
Submitted:
19 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
In Benin, Xanthomonas oryzae pv. oryzae (Xoo) causing bacterial blight (BLB) was first described in 2013 on wild rice Oryza longistaminata. No studies have been conducted on Beninese Xoo strains and resistant rice varieties. The aim of this study is to check whether Xoo has already spread to cultivated varieties and to screen some rice accessions for the main resistance genes detection. To achieve this aim, rice leaves showing typical BLB symptoms were collected from various rice fields in Benin and analyzed by PCR to identify Xoo-specific sequences. In addition, seventy-five rice accessions were examined to identify Xoo resistance genes. The results showed that Xanthomonas oryzae (Xo) had been identified in Banikouara and Malanville. In addition, Sphingomonas sp was detected in several rice fields. A Xo-Sphingomonas Sp co-infection was also noted. 47 of 75 rice accessions examined (62.66%) carried Xoo resistance genes with 3 (4%) and 40 (53.33%) of xa5 and Xa21 respectively. None of the accessions had neither Xa7 nor Xa13 resistance genes. Three accessions had both xa5 and Xa21 genes. In addition, the rice isogenic lines IRBB60 and IRBB21used as positive controls, were found to be susceptible. These results indicate that Xo has evolved from a wild rice to cultivated varieties in northern Benin. Consequently, a varietal improvement program must be implemented with varieties having resistance genes to prevent a BLB pandemic in Benin.
Keywords:
Bacterial blight
; Xanthomonas oryzae pv. oryzae
; molecular characterization
; resistance genes
1. Introduction
Rice is a well-known crop around the world. It is the second-largest cereal grown with 149,000,000 ha and the third in terms of production with 380,000,000 tons, consumed and exported worlwide after wheat and maize. Asian production alone stands at 643 million tonnes (90%) while African production remains very low (24.8 million tonnes). However, this production is limited by numerous biotic and abiotic constraints. Diseases are the major biotic factors influencing rice production. Xanthomonas oryzae causes two important diseases of rice: X. oryzae pv. oryzae (Xoo) causes bacterial leaf blight (BLB), while X. oryzae pv. oryzicola (Xoc) causes bacterial leaf streak (BLS). The yield losses caused vary between 20 and 80% [1,2]. In 2003 and 2009, extensive surveys in three West African countries indicated a high incidence of BLB with race A3 reported in Mali, race A1 and A2 in Burkina Faso, and A1 also reported in Niger [1,2]. In Benin, rice consumption has become high (406,000 tons of paddy rice in 2019) and domestic production has tripled in the last ten years [3]. However, his production is unable to meet demand [4]. In Benin, Xoo was first described in 2013 on wild rice Oryza longistaminata. So far, no study has been done on Beninese Xoo strains to better characterize these strains [5].
To control BLB, cultural, chemical, and biological control approaches have been developed, but none of them has been found to be fully effective in the case of severe disease outbreaks [6,7]. The selection of disease-resistant rice varieties has been found to be the most effective, economical, and least environmentally damaging method for BLB control [8]. Various rice genes conferring resistance to Xoo have been identified genetically and designated in a series from Xa1 to Xa 45, of which nine were identified as recessively inherited. Eleven of them have been characterized as encoding different types of proteins, suggesting multiple mechanisms of R gene-mediated X. oryzae pv. oryzae resistance [9;10]. Some of them have been introgressed into elite rice varieties in Asia. Xa21, Xa5, Xa7, and Xa13, one of the most widely used genes in Asian rice breeding programs are referred to have broad-spectrum because of their resistance to several races of Xanthomonas oryzae Pv. Oryzae.
The Xa21 resistance gene located on chromosome 11 of rice and originating from the ancestral species Oryza longistaminata has attracted the attention of breeders because of its broad-spectrum resistance to to various strains of Xoo. This dominant gene provides stable and durable resistance against Xoo in northern Vietnam and India [11]. It is the most effective gene showing effective resistance to 88% of Xoo strains in India [11]. Unlike other genes that confer resistance only at the adult stage of the plant (e.g., Xa7), resistance mediated by the Xa21 gene increases gradually from the susceptible juvenile stage to full resistance at the later adult stage [12]. The Xa21 gene has shown resistance to six races of Xoo in China and has been used in a breeding program (Huang et al., 1997) to improve resistance to resistance to bacterial blight. Similarly, it was introduced into the cultivar PR106 in India because it showed resistance to 17 strains of Xoo [11]. It is considered the most effective gene for 88% of Xoo strains in India [11]. The recessive xa5 gene was identified in DZ192 [13] and is located on the short arm of chromosome 5 [14]. This gene is a natural mutation of a sensitive allele from which it differs by two nucleotide substitutions to cause the change of a single amino acid at position 39, i.e. glutamine (E) instead of valine (V) (V39E). This mutation confers resistance to a wide range of Xoo strains [15]. Thus, the xa5 gene is one of the most widely used genes in breeding programs in Asia and confers resistance to several strains and races of Xanthomonas oryzae pv. Oryzae. Most of the resistance genes identified do not confer durable resistance, particularly in the current context of global warming [16]. High temperatures reduce the resistance of certain rice varieties to Xoo, such as those carrying the Xa4 gene [17]. On the other hand, the IRBB7 line, which carries the Xa7 gene, shows more effective resistance at high temperatures [17]. In addition, studies have shown that the dominant Xa7 gene confers long-lasting, broad-spectrum resistance to Xoo [16]. The development of this gene in Beninese rice cultivars will therefore help to improve rice production under these changing climatic conditions. The recessive xa13 gene is one of the most widely used genes in Asian breeding programs [18]. This gene induces resistance to the most virulent race of Xoo, unlike most of the R genes reported [19]. These genes can be integrated into sustainable management strategies for vascular bacterial disease of rice [20]. Identifying these genes would therefore represent a crucial step in developing cultivars resistant to Xoo. In Benin, we do not know whether the pathogen has already passed into the grown rice varieties, or if rice varieties are exposed to other bacteria such as Sphingomonas. Whereas the use of resistant varieties, carrying resistance genes, is the only highly effective and environmentally friendly way to control this disease [21], no information is available on these Xoo resistance genes in rice varieties grown in Benin apart from the one we recently initiated. This study aims to identify Beninese Xoo strains, causing vascular bacterial leaf, and screen rice varieties grown in Benin for the major BLB resistance genes Xa5, Xa7, Xa13 and Xa21 genes in order to help ensure food security.
2. Materials and Methods
2.1. Study Area and collection of diseased leaves
Sixty-nine diseased leaves showing typical symptoms of leaf blight, notably yellow-brown discoloration along one of the two leaf blades, changing to brown to dark brown with age, were collected in the three phytogeographical zones of Benin where rice is mainly produced (Guinean, Sudano-Guinean and Sudanese zones). Samples were collected using the random sampling method, from rice plants at the heading and approaching maturity stages, as the disease usually develops well at these stages of plant growth. Diseased leaves were detached and placed in a paper envelope. These envelopes were labelled with the variety name, location, sampling date, rice ecosystem, and geographic coordinates. The samples were finally transported to the laboratory and stored in the refrigerator for further processing. On the other hand, our collection of seventy-six rice accessions grown in Benin and three positive controls IRBB5, IRBB21, and IRBB60 were screened for the resistance genes.

