
Submitted:
20 September 2023
Posted:
21 September 2023
You are already at the latest version
Abstract
Cancer is a complex and heterogeneous disease, influenced by various factors that affect its progression and response to treatment. Although histopathological diagnosis is crucial for identifying and classifying cancer, it may not accurately predict the disease's development and evolution in all cases. To address this limitation, liquid biopsy has emerged as a valuable tool, enabling a more precise and non-invasive analysis of cancer. Liquid biopsy offers the ability to detect tumor DNA fragments, circulating tumor cells, and exosomes released by cancer cells into the bloodstream. Exosomes, in particular, have attracted significant attention in cancer research due to their specific protein composition, which can provide valuable insights into the disease. The protein profile of exosomes often differs from that of normal cells, reflecting the unique molecular characteristics of cancer. Analyzing these proteins can help identify cancer-associated markers that play critical roles in tumor progression, invasion, and metastasis. Ongoing research and clinical validation are essential to advance and effectively utilize protein biomarkers in cancer. Nevertheless, their potential to improve diagnosis and treatment is highly promising. This review discuss several exosome proteins of interest in breast cancer, particularly focusing on studies conducted in mammary tissue and cell lines in humans and experimental animals. Unfortunately, the studies conducted in the canine species are scarce. This emphasis sheds light on the limited research available in this field. Additionally, we present a curated selection of studies that have explored exosomal proteins as potential biomarkers, aiming to achieve the aforementioned benefits in breast cancer diagnosis, prognosis, monitoring, and treatment.
Keywords:
extracellular vesicles
; exosomes
; proteomics
; signaling molecules
; biomarkers
; mammary neoplasia
Introduction
In the realm of cancer research, protein biomarkers have emerged as powerful tools that unlock a deeper understanding of this complex disease. These molecular signposts found in the body offer invaluable insights into cancer detection, diagnosis, prognosis, and treatment. Their great importance relies on their ability to revolutionize personalized medicine, to enhance patient outcomes, and to pave the way for targeted therapies. In this review, we delve into the mammary cancer protein biomarkers described for human and dogs through a proteomic standpoint.
In most of the past four decades and during the most recent years (2010-2019) mammary cancer incidence rates have risen by 0.5% annually[1]. Female breast cancer (BC) has surpassed lung cancer as the most commonly diagnosed cancer in the vast majority of countries, with an estimated 2.3 million new cases yearly, accounting for 1 in 4 cancer cases and for 1 in 6 cancer deaths[2].
In veterinary medicine, canine mammary tumors (CMT) are the most common cancer in female dogs[3], although it occurs mainly in those countries where early female sterilization is not a current practice. Due to the high similarity of CMT to human breast cancers, it represents an excellent experimental model to BC. Also, human BC biomarkers are frequently detectable in cases of CMT[4].
The history of BC is full of attempts to understand the wily nature of this hormone-responsive cancer [5]. A hereditary predisposition may influence screening and follow-up recommendations for high-risk patients, but a specific predisposing gene is identified in less than 30% of cases[6]. On the other hand, there is an important role for microenvironmental and epigenetic changes as well [7].
Traditional methods of cancer detection such as tissue biopsy are not comprehensive enough to capture the entire genomic landscape of breast tumors. However, with the introduction of novel techniques, the application of liquid biopsy has been enhanced, enabling the improvement of various aspects of breast cancer management [8].
Liquid biopsy can be defined as the sampling of components of tumor cells that are released from a tumor and/or metastatic deposits into the biological substances, such as the blood. Such components include circulating tumor cells (CTCs), circulating tumor DNA (ctDNA) or circulating tumor RNA (ctRNA), platelets, and exosomes [9]. Relative to direct tumor biopsies, the easy-to-obtain nature of bioliquids makes them an attractive alternative source and exosomes released from heterogeneous cancer cells in biofluids could provide the potential information of tumors [10]. Therefore, liquid biopsies in breast cancers have yielded promising results, especially in the areas of monitoring treatment response and predicting disease progression or relapse [11].
As one of the most prevalent and devastating diseases affecting women worldwide, early detection and accurate diagnosis of mammary cancer are crucial for improving patient outcomes and implementing effective treatment strategies. Therefore, extensive research efforts have been dedicated to identifying reliable biomarkers that can aid in the early detection, monitoring and treatment.
Breast Cancer biomarkers
The diagnosis and prognosis of the metastatic potential of mammary cancer is currently based on histopathology after surgical excision. However, no reliable histological criteria for metastatic potential are available before metastasis actually occurs and there are major discrepancies between the histological prognosis and the actual outcome of the disease [12]. The enormous heterogeneity of breast cancer represents the major hurdle for accurate diagnosis and curative therapy. Currently, a number of molecular biomarkers have been used in standard clinical practice, including the hormone receptors for breast cancer subtyping and several genes involved in genome maintenance for prediction of breast cancer susceptibility [13,14].
Blood-based biomarkers may offer an alternative non-invasive strategy to improve cancer screening [15]. Promising candidate biomarkers such as proteins, autoantibodies, miRNAs, nucleic acid methylation, metabolites, lipids and extracellular vesicles have shown great potential for detecting BC, including detection at the pre-invasive and early stages of the disease [15].
More recently, clinical cancer proteomics aims at the identification of biomarkers and has resulted in a wealth of information and in the discovery of several new potential tumor biomarkers [16]. Also, as the early identification, diagnosis and treatment of breast cancer evolve, we will move to a time when each patient and their cancer can be assessed to determine unique patient-specific (personalized) approaches to therapy [17].
