
Submitted:
17 September 2023
Posted:
19 September 2023
You are already at the latest version
Abstract
The functioning of forest ecosystems depends on the taxonomic and ecological diversity of soil fungi. Urbanization is increasing worldwide and is regarded a major driver of environmental change altering local species assemblages in urban forest. We investigated whether the degree of urbanization and local forest characteristics affect the soil fungal community in 20 beech forests located along a rural–urban gradient in the city of Basel and its suburbs (Switzerland). We analyzed their soil fungal communities by DNA metabarcoding of the rDNA ITS2 region and related these data to local forest vegetation characteristics and soil properties. The number of fungal OTUs in the 20 forests examined ranged from 170 to 303. Richness, diversity and evenness of fungal OTUs were all significantly affected by the degree of urbanization, but in different ways. OTU richness was highest in forests in areas with a low degree of urbanization and lowest in forests in rural areas. In contrast, the diversity of OTUs increased with increasing degree of urbanization. Different fungal phyla and fungal guilds showed distinct patterns in their relative abundance along the rural-urban gradient. The degree of urbanization reduced the relative abundance of symbiotrophic fungi, but increased that of saprotrophic and pathotrophic fungi. Our results show that urbanization alters soil fungal community, which in turn can lead to changes in forest ecosystems.
Keywords:
urbanization
; soil microbial community
; beech forest
; forest characteristics
; trophic groups
; fungal guilds
1. Introduction
Urbanization is increasing worldwide and is considered a key driver of environmental change [1]. Forests represent one of the most frequent types of green space in urban landscapes [2]. Forests provide a wide range of ecosystem functions, from nutrient recycling and storage, air filtering, temperature regulation, recreation for residents to habitat for native plants and animals [3,4]. However, urban forests are influenced by the environmental conditions of the built-up areas in the surroundings. Several studies have reported changes in abiotic conditions in forest areas, such as an increase in temperature and nitrogen deposition and a decrease in precipitation along a rural–urban gradient [5]. These changes affect habitat quality and thus species richness, species composition and the functional diversity of animals and plants in urban forests [6,7,8], which in turn influence the functioning of ecosystems [9,10].
Soil microbial organisms, especially soil fungi, are a key component of forest ecosystems [11]. In particular, the high taxonomical and ecological diversity as well as the high biomass of soil fungi are important for the proper functioning of forest ecosystems [12]. Among other things, fungi mediate important ecosystem processes such as the carbon and nutrient cycling [13,14]. In forests, three major functional guilds of soil fungi can be distinguished based on their ecological lifestyle: saprotrophic, symbiotrophic and plant pathotrophic fungi [15]. Saprotrophic fungi are decomposers that gain carbon and nutrients by breaking down dead organic matter. Symbiotrophic fungi form close mutualistic associations with plant roots, facilitating nutrient uptake by their host plant in exchange for photosynthetic carbon products. Mycorrhizal fungi, an essential part of the symbiotrophic fungi, have an important function for the establishment and growth of many plants and thus for the natural regeneration and structure of forests [16]. Plant pathotrophic fungi comprise a heterogeneous group of soil fungi that infect roots of various plant and tree species and are mostly non-host specific necrotrophs [17]. Thus, plant pathotrophic fungi have the potential to change the composition of plant and tree species [18]. As a consequence, the altered forest structure can lead to shifts in the composition of soil fungi and thus influence nutrient cycling [19].
Most studies examining the effects of urbanization on the soil microbial community have focused on soil bacteria, while less attention has been directed to soil fungi. The few studies on soil fungal communities in urban habitats have shown contrasting results. Urbanization reduced the abundance and diversity of total soil fungi in some studies [20,21]. However, other studies reported an increase in fungal species richness from rural areas to the inner city [22,23]. Still other studies found only a weak or no effect of urbanization on soil fungal diversity [24,25].
In our study, we used a standardized sampling procedure to investigate whether the degree of urbanization, local forest characteristics or soil properties influence the soil fungal community. We selected 20 beech forests along a rural–urban gradient in the city of Basel and its suburbs (Switzerland). We recorded the abundance and species composition of plants, shrubs and trees in each forest and collected soil samples for analysis of soil fungi and assessment of various physio-chemical soil properties. The soil fungal community was analyzed by DNA metabarcoding of the rDNA ITS2 region [26].
We tested the following hypotheses:
- (1)
- Spores are the main dispersal mode of fungi [27]. Forest patches in urban environments receive reduced numbers of migrating fungal spores, which could result in decreased fungal diversity and altered composition of the soil fungal community [20]. We therefore hypothesize that the species richness of fungi in the soil decreases with increasing degree of urbanization and that increasing urbanization leads to changes in the composition of soil fungi.
- (2)
- The different fungal phyla differ in their susceptibility to changes in biotic and abiotic characteristics of temperate forests [28,29]. Urbanization can change vegetation characteristics and soil properties [30]. We therefore hypothesize that urbanization-induced changes in forest characteristics will affect the different fungal phyla in different ways.
- (3)
- Symbiotrophic fungi are sensitive to disturbances [31]. This may result in a lower abundance and/or species richness of symbiotrophic soil fungi in urban than in rural habitats [32,33]. Based on these findings, we hypothesize that the abundance of symbiotrophic fungi decreases with increasing degree of urbanization.
2. Materials and Methods
2.1. Study Area
The study was carried out in and around the city of Basel in northwestern Switzerland (47°34’N, 7°36’E, elevation: 245–565 m a.s.l.). The study area covers 88.3 km2 and consists of 43.6 km2 (49.4%) residential area, 16.1 km2 (18.3%) agricultural land, 25.5 km2 (28.8%) forest, 2.2 km2 (2.5%) water bodies, and 0.9 km2 (1.0%) other areas [34]. Approximately 315,000 people live in Basel and its suburbs with a population density of about 2,000 inhabitants per km2 [35]. In the study area, the total annual precipitation averages 842–1005 mm and annual mean temperature 10.2–10.9 ˚C (average of the records from 1981 to 2010 [36]).
2.2. Design of the Field Survey
To assess the potential impact of urbanization on the soil fungal community in urban forests, we selected 20 deciduous forests belonging to the Fagetum association [37] along an urbanization gradient (Figure 1). The forests studied ranged in size from 0.23 ha to 337.0 ha and differed in their historical development (Table 1). Fifteen of them are surrounded by settlements and agricultural land and most of them are no longer connected to large continuous forests. Six of these forests are remnants (fragments) of former large continuous forests and six were planted after 1884, while three forests are part of large continuous forests (> 40 ha; Table 1). The remaining five forests are situated in the rural surroundings and are part of large, continuous forests (> 76.2 ha; Table 1). The most abundant tree species in these forests are European beech (Fagus sylvatica), sycamore (Acer pseudoplatanus) and European oak (Quercus robur). The ground vegetation in the forests shows a high richness of vernal geophytes, including Anemone nemorosa, Ranunculus ficaria, Polygonatum multiflorum and Arum maculatum [37]. Most forests were state owned and accessible to the public. Some forests are privately owned but managed by the forestry authorities.
In each forest, we chose an area dominated by European beech (80–90% of all tree individuals) and set up a study plot measuring 10 m x 10 m in its centre. The study plots had a minimum distance of 5 m to the nearest forest edge or to permanent paths to minimize possible edge effects. The forest management in the study plots (time since last thinning and management intensity) was similar among the forests investigated.
As a measure of degree of urbanization, we determined the percentage of sealed areas within a radius of 500 m from the centre of each study plot using satellite images from Google Earth [38] and the pixel count function of Adobe Photoshop (version 10.0.1). Using the same method, we assessed the percentage of forest cover within a radius of 500 m from the centre of each study plot. The percentage of sealed areas around the study plots ranged from 1 to 69% in the forest examined, and the percentage of forest cover was between 1 and 92% (Table 1).
2.3. Forest Vegetation Survey
In order to investigate the species richness and species composition of the forest vegetation, we set up four sampling plots measuring 4 m x 4 m in each corner of the study plots. We determined the plant species richness and the abundance of individual species in the ground vegetation (≤ 40 cm) in a randomly chosen 2 m x 2 m subplot in each of the four sampling plots per forest. Plant species cover was estimated using the Domin scale [39]. To complete the list of plant species in a sampling plot, we recorded any additional species found in the other three subplots. In addition, the number of shrub species (0.4–3 m high) and tree species (diameter at breast height, DBH > 10 cm) was recorded in each study plot. The vegetation surveys were carried out in autumn 2020 and spring 2021.
2.4. Soil Sampling and Physiochemical Properties
In spring 2021, we collected five soil samples (one in each corner and one in the centre) in each sampling plot. We removed the litter layer and used a metal cylinder (diameter: 5.05 cm; soil volume: 100 cm3) to sample the soil to a depth of 5 cm. The five soil samples from a sampling plot were pooled and transported on ice to the laboratory where they were sieved (mesh size: 2 mm). This resulted in a total of 80 soil samples (4 sampling plots x 20 forests). One part of each soil sample was stored at –80 oC until DNA extraction, while the remaining part was stored at –20 oC for determination of physiochemical soil properties.
We determined soil moisture content (%) using the fresh to dry weight ratio and assessed soil pH in distilled water (1:2.5 soil:water) [40]. We assessed the total soil organic matter content (SOM, %) as loss-on-ignition of oven dried soil at 750 °C for 16 h [40]. Total soil nitrogen content (%) was determined using the standard method of Kjeldahl [41]. We also determined the plant-available phosphorus content of soil (μg PO43–/g) using the molybdenum blue method according to Sparks et al. [42].
