
Submitted:
17 September 2023
Posted:
18 September 2023
Read the latest preprint version here
Abstract
Pathogens, especially invasive species, have caused significant global ecological, economic, and social losses in forests. Plant disease research has traditionally focused on direct interactions between plants and pathogens in an appropriate environment. However, recent research indicates that the microbiome can interact with the plant host and pathogens to modulate plant resistance and/or pathogen pathogenicity, and thereby altering the outcome of plant-pathogen interactions. Thus, this presents new opportunities for studying the microbial management of forest diseases. Compared to parallel studies on human and crop microbiomes, research into the forest tree microbiome and its critical role in forest disease progression has lagged. The rapid development of microbiome sequencing and analysis technologies has resulted in the rapid accumulation of a large body of evidence regarding the association between forest microbiomes and diseases. These data will aid the development of innovative, effective, and environmentally sustainable microbial management of forest diseases. Herein, in this review, we summarize the most recent findings on the dynamic structure and composition of forest tree microbiomes in belowground and aboveground plant tissues (i.e., rhizosphere, endosphere, and phyllosphere), as well as their pleiotropic impact on manipulating plant immunity and pathogen pathogenicity, highlighting representative examples of Biological Control Agents used to modulate relevant tree microbiomes. Lastly, we discuss the potential application of forest tree microbiomes in disease control as well as their future prospects and challenges.
Keywords:
microbiome
; rhizosphere
; phyllosphere
; endosphere
; plant immunity
; pathogen suppression
; biological control agents
; woody plants
1. Introduction
Forests provide a multitude of benefits to humans, including climate regulation, water supply and regulation, habitats for biodiversity. Nevertheless, the current condition of global forest resources does not bode well for their conservation [1]. Apart from human deforestation, outbreaks of old diseases and the emergence of new diseases is one of the most influential factors threatening forest resources. Typical examples of devastating diseases include chestnut ink disease (Phytophthora cinnamomi and Phytophthora cambivora) [2], chestnut blight (Cryphonectria parasitica) [3], Dutch elm disease (Ophiostoma ulmi and O. novo-ulmi) [4], sudden oak death (Phytophthora ramorum) [5], pine wilt disease (Bursaphelenchus xylophilus) [6], ash dieback (Hymenoscyphus fraxineus), and laurel wilt (Raffaelea lauricola) [7]. These diseases have led to the disappearance of the forest trees and the degradation of forest ecosystems. The occurrence of forest diseases in nature requires a susceptible host, a pathogen, and suitable environmental conditions (e.g., temperature and humidity) [8,9,10,11], as explicitly stated by the disease triangle theory, a principle of plant pathology. However, studies have traditionally focused on the interaction between a single woody plant and a single pathogen, with little attention paid to the role that environmental factors (e.g., biological and abiotic factors) play in pathogenic colonization and pathogenicity. In this disease triangle, the influence of other woody plant (WP)-colonizing microorganisms, namely the microbiota, on pathogen colonization and plant defense against infection is not considered.
Plant microbiomes exist widely in rhizosphere, phyllosphere, and inside plants. Microbiome-wide association studies (MWAS) have been widely used to investigate the links between the altered microbiome and the development of human diseases, including, but not limited to, type 2 diabetes, rheumatoid arthritis, liver cirrhosis, obesity, and colorectal cancer [12]. Due to its present success in the field of human and animal diseases, MWAS have been broadened to investigate the association between the plant microbiome and disease [13,14]. Accumulating studies suggest that, like the human microbiome, the plant microbiome also plays an important role in promoting plant growth and development, nutrient uptake, and disease defense [15,16]. Plant microbiomes are not static. Their structure and function change in response to pathogen infection, to directly or indirectly regulate the plant immune system, ultimately determining plant health. As a result, some related concepts such as Core Microbiome (CM), Synthetic Microbiome (SynComs), Defense Microbiome (DefenseBiome), and Agricultural Precision Microbiome have evolved. Our understanding of the plant microbiome and its function in plant health has advanced considerably in recent years due to the rapid development of microbiome-related technologies and approaches. The attention from the initial description stage to mechanisms governing the assembly and function of microbiota, and potential applications in integrated plant management programs was shifting. Key drivers affecting the assembly of plant-associated microbiota are identified by microbiome-genome-wide association studies (mGWAS) and metagenomic association studies, which also link certain microbial species to genes involved in plant colonization, plant physiology, and fitness-related features. Metaproteomic and metabolomics technologies have been used to mine and identify key proteins, signaling molecules, hormones, and secondary metabolites in the microbiome, thereby revealing new mechanisms by which the microbiome aids plant defense against disease. In this context, forest pathologists also sense an opportunity to study on the relationships between taxonomic composition and functioning of the WP microbiota and disease control [17,18,19,20,21].
Strengthening the prevention and control of forest diseases and protecting the healthy production of forest trees are major issues facing the development of the forest industry [22]. Trees possess their own unique attributes, such as long generation times, heterogeneous materials, large size, lignification, and complex relationships with other biological populations. As a result, research into the forest tree microbiome has significantly lagged behind that of gramineous and herbaceous plants. Therefore, in-depth and extensive research into forest tree microbiomes will provide new strategies for the introduction of new BCAs for high-efficiency seedling breeding, disease-resistant breeding, and biological control. This will also contribute to the sustainable production of wood and the ability of forest ecosystems to deal with environmental pressure. In this review, we discuss the ways in which the forest tree microbiome (mainly the rhizosphere and endosphere microbiomes) protect plants against disease, various changes that can occur within the forest tree microbiome upon pathogen infection, and prospects for the integration of tree-pathogen-microbiome interactions into forest health management.
2. Tree Rhizosphere Microbiome-Mediated Protection against Pathogens
In the natural environment, WP-microbial interactions (in different forms, namely antagonism, amensalism, parasitism, and symbiosis) occur inside (such as in the leaf endosphere and root endosphere) and on WP surfaces (such as the rhizosphere and phyllosphere) [23,24] (Figure 1). The above- and belowground microbial communities are actively recruited from the surrounding microbial reservoir to promote plant growth and development, enhance stress tolerance, and improve disease resistance. Plant roots provide a unique niche for soil microbial communities, attracting a wide variety of microbial communities which are then distributed in the rhizosphere, roots, and surface. Among them, the rhizosphere is a hotspot of microbial habitat and activity, and an important location for energy flow and material exchange between the plant roots and soil [25,26]. The selective recruitment of microorganisms from bulk soil and their interrelated systems is called the rhizosphere microbiome (Figure 1). The interaction between the rhizosphere microbiome and the plant expands the functional scope of the host plant.
2.1. Soil-Borne Pathogens and Beneficial Microbes in the Tree Rhizosphere
Compared to herbaceous and cereal plants, the rhizosphere zone of woody plants is larger, contains more microorganisms, has increased variety, and is more active. When certain microorganisms expand and reproduce in large numbers, they have the potential to cause root diseases in forest trees. For example, P. cinnamomic is a destructive and widespread soil-borne pathogen that infects woody plant hosts, such as eucalyptus, oaks, chestnuts, and pines. [27]. Ralstonia solanacearum causes bacterial wilt in eucalyptus [28] and fig trees [29]. Ceratocystis sp. causes damping off in many forestry species, including Acacia spp., Eucalyptus spp., and Quercus spp. [30,31]. Vascular wilt is caused by the fungus Verticillium dahliae, a classical soil-borne disease that presents a threat to olive trees [32], smoke trees [33], and many other tree hosts [34].
Approximately 2 – 5% of rhizosphere microorganisms mainly form two types of symbioses [arbuscular mycorrhiza (AM) and ectomycorrhiza (ECM)] with forest roots, which have a significant impact on promoting plant growth and reducing plant diseases [35,36,37,38]. Some can be used as biocontrol agents (bacteria: Bacillus and Pseudomonas, fungi: Trichoderma and yeast, and actinomycetes: Streptomyces) to inhibit pathogen growth and reproduction by competing with pathogens for rhizosphere nutrition and niches, or by secreting siderophile elements, hydrolytic enzymes, volatile organic compounds, and antibiotics, thereby indirectly promoting plant growth [17,39]. For instance, the Pseudomonas fluorescens strain, PICF7, has been applied to olive trees to induce host resistance to Verticillium wilts, caused by the pathogen V. dahliae [40,41]. Some plant growth-promoting rhizobacteria (PGPR) can function as biological triggers to induce resistance, plant disease severity and incidence under greenhouse and field conditions [42].
2.2. Root Exudates and Microbial signal Molecules Affecting Rhizosphere Microbiome of Trees
Trees can change the composition of rhizosphere microbiota by secreting bioactive exudates (e.g., organic acids, amino acids, fatty acids, phenolics, plant growth regulators, nucleotides, sugars, putrescine, sterols, and vitamins) into the rhizosphere region [43,44,45,46]. Root exudates, in addition to serving as carbon and nitrogen substrates for microbial growth, have a variety of impacts on rhizosphere microbes, acting as signaling molecules, attractants, stimulants, but also as inhibitors or repellents [47,48]. For example, Rudrappa, et al. [49] showed that L-malic acid, an intermediate product of the tricarboxylic acid cycle, which is secreted from Arabidopsis roots, selectively signals and recruits the beneficial rhizobacterium Bacillus subtilis in a dose-dependent manner, resulting in a stronger immune response to Pseudomonas syringae pathovar tomato (Pst). Salicylic acid (SA), a defense phytohormone secreted in the Arabidopsis root zone, has been reported by Lebeis, et al. [50] to modulate colonization of the root microbiome by specific bacterial taxa, thereby contributing to the root microbiome composition. Banana root exudates contain fumaric acid, which attracts B. subtilis N11 and promotes biofilm formation [51]. Flavonoids secreted by the roots of the Chinese tallow tree (Triadica sebifera) can increase AM fungal spore germination and colonization and may also impact soil microbes and other soil processes [52].
Since the types and concentrations of root exudates vary among different plant species, different natural populations within the same plant species, and further different plant growth stages, nutritional status, and stress exposure [45,53,54], host genotype play an important role in determining the composition of the rhizosphere microbiome [55]. In addition, rhizosphere microbes can also produce and release signal molecules (e.g., volatile organic compounds, oxalic acid, glucose) or extracellular metabolites (e.g., amino acids, vitamins), which can change both microbe-to-microbe and plant-to-microbe communication in the rhizosphere. For example, fungal hyphae are used by bacteria for migration into soils in an interaction, known as “fungal highways”. This process is a synergistic example of inter-microbial assisted dispersal [56,57]. Further, N-acyl-homoserine lactone (AHL) quorum-sensing (QS) signaling molecules are produced by PGPRs, which are responsible for plant growth promotion [58] and the induction of protection against pathogens [59]. Microbial volatile organic substances (VOCs), including 2,3-butanediol, 3-phenyproprionic acid, chokol K, and 3-octanone, function as chemical weapons against other microbes, as regulators of plant growth and stress resistance, and as signals mediating intra- and interspecies communications [60,61,62].