Table 1.
List of rice accessions used in the study.
| No | Accession code | Department/Town/Village | No | Accession code | Department/Town/Village |
|---|---|---|---|---|---|
| 1 | Bagou 18 | Alibori/Gogounou/Bagou | 46 | Nana30 | Atacora/Cobly/Nanagadé |
| 2 | Gou10 | Alibori/Karimama/Gouroubéri | 47 | Bagou 21 | Alibori/Gogounou/Bagou |
| 3 | Tchaka38 | Atacora/Touncoutouna/Tchakalakou | 48 | Koung65 | Atacora/Wassa Pehonco/Koungarou |
| 4 | Kotch70 | Atacora/Tanguiéta/Kotchessi | 49 | Doko122 | Atlantique/Abomey-Calavi/Dokomey |
| 5 | Tchaka41 | Atacora/Touncoutouna/Tchakalakou | 50 | Kik96 | Donga/Bassila/Kikélé-Lokpa |
| 6 | Kan58 | Atacora/Matéri/Kankini-Séri | 51 | Nippon bar | Alibori/Kandi/Angaradébou |
| 7 | Bagou 25 | Alibori/Gogounou/Bagou | 52 | Ang 6 | Alibori/Kandi/Angaradébou |
| 8 | Gami74 | Borgou/Bembèrèkè/Gamia | 53 | Bagou 23 | Alibori/Gogounou/Bagou |
| 9 | Nana32 | Atacora/Cobly/Nanagadé | 54 | Bagou 28 | Alibori/Gogounou/Bagou |
| 10 | Tchaka33 | Atacora/Touncoutouna/Tchakalakou | 55 | Ang 2 | Alibori/Kandi/Angaradébou |
| 11 | Bagou 19 | Alibori/Gogounou/Bagou | 56 | Tog 5307 | Alibori/Kandi/Angaradébou |
| 12 | Okouta98 | Collines/Bantè/Okouta-Ossè | 57 | Koum53 | Atacora/Boukoumbé/Koumadogou |
| 13 | Kan61 | Atacora/Matéri/Kankini-Séri | 58 | IR64 | Atlantique/Zè/Awokpa |
| 14 | Gou11 | Alibori/Karimama/Gouroubéri | 59 | Gami76 | Borgou/Bembèrèkè/Gamia |
| 15 | Bagou 17 | Alibori/Gogounou/Bagou | 60 | Koud43 | Atacora/Natitingou/Koudengou |
| 16 | Bori84 | Borgou/N’dali/Bori | 61 | Agbab 101 | Collines/Savè/Agbaboué |
| 17 | Koud45 | Atacora/Natitingou/Koudengou | 62 | Gou12 | Alibori/Karimama/Gouroubéri |
| 18 | Kpatab100 | Collines/Savalou/Kpataba | 63 | Ang 16 | Alibori/Kandi/Angaradébou |
| 19 | Koum54 | Atacora/Boukoumbé/Koumadogou | 64 | Tchaka39 | Atacora/Touncoutouna/Tchakalakou |
| 20 | Bagou 22 | Alibori/Gogounou/Bagou | 65 | Koud42 | Atacora/Natitingou/Koudengou |
| 21 | Kan60 | Atacora/Matéri/Kankini-Séri | 66 | Foun15 | Alibori/Banikoara/Founougo |
| 22 | Kan59 | Atacora/Matéri/Kankini-Séri | 67 | Tot82 | Borgou/Nikki/Totorou |
| 23 | Bagou 26 | Alibori/Gogounou/Bagou | 68 | Nana29 | Atacora/Cobly/Nanagadé |
| 24 | Kotch71 | Atacora/Tanguiéta/Kotchessi | 69 | Bori83 | Borgou/N’dali/Bori |
| 25 | Bagou 24 | Alibori/Gogounou/Bagou | 70 | Kotch73 | Atacora/Tanguiéta/Kotchessi |
| 26 | Koum47 | Atacora/Boukoumbé/Koumadogou | 71 | Moroberekan | Atacora/Tanguiéta/Kotchessi |
| 27 | Koud46 | Atacora/Natitingou/Koudengou | 72 | Koum55 | Atacora/Boukoumbé/Koumadogou |
| 28 | Man118 | Mono/Houéyogbé/Manonkpon | 73 | Koud 44 | Atacora/Natitingou/Koudengou |
| 29 | Tchaka36 | Atacora/Touncoutouna/Tchakalakou | 74 | Koum 51 | Atacora/Boukoumbé/Koumadogou |
| 30 | Bagou 27 | Alibori/Gogounou/Bagou | 75 | IR841 | INRAB |
| 31 | Koum49 | Atacora/Boukoumbé/Koumadogou |
Positif controls |
IRBB60 | |
| 32 | NERICA19 | Atacora/Matéri | IRBB5 | ||
| 33 | Okouta97 | Collines/Bantè/Okouta-Ossè | IRBB21 | ||
| 34 | Koung69 | Atacora/Wassa Pehonco/Koungarou | |||
| 35 | ONK93 | Donga/Djougou/Onklou | |||
| 36 | Kotch72 | Atacora/Tanguiéta/Kotchessi | |||
| 37 | Tchal89 | Donga/Ouaké/Tchalinga | |||
| 38 | Dev116 | Couffo/Dogbo/Dévé | |||
| 39 | Koung67 | Atacora/Wassa Pehonco/Koungarou | |||
| 40 | Ang1 | Alibori/Kandi/Angaradébou | |||
| 41 | Koum50 | Atacora/Boukoumbé/Koumadogou | |||
| 42 | ONK93b | Donga/Djougou/Onklou | |||
| 43 | Gami77 | Borgou/Bembèrèkè/Gamia | |||
| 44 | Bagou 20 | Alibori/Gogounou/Bagou | |||
| 45 | 6R2B9 | Alibori/Gogounou/Bagou | |||
2.2. Sowing and sampling of rice leaves for DNA extraction.
Five Paddy rice seeds of each variety of our collection were grown in germination pots in a greenhouse with a controlled environment for better growth conditions. They were watered as required for 10 to 15 days. The youngest leaves at the seedling stage (21 days) were removed with scissors and then wrapped in aluminium foil on which the pot references (name and code of the sample as well as the sampling date) were written. The samples were then stored in the refrigerator at 40°C.
2.3. DNA Extraction from collected leaves showing BLB symptoms and from leaves of cultivated rice accessions of the laboratory of molecular biology and bioinformatics applied to genomic.
