
Submitted:
09 September 2023
Posted:
20 September 2023
You are already at the latest version
Abstract
All allergic responses to food indicate failure of immunological tolerance but it is unclear why cow’s milk and egg (CME) allergies resolve more readily than reactivity to peanuts (PN). We sought to identify differences between PN and CME allergies through constitutive immune status and responses to cognate and non-cognate food antigens. Children with confirmed allergy to CME (n=6), PN (n=18) non-allergic (NA) (n=8) controls were studied. Constitutive secretion of cytokines was tested in plasma and unstimulated mononuclear cell (PBMNC) cultures. Blood dendritic cell (DC) subsets were analyzed alongside changes in phenotypes and soluble molecules in allergen stimulated MNC cultures with or without cytokine neutralization. We observed that in allergic children constitutively high plasma levels IL-1β, IL-2, IL-4, IL-5 and IL-10 but less IL-12p70 than in non-allergic children was accompanied by spontaneous secretion of sCD23, IL-1β, IL-2 IL-4, IL-5, IL-10, IL-12p70, IFN- and TNF- in MNC cultures. Furthermore, blood DC subset counts differed in food allergy. Antigen-presenting cell phenotypic abnormalities were accompanied by higher B and T cell percentages with more Bcl-2 within CD69+ subsets. Cells were generally refractory to antigenic stimulation in vitro but IL-4 neutralization led to CD152 downregulation by CD4+ T cells from PN allergic children responding to PN allergens. Canonical discriminant analyses segregated non-allergic and allergic children by their cytokine secretion patterns, revealing differences and areas of overlap between PN and CME allergies. Despite an absence of recent allergen exposure, indication of in vivo activation, in vitro responses independent of challenging antigen and the presence of unusual costimulatory molecules suggest dysregulated immunity in food allergy. Most important, higher Bcl-2 content within key effector cells implies survival advantage with potential to mount abnormal responses that may give rise to the manifestations of allergy. Here, we put forward the hypothesis that the lack of apoptosis of key immune cell types might be central to the development of food allergic reactions.
Keywords:
food allergy
; cytokines
; immunoregulation
; pediatric allergy
; apoptosis
Introduction
Maintaining oral tolerance is one of the main tasks of the intestinal immune system that is achieved through cross-talk between the microbiota, epithelium and underlying immune system. Defective homeostasis can result in dysregulated immune responses such as those found in food allergy. There are many implied causal factors, including perturbations in composition of the intestinal microbiota [1], an imbalance in Th1/Th2 responses[2], deficient T cell[3] or DC[4] regulatory function, changes in antigen presentation[5], abnormalities in expression of β7 integrin[6] and mutations in the filaggrin[7] or cytokine receptor[8] genes. Multiple factors are likely to be involved and susceptibility of any individual will be determined by any of these, alone or in combination resulting in a failure of oral tolerance. All allergic reactions are recall responses and like initial sensitization events their effective regulation requires T-cell and antigen-presenting cell (APC) interactions involving the CD28/CD152-CD80/CD86 signaling pathway[9].
The CD28/CD152-CD80/CD86 signaling pathway is one of the most important regulators of both Th1 and Th2 responses in vivo[10]. It is essential for the development of allergic reactions to food antigens and crucial for inducing tolerance to them[11]. CD28 and CD86 are largely constitutive on the surface of respective T and antigen presenting cells whereas CD152 and CD80 are upregulated on activation[12]. CD80 interaction with CD152 is crucial for tempering the intensity of initial sensitization events and also the induction of low dose tolerance[12,13]. While the mechanisms by which this occurs is not known, ligation of CD80 on APCs with constitutively expressed CD152 on regulatory T cells (Treg) may activate the indoleamine-2,3-dioxygenase pathway in DCs, leading to the production of immunosuppressive kynurenin[14]. As CD86 favors CD23-CD21 pairing it consequently functions as a selective and potent costimulus for IgE synthesis by activated B cells[15]. CD23 is the low affinity receptor for IgE. It exists in membrane-bound and soluble forms and participates in both positive and negative regulation of IgE synthesis by B and other cell types[16,17,18]. As a multifunctional molecule, CD23 can also promote B cell survival via receptor-mediated inhibition of apoptosis[19] or through upregulation of Bcl-2[20,21]. Like B cells, T and other cells also require ongoing signals to maintain their viability and in an allergic setting IL-4 can upregulate Bcl-2[22,23]. Such deregulation of cell death in key cells is likely to have a significant effect on allergic responses as previously observed in mice[24,25].
With this in mind we have documented expression of these key molecules, as well as sCD23, Bcl-2 and the early activation marker CD69. We have also quantified DC subsets and measured cytokines released spontaneously or in response to antigenic stimulation as these proteins are central to signaling events in any immune response. Some food allergies resolve spontaneously (e.g., CME) while others are less likely to do so (e.g., PN) [26]; thus, it is clear that alongside shared features of immune induction/dysregulation there must also be characteristics that are determined by the nature of the allergen itself in the genetic and immunological context of the individual involved. Our study therefore sought to document phenotypic and functional similarities and differences between PN and CME allergic children, and to compare them with profiles derived for those without any allergies.
Methods
Study population
Children with PN (eighteen, age range 3-12 years of age, average 9.6) or CME (six, age-range 4-13, average 10.2) allergies attending pediatric allergy clinics alongside NA (eight, age-range 4-10, average 8.7) children were invited to participate in the study. Only allergic patients with confirmed mono-allergic status were enrolled to the study. This was determined by using SPT-induced wheal size of 8mm or more and allergen-specific IgE level of 15KUA/L or greater. The parents of all children signed an informed consent form approved by the local Ethics Committee; LREC No:361/01.
Allergens
The preparation of crude peanut (Arachis hypogea) extract was carried out as described previously[27]. Stock solution was at 100mg/mL and it was used at 10mg/mL culture. Egg albumin (Cat. A5503), milk casein (Cat. C3400) and β-lactoglobulin (Cat. L0130) were purchased from Sigma-Aldrich (Dorset, UK), stored at 10mg/mL stock solutions and used at 10µg/mL culture.
Plasma collection and isolation and culture of peripheral blood mononuclear cells
Blood was collected into sodium heparin vacutainer tubes (Becton Dickinson, Oxford, UK) at the relevant center and sent by courier at ambient temperature. On receipt the blood was centrifuged at 700 g for 10 minutes, plasma was collected and frozen at −80OC until use. Peripheral blood mononuclear cells (PBMNC) with a viability >95% were prepared as described previously[28]. 1x106 MNC were cultured in the absence or presence of allergen and with or without neutralizing antibodies added at culture initiation. After 6 days at 37OC in an atmosphere of 5% CO2 in air, culture supernatants were removed, centrifuged at 380 g for 5 minutes and frozen at −80OC until use. Cells were harvested, washed in phosphate buffered saline containing 0.1% sodium azide, 2% fetal bovine serum and 4mM EDTA (PBSAAE) with centrifugation at 380 g for 5 minutes. Washed cells were stained as described below.
Antibodies
Neutralizing antibodies anti-TNF-α (Cat. AB-210-NA), pan-specific TGF-β (Cat. AB-100-NA), anti-IL-6 (AB-206-NA) and anti-IL-10 (Cat. AB-217-NA) were purchased from RnD Systems (Abingdon, UK). Anti-IL-4 (Cat. 554481) was purchased from BD Biosciences (UK). Each was used at 10µg/mL culture. Furthermore, the antibodies for cell phenotyping and the appropriate isotype controls were as follows: HLA-DR-ECD (Cat. PN IM3636), CD19-ECD (Cat. A07770), CD4-ECD (Cat. 6604727) and IgG1-ECD (Cat. A07797) were purchased from Beckman Coulter (UK). The remaining antibodies were purchased from BD Biosciences (UK); CD80-FITC (Cat. 557226), CD83-FITC (Cat. 556910), CD86-FITC (Cat. 555657), CD3-FITC (Cat. 561807), CD83-PE (Cat. 556855), CD152-PE (Cat. 555853), CD86-APC (Cat. 555660), CD69-APC (Cat. 555533), anti-Bcl-2-PE set (Cat.556536), Mouse IgG1-FITC (Cat.555748), mouse IgG1-PE (Cat. 555749), mouse IgG2a-PE (Cat. 555574) and mouse IgG1-APC (Cat. 555751). Antibodies were titrated to find a concentration that gave optimum performance and both antibodies and their isotype controls were used at the same concentration.
Cell surface and intracellular staining
MNC were first stained for surface antigen expression by incubation with the appropriate antibody or isotype control cocktails for 15 minutes at room temperature. After washing once with PBSAAE stained cells were fixed and permeabilized (Fix and Perm kit, Life Technologies, Paisley, UK; GAS004) according to the manufacturer’s instructions. Anti-Bcl-2-FITC or hamster isotype-FITC was added to the permeabilized cells which were then left at room temperature in the dark for 45 minutes. Following one wash with PBSAAE data were acquired on a Beckman Coulter FC500 flow cytometer and analyzed using MXP software. Flow cytometry was also used for quantitative analysis of dendritic cell subsets. To this end, DC subset counts were performed in whole blood as described[29] using a blood dendritic cell enumeration kit purchased from Miltenyi Biotec Ltd. (Cat.130-091-086), that included CD141/BDCA-3-APC (myeloid cDC), and CD303/BDCA-2-FITC for plasmacytoid DC (PDC). Red blood cells were lysed using a red cell lysing buffer (Sigma, UK; Cat. R7757) according to the manufacturer’s instructions. Just before data acquisition by flow cytometry (Beckman Coulter FC500 cytometer) 100µL Flow-Count Fluorospheres (Beckman Coulter, UK) were added to each tube to enable absolute counting.
Detection of sCD23 and cytokine levels
Frozen culture supernatants were thawed rapidly (at 37OC) before analysis. sCD23 levels were measured by enzyme-linked immunosorbent assay (Bender MedSystems) and detected using a Biorad Benchmark Plus microplate spectrophotometer (Biorad, Hertfordshire, UK). Furthermore, cytokines were detected through multiplexed bead-array technology. A human Th1/Th2 10-Plex FlowCytomix™ kit was purchased from Bender MedSystems (Vienna, Austria, now eBioscience®) to detect IL-1β, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IFN-γ and TNF-α and used according to manufacturer’s instructions. Data were acquired on a Beckman Coulter FC500 flow cytometer and analyzed using MXP software (Beckman Coulter, High Wycombe, UK).