2.3. Root Microbiome Enhances Plant Disease Resistance
The interactions between plants and the rhizosphere microbiome can generate a strong selective pressure that shapes the rate and pattern of microbial evolution and eventually influences the composition of the rhizosphere microbiome [63]. In response to pathogen attack, plants can send specific signals that favor the recruitment of beneficial microbes that are able to enhance plant defenses or directly hinder pathogen growth and invasion. This phenomenon is known as the “cry for help hypothesis” [64,65,66].
In comparison to annual and/or herbaceous plants, perennial woody plants can establish long-term associations with their associated microbiome. For most crops used in previous studies, short growth periods limit the stability of the rhizosphere microbiome. Therefore, their role in plant resistance is easily disturbed by environmental conditions [67,68]. As a result, it is difficult to gain additional insights into the importance of plant species selection and the impact of environmental filtering on the composition of the rhizosphere microbiome during short-term growth. Conversely, due to long-term interactions between trees and the soil microbiome, trees can establish relatively stable rhizosphere microbiome characteristics and plant functional traits [47,69] that support resistance to biotic stresses from soil-borne pathogens. Taken together, optimizing the tree rhizosphere microbiome may be an effective way to improve forest health [70,71,72].
3. Tree Rhizosphere Microbiome-Mediated Protection against Pathogens
The phyllosphere (all aboveground compartments of a plant, often leaves) accounts for a substantially larger proportion of a terrestrial plant than the root system. Leaves, with a global leaf envelope estimated to be about 109 km2 [73], represent one of the most rich habitats on earth [74], and are inhabited on (i.e., epiphytic) and inside (i.e., endophytic) by a diverse range of microbes, primarily bacteria, filamentous fungi, and yeast strains [75], followed by protists [76] and bacteriophages [77] (Figure 1). However, in comparison to those of rhizosphere microbes, the functional roles of phyllosphere microbes remain a subject that has been less explored. Emerging evidence suggests that the phyllosphere microbiome also plays a crucial role in plant health and growth [75,78]. Yet, the underlying mechanism through which the phyllosphere microbiome interacts with the host immune system and consequently confers host plant development and defense remains largely unknown.
3.1. Multiple Factors Drive the Colonization of Microbial Aggregates on the Phylloplane
The colonization source of the phyllosphere is relatively less certain. Recent evidence suggests that phylloplane microorganisms are ultimately derived from soil [79]. At the same time, extensive studies are available for the soil and rhizosphere bacterial community on phyllosphere bacterial colonization in Arabidopsis thaliana [80,81,82,83] and maize [84]. Bacteria are the most abundant and diverse microbial group in the phyllosphere [74], but they are less diverse than those in the rhizosphere or soil [85,86]. Izhaki, et al. [87] identified 32 bacterial species associated with Citrus paradisi leaf surfaces. Epiphytic mycobiota on the phylloplane of two deciduous trees and three evergreen shrubs increases during plant growth [88]. In characterizing phyllosphere bacterial communities in a neotropical forest, Kembel, et al. [89] showed that leaves from individual trees were habitat to more than 400 bacterial taxa and that a core microbiome of taxa including Actinobacteria, Alpha-, Beta-, and Gammaproteobacteria, and Sphingobacteria dominated phyllosphere bacterial communities. Generally, the bacterial phyla belonging to Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes were found to be the most abundant in several plant phyllospheres, including A. thaliana, common ash (Fraxinus excelsior), poplar, and other tree leaves [80,82,89,90,91,92,93].
Usually, soil types and properties, climate, the genetic traits of the plant (genotype and phenotype), geographical location, and attack by pathogens, etc. are drivers that contribute to the assembly of the phyllosphere bacterial communities[94,95,96]. However, how these factors affect the microbial communities in the phyllosphere is not fully understood. Moreover, there is still controversy about the main determining drivers. Finkel, et al. [97] found that different species of the salt-excreting desert Tamarix trees (T. aphylla, T nilotica, T. tetragina) grown in the same geographical location possessed highly similar bacterial communities, suggesting that geographical location rather than plant species was found to be the major determinant of phyllosphere bacterial community composition. Afterwards, sampling T. aphylla leaves growing across the Sonoran Desert in the Southwestern United States provided additional evidence of the main impact of geographic distances on phyllosphere bacterial communities [98]. However, some other studies offered contradictory insights that the genetic traits of the plant drive the bacterial community composition on leaf surfaces. For example, regardless of the geographical location where the leaf samples were collected, the phyllosphere bacterial communities associated with Pinus ponderosa were remarkably similar to each other. Additionally, leaves from 56 distinct tree species collected from the same geographical location contained bacterial populations unique to each plant species [93]. Laforest-Lapointe, et al. [99] in a study of leaves of five tree species also demonstrated that host species identity is a stronger driver of temperate tree phyllosphere bacterial communities than forest sites or time.
The fungal microbiota of the phyllosphere consists of two dominant groups: yeast and filamentous fungi [100]. On healthy plant leaves, yeasts are the main foliar epiphytic fungi. Because they contain many pathogens on plant systems, phyllosphere fungi have received a lot of attention. Infectious filamentous fungi must survive in a harsh and unstable phyllosphere environment, such as one with nutrient deprivation, high UV radiation, temperature and humidity fluctuations, etc. [100], and need to escape from various direct and indirect physical damages until conditions are suitable for plant infestation. The filamentous fungal communities in the phyllosphere were highly diverse at all stages of plant growth, including a variety of pathogenic and non-pathogenic fungi such as Cladosporium, Alternaria, Fusarium, Penicillium, Acremonium, Colletotrichum, Mucor, and Aspergillus [88,101,102]. Upon global warming, the beneficial microbiota in the phyllosphere were found to decrease, whereas pathogens were enriched [103,104]. Until now, the main drivers regarding phyllosphere fungal communities have been equally difficult to summarize. For instance, soil is the main driver in determining fungal community composition, diversity, and abundance on oak (Quercus macrocarpa) leaves [105]. Host genetics is a determinant of fungal community assembly on beech (Fagus sylvatica) leaves [100]. Similarly, host genotype is the most influential factor determining the assembly of the foliar fungal microbiome of balsam poplar (Populus balsamifera) [106,107]. In general, the composition of phyllosphere microorganisms is the combined result of flora competition, climate selection, and host selection [93,108,109,110,111,112].
3.2. Stability of Microbial Consortia against Pathogen Perturbation
The leaf surface creates a stressed and unstable conditions, including deficient in nutrient supplies, harmful ultraviolet (UV) radiation, oxidative stress, low water, fluctuating temperature through the day and biotic stress. The phyllosphere associated microbes, no matter whether they are commensal, beneficial, or detrimental, will employ distinct fitness strategies (tolerance or avoidance strategy) to cope with this combination of stresses and eventually for successful colonization [113]. For example, nonpathogenic phyllosphere-colonizing bacteria actively impact on the plant and gain fitness advantages by adjusting plant hormone mimics (i.e., auxins and cytokinins), secreting biosurfactants, and mediating plant pathogen interactions [75,114,115]. Finally, some of these host-adapted microbial colonizers–the epiphytes– stay on the surface of plant organs, while others are able to penetrate further inside the plants and are called endophytes [116]. The stability of the leaf-associated microbial community is known to be closely related to plant health. Recent studies also revealed that plants control phyllosphere microbiota to ensure plant health. Chen, et al. [117] reported that mutations in genes involved in pattern-triggered immunity (PTI), vesicle trafficking and cell death activation cause marked changes in the leaf microbiome of A. thaliana (an overall increased bacterial population size, decreased community diversity, and a shift from a Firmicutes-rich community to a Proteobacteria-rich community) that result in leaf damage (chlorosis and/or necrosis) phenotypes, suggesting that mechanisms by which terrestrial plants control endophytic microbial abundance and diversity in the phyllosphere to prevent dysbiosis and maintain plant health. To our best knowledge, this is the first report regarding the genetic control of microbiome assembly in the phyllosphere and how this affects plant health. However, significant knowledge gaps still exist about this part.
3.3. Phyllosphere-Mediated Resistance to Pathogen Invasion
Plants can employ the “cry for help” strategy whereby they selectively recruit beneficial microbes from the environment that can help to combat pathogen stress [118,119,120]. Such plant-microbe interactions provide a protective effect on plant health [121]. Not only did rhizosphere microbiome responses play an important role in pathogen resistance and plant health [66,122,123], but so did the phyllosphere microbiome. For example, Berg and Koskella [124] found that the phyllosphere microbiota of tomato plants conferred protection against the causal agent of bacterial speck, Pst. Furthermore, the degree of protection depends on the initial dose of microbiota being applied. Similarly, considerable changes in leaf-associated fungal and bacterial communities were observed upon powdery mildew infection in oak (Quercus robur) [125] and upon Huanglongbing (HLB) infection in citrus [126], respectively. The Jakuschkin study further investigated the pathobiontic network of different fungal and bacterial operational taxonomic units that were connected with each other and directly interacting with Erysiphe alphitoides, suggesting that some of these might have been conferring protection to the oak phyllosphere by inhibiting E. alphitoides and thereby reducing the probabilities of powdery mildew occurrence. More recently, Li, et al. [127] reported that the phyllosphere microbiota of infected citrus leaves displayed a more intense microbial network and larger numbers of the native microbes (e.g., Sphingomonas spp.) and the recruited new microbes (e.g., Pantoea asv90 and Methylobacterium asv41) compared to those of uninfected leaves by the melanose pathogen D. citri. Further, Sphingomonas spp. exhibited promising results in protecting the citrus phyllosphere from pathogen invasion via its iron-competition capability. Overall, the roles of phyllosphere microbiomes in the resistance to pathogen invasion and the contribution to plant health have just started to be recognized and provide a significant opportunity to develop microbiome-based tools for disease prevention or prediction.
4. Contribution of Tree Endosphere Microbiome in the Control of Forest Diseases
Endophytes that colonize within plant can confer ecological advantages to the host plants and improve plant fitness, (i.e., improving nutrient uptake, promoting growth and development, imparting tolerance to abiotic stresses, and providing resistance to pathogens) [128]. Endophytes have received attention in relation to improved disease resistance in Poaceae Barnhart and Brassicaceae Burnett plants compared to forest plants. Endophytes secrete specialized metabolites or bioactive compounds that help keep plants away from pathogens by means of antagonism, mycoparasitism, and induction of plant defense responses. For example, endophytic bacteria, produce defensive enzymes (such as polyphenol oxidase, chitinase, β-1,3-glucanase, and peroxidase) [129,130] and phytohormones [indole acetic acid (IAA), ethylene (ET), abscisic acid (ABA), SA, jasmonic acid (JA)] [131], and secondary metabolites (phenols, antibiotic, fungicides) [132,133,134,135], to enable plants to establish a robust resistance against pathogens.