2.5. Soil Fungal Community
We extracted total soil genomic DNA from 0.5–0.6 g soil in triplicate using NucleoSpin Soil kit (Macherey-Nagel, Oensingen, Switzerland) according to the manufacturer’s instructions. The triplicate DNA extracts were combined into one sample and purified using NucleoSpin gDNA Clean-up kit (Macherey-Nagel, Oensingen, Switzerland). We quantified the concentration and purity of DNA using NanoDrop (NanoDrop Technologies Inc., Wilmington, USA), adjusted the sample to 5 ng/µl and stored it at –20 oC.
Fungal communities were analyzed by amplification and sequencing of the internal transcribed spacer 2 region (ITS2) using the primer pair ITS3/ITS4 [43] added with the Illumina adapters. PCR reactions (25 μl) consisted of 5 μl template DNA, 12.5 μl Master Mix (HotStar Taq Master Mix kit; Qiagen, Switzerland), 2.5 μl Primer ITS3 (10 μM), 2.5 μl Primer ITS4 (10 μM) and 2.5 µl sterile water. The amplification was achieved in an Eppendorf Mastercycler Pro (Vaudaux-Eppendorf AG, Schönenbuch, Switzerland) under the following conditions: initial 15 min heat activation step at 94 °C, followed by 35 amplification cycles of denaturation at 94 °C for 40 s, annealing at 55 °C for 40 s and extension at 72 °C for 60 s, with a final extension step at 72 °C for 10 min. PCR reactions were done in triplicate.
PCR products were sent to Microsynth AG (Belgach, Switzerland) for sequencing and analysis. Briefly, PCR products were purified, quantified, and pooled at equimolar concentrations for sequencing on an Illumina Miseq using the 2 x 250 bp paired-end approach. Paired fungal sequences from Row Illumina Miseq were demultiplexed and merged using the USEARCH pipeline [44]. The merged sequences were then quality-filtered and clustered into operational taxonomical units (OTUs) using the USEARCH pipeline and UPARSE [45]. Singletons were removed prior to OTU determination at 97% sequence identity and chimeric representative sequences were removed with UCHIME [46]. Finally, the original sequences were mapped to OTUs at a 97% identity threshold to obtain an OTU table. The taxonomy of each sequence was analyzed by Ribosomal Database Project Classifier [47] against UNITE fungal database [48] and the NCBI/GenBank [49].
2.6. Data Analyses
All statistical analyses were performed in R [50]. To avoid spatial pseudo-replication, we analyzed the data at the forest site level (n = 20) by using the combined data collected in the four sampling plots in each forest.
We used generalized linear mixed models (GLM) with Poisson-distributed errors to analyze the effects of degree of urbanization and forest size (log-transformed) on the number of plant, shrub and tree species. We applied analyses of covariance (ANCOVA) to assess the effects of degree of urbanization and forest size (log-transformed) as co-factor on ground vegetation cover (arcsine square root-transformed) and on soil moisture (%; arcsine square root-transformed), soil pH, SOM (%; Tukey-transformed), total soil nitrogen (%; Tukey-transformed) and plant-available phosphorus (square root-transformed).
To avoid bias due to differences in sequencing numbers between the soil samples, we rarefied the number of sequences of each sample to the lowest value for normalization using the procedure rarefy in the vegan package [51]. All further analyses were conducted with rarefied OTU data. Alpha-diversity, Shannon-diversity and Pielou’s evenness were calculated using the vegan package [51]. Preliminary analyses revealed inter-correlations between several chemical and physical soil properties (soil pH vs. SOM: rs = 0.72, n = 20, p < 0.001; soil pH vs. total soil nitrogen content: rs = 0.51, n = 20, p = 0.022, and soil pH vs. plant-available phosphorus: rs = 0.66, n = 20, p = 0.002). We therefore considered soil pH, soil moisture and the recorded forest vegetation characteristics as co-factors in the subsequent statistical analyses.
We applied generalized linear mixed models (GLM) with Poisson-distributed errors to analyze the effects of degree of urbanization, forest size, percentage of forest cover within a radius of 500 m from the centre of each study plot, vegetation characteristics, and soil properties on fungal OTU richness. Degree of urbanization was included as a fixed factor, forest size (log-transformed), percentage of forest cover within a radius of 500 m from the centre of each study plot (log-transformed), ground vegetation cover (Tukey-transformed) and plant, shrub and tree species richness (all log-transformed), as well as soil moisture and soil pH as cofactors in the GLM models. We used the same GLM models, but with gamma-distributed errors, to assess the effects of degree of urbanization, forest size, percentage of forest cover within a radius of 500 m from the centre of each study plot, vegetation characteristics, and soil properties on both the Shannon diversity and evenness of fungal OTUs.
To visualize differences in OTU composition, we plotted non-metric multidimensional scaling coordinates (NMDS) for first two dimensions based on Bray-Curtis dissimilarities matrices using metaMDS in the vegan package [51]. Permutational multivariate analysis of variance (PERMANOVA) was used to test whether the degree of urbanization affects the composition of fungal OTUs [52]. The same co-factors as in the GLM models were included in the model. All PERMANOVA tests were based on 9999 permutations of the untransformed raw data, using the adonis function in the vegan package [51]. Finally, individual OTU affinity with a given degree of urbanization was determined using indicator species analysis by the multipatt function in the indicspecies R package [53], which tests the significance of the indicator species index through a permutations test with 9999 permutations.
We applied the same GLM models with gamma-distributed errors to assess the impact of degree of urbanization on the relative abundance of Basiodomycota, Chytridiomycota, Ascomycota and Morteriellomycota (all Tukey-transformed). PERMANOVA analyses, as described above, were conducted to test whether the degree of urbanization affects the composition and the relative abundance of fungal phyla. In addition, we used analysis of similarity (ANOSIM) in the vegan package [51] to examine differences in the composition of fungal phyla between the different degrees of urbanization. ANOSIM is a nonparametric permutation procedure that allows comparison of between-group and within-group dissimilarities [54]. The procedure calculates R statistics ranging from −1 to 1. R = 0 indicates completely random grouping, while R = 1 when all replicates within groups are more similar than all replicates between groups.
We also used GLM models with gamma-distributed errors as described above to analyze the impact of degree of urbanization on the relative abundance of saphotrophic (sqrt-transformed), symbiotrophic (Tukey-transformed) and pathotrophic (sqrt-transformed) fungi.
3. Results
3.1. Vegetation Characteristics and Soil Properties
The degree of urbanization affected the species richness of plants and shrubs in different ways (Table 2). The plant species richness of ground vegetation decreased with increasing degree of urbanization and was higher in forests in both rural and low urbanization areas than in forests in moderate and high urbanization areas (Table 2). Shrubs showed the opposite pattern, species richness increased with increasing degree of urbanization (Table 2). Soil pH was higher in forests in moderately and highly urbanized areas than in forests in low urbanized and rural areas (Table 2). Plant-available phosphorus tended to be higher in forests in moderately and highly urbanized areas than in low urbanized and rural areas (Table 1). Interestingly, forest size had no influence on the vegetation characteristics and soil properties examined (all p > 0.15).
3.2. Diversity and Composition of Fungal OTUs
After quality filtering, a total of 1,704,819 sequences were recovered from the soil samples, with an average of 21,100 sequences per sample (range 3813–44,593). To correct for differences in the number of reads, all samples were subsampled to the lowest number of reads, yielding a total of 680 OTUs with 97% sequence identity (Table S1). The number of fungal OTUs in the 20 forests examined ranged from 170 to 303 (mean ± s.e: 239.6 ± 8.5).
Richness, diversity and evenness of fungal OTUs were all significantly affected by the degree of urbanization (Figure 2; Table 3). OTU richness was highest in forests in areas with a low degree of urbanization and lowest in forests in rural areas (Figure 2a). In contrast, the diversity and evenness of OTUs increased with increasing degree of urbanization (Figure 2b,c). The percentage of area covered with forests in the surroundings of the study plots significantly influenced OTU richness (Table 3). In addition, soil moisture affected both the richness and evenness of fungal OTUs (Table 3).
The PERMANOVA analysis showed that the degree of urbanization caused shifts in the composition of fungal OTUs (F3,9 = 1.92, p = 0.004; Figure 3, Table S2). This was because the fungal OTU composition of forests in areas with a high and moderate degree of urbanization differed from the composition in forests in rural areas (ANOSIM: R = 0.274, p = 0.009; Figure 3). PERMANOVA analyses also revealed that the extent of ground vegetation cover influenced OTU composition (F1,9 = 1.89, p = 0.020; Table S2). In addition, the indicator analysis showed that out of a total of 680 OTUs, 46 OTUs were significantly associated with a particular degree of urbanization (Indval > 0.7; p < 0.05; Table S3). Fiveteen OTUs were identified as indictors for forests in rural areas, one OTU for forests in low urbanization areas, two OTUs for forests in moderate urbanization areas and 28 OTUs for forests in high urbanization areas (Table S3).
3.3. Soil Fungal Community
Of the total number of 680 fungal OTUs recorded, 670 OTUs could be assigned to 11 phyla (98.5%) (Table S1). Likewise, 648 fungal OTUs could be assigned to 31 classes (95.3%), 630 to 58 orders (92.6%), 554 to 111 families (81.4%) and 528 to 157 genera (77.6%). Ten OTUs could only be classified at the fungi kingdom level (Table S1). Basidiomycota was the predominant phylum with 48.1% of the OTUs, followed by Ascomycota (32.4%), Morteriellomycota (10.1%) and Chytridiomycota (5.0%). The corresponding figures for the relative abundances were 44.4%, 28.5%, 22.6% and 2.6%. The other phyla included Rozellomycota, Mucoromycota, Olpidiomycota, Cryptomycota, and Glomeromycota, which together accounted for 4.4% of the OTUs and 2.0% of the relative abundance (Table S1).