Statistics
A Linear ANOVA Model with three factors was first assayed. The factors were “type of allergy” with levels “none PN CME”, “Challenging agent” with levels “none PN CME” and “neutralizing antibody” with levels “anti-IL10, anti-IL4, anti-TGFb, anti-IL6 and none”. Prior to the analyses the value of the measurements, V, was transformed into Ln(V+1). Thereafter the model was simplified as a two way ANOVA model with two factors: “type of allergy” with levels “none PN CME” and “challenging agent” with levels “none PN CME”. The interaction between the effects of these two factors was not significant in any case. Least squares means were computed for each effect to estimate significance of the differences between the levels of each factor. Canonical discriminant analyses were performed to find linear functions able to discriminate between non-allergic individuals and individuals allergic to CME or PN. Prior to analyses each variable, V, was transformed as V’ = ln(V+1). Cross-validation error did not increase. All statistical analyses were carried out using SAS 9.3 (SAS Institute Inc., Cary, NC, USA.)
Results
Constitutive plasma cytokines in food allergy
In comparison with the plasma of non-allergic children, there were constitutively higher levels (P<0.05) of IL-1β (Figure 1A), IL-2 (Figure 1B), IL-4 (Figure 1C), IL-5 (Figure 1D) and IL-10 (Figure 1F) in the plasma of CME and PN allergic children but lower levels of IL-12p70 (Figure 1G). There were no differences in plasma concentrations of IL-6 (Figure 1E), IFN-γ (Figure 1H) or TNF-α (Figure 1I) between allergic and non-allergic children. These data suggest either a predisposition towards Th2 responses in food allergy or on-going in vivo stimulation in affected subjects.
Spontaneous in vitro sCD23 release and cytokine secretion by MNC in food allergy
Consistent with the data above, MNC from allergic children cultured for 6 days in the absence of added antigens spontaneously released significantly more (P<0.05) sCD23 (Fig2A), IL-1β (Figure 2B), IL-2 (Figure 2C), IL-4 (Figure 2D), IL-5 (Figure 2E), IL-10 (Figure 2F), IL-12p70 (Figure 2G), IFN-γ (Figure 2H) and TNF-α (Figure 2I) whereas MNC from non-allergic children secreted virtually none of the cytokines. Challenge with CME or PN antigens did not alter the levels of these soluble factors (data not shown) suggesting that the MNC were already activated and refractory to further in vitro stimulation.
Allergy-associated abnormalities in the expression of costimulatory molecules
We examined the expression of CD80, CD83 and CD86 on in vitro unstimulated MNC and those stimulated with PN or CME antigens. In 6-day unstimulated MNC cultures there were significantly more (P<0.05) CD80+ DR− (Figure 3A), CD80+ CD83+ (Figure 3B), CD80+ CD86− (Figure 3C) subsets in PN allergic than non-allergic MNC and significantly less (P<0.05) CD86+ CD80− (Figure 3D) subsets in MNC from both PN and CME allergic children than those from non-allergic children. There were no differences noted between PN and CME allergies. Antigenic stimulation did not alter the expression of costimulatory molecules. These data suggest dysregulated costimulatory molecule expression in allergy which could impact on allergenic responses in vivo.
Differences in the numbers of immune cell subsets
In order to assess if there was an imbalance of myeloid (mDC) and plasmacytoid dendritic cell (pDC) subsets as documented in atopic patients[30] we quantified absolute numbers of these subsets in whole blood of allergic and non-allergic children. There were significantly more (P<0.05) mDC cells in the blood of CME allergic than either PN allergic or non-allergic children (Figure 4). Although the number of pDCs showed a similar trend the differences did not reach statistical significance. These data highlight differences in DC subsets between PN and CME allergies that may have an impact on in vivo allergenic responses and resolution or lack of it.
We then checked whether the differences in mDC and pDC subsets were accompanied by changes in other cellular subsets. To this end, we quantified the percentages of B cells, T cells and APC following culture for 6 days in the absence or presence of PN or CME antigens. Irrespective of antigenic challenge there were no differences in the percentages of B cells between the different study groups (Figure 5A). However, there were significantly more B cells in cultures from CME allergic children responding to PN antigens than CME antigens. T cell percentages were significantly higher (P <0.05) in allergic than in non-allergic subjects (Figure 5B). This may be due to spontaneous activation of T cells in food allergic subjects[8], resulting in their higher numbers. Antigenic challenge did not induce any changes. No differences were apparent in the percentages of APC between allergic and non-allergic children (Figure 5C).
Survival advantage of immune cell subtypes from allergic children