Extracellular vesicles in cancer intercellular communication
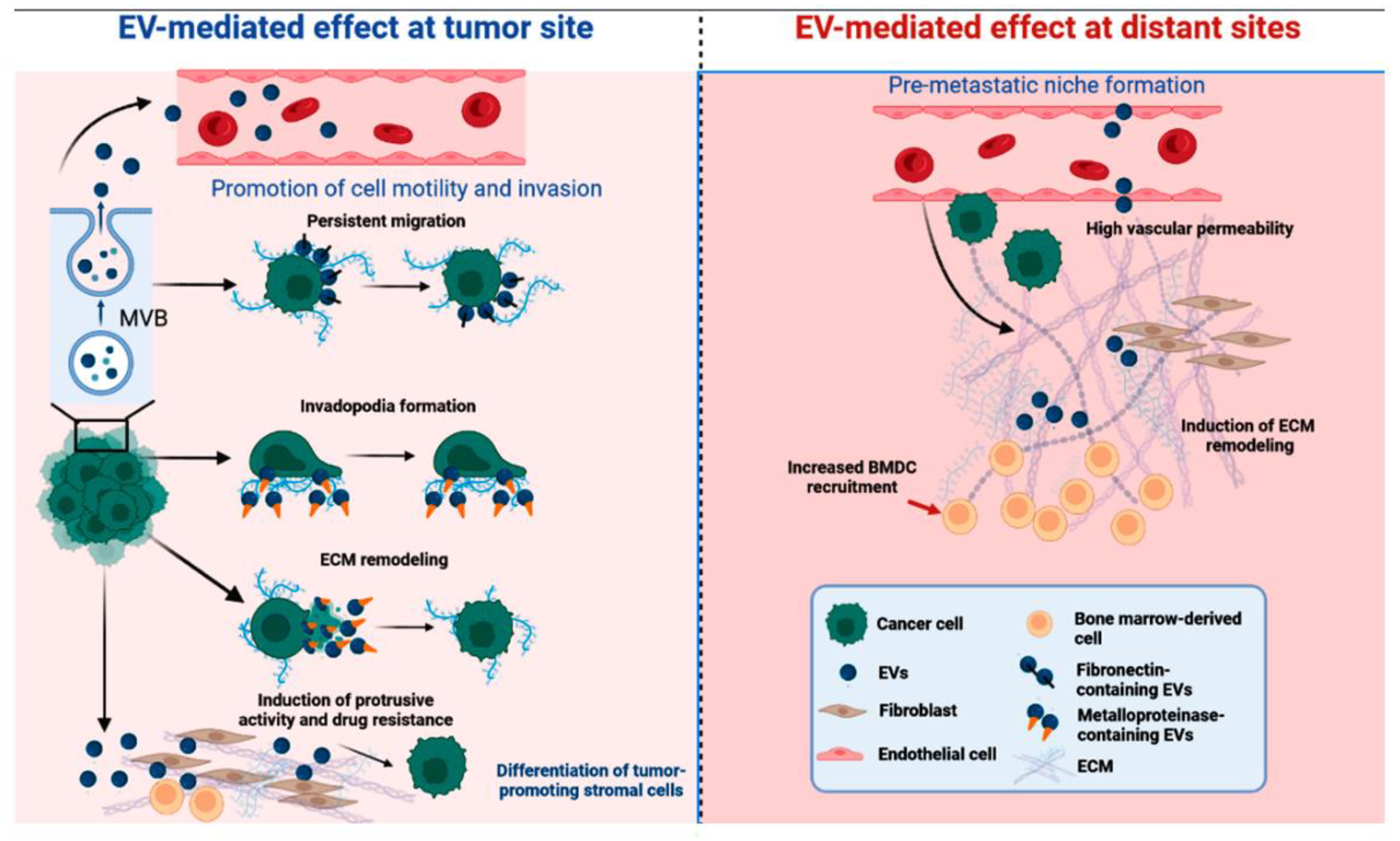
Extracellular vesicles (EVs) derived from cancer cells have received a good deal of attention because of their ability to participate in the horizontal transfer of signaling proteins between cancer cells and to contribute to their invasive activity [18]. EVs, also called exosomes, are small membrane nanoparticles released by tissue cells that contribute to cancer progression and metastases by transporting biologically significant nucleic acids and proteins, including microRNA, mRNA, non-coding RNA, DNA, transcription factors, integrins, signaling molecules and growth factors [18,19]. Small EVs from tumor cells, were demonstrated to fuse preferentially with their resident destination, preparing the niche for metastasis in a process that has been intensely investigated by cancer researchers and seems to be highly related to exosomal integrins. Clinical data have indicated that exosomal integrins could be used to predict organ-specific metastasis [20,21](Figure 1).
EVs shuttle cargo proteins that regulate tumorigenesis and show diagnostic and prognostic potential.
Proteins are among EVs cargo molecules, and they can regulate many processes like the tropism of cells to receive the vesicles, binding and activation of receptors on recipient cells and also the promotion of some reactions inside the EVs [22]. However, EVs do not contain a random array of cargo proteins, but rather a specific array. The range of the EVs biocargo so far reported in the literature has been cataloged in EVpedia (http://evpedia.info), Vesiclepedia (http://www.microvesicles.org), and ExoCarta (http://www.exocarta.org) [23].
Mammary canine EVs proteomic studies are scarce
The identification of exosomal proteins involved in tumor signaling through proteomics has recently been a promising field of research. Despite the growing number of studies on exosomal proteomics in humans, particularly in in vitro studies using cell lines, research on proteomics of exosomes in the canine species is still scarce, especially in mammary carcinoma. So far, the literature has reported proteins found in tissues and body fluids, but not specifically inside extracellular vesicles.
For example, using 2D-DIGE and MALDI-TOF-MS, Klopfleisch and colleagues (2010) [24] identified 21 proteins with significant changes in protein expression between metastasizing and non-metastasizing canine mammary carcinomas. Up-regulated proteins in metastatic carcinomas included proliferating cell nuclear antigen (PCNA), ferritin light chain (FTL), bomapin, tropomyosin 3 (TPM3), thioredoxin-containing domain containing 5 (TXNDC5), adenosin deaminase (ADA), ornithine aminotransferase (OAT), coronin 1A (CORO1A), (RANBP1) RAN-binding protein 1,3-phosphoglycerate dehydrogenase, and eukaryotic translation elongation factor 1 (eEF1).