Different fungal phyla showed distinct patterns in their relative abundance along the rural–urban gradient (Figure 4). The relative abundance of Basidiomycota decreased with increasing degree of urbanization (F3,11 = 5.67, p = 0.009; Figure 4; Table S4). The opposite pattern was found for the relative abundance of Chytridiomycota, which increased with increasing degree of urbanization (F3,14 = 8.63, p < 0. 001; Figure 4; Table S4). Similarly, the relative abundance of Ascomycota tended to increase with increasing degree of urbanization (F3,13 = 2.94, p = 0.08; Figure 4; Table S4). The relative abundance of Morteriellomycota was lowest in forests in rural areas (F3,14 = 4.58, p = 0. 020; Figure 4; Table S4).
The PERMANOVA analysis revealed that the degree of urbanization caused a shift in the composition of fungal phyla (F3,12 = 4.57, p = 0.001; Table S2). Similar to fungal OTUs, the composition of fungal phyla differed between forests in rural areas and forests in areas with moderate or high degrees of urbanization (ANOSIM: R = 0.220, p = 0.043).
3.4. Soil Fungal Functional Composition
The majority of fungal OTUs (76.5%) could be assigned to a specific functional guild (Figure 5 and Figure 6): 255 were saprotrophic fungi (48.9%), accounting for 61.4% of the relative abundance. The corresponding figures for symbiotrophic fungi were 221 (32.4%) and for pathotrophic fungi 44 (6.2%; Table S1). The degree of urbanization influenced the relative abundance of fungal guilds in different ways (Table 4; Figure 5a–c, Figure 6). The relative abundance of symbiotrophic fungi significantly decreased with increasing degree of urbanization (Table 4; Figure 5b), while the relative abundance of saprotrophic fungi tended to increase with increasing degree of urbanization (Table 4; Figure 5a). For pathotrophic fungi (Table 4; Figure 5c), the relative abundance in forests in areas of very low (rural), low and moderate urbanization was similar, but was significantly higher in forests in highly urbanized areas (Figure 5c).
4. Discussion
4.1. Diversity and Composition of Fungal OTUs
We found an average OTU richness of 240 in the 20 forests examined, which is in the range of other studies conducted in different beech forest stands (104 to 1262 OTUs, mean per study site; [55,56,57,58,59,60]). This large variation in soil fungal richness can be explained by the different barcoding regions used for fungal community profiling (e.g. ITS1 versus ITS2) and/or by different soil sampling procedures (e.g. different soil depth and sampling effort).
Our finding that soil fungal richness is negatively affected by the degree of urbanization (Figure 2a) supports the first hypothesis and is in line with the results of some studies reporting negative effects of urbanization on total soil fungal richness [20,21]. However, other studies either showed a positive effect [22,23,61] or found no effect [24,62] of urbanization on soil fungal richness in forests. Furthermore, we showed that both the diversity and evenness of soil fungi were positively related to the degree of urbanization. Similar results have been obtained in other studies assessing the impact of urbanization on soil fungal communities [22,23,32,61].
Urbanization can change forest stand characteristics including plant and tree species richness and soil properties [6,30]. Along with the composition of the surrounding landscape, altered forest characteristics have the potential to influence soil fungal communities [63]. Numerous studies have shown that the richness of tree and plant species has a positive effect on soil fungi richness in grassland and forests (e.g., [64,65]). However, in our study, species richness of soil fungi was not related to tree and plant species richness, ruling out this possibility, although plant species richness decreased along the urbanization gradient, but tree species richness did not (Table 2). Tedersoo et al. [62] reported that soil pH is the most important soil property determining soil fungal species richness and found that this relationship was unimodal, peaking around pH 6.6. Based on this finding, the recorded increase in soil pH along the urbanization gradient should lead to an increase in soil fungal species richness. However, no relationship was found between soil pH and soil fungal richness in our study. The decline in fungal richness along the urbanization gradient in our study could therefore be due to opposing effects of changes in plant species richness and soil pH on soil fungi. Furthermore, our findings that rural forests and forests in highly urbanized areas have a particular OTU composition and specific indicator species were consistent with several studies that urbanization changes the composition of soil fungi [20,21,24]. However, a comparison of our results with the results of other studies is difficult for several reasons. Other studies that quantified the impact of urbanization on soil fungal community either did not describe the particular land use or examined different habitat types. For example, private gardens or parks in urban areas and forests in the rural areas were sampled [20,22,24].
4.2. Soil Fungal Community
The composition of soil fungal community found in our study, with Basiodiomycota as the dominant fungal phyla, followed by Ascomycota and Morteriellomycota, is characteristic for the soil fungal community in beech forests [55,59,66,67]. Consistent with our second hypothesis, we showed that the different fungal phyla were differently affected by the degree of urbanization. The recorded decline in the Basidiomycota abundance along the urbanization gradient is in line with the results of some of the few studies assessing the impact of urbanization on fungal taxonomic identity (e.g. phylum or class). These studies also reported lower abundances of Basiodiomycota in urban areas than in rural areas [20,61]. However, other studies found either a higher abundance of Basidiomycota in urban than in rural habitats [23] or that urbanization had no effect on Basidiomycota abundance [24]. Furthermore, in our study, the abundance of Chytridiomycota increased while that of Morteriellomycota decreased along the rural–urban gradient. Most Chytridiomycota are saprotrophic fungi living on decaying plant material [68]. The higher abundance of Chytridiomycota in urban forest than in rural forests may therefore be due to the higher amount of soil organic matter recorded in urban forest than in rural forests (Table 2; +21%). Furthermore, the decreasing abundance of Morteriellomycota along the urbanization gradient may be caused by changes in both soil properties and vegetation composition. Our finding that the degree of urbanization changed the composition of fungal phyla and classes is consistent with the results of other studies reporting that urbanization alters the composition of fungal phyla or classes [20,61].
4.3. Soil Fungal Functional Composition
In forest soil, fungi represent a key component of soil microbial community, not only in terms of species richness and diversity, but also in terms of their functionalities [69]. In our study, the soil fungal community is dominated by saprotrophic fungi with a relative abundance of 62%, followed by symbiotrophic (32%) and pathotrophic (6%) fungi. A similar soil fungal community composition was also found in different types of temperate beech forests [59,66,70]. Fungal guilds differ in their ecological requirements and can therefore be differently affected by urbanization-related changes in biotic and abiotic environmental factors. In our study, the degree of urbanization reduced the relative abundance of symbiotrophic fungi. The vast majority of the symbiotrophs recorded are ectomycorrhizal fungi (EM fungi; 220 out of a total of 221) (Table S1). Other studies also reported that urbanization negatively affected richness and abundance of EM fungi in different temperate forests [32,61,71], subtropical and tropical forests [68], and in desert vegetation [21]. However, Liu et al. [22] found no urbanization effect on EM fungi and Tan et al. [24] even reported a higher abundance of ectomycorrhizal fungi in urban than in non-urban habitats.
The magnitude of the abundance reduction in EM fungi recorded in our study (40%) was similar to other studies [28–42%; 21,32,61,71]. However, we showed that the abundance of saprotrophic fungi is positively related to the degree of urbanization. This is consistent with the result of Scholier et al. [61]. In contrast, other studies reported higher abundances of saprotrophic fungi in suburban than in urban areas [21,24], or no effect of urbanization on this fungal guild [71]. Overall, ectomycorrhizal and saprotrophic fungi use the same organic matter and therefore compete for a resource, which can result in inhibition of both fungal groups [72]. Saprotrophic fungi have a higher competitive ability for nutrient uptake than EM fungi when carbon resources are not limited [73,74]. Indeed, the higher soil organic matter content (+20%) in forests in highly urbanized areas than in forests in rural areas could be responsible for the opposite changes in the abundance of EM and saprotrophic fungi. Our finding that the abundance of pathogenic fungi increases with increasing urbanization is supported by other studies [24,71]. Competitive interactions can lead to different responses of EM and pathotrophic fungi depending on the degree of urbanization, because EM fungi can act as antibiotic agents to protect plant roots from infection of soil-borne plant pathogens [75,76].
5. Conclusions
Urbanization is progressing worldwide, and in 2050 it is projected that the number of people living in urban areas will increase by 2.5 billion [77]. Forests are the most frequent type of green space in urban landscapes [2], providing residents with a wide range of ecosystem services, from recreational areas, to the recycling and nutrient storage, to air filtering and temperature regulation, as well as habitat for native species [4]. Our study showed that the taxonomic and functional richness and abundance of soil fungi along a rural–urban gradient are influenced by urbanization. This leads to changes in soil fungal community composition, which in turn can affect ecosystem functions such as nutrient cycling or carbon sequestration, all of which are important for maintaining healthy forests. Appropriate management strategies need to be developed and implemented to maintain a high soil microbial community diversity in forests in urban landscapes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Taxonomic status, ecological guilds and abundance data of all OTUs found in 20 forest sites situated in Basel and its surroundings (Switzerland), Table S2: Summary of PERMANOVA analyses examining the effects of degree of urbanization (four classes), forest size, percentage of area covered with forest within 500 m of the centre of the study plots, forest vegetation characteristics, and soil properties on the composition of fungal OTUs and the composition of fungal phyla, Table S3: List of indicator taxa for the different degrees of urbanization (four classes), together with values for positive predictive power (A), sensitivity (B), and the strength of association for each combination (Indval), Table S4: Summary of general linear models (GLM) testing the effects of degree of urbanization (four classes), forest vegetation characteristics and soil properties on the relative abundance of Basidiomycota, Chytridiomycota, Ascomycota and Morteriellomycota.