Endophytes may underscore the induction of defense-related genes and confer pathogens resistance [136]. For example, the fungal leaf endophyte Colletotrichum tropicale induces the expression of hundreds of host defense-related genes in Theobroma cacao, resulting in greater plant immunity [137]. Moreover, Phoma medicaginis, which causes alfalfa leaf spots, is resisted by arbuscular mycorrhizal fungus (AMF) by activating defense activities, including JA, SA, peroxidase (POD), and polyphenol oxidase (PPO) [138]. Pathogen-induced activation of enzyme-coding genes associated with fungal cell-wall degradation and biosynthetic gene clusters encoding non-ribosomal peptide synthases (NRPSs) and polyketide synthases (PKSs) are responsible for disease-suppressive functions in endophytes [66]. The endophytic microbiome can provide an extra layer of defense by selectively enriching microbes with genetic machinery that produces enzymes and secondary metabolites against pathogens [66,71].
Relatively little research has been conducted on the use of biocontrol endophytes to control forest pests and diseases. Most of the limited available studies largely focus on identifying beneficial endophytes that antagonize pathogens and structurally characterizing their metabolic antimicrobial compounds [139,140]. An example of bioactive natural compounds identified from beneficial endophytes of woody perennials include, phomopsolides and the stable α-pyrone isolated from the conifer endophyte Diaporthe maritima, respectively, exhibited antifungal activity towards Microbotryum violaceum and B. subtilis pathogens [141]. In addition, Sumarah, et al. [142] identified five foliar fungal endophytes isolated from Pinus strobus (eastern white pine) that produce potently antifungal compounds. Further investigation revealed that the pyrenophorol in the culture filtrates of the pine endophyte Lophodermium nitens was responsible for suppressing growth in the pine pathogen Cronartium ribicola [143]. More bioactive antimicrobial metabolites (lipopeptides, pyrrolizin, glucoamylase, garcinia cambogia chlorophyll, and chitinase) were shown to inhibit or kill pathogens, which alleviated forest tree diseases [144]. Thus, the capacity of endophytes to alter interactions with pathogens may provide integrated disease management tools. However, the specific mechanisms underlying endophyte-mediated pathogen resistance in these studies are not fully understood.
5. Pathogen Invasion Triggers Innate Immune Responses in Plants
Plants make good use of two types of immune receptors, located on the cell surface and inside the cell, which can recognize invasive microorganisms (e.g., viruses, bacteria, fungi, oomycetes, nematodes, insects) and activate protective immune responses in the plant. However, when pathogens infect host plants, they secrete effectors into the host cells or extracellular space to interfere with host physiological activities to facilitate their infection or colonization. Cell surface-localized immune receptors, also known as “pattern-recognition receptors (PRRs)”, consist mainly of single-transmembrane receptor-like kinases (RLKs) and receptor-like proteins (RLPs) that detect pathogen-associated molecular patterns (PAMPs), such as bacterial flagellins, bacterial elongation factors, fungal chitin polysaccharides, fungal xylanases, and endogenous elicitors. The immunity triggered by PRRs is known as pattern-triggered immunity (PTI). Intracellular immune receptors are mainly a class of receptor proteins containing nucleotide-binding (NB) and nucleotide-binding leucine-rich repeat receptors (NLRs), which recognize cytoplasmic effector proteins to elicit effector-triggered immunity (ETI).
In addition to pathogens, some plant symbiotic microbes also possess microbe-associated molecular patterns (MAMPs) that can stimulate the innate immune response (MAMP-triggered immunity, MTI), which is similar to PTI [145]. When symbiotic microbes are present, plants will actively reduce PRR expression levels and MAMP responsiveness to allow symbiotic microbes to thrive and exert their beneficial effects on host plants [145,146]. For example, in the mycorrhizal symbiosis, the mycorrhizal symbiotic receptor, OsMYR1, and its ligand, CO4, will competitively bind OsCERK1, thus inhibiting the formation of immune receptor complexes between OsCERK1 and OsCEBiP, which weaken PAMP-mediated immune responses to promote its own symbiosis [147]. Therefore, the immune system may also be an alternative route to affect the plant microbiome.
6. The Microbiome Enhances Plant Immunity and Functions as an Extension of the Plant Immune System
Increasing evidence indicates that under pathogen stress, the composition of the plant microbiome changes leading the plant to actively recruit beneficial or protective microbes [148], to aid pathogen resistance and improve plant immunity [95,148,149,150]. Recently, it was demonstrated that infection of citrus trees by Candidatus Liberibacter asiaticus, associated with Huanglongbing (HLB), was shown to drastically alter the composition of citrus communities across the disease spectrum [126]. Moreover, specific compounds in tree root exudates stimulate key rhizosphere bacteria to cooperate in suppressing soil-borne pathogens [72]. The plant microbiome interacts with pathogens directly or indirectly to strengthen pathogen resistance (Figure 2).
6.1. Acting Directly against Pathogens: Direct Interaction between Microbes
Pathogen infected plants can recruit and enrich beneficial microbial communities to directly antagonize infecting pathogens via a variety of methods, such as the "cry for help" strategy. Specific mechanisms include competing with pathogens for ecological niches and nutrients, secretion of antagonistic compounds (such as antibiotics, lysozyme, and volatile substances), parasitism and predation, and inhibition of pathogen signaling systems, among others (Figure 2). For example, The Gram-negative bacterium, Pantoea agglomerans, inhibits the growth of Erwinia amylovora, which causes fire blight disease on apple and pear trees, through its antibiosis activity [151,152]. Bacillus sp. strains secrete lipopeptides (like Surfactin, Iturin, and Fengycin), polyketides, surfactants, and VOCs, among others, to directly kill or suppress pathogen growth, thereby improving the natural soil resistance against pathogens [153,154,155,156]. Some probiotic Bacillus bacteria can also eliminate pathogens by interfering with pathogen QS signaling [157]. Beneficial fungi can effectively inhibit the infection of destructive fungal pathogens such as Sclerotinia sclerotiorum, Fusarium oxysporum, and Rhizoctonia solani through mechanisms of parasitism [158,159,160]. Therefore, the plant microbiome offers a goldmine for antagonists that can directly inhibit pathogens.
6.2. Acting Indirectly against Pathogens: Stimulating or Inducing Plant Immunity
Plant microbiome can act indirectly by priming plant immunity (e.g., ISR) to inhibit pathogen invasion and protect the under- and above-ground parts from encroachment (Figure 2). For example, a disease-resistant synthetic bacterial community was constructed and found to protect plants through a synergistic effect of high-abundance bacteria inhibiting fungal pathogen growth and low-abundance bacteria activating plant ISRs [161]. Foliar infection by the downy mildew pathogen, Hyaloperonospora arabidopsidis, leads to the promotion of three bacterial species (Xanthomonas, Stenotrophomonas, and Microbacterium) in the rhizosphere of Arabidopsis plants, conferring a soil-mediated legacy that provides increased resistance against this pathogen [162]. A set of important studies have provided a wealth of data showing that infection with pathogens in the above- and below-ground plant tissues results in a change in the pattern of root secretions, thereby inducing the selective recruitment of beneficial root-associated microbes, forming soil memory or legacy effects of disease resistance to protect a subsequent population of plants growing in the same soil against the same pathogen [65,162,163]. In general, the recruitment of beneficial microbes in the plant rhizosphere during pathogen invasion is beneficial to better mobilize the role of the rhizosphere microbiome in the plant defense system, enhance the ability to control plant diseases, and even unlock new breeding strategies. However, further research is needed to understand the mechanisms by which plants recruit beneficial microbes to resist disease and how the microbiome initiates ISR.
7. Future Perspectives: Integrating Tree-Pathogen-Microbiome Interactions into Forest Pest Management
7.1. The Challenges of Applying BCAs in Protecting Trees
Compared to annual crops like wheat or potatoes, or herbaceous plants, tree generations can persist for decades. Therefore, strategies to control tree diseases need to be effective over many years, not just during a specific growing season [164]. In plantations, the long lifespan and high economic costs of trees do not always allow for regular switching of cultivated species (e.g., crop rotation). In addition to these limitations in the application of agronomic and cultural practices, the use of chemical treatments attracts growing concerns about environmental pollution and the adverse effects of chemicals on humans and animals. Thus, manipulation of the plant microbiome is considered to be a very promising and environmentally friendly approach to fostering plant protection against pests.
In recent years, the potential for BCAs has been realized as a key part of Integrated Pest Management (IPM). Current research builds on the development of omic approaches, synthetic communities, and microbial network inference to design beneficial microbial consortiums with the potential to provide broad and durable plant protection. Such strategies are also increasingly being adopted in planted forests, despite the fact that just a few instances of success are known. Importantly, adaptation to inner plant tissues and endurance within them for long periods of time make endophytes potentially promising biocontrol agents of long term woody plant diseases [165]. In fact, tree endophytes have been suggested as a valuable tool for sustaining forest health [166]. Effective and/or promising uses of endophytes (bacterial endophytes and fungal endophytes) to control diseases in trees are already in practice.
7.2. The Advantages in Tree Pathogen Inhibition by Multi-Strain BCAs over Single-strain bcas
in several cases of the management of soil-borne crop pathogens, different bca consortia, consisting of two or more microbial strains, appeared to possess enhanced biocontrol activity than single-strain bcas [167,168,169,170]. there is growing interest in the use of multi-strain bcas for controlling forest diseases. a consortium consisting of trichoderma virens gi006 and bacillus velezensis bs006 was found to be more efficient against infection by fusarium wilt of cape gooseberry than the individual strains [171]. a bacterial consortium of pseudomonas aeruginosa drb1 and trichoderma harzianum cbf2 conferred more consistent protection against fusarium wilt of banana than the individual community members [172]. the interaction among the members of multi-strain bcas might lead to their superior stability and efficiency of disease-suppressing effect. microbial interactions within the plant microbiome are essential selective forces that help to establish complex microbial assemblages [173]. consequently, when constructing multi-strain bcas, it is important to pay attention to the microbe−microbe interplay-related elements.