One year later, Suárez-Bonnet and colleagues (2011) [25] examined the expression of 14-3-3 σ, a protein related to cell cycle regulation, in normal, dysplastic and neoplastic canine mammary tissue, to assess the capacity of this protein to act as marker of myoepithelial cells (MECs). The findings indicated that 14-3-3 σ protein is present in both normal and neoplastic canine mammary tissue, with high expression of this antigen in MECs. These results suggested this protein may have a role in enhancing the spreading capacity of canine mammary tumors and can be considered both a prognostic biomarker and a therapeutic target.
Later, Jagarlamudi and colleagues (2014) [26] analyzed another cell cycle-related protein, specifically serum thymidine kinase 1 (sTK1). Its activity and protein levels were significantly higher in CMT than in healthy dogs.
Four years later, Jena and colleagues (2019) [27] reported the detection of Baculoviral inhibitor of apoptosis repeat containing-5 (BIRC5 or survivin) protein biomarker in dog serum, in primary culture of canine mammary tumor cells, as well as in REM-134 canine mammary cancer cell line. They found that serum BIRC5 levels, a protein that inhibits cell death, were 109.83 ± 8.10 pg/mL in dogs with CMT as compared to 44.71 ± 2.61 pg/mL and 30.23 ± 1.32 pg/mL, respectively, in case of dogs with non-cancerous diseases and healthy subjects.
Fortunately, study attendance seemed to improve afterwards. Fhaikrue and colleagues (2020) [28] designed a study to identify protein expression on canine mammary tumors (CMTs) using primary cell culture from benign mixed tumor, simple carcinoma, complex carcinoma, and normal mammary gland through a proteomic approach. Cytokeratin 5 (CK5) and transketolase (TKT), an enzyme that connects pentose phosphate pathway to glycolysis, were identified in benign mixed tumor cells and complex carcinoma cells. In contrast, cytokeratin 18 (CK18) and pyruvate kinase (PKM), the enzyme from last step of glycolysis, were identified in simple carcinoma cells. Moreover, alpha-2-HS-glycoprotein tumor antigen was identified specifically in complex carcinoma cells, while ATP-dependent 6-phosphofructokinase platelet type and elongation factor 2 proteins were observed in benign cells. Such alterations in metabolism-related genes indicate that CMTs undergo metabolic reprogramming from benign to malignant tumors which supplies the demands of proliferating cells, a phenomenon commonly describe in several cancer, including breast cancer [29].
In the same year, Park and colleagues (2020) [30] published their study focused on discovering aggressiveness biomarkers of canine mammary tumors using proteomics on canine normal and cancer plasma samples. They identified 54 proteins more elevated in cancer than in normal plasma, including SERPING1, SERPINA6, and LCAT (lecithin-cholesterol acyltransferase). This study revealed the plasma protein LCAT, a plasma enzyme that esterifies cholesterol, as a biomarker for advanced breast cancer as well as mammary tumors undergoing metastasis.
A little while later, through the comparison of the proteomic profile of canine intratumor cell populations in different degrees of differentiation, Cordeiro and colleagues (2021) [31] demonstrated that the malignant phenotype may have arisen as a consequence of alterations in the expression of key proteins such as FNDC1, A1BG, CANX, HSPA5, and PDIA3 and that most of these variations might be involved in tumor evasion against inflammatory cells, thus facilitating cancer spreading.
Also, the study of Yuan and colleagues (2021) [32] revealed that anterior gradient protein 2 (AGR2), a chaperone and p53 inhibitor involved in cell migration, transformation and metastasis, is overexpressed in canine mammary malignant tumor (MMT) tissues, and elevated levels of extracellular AGR2 in sera of MMT dogs are significantly associated with progression and remote metastasis of MMT and an unfavorable overall patient’s survival rate. Hence, they concluded serum eAGR2 level is significantly associated with an adverse outcome of MMT dogs and holds a predictive potential in MMT prognosis.
As previously documented, research on mammary tumor proteins in the exosomes of dogs is currently non-existent. As a result, there is a pressing need for further investigation in this area. Despite the extensive research conducted on mammary tumors in dogs, there has been relatively little focus on the proteins present in the exosomes derived from these tumors. Therefore, the comprehensive characterization and exploration of the protein cargo within dog mammary tumor exosomes remain largely unexplored.
EVs proteomics may identify novel breast cancer biomarkers
Extensive efforts have been dedicated to identifying reliable biomarkers that can aid in the early detection and monitoring of human mammary cancer. Proteomic analysis of exosomes has gained significant attention in mammary cancer research due to the abundance and diversity of proteins encapsulated within these vesicles. By characterizing the protein composition of exosomes derived from mammary cancer cells, researchers can uncover unique molecular signatures specific to cancerous cells.
In recent decades, advances in proteomic analysis present the main objective of discovering molecular biomarkers for the cancer early detection, characterization of the tumor profile and identification of new therapeutic targets [33]. A particular advantage of the proteome is that not only tissues but also body fluids such as blood, urine and saliva can be used to investigate the molecular correlation of disease and drug action [34,35].
For example, in the study of Khan and colleagues (2014) [36] they demonstrated that survivin and the survivin splice variants are exosomally packaged in the breast cancer patients' sera, mimicking the survivin splice variant pattern that they also report in breast cancer tissues. They concluded that differential expression of exosomal-survivin, particularly survivin-2B, might serve as a diagnostic and/or prognostic marker in early breast cancer patients (Table 1).
Proteomic profiling on exosomes released from three breast cancer cell lines (MCF-7, MDA-MB-231 and Rab27b) identified 85 differentially expressed proteins. In metastatic tumor-derived exosomes, they observed up-regulation of unique set of adhesion proteins (vimentin, galectin-3-binding protein, annexin A1, plectin, protein CYR61, EGF-like repeat and discoidin I-like domain containg protein, filamin-B, protein-glutamine gamma-glutamytransferase 2) [37].