Author Contributions
Conceptualization, H.-P.R. and B.B.; methodology, H.-P.R.; software, H.-P.R.; validation H.-P.R. and B.B.; formal analysis H.-P.R.; investigation, H.-P.R. and B.B.; resources, B.B.; data curation, H.-P.R. and B.B..; writing—original draft preparation, H.-P.R. and B.B.; writing—review and editing, H.-P.R. and B.B.; visualization, H.-P.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data presented in this study are available in the article and its Supporting Information Files (Tables S1–S4).
Acknowledgments
We thank Thomas Boller and Georg Armbruster for constructive comments on the manuscript. Special thanks go to the foresters in charge of permission to carry out the vegetation survey and soil sampling in the forests.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.G.; Bai, X.M.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef]
- Cvejić, R.; Eler, K.; Pintar, M.; Železnikar, Š.; Haase, D.; Kabisch, N.; Strohbach, M. A typology of urban green spaces, ecosystem services provisioning services and demands. 2015, Report D3.1, European Union, Brüssel.
- Dwyer, J.F.; McPherson, E.G.; Schroeder, H.W. , Rowntree, R. Assessing the benefits and costs of the urban forest. Arboric. J. 1992, 18, 227–234. [Google Scholar] [CrossRef]
- Grammatikopoulou, I.; Vackarova, D. The value of forest ecosystem services: A meta-analysis at the European scale and application to national ecosystem accounting. Ecosyst. Serv. 2021, 48. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Boone, C.G.; Groffman, P.M.; Irwin, E.; Kaushal, S.S.; Marshall, V.; McGrath, B.P.; Nilon, C.H.; et al. Urban ecological systems: Scientific foundations and a decade of progress. J. Environ. Manage. 2011, 92, 331–362. [Google Scholar] [CrossRef] [PubMed]
- Melliger, R.L.; Braschler, B.; Rusterholz, H.P.; Baur, B. Diverse effects of degree of urbanisation and forest size on species richness and functional diversity of plants, and ground surface-active ants and spiders. Plos One 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Fenoglio, M.S.; Rossetti, M.R.; Videla, M. Negative effects of urbanization on terrestrial arthropod communities: A meta-analysis. Glob. Ecol. Biogeogr. 2020, 29, 1412–1429. [Google Scholar] [CrossRef]
- Meyer, S.; Rusterholz, H.P.; Baur, B. Saproxylic insects and fungi in deciduous forests along a rural–urban gradient. Ecol. Evol. 2021, 11, 1634–1652. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Vitousek, P.M. The effects of plant composition and diversity on ecosystem processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef]
- Chapin III, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Decon, J.W. Fungal Biology, 4nd ed.; John Willey & Sons: New York, 2005; p. 384. [Google Scholar]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Treseder, K.K.; Lennon, J.T. Fungal traits that drive ecosystem dynamics on land. Microb. Molec. Biol. Rev. 2015, 79, 243–262. [Google Scholar] [CrossRef]
- Baldrian, P. Forest microbiome: diversity, complexity and dynamics. FEMS Microbiol. Ecol. 2017, 41, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z. W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Kadowaki, K.; Yamamoto, S.; Sato, H.; Tanabe, A.S.; Hidaka, A.; Toju, H. Mycorrhizal fungi mediate the direction and strength of plant-soil feedbacks differently between arbuscular mycorrhizal and ectomycorrhizal communities. Community Biol. 2018, 1. [Google Scholar] [CrossRef]
- Gomdola, D.; Bhunjun, C.S.; Hyde, K.D.; Jeewon, R.; Pem, D.; Jayawardena, R.S. Ten important forest fungal pathogens: a review on their emergence and biology. Mycosphere 2022, 13, 612–671. [Google Scholar] [CrossRef]
- Terhonen, E.; Kovalchuk, A.; Zarsav, A.; Asiegbu, F.O. Biocontrol potential of forest tree endophytes. In Endophytes of Forest Trees: Biology and Applications, 2nd Edition; Pirttilä, M.A., Frank, Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 283–318. [Google Scholar] [CrossRef]
- Dobson, A.; Crawley, W. Pathogens and the structure of plant-communities. TREE 1994, 9, 393–398. [Google Scholar] [CrossRef]
- Abrego, N.; Crosier, B.; Somervuo, P.; Ivanova, N.; Abrahamyan, A.; Abdi, A.; Hamalainen, K.; Junninen, K.; Maunula, M.; Purhonen, J.; et al. Fungal communities decline with urbanization – more in air than in soil. ISME J. 2020, 14, 2806–2815. [Google Scholar] [CrossRef]
- Chen, Y.J.; Martinez, A.; Cleavenger, S.; Rudolph, J.; Barberan, A. Changes in soil microbial communities across an urbanization gradient: a local-scale temporal study in the arid Southwestern USA. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Liu, L.; Barberan, A.; Gao, C.; Zhang, Z.C.; Wang, M.; Wurzburger, N.; Wang, X.; Zhang, R.; Li, J.X.; Zhang, J. Impact of urbanization on soil microbial diversity and composition in the megacity of Shanghai. Land Degrad. Dev. 2022, 33, 282–293. [Google Scholar] [CrossRef]
- Whitehead, J.; Roy, J.; Hempel, S.; Rillig, M.C. Soil microbial communities shift along an urban gradient in Berlin, Germany. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Tan, X.L.; Kan, L.; Su, Z.Y.; Liu, X.D.; Zhang, L. The Composition and diversity of soil bacterial and fungal communities along an urban-to-rural gradient in South China. Forests 2019, 10. [Google Scholar] [CrossRef]
- Wang, M.; Yu, S.Q.; Chen, X.H.; Liu, X.D.; Zeng, H.X.; Wu, W.K.; Liu, M.Y.; Su, C.H.; Xu, G.L. Soil microbial community changes in response to the environmental gradients of urbanization in Guangzhou City. Urban Ecosyst. 2022, 25, 1865–1874. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjoller, R.; Koljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers – a user's guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, V.B.; Aguilar-Trigueros, C.A.; Mansour, I.; Rillig, M.C. Fungal dispersal across spatial scales. Annu. Rev. Ecol. Evol. Syst. 2022, 53, 69–85. [Google Scholar] [CrossRef]
- Osburn, E.D.; McBride, S.G.; Aylward, F.O.; Badgley, B.D.; Strahm, B.D.; Knoepp, J.D.; Barrett, J.E. Soil bacterial and fungal communities exhibit distinct long-term responses to disturbance in temperate forests. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Odriozola, I.; Martinovic, T.; Bahnmann, B.D.; Rysanek, D.; Masinova, T.; Sedlak, P.; Merunkova, K.; Kohout, P.; Tomsovsky, M.; Baldrian, P. Stand age affects fungal community composition in a Central European temperate forest. Fungal Ecol. 2020, 48. [Google Scholar] [CrossRef]
- Enescu, R.E.; Dinca, L.; Zup, M.; Davidescu, S.; Vasile, D. Assessment of soil physical and chemical properties among urban and peri-urban forests: a case study from metropolitan area of Brasov. Forests 2022, 13. [Google Scholar] [CrossRef]
- van der Heyde, M.; Ohsowski, B.; Abbott, L.K.; Hart, M. Arbuscular mycorrhizal fungus responses to disturbance are context-dependent. Mycorrhiza 2017, 27, 431–440. [Google Scholar] [CrossRef]
- Schmidt, D.J.E.; Pouyat, R.; Szlavecz, K.; Setälä, H.; Kotze, D. J.; Yesilonis, I.; Cilliers, S.; Hornung, E.; Dombos, M.; Yarwood, S.A. Urbanization erodes ectomycorrhizal fungal diversity and may cause microbial communities to converge. Nat. Ecol. Evol. 2017, 1. [Google Scholar] [CrossRef]
- Shen, H.Y.; Yang, B.S.; Wang, H.; Sun, W.; Jiao, K.Q.; Qin, G.H. Changes in soil ectomycorrhizal fungi community in oak forests along the urban-rural gradient. Forests 2022, 13. [Google Scholar] [CrossRef]
- Arealstatistik Schweiz 2021. https://www.bfs.admin.ch/bfs/de/home/statistiken/raum-umwelt/erhebungen/area.html (accessed on 10 May 2023).
- Statistischer Atlas der Schweiz 2021. Available online: https://www.bfs.admin.ch/bfs/de/home/statistiken/regionalstatistik/atlanten/statistischer-atlas-schweiz.html (accessed on 10 May 2023).
- MeteoSwiss, 2023. Climate Normals 1981–2010. Federal Office of Meteorology and Climatology MeteoSwiss. Available online: www.meteoswiss.admin.ch (accessed on 30 April 2023).
- Burnand, J.; Hasspacher, B. Waldstandorte beider Basel. Quellen und Forschungen zur Geschichte und Landeskunde des Kanton Basel-Landschaft, 2nd ed.; Verlag des Kantons Basel-Landschaft: Liestal, Switzerland, 1999; p. 266. [Google Scholar]
- Google Earth. Available online: https://www.google.com/intl/de/earth (accessed on 12 November 2022).
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; Blackburn Press: Caldwell, NJ, USA, 2002.; p. 547. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific: Oxford, U.K, 1989; p. 368. [Google Scholar]
- Bremner, J.; Mulvaney, C. Total nitrogen. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Methods; Page, A.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis, Part 3, Chemical Methods, 3rd ed.; ASA: Madison, Wisconsin, USA, 1996, p. 1424. [Google Scholar] [CrossRef]
- White, T.J. ; Bruns,T.; Lee, S., Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications. Innis, M.A., Gelfan, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 7, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- R Core Team, 2020. R: a language and environment for statistical computing, Version 3.6.3; R Foundation for Statistical Computing: Vienna. https://www.R-project.org/.
