mol. Biol. Rep. 2018, 45, 1297–1309. [Google Scholar] [CrossRef]
- Rohban, R.; Amoozegar, M.A.; Ventosa, A. Screening and Isolation of Halophilic Bacteria Producing Extracellular Hydrolyses from Howz Soltan Lake, Iran. J. Ind. Microbiol. Biotechnol. 2009, 36, 333–340. [Google Scholar] [CrossRef]
- Cayol, J.L.; Ollivier, B.; Patel, B.K.C.; Prensier, G.; Guezennec, J.; Garcia, J.L. Isolation and Characterization of Halothemothrix Orenii Gen. Int. J. Syst. Bacteriol. 1994, 44, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Sudiartha, G.A.; Imai, T.; Mamimin, C.; Reungsang, A. Effects of Temperature Shifts on Microbial Communities and Biogas Production: An In-Depth Comparison. Fermentation 2023, 9. [Google Scholar] [CrossRef]
- Taroepratjeka, D.A.H.; Imai, T.; Chairattanamanokorn, P.; Reungsang, A. Investigation of Hydrogen-Producing Ability of Extremely Halotolerant Bacteria from a Salt Pan and Salt-Damaged Soil in Thailand. Int. J. Hydrogen Energy 2019. [Google Scholar] [CrossRef]
- Zhou, W.; Imai, T.; Ukita, M.; Sekine, M.; Higuchi, T. Triggering Forces for Anaerobic Granulation in UASB Reactors. Process Biochem. 2006, 41, 36–43. [Google Scholar] [CrossRef]
- Sudiartha, G.A.W.; Imai, T. An Investigation of Temperature Downshift Influences on Anaerobic Digestion in the Treatment of Municipal Wastewater Sludge. J. Water Environ. Technol. 2022, 20, 154–167. [Google Scholar] [CrossRef]
- Zheng, X.J.; Zheng, Y.M.; Yu, H.Q. Influence of NaCl on Hydrogen Production from Glucose by Anaerobic Cultures. Environ. Technol. 2005, 26, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Pierra, M.; Trably, E.; Godon, J.-J.; Bernet, N. Fermentative Hydrogen Production under Moderate Halophilic Conditions. Int. J. Hydrogen Energy 2014, 39, 7508–7517. [Google Scholar] [CrossRef]
- Xiang, L.; Achen, W.; Wenjie, W.; Xuesong, L.; Liuxia, Z.; Guangwen, H.; Daqing, H.; Wenli, C.; Qiaoyun, H. High Salinity Inhibits Soil Bacterial Community Mediating Nitrogen Cycling. Appl. Environ. Microbiol. 2021, 87, e01366–21. [Google Scholar] [CrossRef]
- Liu, H.; Wang, G. Hydrogen Production of a Salt Tolerant Strain Bacillus Sp. B2 from Marine Intertidal Sludge. World J. Microbiol. Biotechnol. 2012, 28, 31–37. [Google Scholar] [CrossRef]
- Kivistö, A.; Santala, V.; Karp, M. Hydrogen Production from Glycerol Using Halophilic Fermentative Bacteria. Bioresour. Technol. 2010, 101, 8671–8677. [Google Scholar] [CrossRef]
- Westerholm, M.; Isaksson, S.; Karlsson Lindsjö, O.; Schnürer, A. Microbial Community Adaptability to Altered Temperature Conditions Determines the Potential for Process Optimisation in Biogas Production. Appl. Energy 2018. [Google Scholar] [CrossRef]
- Tezel, U.; Tandukar, M.; Hajaya, M.G.; Pavlostathis, S.G. Transition of Municipal Sludge Anaerobic Digestion from Mesophilic to Thermophilic and Long-Term Performance Evaluation. Bioresour. Technol. 2014. [Google Scholar] [CrossRef]
- Kanthi Kiran, K.; Chandra, T.S. Production of Surfactant and Detergent-Stable, Halophilic, and Alkalitolerant Alpha-Amylase by a Moderately Halophilic Bacillus Sp. Strain TSCVKK. Appl. Microbiol. Biotechnol. 2008, 77, 1023–1031. [Google Scholar] [CrossRef]
- Moshabaki Isfahani, F.; Tahmourespour, A.; Hoodaji, M.; Ataabadi, M.; Mohammadi, A. Characterizing the New Bacterial Isolates of High Yielding Exopolysaccharides under Hypersaline Conditions. J. Clean. Prod. 2018, 185, 922–928. [Google Scholar] [CrossRef]
- Lim, J.W.; Park, T.; Tong, Y.W.; Yu, Z. Chapter One - The Microbiome Driving Anaerobic Digestion and Microbial Analysis. In; Li, Y., Khanal, S.K.B.T.-A. in B., Eds.; Elsevier, 2020; Vol. 5, pp. 1–61 ISBN 2468-0125.