To extract total genomic DNA from the leaves, the CTAB protocol was performed according to the protocol of Djedatin et al. [22]. Briefly, 200mg of infected rice leaf was weighed and then ground in porcelain mortars with 1000µL of CTAB pre-warmed to a temperature of 65°C. The crushed material was then transferred to a 2ml eppendorf tube, followed by the addition of 50µL of SDS and homogenization. The samples were incubated in an oven at 65°C for 45 minutes and then allowed to cool to room temperature. Then 750µL of CIA (Chloroform Isoamyl Alcohol) was added to each tube followed by gentle inversion shaking for 5 minutes, then centrifuged at 10000rpm for 15 minutes. The supernatant from each sample was collected in a 1.5 ml Eppendorf tube bearing the same label. DNA precipitation was carried out by adding 800µL of isopropanol at cold - 20°C with gentle homogenization by inversion. This was followed by centrifugation at 10,000 rpm for 10 minutes. The aqueous solution was carefully removed so that the pellet was not damaged or lost. The pellet was then washed with 500 µL of 70% ethanol followed by centrifugation at 10000rpm for 10 minutes: this was repeated three times to purify the DNA pellet. The tubes were opened and dried on blotting paper for a good while (overnight). Finally, a suspension of the dried of dried DNA was made by adding 100µL of pure, sterile H2O, then stored at -20°C.
2.4. Control of total genomic DNA quality by electrophoresis
The quality of the DNA was checked by electrophoresis on a 1% agarose gel. Indeed, a mixture of 3 µL of extract of Total DNA and 8µL of 2X loading blue was migrated at 100V for 30 min in Tris Bromate EDTA buffer (TBE). After migration, the gel was put in ethidium bromide (0.1% BET) solution for 15 min then rinsed with distilled water for five min. Then, the gel was visualized on a UV trans-illuminator. Finally, dilution of the DNA was carried out and stored at 4°C for subsequent tests of DNA.
2.5. Polymerase chain reaction (PCR) design methodology for Xanthomonas oryzae pv Oryzae identification.
Bacteria were identified on the leaves showing symptoms of BLB by amplification of X. oryzae and X. oryzae pv orysae Hypothetical protein specific sequences using specific primers [23] listed in Table 2. Samples of diseased leaves with a sequence of 331 Pb different from the expected 272 Pb of Xo are tested with primers specific to Sphingomonas Sp. [24] since previous studies have identified a sequence of approximately 270 Pb which corresponds to a sequence specific to sphingomonas sp .The technique used is based on the PCR protocol described by Fanou et al [25]. PCR was performed using a thermal cycler in 20 µL volumes containing a mixture of 1 × PCR buffer, 0.1 mM dNTPs, 0.2 μM each of forward and reverse primers, 50–200 ng genomic DNA, 2.0–3.0 U of Taq DNA polymerase, with sterilized distilled water added to 50 μL. The thermal cycler program consisted of 95 °C for 5 minutes followed by 25 cycles of 1 min at 94 °C, 1 min at 54 °C and 2 min at 72 °C, and a final extension step at 72 °C for 5 min for X. oryzae. The thermal cycler program consisted of 95 °C for 5 minutes followed by 25 cycles of 1 min at 94 °C, 1 min at 52 °C and 2 min at 72 °C, and a final extension step at 72 °C for 5 min for X. oryzae pv oryzae. For Sphypingomonas sp, the thermal cycler program Composed of 95 °C for 5 minutes followed by 25 cycles of 1 min at 94 °C, 1 min at 55 °C and 2 min at 72 °C, and a final extension step at 72 °C for 5 min.
2.6. Molecular screening of rice accessions cultivated in Benin for the detection of resistance genes to bacterial leaf blight caused by Xanthomonas oryzae pv Oryzae
Polymerase chain reactions (PCR) were performed to identify resistance gene(s) among the selected accessions. Six co-segregating STS markers tightly linked to xa5, Xa7, xa13, and Xa21 [26,27,28,29] were used (Table 3) for the detection of bacterial blight-resistant gene(s). Amplification was carried out in a thermal cycler according to the following programme: pre-denaturation at 95°C for 5min followed by 35 cycles consisting of denaturation at 94°C for 1min, hybridisation at 51°C (for the RM122 marker), 59°C (for the M5 and xa13-prom markers), 55°C and 57°C (for the pTA248 marker) for1min, elongation at 72°C for 2min and final extension at 72°C for 5min.
2.7. Electrophoresis of PCR products
The PCR products and a marker with a molecular weight of 100bp were subjected to agarose gel electrophoresis (at 1.5% for the pTA248 marker, 2% for M5 and xa13-prom and 3% for the RM122 marker) run in a solution of TBE (diluted 0.5X) at 120V for 35min. The gel was then subjected to UV light on a Transilluminator for visualization.
2.8. Data analysis
The bands of the different amplicons were compared with those of the positive controls. Data were scored as ≪+ ≫ and ≪- ≫ signifying the presence and absence of the target genes, respectively. The Excel 2016 MSO spreadsheet (version 2209 Build 16.0.15629.20200) was used to make graphs to better analyze the data.
3. Results and discussion
Bacterial blight is one of the most devastating diseases of rice worldwide and is found both in tropical and temperate regions. Bacterial leaf blight is caused by Xanthomonas oryzae pv. oryzae (Xoo). The disease is ravaging Benin's neighbouring countries, such as Côte d'Ivoire, Niger, Mali and Burkina Faso. In Benin, although the strain was reported in 2013 on the wild variety Oryza longistaminata, none study has been carried out in the country to check whether it has spread to currently cultivated varieties. In this study, leaves showing symptoms of BLB were analyzed by PCR for Xoo-specific sequences. Xanthomonas oryzae was identified in Banikouara and Malanville (Alibori) with the presence of 331bp bands specific to the Hypothetical protein of Xo. For certain leaves collected in the north and south of the country, bands of 272 bp different from those expected were identified on the agarose gel. Since the sequencing of amplicons of this size enabled kini et al [24] to reveal a sequence specific to Sphingomonas sp in their study, we used primers specific to Sphingomonas sp to screen these samples. This confirmed sphingomonas sp (Figure 1 and Figure 2). The test did not reveal the specific Xoo band. This can be explained by the long storage time of the leaves in the freezer. Amplicon sequencing is required to clarify the pathovar of Xo.