When the Bcl-2 content of cultured cells was examined, it seemed that the cells were not refractory to in vitro stimulation as previously thought. Taking this into consideration alongside the allergy-associated constitutive presence of cytokines in plasma and spontaneous release of soluble molecules during in vitro culture we looked for the expression of CD69, a leukocyte activation molecule expressed in situations of chronic inflammation[31]. Furthermore, reduced levels of apoptosis were observed in a mouse model of food allergy. Important, cells escaping T cell-mediated apoptosis when transferred into a naïve animals triggered an antigen specific IgE response suggesting an important role in the genesis of allergic reactions[24]. Thus, we also examined the expression of the pro-survival factor Bcl-2 to ascertain if MNC from allergic children had a survival advantage. We report that Bcl-2 content of CD69+ B cells (Figure 5D) and T cells (Figure 5E) was significantly higher (P<0.05) than that of similar cells from non-allergic donors. The Bcl-2 content of CD69+ APC was comparable between the study subjects (Figure 5F). In all cases challenge with CME or PN antigens did not alter the constitutive Bcl-2 expression (Figure 5E−G). These data suggest that activated T and B cells from allergic children have a survival advantage and that such longevity might influence the duration of a discrete allergic response or persistence of an individual’s allergic sensitivity over time.
The survivability of CD69+ T cells from food allergic children could be mediated through surface expression of CD152. It has been shown that signals induced by CD152 act directly on activated T lymphocytes to confer resistance to cell death mainly in Th2 cells[31,32]. While we found no differences in the expression of cell surface CD152 on CD4+ T cells from allergic and non-allergic children (Figure 6A), there was significantly higher expression (P<0.05) on the surface of CD69+ CD4+ T cells from both CME and PN allergic compared with non-allergic donors (Figure 6B). These data suggest that in food allergy CD152 may also confer longevity to activated CD4+ T cells.
IL-4-mediated regulation of CD152 expression
As cell surface expression of CD152 can be upregulated by cytokines[34] we tested the ability of several neutralizing antibodies (anti-IL-4. -IL-6, -IL-10 and -TGF-β) to alter its expression. The only effect seen was on MNC from PN allergic children challenged with their cognate allergen when neutralization of IL-4 brought cell surface CD152 expression down to the levels seen in non-allergic children (Figure 7). It has been reported that stimulation in the presence of IL-4 induces CD152 expression on B cells[35] and this may also be the case with T cells.
Canonical discriminant analysis
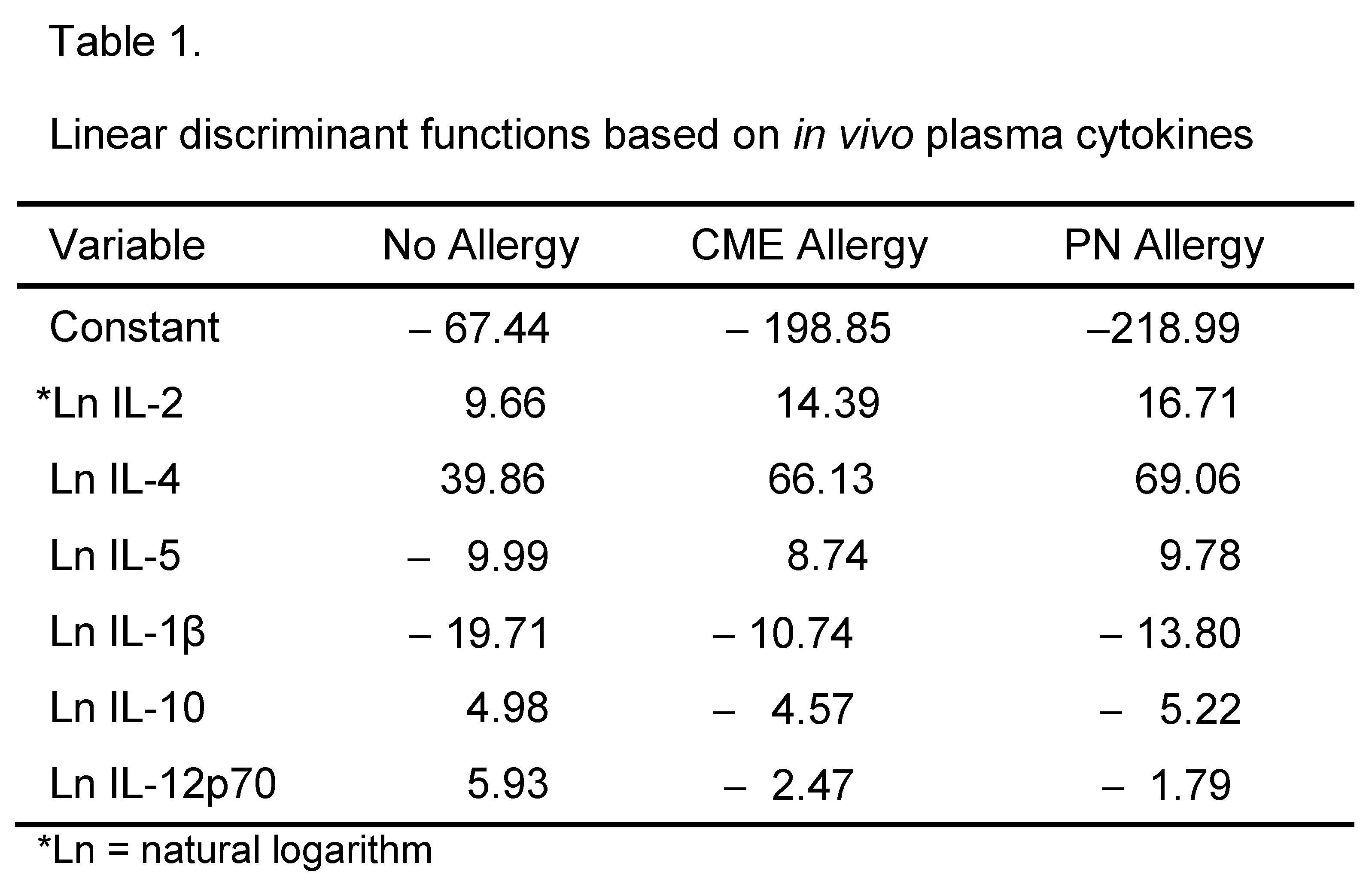
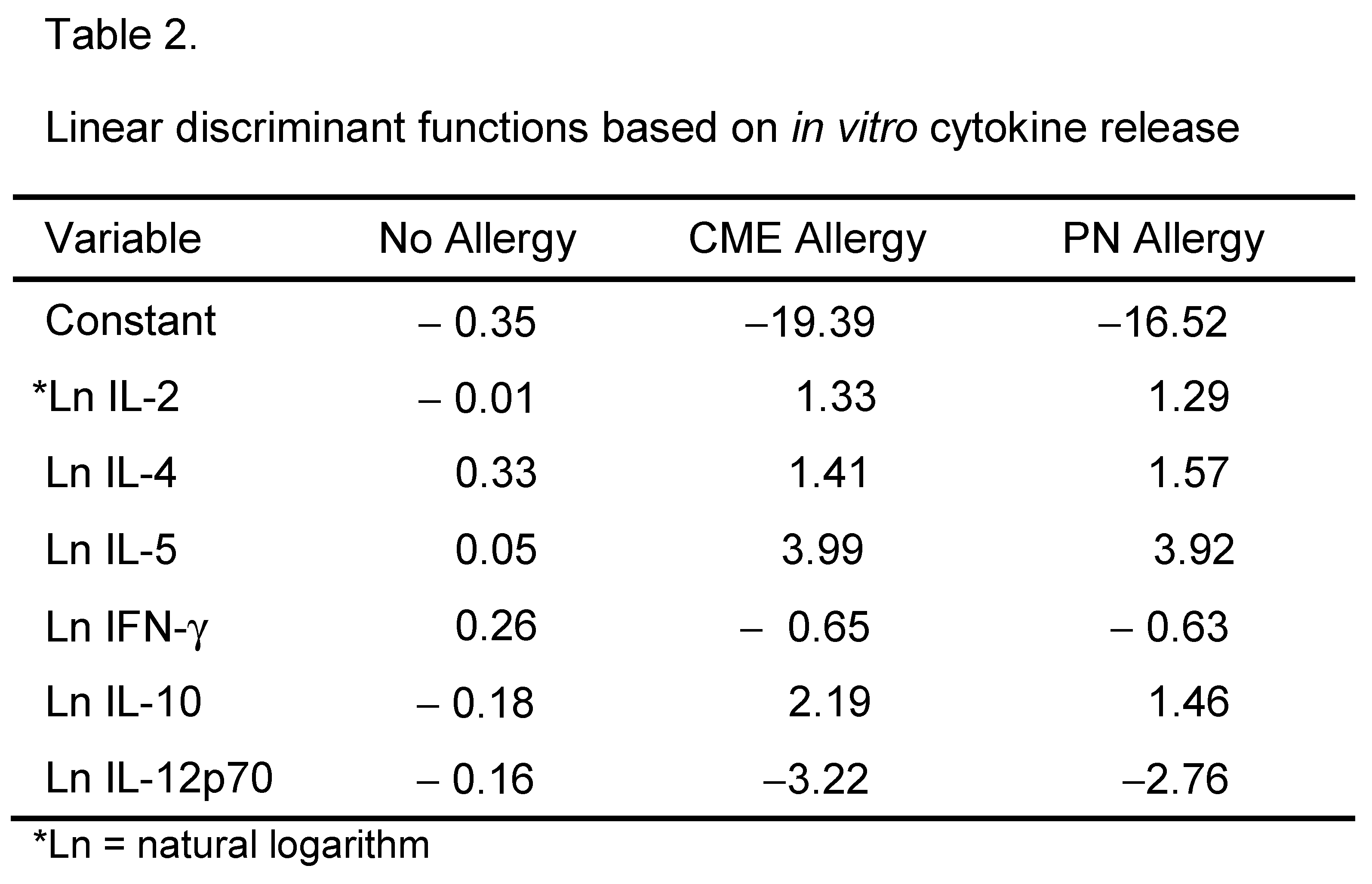
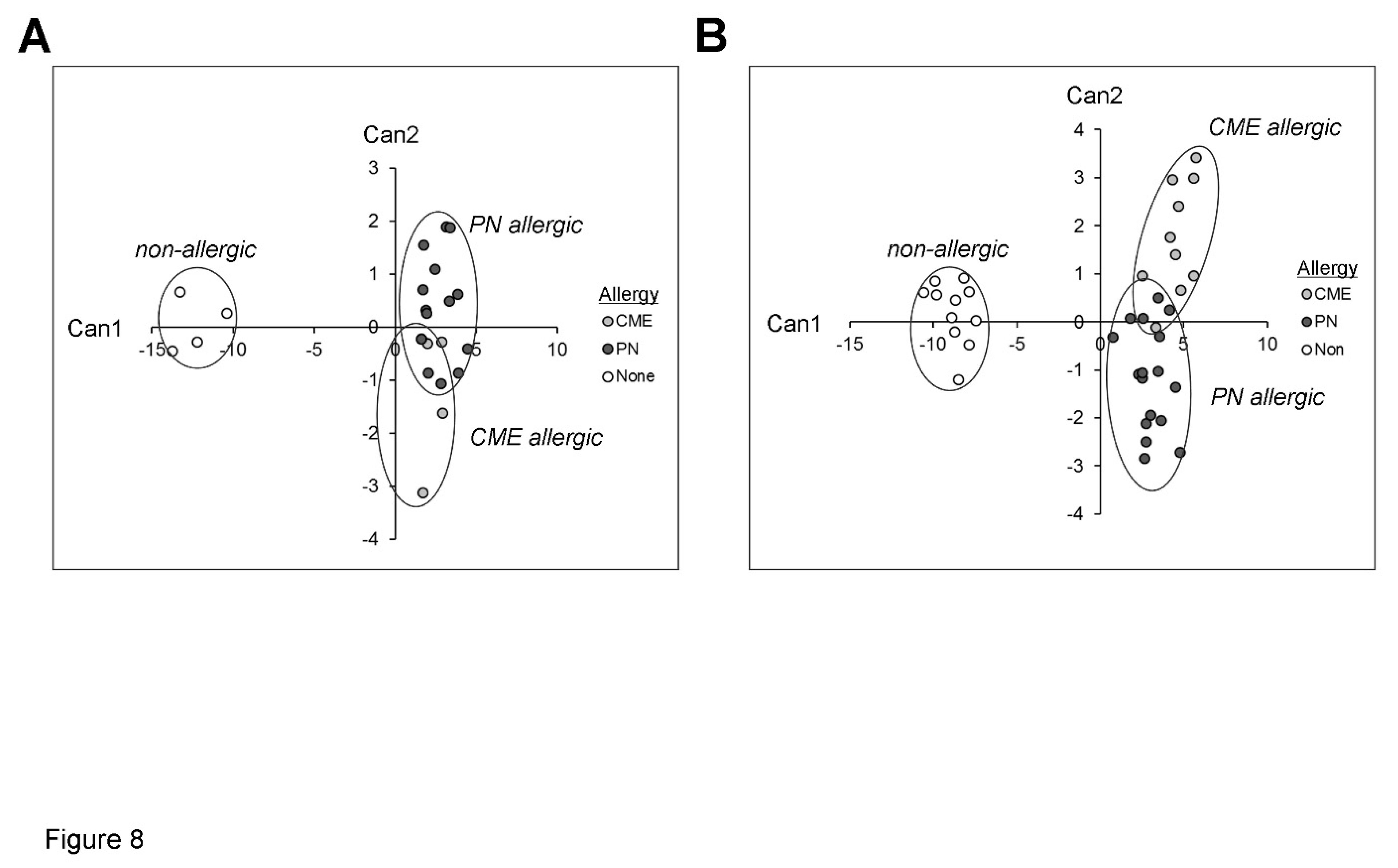
Canonical discriminant analyses were performed to find linear functions able to differentiate between non allergic individuals and individuals allergic to CME or PN. Only subjects with complete data sets have been included in these analyses. Linear discriminant functions based on plasma measurements of in vivo (Table 1) and in vitro (Table 2) cytokine secretion were efficient in discriminating between non-allergic and allergic individuals but not between individuals allergic to PN or CME Figure 8A,B). Nevertheless, there was some separation between CME and PN allergic children alongside overlap between the two allergies reflecting their similarities, differences and variations in severity of their particular allergies.
Discussion
This study suggests that children with CME and PN allergy diverged in their mechanisms of immune regulation/dysregulation and most important both populations displayed survival advantage of key cell types.. We show constitutive presence of Th2, pro-inflammatory and regulatory cytokines in the plasma of allergic children. With regard to regulatory cytokines, IL-10 and TGF-ß are found mainly in the allergen-specific Treg cell population[36] and there is strong evidence that peripheral T-cell regulation has a crucial role in the control of allergy. Our data is in agreement with a previous observation of[37] with regard to IL-4 but the allergic children in our study were older and had virtually no IL-12p70 with higher levels of IL-10, indicative of in vivo Th2 bias. IL-4 has been shown to enhance IL-10 gene expression in Th2 cells[38]. This could represent negative feedback regulation, except that plasma IL-10 levels are comparable between PN tolerant and PN allergic individuals[39]. Furthermore, severe acute anaphylaxis episodes can occur in allergic patients with high IL-10 levels[40]. Indeed, several studies have shown that IL-10 may promote the development of Th2 cells [41,42,43] and facilitate IgE production[44,45] to contribute to allergy-related disease through collaboration with IL-1β[46] which was also higher in our allergic subjects. There is a requirement for IL-1 in Th2 cell activation[47] and IL-1β can induce the production of IL-2[48] which was also higher in the plasma of our allergic children. In turn, IL-2 may be responsible for the increased plasma IL-5 in our allergic subjects[49] and successful oral immunotherapy for peanut allergy is associated with a decrease in both IL-5 and IL-2[50]. IL-5 induces terminal maturation of eosinophils, prolongs eosinophil survival by delaying apoptotic death, possesses eosinophil chemotactic activity, increases eosinophil adhesion to endothelial cells and enhances eosinophil effector functions[51]. So, its presence in plasma suggests an enhanced ability to mount Type I hypersensitivity responses. This poses an important question as to why these cytokines are present in the circulation of allergic children who have avoided eating the offending foods. The answer may lie, at least partly, in molecular cross-reactivity of food allergens with other plant food sources or with aeroallergens in the environment[52,53,54] because most food allergic children have other chronic atopic conditions such as asthma, eczema or allergic rhinitis.