Using proteomic analysis, Blomme and colleagues (2016) [38] validated a novel exosomal protein termed myoferlin, which is related with angiogenesis, metabolism reprogramming and epithelial mesenchymal transition in cancer [39], and demonstrated that myoferlin depletion in cancer cells leads to a significantly modulated exosomes that were functionally deficient; for instance, they had a significantly reduced ability to induce migration and proliferation of those cells.
Vardaki and colleagues (2016) [40] documented significant differences not only in the number of secreted exosomes but most importantly in the protein content of exosomes secreted from metastatic versus non-metastatic tumors. They identified periostin, a protein that on cancer cells seems to bind to integrins triggering Akt/PKB and FAK signaling pathways and increases angiogenesis, invasion, metastasis and cell survival [41].It was found enriched in exosomes secreted by metastatic cells and validated its presence in a pilot cohort of breast cancer patient samples with localized disease or lymph node metastasis.
Hurwitz and colleagues (2016) [42] used sixty National Cancer Institute (NCI-60) cell lines to provide the largest proteomic profile of extracellular vesicles in a single study, identifying a total of 6071 proteins. Only tetraspanin CD81, Alix and HSC70 were found in all samples. Another protein, periostin, was confirmed in both metastatic breast cancer cell lines (MDA-MB-231 and HS 578T) but was not detected in other non-metastatic breast cancer EVs. Despite not composing a canonical pathway, additional proteins including raftilin, fibulin-7 and plasminogen activator inhibitor 1 were found exclusively in EVs from metastatic breast cancer. The great similarity of molecular profiles suggested that EVs reliably represent their progenitor cells and are therefore excellent candidates for biomarkers for clinical strategies to fight cancer.
In the study by Moon and colleagues (2016) [43], fibronectin (FN), a protein that mediates the interation of cells with extracellular matrix (ECM) and downstream factor that promotes metastasis [44], was selected as a biomarker candidate, due to its presence in the surface of EVs secreted from human BC cell lines. FN levels were significantly elevated at all stages of BC and returned to normal after tumor removal. At early stages of BC, another study conducted by Lee and colleagues (2017) [45] reported Developmental Endothelial Locus-1 (DEL-1) as a possible diagnostic tool to distinguish from benign tumors and healthy breast.
Gangoda and colleagues (2017) [46] conducted a comparative proteomic analysis of exosomes isolated from several genetically related mouse breast tumor lines with different metastatic propensity. They observed that metastatic cell-derived exosomes contained proteins capable of promoting cell migration, proliferation, invasion and angiogenesis, while the non-metastatic cell-derived exosomes contained proteins involved in cell-cell/cell-matrix adhesion and polarity maintenance. The metastatic exosomes revealed a distinct set of membrane proteins including ceruloplasmin and metadherin which could presumably aid in targeting the primary cancer cells to specific metastatic sites.
Exosome Annexin A2 was demonstrated by Maji and colleagues (2017) [47] to be an important component of the breast cancer microenvironment signaling by promoting angiogenesis and vascularization via tPA dependent increase in plasmin generation. Additionally, it also induced macrophage activation to create a pre-metastatic aiding for distant organ-specific breast cancer metastasis. The authors concluded exosome-associated Annexin A2 can be exploited as a potential biomarker as well as a therapeutic target for diagnosis and treatment of metastatic breast cancer.
Rontogianni and colleagues (2019) [48] focused on defining EV subtype-specific signatures that could play a role in non-invasive diagnostic testing. To this end, they profiled the proteomes of EVs secreted by BC cell lines and patient serum, with a special emphasis on the TNBC and HER2 subtypes. Some representative TNBC-signature proteins included ephrin type-A receptor 2, DnaJ homolog subfamily A member 1, polyadenylate-binding protein 1, and neuropilin-1, which showed higher expression levels in the patient’s EVs compared with the EVs of HER2-positive patients. Similarly, receptor tyrosine-protein kinase erbB-2, growth factor receptor-bound protein 7, eukaryotic translation initiation factor 3 subunit H and brefeldin A-inhibited guanine nucleotide-exchange protein 2 were the most discriminative protein markers for the HER2-positive patient serum-derived EVs. Their data revealed very distinct proteomic profiles across the different cell line-derived EVs, thus reflecting the unique biology of its breast cancer subtype.
Distinct proteomic content of EVs was also demonstrated in invasive breast cancer cell lines, compared to non-invasive breast cancer cells. EVs produced by the invasive MDA-MB231 cells were significantly enriched for proteins involved in vesicle formation, protein synthesis, proteolysis, and glycolysis. Conversely, EVs produced by MCF10 were significantly enriched by membrane proteins, adhesion molecules, proteins involved in cellular migration, and components of the extracellular matrix (ECM). Based on these differences, the most abundant proteins uniquely identified in MDA-MB231 EVs were those involved in transcriptional regulation (spliceosome, transcription factors, ribosomal proteins, tRNA ligases), proteolysis (proteasome units, pyrophosphatase), EV formation (annexin and vesicle markers LAMP-1 and EEA1), cell cycle (NUMA1), and cell motility and adherence to extracellular matrices (vitronectin, collagen, filamin proteins, and EDIL3) [20].
Dalla and colleagues (2020) [49] also examined the protein contents of EVs derived from three well-known breast cancer cell lines, MCF-7, MDA-MB-231, and T47D. They found seven proteins to be most commonly abundant across the three cell-line-derived EVs. They were actin cytoplasmic 1, pyruvate kinase, glyceraldehyde-3-phosphate dehydrogenase, mitochondrial 60 kDa heat shock protein, mitochondrial ATP synthase subunit alpha, sodium/potassium-transporting ATPase subunit beta-3 and voltage-dependent anion-selective channel protein 2. Interestingly, most of them are metabolism-related protein involved with mitochondrial processes which supports the hypothesis of EVs’ role on metabolic reprogramming.