The use of varieties carrying resistance genes has been considered to be the best method of controlling vascular bacterial disease in rice. The present study consisted of screening of 75 rice accessions collected in Benin in order to identify those possessing the xa5, Xa7, xa13 and Xa21 genes. Microsatellites markers were used because of their proven effectiveness in numerous studies. Indeed, the use of these markers enabled Djedatin et al. [22] to determine genetic diversity between rice varieties grown in the Atacora region. A similar study has enabled to identify and differentiate Xoo-susceptible and Xoo-resistant individuals [30]. The results of these authors show that microsatellites are the markers of choice for these studies. In our work, the markers M5 (XA7), xa13-prom (xa13), pTA248 (XA21) and RM 122 (xa5) were used. Molecular screening of the 75 rice accessions collected in Benin and the three positive controls IRBB5, IRBB21 and IRBB60 revealed the presence of three alleles of the Xa21 gene, two alleles of the xa13 gene, one allele of the Xa7 gene and two alleles of the xa5 gene (Figure 3On the one hand, there are Xoo sensitivity alleles (S) with bands of 730bp for Xa21, 220bp for xa13, 1170bp for Xa7 and 230bp for xa5. On the other hand, we observed Xoo resistance alleles (R) with bands of 1100bp or 925bp for Xa21, 450bp for xa13 and 240bp for xa5. (Figure 3, Table 4). These different band sizes obtained for each of the markers used indicate the polymorphism of the microsatellites. Similar results were obtained by [27,28,31], who used the same markers to amplify fragments of similar size. The results of the present study thus confirm the efficiency of the M5, xa13-prom, pTA248, and RM 122 markers in the selection of BLB-resistant rice varieties in Benin.
This study showed that 47 (62.66%) accessions were resistant to Xoo, including 3 (4%) homozygous for xa5 resistance allele and 40 (53.33%) homozygous and heterozygous for the Xa21 resistance allele On the other hand, 31 (41.33%) had only the 730bp band, 66 (88%) had at least the 230bp band and 75 (100%) had only the 1170bp and 220bp bands, corresponding to the susceptibility alleles indicating the susceptibility of these varieties to the pathogen (Figure 3, Table 4). Similar results were obtained by Dossa et al. [33] who, using the same M5 marker, did not obtain the XA7 resistance allele in any of the O glaberrima accessions tested. Moreover, the xa13 resistance allele was also absent in all the O. glaberrima accessions and they obtained only individuals with the Xa13 susceptibility allele [33], suggesting that O. glaberrima lacks the resistance allele of the Xa7 and Xa13 genes. Our results therefore concur with those of this author and suggest that the accessions tested are probably O. glaberrima. However, these results differ from those of Ullah et al. [34], who obtained high polymorphism in the 57 cultivars tested. This result can be explained by the fact that the accessions used in these studies were different. Among the 47 resistant accessions, Bagou22, Bagou24, Kpatab 100 and Foun15 carry both the xa5 and Xa21 genes (Table 4). The combination of these two genes would confer maximum and long-lasting resistance against a broad spectrum of Xoo pathotypes [35]. The Tchaka36 variety did not show any alleles of the Xa21 gene. Similarly, varieties such as Koum50 and Bagou28 did not show either allele of the xa5 gene (Table 4). This can be explained by the fact that the xa5 and Xa21 genes are not present in all the rice varieties or accessions grown in Benin.
Furthermore, the isogenic line IRBB21 supposed to be positive control in the present study and which according to the literature [33] should present a band of 1100bp or 925bp corresponding to the resistance allele[36], presented sensitivity allele with a band of 730bp (Figure 3D, Table 4). Similarly, the isogenic line IRBB60, considered as a positive control for the xa5 and xa13 genes, showed two bands for each gene instead of just one (Figure 3a and c,Table 4).This result therefore indicates that the IRBB60 line used in this study could be susceptible to vascular bacterial disease in a real environment and cannot be considered as a positive control for the xa5 and xa13 genes. However, it is possible that this is a second resistance allele or another allele that is not involved in BLB resistance. These results differ from previous studies [37,38] which only obtained the 240 bp band for xa5 and the 450 bp band for xa13 respectively. These results can be explained by gene flow during seed multiplication in the field, a problem with the conservation (purity) of these lines, or genetic drift resulting in the loss of the xa5, xa13 and Xa21 resistance alleles by the isogenic lines IRBB60 and IRBB21 respectively. Crossing over events could therefore have generated individuals lacking the gene.
Amplification of Xa21 performed at 55°C produced a multitude of bands of around 500bp and 400bp presented by accessions 4 and 47. On the other hand, amplification of the DNAs of these same individuals at a temperature of 57°C produced only the expected bands. These results suggest that increasing the temperature eliminated all the non-specific sequences that were amplified (Figure 4). In fact, these 500bp bands obtained by [39] in a similar study after an amplification at 55°C and considered as bands corresponding to the sensitivity allele, would be due to a non-specific amplification.
In total, Xo strains must be isolated from leaves and sequenced for better identification of Xoo. In view of the results, there is considerable genetic diversity in resistance to bacterial blight in Benin. Pathogenicity testing of Xoo strains from Benin and those from Burkina Faso, Niger, and Ivory Coast on the resistant accessions identified is necessary to assess the expression of this resistance gene in a real environment and above all the behavior of heterozygous individuals to analyze their resistance level to the plant pathogen
4. Conclusions
This study reveals the presence of Xo in accessions in northern Benin, suggesting a possible spread of the Oryza longistaminata pathogen to cultivated species. It is also clear that Sphingomonas sp infection, which also causes rice leaf scorch, is also taking hold in the country. The SSR markers used have proved effective in identifying cultivars carrying multiple resistance genes, since the epistasis effect has rendered the conventional approach ineffective. The results will be very useful to rice breeders as they will be able to develop resistant varieties easily. Accessions possessing both the xa5 and Xa21 genes can be directly disseminated in Beninese agriculture or used as donor parents for the development of sustainable bacterial-resistant varieties in breeding programmes after confirmation by phenotypic screening. The data obtained reports the absence of two major resistance genes, Xa7, and xa13, which are effective against a broad spectrum of Xoo pathotypes in Beninese rice production. It would be important to introgress these two genes into elite rice varieties produced in Benin in perspective to provide effective control of BLB in the event of an epidemic in the country.
Author Contributions
Conceptualization, C. N. M.N.,L.G.D.; methodology, C.N.M.N,L.G.D., K,D,H.M., A. A. M., K.L. D., B. M.P. F. L., B.W. D. F. B., E.Z., R.G.F. H. and A.H.; Validation, C. N. M.N.,L.G.D., K,D,H.M.,A. A. M. and L.S.B.; funding acquisition, C. N. M.N. and L.G.D.; Investigation, C. N. M.N., K.L. D., B. M.P. F. L., B.W. D. F. B., E.Z., R.G.F. H. and A.H.; data interpretation, C. N. M.N.,L.G.D., K,D,H.M., A. A. M., K.L. D., B. M.P. F. L., B.W. D. F. B., E.Z., R.G.F. H. and A.H.; writing—original draft preparation, C. N. M.N., draft review, L.G.D. ; supervision, L.G.D.; K,D,H.M.,A. A. M. and L.B.-M.; project administration, C. N. M.N.,L.G.D.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received financial support from The World Academy of Sciences (TWAS).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors thank all the research team of the Molecular Biology and Bioinformatics Applied to Genomics Laboratory.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sere, Y.; Onasanya, A.; Verdier, V.; Akator, K.; Ouedraogo, L.S.; Segda, Z.; Mbare, M.M.; Sido, A.Y.; Baso., A. Rice bacterial leaf blight in West Africa: preliminary studies on disease in farmer’s field and screening. Asian J. Plant Sci. 2005, 4, 577–579. [Google Scholar] [CrossRef]
- Gonzalez, C.; Szurek, B.; Manceau, C.; Mathieu, T.; Séré, Y.; Verdier, V. “Molecular and Pathotypic Characterization of New Xanthomonas oryzae Strains from West Africa.”. Mol. Plant Microbe Interact. 2007, 20(5), 534–546. [Google Scholar] [CrossRef]
- FAO; 2020. Available: www.fao.org com. Accessed on 05/12/2023.