A constitutive presence of Th2 cytokines in the plasma of allergic children was accompanied by the spontaneous release of these and other cytokines in MNC cultures. A previous study[8] showed that in the absence of allergen exposure, Th2 cytokine secretion by T cells from food allergic donors is DC dependent. In agreement with our study, they found that no further increase in these cytokines occurred after stimulation with allergens and concluded that T cells from these subjects were already activated in vivo rendering them unresponsive to additional allergenic stimulation in vitro. They found abnormalities in costimulatory molecule expression in eosinophilic oesophagitis but not in food allergy. In our study however, using a different detection panel we did find differences in costimulatory molecule expression between non-allergic versus allergic and PN versus CME allergies. The differences from non-allergic children involved CD80+ subsets with lower expression of other costimulatory molecules as well. Due to the complexities of costimulatory interactions it is impossible to speculate on the impact of these changes on immune responsiveness. However, it is recognized that aside from overlapping functions, CD80 and CD86 have distinct functional identities. CD80 is associated with diminution of immune responses through interaction with its functional ligand CD152, to exert tolerogenic effects when expressed by immature dendritic cells in the absence of CD86 upregulation[55]. Certainly, there were lower percentages of CD80 and CD86 double positive cells following in vitro challenge with either CME or PN antigens. These unusual subsets in our allergic children may impair normal maturation of dendritic cells that has been described in allergy[56]. It is also noteworthy that IL-10 was constitutively high in the plasma of our allergic children and its presence during DC maturation impairs upregulation of costimulatory molecules[57]. On a speculative note, in the presence of such allergic immune dysregulation weakly immunogenic food allergens may pose a problem. It is equally possible that the failure to upregulate CD86 may be due to in vivo activation rendering the cells refractory to further stimulation. One could argue that in vivo activation should yield a higher proportion of CD86+ cells but this may not be so as CD86 has faster association and dissociation rates than CD80[55] making stable complex formation unlikely. CD152-CD80 regulatory interactions are likely to proceed in the absence of CD86. Indeed, CD69+ cells that were present in our allergic participants had more CD152 expressed.
Signals generated by CD152 on activated CD69+ T cells induce resistance to cell death[58]. This notion has a direct bearing on our observation that in allergic children levels of the anti-apoptotic Bcl-2 within CD69+ CD152+ T cells were significantly higher. Dysregulation of apoptosis, a cell death pathway key for the development and the maintenance of homeostasis, is at the core of a variety of diseases [59] Most important, we have previously observed that in a mouse model of food allergy lack of apoptosis occurred during antigen-presentation[24] and that the adoptive transfer of apoptosis-resistant cells triggered antigen-specific response in naïve mice even in absence of antigen administration[60]. Further, the added presence of IL-4, also a viability factor, would serve to sustain high Bcl-2 levels within both B and T cells[61,62].
It is important to highlight that up to date the presence of cells displaying a clear survival advantage in food allergy has not been reported in humans. Longevity of allergic effector T cells could influence both magnitude and duration of responses amplified by a higher frequency of allergen-specific T cells as seen in IB[63] We hypothesize that expanded populations of allergen-specific T cells are maintained by on-going stimulation in vivo through cross reaction with other food proteins and that these cells contribute to exaggerated Type I hypersensitivity responses through resistance to the intrinsic cell death pathway.
Despite the Th2 bias of both CME and PN allergies, IL-4-mediated regulation of CD152 on the surface of CD4+ T cells after challenge with PN was seen in PN and NA donors but not in CME donors. Other differences included sCD23 and IL-1β release, costimulatory molecule expression, DC1 subsets, and a greater survival advantage of CD69+ B cells from PN allergic donors. In view of the plasticity of DCs, the impact of higher numbers in CME allergy is not known. Canonical discriminant analysis confirmed distinction between PN and CME allergies. We suggest that aside from the greater allergenicity of seed storage proteins found in peanuts, there are inherent differences in the regulation of PN versus other food allergies in affected individuals.
Conclusions
In summary, this study suggests the existence of differences in various immunological parameters between different allergies and non-allergic children, the most noticeable being the survival advantage acquired in allergic individuals, through higher Bcl-2 expression in T and B cells. Up to date the latter was observed in a mouse model of food allergy[24] but not described in humans. We are aware of the small number of patients utilized in this study. Thus, the results enable us to formulate the hypothesis that the lack of apoptosis is pivotal to the development of food allergy that will need to be confirmed by a study involving a larger cohort of subjects. The latter will also enable the investigation on the mechanism underlying the allergy-related survival advantage of certain cell types key to the regulation of immune response.
Author Contributions
Conceptualization and supervision CN, KI, MM; investigation KI, DB, RA, FP; Writing original draft KI, CN; Writing – Review & Editing, all authors.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Southampton General Hospital LREC No:361/01.
Informed Consent Statement
Parental consent was obtained from all subjects involved in the study.
Data Availability Statement
Data are available upon request.
Acknowledgements
The authors wish to thank J. Hourihane for the samples and insightful discussions and C Pin for statistical analysis.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
APC, antigen presenting cell, CME, cow’s milk or egg; DC, dendritic cell; ECD, electron coupled dye; EDTA, ethylenediaminetetraacetic acid; FITC, fluorescein isothiocyanate; IFN-β, interferon-beta; Ig, immunoglobulin; IL, interleukin ; MNC, mononuclear cell; PBMNC, peripheral blood mononuclear cell ; PE, phycoerythrin; PN, peanut; sCD23, soluble CD23, Th2, T-helper 2; TNF-α, tumour necrosis factor-alpha
References
- Ling Z, Li Z, Liu X, Cheng Y, Luo Y, Tong X, Yuan L, Sun J, Li L, C. Altered Fecal Microbiota Composition Associated with Food Allergy in Infants. Appl Environ Microbiol 2014, 80, 2546–2554. [Google Scholar] [CrossRef] [PubMed]
- Tomicić S, Fälth-Magnusson K, Böttcher MF. Dysregulated Th1 and Th2 responses in food-allergic children--does elimination diet contribute to the dysregulation? Pediatr Allergy Immunol 2010, 4, 649–655. [Google Scholar]
- Chatila, T. Regulatory T cells: Key Players in Tolerance and Autoimmunity. Endocrinol Metab Clin North Am 2009, 38, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Kwon HK, Lee CG, So JS, Chae CS, Hwang JS, Sahoo A, Nam JH, Rhee JH, Hwang KC, Im SH. Generation of regulatory dendritic cells and CD4+Foxp3+ T cells by probiotics administration suppresses immune disorders. Proc Natl Acad Sci USA 2010, 107, 2159–2164.
- Ruiter B, Shreffler WG. The role of dendritic cells in food allergy. J Allergy Clin Immunol 2012, 129, 921–928. [Google Scholar] [CrossRef]
- Pabst O, Mowat AM. Oral tolerance to food protein. Mucosal Immunol 2012, 5, 232–239. [Google Scholar] [CrossRef]
- Ogrodowczyk A, Markiewicz L, Wróblewska B. Mutations in the filaggrin gene and food allergy. Prz Gastroenterol 2014, 9, 200–207. [Google Scholar]
- Frischmeyer-Guerrerio PA, Guerrerio AL, Chichester KL, Bieneman AP, Hamilton RA, Wood RA, Schroeder JT. Dendritic cell and T cell responses in children with food allergy. Clin Exp Allergy 2011, 41, 61–71. [Google Scholar] [CrossRef]
- Boesteanu AC, Katsikis PD. Memory T cells need CD28 costimulation to remember. Semin Immunol. 2009, 21, 69–77. [Google Scholar] [CrossRef]
- Borriello F, Sethna MP, SD, Schweitzer AN, Tivol EA, Jacoby D, Strom TB, Simpson EM, Freeman GJ, Sharpe AH. B7-1 and B7-2 Have Overlapping, Critical Roles in Immunoglobulin Class Switching and Germinal Center Formation. Immunity 1997, 6, 303–313. [Google Scholar] [CrossRef]
- van Wijk F, Nierkens S, de Jong W, Wehrens EJ, Boon L, van Kooten P, Knippels LM, Pieters R. The CD28/CTLA-4-B7 signaling pathway is involved in both allergic sensitization and tolerance induction to orally administered peanut proteins. J Immunol 2007, 178, 6894–6900. [Google Scholar] [CrossRef] [PubMed]
- Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system of T cell costimulation. Ann Rev Immunol 1996, 14, 233–258. [Google Scholar] [CrossRef] [PubMed]
- van Wijk F, Hoeks S, Nierkens S, Koppelman SJ, van Kooten P, Boon L, Knippels LM, Pieters R. CTLA-4 signaling regulates the intensity of hypersensitivity responses to food antigens, but is not decisive in the induction of sensitization. J Immunol 2005, 174, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Grohmann U, Orabona C, Fallarino F, Vacca C, Calcinaro F, Falorni A, Candeloro P, Belladonna ML, Bianchi R, Fioretti MC, Puccetti P. CTLA-4–Ig regulates tryptophan catabolism in vivo. Nat. Immunol 2002, 3, 1097–1101. [Google Scholar] [CrossRef]
- Jeannin P, Delneste Y, Lecoanet-Henchoz S, Gauchat J-F Jonathan Ellis J, Bonnefoy J-Y. CD86 (B7.2) on human B cells. A functional role in proliferation and selective differentiation into IgE and IgG4-producing cells. J Biol Chem 1997, 272, 15613–15619. [Google Scholar]
- Cooper AM, Hobson PS, utton MR, Kao MW, Drung B, Schmidt B, Fear DJ, Beavil AJ, McDonnel JM, Sutton BJ, Gould HJ. Soluble CD23 Controls IgE Synthesis and Homeostasis in Human B Cells. J Immunol 2012, 188, 3199–3207. [Google Scholar] [CrossRef]
- Gould HJ, Sutton BJ. IgE in allergy and asthma today. Nature Rev Immunol 2008, 8, 205–217. [Google Scholar] [CrossRef]
- Edkins AL, Borland G, Acharya M, CogdellRJ, Ozanne BW, Cushley W. Differential regulation of monocyte cytokine release by αV and β2 integrins that bind CD23. Immunol 2012, 136, 241–251. [Google Scholar] [CrossRef]
- Lindsey, J. White LJ. Bradford W. Ozanne BW, Pierre Graber P, Jean-Pierre Aubry J-P, Jean-Yves Bonnefoy JY, William Cushley W. Inhibition of Apoptosis in a Human Pre-B–Cell Line by CD23 Is Mediated Via a Novel Receptor. Blood 1997, 90, 234–243. [Google Scholar]
- Liu YJ, Cairns JA, Holder MJ, Abbot SD, Jansen KU, Bonnefoy JY, Gordon J, MacLennan I C. Recombinant 25-kDa CD23 and interleukin 1 alpha promote the survival of germinal center B cells: evidence for bifurcation in the development of centrocytes rescued from apoptosis. Eur J Immunol 1991, 21, 1107–1114. [Google Scholar] [CrossRef]
- Cory, S. Regulation of lymphocyte survival by the BCL-2 gene family. Ann Rev Immunol 1995, 13, 513–543. [Google Scholar] [CrossRef] [PubMed]
- Graninger WB, Steiner CW, Graninger MT, Aringer M, Smolen JS. Cytokine regulation of apoptosis and Bcl-2 expression in lymphocytes of patients with systemic lupus erythematosus. Cell Death Diff 2000, 7, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Aronica MA, Goenka S, Boothby M. IL-4-dependent induction of BCL-2 and BCL-X(L)IN activated T lymphocytes through a STAT6- and pi 3-kinase-independent pathway. Cytokine 2000, 12, 578–587. [Google Scholar] [CrossRef]
- Man AL, Bertelli E, Regoli M, Chambers SJ, Nicoletti C. Antigen-specific T cell–mediated apoptosis of dendritic cells is impaired in a mouse model of food allergy. J Allergy Clin Immunol 2004, 113, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Arques JL, Regoli M, Bertelli E, Nicoletti C. Persistence of apoptosis-resistant T cell-activating dendritic cells promotes T helper type-2 response and IgE antibody production. Mol Immunol 2008, 45, 2177–2186. [Google Scholar] [CrossRef] [PubMed]
- Sicherer SH, Food allergy from infancy through adulthood. J Allergy Clin Immunol. 2020, 8, 1854–1864.