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, R.; Stevens, M.H.H.; Wagner, H. Vegan community ecology package. R package version 2.0–10. http://CRAN.R-project.org/package=vegan (accessed on 11 May 2022).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure". Austral Ecology. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Goldmann, K.; Schröter, K.; Pena, R.; Schöning, I.; Schrumpf, M.; Buscot, F.; Polle, A.; Wubet, T. Divergent habitat filtering of root and soil fungal communities in temperate beech forests. Sci. Rep. 2016, 6, 31439. [Google Scholar] [CrossRef]
- Bahnmann, B.; Masinova, T.; Halvorsen, R.; Davey, M.L.; Sedlak, P.; Tomsovsky, M.; Baldrian, P. Effects of oak, beech and spruce on the distribution and community structure of fungi in litter and soils across a temperate forest. Soil Biol. Biochem. 2018, 119, 162–173. [Google Scholar] [CrossRef]
- Asplund, J.; Kauserud, H.; Ohlson, M.; Nybakken, L. Spruce and beech as local determinants of forest fungal community structure in litter, humus and mineral soil. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef] [PubMed]
- Clausing, S.; Likulunga, L.E.; Janz, D.; Feng, H.Y.; Schneider, D.; Daniel, R.; Kruger, J.; Lang, F.; Polle, A. Impact of nitrogen and phosphorus addition on resident soil and root mycobiomes in beech forests. Biol. Fertil. Soils 2021, 57, 1031–1052. [Google Scholar] [CrossRef]
- Gorfer, M.; Mayer, M.; Berger, H.; Rewald, B.; Tallian, C.; Matthews, B.; Sanden, H.; Katzensteiner, K.; Godbold, D.L. High fungal diversity but low seasonal dynamics and ectomycorrhizal abundance in a mountain beech forest. Microb. Ecol. 2021, 82, 243–256. [Google Scholar] [CrossRef]
- Likulunga, L.E.; Perez, C.A.R.; Schneider, D.; Daniel, R.; Polle, A. Tree species composition and soil properties in pure and mixed beech-conifer stands drive soil fungal communities. For. Ecol. Manage. 2021, 502. [Google Scholar] [CrossRef]
- Scholier, T.; Lavrinienko, A.; Brila, I.; Tukalenko, E.; Hindstrom, R.; Vasylenko, A.; Cayol, C.; Ecke, F.; Singh, N.J.; Forsman, J.T.; et al. Urban forest soils harbour distinct and more diverse communities of bacteria and fungi compared to less disturbed forest soils. Molecular Ecology 2023, 32, 504–517. [Google Scholar] [CrossRef]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Drenkhan, R.; Pritsch, K.; Buegger, F.; Padari, A.; Hagh-Doust, N.; Mikryukov, V.; Gohar, D.; et al. Regional-scale in-depth analysis of soil fungal diversity reveals strong pH and plant species effects in Northern Europe. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Christel, A.; Dequiedt, S.; Chemidlin-Prevost-Bouré, N.; Mercier, F.; Tripied, J.; Comment, G.; Djemiel, C.; Bargeot, L.; Matagne, E.; Fougeron, A.; et al. Urban land uses shape soil microbial abundance and diversity. Sci. Total Environ. 2023, 883. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Klironomos, J.N.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I.R. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Cajthaml, T.; Polme, S.; Hiiesalu, I.; Anslan, S.; Harend, H.; Buegger, F.; Pritsch, K.; Koricheva, J.; et al. Tree diversity and species identity effects on soil fungi, protists and animals are context dependent. ISME J. 2016, 10, 346–362. [Google Scholar] [CrossRef]
- Wubet, T.; Christ, S.; Schoning, I.; Boch, S.; Gawlich, M.; Schnabel, B.; Fischer, M.; Buscot, F. Differences in soil fungal communities between European Beech (Fagus sylvatica L.) dominated forests are related to soil and understory vegetation. Plos One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Kameshwar, A.K.S.; Qin, W.S. Systematic review of publicly available non-Dikarya fungal proteomes for understanding their plant biomass-degrading and bioremediation potentials. Bioresour. Bioprocess. 2019, 6. [Google Scholar] [CrossRef]
- Blackwell, M. The Fungi: 1, 2, 3… 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Nacke, H.; Goldmann, K.; Schoning, I.; Pfeiffer, B.; Kaiser, K.; Castillo-Villamizar, G.A.; Schrumpf, M.; Buscot, F.; Daniel, R.; Wubetz, T. Fine spatial scale variation of soil microbial communities under European Beech and Norway Spruce. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Kuang, J.L.; Wang, P.D.; Shu, W.S.; Barberan, A. Associations between human impacts and forest soil microbial communities. Elementa 2020, 8. [Google Scholar] [CrossRef]
- Leake, J.R.; Johnson, D.; Donnelly, D.P.; Muckle, G.E.; Boddy, L.; Read, D.J. Networks of power and influence: the role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning. Can. J. Bot. 2004, 82, 1016–1045. [Google Scholar] [CrossRef]
- Lindahl, B.; Stenlid, J.; Finlay, R. Effects of resource availability on mycelial interactions and P-32 transfer between a saprotrophic and an ectomycorrhizal fungus in soil microcosms. FEMS Microb. Ecol. 2001, 38, 43–52. [Google Scholar] [CrossRef]
- Bödeker, I.T.M.; Lindahl, B.D.; Olson, Å.; Clemmensen, K.E. Mycorrhizal and saprotrophic fungal guilds compete for the same organic substrates but affect decomposition differently. Funct. Ecol. 2016, 30, 1967–1978. [Google Scholar] [CrossRef]
- Marx, D.H. Ectomycorrhizae as biological deterrents to pathogenic root infections. Annu. Rev. Phytopathol. 1972, 10, 429–441. [Google Scholar] [CrossRef]
- Dai, D.; Yang, J.R.; Wu, Y.G.; Zhang, W.H.; Wu, X.; Liu, Y.J.; Xing, H.; Liu, Y. Correlation between fine root traits and pathogen richness depends on plant mycorrhizal types. Oikos 2023, 2023. [Google Scholar] [CrossRef]
- UN Urban Agenda 2016. URL: http://habitat3.org/the-new-urban-agenda (accessed on 18 August 2023).
Figure 1.
Location of the study area in Basel and its surroundings in northwestern Switzerland (upper left corner), and spatial distribution of the forests examined. The line marks the border of Switzerland. Forests were grouped in four classes according to the degree of urbanization of their surroundings: very low urbanized areas (rural, open dots), low urbanized areas (light grey), moderately urbanized areas (dark grey), and highly urbanized areas (black dots).
Figure 1.
Location of the study area in Basel and its surroundings in northwestern Switzerland (upper left corner), and spatial distribution of the forests examined. The line marks the border of Switzerland. Forests were grouped in four classes according to the degree of urbanization of their surroundings: very low urbanized areas (rural, open dots), low urbanized areas (light grey), moderately urbanized areas (dark grey), and highly urbanized areas (black dots).
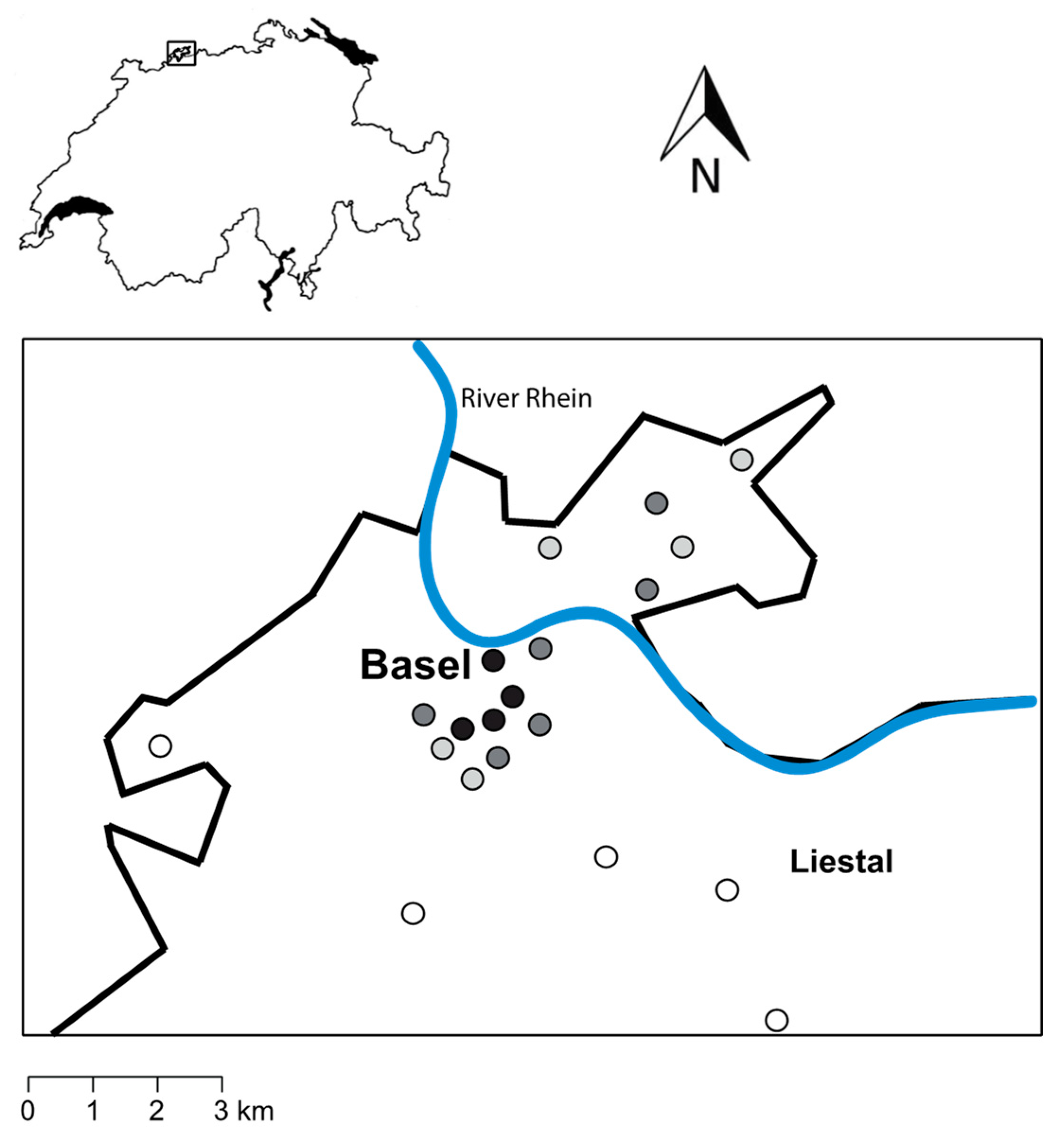
Figure 2.