- Pandey, M. K.; Shobha R., N.; Sundaram, R.; Laha, G.; Madhav, M.; Srinivasa R., K.; Sudharshan, I.; Hari, Y.; Varaprasad, G.; Subba R., L. Improvement of two traditional Basmati rice varieties for bacterial blight resistance and plant stature through morphological and marker-assisted selection. Mol. Breed. 2013, 31, 239-246. [Google Scholar] [CrossRef]
- Afolabi, O.; Amoussa, R.; Bilé, M.; Oludare, A.; Gbogbo, V.; Poulin, L.; Koebnik, R.; Szurek, B.; Silué, D. First Report of Bacterial Leaf Blight of Rice Caused by Xanthomonas oryzae pv. oryzae in Benin. Plant Dis. 2016, 100, 515. [Google Scholar] [CrossRef]
- Islam, Md. R.; Alam, Md. S.; Khan, A. I.; Hossain, I.; Adam, L. R.; Daayf, F. “Analyses of Genetic Diversity of Bacterial Blight Pathogen, Xanthomonas oryzae pv. oryzae Using IS1112 in Bangladesh.” C.R. Biologies 2016, 399–407.
- Mundt, C.C. “Durable Resistance: A Key to Sustainable Management of Pathogens and Pests.”Infect. Genet. Evol. 2014, 27 446–55. [CrossRef]
- Verdier, V. Characterization of new races of Xanthomonas oryzae pv. Oryzae in Mali informs resistance gene deployment. Phytopathology 2020, 110(2), 267-277.
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G. L.; White, F. F.; Yin, Z. R. gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Fu, B.; Yang, H.; Xu, C.; Li, Z; Sanchez, A. Targeting xa13, a recessive gene for bacterial blight resistance in rice. Theor. Appl. Genet 2006, 12, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.S.; Rao, Y.P.; Mohan, S.K.; Verma, J.P. Role of Leptocorisaacuta Thun in the spread of bacterial blight of rice. Curr. Sci. 1976, 45, 11, p 426 427. [Google Scholar]
- Furuya, N.; Taura, S.; Goto., T.; Thuy, B.T.; Ton, P.H.; Tsuchiya, K.; Yoshimura, A. Diversity in virulence of Xanthomonas oryzae pv. oryzae from Northern Vietnam. Jpn. Agric. Res. Q. 2012, 46(4), 329–338. [Google Scholar] [CrossRef]
- Petpisit, V.; Khush, G.S.; Kauffman, H.E. Inheritance of Resistance to Bacterial-Blight in Rice. Crop Sci. 1977, 17, 551–554. [Google Scholar] [CrossRef]
- Blair, M. W.; Garris, A. J.; Iyer, A. S.; Chapman, B.; Kresovich, S.; McCouch, S.R. 2003: High-resolution genetic mapping and candidate gene identification at the xa5 locus for 44 bacterial blight resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2003, 107, 62–73. [Google Scholar] [CrossRef]
- Jiang, G.H.; Xia, Z.H.; Zhou, Y.L.; Wan, J.; Li, D.Y.; Chen, R.S.; Zhai, W.X.; Zhu, L.H. Testifying the rice bacterial blight resistance gene xa5 by genetic complementation and further analyzing xa5 (Xa5) in comparison with its homolog TFIIAγ1. Mol. Genet. Genom 2006, 275(4), 354–366. [Google Scholar] [CrossRef]
- Chen, X.; Liu, P.; Mei, L.; He, X.; Chen, L.; Liu, H.; Shen, S.; Ji, Z.; Zheng, X.; Zhang, Y. (2021. Xa7, a new executor R gene that confers durable and broad-spectrum resistance to bacterial blight disease in rice. Plant Comm. 2021, 2(3), 100143. [Google Scholar] [CrossRef]
- Dossa, G. S.; Quibod, I.; Atienza-Grande, G.; Oliva, R.; Maiss, E.; Vera Cruz, C.; Wydra, K. Rice pyramided line IRBB67 (Xa4/Xa7) homeostasis under combined stress of high temperature and bacterial blight. Sci. Rep 2020, 10(1), 683. [Google Scholar] [CrossRef] [PubMed]
- Khan, M. A.; Naeem, M.; Iqbal, M. Breeding approaches for bacterial leaf blight resistance in rice (Oryza sativa L.), current status and future directions. Eur. J. Plant Pathol. 2014, 139, 27-37. [Google Scholar] [CrossRef]
- Vikal, Y.; Bhatia, D. Genetics and genomics of bacterial blight resistance in rice. Advances in international rice research, 2017,175-213.
- Moumouni, B. Le flétrissement bactérien du riz au Niger : Diversité pathologique d’isolats collectés sur les périmètres irrigués. J. appl. biosci, 2007, 38, 2551-2563. [Google Scholar]
- Priya, L.B.; Ujjal, K.N.; Sharmistha, G.; Gayatri, G.; Shalim, U.; Omar, M.A.; Arafat, A.H.; Alison, M.L.; Yong-Ming, G.; Akbar, H. Introgression of bacterial blight resistance genes in the rice cultivar ciherang: Response against Xanthomonas oryzae pv. oryzae in the F6 generation. Plants 2021, 10, 2048. [Google Scholar]
- Djedatin, G.; Nanoukon, C.; Missihoun, A.; Lomou, M.; Sedah, P.; Agbangla, C. Molecular Identification of Xa4 Resistance Gene to Xanthomonas oryzae pv. Oryzae in Cultivated Rice in Northwest Benin. Asian J. Agric. Res 2022, 15(4), 11–22. [Google Scholar] [CrossRef]
- Lang, J M.; Hamilton, J. P.; Diaz, M. G.Q.; Van Sluys, M.A.M.; Burgos, R.G.; Vera Cruz, C.M.; Buell, Tisserat, C.R., Leach, N.A. “Genomics-Based Diagnostic Marker Development for Xanthomonas oryzae pv. oryzae and X. oryzae pv. oryzicola.” Plant Dis, 2010, 94 (3): 311–19.