7.3. Microbial Interactions Promote Rhizosphere Colonization
A large number of microbe−microbe interactions have documented their positive role in promoting rhizosphere colonization by beneficial microorganisms through boosting biofilm formation, microbial growth, migration inside of the microbiome, and interacting with plant roots. For examples, a five-species biocontrol community was found to form more biofilm than the single strains either under in vitro or in vivo conditions, and thus stimulating the colonization of host roots [174]; Hyphomicrobium spp. is capable of associating with methanotrophs, establishing a rhizospheric microbial association where Hyphomicrobium spp. can remove methanol, which suppress the growth of methanotrophs in the rhizosphere [175]. The syntrophic interactions between multiple microorganisms can promote the growth of rhizosphere microbes by removal of harmful substances; fungal hyphae, also referred to as “fungal highway”, can act as vectors for the dispersion of bacteria in the rhizosphere [176,177], indicating that the migration of microorganisms can be enhanced by the interactions among community members. Thus, the application of multi-strain BCAs that actively interact among their members may increase survival of disease-suppressing microbes, and their adaption to challenging and shifting environmental conditions. As a result, they might be able to maintain their advantageous effects for inhibiting forest diseases.
7.4. Microbial Interactions Suppress Forest Pathogen Growth
Multi-strain BCAs are able to exhibit stronger suppressive efficacy on the growth of pathogens than single-strain BCAs due to the positive impact of interactions within multi-strain BCAs, such as the increased capability to utilize resources [178,179,180,181] and the biosynthesis of antimicrobial compounds [182,183] stimulated or activated through both inter- and intra-domain microbial interactions. contribute to strengthening the inhibition of growth of pathogens.
7.5. Microbial Interactions Induce Enhanced Plant Defense Responses to Pathogens
In addition to the above positive features in controlling diseases by multi-strain BCAs, inducing elevated host immunities to plant pathogens through the biocontrol consortia has been described in many studies. This is mainly directed by activating a number of distinctive metabolic and signaling pathways in order to combat a given disease [168,184,185]. However, it still needs to be better understood how interactions among the multi-strain BCA members might successfully improve specific systemic resistance to pathogens. One possibility is that the interaction between microbes inside the biocontrol consortia may result in the generation of more of certain elicitors and potent compounds that can more efficiently elicit ISR, as already discussed in part 5.
7.6. Future Perspectives of Applying BCAs in Protecting Trees
Because of the inherent characteristics of trees (e.g., large biomass, complicated anatomy, longevity, and perennial nature) and pathogens (e.g., complex life cycles, complex infection), biological control in trees and woody plants presents challenges, difficulties, and limitations. For instance, a tree can be affected by the disease in successive growing seasons, not just during a specific growing season, which can impact the effectiveness of biocontrol strategies. As a result, the utilization of BCAs as a disease control strategy in trees and woody plants has been implemented to a lesser extent than in annual crops and herbaceous species. Despite this, there are many cases where biocontrol applications have been successfully used to enhance forest plantations health and even the emergence of commercial biocontrol products. One example includes Dutch Trig®, which has been used for around 30 years to protect elm trees from infection by the blue stain fungus O. novo-ulmi, which causes Dutch elm disease (DED) [186]. In order to enhance the effectiveness in applying BCAs in mitigating emerging forest diseases, more research is needed to unravel the dynamics of such diseases, how they interact with BCA measures, and what impact they may have on the ecology of a forested environment. Moreover, biological control with BCAs must be combined with other measures (e.g., cultural practices, sanitary cuttings, tree Breeding for disease Resistance, increase tree species diversity) in an integrated disease management approach.
Author Contributions
Conceptualization, Q.X. and X.L.; writing-original draft preparation, Q.X.; writing, review and editing, Q.X., J.Y., S.N., H.L., and X.L.; Funding acquisition, Q.X. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2022YFD2201900, 2021YDF22011202), the National Natural Science Foundation of China (31600512, 32122056, 42011045), the Jiangsu Agriculture Science and Technology Independent Innovation Fund (CX-21-3045), the China Postdoctoral Science Foundation (2021M691605), Postdoctoral Science Foundation of Jiangsu Province (2021K641C), Postgraduate Research & Practice Innovation Program of Jiangsu Province (SJCX23_0350); and Students Practice Innovation and Training Program of Nanjing Forestry University under Grant nos. 2022NFUSPITP0364 and 202310298135Y.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available in individual cited studies in this review.
Acknowledgments
Past colleagues are acknowledged for the insights into “Microbiome-mediated protection against pathogens in woody plants” they have provided through a number of fruitful collaborations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bettenfeld, P.; Fontaine, F.; Trouvelot, S.; Fernandez, O.; Courty, P.-E. Woody plant declines. what’s wrong with the microbiome? Trends Plant Sci. 2020, 25, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Vettraino, A.; Morel, O.; Perlerou, C.; Robin, C.; Diamandis, S.; Vannini, A. Occurrence and distribution of Phytophthora species in European chestnut stands, and their association with Ink Disease and crown decline. Eur. J. Plant Pathol. 2005, 111, 169–180. [Google Scholar] [CrossRef]
- Rigling, D.; Prospero, S. Cryphonectria parasitica, the causal agent of chestnut blight: Invasion history, population biology and disease control. Mol. Plant Pathol. 2018, 19, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M. Rapid evolution of introduced plant pathogens via interspecific hybridization: Hybridization is leading to rapid evolution of Dutch elm disease and other fungal plant pathogens. Bioscience 2001, 51, 123–133. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Garbelotto, M.; Goss, E.M.; Heungens, K.; Prospero, S. Emergence of the sudden oak death pathogen Phytophthora Ramorum. Trends Microbiol. 2012, 20, 131–138. [Google Scholar] [CrossRef]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef]
- Harrington, T.C.; Yun, H.Y.; Lu, S.-S.; Goto, H.; Aghayeva, D.N.; Fraedrich, S.W. Isolations from the redbay ambrosia beetle, Xyleborus glabratus, confirm that the laurel wilt pathogen, Raffaelea lauricola, originated in Asia. Mycologia 2011, 103, 1028–1036. [Google Scholar] [CrossRef]
- Sturrock, R.; Frankel, S.; Brown, A.; Hennon, P.; Kliejunas, J.; Lewis, K.; Worrall, J.; Woods, A. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Prospero, S.; Cleary, M. Effects of host variability on the spread of invasive forest diseases. Forests 2017, 8, 80. [Google Scholar] [CrossRef]
- Holdenrieder, O.; Pautasso, M.; Weisberg, P.J.; Lonsdale, D. Tree diseases and landscape processes: The challenge of landscape pathology. Trends Ecol. Evol. 2004, 19, 446–452. [Google Scholar] [CrossRef]
- Yang, L.-N.; Ren, M.; Zhan, J. Modeling plant diseases under climate change: Evolutionary perspectives. Trends Plant Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jia, H. Metagenome-wide association studies: Fine-mining the microbiome. Nat. Rev. Microbiol. 2016, 14, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Poudel, R.; Jumpponen, A.; Schlatter, D.C.; Paulitz, T.; Gardener, B.M.; Kinkel, L.L.; Garrett, K. Microbiome networks: A systems framework for identifying candidate microbial assemblages for disease management. Phytopathology 2016, 106, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, M.G.; Hartmann, M. Networking in the plant microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Prospero, S.; Botella, L.; Santini, A.; Robin, C. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]
- Cai, S.; Jia, J.; He, C.; Zeng, L.; Fang, Y.; Qiu, G.; Lan, X.; Su, J.; He, X. Multi-omics of pine wood nematode pathogenicity associated with culturable associated microbiota through an artificial assembly approach. Front. Plant Sci. 2022, 12. [Google Scholar] [CrossRef]
- Corinne, V.; Bastien, C.; Emmanuelle, J.; Heidy, S. Trees and Insects Have Microbiomes: Consequences for Forest Health and Management. Curr. For. Rep. 2021, 7, 81–96. [Google Scholar] [CrossRef]
- Asiegbu, F.O. Chapter 22 - Forest microbiome: Challenges and future perspectives. In Forest Microbiology; Asiegbu, F.O., Kovalchuk, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 395–399. [Google Scholar]
- Burke, D.J.; Hoke, A.J.; Koch, J. The emergence of beech leaf disease in Ohio: Probing the plant microbiome in search of the cause. Forest Pathol. 2020, 50, e12579. [Google Scholar] [CrossRef]
- Li, F.; Zi, H.; Sonne, C.; Li, X. Microbiome sustains forest ecosystem functions across hierarchical scales. Eco-Environ. Health 2023, 2, 24–31. [Google Scholar] [CrossRef]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2016, 41, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Asiegbu, F.O.; Kovalchuk, A. Forest Microbiology: Volume 1: Tree Microbiome: Phyllosphere, Endosphere and Rhizosphere; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Kuzyakov, Y. Factors affecting rhizosphere priming effects. J. Plant Nutr. Soil. Sci. 2002, 165, 382–396. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil. 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Hansen, E.M. Phytophthora species emerging as pathogens of forest trees. Curr. For. Rep. 2015, 1, 16–24. [Google Scholar] [CrossRef]
- Coutinho, T.A.; Wingfield, M.J. Ralstonia solanacearum and R. pseudosolanacearum on Eucalyptus: Opportunists or Primary Pathogens? Front. Plant Sci. 2017, 8, 761. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, B.; Liu, P.; Liao, F.; Weng, Q.; Chen, Q. First report of bacterial wilt caused by Ralstonia solanacearum on fig trees in China. For. Pathol. 2016, 46, 256–258. [Google Scholar] [CrossRef]