Richness (a), Shannon diversity (b) and Pielou’s evenness (c) of fungal OTUs in forests in areas of very low urbanization (rural areas; white bars), in forests in low (light grey), moderately (dark grey) and highly urbanized areas (black). Mean values ± s.e. are shown.
Figure 2.
Richness (a), Shannon diversity (b) and Pielou’s evenness (c) of fungal OTUs in forests in areas of very low urbanization (rural areas; white bars), in forests in low (light grey), moderately (dark grey) and highly urbanized areas (black). Mean values ± s.e. are shown.
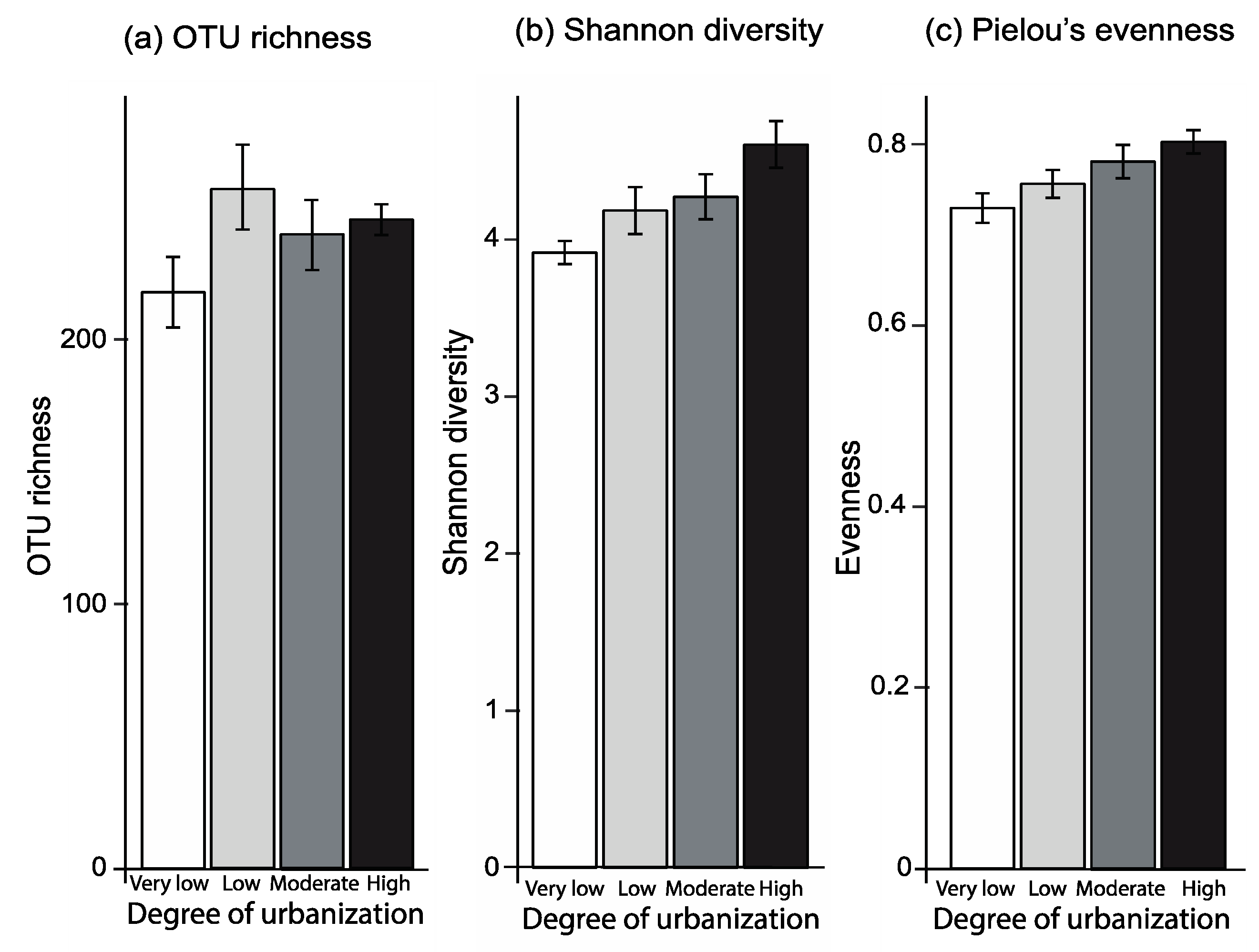
Figure 3.
Non-metric multidimensional scaling (NMDS) ordination plot based on the Bray-Curtis dissimilarities in the composition of fungal OTUs of forests in areas with extremely low urbanization (rural areas, white), and in forests in areas with low urbanization (light grey), forests in areas with moderate urbanization (dark grey) and forests in areas with high urbanization (black).
Figure 3.
Non-metric multidimensional scaling (NMDS) ordination plot based on the Bray-Curtis dissimilarities in the composition of fungal OTUs of forests in areas with extremely low urbanization (rural areas, white), and in forests in areas with low urbanization (light grey), forests in areas with moderate urbanization (dark grey) and forests in areas with high urbanization (black).
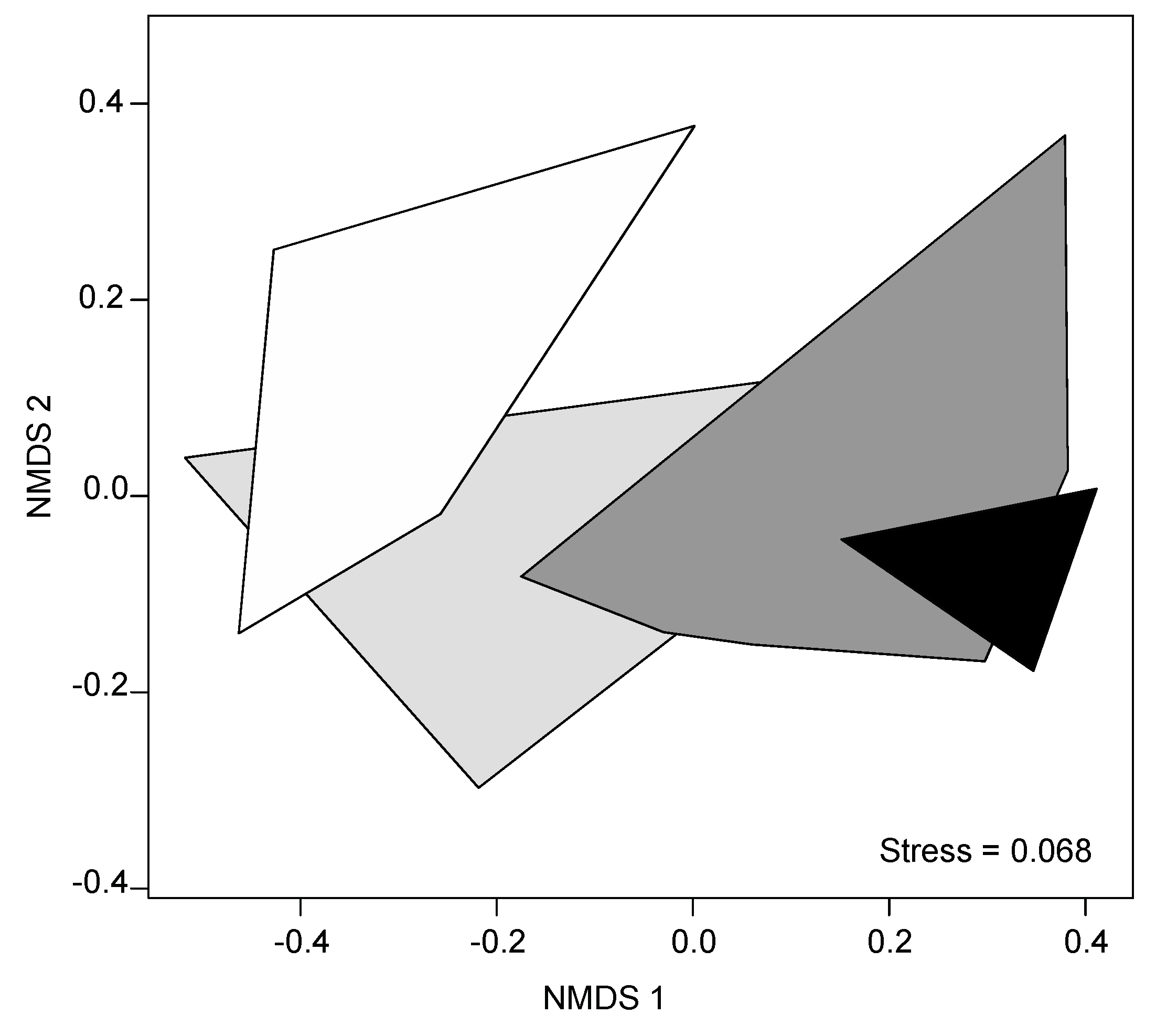
Figure 4.