Furthermore, Risha and colleagues (2020) [50] studied EVs proteomes of MDA-MB-231 and MCF-10A cell lines. While 87 proteins were predicted to be relevant to breast cancer, a total of 16 proteins were mainly involved with cancer metastasis. Three exosomal membrane/surface proteins, glucose transporter 1 (GLUT-1), glypican 1 (GPC-1), disintegrin and metalloproteinase domain-containing protein 10 (ADAM10), were identified as potential breast cancer biomarkers and indicate that distinct molecular mechanisms can be mediated in cancer via EVs, like glucose uptake and ECM remodeling.
Vinik and colleagues (2020) [51] isolated small EVs-enriched fractions from plasma of healthy controls and BC patients at different disease stages before and after surgery. Proteomic analysis of sEV-enriched fractions revealed a signature of seven proteins that differentiated BC patients from healthy individuals, named fibronectin, FAK, MEC1, B-actine, p90RSK_pT573, N-Cadherin and C-Raf, of which FAK and fibronectin displayed high diagnostic accuracy and are related with cell adhesion.
More recently, Li and colleagues (2021) [52] studied the proteomic landscape of serum exosomes derived from ten patients with TNBC and 17 healthy donors. Interestingly, they found that the tetraspanin CD151 expression levels in TNBC-derived serum exosomes were significantly higher than those exosomes from healthy subjects, and they validated their findings with samples from 16 additional donors. Furthermore, they observed that exosomal CD151 facilitated secretion of ribosomal proteins via exosomes while inhibiting exosome secretion of complement proteins. Notably, CD151-deleted exosomes significantly decreased the migration and invasion of TNBC cells.
In a different approach, Patwardhan and colleagues (2021) [53] studied the mechanisms by which ECM stiffening drives breast cancer invasion, demonstrating the role of exosomes in ECM stiffness triggered breast cancer invasiveness. Proteomic analysis of exosomal lysates revealed an enrichment of cell adhesion and cell migration proteins in exosomes from stiff ECM cultures compared to that of soft ones; in these cultures, thrombospondin-1 (THBS1) was identified as a prospective regulator of stiffness-dependent cancer invasion by engaging matrix metalloproteinase and focal adhesion kinase.
Taken together, these data may support the development of new diagnostic tools, but further investigation is still required (Table 1).

Table 1.
List of the main EVs-associated proteins as potential biomarkers for BC.
| Author | Proteins |
|---|---|
| Khan et al. (2014) [36] | Survivin and splice variants |
| Harris et al. (2015) [37] | Vimentin, galectin-3-binding protein, annexin A1, plectin, protein CYR61, EGF-like repeat and discoidin I-like domain containg protein, filamin-B, protein-glutamine gamma-glutamytransferase 2 |
| Blomme et al. (2016) [38] | Myoferlin |
| Vardaky et al. (2016) [40] | Periostin |
| Hurwitz et al. (2016) [42] | Periostin, raftilin (lipid-raft regulating protein), fibulin-7 (adhesion molecule), and plasminogen activator inhibitor 1 (serine protease inhibitor) |
| Moon et al. (2016) [43] | Fibronectin |
| Lee et al. (2017) [45] | Del-1 |
| Gangoda et al. (2017) [46] | Ceruloplasmin, metadherin |
| Maji et al. (2017) [47] | Anexin A2 |
| Rontogianni et al. (2019) [48] | EPHA2, DNAJA1, PABPC1, and NRP1, ERBB2, GRB7, EIF3H and ARFGEF2 |
| Jordan et al. (2020) [20] | Spliceosome, transcription factors, ribosomal proteins, tRNA ligases, proteasome units, pyrophosphatase, annexin and vesicle markers LAMP-1 and EEA1, NUMA1, vitronectin, collagen, filamin proteins, and EDIL3 |
| Dalla et al. (2020) [49] | Actin cytoplasmic 1, pyruvate kinase, glyceraldehyde-3-phosphate dehydrogenase, mitochondrial 60 kDa heat shock protein, mitochondrial ATP synthase subunit alpha, sodium/potassium-transporting ATPase subunit beta-3 and voltage-dependent anion-selective channel protein 2 |
| Risha et al. (2020) [50] | Glucose transporter 1 (GLUT-1), glypican 1 (GPC-1), and disintegrin and metalloproteinase domain-containing protein 10 (ADAM10) |
| Vinik et al. (2020) [51] | Fibronectin, FAK, MEC1, B-actine, p90RSK_pT573, N-Cadherin and C-Raf |
| Li et al. (2021) [52] | CD151 |
| Patwardhan et al. (2021) [53] | Thrombospondin-1 (THBS1) |
Concluding Remarks and Perspectives
In view of the possibility of performing early diagnosis and longitudinal prognostic evaluation, studies focusing on exosomal proteomics of patients diagnosed with breast tumor has grown. Also, one of the most promising applications of cancer protein biomarkers relies on the development of targeted therapies. By identifying specific proteins that drive tumor growth, metastasis, or resistance to treatment, researchers can design drugs or treatment strategies that specifically target these biomarkers. Also, the identification of a group of proteins allows to clarify what type of biological processes are deregulated due to EVs. In the present review, is clear that metabolic reprogramming at distinct points occurs, but also several proteins that mediates cell interaction with ECM, suggesting the key role of EVs on metastasis. This could improve the advances in personalized approach, known as precision medicine, maximizes treatment effectiveness while minimizing side effects on healthy tissues. For obvious reasons, research in the canine species is still scarce, albeit important, since CMT is considered an excellent experimental model for BC. Due to the shorter lifespan of dogs and the rapid progression of CMTs, researchers can gain insights into tumor development, metastasis, and therapeutic responses in a relatively shorter timeframe compared to human studies. On the other hand, liquid biopsy has several advantages over traditional tissue biopsy in the context of breast cancer, since it enables the identification and characterization of tumor-specific genetic alterations and mutations, even at early stages when traditional imaging techniques may not detect the disease. Its non-invasive nature makes it an attractive option for serial sampling, reducing patient discomfort and facilitating longitudinal studies. Therefore, unveiling biomarker proteins carried by the exosomes through liquid biopsy can provide precious information. Indeed, if the biomarker detection occurs before they carry out their tumor signaling, there is a chance to interrupt the pathogenesis of the disease. Biomarker proteins could be modulated at various levels, such as silencing genes, affecting their transcription or even the protein signaling pathway. It would be possible, for example, to block the signaling effect of exosomal metastasis-inducing proteins or even to modulate this effect by introducing protective signaling proteins into the vesicles. Cancer protein biomarkers have revolutionized cancer research and patient care, ushering in a new era of precision medicine. As researchers continue to unravel the intricate landscape of cancer biology, protein biomarkers will remain invaluable in the fight against cancer, bringing hope for improved outcomes, enhanced quality of life, and ultimately, a world free from the burden of this devastating disease.