- Roux, J.; Wingfield, M. Ceratocystis species: Emerging pathogens of non-native plantation Eucalyptus and Acacia species. South. For. A J. For. Sci. 2009, 71, 115–120. [Google Scholar] [CrossRef]
- Barnes, I.; Roux, J.; Wingfield, B.; O’Neill, M.; Wingfield, M. Ceratocystis fimbriata infecting Eucalyptus grandis in Uruguay. Australas Plant Pathol. 2003, 32, 361–366. [Google Scholar] [CrossRef]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Paplomatas, E.J.; Tjamos, E.C. Biological control of Verticillium wilt of olive by Paenibacillus alvei, strain K165. BioControl 2016, 61, 293–303. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Tian, C. Quantitative detection of pathogen DNA of Verticillium wilt on smoke tree Cotinus coggygria. Plant Dis. 2013, 97, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.; Hiemstra, J. A Compendium of Verticillium Wilts in Tree Species; Ponsen and Looijen: Wageningen, The Netherlands, 1998. [Google Scholar]
- Hrynkiewicz, K.; Baum, C. The Potential of Rhizosphere Microorganisms to Promote the Plant Growth in Disturbed Soils; Springer: Berlin/Heidelberg, Germany, 2012; pp. 35–64. [Google Scholar]
- Al-Karaki, G.N. The Role of Mycorrhiza in the Reclamation of Degraded Lands in Arid Environments; Springer: Berlin/Heidelberg, Germany, 2013; pp. 823–836. [Google Scholar]
- Khabou, W.; Hajji, B.; Zouari, M.; Rigane, H.; Abdallah, F.B. Arbuscular mycorrhizal fungi improve growth and mineral uptake of olive tree under gypsum substrate. Ecol. Eng. 2014, 73, 290–296. [Google Scholar] [CrossRef]
- Manaut, N.; Sanguin, H.; Ouahmane, L.; Bressan, M.; Thioulouse, J.; Baudoin, E.; Galiana, A.; Hafidi, M.; Prin, Y.; Duponnois, R. Potentialities of ecological engineering strategy based on native arbuscular mycorrhizal community for improving afforestation programs with carob trees in degraded environments. Ecol. Eng. 2015, 79, 113–119. [Google Scholar] [CrossRef]
- Kerr, A. Biological control of crown gall. Australas. Plant Pathol. 2016, 45, 15–18. [Google Scholar] [CrossRef]
- Prieto, P.; Navarro-Raya, C.; Valverde-Corredor, A.; Amyotte, S.G.; Dobinson, K.F.; Mercado-Blanco, J. Colonization process of olive tissues by Verticillium dahliae and its in planta interaction with the biocontrol root endophyte Pseudomonas fluorescens PICF7. Microb. Biotechnol. 2009, 2, 499–511. [Google Scholar] [CrossRef]
- Martínez-García, P.M.; Ruano-Rosa, D.; Schilirò, E.; Prieto, P.; Ramos, C.; Rodríguez-Palenzuela, P.; Mercado-Blanco, J. Complete genome sequence of Pseudomonas fluorescens strain PICF7, an indigenous root endophyte from olive (Olea europaea L.) and effective biocontrol agent against Verticillium dahliae. Stand. Genomic Sci. 2015, 10, 1–7. [Google Scholar] [CrossRef]
- Wook eYang, J. ISR meets SAR outside: Additive action of the endophyte Bacillus pumilus INR7 and the chemical inducer, benzothiadiazole, on induced resistance against bacterial spot in field-grown pepper. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Robert, C.A.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; Van Der Heijden, M.G. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Baetz, U.; Martinoia, E. Root exudates: The hidden part of plant defense. Trends Plant Sci. 2014, 19, 90–98. [Google Scholar] [CrossRef]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Glavina del Rio, T.; Jones, C.D.; Tringe, S.G. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of different plant root exudates and their organic acid components on chemotaxis, biofilm formation and colonization by beneficial rhizosphere-associated bacterial strains. Plant Soil. 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Tian, B.; Pei, Y.; Huang, W.; Ding, J.; Siemann, E. Increasing flavonoid concentrations in root exudates enhance associations between arbuscular mycorrhizal fungi and an invasive plant. ISME J. 2021, 15, 1919–1930. [Google Scholar] [CrossRef]
- Bai, B.; Liu, W.; Qiu, X.; Zhang, J.; Zhang, J.; Bai, Y. The root microbiome: Community assembly and its contributions to plant fitness. J. Integr. Plant Biol. 2022, 64, 230–243. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; Mendes, R.; Raaijmakers, J.M. Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant Mol. Biol. 2016, 90, 635–644. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef]
- Warmink, J.; Nazir, R.; Corten, B.; Van Elsas, J. Hitchhikers on the fungal highway: The helper effect for bacterial migration via fungal hyphae. Soil. Biol. Biochem. 2011, 43, 760–765. [Google Scholar] [CrossRef]
- Kohlmeier, S.; Smits, T.H.; Ford, R.M.; Keel, C.; Harms, H.; Wick, L.Y. Taking the fungal highway: Mobilization of pollutant-degrading bacteria by fungi. Environ. Sci. Technol. 2005, 39, 4640–4646. [Google Scholar] [CrossRef]
- Barriuso, J.; Ramos Solano, B.; Fray, R.G.; Cámara, M.; Hartmann, A.; Gutiérrez Mañero, F.J. Transgenic tomato plants alter quorum sensing in plant growth-promoting rhizobacteria. Plant Biotechnol. J. 2008, 6, 442–452. [Google Scholar] [CrossRef]
- Schenk, S.T.; Schikora, A. AHL-priming functions via oxylipin and salicylic acid. Front. Plant Sci. 2015, 5, 784. [Google Scholar] [CrossRef]
- Bitas, V.; Kim, H.-S.; Bennett, J.W.; Kang, S. Sniffing on microbes: Diverse roles of microbial volatile organic compounds in plant health. Mol. Plant Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef]
- Schmidt, R.; Cordovez, V.; De Boer, W.; Raaijmakers, J.; Garbeva, P. Volatile affairs in microbial interactions. ISME J. 2015, 9, 2329–2335. [Google Scholar] [CrossRef]
- Audrain, B.; Farag, M.A.; Ryu, C.-M.; Ghigo, J.-M. Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 2015, 39, 222–233. [Google Scholar] [CrossRef]
- Cosetta, C.M.; Wolfe, B.E. Causes and consequences of biotic interactions within microbiomes. Curr. Opin. Microbiol. 2019, 50, 35–41. [Google Scholar] [CrossRef]
- Li, P.; Zhu, Z.; Zhang, Y.; Xu, J.; Wang, H.; Wang, Z.; Li, H. The phyllosphere microbiome shifts toward combating melanose pathogen. Microbiome 2022, 10, 1–17. [Google Scholar] [CrossRef]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; De Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.; Gomez-Exposito, R.; Elsayed, S.S. Pathogen-induced activation of disease-suppressive functions in the endophytic root microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Kokalis-Burelle, N.; McSorley, R.; Wang, K.-H.; Saha, S.K.; McGovern, R.J. Rhizosphere microorganisms affected by soil solarization and cover cropping in Capsicum annuum and Phaseolus lunatus agroecosystems. Appl. Soil. Ecol. 2017, 119, 64–71. [Google Scholar] [CrossRef]
- Smalla, K.; Wieland, G.; Buchner, A.; Zock, A.; Parzy, J.; Kaiser, S.; Roskot, N.; Heuer, H.; Berg, G. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: Plant-dependent enrichment and seasonal shifts revealed. Appl. Environ. Microbiol. 2001, 67, 4742–4751. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Blanco, J.; Abrantes, I.; Barra Caracciolo, A.; Bevivino, A.; Ciancio, A.; Grenni, P.; Hrynkiewicz, K.; Kredics, L.; Proença, D.N. Belowground microbiota and the health of tree crops. Front. Microbiol. 2018, 9, 1006. [Google Scholar] [CrossRef]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Yu, L.; Zi, H.; Zhu, H.; Liao, Y.; Li, X. Rhizosphere microbiome of forest trees determines their resistance to soil-borne pathogens. Reserch Square. 2022. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef]
- Stone, B.W.; Weingarten, E.A.; Jackson, C.R. The role of the phyllosphere microbiome in plant health and function. Annu. Plant Rev. Online 2018, 533–556. [Google Scholar] [CrossRef]
- Sapp, M.; Ploch, S.; Fiore-Donno, A.M.; Bonkowski, M.; Rose, L.E. Protists are an integral part of the Arabidopsis thaliana microbiome. Environ. Microbiol. 2018, 20, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Balogh, B.; Nga, N.T.T.; Jones, J.B. Relative level of bacteriophage multiplication in vitro or in phyllosphere may not predict in planta efficacy for controlling bacterial leaf spot on tomato caused by Xanthomonas perforans. Front. Microbiol. 2018, 9, 2176. [Google Scholar] [CrossRef]
- Ritpitakphong, U.; Falquet, L.; Vimoltust, A.; Berger, A.; Métraux, J.P.; L'Haridon, F. The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol. 2016, 210, 1033–1043. [Google Scholar] [CrossRef]
- Xiong, C.; Zhu, Y.G.; Wang, J.T.; Singh, B.; Han, L.L.; Shen, J.P.; Li, P.P.; Wang, G.B.; Wu, C.F.; Ge, A.H. Host selection shapes crop microbiome assembly and network complexity. New Phytol. 2021, 229, 1091–1104. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Horton, M.W.; Bergelson, J. Bacterial communities associated with the leaves and the roots of Arabidopsis thaliana. PLoS ONE. 2013, 8, e56329. [Google Scholar] [CrossRef] [PubMed]
- Maignien, L.; DeForce, E.A.; Chafee, M.E.; Eren, A.M.; Simmons, S.L. Ecological succession and stochastic variation in the assembly of Arabidopsis thaliana phyllosphere communities. mBio 2014, 5, e00682–00613. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Müller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Münch, P.C.; Spaepen, S.; Remus-Emsermann, M. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef]
- Müller, T.; Ruppel, S. Progress in cultivation-independent phyllosphere microbiology. FEMS Microbiol. Ecol. 2014, 87, 2–17. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Delmotte, N.; Knief, C.; Chaffron, S.; Innerebner, G.; Roschitzki, B.; Schlapbach, R.; von Mering, C.; Vorholt, J.A. Community proteogenomics reveals insights into the physiology of phyllosphere bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 16428–16433. [Google Scholar] [CrossRef]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; Von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Izhaki, I.; Fridman, S.; Gerchman, Y.; Halpern, M. Variability of bacterial community composition on leaves between and within plant species. Curr. Microbiol. 2013, 66, 227–235. [Google Scholar] [CrossRef]
- Inácio, J.; Pereira, P.; Carvalho, d.M.; Fonseca, A.; Amaral-Collaco, M.; Spencer-Martins, I. Estimation and diversity of phylloplane mycobiota on selected plants in a mediterranean–type ecosystem in Portugal. Microb. Ecol. 2002, 44, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef]