Relative abundance of fungal communities at the phylum level in forests in areas with very low urbanization (rural areas), and in forests in areas with low, moderate and high urbanization. Mean values ± s.e. are shown.
Figure 4.
Relative abundance of fungal communities at the phylum level in forests in areas with very low urbanization (rural areas), and in forests in areas with low, moderate and high urbanization. Mean values ± s.e. are shown.
Figure 5.
Relative abundance of saprotrophic (a), symbiotrophic (b), and pathotrophic fungi (c) in forests in extremely low urbanized areas (rural areas; white bars), and in forests in low (light grey), moderately (dark grey) and highly urbanized areas (black). Mean values ± s.e are shown. Different letters indicate differences among the four urbanization classes based on Tukey HSD post hoc tests (p < 0.05).
Figure 5.
Relative abundance of saprotrophic (a), symbiotrophic (b), and pathotrophic fungi (c) in forests in extremely low urbanized areas (rural areas; white bars), and in forests in low (light grey), moderately (dark grey) and highly urbanized areas (black). Mean values ± s.e are shown. Different letters indicate differences among the four urbanization classes based on Tukey HSD post hoc tests (p < 0.05).
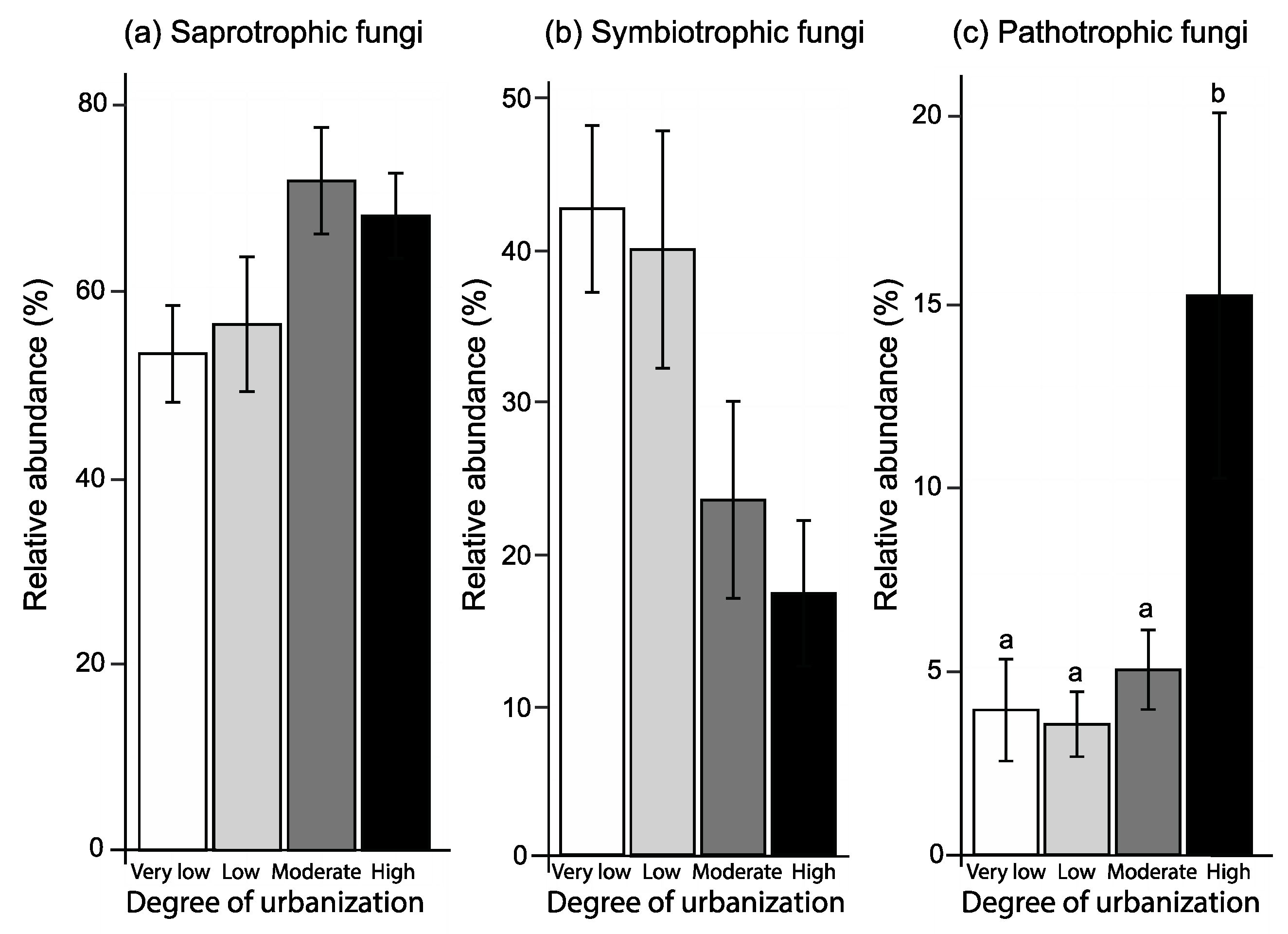
Figure 6.
Relative abundance of different fungal guilds for forests in very low (rural), low, moderately and highly urbanized areas. Mean values ± s.e. of the various fungal guilds are shown.
Figure 6.
Relative abundance of different fungal guilds for forests in very low (rural), low, moderately and highly urbanized areas. Mean values ± s.e. of the various fungal guilds are shown.
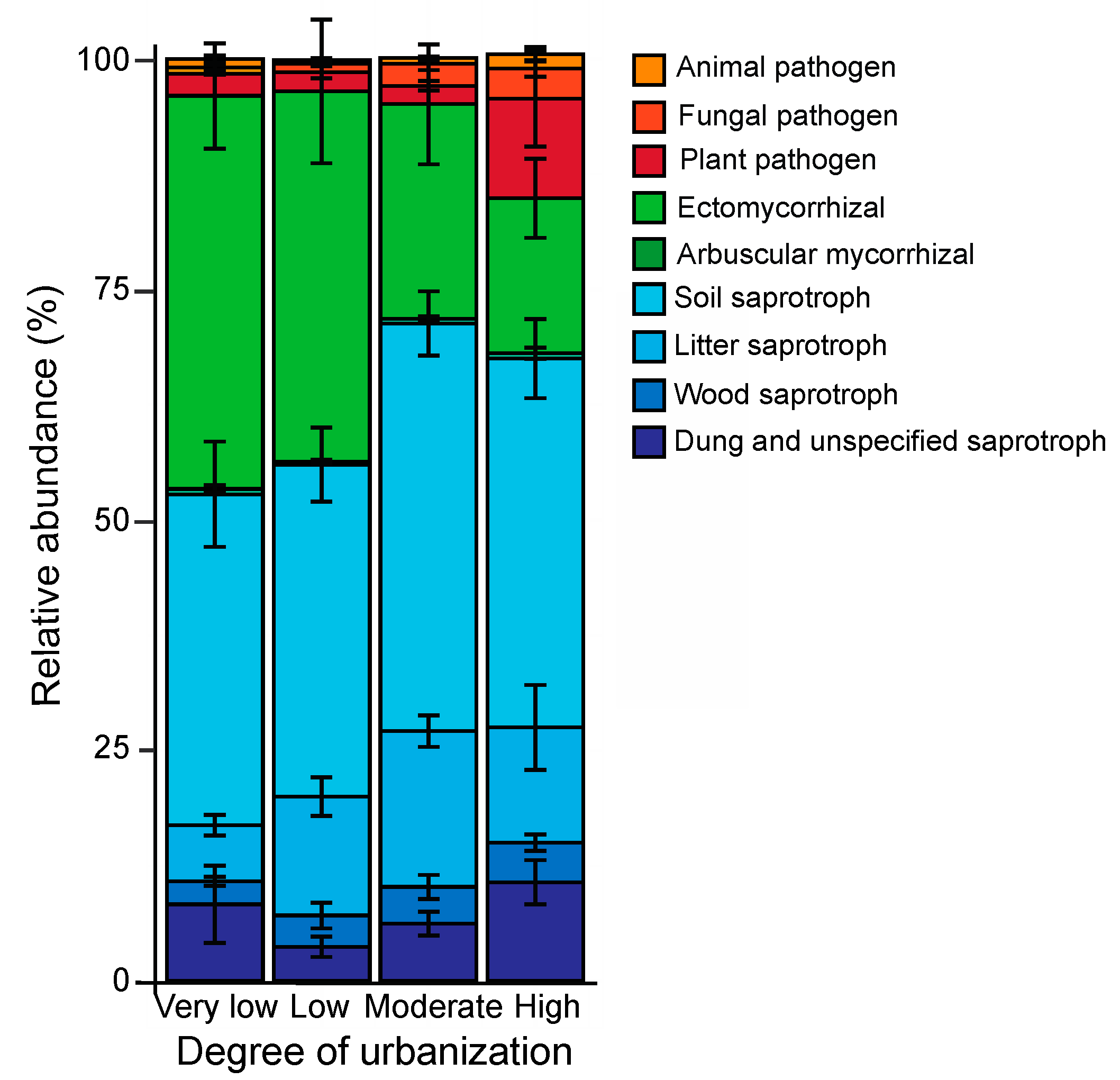

Table 1.
Characteristics of the 20 forest sites examined in the city of Basel (Switzerland) and its surroundings.
Table 1.