- Clemente A, Chambers SJ, Lodi F, Nicoletti C, Brett GM. Use of indirect competitive ELISA for the detection of Brazil nut in food products. Food Contr 2004, 15, 65–69. [Google Scholar] [CrossRef]
- Ivory K, Chambers SJ, Pin C, Prieto E, Arques JL, Nicoletti C. Oral delivery of Lactobacillus casei Shirota modifies allergen-specific immune responses in allergic rhinitis. Clin Exp Allergy 2008, 38, 1282–1289. [Google Scholar] [CrossRef]
- Riemann, D. , Turzer S, Ganchev G, Schütte W, Seliger B, Möller. Monitoring Blood Immune Cells in Patients with Advanced Small Cell Lung Cancer Undergoing a Combined Immune Checkpoint Inhibitor/Chemotherapy. Biomolecules 2023, 13, 190. [Google Scholar] [CrossRef]
- .Uchida Y, Kurasawa K, Nakajima H, Nakagawa N, Tanabe E, Sueishi M, Saito Y, Iwamoto I. Increase of dendritic cells of type 2 (DC2) by altered response to IL-4 in atopic patients. J Allergy Clin Immunol 2001, 108, 1005–1011. [Google Scholar] [CrossRef]
- Sancho D, Gomez M, Sanchez-Madrid F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol 2005, 26, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Pandiyan P, Gärtner G, Soezeri O, Radbruch A, Schulze-Osthoff K, Brunner-Weinzierl MC. CD152 (CTLA-4) determines the unequal resistance of Th1 and Th2 cells against activation-induced cell death by a mechanism requiring PI3 kinase function. J Exp Med 2004, 199, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Rudd CE, Taylor A, Schneider H. CD28 and CTLA-4 coreceptor expression and signal transduction. Immunol Rev 2009, 229, 12–26. [Google Scholar] [CrossRef]
- Alegre ML, Noel PJ, Eisfelder BJ, Chuang E, Clark MR, Reiner SL, Thompson CB. Regulation of surface and intracellular expression of CTLA4 on mouse T cells. J Immunol 1996, 157, 4762–4770. [Google Scholar] [CrossRef]
- Pioli, C, Gatta L, Ubaldi V, Doria G. Inhibition of IgG1 and IgE production by stimulation of the B cell CTLA-4 receptor. J. Immunol. 2000, 165, 5530–5536. [Google Scholar] [CrossRef] [PubMed]
- Akdis CA, Blesken T, Akdis M, Wüthrich B, Blaser K. Role of interleukin-10 in specific immunotherapy. J. Clin Invest 1998, 102, 98–106. [Google Scholar] [CrossRef]
- Dang TD, Tang MLK, Koplin JJ, Licciardi PV, Eckert JK, Tan T, Gurin LC, Ponsonby A-L, Allen KL. Characterization of plasma cytokines in an infant population cohort of challenge-proven food allergy. Allergy 2013, 68, 1333–1340. [Google Scholar]
- Schmidt-Weber CB, Alexander SI, Henault LE, James L, Lichtman AH. IL-4 Enhances IL-10 Gene Expression in Murine Th2 Cells in the Absence of TCR Engagement. J Immunol 1999, 162, 238–244. [Google Scholar] [CrossRef]
- Lotoski LC, Simons FER, Chooniedass R, Liem J, Ostopowich, Becker AB, HayGlass KT. Are Plasma IL-10 Levels a Useful Marker of Human Clinical Tolerance in Peanut Allergy? PLoS One 2010, 5, e11192. [Google Scholar]
- Stone SF, Cotterell C, Isbister GK, Holdgate A, Brown SG. Elevated serum cytokines during human anaphylaxis: Identification of potential mediators of acute allergic reactions. J Allergy Clin Immunol 2009, 124, 786–792. [Google Scholar] [CrossRef]
- Palma JP, Yauch RL, Kang HK, Lee HG, Kim BS. Preferential induction of IL-10 in APC correlates with a switch from Th1 to Th2 response following infection with a low pathogenic variant of Theiler’s virus. J. Immunol 2002, 168, 4221–4230. [Google Scholar] [CrossRef] [PubMed]
- Laouini D, Alenius H, Bryce P, Oettgen H, Tsitsikov E, Geha RS. IL-10 is critical for Th2 responses in a murine model of allergic dermatitis. J Clin Invest 2003, 112, 1058–1066. [Google Scholar] [CrossRef]
- Kobayashi N, Nagumo H, Agematsu K. IL-10 enhances B-cell IgE synthesis by promoting differentiation into plasma cells, a process that is inhibited by CD27/CD70 interaction. Clin Exp Immunol 2002, 129, 446–452. [Google Scholar] [CrossRef]
- Punnonen J, Kainulainen L, Ruuskanen O, Nikoskelainen J, Arvilommi H. IL-4 synergizes with IL-10 and anti-CD40 MoAbs to induce B-cell differentiation in patients with common variable immunodeficiency. Scand J Immunol 1997, 45, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Uejima Y, Takahashi K, Komoriya K, Kurozumi S, and Ochs HD. Effect of interleukin-10 on anti-CD40- and interleukin-4-induced immunoglobulin E production by human lymphocytes. Int Arch Allergy Appl Immunol 1996, 110, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Nakae S, Komiyama Y, Yokoyama H, Nambu A, Umeda M, Iwase M, Homma I, Sudo K, Horai R, Asano M, Iwakura Y. IL-1 is required for allergen-specific Th2 cell activation and the development of airway hypersensitivity response. Int Immunol 2003, 15, 483–490. [Google Scholar] [CrossRef]
- Dinarello, CA. Biologic basis for interleukin-1 in disease. Blood 1996, 87, 2095–2147. [Google Scholar] [CrossRef]
- van Haelst Pisani C, JS Kovach JS, Kita H, Leiferman KM, Gleich GJ, Silver JE, Dennin R, Abrams JS. Administration of interleukin-2 (IL-2) results in increased plasma concentrations of IL-5 and eosinophilia in patients with cancer. Blood 1991, 78, 1538–1544. [Google Scholar] [CrossRef]
- Blumchen K, Ulbricht H, Staden U, Dobberstein K, Beschorner J, de Oliveira LC, Shreffler WG, Sampson HA, Niggemann B, Wahn U, Beyer K. Oral peanut immunotherapy in children with peanut anaphylaxis. J Allergy Clin Immunol 2010, 126, 83–91. [Google Scholar] [CrossRef]
- Kouro T, Takatsu K. IL-5- and eosinophil-mediated inflammation: from discovery to therapy. Int Immunol 2009, 2, 1303–1309. [Google Scholar]
- Bublin M, Breiteneder H. Cross-Reactivity of Peanut Allergens. Curr Allergy Asthma Rep 2014, 14, 426. [Google Scholar] [CrossRef] [PubMed]
- Rozenfeld P, Docena GH, Añón MC, Fossati CA. Detection and identification of a soy protein component that cross-reacts with caseins from cow’s milk. Clin Exp Immunol 2002, 130, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Popescu F-D. Cross-reactivity between aeroallergens and food allergens. World J Methodol 2015, 5, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Sansom DM, Manzotti CN, Zheng Y. What’s the difference between CD80 and CD86? Trends Immunol 2003, 24, 313–318.
- Casas R, Skarsvik S, Lindström A, Zetterström O, Duchén K. Impaired maturation of monocyte-derived dendritic cells from birch allergic individuals in association with birch-specific immune responses. Scand J Immunol 2007, 66, 591–598. [Google Scholar] [CrossRef]
- Hubo M, Trinschek B, Kryczanowsky F, Tuettenberg A, Steinbrink K, Jonuleit H. Costimulatory Molecules on Immunogenic Versus Tolerogenic Human Dendritic Cells. Front Immunol 2013, 4, 82. [Google Scholar] [CrossRef]
- Pandiyan P, Gärtner D, Soezeri O, Radbruch A, Schulze-Osthoff K, Brunner-Weinzierl MC. CD152 (CTLA-4) determines the unequal resistance of Th1 and Th2 Cells against activation-induced cell death by a mechanism requiring PI3 kinase function. J Exp Med 2004, 199, 831–842. [Google Scholar] [CrossRef]
- Aronica MA, Goenka S, Boothby M. IL-4-dependent induction of BCL-2 and BCL-X(L)IN activated T lymphocytes through a STAT6- and PI3-kinase-independent pathway. Cytokine 2000, 12, 578–587. [Google Scholar] [CrossRef]
- Singh, R. , Letai, A, Sarosiek, K. Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nat Rev Mol Cell Biol 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Chambers SJ, Bertelli E, Winterbone MS, Regoli M, Man AL, Nicoletti C. Adoptive transfer of dendritic cells from allergic mice induces specific immunoglobulin E antibody in naïve recipients in absence of antigen challenge without altering the T helper 1/T helper 2 balance. Immunology. 2004, 112, 72–79. [Google Scholar] [CrossRef]
- Graninger WB, Steiner CW, Graninger MT, M Aringer M, Smolen JS. Cytokine regulation of apoptosis and Bcl-2 expression in lymphocytes of patients with systemic lupus erythematosus. Cell Death Differ 2000, 7, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Berin MC, Sampson HA. Food allergy: an enigmatic epidemic. Trends Immunol 2013, 34, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Mudter J, Neurath MF. Apoptosis of T cells and the control of inflammatory bowel disease: therapeutic implications. Gut. 2007, 56, 293–303. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Constitutive plasma cytokines. (A) IL-1β, (B) IL-2, (C) IL-4, (D) IL-5, (E) IL-6, (F) IL-10, (G) IL-12p70, (H) IFN-γ, (I) TNF-α were evaluated by multiplex bead technology in the plasma of non-allergic children and those with CME or PN allergies. Data were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
Figure 1.