- Griffiths, S.M.; Galambao, M.; Rowntree, J.; Goodhead, I.; Hall, J.; O’Brien, D.; Atkinson, N.; Antwis, R.E. Complex associations between cross-kingdom microbial endophytes and host genotype in ash dieback disease dynamics. J. Ecol. 2020, 108, 291–309. [Google Scholar] [CrossRef]
- Ulrich, K.; Becker, R.; Behrendt, U.; Kube, M.; Ulrich, A. A comparative analysis of ash leaf-colonizing bacterial communities identifies putative antagonists of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 966. [Google Scholar] [CrossRef]
- Durand, A.; Maillard, F.; Alvarez-Lopez, V.; Guinchard, S.; Bertheau, C.; Valot, B.; Blaudez, D.; Chalot, M. Bacterial diversity associated with poplar trees grown on a Hg-contaminated site: Community characterization and isolation of Hg-resistant plant growth-promoting bacteria. Sci. Total Environ. 2018, 622, 1165–1177. [Google Scholar] [CrossRef]
- Redford, A.J.; Bowers, R.M.; Knight, R.; Linhart, Y.; Fierer, N. The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ. Microbiol. 2010, 12, 2885–2893. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S. The soil microbiome influences grapevine-associated microbiota. mBio 2015, 6, e02527–e02514. [Google Scholar] [CrossRef]
- Copeland, J.K.; Yuan, L.; Layeghifard, M.; Wang, P.W.; Guttman, D.S. Seasonal community succession of the phyllosphere microbiome. Mol. Plant Microbe Interact. 2015, 28, 274–285. [Google Scholar] [CrossRef]
- Finkel, O.M.; Burch, A.Y.; Lindow, S.E.; Post, A.F.; Belkin, S. Geographical location determines the population structure in phyllosphere microbial communities of a salt-excreting desert tree. Appl. Environ. Microbiol. 2011, 77, 7647–7655. [Google Scholar] [CrossRef] [PubMed]
- Finkel, O.M.; Burch, A.Y.; Elad, T.; Huse, S.M.; Lindow, S.E.; Post, A.F.; Belkin, S. Distance-decay relationships partially determine diversity patterns of phyllosphere bacteria on Tamrix trees across the Sonoran Desert. Appl. Environ. Microbiol. 2012, 78, 6187–6193. [Google Scholar] [CrossRef] [PubMed]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Host species identity, site and time drive temperate tree phyllosphere bacterial community structure. Microbiome 2016, 4, 1–10. [Google Scholar] [CrossRef]
- Cordier, T.; Robin, C.; Capdevielle, X.; Desprez-Loustau, M.-L.; Vacher, C. Spatial variability of phyllosphere fungal assemblages: Genetic distance predominates over geographic distance in a European beech stand (Fagus sylvatica). Fungal Ecol. 2012, 5, 509–520. [Google Scholar] [CrossRef]
- Arnold, A.E.; Maynard, Z.; Gilbert, G.S.; Coley, P.D.; Kursar, T.A. Are tropical fungal endophytes hyperdiverse? Ecol. Lett. 2000, 3, 267–274. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Sheikh, I.; Dhiman, A.; Yadav, N.; Yadav, A.N.; Rastegari, A.A.; Singh, K.; Saxena, A.K. Endophytic fungi: Biodiversity, ecological significance, and potential industrial applications. In Recent Advancement in White Biotechnology through Fungi; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–62. [Google Scholar]
- Perreault, R.; Laforest-Lapointe, I. Plant-microbe interactions in the phyllosphere: Facing challenges of the anthropocene. ISME J. 2022, 16, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Xiong, C.; Wei, Z.; Chen, Q.L.; Ma, B.; Zhou, S.Y.D.; Tan, J.; Zhang, L.M.; Cui, H.L.; Duan, G.L. Impacts of global change on the phyllosphere microbiome. New Phytol. 2022. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K. Massively parallel 454 sequencing indicates hyperdiverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol. 2009, 184, 438–448. [Google Scholar] [CrossRef]
- Balint, M.; Bartha, L.; O'Hara, R.B.; Olson, M.S.; Otte, J.; Pfenninger, M.; Robertson, A.L.; Tiffin, P.; Schmitt, I. Relocation, high-latitude warming and host genetic identity shape the foliar fungal microbiome of poplars. Mol. Ecol. 2015, 24, 235–248. [Google Scholar] [CrossRef]
- Balint, M.; Tiffin, P.; Hallstroem, B.; O'Hara, R.B.; Olson, M.S.; Fankhauser, J.D.; Piepenbring, M.; Schmitt, I. Host genotype shapes the foliar fungal microbiome of balsam poplar (Populus balsamifera). PLoS ONE. 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Herrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F. Genomic features of bacterial adaptation to plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Junker, R.R.; Tholl, D. Volatile organic compound mediated interactions at the plant-microbe interface. J. Chem. Ecol. 2013, 39, 810–825. [Google Scholar] [CrossRef]
- Cosoveanu, A.; Gimenez-Mariño, C.; Cabrera, Y.; Hernandez, G.; Cabrera, R. Endophytic fungi from grapevine cultivars in Canary Islands and their activity against phytopatogenic fungi. Int. J. Agric. Crop Sci. 2014, 7, 1497. [Google Scholar]
- Balint-Kurti, P.; Simmons, S.J.; Blum, J.E.; Ballaré, C.L.; Stapleton, A.E. Maize leaf epiphytic bacteria diversity patterns are genetically correlated with resistance to fungal pathogen infection. Mol. Plant Microbe Interact. 2010, 23, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Ali, B.; Sabri, A.; Ljung, K.; Hasnain, S. Auxin production by plant associated bacteria: Impact on endogenous IAA content and growth of Triticum aestivum L. Lett. Appl. Microbiol. 2009, 48, 542–547. [Google Scholar] [CrossRef]
- Burch, A.Y.; Zeisler, V.; Yokota, K.; Schreiber, L.; Lindow, S.E. The hygroscopic biosurfactant syringafactin produced by Pseudomonas syringae enhances fitness on leaf surfaces during fluctuating humidity. Environ. Microbiol. 2014, 16, 2086–2098. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Chen, T.; Nomura, K.; Wang, X.; Sohrabi, R.; Xu, J.; Yao, L.; Paasch, B.C.; Ma, L.; Kremer, J.; Cheng, Y. A plant genetic network for preventing dysbiosis in the phyllosphere. Nature 2020, 580, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.; Pieterse, C.M.; de Jonge, R.; Berendsen, R.L. The soil-borne legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, J.; Carvalhais, L.C.; Percy, C.D.; Prakash Verma, J.; Schenk, P.M.; Singh, B.K. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytol. 2021, 229, 2873–2885. [Google Scholar] [CrossRef]
- Cordovez, V.; Dini-Andreote, F.; Carrión, V.J.; Raaijmakers, J.M. Ecology and evolution of plant microbiomes. Annu. Rev. Microbiol. 2019, 73, 69–88. [Google Scholar] [CrossRef]
- Getzke, F.; Thiergart, T.; Hacquard, S. Contribution of bacterial-fungal balance to plant and animal health. Curr. Opin. Microbiol. 2019, 49, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Casa Vargas, J.M.; Schlatter, D.C.; Hagerty, C.H.; Hulbert, S.H.; Paulitz, T.C. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome 2021, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.-J.; Kong, H.G.; Choi, K.; Kwon, S.-K.; Song, J.Y.; Lee, J.; Lee, P.A.; Choi, S.Y.; Seo, M.; Lee, H.J. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat. Biotechnol. 2018, 36, 1100–1109. [Google Scholar] [CrossRef]
- Berg, M.; Koskella, B. Nutrient-and dose-dependent microbiome-mediated protection against a plant pathogen. Curr. Biol. 2018, 28, 2487–2492. [Google Scholar] [CrossRef]
- Jakuschkin, B.; Fievet, V.; Schwaller, L.; Fort, T.; Robin, C.; Vacher, C. Deciphering the pathobiome: Intra-and interkingdom interactions involving the pathogen Erysiphe alphitoides. Microb. Ecol. 2016, 72, 870–880. [Google Scholar] [CrossRef]
- Ginnan, N.A.; Dang, T.; Bodaghi, S.; Ruegger, P.M.; McCollum, G.; England, G.; Vidalakis, G.; Borneman, J.; Rolshausen, P.E.; Roper, M.C. Disease-induced microbial shifts in citrus indicate microbiome-derived responses to huanglongbing across the disease severity spectrum. Phytobiomes J. 2020, 4, 375–387. [Google Scholar] [CrossRef]
- Li, P.-D.; Zhu, Z.-R.; Zhang, Y.; Xu, J.; Wang, H.; Wang, Z.; Li, H. The phyllosphere microbiome shifts toward combating melanose pathogen. Microbiome 2022, 10, 1–17. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Vescio, K.; Mitter, B.; Trognitz, F.; Ma, L.-J.; Sessitsch, A. Ecology and genomic insights into plant-pathogenic and plant-nonpathogenic endophytes. Annu. Rev. Phytopathol. 2017, 55. [Google Scholar] [CrossRef]
- Baiyee, B.; Pornsuriya, C.; Ito, S.-i.; Sunpapao, A. Trichoderma spirale T76-1 displays biocontrol activity against leaf spot on lettuce (Lactuca sativa L.) caused by Corynespora cassiicola or Curvularia aeria. Biol. Control. 2019, 129, 195–200. [Google Scholar] [CrossRef]
- Loc, N.H.; Huy, N.D.; Quang, H.T.; Lan, T.T.; Thu Ha, T.T. Characterisation and antifungal activity of extracellular chitinase from a biocontrol fungus, Trichoderma asperellum PQ34. Mycology 2020, 11, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yassin, M.T.; Mostafa, A.A.-F.; Al-Askar, A.A.; Sayed, S.R.; Rady, A.M. Antagonistic activity of Trichoderma harzianum and Trichoderma viride strains against some fusarial pathogens causing stalk rot disease of maize, in vitro. Egypt. J. Biol. Pest. Co. 2021, 33, 101363. [Google Scholar] [CrossRef]
- Yan, L.; Khan, R.A.A. Biological control of bacterial wilt in tomato through the metabolites produced by the biocontrol fungus, Trichoderma harzianum. Egypt. J. Biol. Pest. Co. 2021, 31, 1–9. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Nita, M.; Ishii, T.; Watanabe, M.; Noutoshi, Y. Biological control agent Rhizobium (= Agrobacterium) vitis strain ARK-1 suppresses expression of the essential and non-essential vir genes of tumorigenic R. vitis. BMC Res. Notes. 2019, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tao, Y.; Tao, X.; Su, Q.; Cai, J.; Qin, C.; Ding, W.; Li, C. Sesquiterpenes with phytopathogenic fungi inhibitory activities from fungus Trichoderma virens from Litchi chinensis Sonn. J. Agric. Food Chem. 2019, 67, 10646–10652. [Google Scholar] [CrossRef]
- Mengistu, A.A. Endophytes: Colonization, behaviour, and their role in defense mechanism. Int. J. Microbiol. 2020, 2020. [Google Scholar] [CrossRef]
- Mejía, L.C.; Herre, E.A.; Sparks, J.P.; Winter, K.; García, M.N.; Van Bael, S.A.; Stitt, J.; Shi, Z.; Zhang, Y.; Guiltinan, M.J. Pervasive effects of a dominant foliar endophytic fungus on host genetic and phenotypic expression in a tropical tree. Front. Microbiol. 2014, 5, 479. [Google Scholar] [CrossRef]