- O-Thong, S.; Mamimin, C.; Kongjan, P.; Reungsang, A. Chapter Six - Two-Stage Fermentation Process for Bioenergy and Biochemicals Production from Industrial and Agricultural Wastewater. In; Li, Y., Khanal, S.K.B.T.-A. in B., Eds.; Elsevier, 2020; Vol. 5, pp. 249–308 ISBN 2468-0125.
- Hung, K.-S.; Liu, S.-M.; Tzou, W.-S.; Lin, F.-P.; Pan, C.-L.; Fang, T.-Y.; Sun, K.-H.; Tang, S.-J. Characterization of a Novel GH10 Thermostable, Halophilic Xylanase from the Marine Bacterium Thermoanaerobacterium Saccharolyticum NTOU1. Process Biochem. 2011, 46, 1257–1263. [Google Scholar] [CrossRef]
- Bhalla, A.; Bansal, N.; Kumar, S.; Bischoff, K.M.; Sani, R.K. Improved Lignocellulose Conversion to Biofuels with Thermophilic Bacteria and Thermostable Enzymes. Bioresour. Technol. 2013, 128, 751–759. [Google Scholar] [CrossRef]
- Cheng, H.-H.; Whang, L.-M. Resource Recovery from Lignocellulosic Wastes via Biological Technologies: Advancements and Prospects. Bioresour. Technol. 2022, 343, 126097. [Google Scholar] [CrossRef]
- Chen, J.; Wu, H.; Zhang, W.; Mu, W. Ribose-5-Phosphate Isomerases: Characteristics, Structural Features, and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 6429–6441. [Google Scholar] [CrossRef]
- Naik, R.A.; Rawat, D.; Ahi, J.D.; Koiri, R.K. Ameliorative Effect of Piracetam on Emamectin Benzoate Induced Perturbations in the Activity of Lactate Dehydrogenase in Murine System. Adv. Redox Res. 2021, 3, 100019. [Google Scholar] [CrossRef]
- Andreeßen, C.; Wolf, N.; Cramer, B.; Humpf, H.-U.; Steinbüchel, A. In Vitro Biosynthesis of 3-Mercaptolactate by Lactate Dehydrogenases. Enzyme Microb. Technol. 2018, 108, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lu, Y.; Wang, L.; Zhang, C.; Yang, C.; Xing, X. Disruption of Lactate Dehydrogenase and Alcohol Dehydrogenase for Increased Hydrogen Production and Its Effect on Metabolic Flux in Enterobacter Aerogenes. Bioresour. Technol. 2015, 194, 99–107. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram of a vial during incubation.