Constitutive plasma cytokines. (A) IL-1β, (B) IL-2, (C) IL-4, (D) IL-5, (E) IL-6, (F) IL-10, (G) IL-12p70, (H) IFN-γ, (I) TNF-α were evaluated by multiplex bead technology in the plasma of non-allergic children and those with CME or PN allergies. Data were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
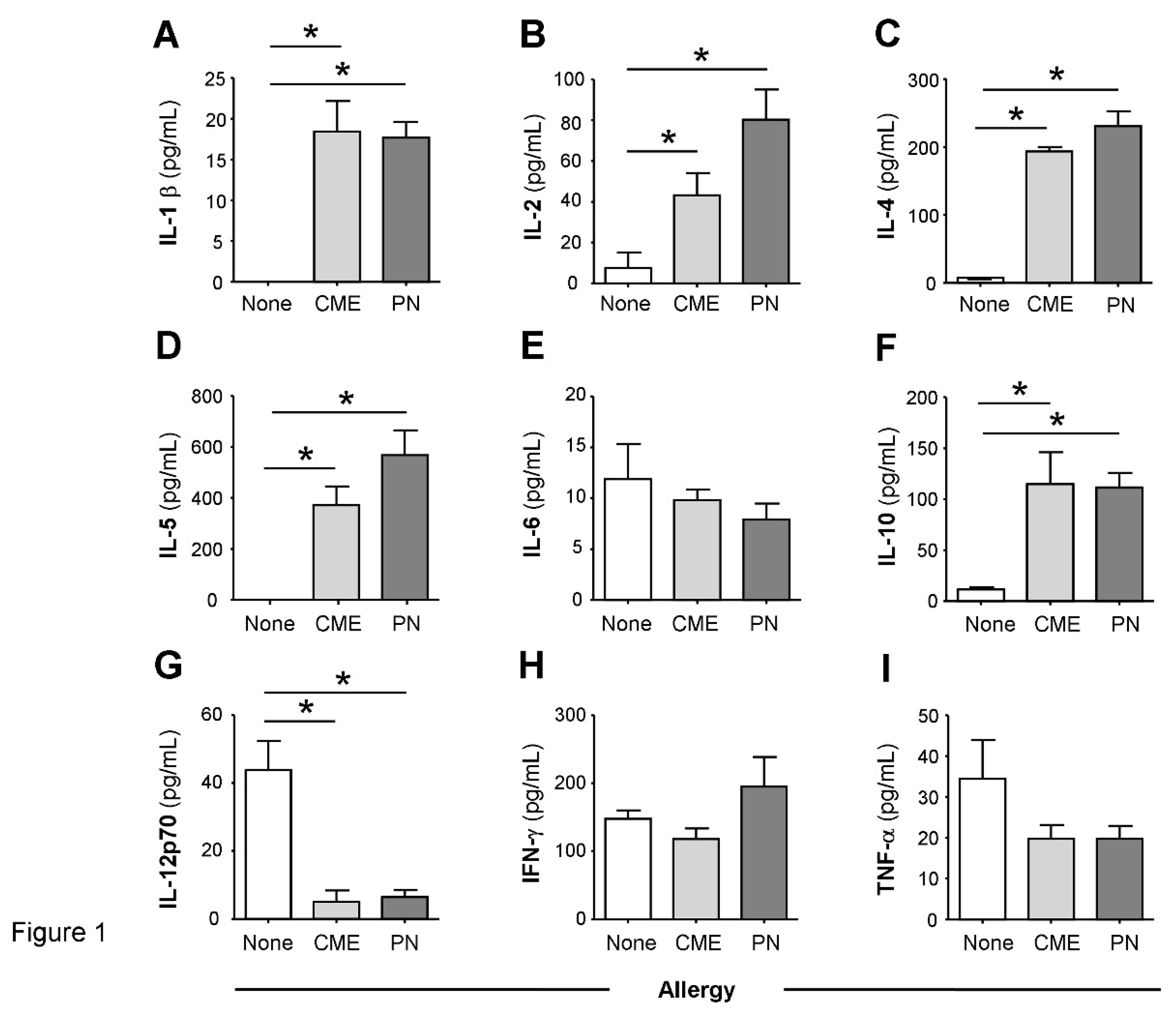
Figure 2.
Spontaneous in vitro sCD23 and cytokine secretion. MNC from non-allergic children or those with CME or PN allergies were cultured for six days without allergenic stimulation. Culture supernatants were collected and their levels of (A) sCD23 were detected by ELISA while cytokines (B) IL-1β, (C) IL-2), (D) IL-4, (E) IL-5, (F) IL-10, (G) IL-12p70, (H) IFN-γ and (I) TNF-α were measured by multiplex bead technology and flow cytometry. Data are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
Figure 2.
Spontaneous in vitro sCD23 and cytokine secretion. MNC from non-allergic children or those with CME or PN allergies were cultured for six days without allergenic stimulation. Culture supernatants were collected and their levels of (A) sCD23 were detected by ELISA while cytokines (B) IL-1β, (C) IL-2), (D) IL-4, (E) IL-5, (F) IL-10, (G) IL-12p70, (H) IFN-γ and (I) TNF-α were measured by multiplex bead technology and flow cytometry. Data are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
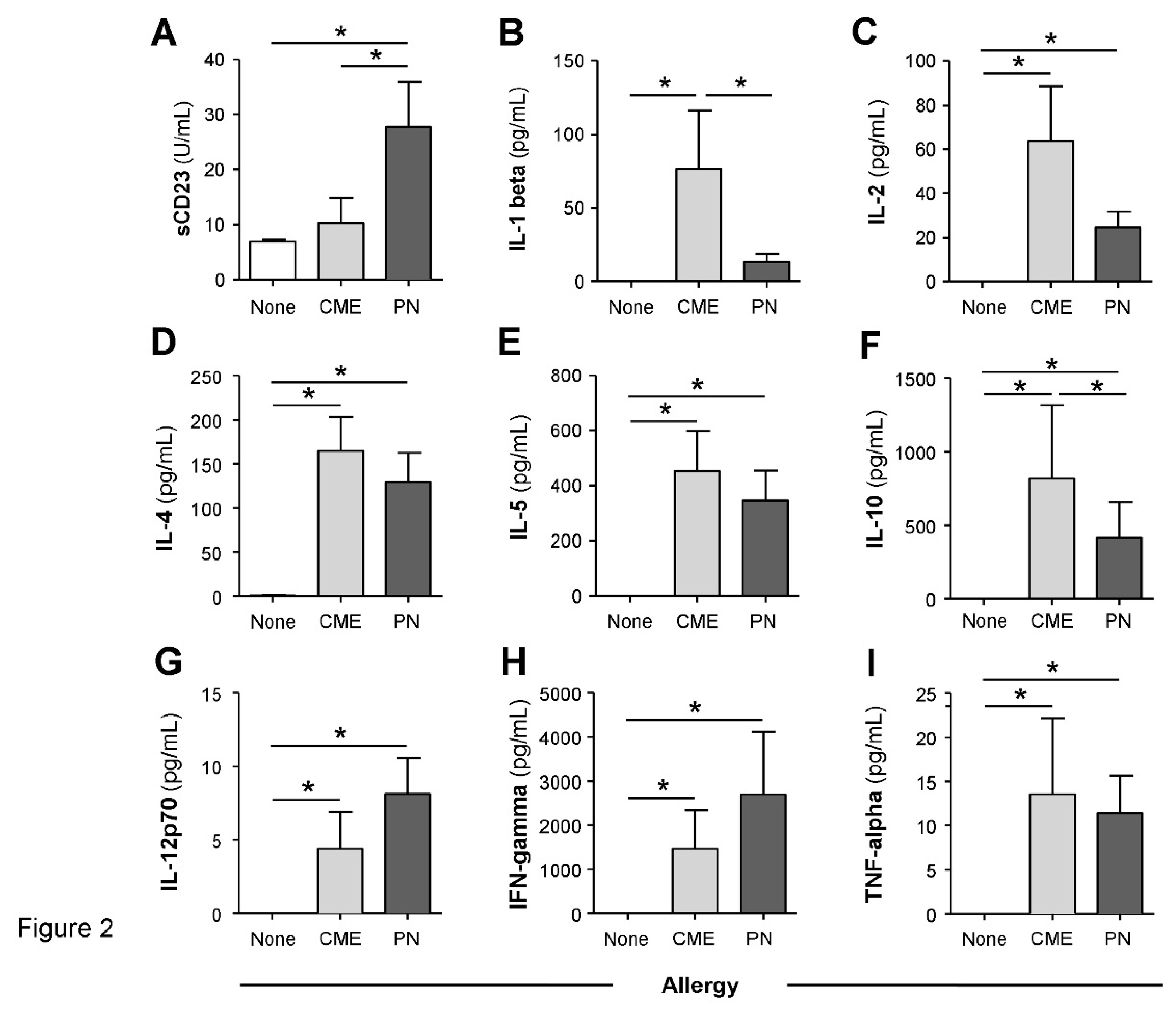
Figure 3.
Expression of costimulatory molecules. MNC from non-allergic children and those with CME or PN allergies were cultured without allergenic stimulation. At day 6 cells were harvested, washed and stained with CD80-FITC, CD83-PE, CD86-APC and HLA-DR-ECD. Data for the percentages of (A) CD80+HLA DR−, (B) CD80+ CD83+, (C) CD80+ CD86− and (D) CD86+ CD80−.were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey.
Figure 3.
Expression of costimulatory molecules. MNC from non-allergic children and those with CME or PN allergies were cultured without allergenic stimulation. At day 6 cells were harvested, washed and stained with CD80-FITC, CD83-PE, CD86-APC and HLA-DR-ECD. Data for the percentages of (A) CD80+HLA DR−, (B) CD80+ CD83+, (C) CD80+ CD86− and (D) CD86+ CD80−.were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey.