- Li, Y.; Duan, T.; Nan, Z.; Li, Y. Arbuscular mycorrhizal fungus alleviates alfalfa leaf spots caused by Phoma medicaginis revealed by RNA-seq analysis. J. Appl. Microbiol. 2021, 130, 547–560. [Google Scholar] [CrossRef]
- Busby, P.E.; Peay, K.G.; Newcombe, G. Common foliar fungi of Populus trichocarpa modify Melampsora rust disease severity. New Phytol. 2016, 209, 1681–1692. [Google Scholar] [CrossRef]
- Ren, J.H.; Ye, J.R.; Liu, H.; Xu, X.L.; Wu, X.Q. Isolation and characterization of a new Burkholderia pyrrocinia strain JK-SH007 as a potential biocontrol agent. World J. Microbiol. Biotechnol. 2011, 27, 2203–2215. [Google Scholar] [CrossRef]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef]
- Sumarah, M.W.; Kesting, J.R.; Sørensen, D.; Miller, J.D. Antifungal metabolites from fungal endophytes of Pinus strobus. Phytochemistry 2011, 72, 1833–1837. [Google Scholar] [CrossRef]
- Sumarah, M.W.; Walker, A.K.; Seifert, K.A.; Todorov, A.; Miller, J.D. Screening of fungal endophytes isolated from eastern white pine needles. In The Formation, Structure and Activity of Phytochemicals; Springer: Berlin/Heidelberg, Germany, 2015; pp. 195–206. [Google Scholar]
- Segaran, G.; Sathiavelu, M. Fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101284. [Google Scholar] [CrossRef]
- Zhou, F.; Emonet, A.; Dénervaud Tendon, V.; Marhavy, P.; Wu, D.; Lahaye, T.; Geldner, N. Co-incidence of damage and microbial patterns controls localized immune responses in roots. Cell 2020, 180, 440–453. [Google Scholar] [CrossRef]
- Hacquard, S.; Spaepen, S.; Garrido-Oter, R.; Schulze-Lefert, P. Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 2017, 55, 565–589. [Google Scholar] [CrossRef]
- Zhang, C.; He, J.; Dai, H.; Wang, G.; Zhang, X.; Wang, C.; Shi, J.; Chen, X.; Wang, D.; Wang, E. Discriminating symbiosis and immunity signals by receptor competition in rice. Proc. Natl. Acad. Sci. USA 2021, 118, e2023738118. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Bakker, P.A.; Berendsen, R.L.; Van Pelt, J.A.; Vismans, G.; Yu, K.; Li, E.; Van Bentum, S.; Poppeliers, S.W.; Gil, J.J.S.; Zhang, H. The soil-borne identity and microbiome-assisted agriculture: Looking back to the future. Mol. Plant. 2020, 13, 1394–1401. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.; Poinssot, B.; Pozo, M.J. Priming: Getting ready for battle. Mol. Plant Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef]
- Stockwell, V.; Johnson, K.; Sugar, D.; Loper, J. Antibiosis contributes to biological control of fire blight by Pantoea agglomerans strain Eh252 in orchards. Phytopathology 2002, 92, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Pusey, P.; Stockwell, V.; Reardon, C.; Smits, T.; Duffy, B. Antibiosis activity of Pantoea agglomerans biocontrol strain E325 against Erwinia amylovora on apple flower stigmas. Phytopathology 2011, 101, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shao, J.; Li, B.; Yan, X.; Shen, Q.; Zhang, R. Contribution of bacillomycin D in Bacillus amyloliquefaciens SQR9 to antifungal activity and biofilm formation. Appl. Environ. Microbiol. 2013, 79, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, R.; Wang, D.; Qiu, M.; Feng, H.; Zhang, N.; Shen, Q. Enhanced control of cucumber wilt disease by Bacillus amyloliquefaciens SQR9 by altering the regulation of its DegU phosphorylation. Appl. Environ. Microbiol. 2014, 80, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yuan, J.; Zhang, J.; Shen, Z.; Zhang, M.; Li, R.; Ruan, Y.; Shen, Q. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils. 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Yuan, J.; Raza, W.; Shen, Q.; Huang, Q. Antifungal activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp. cubense. Appl. Environ. Microbiol. 2012, 78, 5942–5944. [Google Scholar] [CrossRef]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.-S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Tian, B.; Xie, J.; Fu, Y.; Cheng, J.; Li, B.; Chen, T.; Zhao, Y.; Gao, Z.; Yang, P.; Barbetti, M.J. A cosmopolitan fungal pathogen of dicots adopts an endophytic lifestyle on cereal crops and protects them from major fungal diseases. ISME J. 2020, 14, 3120–3135. [Google Scholar] [CrossRef]
- Zhang, J.; Miao, Y.; Rahimi, M.J.; Zhu, H.; Steindorff, A.; Schiessler, S.; Cai, F.; Pang, G.; Chenthamara, K.; Xu, Y. Guttation capsules containing hydrogen peroxide: An evolutionarily conserved NADPH oxidase gains a role in wars between related fungi. Environ. Microbiol. 2019, 21, 2644–2658. [Google Scholar] [CrossRef]
- Zhang, J.; Bayram Akcapinar, G.; Atanasova, L.; Rahimi, M.J.; Przylucka, A.; Yang, D.; Kubicek, C.P.; Zhang, R.; Shen, Q.; Druzhinina, I.S. The neutral metallopeptidase NMP1 of Trichoderma guizhouense is required for mycotrophy and self-defence. Environ. Microbiol. 2016, 18, 580–597. [Google Scholar] [CrossRef]
- Li, Z.; Bai, X.; Jiao, S.; Li, Y.; Li, P.; Yang, Y.; Zhang, H.; Wei, G. A simplified synthetic community rescues Astragalus mongholicus from root rot disease by activating plant-induced systemic resistance. Microbiome 2021, 9, 217. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; De Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Cox, C.; Garrett, K.; Bockus, W. Meeting the challenge of disease management in perennial grain cropping systems. Renew. Agric. Food Syst. 2005, 20, 15–24. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; JJ Lugtenberg, B. Biotechnological applications of bacterial endophytes. Curr. Biotechnol. 2014, 3, 60–75. [Google Scholar] [CrossRef]
- Pautasso, M.; Schlegel, M.; Holdenrieder, O. Forest health in a changing world. Microb. Ecol. 2015, 69, 826–842. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, M.; Freilich, S. Prospects for biological soilborne disease control: Application of indigenous versus synthetic microbiomes. Phytopathology 2017, 107, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Sarma, B.K.; Yadav, S.K.; Singh, S.; Singh, H.B. Microbial consortium-mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil. Biol. Biochem. 2015, 87, 25–33. [Google Scholar] [CrossRef]
- Vorholt, J.A.; Vogel, C.; Carlström, C.I.; Müller, D.B. Establishing causality: Opportunities of synthetic communities for plant microbiome research. Cell Host Microbe. 2017, 22, 142–155. [Google Scholar] [CrossRef]
- Woo, S.L.; Pepe, O. Microbial consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-García, L.F.; González-Almario, A.; Cotes, A.M.; Moreno-Velandia, C.A. Trichoderma virens Gl006 and Bacillus velezensis Bs006: A compatible interaction controlling Fusarium wilt of cape gooseberry. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.F.; Saidi, N.B.; Vadamalai, G.; Teh, C.Y.; Zulperi, D. Effect of bioformulations on the biocontrol efficacy, microbial viability and storage stability of a consortium of biocontrol agents against Fusarium wilt of banana. J. Appl. Microbiol. 2019, 127, 544–555. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Santhanam, R.; Menezes, R.C.; Grabe, V.; Li, D.; Baldwin, I.T.; Groten, K. A suite of complementary biocontrol traits allows a native consortium of root-associated bacteria to protect their host plant from a fungal sudden-wilt disease. Mol. Ecol. 2019, 28, 1154–1169. [Google Scholar] [CrossRef]
- Liechty, Z.; Santos-Medellín, C.; Edwards, J.; Nguyen, B.; Mikhail, D.; Eason, S.; Phillips, G.; Sundaresan, V. Comparative analysis of root microbiomes of rice cultivars with high and low methane emissions reveals differences in abundance of methanogenic archaea and putative upstream fermenters. mSystems 2020, 5. [Google Scholar] [CrossRef]
- Kohlmeier, S.; Smits, T.H.M.; Ford, R.M.; Keel, C.; Harms, H.; Wick, L.Y. Taking the fungal highway: mobilization of pollutant-degrading bacteria by fungi. Environ. Sci. Technol. 2005, 39, 4640–4646. [Google Scholar] [CrossRef]
- Warmink, J.A.; Nazir, R.; Corten, B.; van Elsas, J.D. Hitchhikers on the fungal highway: The helper effect for bacterial migration via fungal hyphae. Soil. Biol. Biochem. 2011, 43, 760–765. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, T.; Friman, V.-P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic network architecture of root-associated bacterial communities determines pathogen invasion and plant health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wei, Z.; Friman, V.-P.; Gu, S.-h.; Wang, X.-f.; Eisenhauer, N.; Yang, T.-j.; Ma, J.; Shen, Q.-r.; Xu, Y.-c.; et al. Probiotic diversity enhances rhizosphere microbiome function and plant disease suppression. mBio 2016, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Wei, Z.; Shao, Z.; Friman, V.-P.; Cao, K.; Yang, T.; Kramer, J.; Wang, X.; Li, M.; Mei, X.; et al. Competition for iron drives phytopathogen control by natural rhizosphere microbiomes. Nat. Microbiol. 2020, 5, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Traxler, M.F.; Watrous, J.D.; Alexandrov, T.; Dorrestein, P.C.; Kolter, R. Interspecies interactions stimulate diversification of the Streptomyces coelicolor secreted metabolome. mBio 2013, 4, 10–1128. [Google Scholar] [CrossRef]
- Ola, A.R.B.; Thomy, D.; Lai, D.; Brötz-Oesterhelt, H.; Proksch, P. Inducing secondary metabolite production by the endophytic fungus Fusarium tricinctum through coculture with Bacillus subtilis. J. Nat. Prod. 2013, 76, 2094–2099. [Google Scholar] [CrossRef]
- Pishchany, G.; Mevers, E.; Ndousse-Fetter, S.; Horvath, D.J.; Paludo, C.R.; Silva-Junior, E.A.; Koren, S.; Skaar, E.P.; Clardy, J.; Kolter, R. Amycomicin is a potent and specific antibiotic discovered with a targeted interaction screen. Proc. Natl. Acad. Sci. USA 2018, 115, 10124–10129. [Google Scholar] [CrossRef]
- Jain, A.; Singh, S.; Kumar Sarma, B.; Bahadur Singh, H. Microbial consortium–mediated reprogramming of defence network in pea to enhance tolerance against Sclerotinia sclerotiorum. J. Appl. Microbiol. 2012, 112, 537–550. [Google Scholar] [CrossRef]
- Alizadeh, H.; Behboudi, K.; Ahmadzadeh, M.; Javan-Nikkhah, M.; Zamioudis, C.; Pieterse, C.M.J.; Bakker, P.A.H.M. Induced systemic resistance in cucumber and Arabidopsis thaliana by the combination of Trichoderma harzianum Tr6 and Pseudomonas sp. Ps14. Biol. Control 2013, 65, 14–23. [Google Scholar] [CrossRef]
- Postma, J.; Goossen-van de Geijn, H. Twenty-four years of Dutch Trig® application to control Dutch elm disease. BioControl 2016, 61, 305–312. [Google Scholar] [CrossRef]
Figure 1.