- Kini, K.; Agnimonhan, R.; Dossa, R.; Soglonou, B.; Gbogbo, V.; Ouedraogo, I.; Kpemoua, K.; Traoré, M.; Silue, D. First report of Sphingomonas sp. causing bacterial leaf blight of rice in Benin, Burkina Faso, The Gambia, Ivory Coast, Mali, Nigeria, Tanzania and Togo. New Dis. Rep. 2027, 35, 32. [Google Scholar] [CrossRef]
- Fanou, A. A.; Missihoun,A.A.; Sovegnon,P.; Behoundja-kotoko,O.; Baimey, H.; Agbangla C. Molecular genetic identification of viruses affecting pepper crop (Capsicum spp.) in Western North of Benin. Int. J. Curr. Res. Biosci. Plant Biol. 2019, 6(3), 9-14. [CrossRef]
- Chen, X.; Temnykh, S.; Xu, Y.; Cho, Y.; McCouch, S. Development of a microsatellite framework map providing genome-wide coverage in rice (Oryza sativa L.). Theor. Appl. Genet, 2017, 95, 553-567. [Google Scholar] [CrossRef]
- Porter, B.; Chittoor, J.; Yano, M.; Sasaki, T.; White, F. Development and mapping of markers linked to the rice bacterial blight resistance gene Xa7. Crop Sci, 2003, 43(4), 1484-1492. [CrossRef]
- Hajira, S. K.; Sundaram, R. M.; Laha, G. S.; Yugander, A.; Balachandran, S. M.; Viraktamath, B. C.; Sujatha, K.; Balachiranjeevi, C. H.; Pranathi, K.; Anila, M. A single-tube, functional marker-based multiplex PCR assay for simultaneous detection of major bacterial blight resistance genes Xa21, xa13 and xa5 in rice. Rice Sci, 2016, 23(3), 144-151. [CrossRef]
- Yan-Chang, L.; Shou-hai, W.; Cheng-quan, L.; Shuang, W.; De-zheng, W.; Shi-yun, D. Improvement of resistance to bacterial blight by marker-assisted selection in a wide compatibility restorer line of hybrid rice. Rice Sci 2004, 11(5-6), 231. [Google Scholar]
- Ullah, I.; Jamil, S.; Iqbal, M.; Shaheen, H.; Hasni, S.; Jabeen, S.; Mehmood, A.; Akhter, M. Detection of bacterial blight resistance genes in basmati rice landraces. Genet Mol Res 2012, 11(3), 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Sabar, M.; Bibi, T.; Farooq, H. U.; Haider, Z.; Naseem, I.; Mahmood, A.; Akhter, M. Molecular screening of rice (Oryza sativa L.) germplasm for Xa4, xa5 and Xa21 bacterial leaf blight (BLB) resistant genes using linked marker approach. Afr. J. Biotechnol 2016, 15(41), 2317–2324. [Google Scholar]
- Kamhun, W.; Pheng-am, S.; Uppananchai, T.; Ratanasut, K.; Rungrat, T. Effects of nitrogen levels on sucrose content, disease severity of Xanthomonas oryzae pv. Oryzae and yield of hybrid rice (BC4F5). Agric. Nat. Resour. 2022, 56(5), 909–916. [Google Scholar]
- Dossa, G. S. , Oliva, R., Maiss, E., Vera Cruz, C., & Wydra, K. High temperature enhances the resistance of cultivated African rice, Oryza glaberrima, to bacterial blight. Plant Dis. 2016, 100(2), 380–387. [Google Scholar] [CrossRef]
- Ullah, I. , Jamil, S., Iqbal, M., Shaheen, H., Hasni, S., Jabeen, S., Mehmood, A., & Akhter, M. Detection of bacterial blight resistance genes in basmati rice landraces. Genet Mol Res. 2012, 11(3), 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.P.; Jeung, J.U.; Noh, T.H.; Cho, Y.C.; Park, S.H.; Park, H.S.; Shin, M.S.; Kim, C.K.; Jena, K. K;. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice, 2013, 6, 1-11. [Google Scholar] [CrossRef] [PubMed]
- Ronald, P. C.; Albano, B.; Tabien, R.; Abenes, L.; Wu, K.; McCouch, S.; Tanksley, S. D. Genetic and physical analysis of the rice bacterial blight disease resistance locus, Xa21. Mol. Genet. Genom. 1992, 236, 113-120. [Google Scholar] [CrossRef]
- Gonzalez, C.; Szurek, B.; Manceau, C.; Mathieu, T.; Séré, Y.; Verdier, V. Molecular and pathotypic characterization of new Xanthomonas oryzae strains from West Africa. Mol. Plant Microbe Interact. 2007, 20(5), 534–546. [Google Scholar] [CrossRef]
- Ramalingam, J.; Raveendra, C.; Savitha, P.; Vidya, V.; Chaithra, T. L.; Velprabakaran, S.; Saraswathi, R.; Ramanathan, A.; Arumugam Pillai, M. P.; Arumugachamy, S. Gene pyramiding for achieving enhanced resistance to bacterial blight, blast, and sheath blight diseases in rice. Front. Plant Sci., 2020, 11, 591457. [Google Scholar] [CrossRef]
- Krishnakumar, R.; Kumaravadivel, N. Marker-assisted selection for biotic stress (Bacterial leaf blight and gall midge) tolerance in Bc4F4 generation of rice (Oryza sativa L.). Electron. J. Plant Breed. 2018, 9(1), 275–282. [Google Scholar] [CrossRef]
Figure 1.
Map showing the collection area for rice leaves showing symptoms of BLB and identification areas of Xanthomonas oryzae and Sphingomonas Sp in the three zones.
Figure 1.
Map showing the collection area for rice leaves showing symptoms of BLB and identification areas of Xanthomonas oryzae and Sphingomonas Sp in the three zones.

Figure 2.
gel photos showing bands specific to Xo and Sphingomonas sp.A: bands of 312pb of Xo Hypothetical protein. B: Bands of 272pb different from those of Xo. C: Bands of 435pb specific to sphingomonas.
Figure 2.
gel photos showing bands specific to Xo and Sphingomonas sp.A: bands of 312pb of Xo Hypothetical protein. B: Bands of 272pb different from those of Xo. C: Bands of 435pb specific to sphingomonas.