Characteristics of the 20 forest sites examined in the city of Basel (Switzerland) and its surroundings.
| Forest | Coordinates | Historical development1 |
Forest vegetation2 | Elevation (m a.s.l.) | Exposure3 | % cover of sealed area (r = 500 m) |
Degree of urbanization4 | Forest area (ha) | % cover of forest (r = 500 m) |
|---|---|---|---|---|---|---|---|---|---|
| BS1 | 47° 33' 13" N 7° 36' 17" E |
Planted | Galio-Fagetum Pulmonarietosum | 363 | WNW | 59 | 4 | 0.33 |
2 |
| BS2 | 47° 33' 14" N 7° 36' 49" E |
Fragment | Galio Odorati-Fagetum Cornetosum | 262 | NE | 39 | 3 | 1.42 | 3 |
| BS3 | 47° 33' 55" N 7° 38' 41" E |
Planted | Galio Odorati-Fagetum Pulmonarietosum | 319 | NNW | 30 | 3 | 0.41 | 56 |
| BS4 | 47° 32' 12" N 7° 36' 6" E |
Fragment | Galio Odorati-Fagetum Cornetosum | 321 | NE | 54 | 4 | 1.16 | 13 |
| BS5 | 47° 32' 04.6" N 7° 31' 16.2" E |
Forest | Galio-Fagetum Pulmonarietosum | 351 | – | 1 | 1 | 76.2 | 45 |
| BS6 | 47° 34' 53" N 7° 38' 52" E |
Planted | Galio Odorati-Fagetum | 283 | – | 33 | 3 | 0.33 | 1 |
| BS7 | 47° 32' 18" N 7° 35' 39" E |
Planted | Aro-Fagetum | 325 | NE | 43 | 3 | 0.23 | 6 |
| BS8 | 47° 31' 49" N 7° 35' 49" E |
Fragment | Galio Odorati-Fagetum Typicum | 370 | E | 23 | 2 | 2.70 | 11 |
| BS9 | 47° 31' 55 N 7° 36' 6" E |
Fragment | Galio Odorati-Fagetum Typicum | 338 | NW | 44 | 3 | 2.10 | 19 |
| BS10 | 47° 34' 20" N 7° 37' 6" E |
Forest | Galio-Carpinetum Corydalidetosum | 269 | – | 25 | 2 | 2.53 | 35 |
| BS11 | 47° 29' 11" N 7° 40' 43" E |
Forest | Galio-Fagetum Pulmonarietosum | 565 | – | 3 | 1 | 186.4 | 92 |
| BS12 | 47° 34' 29" N 7° 39' 58" E |
Forest | Galio Odorati-Fagetum Cornetosum | 450 | NW | 10 | 2 | 5.15 | 37 |
| BS13 | 47° 35' 18” N 7° 40' 20” E |
Forest | Galio Odorati-Fagetum Cornetosum | 473 | SW | 13 | 2 | 3.42 | 54 |
| BS14 | 47° 32' 31" N 7° 35' 2" E |
Fragment | Galio Odorati-Fagetum Cornetosum | 299 | NNE | 35 | 3 | 1.95 | 6 |
| BS15 | 47° 30' 53" N 7° 38' 11" E |
Forest | Galio-Fagetum Pulmonarietosum | 418 | – | 2 | 1 | 79.0 | 43 |
| BS16 | 47° 30' 31" N 7° 40' 04" E |
Forest | Aro-Fagetum | 454 | – | 1 | 1 | 337.0 | 66 |
| BS17 | 47° 32' 43" N 7° 36' 27" E |
Planted | Aro-Fagetum | 276 | – | 69 | 4 | 0.37 | 2 |
| BS18 | 47° 30' 18" N 7° 34' 46" E |
Forest | Galio Odorati-Fagetum Typicum | 380 | – | 2 | 1 | 237.7 | 59 |
| BS19 | 47° 34' 23" N 7° 39' 16" E |
Planted | Galio Odorati-Fagetum Typicum | 380 | – | 9 | 2 | 1.28 | 38 |
| BS20 | 47° 32' 14" N 7° 35' 26" E |
Fragment | Galio Odorati-Fagetum Cornetosum | 326 | E | 56 | 4 | 0.89 | 5 |
1 Forest = part of a large continuous forest; fragment = remnant of a former large continuous forest; planted = trees planted after 1884. 2 Plant association following Burnand and Hasspacher [37]. 3 Exposure was determined for forest sites situated on slopes. “–“ indicates flat forest sites. 4 Four classes: 1 = very low (cover of sealed area <5% within a radius of 500 m); 2 = low (5.1–25%); 3 = moderate (25.1– 45%); 4 = high (>45%).
Table 2.
Vegetation characteristics and soil properties of the 20 deciduous forests examined along an urbanization gradient.
Table 2.
Vegetation characteristics and soil properties of the 20 deciduous forests examined along an urbanization gradient.
| Degree of urbanization | |||||
|---|---|---|---|---|---|
|
Very low (rural) (n = 5) |
Low (n = 5) |
Moderate (n = 6) |
High (n = 4) |
p | |
| Forest vegetation characteristics | |||||
| Ground vegetation cover (%) | 67.2 ± 9.5 | 56.2 ± 11.7 | 78.0 ± 18.1 | 73.9 ± 15.9 | N.S. |
| Plant species richness1 | 9.5 ± 1.4a | 6.8 ± 0.6a | 5.6 ± 0.4b | 5.2 ± 0.2b | 0.005 |
| Shrub species richness2 | 4.0 ± 0.7a | 4.0 ± 0.9a | 6.0 ± 0.8b | 7.5 ± 0.3b | 0.034 |
| Tree species richness2 | 3.8 ± 0.6 | 3.8 ± 0.5 | 3.8 ± 0.6 | 2.5 ± 0.8 | N.S. |
| Soil properties | |||||
| Soil moisture (%) | 31.4 ± 2.3 | 29.0 ± 1.5 | 28.8 ± 1.6 | 25.7 ± 2.1 | N.S. |
| Soil pH | 5.6 ± 0.4a | 5.7 ± 0.4a | 6.6 ± 0.2b | 7.2 ± 0.1b | 0.004 |
| SOM (%) | 18.3 ± 6.9 | 12.7 ± 1.9 | 16.6 ± 2.2 | 22.2 ± 3.4 | N.S. |
| Total soil organic nitrogen (%) | 0.298 ± 0.071 | 0.282 ± 0.034 | 0.313 ± 0.033 | 0.381 ± 0.047 | N.S. |
| Plant-available phosphorus3 | 26.9 ± 5.0a | 19.4 ± 3.9a | 35.8 ± 8.5b | 46.4 ± 5.3b | 0.07 |
1 Number of plant species per 4 m2. 2 Number of shrub and tree species per 100 m2. 3 μg PO43–/g. Mean values ± s.e. are shown. n refers to the number of forests studied. Different letters indicate differences between the four urbanization classes based on Tukey HSD post hoc tests (p < 0.05). N.S., not significant. P-values resulted from ANCOVA or GLM analyses (see detailed description in 2.6. Data Analyses).
Table 3.
Summary of general linear models (GLM) analysing the effects of degree of urbanization (four classes), forest size, percentage of area covered with forest within 500 m of the centre of the study plots, forest vegetation characteristics and soil properties on richness, diversity and evenness of fungal OTUs.
Table 3.
Summary of general linear models (GLM) analysing the effects of degree of urbanization (four classes), forest size, percentage of area covered with forest within 500 m of the centre of the study plots, forest vegetation characteristics and soil properties on richness, diversity and evenness of fungal OTUs.
| OTU richness | Shannon diversity | Pielou’s evenness | |
|---|---|---|---|
| Degree of urbanization | Chi23,16 = 16.79,p < 0.001 | F3,14 = 4.22, p = 0.026 | F3,11 = 5.14, p = 0.018 |
| Forest area (ha)1 | Chi21,15 = 1.47, p = 0.226 | – | F1,11 = 3.51, p = 0.088 |
| % forest within 500 m1 | Chi21,14 = 5.90, p = 0.015 | F1,14 = 1.82, p = 0.197 | F1,11 = 1.08, p = 0.322 |
| Plant species richness1 | – | – | F1,11 = 2.82 p = 0.123 |
| Tree species richness1 | Chi21,13 = 1.82, p = 0.177 | – | – |
| Soil moisture | Chi21,12 = 8.20, p = 0.004 | F1,14 = 1.68, p = 0.216 | F1,11 = 5.05, p = 0.046 |
| Soil pH | – | – | F1,11 = 1.23, p = 0.290 |
1 log transformed. Significant p-values (< 0.05) are indicated in bold. – Removed from the model by the stepwise reduction procedure.
Table 4.
Summary of general linear models (GLM) analyzing the effects of degree of urbanization (four classes), forest vegetation characteristics and soil properties on the relative abundance of saprotrophic, symbiotrophic and pathotrophic fungi.
Table 4.
Summary of general linear models (GLM) analyzing the effects of degree of urbanization (four classes), forest vegetation characteristics and soil properties on the relative abundance of saprotrophic, symbiotrophic and pathotrophic fungi.
| Relative abundance of | |||
|---|---|---|---|
| Saprotrophic fungi | Symbiotrophic fungi | Pathotrophic fungi | |
| Degree of urbanization | F3,13 = 3.16, p = 0.061 | F3,12 = 4.37, p = 0.026 | F3,13 = 5.24, p = 0.014 |
| Ground vegetation cover (%) | F1,13 = 1.47, p = 0.247 | F1,12 = 2.08, p = 0.175 | F1,13 = 1.25, p = 0.284 |
| Plant species richness1 | F1,13 = 2.55, p = 0.141 | F1,12 = 1.22, p = 0.290 | – |
| Tree species richness1 | – | F1,12 = 1.85, p = 0.198 | – |
| Soil moisture (%) | F1,13 = 4.10, p = 0.046 | – | F1,13 = 3.46, p = 0.084 |
| Soil pH | – | F1,12 = 3.17, p = 0.100 | F1,13 = 2.20, p = 0.162 |
1 log transformed. Significant p-values (< 0.05) are indicated in bold. – Removed from the model by the stepwise reduction procedure.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