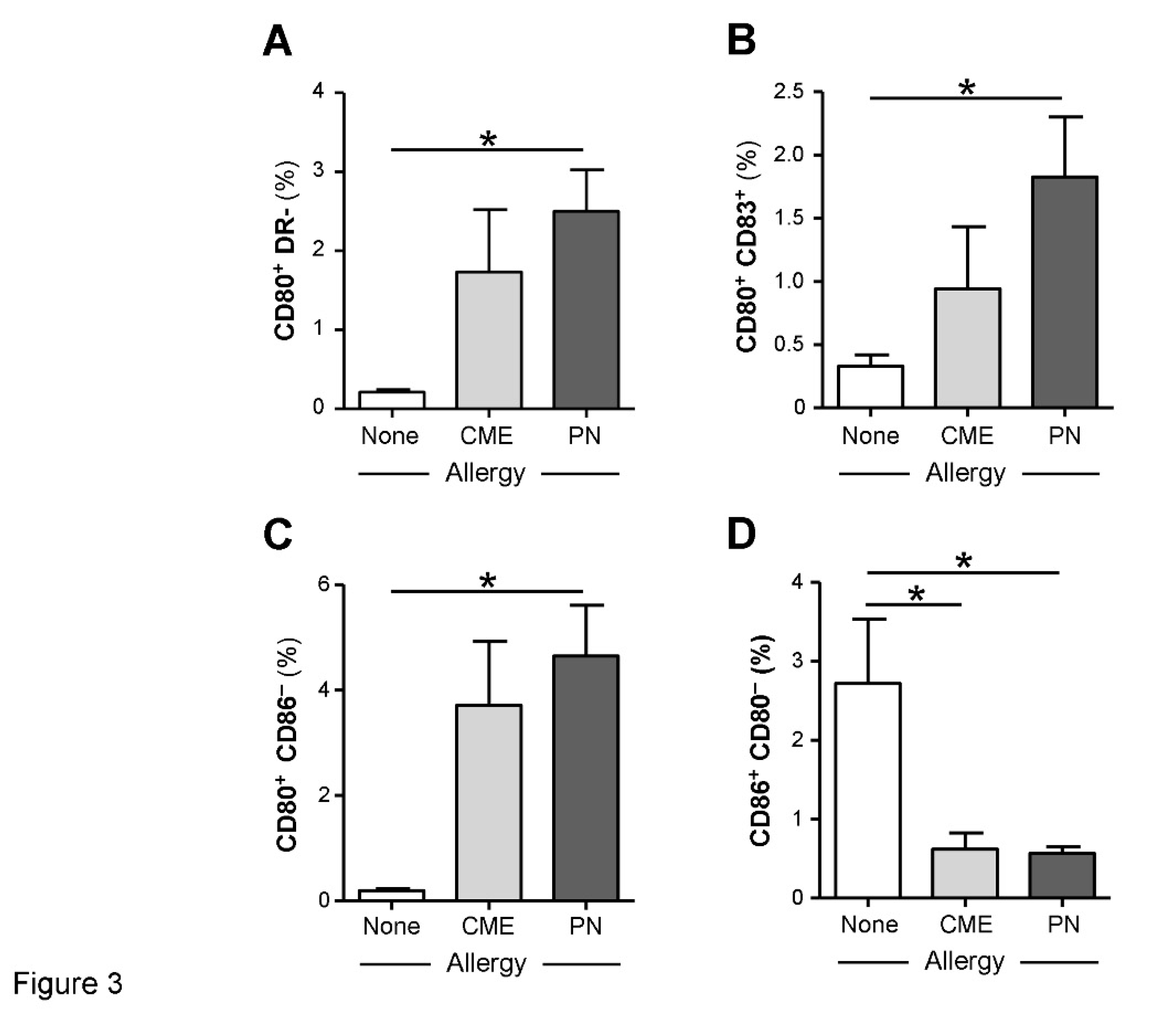
Figure 4.
Dendritic cell subset enumeration. Dendritic cells were quantified in heparinized whole blood from non-allergic children and those with CME or PN allergies. Data were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
Figure 4.
Dendritic cell subset enumeration. Dendritic cells were quantified in heparinized whole blood from non-allergic children and those with CME or PN allergies. Data were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
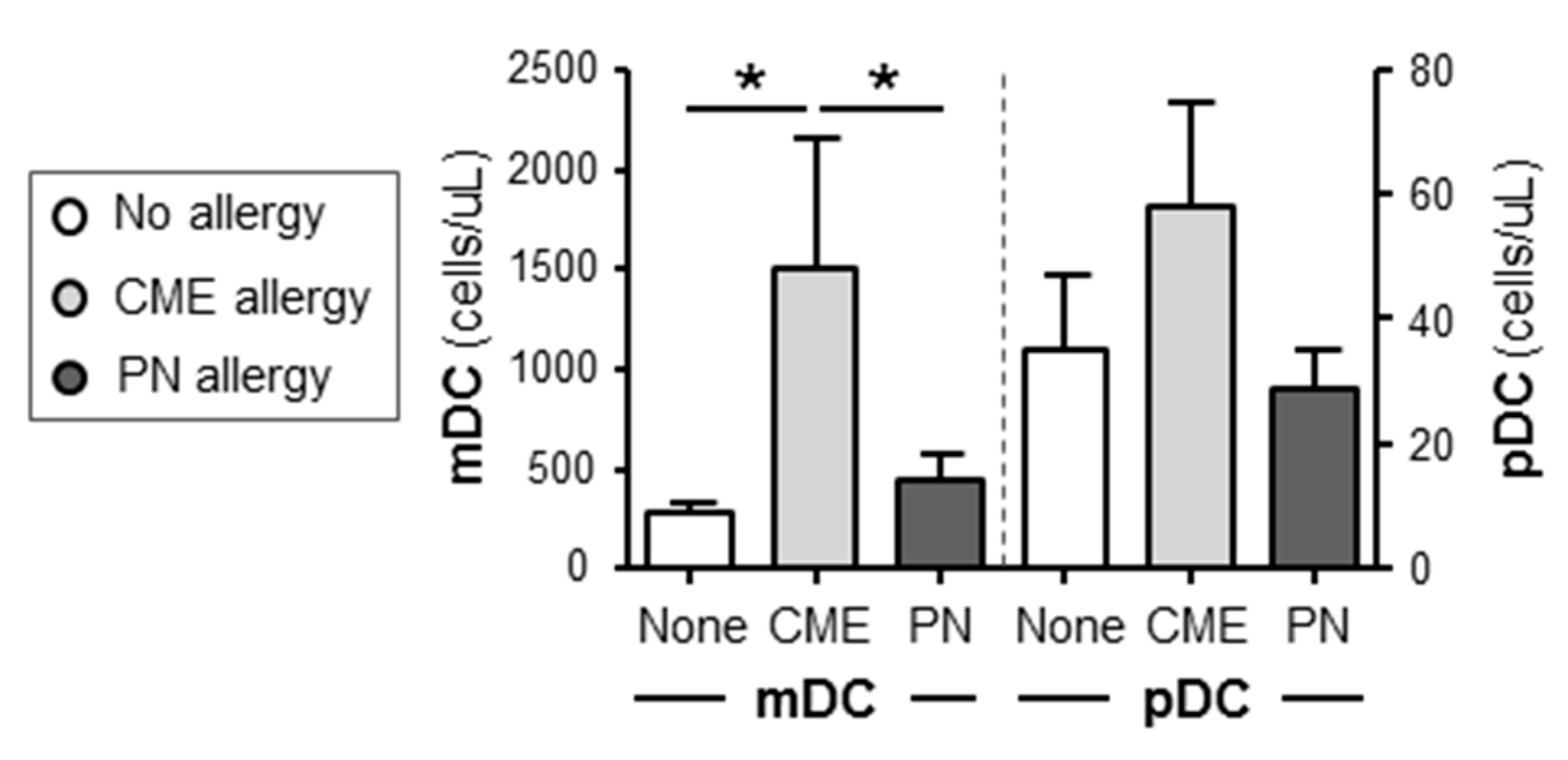
Figure 5.
Percentages of B cells, T cells, APCs and their Bcl-2 content. MNC from non-allergic children and those with CME or PN allergies were first stained with each of two panels for cell surface antigens, both containing CD69-APC, CD80-FITC, CD83-FITC and CD86-FITC except that Panel 1 had CD3-ECD and Panel 2 had CD19-ECD as lineage marker. This was followed by intracellular staining with Bcl-2-PE. Percentages of (A) B cells, B) T cells and (C) APC are shown alongside MFI values for Bcl-2 content within CD69+ (D) B cells, (E) T cells and (F) APC. All data were acquired by flow cytometry and are seen as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
Figure 5.
Percentages of B cells, T cells, APCs and their Bcl-2 content. MNC from non-allergic children and those with CME or PN allergies were first stained with each of two panels for cell surface antigens, both containing CD69-APC, CD80-FITC, CD83-FITC and CD86-FITC except that Panel 1 had CD3-ECD and Panel 2 had CD19-ECD as lineage marker. This was followed by intracellular staining with Bcl-2-PE. Percentages of (A) B cells, B) T cells and (C) APC are shown alongside MFI values for Bcl-2 content within CD69+ (D) B cells, (E) T cells and (F) APC. All data were acquired by flow cytometry and are seen as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey. .
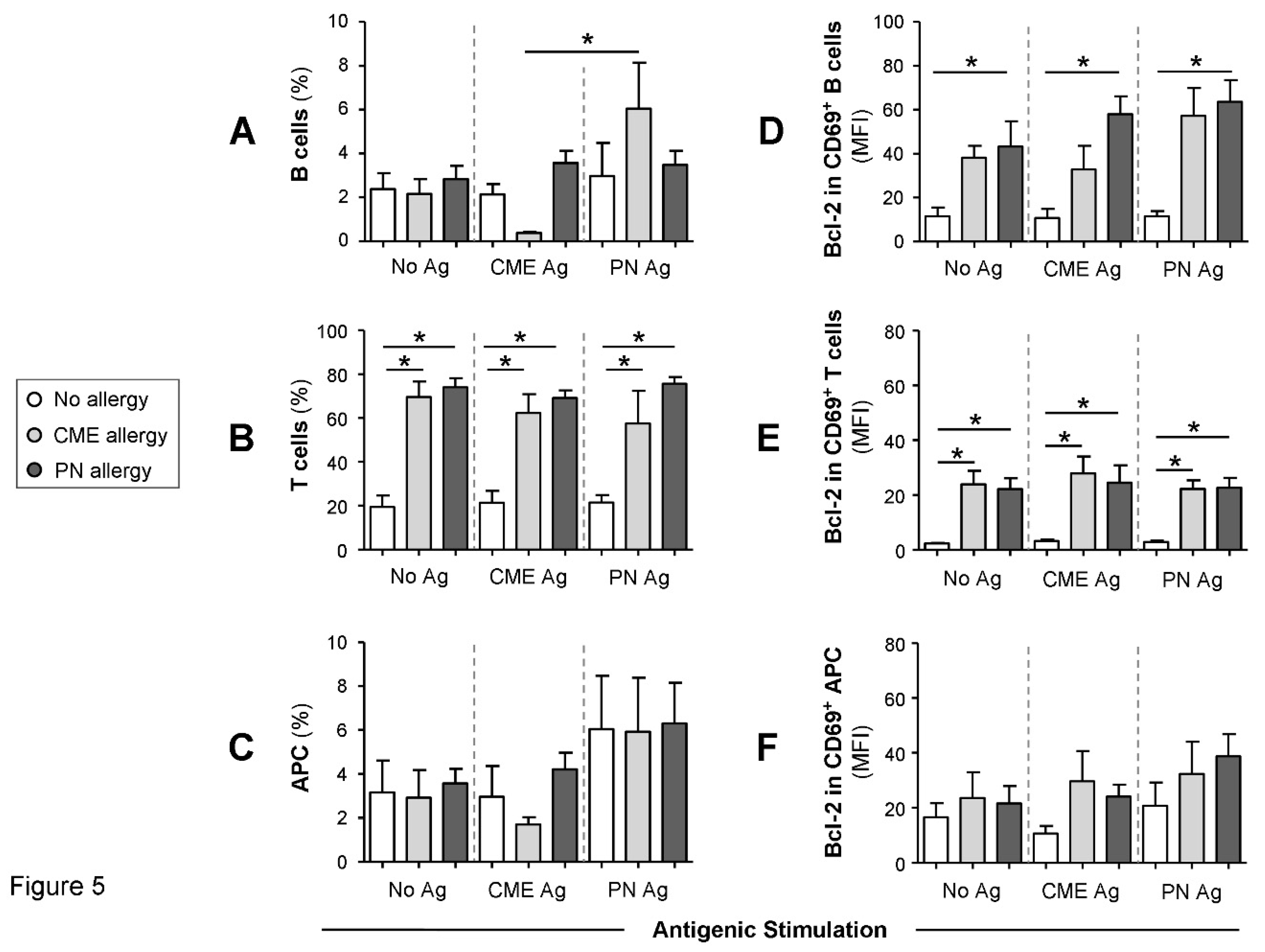
Figure 6.