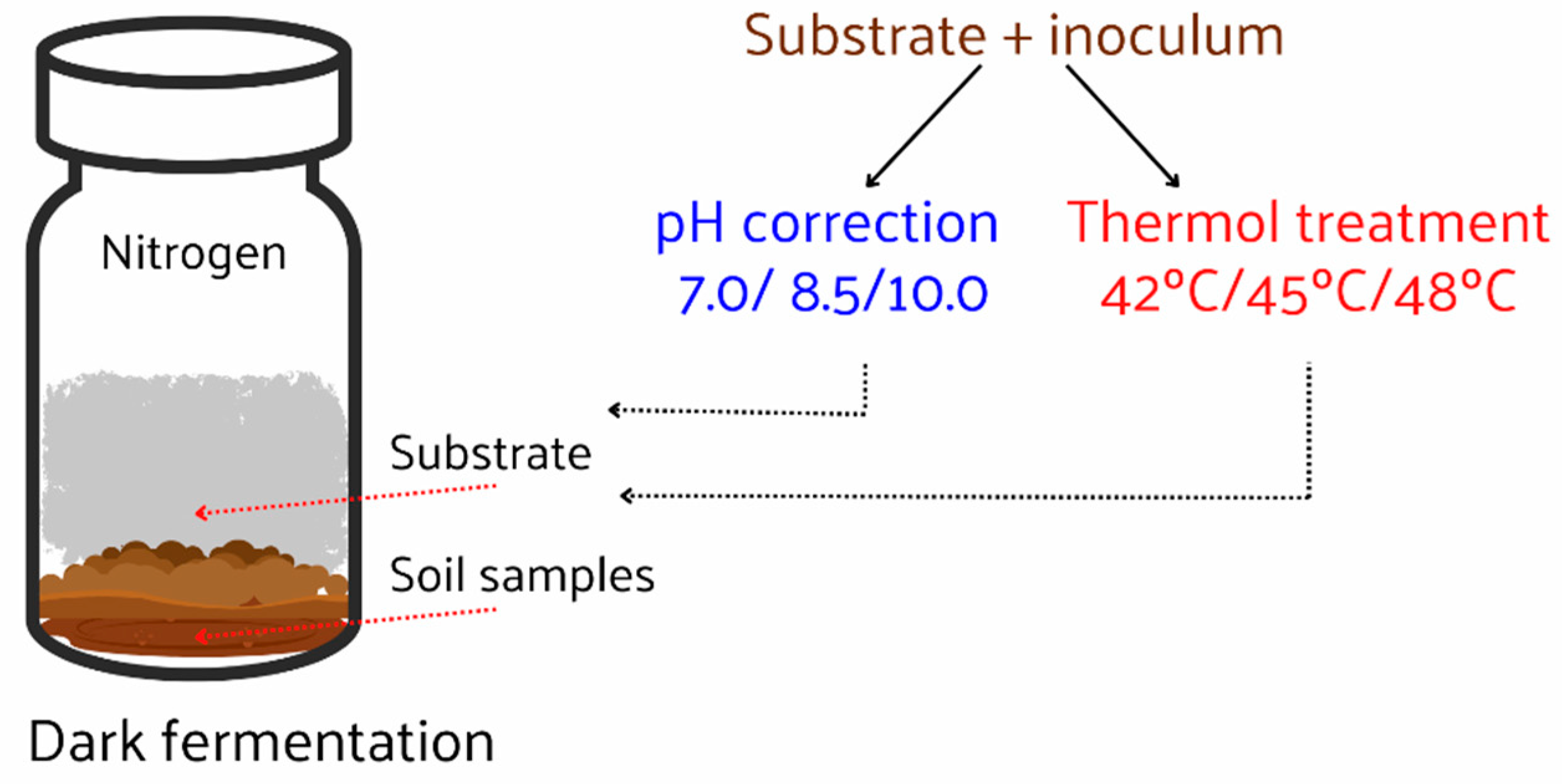
Figure 2.
Observation of Hydrogen Production at Different Temperatures.