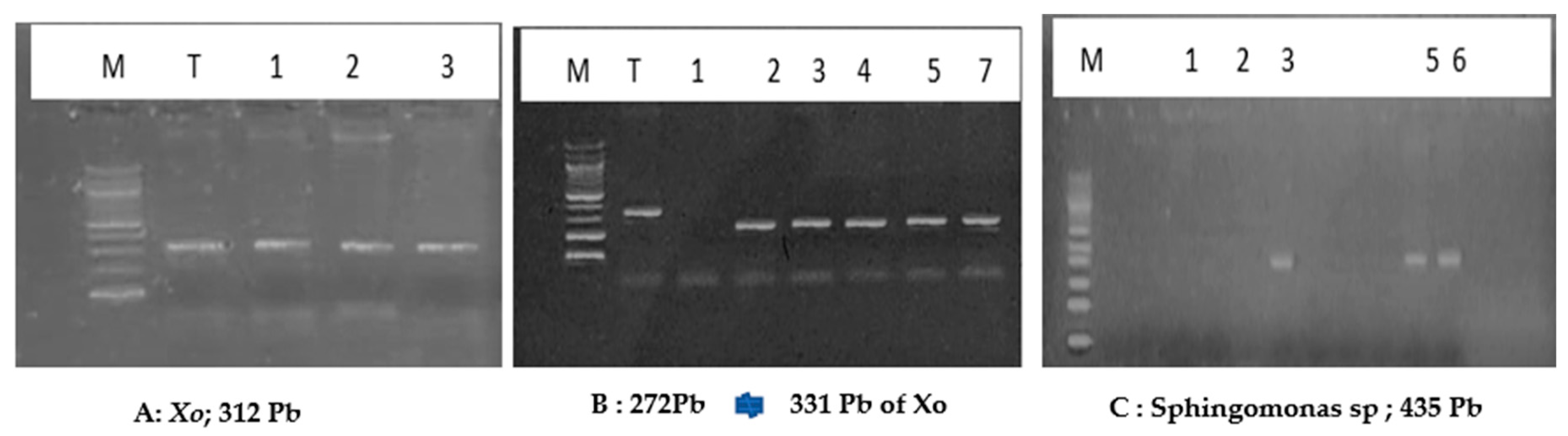
Figure 3.
Representative gel pictures showing amplification patterns generated by different STS markers used in the study. A: MR122 Marker (xa5 gene linked); B: M5 marker (Xa7 gene linked); C: pTA248 marker (Xa21gene linked); D: xa13-prom marker (xa13 gene linked).
Figure 3.
Representative gel pictures showing amplification patterns generated by different STS markers used in the study. A: MR122 Marker (xa5 gene linked); B: M5 marker (Xa7 gene linked); C: pTA248 marker (Xa21gene linked); D: xa13-prom marker (xa13 gene linked).
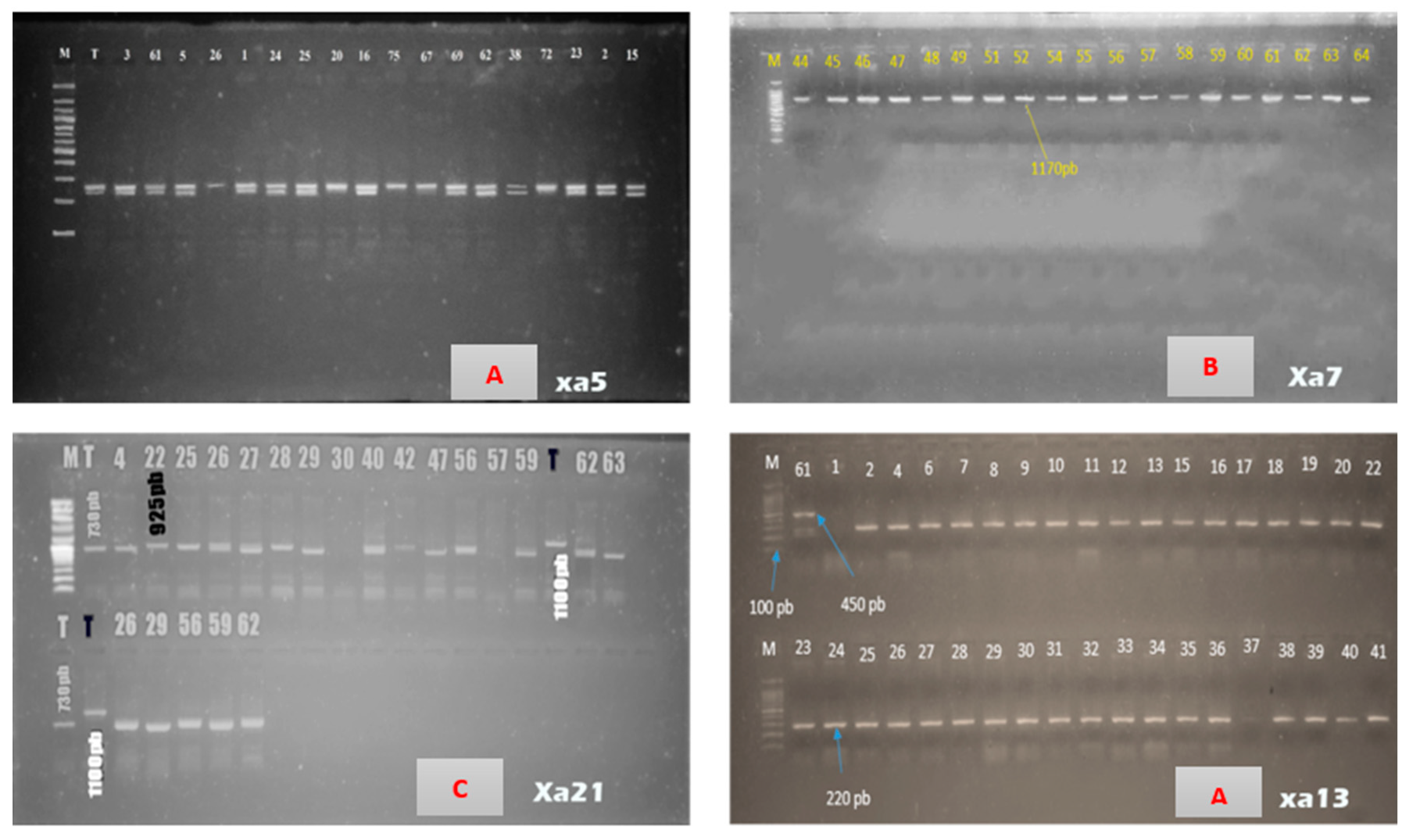
Figure 4.
Representative gel pictures showing amplification patterns generated by pTA248 marker (Xa21 gene linked) at 55°C and 57°C. A: Amplification at 55°C.B: Amplification at 57°C.
Figure 4.
Representative gel pictures showing amplification patterns generated by pTA248 marker (Xa21 gene linked) at 55°C and 57°C. A: Amplification at 55°C.B: Amplification at 57°C.