Cell surface expression of CD152 within CD69+ CD4+ T cells. MNC from non-allergic children and those with CME or PN allergies were stained with CD3-FITC, CD152-PE, CD4-ECD and CD69-APC. Cells were gated using scatter profiles and CD3 expression to identify T cells. Data for the percentages of CD152+ cells within (A) CD4+ or (B) CD4+ CD69+ were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey.
Figure 6.
Cell surface expression of CD152 within CD69+ CD4+ T cells. MNC from non-allergic children and those with CME or PN allergies were stained with CD3-FITC, CD152-PE, CD4-ECD and CD69-APC. Cells were gated using scatter profiles and CD3 expression to identify T cells. Data for the percentages of CD152+ cells within (A) CD4+ or (B) CD4+ CD69+ were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey.
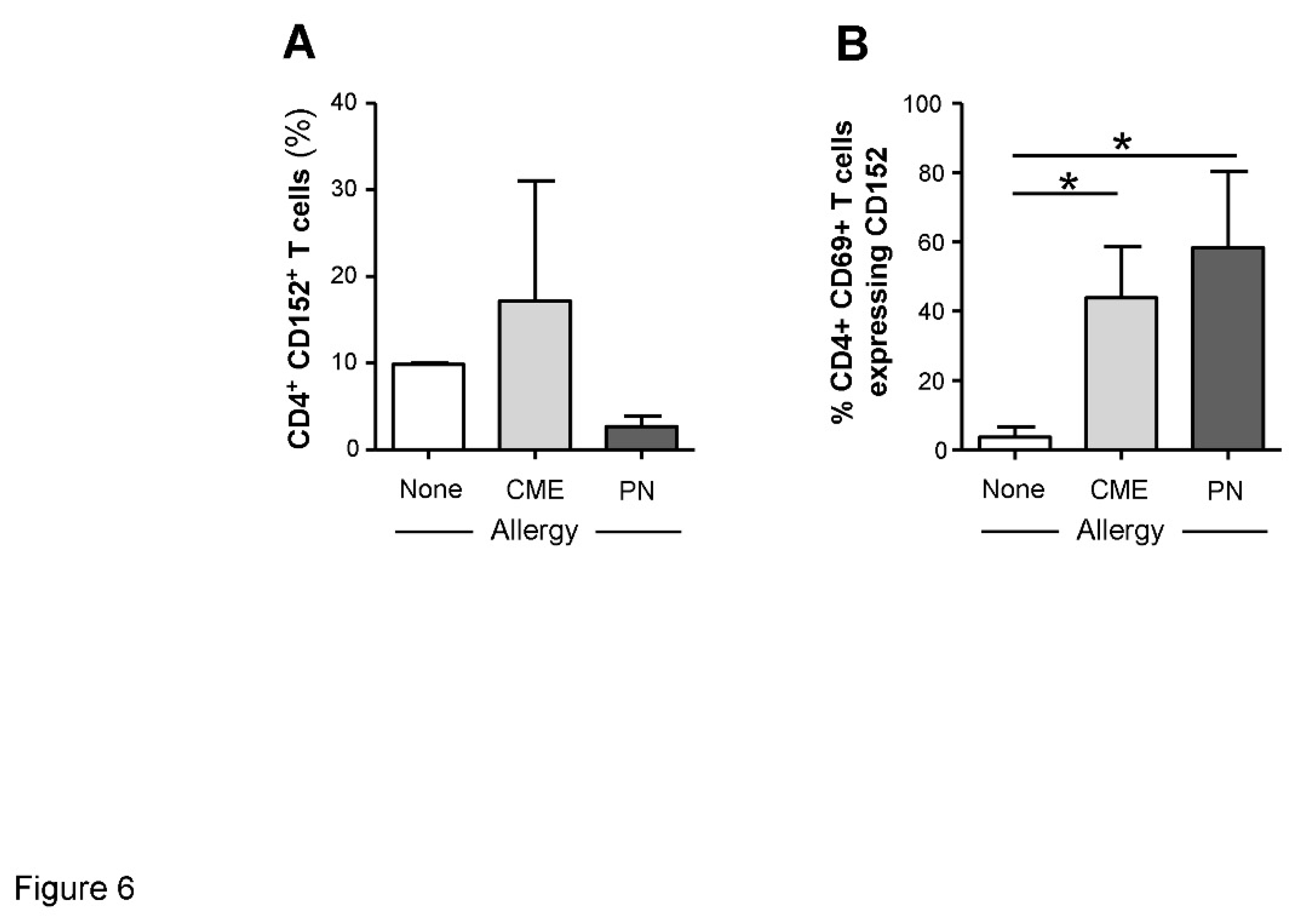
Figure 7.
Cytokine-mediated regulation of CD152 expression. MNC from non-allergic children and those with CME or PN allergies were cultured in the presence or absence of PN antigens (Ag) and with or without an IL-4 neutralizing antibody. At day 6 cells were harvested, washed and stained with CD3-FITC, CD152-PE and CD4-ECD. Data for the percentages of positive cells were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey.
Figure 7.
Cytokine-mediated regulation of CD152 expression. MNC from non-allergic children and those with CME or PN allergies were cultured in the presence or absence of PN antigens (Ag) and with or without an IL-4 neutralizing antibody. At day 6 cells were harvested, washed and stained with CD3-FITC, CD152-PE and CD4-ECD. Data for the percentages of positive cells were acquired by flow cytometry and are shown as means ± s.e.m; *P < 0.05, analyzed using Anova with adjustment for multiple comparisons using Tukey.
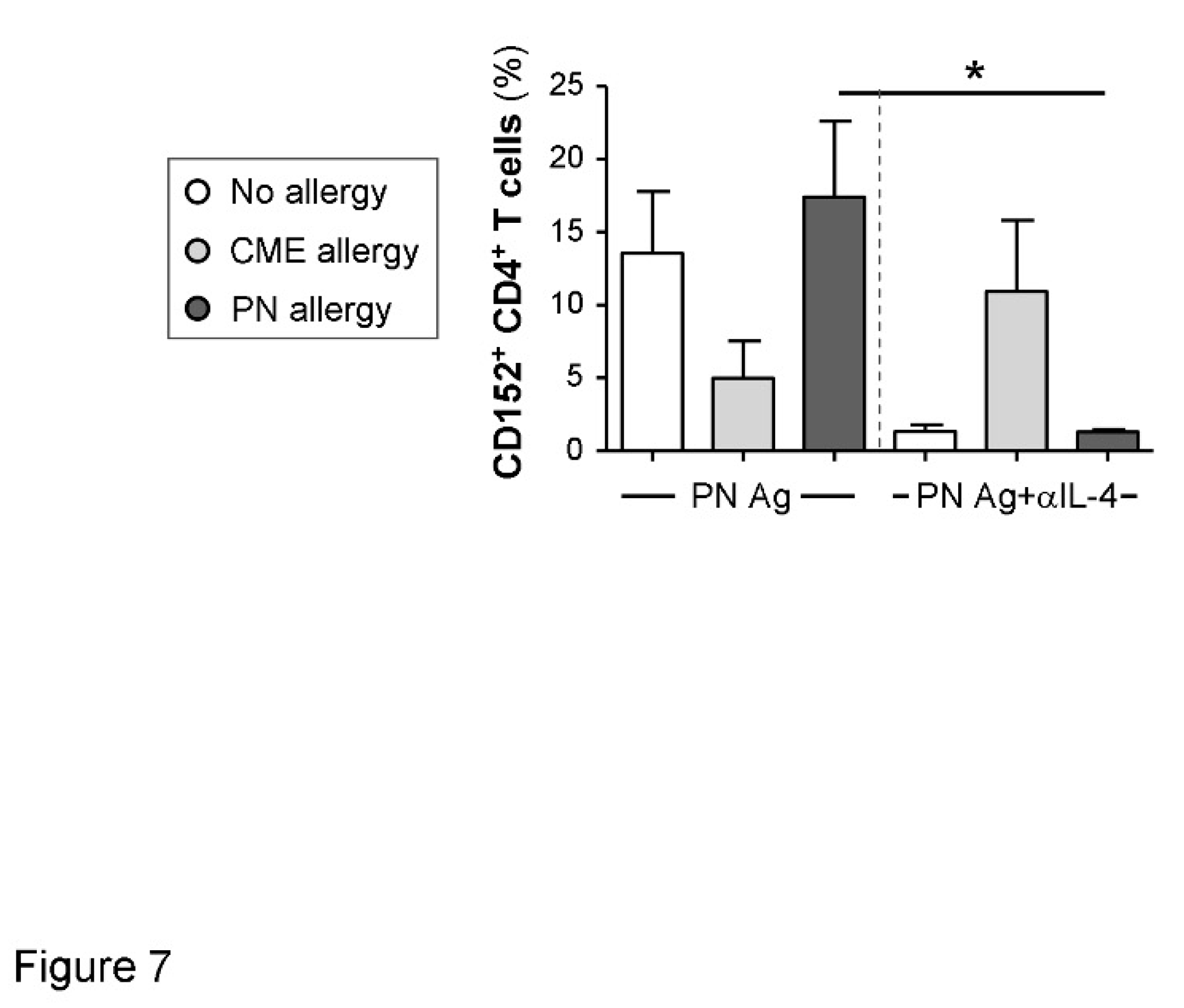
Figure 8.
Canonical coefficients of non-allergic individuals and individuals allergic to CME or PN. Linear discriminant functions were based on cytokines secreted (A) in vivo and therefore constitutively present in plasma or (B) released following 6-day MNC culture in vitro.
Figure 8.
Canonical coefficients of non-allergic individuals and individuals allergic to CME or PN. Linear discriminant functions were based on cytokines secreted (A) in vivo and therefore constitutively present in plasma or (B) released following 6-day MNC culture in vitro.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