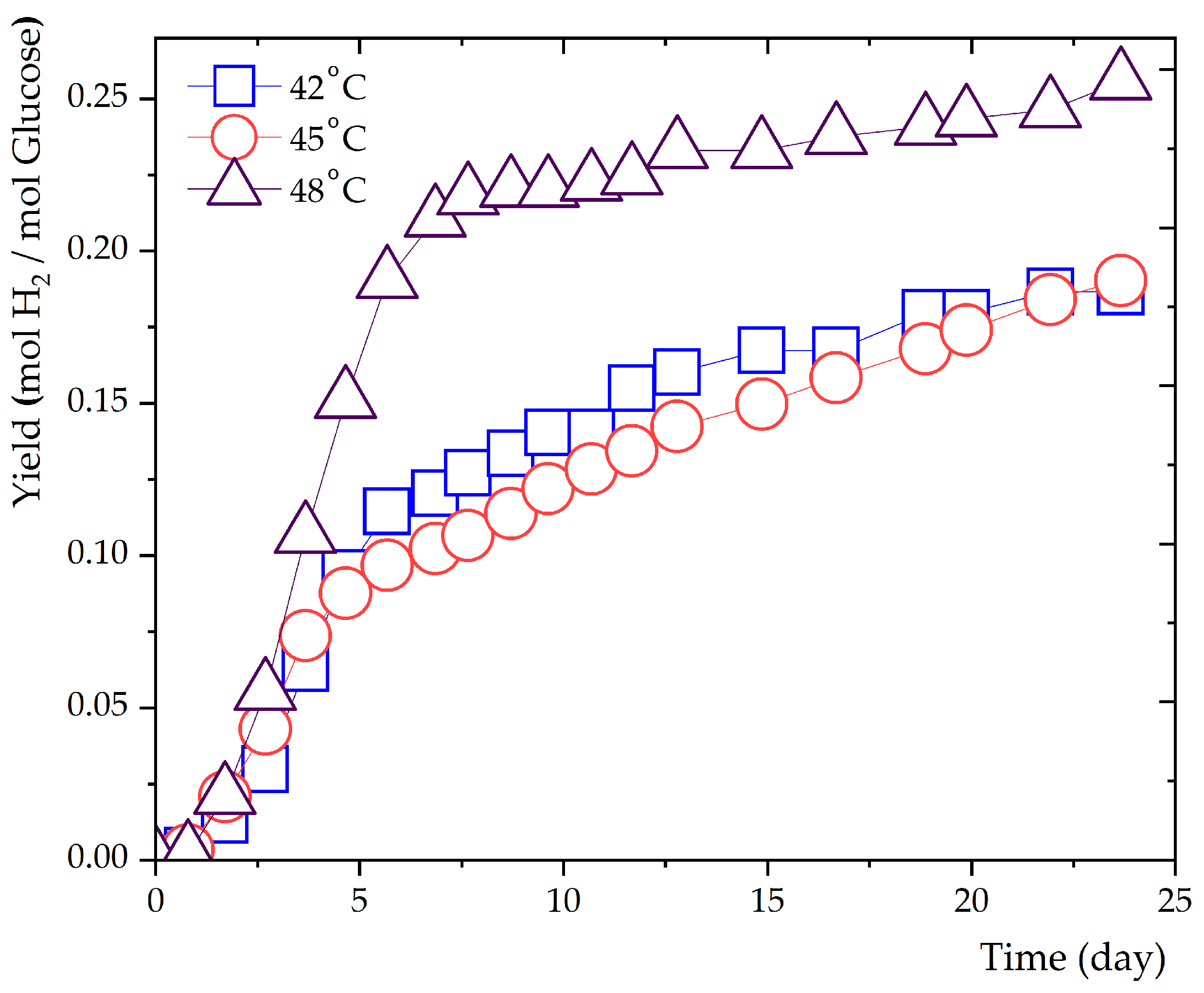
Figure 3.
Observation of hydrogen production and cumulative gas generation at 48 °C.