The phyllosphere, endosphere, and rhizosphere are depicted schematically as the major habitats for the diverse microflora that compose plant-associated communities.
Figure 1.
The phyllosphere, endosphere, and rhizosphere are depicted schematically as the major habitats for the diverse microflora that compose plant-associated communities.
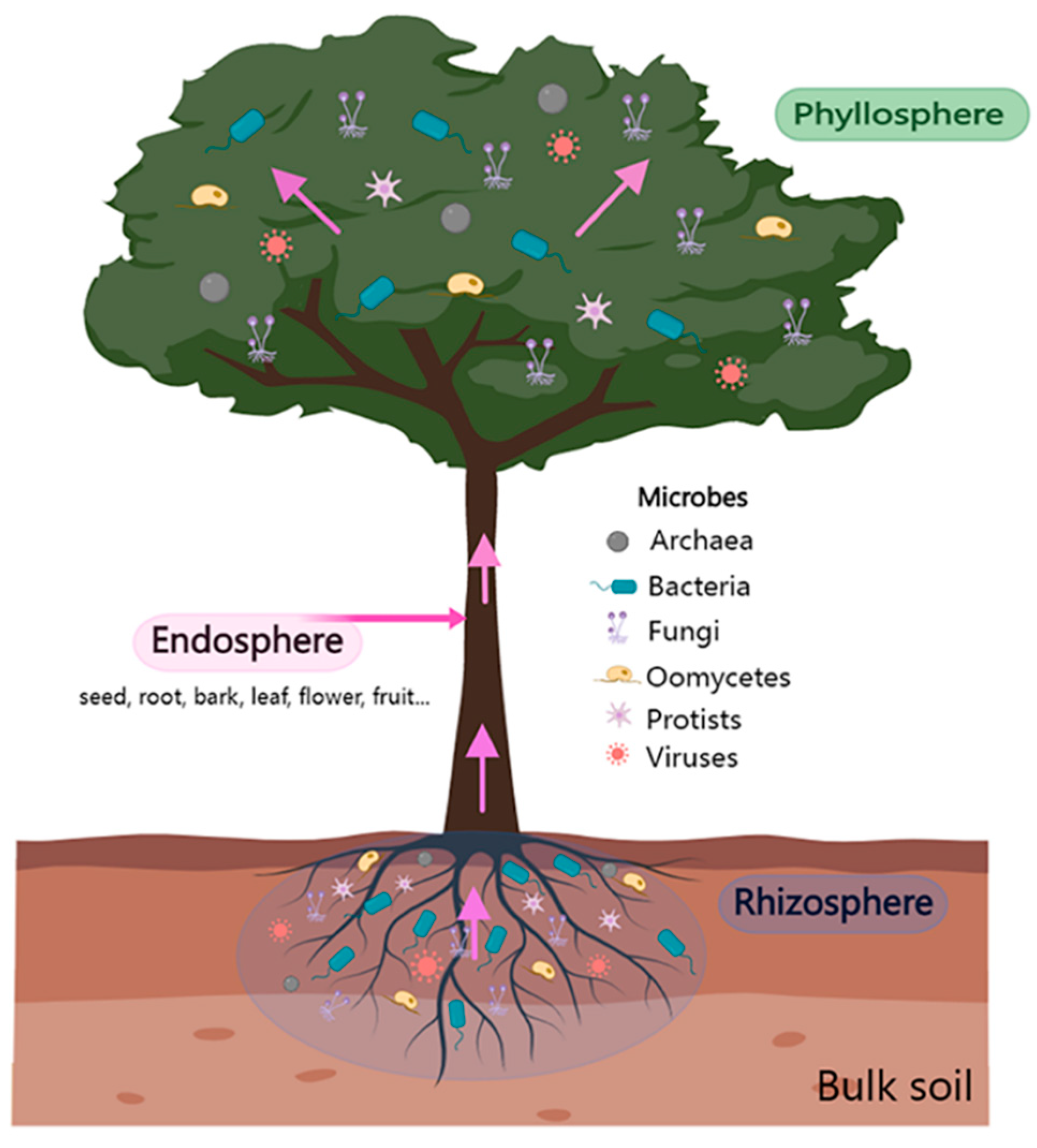
Figure 2.
Both interactions of the microbiome with the host and pathogens contribute to plant protection against pathogens. Plants infected by pathogens can recruit and enrich beneficial microbial communities to directly antagonize the pathogens via a variety of methods, such as the "cry for help" strategy. Specific mechanisms include secretion of antagonistic compounds (such as antibiotics, lysozyme, and volatile substances), competition with pathogens for ecological niche and nutrients, inhibition of pathogens’ signaling systems, parasitism and predation, and so on, thus directly inhibiting pathogen proliferation and infection. On the other hand, the plant microbiome can also act indirectly by priming plant immunity, such as induced systemic resistance (ISR) triggered by microbe-associated molecular patterns (MAMPs), defense hormones, plant growth-promoting rhizobacteria (PGPRs), and so on, to inhibit pathogen invasion and protect the underground and above-ground parts from encroachment.
Figure 2.
Both interactions of the microbiome with the host and pathogens contribute to plant protection against pathogens. Plants infected by pathogens can recruit and enrich beneficial microbial communities to directly antagonize the pathogens via a variety of methods, such as the "cry for help" strategy. Specific mechanisms include secretion of antagonistic compounds (such as antibiotics, lysozyme, and volatile substances), competition with pathogens for ecological niche and nutrients, inhibition of pathogens’ signaling systems, parasitism and predation, and so on, thus directly inhibiting pathogen proliferation and infection. On the other hand, the plant microbiome can also act indirectly by priming plant immunity, such as induced systemic resistance (ISR) triggered by microbe-associated molecular patterns (MAMPs), defense hormones, plant growth-promoting rhizobacteria (PGPRs), and so on, to inhibit pathogen invasion and protect the underground and above-ground parts from encroachment.
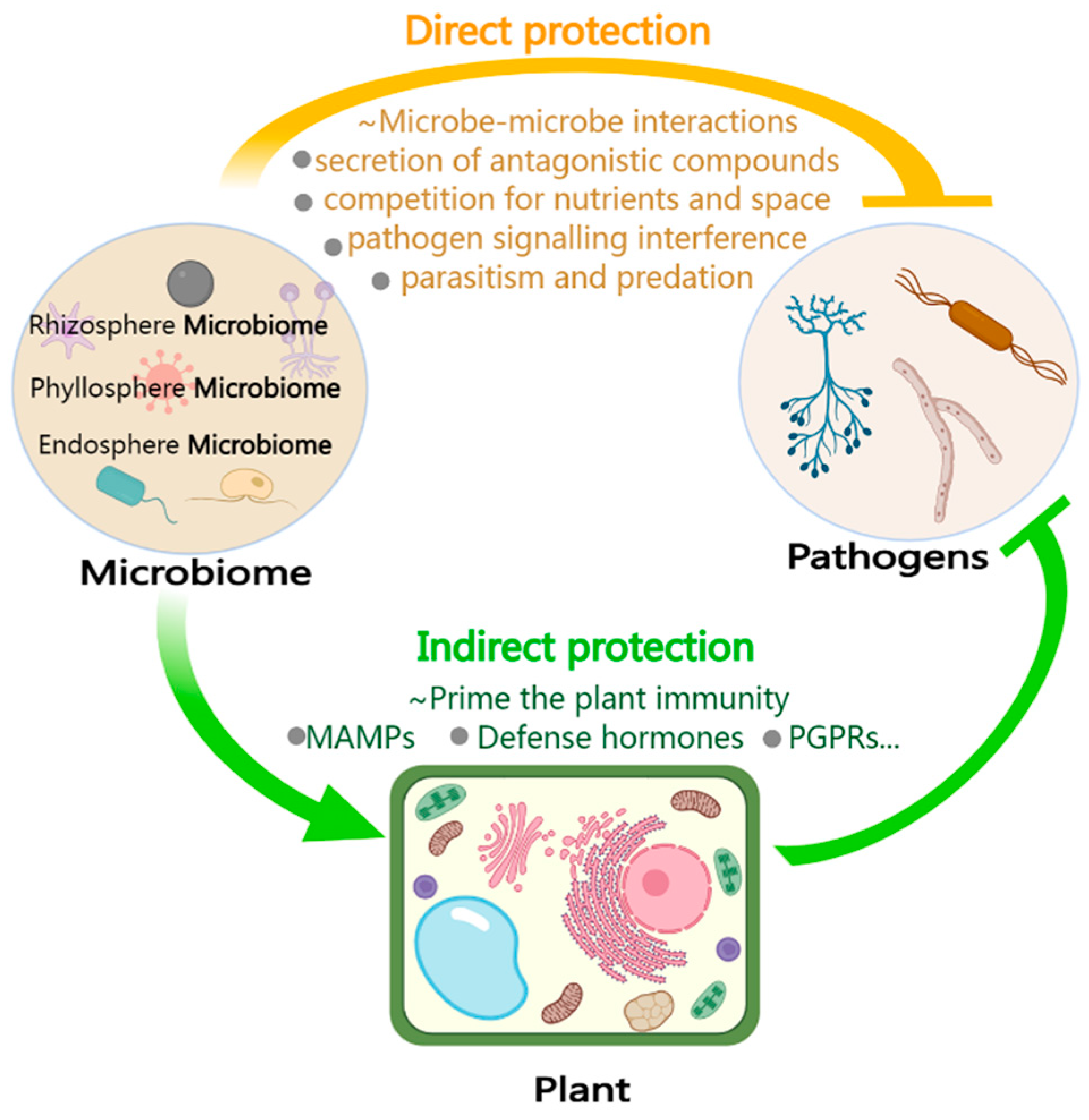
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