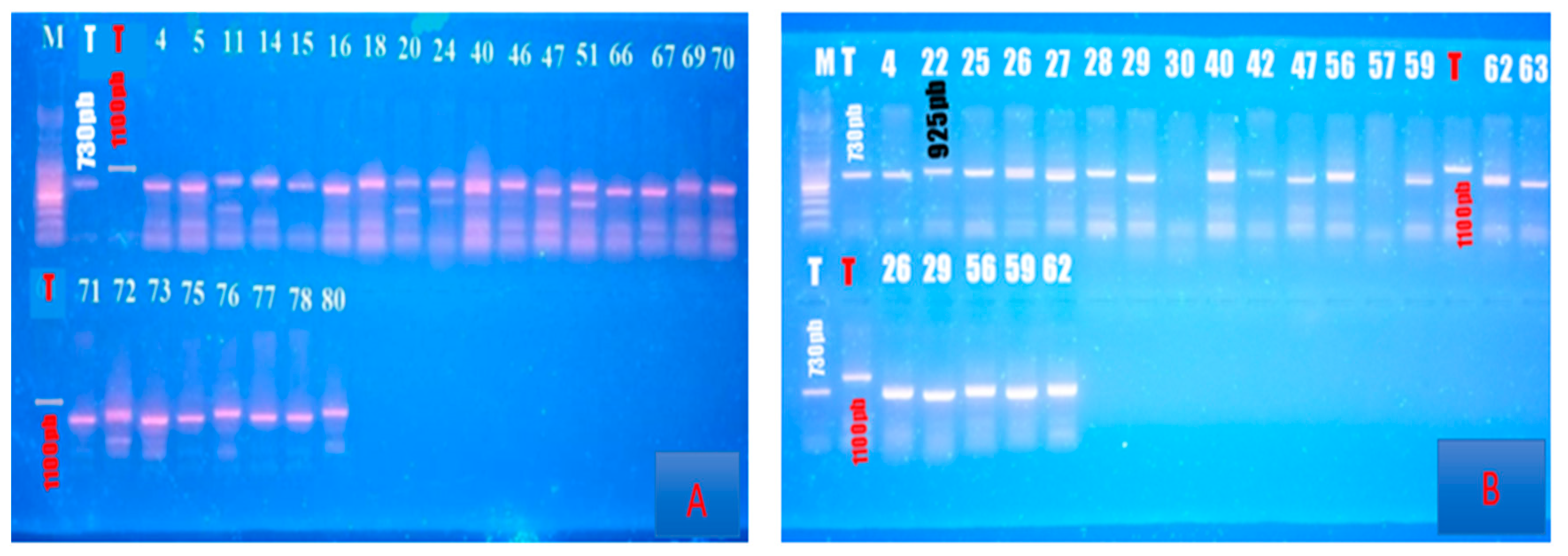
Table 2.
Sequences of primers used for identification of Xanthomonas oryzae, Xanthomonas oryzae pv oryzae and Sphingomonas sp.
Table 2.
Sequences of primers used for identification of Xanthomonas oryzae, Xanthomonas oryzae pv oryzae and Sphingomonas sp.
| Target genes | Primers sequence (5’-3’) | Size (bp) | References | |
|---|---|---|---|---|
| Hypothetical protein (X. orysae) | F | CATCGTTAGGACTGCCAGAAG | 331 | [23] |
| R | GTGAGAACCACCGCCATCT | |||
| Hypothetical protein (X. oryzae pv. Oryzae) | F | GCCGCTAGGAATGAGCAAT | 162 | [24] |
| R | GCGTCCTCGTCTAAGCGATA | |||
| Sphingomonas sp | F | CGGCTGCTAATACCGGATGAT | 435 | [24] |
| R | AGGCAGTTCTGGAGTTGAGC | |||
Table 3.
Characteristics of microsatellite markers used to search for resistance genes.
| Genes | chromosome | Marqueur types / Nom | Primers sequence (5’-3’) | Resistance allele (pb) |
Susceptibility allele (pb) | References | |
|---|---|---|---|---|---|---|---|
| xa5 | 5 |
STS/ RM 122 |
F | GAGTCGATGTAATGTCATCAGTGC | 240pb | 230pb | [26] |
| R | GAAGGAGGTATCGCTTTGTTGGAC | ||||||
| Xa7 | 6 | STS/M5 | F | CTGGATACGGAACCTTCTAAC | 294pb | 1170pb | [27] |
| xa13 | 8 | STS/xa13-prom | R | AGAGAACCTTCTCCTTCAGTG | |||
| F | GGCCATGGCTCAGTGTTTAT | 450pb | 220pb | [28] | |||
| Xa21 | 11 | STS/ pTA248 | R | GAGTCCAGCTCTCCAAATG | |||
| F | AGACGCGAAGGGTGGTTCCCGA | 925pb | 730pb | [29] | |||
| R | AGACGCGGTAATCGAAGATGAAA | ||||||
Table 4.
Types, genotypes and phenotypes of the rice varieties analysed.
| xA5 | Xa21 | Xa7 | xa13 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Department | Samples | Resistanceallele (240pb) | Susceptibility allele (230pb) |
Genotypes | Resistance allele(1100pb) |
Resistance allele (925pb) |
Susceptibility allele (730pb) |
Genotypes | Resistance Allele (294pb) |
Susceptibility Allele (1170pb) |
Genotypes | Resistance allele (450pb) |
Susceptibility allele (220pb) |
Genotypes |
| Alibori | Bagou 18 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Gou 10 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 25 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 19 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Gou 11 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 17 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 22 | + | - | xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 26 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 24 | + | - | xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 27 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Ang 1 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 20 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 21 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Ang 6 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 23 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bagou 28 | - | - | Ø | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Ang 2 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koum 53 | - | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koud 43 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Gou 12 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Ang 16 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Foun 15 | + | - | xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Borgou | Gami 74 | + | - | xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Bori 84 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Gami 77 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Gami 76 | + | + | Xa5/Xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Tot 82 | + | - | xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Bori 83 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Atacora | Koum 51 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Tchaka 38 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kotch 70 | + | - | xa5/xa5 | - | - | + | xa21/xa21 | + | xa7/xa7 | - | + | Xa13/Xa13 | ||
| Tchaka 41 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kan 58 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Nana 32 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Tchaka 33 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kan 61 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koud 45 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koum 54 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kan 60 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kan 59 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kotch 71 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koum 47 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koud 46 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | ||
| Tchaka 36 | + | + | Xa5/xa5 | - | - | - | ϕ | - | + | xa7/xa7 | - | + | ||
| Koung 69 | - | + | Xa5/Xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | ||
| Kotch 72 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koung 67 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koum 50 | - | - | Ø | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Nana 30 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Nana 29 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koud 44 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kotch 73 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koum 55 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koum 49 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koung 65 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| NERICA 19 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Tchaka 39 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Koud 42 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Donga | ONK 93 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Tchal 89 | - | + | Xa5/Xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| ONK 93b | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Kik 96 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Colline | Okouta 98 | + | + | Xa5/xa5 | - | + | + | Xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Kpatab 100 | + | - | xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Okouta 97 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Agbab 101 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Mono | Man 118 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Couffo | Dev 116 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| Atlantique | Doko 122 | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 |
| 6R2B9 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| IR841 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | ||||||||
| IR64 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | ||||||||
| Moroberekan | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Nippon bar | + | + | Xa5/xa5 | - | + | - | Xa21/Xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
| Tog 5307 | + | + | Xa5/xa5 | - | - | + | xa21/xa21 | - | + | xa7/xa7 | - | + | Xa13/Xa13 | |
|
Positif controls |
IRBB5 | + | - | xa5/xa5 | ||||||||||
| IRBB60 | + | + | Xa5/xa5 | + | - | - | Xa21/Xa21 | + | + | xa13/Xa13 | ||||
| IRBB21 | - | - | + | xa21/xa21 | ||||||||||
Legend: +: Presence; -: Absence; ∅: Absence of gene.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