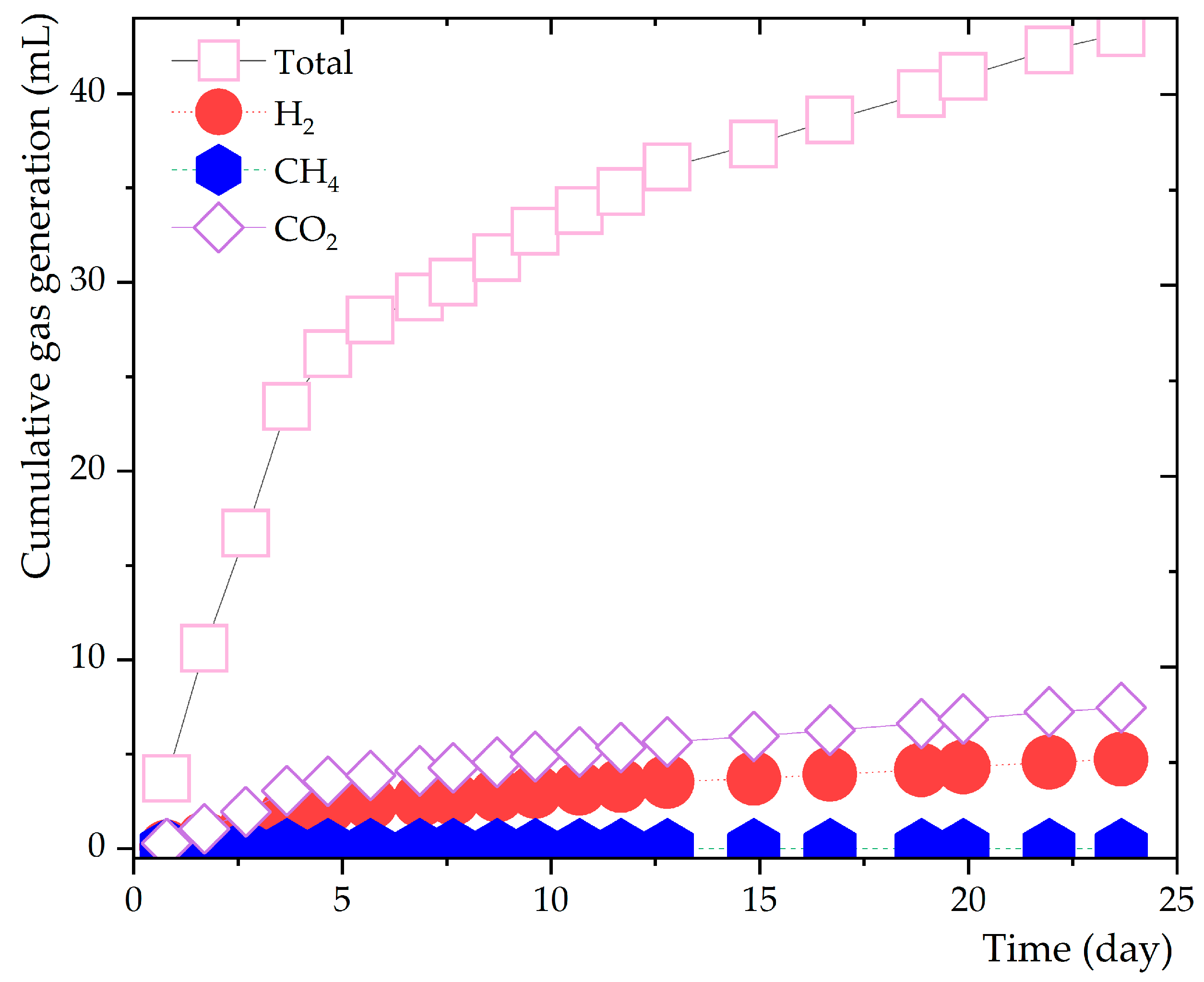
Figure 4.
Hydrogen Integration Across Various pH Conditions and Temperatures.
Figure 5.
PCR-DGGE Band Profiles for Soil Samples.