Author Contributions
AAN conceived the review, compiled the literature and wrote the manuscript. GHT compiled the literature and wrote the manuscript. LGAC prepared figures for the article, guided the insertions and the scope of the review. DAPCZ conceived the review and checked all drafts and the final version of the report. All authors significantly contributed and approved the final version of the manuscript.
Funding
The authors are grateful to the São Paulo Research Foundation (FAPESP) and the National Council for Scientific and Technological Development (CNPq). The authors received financial support as follows: GHT (FAPESP grant number 2022/00890-5), DAPCZ (FAPESP grant number 2020/12970-8; CNPq Process number 307288/2022-6), and LGAC (CNPq process number 304108/2020-0).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Sorenmo, K. Canine mammary gland tumors. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 573–596. [Google Scholar] [CrossRef]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef]
- Lakhtakia, R. A Brief History of Breast Cancer: Part I: Surgical domination reinvented. Sultan Qaboos Univ. Med. J. 2014, 14, e166–e169. [Google Scholar] [PubMed]
- Shiovitz, S.; Korde, L.A. Genetics of breast cancer: a topic in evolution. Ann. Oncol. 2015, 26, 1291–1299. [Google Scholar] [CrossRef]
- Polyak, K. Breast cancer: origins and evolution. J. Clin. Invest. 2007, 117, 3155–3163. [Google Scholar] [CrossRef] [PubMed]
- Alimirzaie, S.; Bagherzadeh, M.; Akbari, M.R. Liquid biopsy in breast cancer: A comprehensive review. Clin. Genet. 2019, 95, 643–660. [Google Scholar] [CrossRef]
- Freitas, A.J.A.d.; Causin, R.L.; Varuzza, M.B.; Calfa, S.; Hidalgo Filho, C.M.T.; Komoto, T.T.; Souza, C.d.P.; Marques, M.M.C. Liquid biopsy as a tool for the diagnosis, treatment, and monitoring of breast cancer. Int. J. Mol. Sci. 2022, 23, 9952. [Google Scholar] [CrossRef]
- Halvaei, S.; Daryani, S.; Eslami-S, Z.; Samadi, T.; Jafarbeik-Iravani, N.; Bakhshayesh, T.O.; Majidzadeh-A, K.; Esmaeili, R. Exosomes in cancer liquid biopsy: A focus on breast cancer. Mol. Ther. Nucleic Acids 2018, 10, 131–141. [Google Scholar] [CrossRef]
- Tay, T.K.Y.; Tan, P.H. Liquid biopsy in breast cancer: A focused review. Arch. Pathol. Lab. Med. 2021, 145, 678–686. [Google Scholar] [CrossRef]
- da Costa, A.; Lenze, D.; Hummel, M.; Kohn, B.; Gruber, A.D.; Klopfleisch, R. Identification of six potential markers for the detection of circulating canine mammary tumour cells in the peripheral blood identified by microarray analysis. J. Comp. Pathol. 2012, 146, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, J.; Hu, G. Biomarker studies in early detection and prognosis of breast cancer. Adv. Exp. Med. Biol. 2017, 1026, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Gam, L.-H. Breast cancer and protein biomarkers. World J. Exp. Med. 2012, 2, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Loke, S.Y.; Lee, A.S.G. The future of blood-based biomarkers for the early detection of breast cancer. Eur. J. Cancer 2018, 92, 54–68. [Google Scholar] [CrossRef]
- Celis, J.E.; Gromov, P.; Cabezón, T.; Moreira, J.M.A.; Ambartsumian, N.; Sandelin, K.; Rank, F.; Gromova, I. Proteomic characterization of the interstitial fluid perfusing the breast tumor microenvironment: a novel resource for biomarker and therapeutic target discovery. Mol. Cell. Proteomics 2004, 3, 327–344. [Google Scholar] [CrossRef]
- Duan, S.; Buxton, I.L.O. Evolution of medical approaches and prominent therapies in breast cancer. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Antonyak, M.A.; Li, B.; Boroughs, L.K.; Johnson, J.L.; Druso, J.E.; Bryant, K.L.; Holowka, D.A.; Cerione, R.A. Cancer cell-derived microvesicles induce transformation by transferring tissue transglutaminase and fibronectin to recipient cells. Proc Natl Acad Sci USA 2011, 108, 4852–4857. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by extracellular vesicles: where we are and where we need to go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Jordan, K.R.; Hall, J.K.; Schedin, T.; Borakove, M.; Xian, J.J.; Dzieciatkowska, M.; Lyons, T.R.; Schedin, P.; Hansen, K.C.; Borges, V.F. Extracellular vesicles from young women’s breast cancer patients drive increased invasion of non-malignant cells via the Focal Adhesion Kinase pathway: a proteomic approach. Breast Cancer Res. 2020, 22, 128. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; Singh, S.; Williams, C.; Soplop, N.; Uryu, K.; Pharmer, L.; King, T.; Bojmar, L.; Davies, A.E.; Ararso, Y.; Zhang, T.; Zhang, H.; Hernandez, J.; Weiss, J.M.; Dumont-Cole, V.D.; Kramer, K.; Wexler, L.H.; Narendran, A.; Schwartz, G.K.; Healey, J.H.; Sandstrom, P.; Labori, K.J.; Kure, E.H.; Grandgenett, P.M.; Hollingsworth, M.A.; de Sousa, M.; Kaur, S.; Jain, M.; Mallya, K.; Batra, S.K.; Jarnagin, W.R.; Brady, M.S.; Fodstad, O.; Muller, V.; Pantel, K.; Minn, A.J.; Bissell, M.J.; Garcia, B.A.; Kang, Y.; Rajasekhar, V.K.; Ghajar, C.M.; Matei, I.; Peinado, H.; Bromberg, J.; Lyden, D. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Jabalee, J.; Towle, R.; Garnis, C. The role of extracellular vesicles in cancer: cargo, function, and therapeutic implications. Cells 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-X.; Gires, O. Tumor-derived extracellular vesicles in breast cancer: From bench to bedside. Cancer Lett. 2019, 460, 54–64. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Klose, P.; Weise, C.; Bondzio, A.; Multhaup, G.; Einspanier, R.; Gruber, A.D. Proteome of metastatic canine mammary carcinomas: similarities to and differences from human breast cancer. J. Proteome Res. 2010, 9, 6380–6391. [Google Scholar] [CrossRef]
- Suárez-Bonnet, A.; Herráez, P.; Martín de las Mulas, J.; Rodríguez, F.; Déniz, J.M.; Espinosa de los Monteros, A. Expression of 14-3-3 σ protein in normal and neoplastic canine mammary gland. Vet. J. 2011, 190, 345–351. [Google Scholar] [CrossRef]
- Jagarlamudi, K.K.; Westberg, S.; Rönnberg, H.; Eriksson, S. Properties of cellular and serum forms of thymidine kinase 1 (TK1) in dogs with acute lymphocytic leukemia (ALL) and canine mammary tumors (CMTs): implications for TK1 as a proliferation biomarker. BMC Vet. Res. 2014, 10, 228. [Google Scholar] [CrossRef] [PubMed]
- Jena, S.C.; Shrivastava, S.; Saxena, S.; Kumar, N.; Maiti, S.K.; Mishra, B.P.; Singh, R.K. Surface plasmon resonance immunosensor for label-free detection of BIRC5 biomarker in spontaneously occurring canine mammary tumours. Sci. Rep. 2019, 9, 13485. [Google Scholar] [CrossRef]
- Fhaikrue, I.; Srisawat, W.; Nambooppha, B.; Pringproa, K.; Thongtharb, A.; Prachasilchai, W.; Sthitmatee, N. Identification of potential canine mammary tumour cell biomarkers using proteomic approach: Differences in protein profiles among tumour and normal mammary epithelial cells by two-dimensional electrophoresis-based mass spectrometry. Vet. Comp. Oncol. 2020, 18, 787–795. [Google Scholar] [CrossRef]
- Tamarindo, G.H.; Novais, A.A.; Chuffa, L.G.A.; Zuccari, D.A.P.C. Metabolic alterations in canine mammary tumors. Animals (Basel) 2023, 13. [Google Scholar] [CrossRef]
- Park, H.-M.; Kim, H.; Kim, D.W.; Yoon, J.-H.; Kim, B.-G.; Cho, J.-Y. Common plasma protein marker LCAT in aggressive human breast cancer and canine mammary tumor. BMB Rep. 2020, 53, 664–669. [Google Scholar] [CrossRef]
- Cordeiro, Y.G.; Mulder, L.M.; van Zeijl, R.J.M.; Paskoski, L.B.; van Veelen, P.; de Ru, A.; Strefezzi, R.F.; Heijs, B.; Fukumasu, H. Proteomic Analysis Identifies FNDC1, A1BG, and Antigen Processing Proteins Associated with Tumor Heterogeneity and Malignancy in a Canine Model of Breast Cancer. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Yuan, S.H.-C.; Chang, S.-C.; Huang, Y.; Liu, H.-P. Serum Level of Tumor-Overexpressed AGR2 Is Significantly Associated with Unfavorable Prognosis of Canine Malignant Mammary Tumors. Animals (Basel) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Gast, M.-C.W.; Schellens, J.H.M.; Beijnen, J.H. Clinical proteomics in breast cancer: a review. Breast Cancer Res. Treat. 2009, 116, 17–29. [Google Scholar] [CrossRef]
- Hondermarck, H.; Tastet, C.; El Yazidi-Belkoura, I.; Toillon, R.-A.; Le Bourhis, X. Proteomics of breast cancer: the quest for markers and therapeutic targets. J. Proteome Res. 2008, 7, 1403–1411. [Google Scholar] [CrossRef]
- Latterich, M.; Abramovitz, M.; Leyland-Jones, B. Proteomics: new technologies and clinical applications. Eur. J. Cancer 2008, 44, 2737–2741. [Google Scholar] [CrossRef]
- Khan, S.; Bennit, H.F.; Turay, D.; Perez, M.; Mirshahidi, S.; Yuan, Y.; Wall, N.R. Early diagnostic value of survivin and its alternative splice variants in breast cancer. BMC Cancer 2014, 14, 176. [Google Scholar] [CrossRef]
- Harris, D.A.; Patel, S.H.; Gucek, M.; Hendrix, A.; Westbroek, W.; Taraska, J.W. Exosomes released from breast cancer carcinomas stimulate cell movement. PLoS ONE 2015, 10, e0117495. [Google Scholar] [CrossRef]
- Blomme, A.; Fahmy, K.; Peulen, O.; Costanza, B.; Fontaine, M.; Struman, I.; Baiwir, D.; de Pauw, E.; Thiry, M.; Bellahcène, A.; Castronovo, V.; Turtoi, A. Myoferlin is a novel exosomal protein and functional regulator of cancer-derived exosomes. Oncotarget 2016, 7, 83669–83683. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, B.; Zhao, C.; Ba, Z.; Xu, H.; Yan, X.; Liu, W.; Zhu, B.; Wang, L.; Ren, C. Myoferlin, a multifunctional protein in normal cells, has novel and key roles in various cancers. J. Cell. Mol. Med. 2019, 23, 7180–7189. [Google Scholar] [CrossRef] [PubMed]