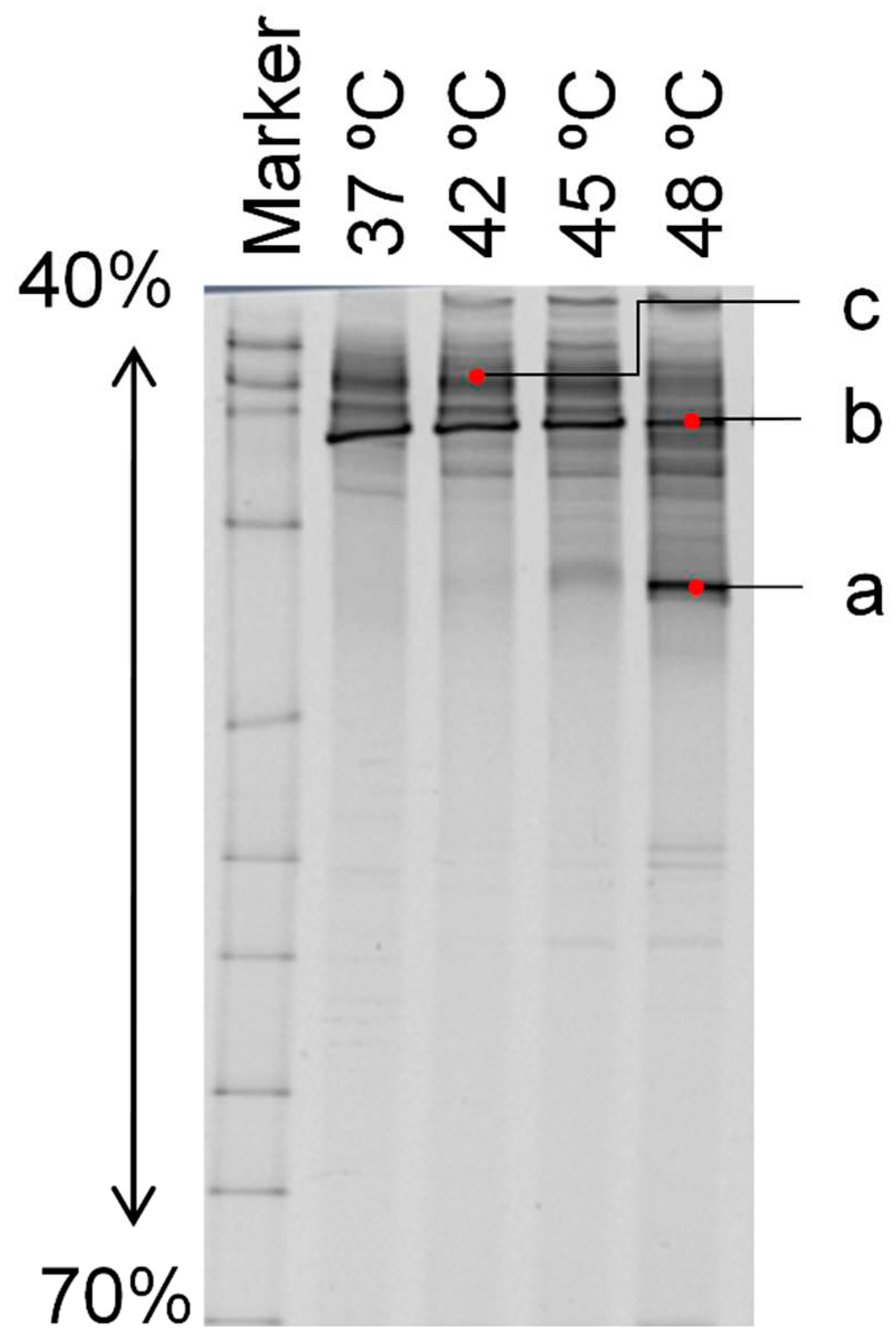
Figure 6.
Three dominant species in the samples after three months, and one year incubation.
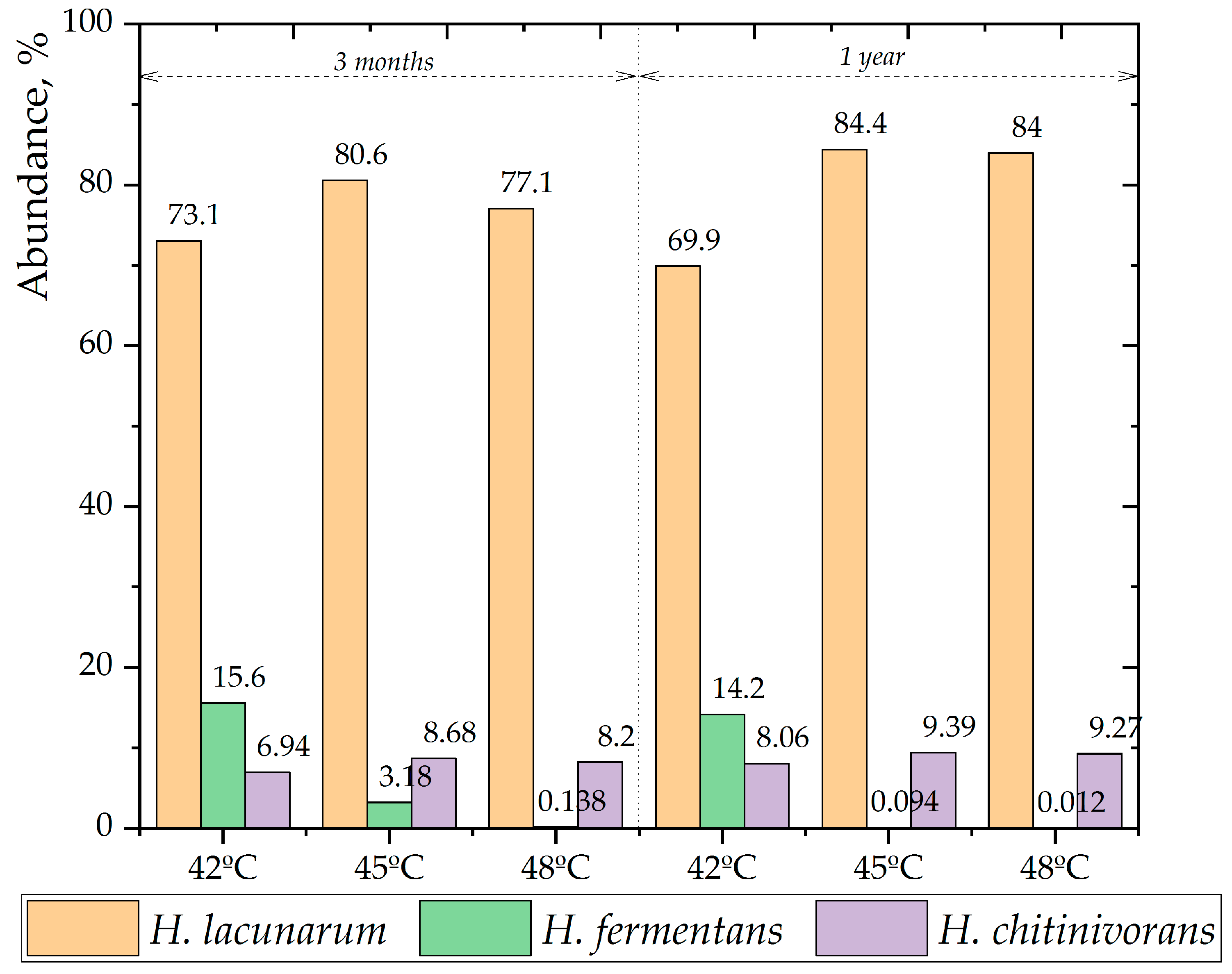
Figure 7.
Relative abundance of microbial communities in three different temperature treatment to genus level.
Figure 7.
Relative abundance of microbial communities in three different temperature treatment to genus level.
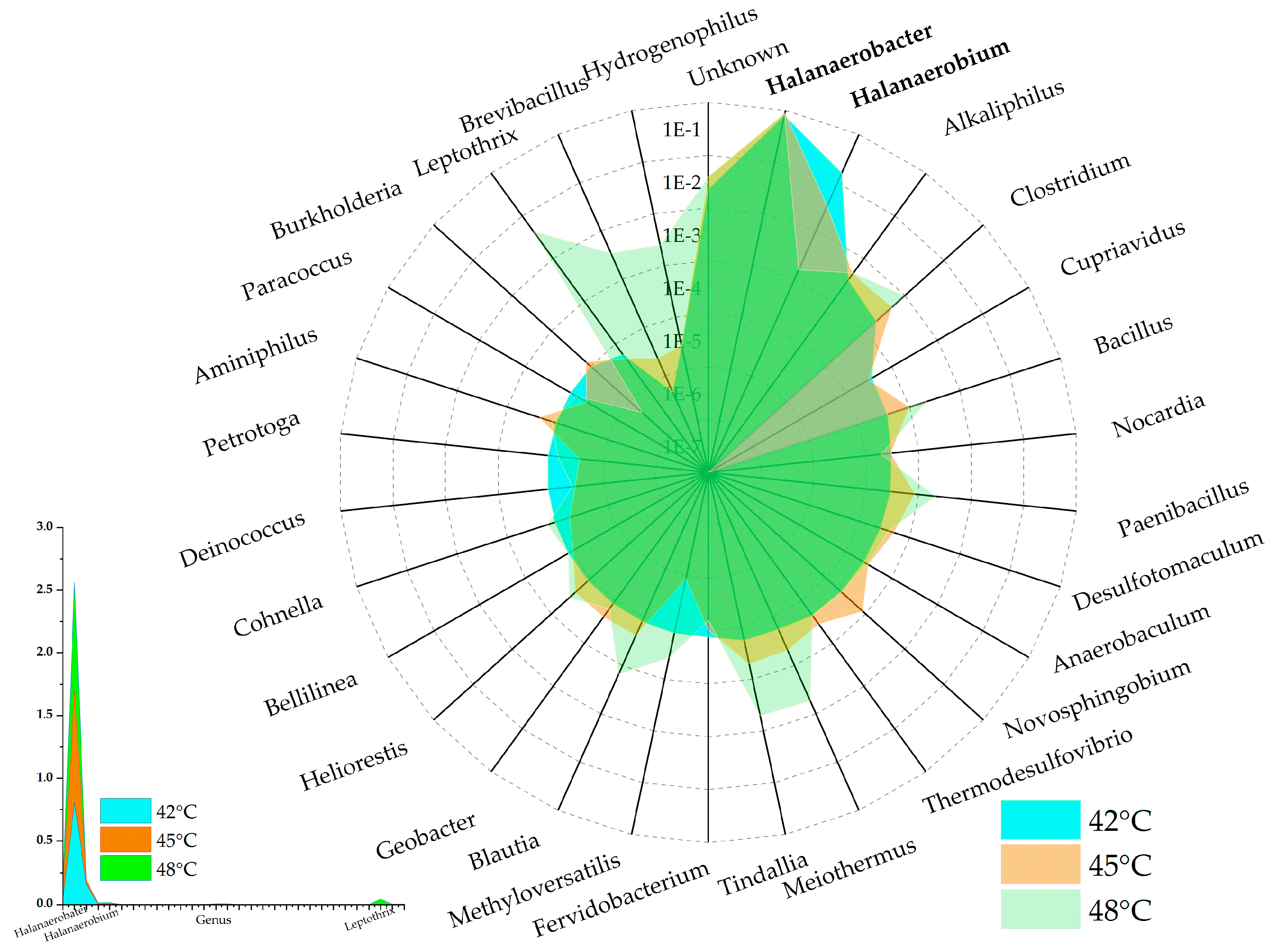

Table 1.
Sequence Analysis Identification Results for Soil Samples.
| Band | Related species | Homologous |
|---|---|---|
| a | Halanaerobacter lacunarum | 100% |
| b | Halanaerobiumfermentans | 100% |
| c | Halanaerobium saccharolyticum subsp. Senegalense | 94.29% |
Table 2.
Key Enzymes and Reactions in High-Salt Soil Anaerobic Digestion.
| AD Phase | Enzymes (EC) | Reaction | Relative abundance (%) | ||
|---|---|---|---|---|---|
| 42 °C | 45 °C | 48 °C | |||
| Hydrolysis | Xylan 1,4-beta-xylosidase(EC: 3.2.1.37) | D-Xylose + 1,4-beta-D-Xylan <=> 1,4-beta-D-Xylan + H2O | 16.43 | 4.66 | 2.44 |
| D-ribose-5-phosphate aldose-ketose isomerase (EC: 5.3.1.6) | D-Ribose 5-phosphate <=> D-Ribulose 5-phosphate | 16.30 | 4.50 | 6.91 | |
| Acidogenesis | L-lactate dehydrogenase (EC: 1.1.1.27) | (S)-Lactate + NAD+ <=> Pyruvate + NADH + H+ | 16.21 | 4.33 | 2.47 |
| Methanogenesis | CoB-CoM Ferredoxin: H2 reductase (EC: 1.8.98.5) | Coenzyme M 7-mercaptoheptanoylthreonine-phosphate heterodisulfide + Dihydromethanophenazine <=> Coenzyme B + Coenzyme M + Methanophenazine | 0.328 | 0.709 | 1.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