- Vardaki, I.; Ceder, S.; Rutishauser, D.; Baltatzis, G.; Foukakis, T.; Panaretakis, T. Periostin is identified as a putative metastatic marker in breast cancer-derived exosomes. Oncotarget 2016, 7, 74966–74978. [Google Scholar] [CrossRef] [PubMed]
- Dorafshan, S.; Razmi, M.; Safaei, S.; Gentilin, E.; Madjd, Z.; Ghods, R. Periostin: biology and function in cancer. Cancer Cell Int. 2022, 22, 315. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, S.N.; Rider, M.A.; Bundy, J.L.; Liu, X.; Singh, R.K.; Meckes, D.G. Proteomic profiling of NCI-60 extracellular vesicles uncovers common protein cargo and cancer type-specific biomarkers. Oncotarget 2016, 7, 86999–87015. [Google Scholar] [CrossRef]
- Moon, P.-G.; Lee, J.-E.; Cho, Y.-E.; Lee, S.J.; Jung, J.H.; Chae, Y.S.; Bae, H.-I.; Kim, Y.-B.; Kim, I.-S.; Park, H.Y.; Baek, M.-C. Identification of Developmental Endothelial Locus-1 on Circulating Extracellular Vesicles as a Novel Biomarker for Early Breast Cancer Detection. Clin. Cancer Res. 2016, 22, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-C.; Yang, C.-H.; Cheng, L.-H.; Chang, W.-T.; Lin, Y.-R.; Cheng, H.-C. Fibronectin in cancer: friend or foe. Cells 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, J.; Jung, J.H.; Park, H.Y.; Moon, P.-G.; Chae, Y.S.; Baek, M.-C. Exosomal Del-1 as a Potent Diagnostic Marker for Breast Cancer: Prospective Cohort Study. Clin. Breast Cancer 2021, 21, e748–e756. [Google Scholar] [CrossRef]
- Gangoda, L.; Liem, M.; Ang, C.-S.; Keerthikumar, S.; Adda, C.G.; Parker, B.S.; Mathivanan, S. Proteomic Profiling of Exosomes Secreted by Breast Cancer Cells with Varying Metastatic Potential. Proteomics 2017, 17. [Google Scholar] [CrossRef]
- Maji, S.; Chaudhary, P.; Akopova, I.; Nguyen, P.M.; Hare, R.J.; Gryczynski, I.; Vishwanatha, J.K. Exosomal annexin II promotes angiogenesis and breast cancer metastasis. Mol. Cancer Res. 2017, 15, 93–105. [Google Scholar] [CrossRef]
- Rontogianni, S.; Synadaki, E.; Li, B.; Liefaard, M.C.; Lips, E.H.; Wesseling, J.; Wu, W.; Altelaar, M. Proteomic profiling of extracellular vesicles allows for human breast cancer subtyping. Commun. Biol. 2019, 2, 325. [Google Scholar] [CrossRef]
- Dalla, P.V.; Santos, J.; Milthorpe, B.K.; Padula, M.P. Selectively-Packaged Proteins in Breast Cancer Extracellular Vesicles Involved in Metastasis. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Risha, Y.; Minic, Z.; Ghobadloo, S.M.; Berezovski, M.V. The proteomic analysis of breast cell line exosomes reveals disease patterns and potential biomarkers. Sci. Rep. 2020, 10, 13572. [Google Scholar] [CrossRef]
- Vinik, Y.; Ortega, F.G.; Mills, G.B.; Lu, Y.; Jurkowicz, M.; Halperin, S.; Aharoni, M.; Gutman, M.; Lev, S. Proteomic analysis of circulating extracellular vesicles identifies potential markers of breast cancer progression, recurrence, and response. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, X.; Yang, S.; Pi, H.; Li, Z.; Yao, P.; Zhang, Q.; Wang, Q.; Shen, P.; Li, X.; Ji, J. Proteomic Landscape of Exosomes Reveals the Functional Contributions of CD151 in Triple-Negative Breast Cancer. Mol. Cell. Proteomics 2021, 20, 100121. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, S.; Mahadik, P.; Shetty, O.; Sen, S. ECM stiffness-tuned exosomes drive breast cancer motility through thrombospondin-1. Biomaterials 2021, 279, 121185. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Diverse cellular signalig of exosomes (EVs) are described to influence the establishment and spread of cancer. EV-mediated effects may occur at tumor site (eg. persistent migration, invadopodia formation, ECM remodeling, induction of protrusive activity and drug resistance and differentiation of tumor-promoting stromal cells) and at distant sites (eg. high vascular permeability, induction of ECM remodeling and increased BMDC recruitment) [19]. EV- extracellular vesicle; MVB – multivesicular bodies; ECM – extracellular matrix; BMDC – bone marrow-derived cell.
Figure 1.
Diverse cellular signalig of exosomes (EVs) are described to influence the establishment and spread of cancer. EV-mediated effects may occur at tumor site (eg. persistent migration, invadopodia formation, ECM remodeling, induction of protrusive activity and drug resistance and differentiation of tumor-promoting stromal cells) and at distant sites (eg. high vascular permeability, induction of ECM remodeling and increased BMDC recruitment) [19]. EV- extracellular vesicle; MVB – multivesicular bodies; ECM – extracellular matrix; BMDC – bone marrow-derived cell.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



